Embed Size (px)
Citation preview

Morphological Features of NeuronsContaining Calcium-Binding Proteins
in the Human Striatum
LUCIA PRENSA, JOSE MANUEL GIMENEZ-AMAYA, AND ANDRE PARENT*Laboratoire de Neurobiologie, Centre de Recherche Universite Laval Robert-Giffard,
Beauport, Quebec, Canada G1J 2G3
ABSTRACTAn immunohistochemical approach was used to characterize the morphological pheno-
type of neurons containing the calcium-binding proteins calretinin (CR), parvalbumin (PV), orcalbindin-D28k (CB) in the normal human striatum. The protein CR occurs in at least fourmorphologically distinct types of neurons. Apart from the numerous medium-sized aspinyinterneurons and the less abundant giant aspiny interneurons, CR also labels somemedium-sized spiny neurons morphologically identical to striatal projection neurons. Thisfinding indicates that CR is not only confined to striatal interneurons but also may be involvedin the function of certain projection neurons. Some small and peculiar bushy-like aspinyneurons also are enriched with CR. These neurons could correspond to the dwarf orneurogliform neurons first described by Ramon y Cajal (1911). Three types of PV-immunoreactive striatal neurons can be visualized in the human striatum: 1) the commonmedium-sized aspiny leptodendritic neurons, 2) some smaller and profusely arborized aspinyneurons, and 3) a few large and intensely stained neurons with conspicuously beaded andpoorly branched dendrites. The protein CB labels virtually all medium-sized spiny projectionneurons located in the striatal matrix but also identifies a small subset of large and moreintensely immunostained aspiny neurons. The latter finding indicates that CB is not entirelyconfined to striatal projection neurons but also may play a role in local circuit neurons. Thesenormative data should help our understanding of the chemical anatomy of the humanstriatum in both health and disease. J. Comp. Neurol. 390:552–563, 1998.r 1998 Wiley-Liss, Inc.
Indexing terms: human basal ganglia; striatal chemospecific neurons; calretinin; parvalbumin;
calbindin-D28k; neurodegenerative disorders
The calcium-binding proteins of the ‘‘EF-hand’’ family,which include calretinin (CR), parvalbumin (PV), andcalbindin-D28k (CB), are widely distributed within thecentral nervous system. These proteins play a crucial rolein the maintenance of intracellular calcium homeostasis(Goodman et al., 1979; Kretsinger, 1980; Rogers, 1987;Persechini et al., 1989; Heizmann and Hunziker, 1991)and also have served as useful markers of different neuro-nal subsets in the brain (Baimbridge and Miller, 1982;Baimbridge et al., 1992; Andressen et al., 1993; Bennettand Bolam, 1993a,b; Heras et al., 1994; Hontanilla et al.,1994, 1995, 1997; Parent et al., 1996b). In the striatum,CR, PV, and CB label distinct neuronal populations. Forexample, CB is considered a reliable marker of the multi-tude of medium-sized spiny projection neurons embeddedin the striatal matrix (Gerfen et al., 1985; DiFiglia et al.,1989; Gerfen, 1992), whereas PV labels the major g-aminobutyric acid (GABA) interneurons of the striatum
(Cowan et al., 1990; Kita et al., 1990; Kawaguchi et al.,1995). In the rat, CR is localized in a small population ofstriatal interneurons (Jacobowitz and Winsky, 1991; Resi-bois and Rogers, 1992; Bennett and Bolam, 1993a), whereasin human and nonhuman primates, the same proteinoccurs in numerous medium-sized interneurons as well asin a smaller subset of large local circuit neurons (Fortinand Parent, 1994; Parent et al., 1995a).
Grant sponsor: Medical Research Council; Grant numbers: MT-5781,PG-13296; Grant sponsor: Killam Foundation of Canada; Grant sponsor:Comunidad de Madrid; Grant sponsor: FIS; Grant number: 96/0488.
*Correspondence to: Andre Parent, Centre de Recherche, UniversiteLaval Robert-Giffard, 2601 de la Canardiere, F-6500, Beauport, Quebec,Canada G1J 2G3. E-mail: [email protected]
Received 12 June 1997; Revised 24 August 1997; Accepted 1 September1997
THE JOURNAL OF COMPARATIVE NEUROLOGY 390:552–563 (1998)
r 1998 WILEY-LISS, INC.

In the present study, an immunohistochemical approachwas applied to postmortem human brain material fromnormal human individuals to investigate the morphologi-cal features of each type of striatal neuron containingeither CR, PV, or CB. This investigation also reports newtypes of neurons enriched in calcium-binding proteins,whose existence shed new light on our current knowledgeof the chemical anatomy of the human striatum.
MATERIALS AND METHODS
Subjects
The present observations are based on the analysis ofpostmortem material obtained from six normal individu-als (Table 1), with no clinical or pathological evidence ofneurological and psychiatric disorders. The material waskindly provided by Dr. Michel Marois, Service de Pathol-ogy, Hopital Saint-Francois-d’Assise, Quebec, and our pro-tocol was approved by Laval University Committee onEthics and Research. After postmortem delays that rangedfrom 6 to 20 hours, the brains were sliced unfixed into0.5-cm-thick slabs that were fixed by immersion at 4°C,either for 2 days in 4% paraformaldehyde or 3 or 4 days inbuffered formalin (3.75% formaldehyde with 1–1.5% metha-nol, pH 7.4). The slabs were then stored at 4°C in 0.1 Mphosphate-buffered saline (PBS, pH 7.4) with 15% sucroseand 0.1% sodium azide. The portion of the striatumincluded in the slabs extended rostrocaudally from theanterior commissure to the mammillary body level.
Tissue preparation
The slabs were cut with a freezing microtome into50-µm-thick coronal sections that were serially collected incold PBS and placed for 30 minutes at room temperaturein a solution containing an equal volume of PBS and 3%hydrogen peroxide to eliminate endogenous peroxidaseactivity. Series of adjacent sections were then processedimmunohistochemically for the visualization of CR, PV, orCB. The protocol used for the three calcium-binding pro-teins was identical. After three rinses of 10 minutes each,the sections were preincubated for 30 minutes in a solutioncontaining either 5% normal goat serum (for CR) or 5%normal horse serum (for PV and CB) and 0.1% of TritonX-100. They were then incubated overnight at 4°C in asolution containing 5% normal goat or horse serum and0.1% of Triton X-100 plus either anti-CR (SWant, Bell-inzona, Switzerland; dilution 1:2,000), anti-PV (Sigma, St.Louis, MO; dilution 1:2,000) or anti-CB (Sigma; dilution1:2,500) antisera. The polyclonal anti-CR antiserum wasproduced in rabbit by immunization with recombinanthuman CR and did not cross-react with calbindin D-28k or
other known calcium-binding proteins (Schwaller et al.,1993). The anti-PV and anti-CB antibodies were highlyspecific mouse monoclonal antibodies (clone 235 and 300,respectively; Celio et al., 1988; Celio, 1990). The anti-PVantibody was produced by using carp muscle PV as theimmunogen. This mouse monoclonal antibody was di-rected against an epitope at the first Ca12-binding site andreacted in a highly specific fashion with Ca12-bound PV inbrain tissue from several species, including human, and itdid not cross-react with other members of the EF-handfamily of calcium-binding proteins (Celio et al., 1988). Theanti-CB antibody was produced with CB purified fromchicken gut as the immunogen. This antibody reacted in ahighly specific manner with CB in several species, includ-ing humans, and did not cross-react with calbindin-D9knor with CR (Celio, 1990).
After three rinses of 10 minutes each in PBS, thesections were incubated for 1 hour at room temperature inthe secondary antibodies, which were biotinylated goatIgG (for CR) or biotinylated horse IgG (for PV and CB).Then, after three more rinses in PBS, the sections werereincubated for 1 hour at room temperature in 2% avidin-biotin complex (ABC, Vector Labs, Burlingame, CA) accord-ing to the method of Hsu et al. (1981). The sections werethen washed twice in PBS and once in 0.05 M Tris buffer(pH 7.6). The bound peroxidase was revealed by placingthe sections in a medium containing 0.05% 3,38-diamino-benzidine tetrahydrochloride (DAB; Sigma) and 0.003%hydrogen peroxide (H202, 30%) in 0.05 M Tris buffer (pH7.6) at room temperature. The reaction was stopped afterapproximately 5 minutes by extensive washing in PBS. Allimmunostained sections were mounted on gelatin-coatedslides that were air dried, rinsed in distilled water, anddehydrated through passages in ascending grades of alco-hol. Sections were cleared in toluene and coverslipped withPermount, and some of them were treated as above exceptthat the primary antibody was omitted from the incuba-tion medium. These sections remained virtually free ofimmunostaining and served as controls.
Data analysis
The sections were first carefully examined with a Leitzmicroscope to determine the location, approximate num-bers, and morphological features of each type of neuronsexpressing CR, PV, or CB in the human striatum. Eachkind of neuron containing a calcium-binding protein wasthen drawn at 3100 with a microscope equipped with acamera lucida. The large original drawings were reducedwith a photocopier (Toshiba 2550) and then scanned with aMacintosh Quadra-900 computer. The final rendering ofeach drawing was made by using Canvas software (DenebaSystems, Inc., Miami, FL).
RESULTS
Calretinin-immunoreactive neurons
The most common type of CR-immunoreactive (ir) neu-rons found in the human striatum has a medium-sized,oval, or triangular perikaryon (maximum diameter rang-ing from 9 to 18 µm), with two to three thick and smoothprimary dendrites that branch infrequently (Figs. 1A, 3A).The length of the stout primary dendrites is approximately40 µm. The secondary and tertiary dendrites are slightly
TABLE 1. Subject’s Characteristics
Case SexAge
(years)Postmortemdelay (hours) Cause of death
1 M 54 12 Myocardial infarction2 F 25 20 Closed head injury with basal and
convexity fractures3 M 35 6 Head injuries with lacerations of the
scalp4 F 95 18 Rectal carcinoma5 M 21 10 Polytraumatism following car acci-
dent6 F 63 18 Polytraumatism following car acci-
dent
CALCIUM-BINDING PROTEINS IN THE HUMAN STRIATUM 553

Fig. 1. Camera lucida drawings of medium-sized (A) and large-sized (B) CR-ir striatal neurons. The dot in each schema indicates theexact location of the neuron that has been drawn (same for Figs. 2, 4,
and 5). AC, anterior commissure; CD, caudate nucleus; GP, globuspallidus; GPe, external segment of GP; GPi, internal segment of GP;IC, internal capsule; PUT, putamen; TH, thalamus.

varicose and could be followed as far as 200 µm within thestriatal neuropil. These neurons are scattered throughoutthe striatum, but they tend to be more slightly numerousin the caudate nucleus than in the putamen. The secondmost common type of CR-ir striatal neurons possesses avery large globular or triangular perikaryon (22–44 µm),from which emerge numerous thin varicose dendrites thatbranch frequently (Figs. 1B, 3A). Many of the second- andthird-order dendrites curve back toward the cell body, thuscontributing to the very dense aspect of the dendriticdomain of these neurons, which has a radius of approxi-mately 800 µm. These large CR-ir neurons occur through-out the striatum, but, in contrast to the more abundantmedium-sized CR-ir neurons, they tend to be more numer-ous in the putamen than in the caudate nucleus. The axonof both the medium- and large-sized aspiny CR-ir could notbe detected with certainty in the present material. Interest-ingly, medium-sized CR-ir neurons often form small (5–10neurons) clusters in the caudate nucleus, whereas thesame arrangement occurs for the large-sized CR-ir neu-rons in the putamen.
In addition to the medium- and large-sized neuronsdescribed above, which represent by far the majority ofCR-ir striatal neurons, two other types of morphologicallydifferent CR-ir neurons are found in the human striatum.First, a small population of spiny CR-ir neurons occurs inboth the caudate nucleus and the putamen. These neuronshave a round or elongated medium-sized (16–21 µm)perikaryon, from which emerge several thin and moder-ately branched dendrites that fan out in all directionswithin a radius of approximately 700 µm (Fig. 2A). Theproximal portion of these dendrites is smooth, but theirdistal portion is clearly laden with sessile or pedunculatedspines (Figs. 2A, 3A–C). Only a few of these spiny CR-irneurons could be visualized per section of the humanstriatum, and they tend to be slightly more frequent in theputamen than in the caudate nucleus. Second, a popula-tion of small CR-ir neurons with a very peculiar morphol-ogy could be detected in both the caudate nucleus and theputamen (Figs. 2B; 3D,E). Their perikarya (12–16 µm)display a high degree of polymorphism and give rise tomultitudinous short and smooth dendrites that branchedfrequently. The high degree of dendritic branching gives tothese neurons a typical bushy appearance (Fig. 3E). Theirsomatodendritic domain does not exceed 250 µm. Thenumber of these small aspiny CR-ir neurons varies fromone striatum to the other, but in all cases they are morefrequent in the caudate nucleus than in the putamen. Inthe most densely populated striata, their frequency rangesfrom 14 to 21 neurons per section. As for the first two typesof CR-ir neurons, the axon of the medium-sized spiny andsmall aspiny CR-ir striatal neurons could not be traced inthe present material.
Parvalbumin-immunoreactive neurons
Three types of PV-ir neurons could be visualized in thehuman striatum. The most common PV-ir neurons haveround or polygonal perikarya of medium size (18–27 µm)from which emerge two to three smooth dendrites (Figs.4A, 6A). Some of these dendrites arborize close to the cellbody, whereas others branched infrequently and could befollowed for as far as 1,000 µm within the neuropil. Theseneurons are slightly more numerous in the putamen thanin the caudate nucleus. They are more uniformly distrib-uted in the caudate nucleus than in the putamen, where
they predominate in the lateral half of the structure. Adifferent type of PV-ir neurons is also present in areas ofthe human striatum where the neuropil displays a PVimmunostaining that is less intense than in other striatalsectors. This second type of PV-ir neurons has a small(12–17 µm) polygonal perikaryon emitting numerous thindendrites that branched infrequently and rather far fromthe cell body (Figs. 4B, 6B). Their dendrites are typicallydevoid of spines, display a slightly varicose appearance,and extend as far as 800 µm within the neuropil. Theseneurons are very rare (2–4 neurons per section) and couldbe visualized principally in areas of the caudate nucleusand putamen where the PV-ir neuropil is particularlyweak. Finally, a few large (25–27 µm) and intenselyimmunostained PV-ir neurons could be found in the stria-tum. Their polygonal perikarya give rise to several con-spicuously beaded and poorly branched dendrites thatradiate for approximately 700 µm from the cell body (Figs.5A; 6C,D). These large aspiny PV-ir neurons are rare andcould be visualized only in the putamen.
Calbindin-immunoreactive neurons
The human striatum contains multitudinous medium-sized neurons that display a weak and granular CB-ir (Fig.6E). These neurons have a medium-sized (18–24 µm)round or oval cell body, but because of the weak immuno-staining, no processes could be seen to emerge from theperikarya. They constitute by far the densest population ofneurons containing calcium-binding proteins in the entirehuman striatum. They abound throughout both the cau-date nucleus and the putamen, except for some oval orelongated striatal areas of approximately 600–800 µm indiameter that are rather devoid of such CB-ir neurons. Amuch smaller population of larger and more intenselyimmunostained CB-ir neurons also occurs in the humanstriatum (Figs. 5B, 6E). Although present at all rostrocau-dal levels of the striatum, these neurons are particularlyeasy to delineate in the caudal half of the putamen wherethe overall intensity of CB immunostaining is significantlyweaker than that more rostrally. This type of neurons hasan oval perikaryon (20–26 µm) from which emerge two tothree primary dendrites that branch infrequently. Thesedendrites have a smooth surface and some could befollowed as far as 600 µm within the neuropil. The largeand intensely CB-ir neurons are much less frequent thanthe medium-sized weakly CB immunostained neuronsthat populate virtually the entire striatum. Their numbervaries from 150 to 200 per section, and their axon is notclearly distinguishable from their thin dendrites.
DISCUSSION
The present study has provided a detailed description ofthe morphological characteristics of different types ofneurons containing either CR, PV, and CB that wereencountered in the human striatum. These findings inhumans will now be compared with data obtained inrodents and nonhuman primates to obtain a better knowl-edge of the chemical anatomy of the mammalian striatum.
Calretinin-immunoreactive neurons
Striatal neurons are usually subdivided into two majorcategories: 1) the medium-sized projection neurons, which
CALCIUM-BINDING PROTEINS IN THE HUMAN STRIATUM 555

Fig. 2. Camera lucida drawings of spiny (A) and neurogliform (B) CR-ir striatal cells. For abbrevia-tions, see Figure 1.
556 L. PRENSA ET AL.

represent more than 90–95% of the total striatal neuronalpopulation and whose dendrites are characteristicallyladen with spines and 2) the local circuit neurons, whichcomprise both medium- and large-sized elements withtypically smooth or varicose dendrites (Parent, 1986,1996). Hence, because of their smooth dendrites, both thenumerous medium-sized CR-ir neurons and the less abun-
dant large-sized CR-ir neurons present in the humanstriatum are likely to belong to the category of striatalinterneurons. These two types of chemospecific neurons,however, appear to correspond to two entirely differentsubsets of striatal interneurons.
The medium-sized CR-ir neurons, which represent lessthan 3% of the total neuronal population in humans
Fig. 3. Photomicrographs illustrating the four types of calretininimmunoreactive (CR-ir) neurons. A: Low-power view showing a me-dium-sized spiny CR-ir neuron (arrow) surrounded by numerouslarge- and medium-sized aspiny CR-ir neurons in the putamen. B:High-power view of a spiny CR-ir neuron in the caudate nucleus. C:Secondary dendrites of a spiny CR-ir neuron. D: Low-power view of a
neurogliform (arrow) and several large and medium-sized CR-irstriatal cells in the putamen. E: High-power view of a neurogliformcell in the caudate nucleus. Note the multitude of processes thatbranch profusely, close to the cell body. Scale bars 5 200 µm in A,100 µm in B, 15 µm in C, 200 µm in D, 50 µm in E.
CALCIUM-BINDING PROTEINS IN THE HUMAN STRIATUM 557
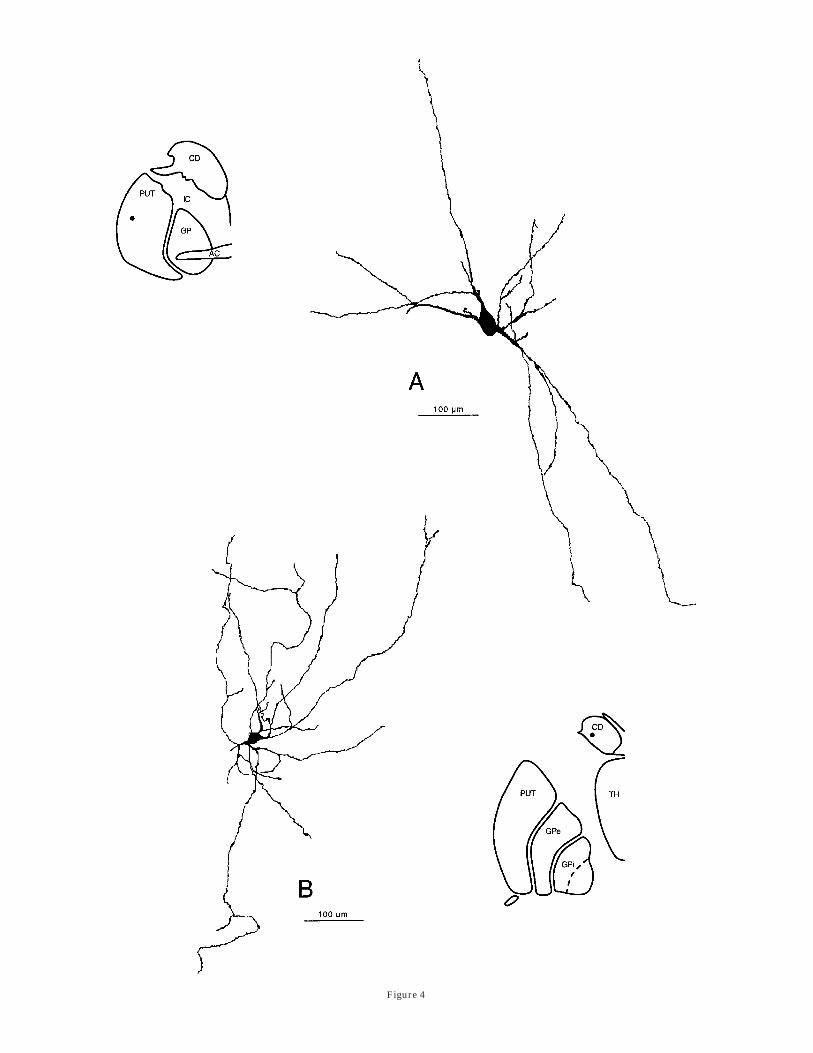
Figure 4

(Parent et al., 1995a), most likely correspond to a similartype of interneurons described previously in the rat stria-tum (Jacobowitz and Winsky, 1991; Resibois and Rogers,1992; Bennett and Bolam, 1993a). Light and electronmicroscopic studies, as well as colocalization and retro-grade labeling experiments, have revealed that medium-sized CR-ir neurons in rodent striatum represent a uniquepopulation of interneurons. These neurons have aspinydendrites, are positive for GABA, but not for cholineacetyltransferase (ChAT), CB, PV, and b-nicotinamideadenine dinucleotide phosphate-diaphorase (NADPH-d)and do not project outside the striatum (Bennett andBolam, 1993a; Kubota et al., 1993; Kawaguchi et al., 1995;Figueredo-Cardenas et al., 1996a,b). In humans, the me-dium-sized CR-ir striatal interneurons are also likely torepresent a distinct subset of neurons that do not colocal-ize with any of the known markers of striatal neurons,except perhaps GABA (Parent et al., 1996a). Furthermore,recent studies have shown that the medium-sized CR-irstriatal interneurons, but not the large-sized ones, areselectively spared in Huntington’s disease (Cicchetti andParent, 1996a).
According to their morphological features, the less nu-merous large-sized CR-ir neurons appear to correspond tothe large aspiny type II cells described from monkey Golgimaterial (DiFiglia et al., 1976). These neurons are believedto correspond to the giant cholinergic striatal interneuronsin humans (Parent et al., 1984; Selden et al., 1994), andcolocalization studies indeed reveal that the majority ofthe large-sized CR-ir express ChAT (Parent et al., 1996a).These findings indicate that, in contrast to the uniquenessof the subset of medium-sized CR-ir interneurons, mostlarge-sized CR-ir belong to the population of large choliner-gic striatal interneurons in humans. The presence oflarge-sized CR-ir neurons may be a typical feature of theprimate striatum because such neurons occur in monkeys(Fortin and Parent, 1994), but not in rodents (Bennett andBolam, 1993a; Figueredo-Cardenas et al., 1996a). Alsoworth noting is the fact that virtually all large-sized CR-irneurons, but very few medium-sized CR-ir neurons, ex-press substance P (neurokinin-1, NK-1) receptor in mon-key and human striatum (Parent et al., 1995b; Cicchetti etal., 1996b). Because most large CR-ir neurons are, in fact,cholinergic neurons and that all of them express NK-1receptors, the large cholinergic neuron stands out as amajor target of substance P-containing fibers in the hu-man striatum, as it is also the case in the rat (Bolam et al.,1986; Gerfen, 1991; Martone et al., 1992).
In addition to the medium- and large-sized aspiny CR-irneurons described above, there exist at least two othertypes of CR-ir neurons in the human striatum. Perhapsthe most surprising finding of the present study was thediscovery of some medium-sized CR-ir neurons that dis-play the typical morphology of projection neurons, withdendrites heavily laden with spines. On the basis of theirmorphological characteristics, these neurons correspondbest to the spiny type I neurons of the Golgi classification
proposed by DiFiglia et al. (1976) for the monkey striatum.These neurons are considered as the typical projectionneurons of the striatum (Parent, 1986), and they can easilybe identified by their content in CB (see Calbindin-Immunoreactive Neurons section). Thus, although thevast majority of striatal medium-sized CR-ir neurons canbe considered as interneurons, the results of present studyreveal that some of the medium-sized CR-ir neurons couldact as projection neurons in the human striatum. Smallpolymorphic and bushy-like CR-ir cells are another pecu-liar type of CR-ir neurons that could be visualized in thehuman striatum. The characteristic morphological fea-tures of these cells indicate that they may correspond tothe dwarf or neurogliform neurons described in the stria-tum of humans (Ramon y Cajal, 1911), cats (Kemp andPowell, 1971), and monkeys (Fox et al., 1974; DiFiglia etal., 1976). The exact nature of these cells is still unknown,but Golgi studies suggest that they significantly differ bothin size and morphology from the glial cells of the striatum(Fox et al., 1974; DiFiglia et al., 1976). Because their axoncould not be identified with certainty, the role that thesesmall bushy CR-ir elements may play at striatal levelremains obscure.
Parvalbumin-immunoreactive neurons
The most frequent striatal PV-ir neurons present in thehuman striatum are morphologically similar to the me-dium-sized aspiny PV-ir neurons that occur in the stria-tum of rats (see review by Kawaguchi et al., 1995),monkeys (Cote et al., 1991; Yelnik et al., 1991), andhumans (Waldvogel and Faull, 1993; Ferrer et al., 1994).These neurons correspond to the intensely GABA- andGAD67-immunoreactive striatal neurons (Kita et al., 1990;Cowan et al., 1990) and are considered as the majorinhibitory interneurons in the rodent striatum (Kawagu-chi et al., 1995). They are the prototype of what Yelnik andcolleagues (1991) refer to as leptodendritic striatal neu-rons.
In addition to these medium-sized PV-ir neurons, asmaller number of small and large aspiny PV-ir neuronsare present in the human striatum. The smaller neuronsare more profusely arborized than the most commonmedium-sized CR-ir and occur principally in areas wherethe immunostaining intensity of the PV-ir neuropil isparticularly weak, which correspond to enkephalin-richstriosomes in human (Waldvogel and Faull, 1993). In therat, the somatodendritic domain of PV-ir neurons that liewithin the striosomal compartment is said to be morpho-logically different from that of PV-ir neurons located in theextrastriosomal matrix compartment (Bennett and Bolam,1994a). The large PV-ir neurons are similar to the fewlarge and intensely stained PV-ir neurons with typicallybeaded dendrites visualized in a previous study of thehuman striatum (Waldvogel and Faull, 1993). Such smalland large PV-ir neurons have never been observed in therodent striatum.
Calbindin-immunoreactive neurons
Both the caudate nucleus and the putamen in humanare densely populated by medium-sized neurons display-
Fig. 4. Camera lucida drawings of an example of one of the mostcommon types of medium-sized PV-ir striatal neurons (A) and of asmaller and more highly branched PV-ir neuron that occurs in striatalareas displaying a rather weak PV immunostaining (B). For abbrevia-tions, see Figure 1.
CALCIUM-BINDING PROTEINS IN THE HUMAN STRIATUM 559

Fig. 5. Camera lucida drawings of a large PV-ir neuron (A) and a large CB-ir neuron (B). Forabbreviations, see Figure 1.
560 L. PRENSA ET AL.

ing a weak and granular CB immunoreactivity. On thebasis of their size, number, and morphology, these neuronsmost likely correspond to the spiny projections neurons,which abound particularly in the matrix compartment ofthe striatum (Gerfen et al., 1985; Bennett and Bolam,1993b, 1994a; Selden et al., 1994). The second type of CB-irneurons that occur in the human striatum are much lessnumerous, but more intensely immunostained than themore common medium-sized CB-ir neurons. They appear
to be equivalent to the intensely stained aspiny CB-irneurons present in the striatum of rats (Roberts andDiFiglia, 1990; Bennett and Bolam, 1993b; Kubota et al.,1993) and humans (Kiyama et al., 1990; Selden et al.,1994). In contrast to the weakly stained spiny medium-sized CB-ir neurons, these larger aspiny CB-ir neurons arerelatively spared in the Huntington’s disease striatum(Kiyama et al., 1990), and a significant proportion of them(20–40%) coexpress NADPH-d (Bennett and Bolam, 1993b;
Fig. 6. Photomicrographs illustrating the three types of PV-irneurons and the large CB-ir neurons in the human striatum. A:Low-power view of the putamen illustrating three medium-sizedaspiny neurons corresponding to the most frequent type of PV-irneurons. B: Example of a smaller and more densely ramified PV-irneuron present in an area of the caudate nucleus that displays
particularly weak PV immunostaining. C: Example of a large PV-irneuron in the putamen. D: High-power view of a conspicuously beadeddendrite belonging to a large PV-ir neuron. E: Low-power view of twolarge and intensely stained CB-ir neurons lying in the putamen amongnumerous smaller and more weakly stained CB-ir neurons. Scalebars 5 100 µm in A, 50 µm in B, 100 µm in C, 15 µm in D, 200 µm in E.
CALCIUM-BINDING PROTEINS IN THE HUMAN STRIATUM 561

Kubota et al., 1993; Selden et al., 1994). These data revealthat at least a small population of striatal CB-ir neuronscould act as local circuit neurons in the human striatum.These neurons could be either a subset of the population ofinterneurons characterized by their immunoreactivity forsomatostatin, neuropeptide Y, and NADPH-d (DiFigliaand Aronin, 1982; Takagi et al., 1983; Vincent and Johans-son, 1983; Vincent et al., 1983; Smith and Parent, 1986) orconstitute a distinct population of striatal neurons.
CONCLUSIONS
The present study has provided a detailed account of themorphological characteristics of the various types of neu-rons displaying immunoreactivity for one of the threeprincipal members of the EF-hand family of calcium-binding proteins, that is, CR, PV, and CB in the humanstriatum. This investigation has brought new informationon the morphological organization of the somatodendriticdomain of already known chemospecific neurons and,moreover, has allowed us to characterize hitherto un-known types of striatal neurons expressing calcium-binding proteins. These normative data complement ourstill limited knowledge of the chemical anatomy of thenormal human striatum. They should also help our under-standing of the major changes that occur at the striatallevel in certain neurodegenerative diseases that specifi-cally involve the human striatum, such as Huntington’sdisease.
ACKNOWLEDGMENTS
The authors thank Carole Emond and Lisette Bertrandfor skillful technical assistance. We also thank Dr. MichelMarois for having provided the postmortem material usedin the present study. L. Prensa was on leave of absencefrom the Departamento de Morfologıa, U.A.M., Madridand the Departamento de Anatomıa, Universidad deNavarra, Pamplona, Spain. J.M. Gimenez-Amaya’s cur-rent addresses are the same as those of L. Prensa.
LITERATURE CITED
Andressen, C., I. Blumcke, and M.R. Celio (1993) Calcium-binding proteins:Selective markers of nerve cells. Cell Tissue Res. 271:181–208.
Baimbridge, K.G., and J.J. Miller (1982) Immunohistochemical localizationof calcium-binding protein in the cerebellum, hippocampal formationand olfactory bulb of the rat. Brain Res. 245:223–229.
Baimbridge, K.G., M.R. Celio, and J.H. Rogers (1992) Calcium bindingproteins in the nervous system. Trends Neurosci. 15:303–308.
Bennett, B.D., and J.P. Bolam (1993a) Characterization of calretinin-immunoreactive structures in the striatum of the rat. Brain Res.609:137–148.
Bennett, B.D., and J.P. Bolam (1993b) Two populations of calbindinD28k-immunoreactive neurones in the striatum of the rat. Brain Res.610:305–310.
Bennett, B.D., and J.P. Bolam (1994a) Localization of calcium bindingproteins in the neostriatum. In G. Percheron, J.S. Mckenzie, and J.Feger (eds): New Ideas and Data on Structure and Forebrain, The BasalGanglia IV, Vol. 41. New York: Plenum Press, pp. 21–24.
Bennett, B.D., and J.P. Bolam (1994b) Localization of parvalbumin-immunoreactive structures in primate caudate-putamen. J. Comp.Neurol. 347:340–356.
Bolam, J.P., C.A. Ingham, P.N. Izzo, A.I. Levey, D.B. Rye, A.D. Smith, andB.H. Wainer (1986) Substance P-containing terminals in synapticcontact with cholinergic neurons in the neostriatum and basal fore-brain: A double immunocytochemical study in the rat. Brain Res.397:279–289.
Celio, M.R. (1990) Calbindin D28k and parvalbumin in the rat nervoussystem. Neuroscience 35:375–475.
Celio, M.R., W. Baier, P. de Viragh, E. Scharer, and C. Gerday (1988)Monoclonal antibodies directed against the calcium binding proteinparvalbumin. Cell Calcium 9:81–86.
Cicchetti, F., and A. Parent (1996) Striatal interneurons in Huntington’sdisease: Selective increase in the density of calretinin-immunoreactivemedium-sized neurons. Mov. Disord. 11:619–626.
Cicchetti, F., P.V. Gould, and A. Parent (1996) Sparing of striatal neuronscoexpressing calretinin and substance P (NK1) receptor in Huntington’sdisease. Brain Res. 730:232–237.
Cote, P.Y., A.F. Sadikot, and A. Parent (1991) Complementary distributionof calbindin D-28k and parvalbumin in the basal forebrain and mid-brain of the squirrel monkey. Eur. J. Neurosci. 3:1316–1329.
Cowan, R.L., C.J. Wilson, P.C. Emson, and C.W. Heizmann (1990) Parvalbu-min-containing GABAergic interneurons in the rat neostriatum. J.Comp. Neurol. 302:197–205.
DiFiglia, M., and N. Aronin (1982) Ultrastructural features of immunoreac-tive somatostatin neurons in the rat caudate nucleus. J. Neurosci.2:1267–1274.
TABLE 2. Neurons Containing Calretinin, Parvalbumin, and Calbindin in Human Striatum
Proteins Neuronal type Perikarya Dendrites DistributionPresence in
other species
Calretinin Medium-aspiny (9–18 µm) Oval or triangular 2–3 thick, smooth, and infrequentlybranched
Scattered, more numerous in caudatethan in putamen
Rat,1 monkey2
Large-aspiny (22–44 µm) Globular or triangular Numerous thin, varicose, and fre-quently branched
Scattered, more numerous in putamenthan in caudate
Monkey2
Medium-spiny (16–21 µm) Round or elongated Numerous thin, spiny, and moderatelybranched
Only a few per section, more frequentin putamen than in caudate
—
Medium-aspiny (12–16 µm) High polymorphism Multitudinous short, smooth, and fre-quently branched
Only a few per section, more frequentin caudate than in putamen
—
Parvalbumin Medium-aspiny (18–27 µm) Round or polygonal 2–3 smooth and irregularly branched Scattered, more numerous in putamenthan in caudate
Rat,3 monkey4
Medium-aspiny (12–17 µm) Polygonal Numerous thin, varicose, and infre-quently branched
Only a few per section, exclusively inareas devoid of PV-ir neuropil
—
Large-aspiny (25–27 µm) Polygonal Numerous conspicously beaded andinfrequently branched
Only a few per section, exclusively inthe putamen
—
Calbindin Medium-spiny (18–24 µm) Round or oval No processes can be seen to emergefrom the perikarya
Scattered, highly abundant in thematrix
Rat,5 monkey5
Large-aspiny (20–26 µm) Oval 2–3 thin, smooth and infrequentlybranched
Scattered, moderately abundant in thecaudate and putamen
Rat6
1Bennett and Bolam, 1993a.2Fortin and Parent, 1994.3Kawaguchi et al., 1995.4Cote et al., 1991.5Gerfen et al., 1985.6Bennett and Bolam 1993b.—No equivalent was found in other species.
562 L. PRENSA ET AL.

DiFiglia, M., P. Pasik, and T.A. Pasik (1976) Golgi study of neuronal typesin the neostriatum of monkeys. Brain Res. 114:245–256.
DiFiglia, M., S. Christakos, and N. Aronin (1989) Ultrastructural localiza-tion of calbindin-D28k in the rat and monkey basal ganglia, includingsubcellular distribution with colloidal gold labeling. J. Comp. Neurol.279:653–665.
Ferrer, I., J. Kulisevsky, G. Gonzalez, A. Escartın, A. Chivite, and R. Casas(1994) Parvalbumin-immunoreactive neurons in the cerebral cortexand striatum in Huntington’s disease. Neurodegeneration 3:169–173.
Figueredo-Cardenas, G., L. Medina, and A. Reiner (1996a) Calretinin islargely localized to a unique population of striatal interneurons in rats.Brain Res. 709:145–150.
Figueredo-Cardenas, G., M. Morello, G. Sancesario, G. Bernardi, and A.Reiner (1996b) Colocalization of somatostatin, neuropeptide Y, neuro-nal nitric oxide synthase and NADPH-diaphorase in striatal interneu-rons in rats. Brain Res. 735:317–324.
Fortin, M., and A. Parent (1994) Patches in the striatum of squirrelmonkeys are enriched with calretinin fibers but devoid of calretinin cellbodies. Neurosci. Lett. 182:51–54.
Fox, C.A., I.J. Lu Qui, and J.A. Rafols (1974) Further observations onRamon y Cajal’s ‘‘dwarf ’’ or ‘‘neurogliaform’’ neurons and the oligoden-droglia in the primate striatum. J. Hirnforsch. 15:517–527.
Gerfen, C.R. (1991) Substance P (neurokinin-1) receptor mRNA is selec-tively expressed in cholinergic neurons in the striatum and basalforebrain. Brain Res. 556:165–170.
Gerfen, C.R. (1992) The neostriatal mosaic: Multiple levels of compartmen-tal organization. Trends Neurosci. 15:133–139.
Gerfen, C.R., K.G. Baimbridge, and J.J. Miller (1985) The neostriatalmosaic: Compartmental distribution of calcium binding protein andparvalbumin in the basal ganglia of the rat and monkey. Proc. Natl.Acad. Sci. USA 82:8780–8784.
Goodman, M., J.F. Perchere, J. Haiech, and J.G. Demaille (1979) Evolution-ary diversification of structure and function in the family of intracellu-lar calcium-binding proteins. J. Mol. Evol. 13:331–352.
Heizmann, C.W., and W. Hunziker (1991) Intracellular calcium-bindingproteins: More sites than insights. Trends Biol. Sci. 16:98–103.
Heras, S. de las, B. Hontanilla, E. Mengual, and J.M. Gimenez-Amaya(1994) Immunohistochemical distribution of calbindin D-28k and parv-albumin in the head of the caudate nucleus and the substantia nigra ofthe cat. J. Morphol. 221:291–307.
Hontanilla, B., A. Parent, and J.M. Gimenez-Amaya (1994) Compartmentaldistribution of parvalbumin and calbindin D-28k in rat globus pallidus.Neuroreport 5:2269–2272.
Hontanilla, B., A. Parent, and J.M. Gimenez-Amaya (1995) Heterogeneousdistribution of neurons containing calbindin D-28k and/or parvalbuminin the rat red nucleus. Brain Res. 696:121–126.
Hontanilla, B., A. Parent, and J.M. Gimenez-Amaya (1997) Parvalbuminand calbindin D-28k in the entopeduncular nucleus, subthalamicnucleus and substantia nigra of the rat as revealed by double immuno-histochemical methods. Synapse 25:359–367.
Hsu, S.M., L. Raine, and H. Fanger (1981) The use of avidin-biotinperoxidase complex (ABC) in immunoperoxidase techniques: A compari-son between ABC and unlabeled antibody (PAP) procedures. J. Histo-chem. Cytochem. 29:577–580.
Jacobowitz, D.M., and L. Winsky (1991) Immunocytochemical localizationof calretinin in the forebrain of the rat. J. Comp. Neurol. 304:198–218.
Kawaguchi, Y., C.J. Wilson, S.J. Augood, and P.C. Emson (1995) Striatalinterneurons: Chemical, physiological and morphological characteriza-tion. Trends Neurosci. 18:527–535.
Kemp, J.A., and T.P.S. Powell (1971) The structure of the caudate nucleus ofthe cat: Light and electron microscopy. Philos. Trans. R. Soc. Lond.[Biol.] 262:383–401.
Kita, H., T. Kosaka, and C.W. Heizmann (1990) Parvalbumin-immunoreac-tive neurons in the rat neostriatum: A light and electron microscopicstudy. Brain Res. 536:1–15.
Kiyama, H., A. Seto-Ohshima, and P.C. Emson (1990) Calbindin D28k as amarker for the degeneration of the striatonigral pathway in Hunting-ton’s disease. Brain Res. 536:1–15.
Kretsinger, R.H. (1980) Structures and evolution of calcium-modulatedproteins. CRC Crit. Rev. 8:119–138.
Kubota, Y., S. Mikawa, and Y. Kawaguchi (1993) Neostriatal GABAergicinterneurones contain NOS, calretinin or parvalbumin. Neuroreport5:205–208.
Martone, M.E., D.M. Armstrong, S.J. Young, and P.M. Groves (1992)Ultrastructural examination of enkephalin and substance P input tocholinergic neurons within the rat neostriatum. Brain Res. 594:253–262.
Parent, A. (1986) Comparative Neurobiology of the Basal Ganglia. NewYork: John Wiley & Sons.
Parent, A. (1996) Carpenter’s Human Neuroanatomy. Baltimore, MD:Williams and Wilkins.
Parent, A., C. Csonka, and P. Etienne (1984) The occurrence of largeacetylcholinesterase-containing neurons in human neostriatum as dis-closed in normal and Alzheimer-diseased brains. Brain Res. 291:154–158.
Parent, A., F. Cicchetti, and T.G. Beach (1995a) Calretinin-immunoreactiveneurons in the human striatum. Brain Res. 674:347-351.
Parent, A., F. Cicchetti, and T.G. Beach (1995b) Striatal neurones display-ing substance P (NK1) receptor immunoreactivity in human andnon-human primates. Neuroreport 6:721–724.
Parent, A., F. Cicchetti, and T.G. Beach (1996a) Immunocytochemicalcharacterization of neurons expressing calretinin in human striatum.Brain Pathol. 6:355.
Parent, A., M. Fortin, P.-Y.Cote, and F. Cicchetti (1996b) Calcium-bindingproteins in primate basal ganglia. Neurosci. Res. 25:309–334.
Persechini, A., N.D. Moncrief, and R.H. Kretsinger (1989) The EF-handfamily of calcium-modulated proteins. Trends Neurosci. 12:462–467.
Ramon y Cajal, S. (1911) Histologie du Systeme Nerveux de l’ Homme et desVertebres, Vol. II (transl. by L. Azoulay). Paris, Maloine pp. 504–518.
Resibois, A., and J.H. Rogers (1992) Calretinin in rat brain: An immunohis-tochemical study. Neuroscience 46:101–134.
Rogers, J.H. (1987) Calretinin: A gene for a novel calcium-binding proteinexpressed principally in neurons. J. Cell Biol. 105:1343–1353.
Roberts, R.C., and M. DiFiglia (1990) Long-term survival of GABA-,enkephalin-, NADPH-diaphorase- and calbindin-d28k-containing neu-rons in fetal striatal grafts. Brain Res. 532:151–159.
Schwaller, B., P. Buchwald, I. Blumcke, M.R. Celio, and W. Hunziker (1993)Characterization of a polyclonal antiserum against the purified humanrecombinant calcium binding protein calretinin. Cell Calcium 14:639–648.
Selden, N., C. Geula, L. Hersh, and M.M. Mesulam (1994) Human striatum:Chemoarchitecture of the caudate nucleus, putamen and ventral stria-tum in health and Alzheimer’s disease. Neuroscience 60:621–636.
Smith, Y., and A. Parent (1986) Neuropeptide Y-immunoreactive neurons inthe striatum of the cat and monkey: Morphological characteristics,intrinsic organization and co-localization with somatostatin. Brain Res.372:241–252.
Takagi, H., P. Somogyi, J. Somogyi, and A.D. Smith (1983) Fine structuralstudies on a type of somatostatin-immunoreactive neuron and itssynaptic connections in the rat neostriatum: A correlated light andelectron microscopic study. J. Comp. Neurol. 214:1–16.
Vincent, S.R., and O. Johansson (1983) Striatal neurons containing bothsomatostatin- and avian pancreatic polypeptide (APP)-like immunore-activities and NADPH-diaphorase activity: A light and electron micro-scopic study. J. Comp. Neurol. 217:264–270.
Vincent, S.R., W.A. Staines, and H.C. Fibiger (1983) Histochemical demon-stration of separate populations of somatostatin and cholinergic neu-rons in the rat striatum. Neurosci. Lett. 35:111–144.
Waldvogel, H.J., and R.L.M. Faull (1993) Compartmentalization of parval-bumin immunoreactivity in the human striatum. Brain Res. 610:311–316.
Yelnik, J., C. Francois, G. Percheron, and D. Tande (1991) MorphologicalTaxonomy of the Neurons of the Primate Striatum. J. Comp. Neurol.313:273–294.
CALCIUM-BINDING PROTEINS IN THE HUMAN STRIATUM 563