Embed Size (px)
Citation preview

Morphological variability of the ParatethyanOligocene–Miocene small reticulofenestrid coccolitesand its paleoecological and paleogeographical implications
KATARÍNA HOLCOVÁ
Holcová, K. 2013. Morphological variability of the Paratethyan small reticulofenestrid coccolites and its paleoecologicaland paleogeographical implications. Acta Palaeontologica Polonica 58 (3): 651–668.
The analysis of size changes (length of placoliths, their width, length of central opening and its width) in ellipticalreticulofenestrids from the NP25–NN5 zonal interval of the Central Paratethys allowed to dicriminate two size catego−ries of placoliths: (i) small Reticulofenestra minuta (< 3.5 μm); (ii) Reticulofenestra haqii–pseudoumbilicus group(4–10.0 μm). The latter group appeared for the first time (FO, first occurrence) in the upper Egerian (size 4–7 μm) withthe size of placoliths in this plexus increasing gradually. The FO of R. pseudoumbilicus > 8.0 μm has been establishedin the Zone NN2 while its FCO (first common occurrence) in the Zone NN5. This study shows that the FOs of size−defined morphotypes of the R. haqii–pseudoumbilicus group differ in the Central Paratethys and oceanic realm.Blooms of R. minuta at the Oligocene–Miocene boundary and in the Early Middle Miocene may be correlated with theincoming of warm water into the higher latitude Central Paratethys basins during connection with the MediterraneanSea. Transgression favored the expansion of near−shore areas associated probably by some short−time oscillations ofsalinity. The FO of R. haqii–pseudoumbilicus group and the FO of R. pseudoumbilicus > 8.0 μm can be correlated withthe opening of new pathways between the Mediterranean and the Central Paratethys. Gradual size changes in theR. haqii–pseudoumbilicus group probably reflect climatic changes: the decrease of coccolith size in the late Egerianmay reflect cooling (Mi1 event) while the increase in coccolith size in the interval from the FO of Helicosphaeraampliaperta to the FO of Sphenolithus heteromorphus occurred due to warming. Two size categories of placoliths inthe R. haqii– pseudoumbilicus group (3.5–6 μm and 6–8 μm) recorded in the interval from Zone NN1 to the lower partof the Zone NN2 may represent seasonal populations.
Key words: Calcareous nannofossils, size changes, paleoecology, Oligocene, Miocene, Central Paratethys.
Katarína Holcová [[email protected]], Department of Geology and Paleontology, Charles University in Prague,Albertov 6, 128 43 Praha 2, Czech Republic.
Revised version received 6 September 2011, accepted 31 December 2011, available online 12 January 2012.
Copyright © 2013 K. Holcová. This is an open−access article distributed under the terms of the Creative Commons Attri−bution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original au−thor and source are credited.
Introduction
The collision of the African/Apulian/Arabian plates and Eur−asian continent began during the Eocene, and resulted in theuplift and emergence of the Alpine chains and the break−up ofthe Tethyan Realm into the Mediterranean and Paratethyandomains (Popov et al. 2004). This resulted in the differentia−tion of the Paratethys as a biogeographic entity. The firstendemic Paratethyan assemblages of mollusks, calcareousnannoplankton and foraminifera appeared in the Oligocene(Báldi 1986). The subsequent history of the Paratethys in−volved periods of isolation from the adjacent MediterraneanSea and Indian Ocean during which endemic biota evolved,and periods of oceanic communication during exchanges oc−curred with the faunas of the Mediterranean and the Indo−Pacific provinces. Additionally episodical paleogeographic
changes, global climatic variations, local oscillations of salin−ity, oxygen content and globally and locally affected sea−levelchanges also influenced the evolution of Paratethyan ecosys−tems (Rögl 1998; Kováč 2000; Popov et al. 2004; Piller et al.2007; etc.). Interactions of the local geodynamic and theglobal climatic factors lead to major problems in their bio−stratigraphical, paleoecological and paleogeographical inter−pretation. As a consequence of periodical paleogeographicchanges affecting them, the morphological changes (size) ofCentral Paratethys populations may be local, differing thosefrom contemporaneous oceanic populations.
Coccolithophorids were widespread autotrophs in marineParatethyan ecosystems and are of great biostratigraphical im−portance. Besides Coccolithus pelagicus, reticulofenestridsare the most common components of the assemblages andtheir detailed study can provide important data for reconstruct−
http://dx.doi.org/10.4202/app.2009.0006Acta Palaeontol. Pol. 58 (3): 651–668, 2013

ing the factors that have induced morphologic changes in cal−careous nannoplankton. Reticulofenestra bisecta, R. abisectaand Cyclicargolithus floridanus are common in the late Oligo−cene while in the lower and early Middle Miocene, reticulo−fenestrids are represented by Reticulofenestr minuta, R. haqii,and R. pseudoumbilicus together with endemic R. excavata(Lehotayová 1975, 1984; Marunteanu 1992; Andreyeva−Gri−gorovich et al. 1997; Holcová 2005). The aim of this study isto show the relationship between size changes in reticulo−fenestrids and paleogeographic and paleoecologic evolutionin an intracontinental basin.
Abbreviations.—FCO, first common occurrence; FO, first oc−currence; LCO, last common occurrence; LO, last occurrence.
Geological settingThe area of the Central Paratethys studied here includes thePannonian Basin system and the Carpathian Foreland basins(Seneš 1961).
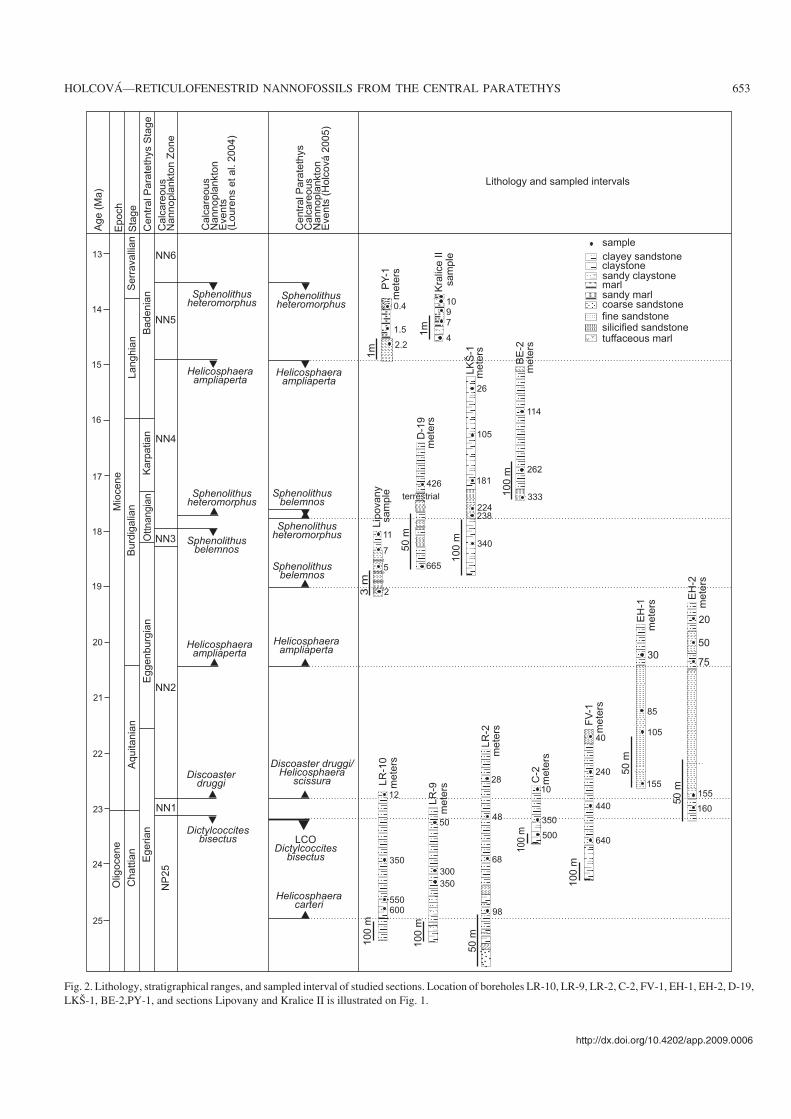
The majority of samples were taken from the South SlovakBasin (Fig. 1). Sediments filling of this geomorphological unitwere deposited in the northern part of the Buda Basin duringthe Late Oligocene (Kiscellian Stage) to the Chattian andlower Aquitanian (Egerian Stage), in the Fiľakovo/PétervásaraBasin during the upper Aquitanian and lower Burdigalian(Eggenburgian Stage) and in the Novohrad/Nógrád basin inthe upper Burdigalian and Langhian (Ottnangian, Karpatian,and early Badenian; Vass 1996). These three basins form thenorthern part of the Pannonian Basin system. Biostratigraphy,lithostratigraphy and sedimentology of this areawere summa−rized in Vass et al. (1979, 1989, 2007). Lithostratigraphic unitshave been defined by Vass and Elečko (1982) and correlationwith standard nannoplankton zones (Martini 1971) has beenestablished by Lehotayová (1982). The paleogeographicalmaps for every Central Paratethys stage have been producedby Vass et al. (1979, 1989, 2007). Vass et al. (1993), Márton etal. (1995), and Vass (1996) delineated the most important tec−tonic events and local sea−level changes; the latter by havebeen correlated with the global sea level curve of Haq et al.(1988) by Vass (1995).
The Middle Miocene samples have been collected fromsections in the Carpathian Foredeep. These sections are de−scribed in detail by Zágoršek et al. (2008, 2009) and Zágoršekand Holcová (2009; Fig. 1).
The study interval from the Oligocene/Miocene boundaryinterval to the lower Middle Miocene (zones NP25–NN5) cor−responds to the Egerian, Eggenburgian, Ottnangian, Karpa−tian, and Lower Badenian stages of the Central Paratethys(Popov et al. 2004; Harzhauser and Piller 2007; Hohenegger etal. 2009).
652 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
R.Sobota
Vepor
0 10 20 km
Hajnaèka
Fi¾akovo
JesenskéLR-9
C-2
Luèenec
BE-2 Lipovany
LR-10
BOHEMIAN MASSIVE
VIENNA
BASINZnojmo
Zlín
Ostrava
SLOVA
KREPUBLI
C
PO
LAN
D
AUSTRIA
Kralice n.O.
PY-1
CARPATHIAN
FOREDEEP
LKŠ-1EH-1
EH-2FV-1
0 50 100 km
0 50 100 km
Torna¾a
LR-2
Brno
Kralice S
VIENNA
BASIN
DANUBEBASIN
Budapest Bebrecen
Lviv
Kraków FOREDEEPOUTER FLYSCH CARPATHIANS
WESTERN CARPATHIANSTRANSCARPATHIAN
BASIN
Hodonín
Olomouc
Brno
Bohemian Massif�dánice Unit
Pouzdøany Unit
Raèa UnitBílé Karpaty Unit
PY-boreholes
relicts of the Lower Miocene sediments
Silesian and Subsilesian units
Bystrica Unit
Pre-Cenozoic units
Nógrád/Novohrad Basin
Hungarian Paleogene Basin
Karpatian/Badenian ryolite tuff
Fi¾akovo/Pétervására Basin
Middle Miocene volcanics
Szecsény and Putnok Schlierterrigenous Upper MiocenesedimentsEggenburgian ryodacite tuff
Upper Miocene–Pleistocenebasalts
Fig. 1. A. Sketch of the main Middle European geological units (after Kováč 2000). B. Moravian part of the Carpathian Foredeep with location of boreholePY−1 near village Přemyslovice; and section Kralice (KRA), geological situation according Chlupáč et al. (2002). C. Eastern part of the South Slovak Basinwith location of boreholes LKŠ−1, BE−2, FV−1, EH−1, and EH−2 drilled by State Geological Institute of Dionýz Štúr; boreholes LR−9 and LR−10 drilled by“Geologický prieskum n.p.” regional centrum Rožňava and section Lipovany (LIP). Geological situation after Halásová et al. (1996).
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 653
Olig
oce
ne
Mio
cen
e
Ch
att
ian
Aq
uita
nia
nB
urd
iga
lian
La
ng
hia
nS
err
ava
llia
n
Eg
eria
nE
gg
en
bu
rgia
nO
ttnan
gian
Ka
rpa
tian
Ba
de
nia
n
NP
25
NN1
NN2
NN3
NN4
NN5
NN6
Ag
e(M
a)
Ep
och
Sta
ge
Ce
ntr
al P
ara
teth
ysS
tag
e
Ca
lca
reo
us
Na
nn
op
lan
kto
nZ
on
e
13
14
15
16
17
18
19
20
21
22
23
24
25
1m
1m
10
Kra
lice
II
7
4
9
claystoneclayey sandstone
sandy claystone
sandy marlmarl
coarse sandstonefine sandstone
10
0m
10
0m
50
m
50
m
50
m
10
0m
10
0m
100
m
10
0m
LR
-9
LR
-2
LR
-10
C-2
FV
-1
EH
-1
EH
-2
terrestrial
D-1
9
LK
Š-1
BE
-2
600
350
500640
440
240
85
105
155155
160
75
50
665
340
333
262
114
0.4
1.5
2.2
238224
181
105
26
426
20
40
30
350
550
350300
28
12
50
10
98
68
48
Lip
ova
ny
3m
sam
ple
11
7
5
2
PY
-1
sam
ple
sample
silicified sandstonetuffaceous marl
mete
rs
mete
rs
mete
rs
mete
rs
mete
rs mete
rs
mete
rs
mete
rs
mete
rs
mete
rs
mete
rs
Sphenolithusheteromorphus
Sphenolithusheteromorphus
Sphenolithusheteromorphus
Sphenolithusheteromorphus
Sphenolithusbelemnos
Sphenolithusbelemnos
Sphenolithusbelemnos
Helicosphaeraampliaperta
Helicosphaeraampliaperta
Helicosphaeraampliaperta
Helicosphaeraampliaperta
Discoasterdruggi
Discoaster druggi/Helicosphaera
scissura
Dictylcoccitesbisectus LCO
Dictylcoccitesbisectus
Ca
lca
reo
us
Na
nn
op
lan
kto
nE
vents
(Lo
ure
ns
eta
l.2
00
4)
Ce
ntr
al P
ara
teth
ysC
alc
are
ou
sN
an
no
pla
nkt
on
Eve
nts
(Ho
lco
vá2
00
5)
Helicosphaeracarteri
Lithology and sampled intervals
50
m
Fig. 2. Lithology, stratigraphical ranges, and sampled interval of studied sections. Location of boreholes LR−10, LR−9, LR−2, C−2, FV−1, EH−1, EH−2, D−19,LKŠ−1, BE−2,PY−1, and sections Lipovany and Kralice II is illustrated on Fig. 1.

Material and methodsA total of 49 samples taken from 13 sections have been studied.The set of samples were chosen from about 500 of samplesanalysed in the study area in previous years (e.g., Holcová2001, 2005). Nannofossils in selected samples were commonto abundant (10–30 specimens in visual field of microscope)and well preserved without apparent diagenetic changes (dis−solution, recrystallization). Relative abundances of reticulo−fenestrids in studied samples varied from 10 to 30% with ex−ception of samples from the zones NN1 and NN5 with morethan 50% of reticulofenestrids. Besides Langhian samples(Zone NN5) were dominated by Coccolithus pelagicus in thestudied assemblages. The location of the sections, their lithol−ogy and stratigraphical ranges are summarized in Figs. 1 and 2.
Calcareous nannofossils were examined and photographedusing a light microscope and 1000× magnification with bothbright field and crossed polarized light. Smear slides were pre−pared from about 1 cc of rock sample (claystone, siltstone, andfine sandstone) using the method of Švábenická (2002). Thefirst 50 specimens of reticulofenestrids were measured in ev−ery sample. Placoliths of Reticulofenestra spp. were measuredfrom the digital microphotographs. In total, 2447 specimenswere analyzed. The following characters were measured foreach placolith: length (the largest diameter of placoliths),width (diameter of placolith perpendicular to the length of theplacolith), length of central opening (along the length of theplacolith) and its width (across the width of the placolith)(Fig. 3).
The measurements were analyzed by simple statisticalmethods using STATISTICA software. Frequency histo−grams were used for the first graphical evaluation of the dis−tribution type of a simple biometric parameter. XY−plotsand the Pearson correlation coefficient were used for evalu−ation of correlations between two parameters.
The size−dependent classification of Reticulofenestraminuta–haqii–pseudoumbilicus group (Young 1999) has beenused as an initial taxonomic concept (Reticulofenestra minuta:< 3 μm, R. haqii: 3–5 μm, and R. pseudoumbilicus > 5 μm).This classification is broadly accepted also in the Central Para−
tethys (Ćorić and Švábenická 2004; Tomanová Petrová andŠvábenická 2007; Ćorić and Hohenegger 2008). However, thesmall−sized placoliths identified like Reticulofenestra minutamay represent two or more species which cannot be distin−guished under the light microscope (Haq 1980).
To analyze size changes in reticulofenestrids over time, thestudied interval was divided into seven intervals on the basisof six calcareous nannoplankton events. The age of thesebioevents in the Central Paratethys may differ from their agein the world ocean (Berggren et al. 1995; Lourens et al. 2004)because new taxa were only able to migrate into the CentralParatethys when communication existed between them.
(i) The FO of Helicospheara carteri was recorded withinZone NP 25 in the study area (Holcová 2005).
(ii) The LCO of Reticulofenestra bisecta. The extinction ofthis species is often used to approximate the Oligocene/Mio−cene boundary (Berggren et al. 1995; Young 1999). However,in the Buda Basin, rare specimens of R. bisecta occur in zonesNN1 and NN2 (Holcová 2005). These are probably reworkedspecimens and therefore only the LCO of R. bisecta was usedto characterize the NP25/NN1 zonal boundary.
(iii) The FO of Helicosphaera scissura and the FO ofDiscoaster druggii are isochronous events in the Central Para−tethys (Holcová 2005). Helicosphaera scissura is, however,more abundant and its FO is more easily determined.
(iv) The FO of H. ampliaperta was recorded in Zone NN2in the Central Paratethys (Marunteanu 1992; Holcová 2002,2005).
(v) The FO of S. belemnos was established to lie at theNN2/NN3 boundary in the Central Paratethys (Marunteanu1992, Andreyeva−Grigorovich et al. 1997).
(vi) The FO of S. heteromorphus/LO of S. belemnos: Inthe Central Paratethys both species briefly co−occur.
(vii) For comparison reticulofenestrids from Zone NN5were studied. This zone is characterized by the absence ofH. ampliaperta and the presence of S. heteromorphus (Mar−tini 1971).
Seven samples were intended to analyze from every inter−val. However, from the LO of Helicosphaera ampliaperta tothe FO of Sphenolithus heteromorhus (upper part of the ZoneNN2 and Zone NN3), terrigenous sedimentation dominatedin the study area. Therefore only limited number of sampleswith well preserved and abundant nannofossils was found inthis interval. On the other hand, longer intervals with varie−gated marine facies (zones NP25 and NN4) were sampled tomore detail.
Results
Morphologic variability of placoliths
Size and length/width−ratio of placoliths and size of centralopening.—The length of the placoliths varies from 1.6 to10 μm, their width from 1 to 9 μm. The length−width scatterplot and the correlation coefficient (0.95) between the length
654 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
lp
wp
lowo
Fig. 3. Dimensions measured in placoliths. Abbreviations: lo, length of cen−tral opening; lp, length of placoliths; wo, width of central opening; wp,width of placoliths.
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 655
r = 0.95
r = 0.83
r = 0.79
1
1
2
2
0
0
3
3
4
4
5
5
6
6
7
7
8
8
9
9
1
2
3
4
5
6
7
8
Length of nannoliths (um)
Length of nannoliths (um)
Length of central opening ( m)μ
Wid
tho
fn
an
no
lith
s(u
m)
Wid
tho
fce
ntr
al o
pe
nin
g(u
m)
Le
ng
tho
fce
ntr
al o
pe
nin
g(μ
m)
10
10
0
0
0
1
1
2
2
3
3
1 2 3 4
n = 2447
n = 2447
n = 2447
R. minuta <3μm
R. haqii–pseudoumbilicus group 3–6μm
R. haqii–pseudoumbilicus group > 8.5 μm
R. haqii–pseudoumbilicus group 6–8.5 μm
Nu
mb
er
ofsp
eci
me
ns
Nu
mb
er
ofsp
eci
me
ns
Nu
mb
er
ofsp
eci
me
ns
Length of central opening (μm)
Length of central opening/lenght of nannolith
Length of nannolith (μm)
Reticulofenestra minuta Reticulofenestra haqii–pseudoumbilicus group R. haqii–pseudoumbilicus group
250
200
150
100
50
0
250
200
150
100
50
0
350
300
250
200
150
100
50
0
R. minuta
n = 2447
n = 2447
n = 2447
0.3
0–
0.5
00
.50
–0
.65
0.6
5–
0.8
00
.80
–0
.95
0.9
5–
1.1
01
.10
–1
.25
1.2
5–
1.4
01
.40
–1
.55
1.5
5–
1.7
01
.70
–1
.85
1.8
5–
2.0
02
.00
–2
.15
2.1
5–
2.3
02
.30
–2
.45
2.4
5–
2.6
02
.60
–2
.75
2.7
5–
2.9
02
.90
–3
.05
3.0
5–
3.2
03
.20
–3
.35
3.3
5–
3.5
0
1.6
6–
2.0
02
.00
–2
.33
2.3
3–
2.6
62
.66
–3
.00
3.0
0–
3.3
33
.33
–3
.66
3.6
6–
4.0
04
.00
–4
.33
4.3
3–
4.6
64
.66
–5
.00
5.0
0–
5.3
35
.33
–5
.66
5.6
6–
6.0
06
.00
–6
.33
6.3
3–
6.6
66
.66
–7
.00
7.0
0–
7.3
37
.33
–7
.66
7.6
6–
8.0
08
.00
–8
.33
8.3
3–
8.6
68
.66
–9
.00
9.0
0–
9.3
39
.33
–9
.66
9.6
6–
10
.00
0.1
5–
0.1
7
0.1
7–
0.1
9
0.1
9–
0.2
1
0.2
1–
0.2
3
0.2
3–
0.2
5
0.2
5–
0.2
7
0.2
7–
0.2
9
0.2
9–
0.3
1
0.3
1–
0.3
3
0.3
3–
0.3
5
0.3
5–
0.3
7
0.3
7–
0.3
9
0.3
9–
0.4
1
0.4
1–
0.4
3
0.4
3–
0.4
5
0.4
5–
0.4
7
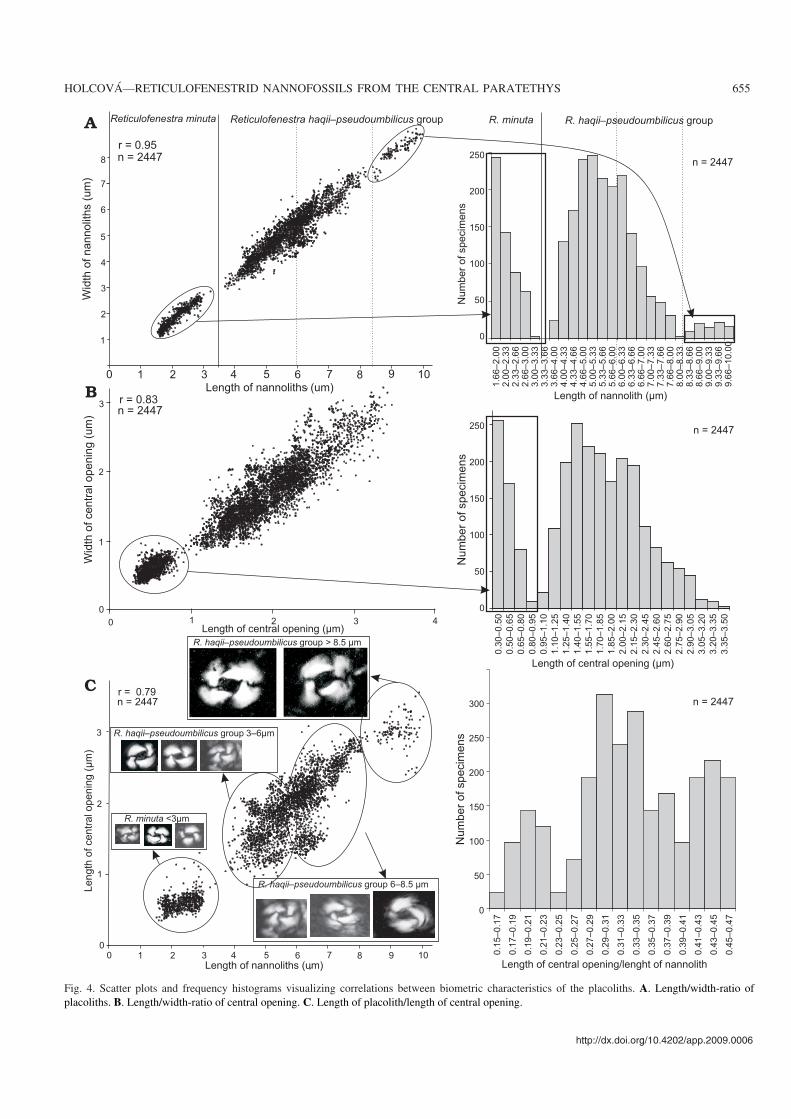
Fig. 4. Scatter plots and frequency histograms visualizing correlations between biometric characteristics of the placoliths. A. Length/width−ratio ofplacoliths. B. Length/width−ratio of central opening. C. Length of placolith/length of central opening.

and width of the placoliths indicate a very high correlation be−tween these two parameters (Fig. 4A). The distribution ofthese characters is polymodal and shows three groups that arealso distinct on the width−length plot (Fig. 4A): (i) smallplacoliths (1.6–3 μm); (ii) medium−size placoliths (3.5–8 μm),and (iii) large placoliths (8–10 μm). The small placoliths con−stitute a distinct size group, but transitional size categories oc−cur between the medium−size and large placoliths.
Relative size and length/width−ratio of central opening.—The size of the central opening varied from 0.35 to 3.5 μm.The correlation between the length and width of the centralopening is high (0.83) with two groups in which the lengthare (i) 0.35–1 μm and (ii) 1–3.5 μm (Fig. 4B).
The relative size of the central opening was expressed as aratio between the size of the placolith and the size of the cen−tral opening (Fig. 4C). The ratio varies from 0.15 to 0.48, andshows a polymodal distribution with three groups: (i) placo−liths with narrow central opening (size of placolith/size ofcentral opening varies from 0.15 to 0.24), the placoliths withnarrow central opening are small and reach maximum diam−eter 7.5 μm; (ii) placoliths with medium central opening (sizeof placolith/size of central opening from 0.25 to 0.40) wererecorded in all size categories of placoliths; (iii) placolithswith wide central opening (size of placolith/size of centralopening from 0.40 to 0.48) were not recorded among thesmallest placoliths (1.6–3 μm).
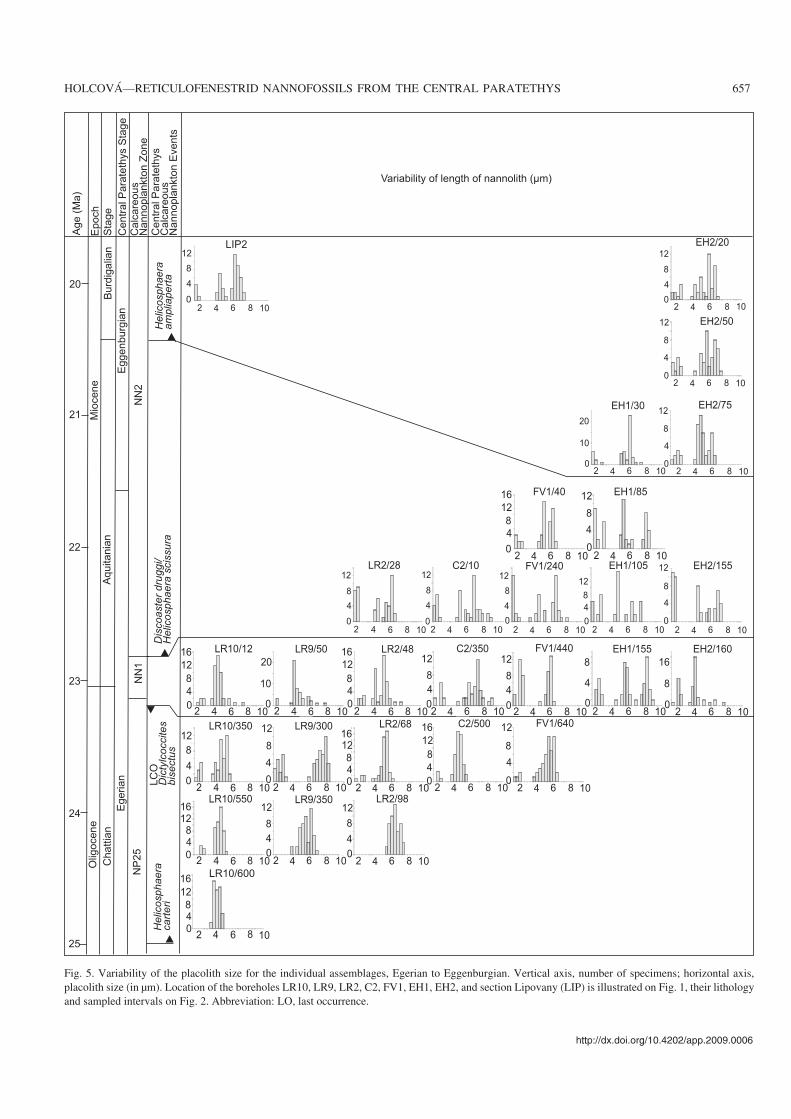
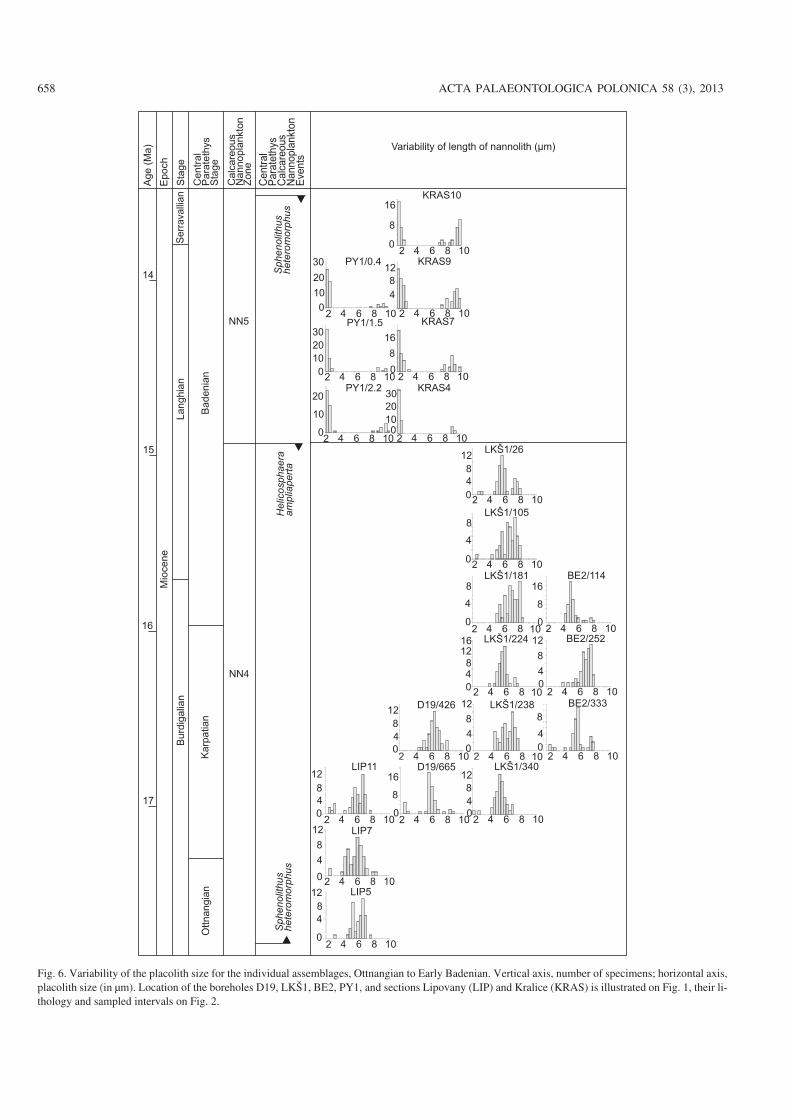
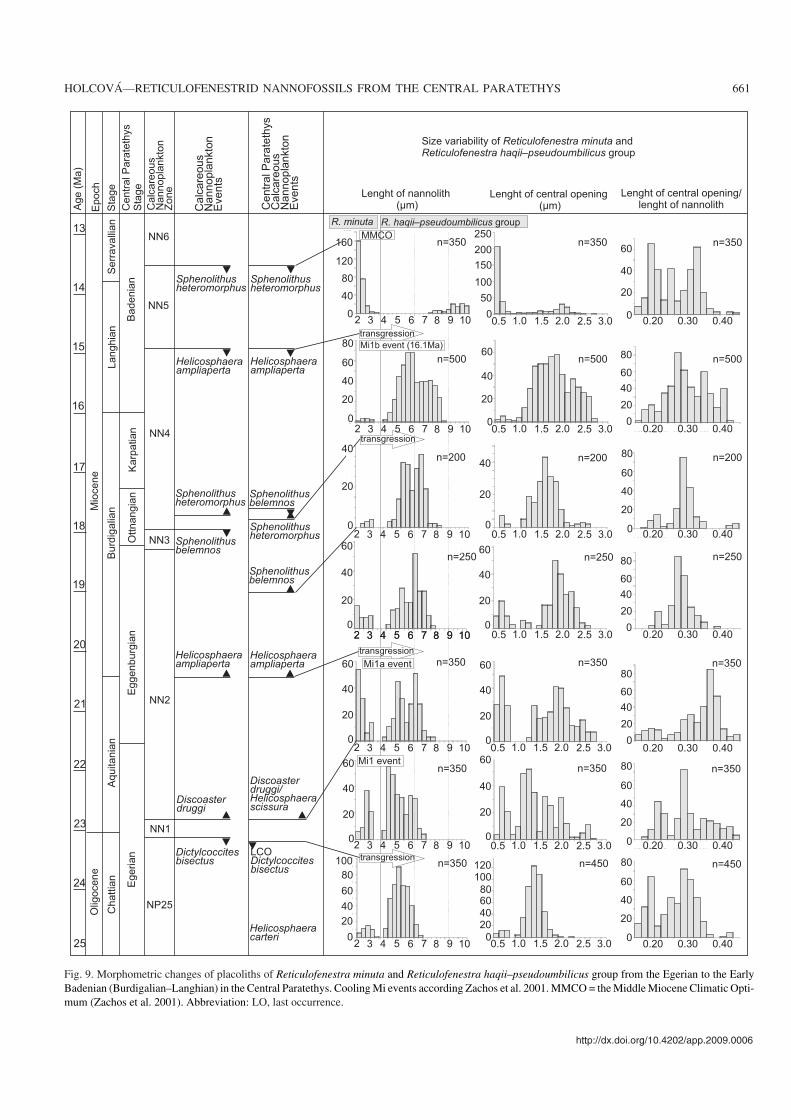
Size changes over timeCorrelations between the parameters analyzed here for placo−liths and the age of samples were investigated on the basis ofcorrelation coefficient. A negative correlation was recordedbetween the length of the placoliths (p = 0.01) and their age.The shift in the total size of placoliths and that of their centralopening over time is illustrated in Figs. 5–8 for individualsamples. A summary of the data is showed in Fig. 9. Fromthese data we may conclude the following:
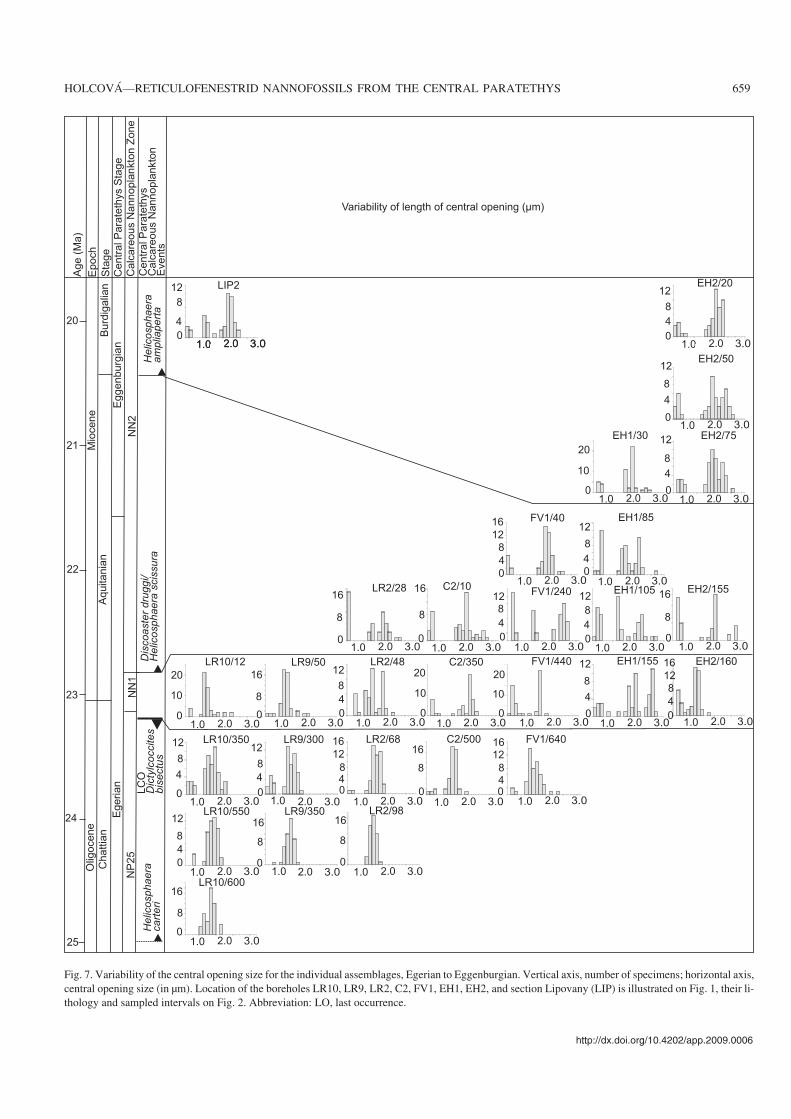
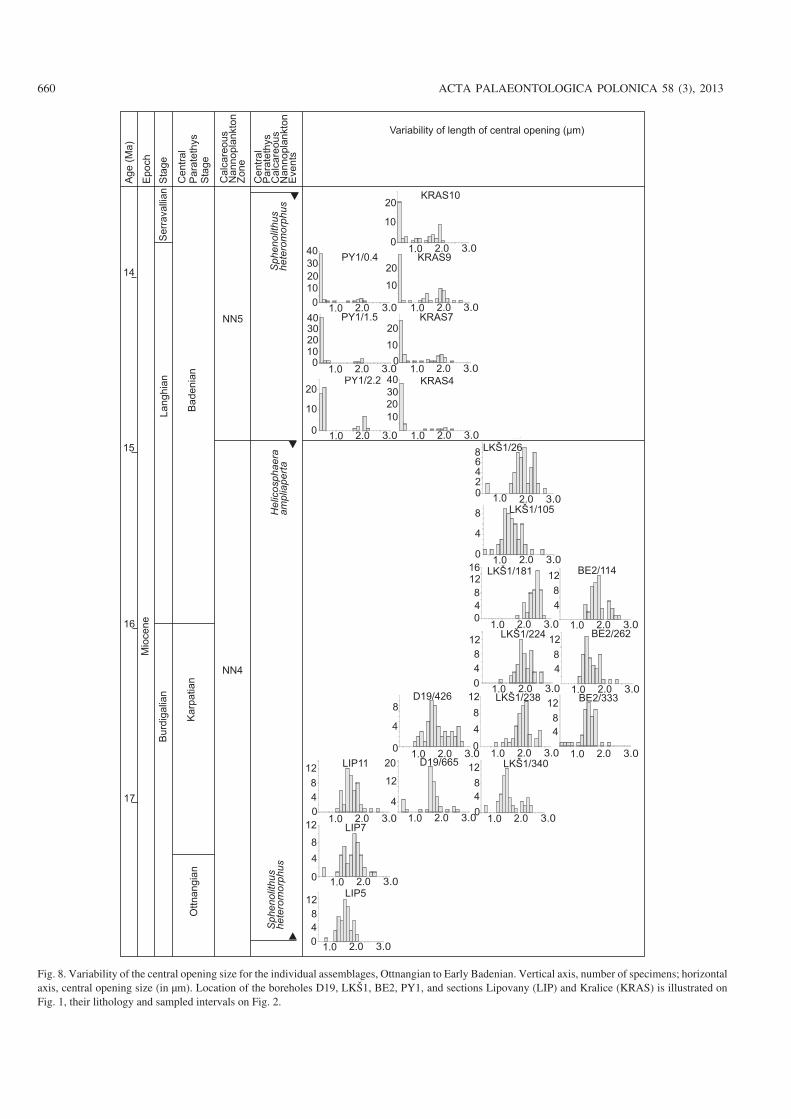
(i) Small placoliths (1.6–3 μm) constitute a well separatedgroup in all assemblages, independent of age. However, theyare abundant in two intervals: (1) in the lowest Miocene(Zone NN1 and lower part of Zone NN2 below the FO of H.ampliaperta), in these intervals they account for 30–70% ofthe nannoplankton assemblages; (2) in the lower MiddleMiocene (Zone NN5), where they account for 50–95% of theassemblages.
(ii) The size of the oldest placoliths in the R. haqii–pseudo−umbilicus group (upper Zone NP25) varies from 4.0–7.0 μm.Than the size of placoliths in the group slightly increasedthough this trend may differ in some individual samples (Figs.5, 6). In the Zone NN3, the size varies from 4.0–8.0 μm.
(iii) Bimodal size distribution was recorded firstly duringZone NN1; it dominates in the lower part of the NN2 Zone(Figs. 9, 10).
(iv) Wide variations in the relative size of the centralopening characterize interval from zones NP25 to lowerpart of NN2; generally, the relative size of the central open−
ing increases (Fig. 9). At the level of the FO of H. ampli−aperta, larger R. haqii (6–7 μm) with small central opening(1.7–2.0 μm) appeared, and relative size of central openingat this level decreased (Fig. 9).
(v) Broad variability of size of placolith as well as of thecentral opening characterizes the assemblages in Zone NN4(Fig. 9).
(vi) Large placoliths (> 8 μm) occur first at the Zone NN5(Fig. 9).
Discussion
Taxonomic implications
The size of placoliths is commonly used for classification ofReticulofenestra species. Using principal component analysis,Backman (1980) showed that the majority of morphologicalvariation in Reticulofenestra can be explained by variation inthe size of placoliths. However, the degree of closure of thecentral area can also be used for classification of Reticulo−fenestra (Backman 1978, 1980; Pujos 1985). Young (1990)suggested that this latter character is an unstable ecopheno−typic character, and based his classification of the Late Mio−cene–Pliocene Reticulofenestra mainly on the length of theplacoliths.
Young (1999) also differentiated the Early Miocene R.minuta–haqii–pseudoumbilicus group on the basis of overallsize of the placoliths. The taxonomic difference between R.minuta and R. haqii was based on a superior limit size of3 μm for the former. This criterion is well applicable to thereticulofenestrids of the Central Paratethys among which 3.1to 3.7 μm long placoliths have not been observed (Fig. 4A).
Two different size limits have been used to separateR. haqii from R. pseudoumbilicus: Backman (1980) pro−posed an upper size limit of 5 μm for R. haqii, whereas Rioet al. (1990) and Raffi et al. (1995) proposed 7 μm. In anycase, R. haqii and R. pseudoumbilicus represent a continuousplexus in world ocean and therefore Young (1990) classifiedthem as one species, R. pseudoumbilicus, with two varieties:R. p. pseudoumbilicus and R. p. haqii. Nevertheless, separa−tion of two species at 5 μm has prevailed.
As in the oceanic realm, the length of the placoliths in theR. haqii–pseudoumbilicus group changes continuously in theCentral Paratethys. Limit of 5 μm has also been used for sep−aration of R. pseudoumbilicus and R. haqii (Ćorić and Švábe−nická 2004; Tomanová Petrová and Švábenická 2007; Ćorićand Hohenegger 2008). In addition, Ćorić and Hohenegger(2008) have distinguished two size groups among R. pseudo−umbilicus, one between 5–7 μm, the other > 7 μm. In studiedmaterial, boundary 8 μm seems to be pronounced. It appearsthat separation of group > 8 μm is specific only for studied setof samples because qualitative examination of placolithsfrom the other Central Paratethys sections of Zone NN5 andNN6 showed gradual changes in size of R. pseudoumbilicusfrom 5 to 10 μm (Jamrich and Halásová 2010).
656 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 657
0
2
4
6
8
10
12
14
16
18
LR10/600
0
2
4
6
8
10
12
14
16
18
LR10/550
0
2
4
6
8
10
12
14
LR10/350
0
2
4
6
8
10
12
14
LR9/350
0
2
4
6
8
10
12
14
16
LR10/12
0
2
4
6
8
10
12
14
FV1/440
0
1
2
3
4
5
6
7
8
9
10
EH1/155
0
2
4
6
8
10
12
14
16
18
20
EH2/160
0
2
4
6
8
10
12
14
16
EH2/155
0
2
4
6
8
10
12
EH1/85
0
2
4
6
8
10
12
14
FV1/240
0
2
4
6
8
10
12
EH2/75
0
5
10
15
20
25
EH1/30
0
2
4
6
8
10
12
EH2/50
2
2 2
4
4 4
8
8 8
10
100
2
4
6
8
10
12
14
LR2/98
0
2
4
6
8
10
12
LR9/300
0
2
4
6
8
10
12
14
16
18
LR2/68
0
2
4
6
8
10
12
14
16
C2/500
0
2
4
6
8
10
12
FV1/640
0
5
10
15
20
25
LR9/50
0
2
4
6
8
10
12
14
16
LR2/48
0
2
4
6
8
10
12
14
C2/350
0
2
4
6
8
10
12
14
LR2/28
0
2
4
6
8
10
12
14
C2/10
0
2
4
6
8
10
12
EH2/155
0
2
4
6
8
10
12
14
16
FV1/40
0
2
4
6
8
10
12
14
EH2/20
4
4
44
4
4
4
4
4 4 44
4
4
4 4
4
4
4
4
4
8
8
8
84
8
8
8
10
10
8
8
8 8 88
8
8
8 8
8
8
8
8
8
12
12
1212
12
12
20
20
12
12
12 1212
12
12
12 12
12
12
16
16
168
16
16
16
0
0
0
00
0
0
0
0
0
0
0 0 00
0
0
EH1/105
FV1/40 EH1/85
EH1/30 EH2/75
EH2/50
EH2/20
0
0 0 0
0
0
0
0
EH2/155
4
4
8
8
12
12
0
0
He
lico
sp
ha
era
am
plia
pe
rta
Dis
co
aste
rd
rug
gi/
He
lico
sp
ha
era
scis
su
ra
LC
OD
icty
lco
ccite
sb
ise
ctu
s
Olig
oce
ne
Mio
cen
eC
ha
ttia
nA
qu
itan
ian
Bu
rdig
alia
nE
ge
ria
nE
gg
en
bu
rgia
n
NP
25
NN
1N
N2
Ag
e(M
a)
Ep
och
Sta
ge
Ce
ntr
al P
ara
teth
ysS
tag
eC
alc
are
ou
sN
an
no
pla
nkt
on
Zo
ne
Ce
ntr
al P
ara
teth
ysC
alc
are
ou
sN
an
no
pla
nkt
on
Eve
nts
20
21
22
23
24
25
He
lico
sp
ha
era
cart
eri
EH2/160EH1/155FV1/440C2/350LR2/48LR9/50LR10/12
12 1216LR9/300LR10/350 LR2/68 C2/500 FV1/640
LR2/98LR9/350LR10/550
FV1/240C2/10LR2/28
12
Variability of length of nannolith (μm)
0
2
4
6
8
10
12
14
LIP2
4
8
12
0
LIP2
6
6 6 10LR10/600
2 4 86 10
2
2
2
22
2
2
2
2
2
2
2
2
4
4
4
44
4
4
4
4
4
4
4
4
8
8
8
88
8
8
8
8
8
8
8
8
10
10
10
1010
6
6
6
66
6
6
6
6
6
6
6
6
10
10
10
10
10
10
10
10
2
2
2
2
2
2
2
2
2
22
4
4
4
4
4
4
4
4
4
44
8
8
8
8
8
8
8
8
8
88
6
6
6
6
6
6
6
6
6
66
10
10
10
10
10
10
10
10
10
1010
16
12
Fig. 5. Variability of the placolith size for the individual assemblages, Egerian to Eggenburgian. Vertical axis, number of specimens; horizontal axis,placolith size (in μm). Location of the boreholes LR10, LR9, LR2, C2, FV1, EH1, EH2, and section Lipovany (LIP) is illustrated on Fig. 1, their lithologyand sampled intervals on Fig. 2. Abbreviation: LO, last occurrence.
658 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
0
2
4
6
8
10
12
BE2/262
0
2
4
6
8
10
12
14
16
BE2/333
0
2
4
6
8
10
12
14
16
18
20
BE2/262
Sp
he
no
lith
us
he
tero
mo
rph
us
Sp
he
no
lith
us
he
tero
mo
rph
us
He
lico
sp
ha
era
am
plia
pe
rta
La
ng
hia
nS
erra
valli
an
Ott
na
ng
ian
Ka
rpa
tian
Ba
de
nia
n
NN4
NN5
14
15
16
170
2
4
6
8
10
12
14
LKŠ1/340
0
2
4
6
8
10
12
LKŠ1/238
0
2
4
6
8
10
12
14
16
LKŠ1/224
0
1
2
3
4
5
6
7
8
9
10
LKŠ1/181
0
1
2
3
4
5
6
7
8
9
10
LKŠ1/105
0
2
4
6
8
10
12
14
LKŠ1/26
2 3 4 5 6 7 8 9
0
2
4
6
8
10
12
LIP7
0
2
4
6
8
10
12
14
LIP11
0
2
4
6
8
10
12
14
16
18
20
D19/665
0
2
4
6
8
10
12
14
D19/426
0
5
10
15
20
25
PY1/2.2
0
5
10
15
20
25
30
35
PY1/1.5
0
5
10
15
20
25
30
PY1/0.4
0
5
10
15
20
25
30
35
KRAS4
0
2
4
6
8
10
12
14
16
18
20
KRAS7
0
2
4
6
8
10
12
14
KRAS9
0
2
4
6
8
10
12
14
16
18
KRAS10
4 4
8
10
10
10
10
8
8
8
8
8
20
20
20
20
16
30
30
30
16
16
16
16
0
0
0
0
0
0
0
0
0
0
LIP5
LKŠ1/224
KRAS4PY1/2.2
BE2/114LKŠ1/181
KRAS7PY1/1.5
KRAS9PY1/0.4
LKŠ1/105
KRAS10
LKŠ1/26
0
2
4
6
8
10
12
14
LIP5
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
2
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
4
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
6
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
8
10
10
10 10
10
10
10
10
10
10
10
10
10
10
10
10
4
4
4
4
4
4
4
4
4 44
8
8
8
8
8
8
8
8
8 88
12
12
1212
12
12
1212
1212
0
0
0
0
0
0
0
0 00
Mio
cen
e
Bu
rdig
alia
n
Ag
e(M
a)
Ep
och
Sta
ge
Ce
ntr
al
Pa
rate
thys
Sta
ge
Ca
lca
reo
us
Na
nn
op
lan
kto
nZ
on
e
Cen
tral
Par
atet
hys
Cal
care
ous
Nan
nopl
ankt
onE
vent
s
Variability of length of nannolith (μm)
LIP7
LKŠ1/340
BE2/333LKŠ1/238D19/426
LIP5
BE2/252
LIP11 D19/665
10
10
10
10
10
Fig. 6. Variability of the placolith size for the individual assemblages, Ottnangian to Early Badenian. Vertical axis, number of specimens; horizontal axis,placolith size (in μm). Location of the boreholes D19, LKŠ1, BE2, PY1, and sections Lipovany (LIP) and Kralice (KRAS) is illustrated on Fig. 1, their li−thology and sampled intervals on Fig. 2.
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 659
16
He
lico
sp
ha
era
am
plia
pe
rta
Dis
co
aste
rd
rug
gi/
He
lico
sp
ha
era
scis
su
raL
CO
Dic
tylc
occite
sb
ise
ctu
s
Olig
oce
ne
Mio
cen
eC
ha
ttia
nA
qu
itan
ian
Bu
rdig
alia
nE
ge
ria
nE
gg
en
bu
rgia
nN
P2
5N
N1
NN
2
Ag
e(M
a)
Ep
och
Sta
ge
Ce
ntr
al P
ara
teth
ysS
tag
eC
alc
are
ou
sN
an
no
pla
nkt
on
Zo
ne
Ce
ntr
al P
ara
teth
ysC
alc
are
ou
sN
an
no
pla
nkt
on
Eve
nts
20
21
22
23
24
25
He
lico
sp
ha
era
ca
rte
ri
LR2/68 C2/500
Variability of length of central opening (μm)
0
2
4
6
8
10
12
14
16
18
20
LR10/600
8
16
0
0
2
4
6
8
10
12
14
LR10/550
0
2
4
6
8
10
12
14
16
18
20
LR9/350
0
2
4
6
8
10
12
14
16
18
20
LR2/98
48 88
12 1616
0 00
0
2
4
6
8
10
12
LR10/350
0
2
4
6
8
10
12
14
LR9/300
0
2
4
6
8
10
12
14
16
LR2/68
0
2
4
6
8
10
12
14
16
18
20
C2/500
0
2
4
6
8
10
12
14
16
FV1/640LR9/300
4 4 4 4
8 8 8 88
12 1212 1216 16
16
00 0 00
LR2/68 C2/500 FV1/640
0
5
10
15
20
25
LR10/12
0
5
10
15
20
25
FV1/440
0
2
4
6
8
10
12
EH1/155
0
2
4
6
8
10
12
14
16
EH2/160
0
2
4
6
8
10
12
14
16
18
20
LR9/50
0
2
4
6
8
10
12
14
LR2/48
0
5
10
15
20
25
C2/350
4 4 4
8 810 10 108
8
1212
121616
0 00 0
20 20 20
0 00
0
2
4
6
8
10
12
14
FV1/240
0
2
4
6
8
10
12
14
16
18
EH2/155
0
2
4
6
8
10
12
14
EH1/105
0
2
4
6
8
10
12
14
16
18
LR2/28
0
2
4
6
8
10
12
14
16
C2/10
4 4
8 888 8
12 12 161616
0 0 00 0
LR2/28 C2/10 EH2/155
0
2
4
6
8
10
12
14
16
FV1/40
0
2
4
6
8
10
12
14
EH1/85
44
88
121216
00
FV1/40 EH1/85
0
2
4
6
8
10
12
EH2/75
0
5
10
15
20
25
EH1/30
4
810
12
0 0
20EH1/30 EH2/75
0
2
4
6
8
10
12
EH2/50
4
8
12
0
EH2/50
0
2
4
6
8
10
12
14
EH2/20
48
12
0
EH2/20
0
2
4
6
8
10
12
14
LIP2
4
812
01.0 3.0
LIP2
LR10/600
LR10/550 LR9/350 LR2/98
LR10/350
LR10/12 LR9/50 LR2/48 C2/350 FV1/440 EH1/155 EH2/160
FV1/240
2.01.0 1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
3.0 3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
2.0 2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
EH1/105
Fig. 7. Variability of the central opening size for the individual assemblages, Egerian to Eggenburgian. Vertical axis, number of specimens; horizontal axis,central opening size (in μm). Location of the boreholes LR10, LR9, LR2, C2, FV1, EH1, EH2, and section Lipovany (LIP) is illustrated on Fig. 1, their li−thology and sampled intervals on Fig. 2. Abbreviation: LO, last occurrence.
660 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
Sp
he
no
lith
us
he
tero
mo
rph
us
Sp
he
no
lith
us
he
tero
mo
rph
us
He
lico
sp
ha
era
am
plia
pe
rta
La
ng
hia
nS
erra
valli
an
Ott
na
ng
ian
Ka
rpa
tian
Ba
de
nia
n
NN4
NN5
14
15
16
17
Mio
cen
e
Bu
rdig
alia
n
Ag
e(M
a)
Ep
och
Sta
ge
Ce
ntr
al
Pa
rate
thys
Sta
ge
Ca
lca
reo
us
Na
nn
op
lan
kto
nZ
on
e
Ce
ntr
al
Pa
rate
thys
Ca
lca
reo
us
Na
nn
op
lan
kto
nE
ven
ts
Variability of length of central opening (μm)
0
2
4
6
8
10
12
LIP7
0
2
4
6
8
10
12
14
LIP11
0
2
4
6
8
10
12
14
16
18
20
D19/665
0
1
2
3
4
5
6
7
8
9
10
D19/426
0
2
4
6
8
10
12
14
LKŠ1/340
0
2
4
6
8
10
12
LKŠ1/238
0
2
4
6
8
10
12
14
LKŠ1/224
0
2
4
6
8
10
12
14
16
LKŠ1/181
0
1
2
3
4
5
6
7
8
9
10
LKŠ1/105
0
1
2
3
4
5
6
7
8
9
10
LKŠ1/26
0
2
4
6
8
10
12
14
BE2/333
0
2
4
6
8
10
12
14
BE2/262
0
2
4
6
8
10
12
14
BE2/114
0
5
10
15
20
25
PY1/2.2
0
5
10
15
20
25
30
35
40
45
PY1/1.5
0
5
10
15
20
25
30
35
40
PY1/0.4
0
5
10
15
20
25
30
35
40
KRAS4
0
5
10
15
20
25
30
KRAS7
0
5
10
15
20
25
30
KRAS90
5
10
15
20
25
KRAS10
BE2/262
KRAS4
KRAS9
KRAS10
PY1/2.2
PY1/0.4
LKŠ1/105
LKŠ1/26
4 44
4
4 4
4
4
4
10
10
10
10
10
4
2
88
8
8 8
8
8
8
10
20
2020
20
20
10
8
4
4
4
121212
1212
12 12
12
12
30
30
30
12
6
16
8
8
8
20
00
0
0
0
20
40
40
40
20
0
0
0
0 0
0
0
0
LIP7
0
2
4
6
8
10
12
14
LIP5
48
12
0
LIP5
LIP11 LKŠ1/340D19/665
LKŠ1/238 BE2/333
LKŠ1/181
KRAS7PY1/1.5
D19/426
BE2/114
0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
1.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
2.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
3.0
LKŠ1/224
Fig. 8. Variability of the central opening size for the individual assemblages, Ottnangian to Early Badenian. Vertical axis, number of specimens; horizontalaxis, central opening size (in μm). Location of the boreholes D19, LKŠ1, BE2, PY1, and sections Lipovany (LIP) and Kralice (KRAS) is illustrated onFig. 1, their lithology and sampled intervals on Fig. 2.
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 661
0
50
100
150
200
250 n=350
0
10
20
30
40
50
60
70 n=500
0
5
10
15
20
25
30
35
40
45
50 n=200
0
10
20
30
40
50
60 n=250
0
5
10
15
20
25
30
35
40 n=200
0
10
20
30
40
50
60
n=250
0
10
20
30
40
50
60 n=350
0
10
20
30
40
50
60 n=350
0
10
20
30
40
50
60
70
80
90
100 n=350
0
20
40
60
80
100
120
140 n=450
0
10
20
30
40
50
60 n=350
0
10
20
30
40
50
60 n=350
0
10
20
30
40
50
60
70
80 n=500
0
20
40
60
80
100
120
140
160
180 n=350
2 3 4 5 6 7 8 9 10 0.5 1.0 1.5 2.0 2.5 3.0
Lenght of central opening(μm)
Lenght of nannolith(μm)
transgression
R. minuta
MMCO
n=200
n=250
n=350 n=450
n=350
n=350
n=200
n=500
n=350
0
10
20
30
40
50
60
70
80
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=450
0
10
20
30
40
50
60
70
80
90
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=350
0
10
20
30
40
50
60
70
80
90
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=250
0
10
20
30
40
50
60
70
80
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=200
0
10
20
30
40
50
60
70
80
90
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=500
0
10
20
30
40
50
60
70
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=350
0
10
20
30
40
50
60
70
80
90
0.16 0.20 0.24 0.28 0.32 0.36 0.40 0.44
n=350
n=450
n=350
n=350
n=250 n=250
n=200
n=500
n=350
Lenght of central opening/lenght of nannolith
Sphenolithusheteromorphus
Sphenolithusheteromorphus
Sphenolithusheteromorphus
Sphenolithusheteromorphus
Sphenolithusbelemnos
Sphenolithusbelemnos
Sphenolithusbelemnos
Helicosphaeraampliaperta
Helicosphaeraampliaperta
Helicosphaeraampliaperta
Helicosphaeraampliaperta
Discoasterdruggi
Discoasterdruggi/Helicosphaerascissura
Dictylcoccitesbisectus
LCODictylcoccitesbisectus
Olig
oce
ne
Mio
cen
e
Ch
att
ian
Aq
uita
nia
nB
urd
iga
lian
La
ng
hia
nS
err
ava
llia
n
Eg
eria
nE
gg
en
bu
rgia
nO
ttn
an
gia
nK
arp
atia
nB
ad
en
ian
NP25
NN1
NN2
NN3
NN4
NN5
NN6
Ag
e(M
a)
Ep
och
Sta
ge
Ce
ntr
al P
ara
teth
ysS
tag
eC
alc
are
ou
sN
an
no
pla
nkt
on
Zo
ne
Ca
lca
reo
us
Na
nn
op
lan
kto
nE
vents
Ce
ntr
al P
ara
teth
ysC
alc
are
ou
sN
an
no
pla
nkt
on
Eve
nts
13
14
15
16
17
18
19
20
21
22
23
24
25
Helicosphaeracarteri
Size variability of Reticulofenestra minutaReticulofenestra haqii–pseudoumbilicus
andgroup
n=350
0.20 0.30 0.40
R. haqii–pseudoumbilicus group
10080
60
40
20
0
10080604020
0
120
60
40
20
0
80
60
40
20
0
80
60
40
20
0
80
60
40
20
0
80
60
40
20
0
80
60
40
20
0
8060
40
20
0
80
60
40
20
0
60
40
20
0
60
40
20
0
60
40
20
0
Mi1 eventn=350
transgression
n=500
transgression
transgression40
20
0
60
40
20
0
60
40
20
0
60
40
20
0
40
20
0
Mi1b event (16.1Ma)
60
40
20
0
n=350160
80
40
0
120200
100
50
0
150
250
0.20 0.30 0.40
0.20 0.30 0.40
0.20 0.30 0.40
0.20 0.30 0.40
0.20 0.30 0.40
0.20 0.30 0.40
0.5 1.0 1.5 2.0 2.5 3.0
0.5 1.0 1.5 2.0 2.5 3.0
0.5 1.0 1.5 2.0 2.5 3.0
0.5 1.0 1.5 2.0 2.5 3.0
0.5 1.0 1.5 2.0 2.5 3.0
0.5 1.0 1.5 2.0 2.5 3.0
2 3 4 5 6 7 8 9 10
2 3 4 5 6 7 8 9 10
2 3 4 5 6 7 8 9 10
2 3 4 5 6 7 8 9 10
2 3 4 5 6 7 8 9 10
2 3 4 5 6 7 8 9 10
2 3 4 5 6 7 8 9 10
Mi1a event
Fig. 9. Morphometric changes of placoliths of Reticulofenestra minuta and Reticulofenestra haqii–pseudoumbilicus group from the Egerian to the EarlyBadenian (Burdigalian–Langhian) in the Central Paratethys. Cooling Mi events according Zachos et al. 2001. MMCO = the Middle Miocene Climatic Opti−mum (Zachos et al. 2001). Abbreviation: LO, last occurrence.
In conclusion, a satisfactory size has not been found uponwhich to distinguish species in the R. haqii–pseudoumbilicusgroup and its taxonomy is simply a matter of convention.
Size changes in the Late Oligocene–EarlyMiocene Reticulofenestra and paleoecologic andpaleogeographic events in the Central Paratethys
Blooms of Reticulofenestra minuta.—Two blooms of thesmall R. minuta were observed in the studied material (Figs. 9,11): (i) the oldest one in the earliest Miocene (Biochron NN1and early part of Biochron NN2), is described for the first time.It is probably a local event in the northern part of the Panno−nian Basin; (ii) the early Middle Miocene (NN5) event was al−ready documented; it occurred throughout the Central Para−tethys (Ćorić and Švábenická 2004; Ćorić and Hohenegger2008, Spezzaferi et al. 2009). In addition to these two wide−spread blooms, sporadical blooms of R. minuta have also beendescribed in the Central Paratethys, having occurred duringthe Eggenburgian (Holcová 2002, 2005) and the Karpatian(Spezzaferi and Ćorić 2001; Švábenická et al. 2003).
Besides the Central Paratethys, blooms of R. minuta havebeen described from several stratigraphical levels in the Mio−cene from the middle to high latitudes (e.g., Cita et al. 1978;Backman 1980). These blooms are heterochronous with theCentral Paratethys ones and reflect local paleoenvironmentalevents (e.g., Messinian salinity crisis; Cita et al. 1978; Wadeand Bown 2006).
Interpretation of the causes of the blooms is ambiguousdue to inconsistent data about the ecological requirements ofsmall Reticulofenestra. Generally, the species is opportunis−tic and characterizes assemblages from near−shore environ−ment (Haq 1980). Their blooms are connected with environ−mental stress characterized by rapid environmental changes(Wade and Bown 2006).
Gartner et al. (1983) suggested that variations in the relativeabundance of R. minuta correlate with changes in nutrient dy−namics. Wells and Okada (1997), Flores et al. (1997), Boll−mann et al. (1998) and Kameo (2002) regard small Reticulo−fenestra spp. as eutrophic species while Hallock (1987), Beau−fort and Aubry (1992), Ćorić and Rögl (2004) suggested thatblooms of small Reticulofenestra indicate oligotrophic warmwater. Wade and Bown (2006) showed that R. minuta mayhave tolerated the brackish to hypersaline, high productivityenvironments, which prevailed immediately before and afterthe evaporite deposition during the Messinian salinity crisis.
The two conspicuous blooms of small R. minuta in theCentral Paratethys occurred during transgression events whenthe Slovenian corridor permitted incursions of warm waterinto the Paratethys (Rögl 1998, 1999; Popov et al. 2004).These events are marked by horizons with larger Foramin−ifera—Miogypsina and Lepidocyclina in the lowermost Mio−cene (Váňová 1975), Amphistegina and Planostegina in theLower Badenian—and replacement of small Globigerinafauna by diversified assemblages with large−sized warm−
water species—Globigerinoides in the Lower Miocene andGlobigerinoides, Praeorbulina, and Orbulina in the MiddleMiocene). At the beginning of the blooms, R. minuta replacesCoccolithus pelagicus, which is an indicator of cold and nutri−ent−rich waters (McIntyre and Be 1967; Rahman and Roth1990). Among the benthic foraminifera, the abundance of ag−glutinated foraminifera decreases (Vass et al. 1983; Holcová2001; Spezzaferi et al. 2009). The paleoenvironmental turn−over correlatable with the Middle Miocene bloom of R. minutahas been well explained: Upwelling regime with cooler sur−face waters and partly dysoxic bottom conditions of the latestEarly Miocene were gradually changed to the warm−watercondition in the early Middle Miocene and antiestuarine circu−lation with evaporation prevailing over fresh−water input(Brzobohatý 1987; Ćorić and Rögl 2004; Báldi 2006; Spezza−ferri et al. 2009). Comparable circulation turnover due to mix−ing of in situ cold and incoming warm water masses andpaleogeographical changes may be expected also in the earli−est Miocene in the South Slovak Basin (Halásová et al. 1996).
Although the incoming of warm−water elements in the ma−rine realm is specific of the two transgressive events, the Earlyand Middle Miocene conditions in the Central Paratethys sub−stantially differed from the global paleoclimatic situation. TheEarly Miocene event was marked with the appeareance ofarctotertiary elements among palynomorphs (Vass et al. 1988;Planderová 1990) indicative of a global cooling correlatedwith the Mi1 and Mi1a events. In contrast, the Middle Mio−cene event can be correlated with the Middle Miocene Clima−tic Optimum (Gonera et al. 2000; Bicchi et al. 2003; Böhme2003). Short−term oscillations in the abundance of R. minutaduring the two blooms were observed (Ćorić and Hohenegger2008; own unpublished data). These may indicate unstable
662 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
Le
ng
tho
fce
ntr
al o
pe
nin
g(μ
m)
4 5 6 7 830.5
1.0
1.5
2.0
2.5
3.0
Length of nannolith (μm)
Winter morphotype Summer morphotype
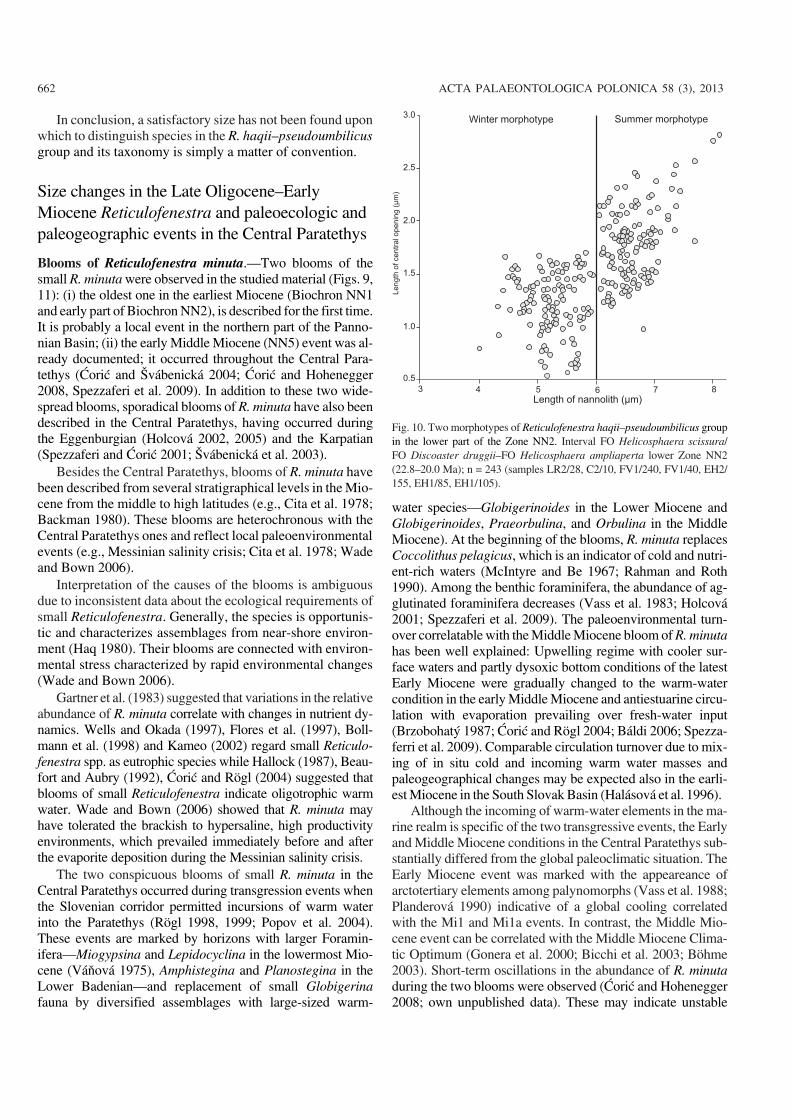
Fig. 10. Two morphotypes of Reticulofenestra haqii–pseudoumbilicus groupin the lower part of the Zone NN2. Interval FO Helicosphaera scissura/FO Discoaster druggii–FO Helicosphaera ampliaperta lower Zone NN2(22.8–20.0 Ma); n = 243 (samples LR2/28, C2/10, FV1/240, FV1/40, EH2/155, EH1/85, EH1/105).

conditions with short−term oscillations of paleoenvironmentalparameters mainly salinity and nutrients. These oscillationshave been interpreted in the Biochron NN5 by Hohenegger etal. (2008): warm intervals characterized by high terrigenousinput and higher seasonality were alternated with cold periodswith lower terrigenous input and lower seasonality. Reticule−
fonestram minuta strongly dominated in the near−shore faciesduring warm intervals (more than 80%) where high terri−genous input during wet seasons may cause seasonal oscilla−tions of salinity in the upper layer of water column (own un−published data).
The expansion of near−shore facies as a result of transgres−
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 663
Ce
ntr
al
Pa
rate
thys
pa
leo
ge
og
rap
hic
al e
ven
ts(R
ög
l 19
98
,1
99
9,
Po
po
ve
ta
l.2
00
4,
Pill
er
eta
l.2
00
7)
Ce
ntr
al P
ara
teth
ysst
ag
es
Eg
ge
nb
urg
ian
Eg
eria
nK
arp
atia
nB
aden
ian
1
2
3
6
5
Inte
rva
ls(t
his
wo
rk)
Ott
na
ng
ian
Re
ticu
lofe
ne
strid
sb
iom
etr
ica
l eve
nts
(th
isw
ork
)
Glo
ba
lse
a-le
velc
ycle
s(H
ard
en
bo
leta
l.1
99
8)
Bur 2
Bur 3
Bur 4
Bur 5/Lan1
19.1
Clim
atic
cha
ng
es
(Kü
rsch
ne
re
ta
l.2
00
8;
Teo
do
rid
ise
ta
l.2
00
7)
20
21
22
23
24
25
19
Ag
e(M
a)
ap
pro
xim
ativ
ely
18
17
16
15
FO of H. ampliaperta
FO of S. belemnos
S. heteromorphusFO of
FO of H. carteri
FO ofFO
D. druggii/H. scissura
LCO of R. bisecta
Sloveniancorridor
Slovenian corridor
Sloveniancorridor
R.m
inu
tab
loo
m
R.minutabloom
slig
htin
cre
ase
ofn
an
no
lith
size
bim
od
als
ize
dis
trib
utio
no
fp
laco
lith
s?
win
ter
an
dsu
mm
er
mo
rph
oty
pe
s
FO of R. haqii
Mi1a
Mi1
Mi1b
Mi2
4
Ch 3
Ch 4/Aq 1
Aq 2
Aq 3/Bur 1
Ch
att
ian
Aq
uita
nia
nB
urd
iga
lian
La
ng
hia
nS
tag
e
Ca
lca
reo
us
na
nn
op
lan
kto
nzo
ne
s
NP25
NN1
NN2
NN3
NN4
NN5
23.1
22.8
~ 25
Use
dca
lca
reo
us
na
nn
op
lan
kto
nb
iost
ratig
rap
hic
al
eve
nts
17.7
caesing ofthe marine
pathwayvia the Alpine
Foredeep andthe Rhine Graben
succesiveopening of
the pathwayvia the Alpine
Foredeep
broadconnection
to theVenetian
Basin,
via the RhineGraben,
the AlpineForedeepand
Ser 1
non-studiedinterval
MMCO
bro
ad
size
an
dsh
ap
eva
ria
bili
tyo
fn
an
no
lith
s
FCO of>7 μmR. pseudoumbilicus
FO of
with smallcentral opening
R. pseudoumbilicus6–7 μm
isolation
20.5
15.97 FO of P. circularis
local transgression
transgression
transgression
transgression
transgression
14.9 LO of H. ampliaperta
FCO of
>8.0 μmR. pseudoumbilicus
transgression
Fig. 11. Correlations between reticulofenestrid biometric events, local and global sea−level changes and paleogeographical events in the Egerian–Early Badenian(Burdigalian–Langhian) of the Central Paratethys. Abbreviations: FCO, first common occurrence; FO, first occurrence; LCO, last common occurrence; LO, lastoccurrenceFCO; MMCO, the Middle Miocene Climatic Optimum; D., Discoaster; H., Helicosphaera; P., Praeorbulina; R., Reticulofenestra; S., Sphenolithus.
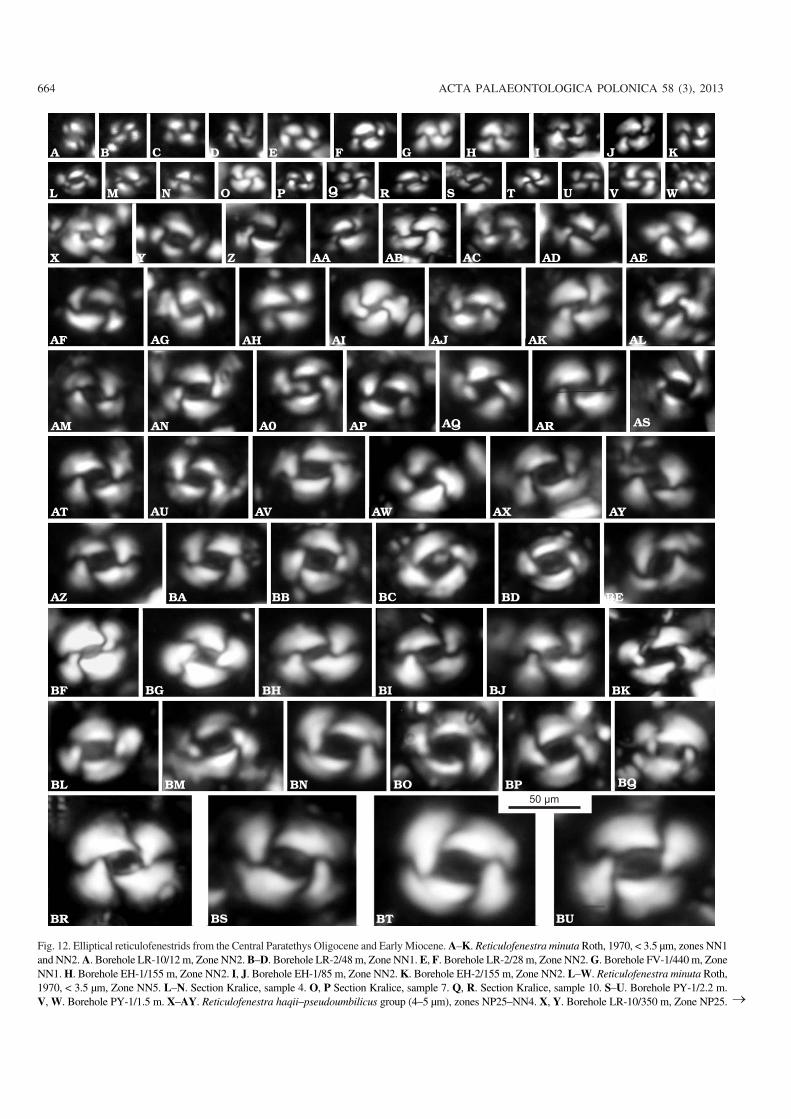
664 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013
A
L
X
AF AG AH AI AJ AK AL
AM AN
AZ BA BB BC BD BE
BF BG BH BI BJ BK
BL
BR BS BT BU
BM BN BO BP BQ
AW AX AY
AP AQ AR AS
Y Z AA AB AC AD AE
M N O P Q R T
B C D E F HG
S U V W
I J K
A0
AT AU AV
50 µm
Fig. 12. Elliptical reticulofenestrids from the Central Paratethys Oligocene and Early Miocene. A–K. Reticulofenestra minuta Roth, 1970, < 3.5 μm, zones NN1and NN2. A. Borehole LR−10/12 m, Zone NN2. B–D. Borehole LR−2/48 m, Zone NN1. E, F. Borehole LR−2/28 m, Zone NN2. G. Borehole FV−1/440 m, ZoneNN1. H. Borehole EH−1/155 m, Zone NN2. I, J. Borehole EH−1/85 m, Zone NN2. K. Borehole EH−2/155 m, Zone NN2. L–W. Reticulofenestra minuta Roth,1970, < 3.5 μm, Zone NN5. L–N. Section Kralice, sample 4. O, P Section Kralice, sample 7. Q, R. Section Kralice, sample 10. S–U. Borehole PY−1/2.2 m.V, W. Borehole PY−1/1.5 m. X–AY. Reticulofenestra haqii–pseudoumbilicus group (4–5 μm), zones NP25–NN4. X, Y. Borehole LR−10/350 m, Zone NP25. �

sion, the changes in water circulation due to the mixing of in−coming warmer and in situ colder water masses and, probably,unstable conditions as a result of short term (?seasonal) oscil−lations of ecological parameters (such as salinity in the upperlayers of water column) may have produced conditions fa−vourable to the blooms of small−sized Reticulofenestra.
Size changes of the Reticulofenestra haqii–pseudoumbi−licus group.— Discrepancies between the FOs of individualsize categories in the R. haqii–pseudoumbilicus group (seechapter: Size changes over time) in the Central Paratethysand in the oceanic realm is notable for interpretation of localvs. global character of size changes in the group. The follow−ing discrepancies have been recorded:
(i) The FO of R. haqii (> 4 μm) was observed in the stud−ied area in Zone NP25 (Holcová 2005) whereas in the worldocean it lies at the NN1/NN2 boundary (Young 1999).
(ii) The FO of R. pseudoumbilicus (> 5 μm) is given in ZoneNN4 in the world ocean (Young 1999). In the Central Para−tethys, Marunteanu (1999) and Chira (2004) reported this spe−cies from near the NN1/NN2 boundary, Andreyeva−Grigo−rovich et al. (2008) from Zone NN2 and Molčíková and Stra−ník (1987) from Zone NN1. Holcová (2005, this paper) de−scribe the FO of specimens > 5 μm together with FO of R. haqii> 4 μm in the uppermost part of the NP25 Zone (Figs. 5, 6, 9).
(iii) The FO of R. pseudoumbilicus > 7 μm is given mostoften being a Middle Miocene event. In the Mediterranean, theFO has been described from Zone NN6 (Fornaciari and Rio1996), and the FCO of R. pseudoumbilicus > 7.0 μm to definethe z ones MNN6a and MNN6b (Fornaciari et al. 1996); in theEastern equatorial Pacific the event was recorded near theNN5/NN6 boundary (Raffi and Flores 1995). However, Howeand Sblendorio−Levy (1998) described the FO of R. pseudo−umbilicus > 7 μm from the upper part of Zone NN 2 in the At−lantic Ocean near Madeira. This correlates with its FO in theCentral Paratethys in Zone NN2 (Molčíková and Straník1987; this paper). The FCO of R. pseudoumbilicus > 7 μm inthe Central Paratethys has been recorded from the NN4 Zone.(Molčíková 1983; Švábenická et al. 2003; Eva Halásová, per−sonal communication 2009; this paper).
The Central Paratehys FOs of all three size categories in R.haqii–pseudoumbilicus group occurred earlier in the globalocean (including the Mediterranean realm) and cannot be usedfor interregional biostratigraphic correlations.
The evolution of size in the R. haqii–pseudoumbilicusgroup in the Central Paratethys would suggest an earlier ap−
pearance of larger specimens owing to specific conditions inan intracontinental basin. However, the most important mor−phological changes in the R haqii–pseudoumbilicus group canbe correlated with the opening of new pathways between theCentral Paratethys and the Mediterranean and the immigrationof incoming species in the Central Paratethys (Fig. 11). First,the FO of R. haqii is associated with the reopening of sea−waysafter the Kiscellian anoxia, which is marked by the immigra−tion of foraminifera (Holcová 2008). Second at the upper partof Zone NN2 marked by the FO of H. ampliaperta, largerspecimens of R. haqii (6–7 μm) with small central opening(1.5–1.7 μm) were recorded. The FO of H. ampliaperta can becorrelated with transgression from the Mediterranean via thePre−Alpine passage (Rögl 1998; Popov et al. 2004). The trans−gression is characterized by substantial faunal turnovermarked by numerous first occurrences of species of westernMediterranean origin (Kroh and Harzhauser 1999; Vávra1979; Mandic and Steininger 2003; Piller et al. 2007). Third,the FCO of R. pseudoumbilicus > 7 μm at the base of ZoneNN4 correlates with the transgression and paleogeographic re−organization and the establishment of a new broad connectionvia the “Trans−Tethyan trench corridor”. The connection en−abled faunal exchange between the Mediterranean and Para−tethys (Rögl 1998; Rögl et al. 2003; Popov et al. 2004).Fourth, the FO of R. pseudoumbilicus > 8 μm is associatedwith the reopening of the Mediterranean–Indo−Pacific sea−way (Rögl and Steininger 1983; Rögl 1998, 1999; Popov et al.2004). This large transgression caused rapid immigration ofthe marine fauna (e.g., Harzhauser and Piller 2007; Holcová2008) termed the EBBE, or Early Badenian−Build−up−Event(Harzhauser and Piller 2007).
Slight gradual size changes in the R. haqii–pseudoumbi−licus group were recorded during periods without paleogeo−graphical changes (Fig. 11). The decrease in the placoliths sizein the late Egerian (upper NP25 to NN1 zonal interval) maycorrespond with cooling interpreted from occurrence of arcto−tertiary elements among palynomorphs (Planderová 1990).Also Kürschner et al. (2008) hypothesized a cooling duringthis interval correlatable with the Mi1/1a event.
The size of placoliths slightly increased in the Eggen−burgian, although this increase is not general (Figs. 5, 6, 9).This size increase can be correlated with warming (Kürschneret al. 2008).
Bimodal size distribution in the R. haqii–pseudoumbi−licus group was recorded firstly during the Zone NN1 and inthe lower part of the Zone NN2 (Fig. 9, 10). The smaller
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 665
Z, AA. Borehole LR−9/350 m, Zone NP25. AB. Borehole LR−2/98 m, Zone NP25. AC–AE. Borehole C−2/500 m, Zone NP25. AF, AG. Borehole EH2/160 m,Zone NN1. AH, AI. Borehole EH−1/105 m, Zone NN2. AJ, AK. Borehole EH−1/85 m, Zone NN2. AL. Borehole EH−2/75 m, Zone NN2. AM, AN. BoreholeLKŠ−1/340 m, Zone NN3. AO–Q. Borehole LKŠ−1/181 m, Zone NN4. AR, AS. Borehole LKŠ−1/105 m, Zone NN4. AT–BE Reticulofenestra haqii–pseudoumbilicus group (5–7 μm), zones NP25–NN4. AT, AU. Borehole LR−10/350 m, Zone NP25. AV. Borehole LR−9/350 m, Zone NP25. AW, AX. Bore−hole C−2/500 m, Zone NP25. AY. Borehole EH2/160 m, Zone NN1. AZ, BA. Borehole EH−1/105 m, Zone NN2. BB, BC. Borehole EH−1/85 m, Zone NN2.BD, BE. Borehole EH−2/155 m, Zone NN2. BF–BK. Reticulofenestra haqii–pseudoumbilicus group (6–7 m) narrow central opening, Zone NN2. BF, BG.Borehole EH−1/155 m, Zone NN2. BH, BI. Borehole EH−1/105 m, Zone NN2. BJ, BK. Borehole EH−2/85m, Zone NN2. BL–BQ. Reticulofenestrahaqii–pseudoumbilicus group (6–8 μm), Zone NN4. BL, BM. Borehole LKŠ−1/105 m, Zone NN4. BN, BO. Borehole LKŠ−1/26 m, Zone NN4. BP, BQ Bore−hole BE−2/114 m, Zone NN4. BR, BU. Reticulofenestra haqii–pseudoumbilicus group (> 8 μm), Zone NN5. BR. Borehole PY−1/2.2 m. BS. BoreholePY−1/1.5 m. BT, BU. Section Kralice, sample 4.

morphotype (< 6 μm) is characterized by a small centralopening 0.5–1.5 μm. Larger morphotype (> 6 μm) havelarger central openings (Fig. 10).
Backman (1980) and Beaufort and Aubry (1992) de−scribed a dependence of the morphometric variability of R.pseudoumbilicus on seasonal or latitudinal temperature dif−ferences during the Miocene and Pliocene. According toBackman (1980) low latitude individuals are characterizedby a larger central opening and can be slightly larger. Highlatitude individuals are distinguished by a smaller centralopening. Beaufort and Aubry (1992) have proposed alterna−tions of winter and summer morphotypes in the Lower Mio−cene of high−latitude populations. The occurrence of two sizecategories (3.5–6 μm and 6–8 μm) in the late Egerian–earlyEggenburgian of analysed samples may also represent sea−sonal morphotypes: the smaller morphotype could representthe winter morphotype while the larger morphotype with alarger central opening could represent the summer morpho−type (Fig. 10) This assumed seasonality has been corrobo−rated by palaeoclimate estimates based on plant macrofossilspreserved in the Břešťany Clay from the Most Basin (Teo−doridis and Kvaček 2006; Kvaček and Teodoridis 2007).These studies have shown a significant difference between aColdest Month Mean Temperature (CMMT) and WarmestMonth Mean Temperature (WMMT), which is 15–20�C(Teodoridis 2007; Teodoridis et al. 2011).
ConclusionsBased on biometric analysis, two size groups of ellipticalreticulofenestrids from the NP25–NN5 zonal interval weredistinguished. One group corresponds to the small R. minuta(< 3 μm); the other is the R. haqiii–pseudoumbilicus group(3.5–10 μm). A clear criterion upon which to define species inthis group has not been found, and species taxonomy (R. haqiivs. R. pseudoumbilicus) can only be conventional (Fig. 12).
Two blooms of small R minuta were observed. The earliestMiocene bloom (NN1 and lower part of Zone NN2) is proba−bly a local event for the northern part of the Pannonian Basin.The early Middle Miocene bloom (Zone NN5) was alreadyknown and has a global occurrence in the Central Paratethysbasins. These blooms can be correlated with the expansion ofnear−shore facies associated with transgression, the incursionsof warm water into the Paratethys, which may cause changesin water circulation due to the mixing of warmer and colderwater masses. Unstable conditions with short−term oscillationsof ecological parameters (probable salinity in the upper layersof water column) can be also expected.
The FOs of all three size categories in R. haqii–pseudo−umbilicus group (3–5 μm, 5–7 μm, > 7 μm) occurred earlierin the Central Paratehys than in the global ocean and theMediterranean region. They cannot be used for interregionalbiostratigraphical correlations.
The slight gradual changes in size in the R. haqii–pseudo−umbilicus group can be correlated with paleoclimatic trends.
The decrease in size of the placoliths in the late Egerian (up−per Zone NP25 and Zone NN1) may result from cooling dur−ing the Mi1/1a event.
The size of placoliths slightly increased from the Eggen−burgian to the Badenian; the interval can be generally charac−terized by warming.
The two morphotypes occurring in the late Egerian–Early Eggenburgian may represent seasonal morphotypes.The smaller morphotype could represent the winter morpho−type while the larger morphotype with a slightly larger cen−tral opening could represent the summer morphotype.
AcknowledgementsI would like to acknowledge the helpful discussion on Oligocene andMiocene paleoclimate by Zlatko Kvaček and Vasilis Teodoridis (bothCharles University in Prague, Czech Republic). I thank the Marie−PierreAubry (Rutgers University, New Jersey, USA), and Jörg Mutterlose(Ruhr−Universität Bochum, Germany) for their comments and correc−tions, which substantially improved the quality of this manuscript. Thisresearch was supported by grants no. MSM0021620855 and GAČR205/09/0103.
ReferencesAndreyeva−Grigorovich, A.S., Kulchytsky, Y.O., Gruzman, A.D., Lozonyak,
P.Y., Petrashkevich, M.I., Portnyagina, L.O., Ivanina, A.V., Smirnov,S.E., Trofimovich, N.A., and Savitskaya, N.A. 1997. Regional strati−graphic scheme of Neogene formations of the Central Paratethys ofUkraine. Geologica Carpathica 48: 123–136.
Andreyeva−Grigorovich, A.S., Oszczypko, N., Slaczka, A., Oszczypko−Clowes, M., Savitskaya, N.A., and Trofimovich, N.A. 2008. New dataon the stratigraphy of the folded Miocene Zone at the front of the Ukrai−nian Outer Carpathians. Acta Geologica Polonica 58: 325–354.
Backman, J. 1978. Late Miocene–Early Pliocene nannofossil biochronologyand biogeography in the Vera Basin, SE Spain. Stockholm Contributionsin Geology 32: 93–114.
Backman, J. 1980. Miocene–Pliocene nannofossils and sedimentation ratesin the Hatton−Rockall Basin, NE Atlantic Ocean. Stockholm Contribu−tions in Geology 36: 1–91.
Báldi, K. 2006. Paleoceanography and climate of the Badenian (Middle Mio−cene, 16.4–13.0 Ma) in the Central Paratethys based on foraminifera andstable isotope (�18O and �13C) evidence. International Journal of EarthScience (Geologische Rundschau) 95: 119–142.
Báldi T. 1986. Mid−tertiary Stratigraphy and Paleogeographic Evolution ofHungary. 201 pp. Akademiai Kiado, Budapest.
Beaufort, L. and Aubry, M.−P. 1992. Palaeoceanographic implications of a17 m.y. long record of high−latitude Miocene calcareous nannoplanktonfluctuations. Proceeding of Ocean Drilling Program, Scientific Results120: 530–549.
Berggren, W.A., Kent, D.V., Swisher III., C.C., and Aubry, M.−P. 1995.A revised cenozoic geochronology and chronostratigraphy. In: W.A.Berggren, D.V. Kent, and J. Hardenbol (eds.), Geochronology, TimeScale and Global Stratigraphic Correlations: A Unified TemporalFramework for an Historical Geology. Society of Economic Paleontol−ogists and Mineralogists, Special Publication 54: 129–212.
Bicchi, E., Ferrero, E., and Gonera, M. 2003. Palaeoclimatic interpretationbased on Middle Miocene planktonic Foraminifera: the Silesia Basin(Paratethys) and Monferrato (Tethys) records. Palaeogeography, Palaeo−climatology, Palaeoecology 196: 265–303.
Böhme, M. 2003. The miocene climatic optimum: evidence from ectothermic
666 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013

vertebrates of Central Europe. Palaeogeography, Palaeoclimatology,Palaeoecology 195: 389–401.
Bollmann, J., Baumann, K.H., and Thierstein, H.R. 1998. Global domi−nance of Gephyrocapsa coccoliths in the late Pleistocene: Selectivedissolution, evolution, or global environmental change? Paleoceano−graphy 13: 517–529.
Brzobohatý, R. 1987. Contribution to paleogeography of the Miocene bas−ins of the Central Paratethys from otolith fauna. Miscellanea Micro−palaeontologica II/2, Knihovnička zemního plynu a nafty 6b: 101–111.
Chira, C. 2004. Early Miocene calcareous nannofossils assemblages fromTransylvania. Acta Paleontologica Romaniae 4: 81–88.
Chlupáč, I., Brzobohatý, R., Kovanda, J., and Stráník, Z. 2002. Geologickáminulost České republiky. 436 pp. Academia, Praha.
Cita, M.B., Wright, R.C., Ryan, W.B.F., and Longinelli, A. 1978. Messinianpaleoenvironments, Initial Reports of the Deep Sea Drilling Project42A: 1003–1035.
Ćorić, S. and Hohenegger, J. 2008. Quantitative analyses ofcalcareous nanno−plankton assemblages from the Baden−Sooss section (Middle Miocene ofVienna Basin, Austria). Geologica Carpathica 59: 447–460.
Ćorić, S. and Rögl, F. 2004. Roggendorf−1 borehole, a key−section for LowerBadenian transgressions and the stratigraphic positin of the Grund Forma−tion (Molasse Basin, Lower Austria). Geologica Carpathica 55: 165–178.
Ćorić, S. and Švábenická, L. 2004. Calcareous nannofossil biostratigraphyof the Grund Formation (Molasse Basin, Lower Austria). GeologicaCarpathica 55: 147–155.
Flores, J.A., Sierro, F.S., Francés, G., Vasquez, A., and Zamarreno, I. 1997.The last 100,000 years in the western Mediterranean: Sea surface waterand frontal dynamics as revealed by coccolithophores. Marine Micro−paleontology 29: 351–366.
Fornaciari, E. and Rio, D. 1996. Latest Oligocene to early middle Miocenequantitative calcareous nannofossil biostratigraphy in the Mediterra−nean region. Micropaleontology 42: 1–36.
Fornaciari, E., Di Stefano, A., Rio, D., and Negri, A. 1996. Middle Miocenequantitative calcareous nannofossil biostratigraphy in the Mediterra−nean region. Micropaleontology 42: 37–63.
Gartner, S., Chen, M.P., and Stanton, R.J. 1983. Late Neogene nanofossilbiostratigraphy and paleoceanography of the northeastern Gulf of mex−ico and adjacent areas. Marine Micropaleontology 8: 17–50.
Gonera, M., Peryt, T.M., and Durakiewicz, T. 2000. Biostratigraphical andpaleoenvironmental implications of isotopic studies (18O, 13C) of Mid−dle Miocene (Badenian) foraminifers in the Central Paratethys. TerraNova 12: 231–238.
Halásová, E., Hudáčková , N., Holcová, K., Vass, D., Elečko, M., and Pere−szlényi, M. 1996. Sea ways connecting the Fiľakovo−Pétervasara Basinwith the Eggenburgian/Burdigalian open sea. Slovak Geological Maga−zine 2: 125–136.
Hallock, P. 1987. Fluctuations in the trophic resource continuum: a factor inglobal diversity cycles? Paleooceanography 2: 457–471.
Haq, B.U. 1980. Biogeographic history of the Miocene calcareous nanno−plankton and paleoceanography of the Atlantic Ocean. Micropaleonto−logy 26: 414–443.
Haq, B.U., Hardenbol, J., and Vail, P.R. 1988. Mesozoic and Cenozoicchronostratigraphy and cycles of sea−level change. Society of EconomicPaleontologists and Mineralogists, Special Publication 42: 71–108.
Harzhauser, M. and Piller, W.E. 2007. Benchmark data of a changingsea—palaeogeograpgy, palaebiogeography and events in the CentralParatethys during the Miocene. Palaeogeography, Palaeoclimatology,Palaeoecology 253: 8–31.
Hohenegger, J., Andersen, N., Báldi, K., Ćorić, S,, Pervesler, P., Rupp, C., andWagreich, M. 2008. Paleoenvironment of the Early Badenian (MiddleMiocene) in the southern Vienna Basin (Austria)—multivariate analysisof the Baden−Sooss section. Geologica Carpathica 59: 461–487.
Hohenegger, J., Coric, S., Khatun, M., Pervesler, P., Rögl, F., Rupp, Ch.,Selge, A., Uchman, A., and Wagreich, M. 2009. Cyclostratigraphic dat−ing in the Lower Badenian (Middle Miocene) of the Vienna Basin (Aus−tria)—the Baden−Sooss core. International Journal of Earth Sciences98: 915–930.
Holcová, K. 2001. New methods in foraminiferal and calcareous nanno−
plankton analysis and evolution of Oligocene and Miocene basins of theSouthern Slovakia. Slovak Geological Magazine 7: 19–41.
Holcová, K. 2002. Calcareous nannoplankton from the Eggenburgian strato−types (Lower Miocene, Central Paratethys). Geologica Carpathica 53:381–390.
Holcová, K. 2005. Quantitative calcareous nannoplankton biostratigraphyof the Oligocene/Miocene boundary interval in the northern part of theBuda Basin (Central Paratethys). Geological Quarterly 49: 263–274.
Holcová, K. 2008. Foraminiferal species diversity in the Central Paratethys—a reflection of global or local events? Geologica Carpathica 59: 71–85.
Howe, R.W. and Sblendorio−Levy, J. 1998. Calcareous nannofossil bio−stratigraphy and sediment accumulation of turbidite sequences on the Ma−deira abyssal plain, sites 950–952. In: P.P.E. Weaver, H.U. Schmincke,and J.V. Firth (eds.), Proceeding of the Ocean Drilling Program, Scien−tific Results 157: 501–520.
Jamrich, M. and Halásová, E. 2010. The evolution of the Late Badenian calcar−eous nannofossil assemblages as a reflexion of the palaeoenvironmentalchanges of the Vienna Basin (Devínska Nová Ves−clay pit). Acta Geo−logica Slovaca 2: 123–140.
Kameo, K. 2002. Late Pliocene Caribbean surface water dynamics and cli−matic changes based on calcareous nannofossil records. Palaeogeogra−phy, Palaeoclimatology, Palaeoecology 179: 211–226.
Kováč, M. 2000. Geodynamic, Paleogeographical and Structural Develop−ment of the Carpathian–Pannonian Region During the Miocene. 202 pp.Veda, Bratislava.
Kroh, A. and Harzhauser, M. 1999. An echinoderm fauna from the LowerMiocene of Austria: Paleoecology and implications for Central Para−tethys paleogeography. Annalen des Naturhistorischen Museums inWien 101A: 145–191.
Kürschner, W.M., Kvaček, Z., and Dilcher, D.L. 2008. The impact of Mio−cene atmospheric carbon dioxine fluctuations on climate and the evolu−tion of terrestrial ecosystems. Proceedings of the National Academy ofSciences of the United States of America 105: 449–453.
Kvaček, Z. and Teodoridis, V. 2007. Tertiary macrofloras of the BohemianMassif: a review with correlations within Boreal and Central Europe.Bulletin of Geosciences 82: 383–408.
Lehotayová, R. 1975. Reticulofenestra excavata n. sp. from the Eggen−burgian to Karpatian of Central Slovakia. Západné Karpaty, sériapaleontológia 2: 39–40.
Lehotayová, R. 1982. Miocene nannoplankton zones in west Carpathians.Západné Karpaty, séria paleontológia 8: 91–110.
Lehotayová, R. 1984. Lower Miocene calcareous nannoflora of the WestCarpathians. Západné Karpaty, séria paleontológia 9: 99–110.
Lourens, L., Hilgen, F., Shackleton, N.J., Laskar, J., and Wilson, D. 2004. TheNeogene period. In: F. Gradstein, J. Ogg, and A. Smith (eds.), A Geologi−cal Time Scale, 409–440. Cambridge University Press, Cambridge.
Mandic, O. and Steininger, F.F. 2003. Computer−based mollusc stratigra−phy—a case study from the Eggenburgian (Lower Miocene) type region(NE Austria). Palaeogeography, Palaeoclimatology, Palaeoecology197: 263–291.
Martini, E. 1971. Standard Tertiary and Quaternary calcareous nanoplanktonzonation. In: A. Farinacci (ed.), Proceeding of 2nd Planktonic Confer−ence, Roma 1970, Roma, 739–785. Tecnoscienza, Rome.
Márton, E., Vass, D., and Túnyi, I. 1995. The Late Tertiary rotation of themegaunit Pelsö and the adjecent West Carpathians area. KnihovničkaZemní Plyn a Nafta 16: 97–108.
Marunteanu, M. 1992. Distribution of the Miocene calcareous nannofossilsin the Intra− and Extra− Carpathian areas of Rumania. KnihovničkaZemní Plyn Nafta 14b (2): 247–262.
Marunteanu, M. 1999. Litho−and biostratigraphy (calcareous nannoplankton)of the Miocene deposits from the Outer Moldavides. Geologica Carpa−thica 50: 313–324.
McIntyre, A. and Be, A.W.H. 1967. Coccolithus neohelis sp.n. a coccolithfossil type in contemporary seas. Deep−Sea Research 14: 369–371.
Molčíková, V. 1983. The Karpatian and Lower Badenian Calcareous nanno−flora of the Carpathian Foredeep in Moravia. Knihovnička Zemní Plyn aNafta 4: 283–304.
Molčíková, V. and Straník, Z. 1987. Calcareous nannoplankton from the
http://dx.doi.org/10.4202/app.2009.0006
HOLCOVÁ—RETICULOFENESTRID NANNOFOSSILS FROM THE CENTRAL PARATETHYS 667

Ždánice−Hustopeče Formation and their relation to underlying forma−tions. Knihovnička Zemní Plyn a Nafta 6b: 59–76.
Piller, W.E., Harzhauser, M., and Mandic, O. 2007. Miocene CentralParatethys stratigraphy—current status and future directions. Stratigra−phy 4: 151–168.
Planderová, E. 1990. Miocene Microflora of Slovak Central Paratethys andits Biostratigraphical Significance. 144 pp. Geologický Ústav DionýzaŠtúra, Bratislava.
Popov, S.V., Rögl, F., Rozanov, A.Y., Steininger, F.F., Shcherba, I.G., andKováč, M. 2004. Lithological−Paleogeographic maps of Paratethys.Courier Forschungsinstitut Senckenberg 250: 1–46.
Pujos, A. 1985. Cenozoic nannofossils, central equatorial Pacific, deep seaDrilling Project Leg 85. In: L. Mayer and F. Theyer (eds.), Initial Re−ports of the Deep Sea Drilling Project 85: 581–608.
Raffi, I. and Flores, J.A. 1995. Pleistocene through Miocene calcareousnannofossils from Eastern Equatorial Pacific Ocean (Leg 138). In: N.G.Pisias, L.A. Mayer, T.R. Janecek, A. Palmer−Julson, and T.H. vanAndel (eds.), Proceedings of the Ocean Drilling Program, ScientificResults 138: 233–286.
Raffi, I., Rio, D., d`Atri, A., Fornaciari, E., and Rochetti, S. 1995. Quantita−tive distribution patterns and biomagnetostratigraphy of middle and lateMiocene calcareous nannofossils from western equatorial Indian andPacific Oceans (Legs 115, 130, and 138). In: N.G. Pisias, L.A. Mayer,T.R. Janecek, A. Palmer−Julson, and T.H. van Andel (eds.), Proceedingof Oceanic Drilling Project, Scientific Results 138: 479–502.
Rahman, A. and Roth, P.H. 1990. Late Neogene paleoceanography andpaleoclimatology of the Gulf of Aden region based on calcareousnannofossils. Paleoceanography 5: 91–107.
Rio, D., Fornaciari, E., and Raffi, I. 1990. Late Oligocene through earlyPleistocene calcareous nannofossils from western equatorial IndianOcean (Leg 115). Proceedings of the Ocean Drilling Program, Scien−tific Results 115: 175–221.
Rögl, F. 1998. Paleogeographic considerations for Mediterranean and Para−tehys seaways (Oligocene to Miocene). Annalen des NaturhistorischeMuseum in Wien 99A: 279–310.
Rögl, F. 1999. Mediterranean and Paratethys. facts and hypotheses of anOligocene to Miocene paleogeography (short overview). GeologicaCarpathica 50: 339–349.
Rögl, F. and Steininger, F.F. 1983. Vom Zerfall der Tethys zu Mediterranund Paratethys—Die neogene Palaegeographie and Palinspastik deszirkummediterranen Raumes. Annalen des Naturhistorische Museum inWien 85/A: 135–164.
Rögl, F., Ćorić, S., Daxner−Höck, G., Harzhauser, M., Mandić, O., Švá−benická, L., and Zorn, I. 2003. Correlation of the Karpatian Stage. In: R.Brzobohatý, I. Cicha, M. Kováč, and F. Rögl (eds.), The Karpatian—aLower Miocene Stage of the Central Paratethys, 27—35. Masaryk Uni−versity, Brno.
Seneš, J. 1961. Paläogeographie des Westkarpatischen Raumes in Beziehungzur übringen Paratethys in Miozän. Geologické práce 60: 160–194.
Spezzaferri, S. and Ćorić, S. 2001. Ecology of Karpatian (Early Miocene) fora−minifera and calcareous nannoplankton from Laa an der Thaya, LowerAustria: a statistical approach. Geologica Carpathica 52: 361–374.
Spezzaferri, S., Ćorić, S., and Stingl, K. 2009. Palaeoenvironmental recon−struction of the Karpatian–Badenian (Late Burdigalian–Early Langhian)transition in the Central Paratethys. A case study from theWagna Section(Austria). Acta Geologica Polonica 59: 523–544.
Švábenická, L. 2002. Calcareous nannofossils of the Upper Karpatian andLower Badenian Deposits in the Carpathian Foredeep, Moravia (CzechRepublic). Geologica Carpathica 53: 197–210.
Švábenická, L., Coric, S., Andreyeva−Grigorovich, A.S., Halásová, E., Ma−runteanu, M., Nagymarosi, A., and Oszcypko−Clowes, M. 2003. CentralParatethys Karpatian Calcareous Nannofossils. R. Brzobohatý, I. Cicha,M. Kováč, and F. Rögl (eds.), The Karpatian—a Lower Miocene Stage ofthe Central Paratethys, 151–168. Masaryk University, Brno.
Teodoridis, V. 2007. Overview of macrofloras from the Most Basin (CzechRepublic) and their phytostratigraphical correlation within Central andBoreal Europe. In: Organizing Committee in Bonn (eds.), Terra Nostra
2, IPC−XII /IOPC−VIII Bonn, Germany 2008, Abstract Volume, 280.GeoUnion Alfred−Wegener−Stiftung, Berlin.
Teodoridis, V. and Kvaček, Z. 2006. Palaeobotanical research of the EarlyMiocene deposits overlying the main coal seam (Libkovice and LomMbs.) in the Most Basin (Czech Republic). Bulletin of Geosciences 81:93–113.
Teodoridis, V., Mazouch, P., Spicer, R.A., and Uhl, D. 2011. RefiningCLAMP—Investigations towards improving the Climate Leaf AnalysisMultivariate Program. Palaeogeography, Palaeoclimatology, Palaeo−ecology 299: 39–48.
Tomanová Petrová, P. and Švábenická, L. 2007. Lower Badenian biostrati−graphy and paleoecology: a case study from the Carpathian Foredeep(Czech Republic). Geologica Carpathica 58: 333–352.
Váňová, M. 1975. Lepidocyclina and Miogypsina from the facistratotype lo−calities Budikovany and Bretka (South Slovakia). In: T. Báldi and J.Seneš (eds.), OM—Egerien, Chronostratigraphie und Neostratotypen,315–339. Slovak Academy of Sciences, Bratislava.
Vass, D. 1995. Global sea level changes reflected on Northern margin of theHungarian Paleogene the Filakovo and Novohrad (Nograd) LowerMiocene Basin (South Slovakia). Mineralia slovaca 27: 193–206.
Vass, D. 1996. The origin and disappearance of Hungarian Paleogene Bas−ins and short−term Lower Miocene Basin in Northern Hungary andSouthern Slovakia. Slovak Geological Magazine 1: 81–95.
Vass, D. and Elečko, M. 1982. Litostratigraphic units of the Kiscellian−Eggenburgian of the Rimava Basin and Cerová Upland. GeologickéPráce, Správy 77: 111–124.
Vass, D., Elečko, M., Pristaš, J., Lexa, J., Hanzel, V., Mordlitba, I., Jánová, V.,Bodnár, J., Husák, Ľ., Filo, M., Májovský, J., and Linkeš, V. 1989. Geol−ogy of the Rimavska Kotlina Depression. 160 pp. Geologický ÚstavDionýza Štúra, Bratislava.
Vass, D., Hók, J., Kováč, P., and Elečko, M. 1993. Succesion of the Paleo−gene and Neogene tectonic events in the light of tectonical analysis.Mineralia Slovaca 25: 79–82.
Vass, D., Konečný, V., and Šefara, J. 1979. Geology of the Ipeľ basin andKrupinská Planina Upland. 227 pp. Geologický Ústav Dionýza Štúra,Bratislava.
Vass, D., Elečko, M., Konečný, V., Krippel, M., Kubeš, P., Lexa, J., Pristaš,J., Zakovič, M., Vozár, J., Vozárová, A., Bodnár, J., Husák, Ľ., Filo, M.,Lacika, J., and Linkeš, V. 2007. Geology of Lučenecká kotlina Depres−sion and Cerová vrchovina Upland. 284 pp. Geologický Ústav DionýzaŠtúra, Bratislava.
Vávra, N. 1979. Die Bryozoenfaunen des österreichischen Tertiärs. NeuesJahrbuch für Geologie und Paläontologie, Abhandlungen 157: 366–392.
Wade, B.S. and Bown, P.R. 2006. Calcareous nannofossils in extreme envi−ronments: the Messinian Salinity Crisis, Polemi Basin, Cyprus. Palaeo−geography, Palaeoclimatology, Palaeoecology 233: 271–286.
Wells, P. and Okada, H. 1997. Response of nannoplankton to major changesin seasurface temperatrure and movements of hydrological fronts oversite DSDP 594 (south Chatham Rise, southeastern New Zealand), dur−ing the last 130 kyr. Marine Micropalaeontology 32: 341–363.
Young, Y.R. 1990. Size variations of neogene Reticulofenestra coccolithsfrom Indian DSDP cores. Journal of Micropaleontology 9: 71–86.
Young, Y. 1999. Neogene. In: P.R. Bown (ed.), Calcareous NannofossilBiostratigraphy, 225–265. Cambridge University Press, Cambridge.
Zachos, J.M., Pagani, L., Sloan, E.T., and Billups, K. 2001. Trends, rhythms,and aberrations in global climate 65 Ma to present. Science 292: 686–693.
Zágoršek, K., Holcová, K., and Trasoň, T. 2008. Bryozoan event from Mid−dle Miocene (Early Badenian) lower neritic sediments from the localityKralice nad Oslavou (Central Paratethys, Moravian part of the Carpa−thian Foredeep). International Journal of Earth Sciences (GeologischeRundschau) 97: 835–850.
Zágoršek, K., Nehyba, S., Holcová, K., Hladiková, Š., and Kroh A. 2009.Fauna from the Middle Miocene (Early Badenian) sediments from thesection Kralice nad Oslavou (Central Paratethys, Moravian part of theCarpathian Foredeep). Bulletin of Geosciences 84: 465–496.
Zágoršek, K. and Holcová, K. 2009: The oldest Early Badenian Bryozoaevent in the Carpathian Foredeep from the Přemyslovice (PY−1, PY−2)boreholes. Přírodovědné Studie Muzea Prostějovska 10–11: 53–59.
668 ACTA PALAEONTOLOGICA POLONICA 58 (3), 2013





























