Embed Size (px)
Citation preview


Morphogenesis Model for Systematic Simulationof Forms’ Co-evolution with Constraints:
Application to Mitosis
Abdoulaye Sarr, Alexandra Fronville, and Vincent Rodin
UMR CNRS 6285, Lab-STICC, CID, IHSEVComputer Science Department
Universite de Brest20 Avenue Le Gorgeu, Brest, France
{abdoulaye.sarr,alexandra.fronville,vincent.rodin}@univ-brest.fr
Abstract. We present a new approach to understand forms’ emergencein a cellular system. We set the hypothesis that beyond the influence ofmechanical forces and gene expression, constraints applied to the cellsover time play a key role in the acquisition of specific shape. We considerthat these constraints are the fundamental principles and basic causeof morphogenesis. In our model, it’s due to these constraints that cellschoose a particular direction while dividing, migrate or die. Our approachof morphogenesis based on constraints has been used to get effectivelyfor a given form all possible evolutions by growth at latter times. Suchwork ensures to do some pattern prediction.
Keywords: Developmental systems, Virtual biology, Morphogenesis,Mathematical morphology, Viability theory.
1 Introduction
Facing the experimental complexity, understanding of the living is more andmore focused on in silico models. The convenience of implementation of a vir-tual lab has made possible the formulation and testing of many hypotheses inBiology, particularly in Morphogenesis. This gave birth to a multitude of mod-els in this area. But we note that they mostly aim to make an integration ofthe dynamic interactions between different spatial and/or temporal scales. Thisapproach introduces some complexity in models that limits their understandingand effectiveness with respect to their purpose. Thus, we consider that the cellmust be the focus, which determines both causalities and downgrades. In otherwords, as a first step of a better understanding, observations must be restrictedto a single spatial and temporal scale of the biological organization.
Thanks to advances in microscopy and imaging, very detailed data on com-ponents and structures of living organisms are now available. Melani and al.achieved a tracking of cell nuclei and the identification of cell divisions in livezebra fish embryos using 3D+time images acquired by confocal laser scanning
A.-H. Dediu et al. (Eds.): TPNC 2014, LNCS 8890, pp. 231–242, 2014.c© Springer International Publishing Switzerland 2014

232 A. Sarr, A. Fronville, and V. Rodin
microscopy [5]. While the zebrafish embryo contains a few pairs of cells, wenoticed:
1. a geometrical segmentation during cellular proliferation2. a specific arrangement of cells at each stage
Theses observations allow us to formulate a set of principles so as to propose amodel. First, the noticed geometrical segmentation allow us to adopt a 2D model,discrete in time and space to study the forms appearing in the early stages ofmorphogenesis. Besides, we make the hypothesis that we can define a morpho-logical dynamic based only on spatial constraints of the cells. Furthermore, weassume that this dynamic is the fundamental principle of morphogenesis and istherefore able to describe all evolutions of a form, both those that modify it andthose that maintain it.
The main contribution of this paper is to present the formalization and imple-mentation of such a model. It’s organized as follows, section 2 provides a quickoverview of some existing models and finishes by our positioning with respect tothese models. Section 3 presents the mathematical morphological dynamic rely-ing on the viability theory [1]. Then we present in section 4 a system based onthat model which allow to construct and reach over time all possible evolutions ofa given form. Section 5 studies the coupling between constraints and organisms(co-evolution) with various types of constraints considered by epigenetics as veryimportant to understand the development of living. Finally section 6 concludesthis paper by giving some applications of this work and then highlighting theperspectives we should address in coming work.
2 Related Works
Morphogenesis is an important research field within Developmental Biology. Itcan be defined as the set of processes that generates shapes in the embryo.There exists many models in the area depending on the main factors consideredin biological form creation and also the studied organisms (prokaryotes, animals,plant etc.).
Tensegrity model is for example interested in cells’ shape changing. This modelconsiders biomechanical forces between cells and the extracellular matrix. Thestretching of cells adhering to the extracellular matrix may result from localreshuffle in this latter. According to this model, growth-generated strains andpressures in developing tissues regulate morphogenesis throughout development[3]. It is therefore the biomechanical forces which play the key role. For exam-ple by modulating cell differentiation, influencing the direction of division ordeforming tissues. However, the question of cell diversity even arises before theacquisition of shape [7]. Indeed, when the embryo has only a few pairs of cells,we can see already a diversification of biochemical content or a diversificationof embryonic cells morphology. That may be the result of genetic and molecu-lar interactions. Indeed, the emergence of forms also stems from the acquisitionof differential properties, of cell mobility and gene expression throughout celldevelopment.

Systematic Simulation of Forms’ Co-evolution 233
Among the mechanisms of cell morphogenesis, we have also artificial Regu-latory Networks. They define a series of regulatory genes and structural genes.The firsts consists of a network of rules determining the evolution of the systemand the latter are intended to each generate a simple specific pattern. They canbe seen as a dynamic system following different trajectories in a state space [4].However, even if the detailed knowledge of genomic sequences allow to determinewhere and when different genes are expressed in the embryo, it is insufficient tounderstand how the organism emerge [6].
So, focusing only on the cellular constraints, we tried to define a mechanismof morphogenesis including the cellular dynamics and allowing to describe andget all possible evolutions of a form over time.
3 Mathematical Morphological Dynamic
In Mathematics, the viability theory offers concepts and methods to control adynamical system in a given fixed environment, in order to maintain it in a setof constraints of viability. Applied to Morphogenesis, this means that we shouldhave at least one co-viable evolution of the cells’ state and their environmentbased on each state-environment pair. This formalization allows us to establishrules in terms of cell plus action. The application of such rules by a cell, onthe one hand is subject to the observance of an array of constraints and, on theother hand causes some biological effects both on the cell and the overall form.
K ⊂ P(X)1 denotes the morphological environment2 (X = 2 denotes the setof containment cells, contained in the complement of vitellus 3).
Cells x ∈ X ∪ ∅ are either characterized by their position (living cells) or bytheir death made of tissues L which are subsets of cells (L ∈ P(X)).
The subset of eight genetic actions d of cells is:
A := {(1, 0, 0), (−1, 0, 0), (0, 1, 0), (0,−1, 0), (0, 0, 1), (0, 0,−1), (0, 0, 0), ∅}
A is made of the six geometric directions, the origin and the empty set. Here,we restrict morphogenesis in the plan:
A := {(0, 1), (0,−1), (1, 0), (−1, 0), (0, 0), ∅}
For convenience, we replace (0, 1), (0,−1), (1, 0), (−1, 0), (0, 0) and ∅ respec-tively by 1, 2, 3, 4, 5 and 6.
A := {1, 2, 3, 4, 5, 6}
These genetic actions allow to describe cells’ behaviour:
1 Supplied with the structure of max-plus algebra for the operation ∪ and + (whereK + ∅ := ∅ with K a cell tissue).
2 For instance, K := {K ⊂ M} is the family of subsets contained in a given subset M .3 In biology, the vitellus is the energy reserves used by the embryos during its devel-opment.

234 A. Sarr, A. Fronville, and V. Rodin
1. Transitions x &→ x+ d, where d ∈ {1, 2, 3, 4} (action)2. Quiescence x &→ x+ 5 = x (no action)3. Apoptosis x &→ x+ 6 = 6 (programmed cell death)
This injunction (d!, d") is described by the genetic inclusion
x !{x+ d!, x+ d"
}
where the mother cell x
– first migrates from x to x+ d! using the migration action d! ∈ A at a newposition (including x and {6}),
– then divides, giving birth to a cell at position x+d" using the division actiond" ∈ A \ {5} .
The composition of these actions produce a mother-daughter cell pair
{x+ d!, x+ d"}.
Hence the basic behaviours of the mother cell are described by:
1. sterile migration by taking d! ∈ A and d" = 62. stationary division by taking d! = 5 and d" ∈ A3. migrating division by taking d! ∈ A \ {5} and d" ∈ A \ {5}
3.1 Gene and Status Expression
To define the genetic actions that cells must take in each time, we intro-duce genetic processes. They indicate the behaviours of cells over time whiletaking into account constraints they must face. The chosen actions in geneticprocesses is the basis of the morphological dynamic and lead to state changesfor the cells. The possible states we define here are: proliferating, divided andquiescent cells. At the beginning of cycles (duration time for all cells to do anaction), all cells are marked “proliferating”. When a mitosis occurs, the mothercell is marked “divided” while the daughter cell is marked “quiescent”. And onlyproliferating cells are enabled to make mitosis. Since we have associated a colorto each genetic process, cells are coloured with respect to their current ap-plied genetic process (gene expression) and in a color level according to theirstate (status expression). The color level codes are:
– light for the “divided”– intermediate for the “proliferating”– dark form the “quiescent”
Lifts from a genetic process to another make differentiation occurs for bothmother and new created daughter cell. They adopt the associated color of thenew genetic process. This mechanism ensures gene expression in forms.
In order to define the genetic process, we introduce the set of permutationsσ of the set A of genetic actions. A genetic process is the ordered sequenceof actions dσ := {dσ(1), ..., dσ(8)} ∈ A. We denote by G the subset of genetic

Systematic Simulation of Forms’ Co-evolution 235
processes of the actions in A (subset of permutation of six elements). Operatinga genetic process under a given criterion, either for migration or for division,means that the process scans successively x + dσ(1), ..., x + dσ(8) until the fisttime when the criterion is satisfied. Since the empty set ∅ (action 6) belongs toany genetic process, any sequence of operations stops before the six geneticactions of the genetic process.
Before establishing the morphological evolutionary mechanism, we distinguish:
– the calendar (or algorithmic) time n ≥ 0 that identifies the cycles– and at each time n, a process time j = 1, ..., jm are the time process for each
cell. That’s why a cycle lasts as long as there are cells. This is mostly due tothe requirement that at each process time, a cell pair cannot occupy the sameposition.
We can now define the genetic regulon. It’s a map4 associating with eachtriple (n, L, x) the pair (G!(n, L, x), G"(n, L, x)) ∈ G×G of genetic processessatisfying the non-overlapping property.
∀x ∈ L,i is the time process when we first have x+G!(n, L, x)(i) ∈ x ∪ "L,ji is the time process when we first have x+G"(n, L, x)(ji) ∈{x \ x+G!(n, L, x)(i)
}∪ "L,
The first property describes the migration of the mother cell, it can stay at thesame position or move to an unoccupied position in the morphological environ-ment. The second property describes the division and the birth of the daughtercell (it cannot take the position of any existing cell).
In what follows, we will describe the morphological evolutionary mechanism.We first define the local morphological dynamics and then the global morpho-logical dynamics.
3.2 Local Morphological Dynamics
The genetic regulons (G!, G") are assumed either to be given or constructedto respectively regulate or describe viable evolutions. They are the input of thecontrolled morphological inclusion we are about to define. First, the map Hdefined by:
H(G!, G")(n, L, x) := (x+G!(n, L, x)(i), x+G"(n, L, x)(ji))
associates with any pair (G!, G") of genetic processes the mother-daughtercell pair and the transitions transform ϕ(n, L, x;G!, G") of the subset L (rep-resenting the cell tissue at time n).
ϕ(n, L, x;G!, G") := L ∪H(G!, G")(n, L, x)
It transforms L at time n after transitions of the cell x ∈ L.4 A single-valued map for the time, since no other parameters, biological ones, areinvolved at the stage of this study.

236 A. Sarr, A. Fronville, and V. Rodin
3.3 Global Morphological Dynamics
Let Kn ∈ P(x) be a subset constructed, described and coded by an ordered list(x1, . . . , xpKn
). Hence, we construct φ in the following way:Kn being given, we define the sequence Kn(x1) := ϕ(n,Kn, x1;G!, G").
∀p = 2, . . . , pKn , Kn(x1, . . . , xp) := ϕ(n,Kn(x1, . . . , xp−1), xp;G!, G").
Thus we can set φ(n,Kn) := ϕ(Kn(x1, . . . , xpKn);G!, G").
4 Sets of Evolutions
Giving a mathematical formalization of cells’ actions and behaviours, definingthe gene and status expressions and describing the morphological dynamics, weaim now to use this mathematical foundation to describe the possible evolutionsof a form by growth. Indeed, we don’t take into account migration and death.Hence, given an initial form, we construct and generate exhaustively its all evo-lutions at later times. It’s also possible to keep only evolutions passing throughspecific paths while generating sets(see figure 1). To do so, a filter catalogue isset to be applied to some given sets of evolutions.
Fig. 1. Construction of the sets of evolutions over time

Systematic Simulation of Forms’ Co-evolution 237
4.1 Implementation
The program is developed in C++, sets are represented with the library BoostGraph and the algorithm operates a redundancy control in sets to ensure thateither a form or its different geometrical transformations (symmetries, rotationsand translations) are stored only once (reduced a set by 87%). Since the sets’size increase asymptomatically, we have also developed a parallel implementationwith Boost Thread, that allowed us a significant gain in execution time (morethat 30x faster). The program offers a view on all details of each evolution:
– at each time which cell were created,– which one created it– and by which genetic action
Besides, while constructing evolutions, genetic processes are also constructedand thus allow to describe each evolution. Therefore, in the output results viewedwith Scilab, we display the genetic process of each evolution. Furthermore,gene and status expressions can be observed in the evolutions by their colors andcolor levels. In the following figures for example, are represented the evolutionsof a single cell when it becomes 2, 4, 8 and 16 cells.
Fig. 2. Set of evolutions at 2 cells (size 1)
Fig. 3. Set of evolutions at 4 cells (size 4)

238 A. Sarr, A. Fronville, and V. Rodin
Fig. 4. Set of evolutions at 8 cells (size 61)
Fig. 5. One of the evolutions at 16 cells. The size of the entire set is 1029
5 Co-evolution between Forms and Constraints
A Multicellular organism is a complex system which can be defined as a compo-sition of a significant number of elements interacting locally to produce a globalbehaviour. According to Doursat [2], whether inanimate structures or living or-ganisms, all processes of form emergence are instances of decentralized morpho-logical self-organization. When cells evolve, they modify their organism which inits turn impacts their behaviour. This is what biologists mean by co-evolutionwith constraints. Epigenetics considers that this coupling between organism andconstraints can not be ignored in understanding the development of living or-ganisms [8]. These constraints may arise from the environment, the dynamics orthe form itself. To highlight the importance of this interaction between formsand constraints, we’ll study two cases:
Systematic Simulation of Forms’ Co-evolution 239
1. a co-evolution of commuting parts of a same form;2. and form’s evolutions in a very restrictive morphological environment
5.1 A Commutative Growth
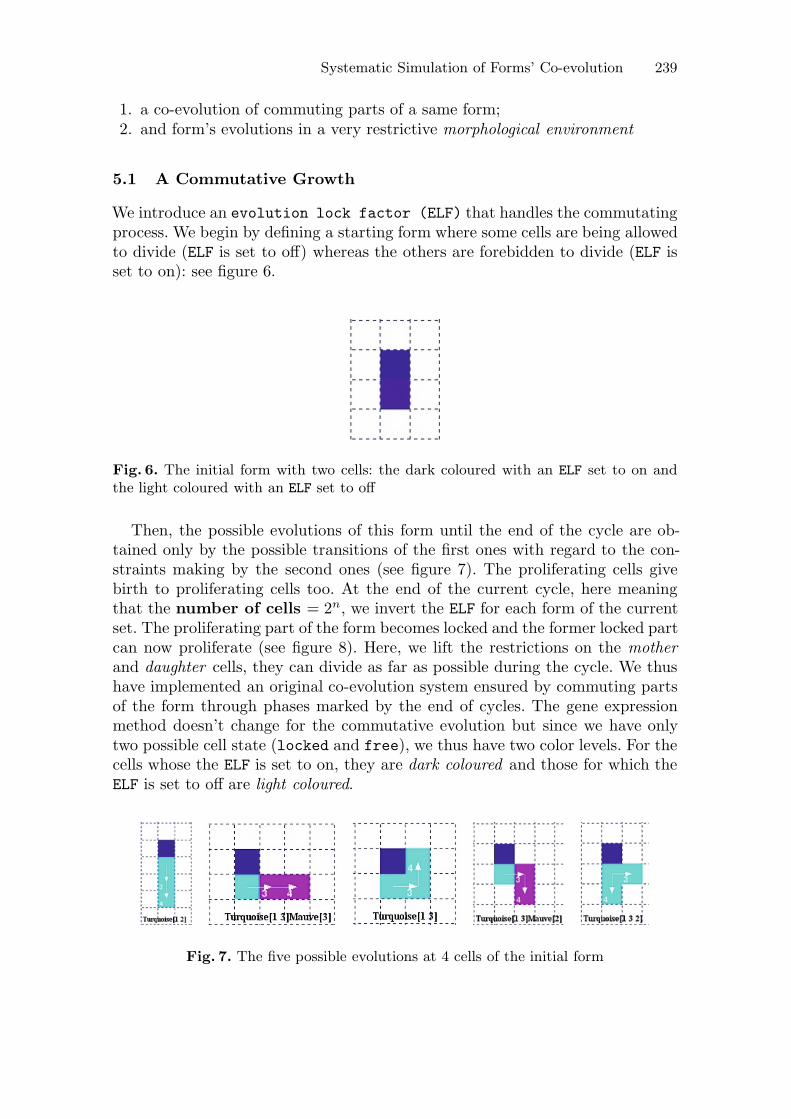
We introduce an evolution lock factor (ELF) that handles the commutatingprocess. We begin by defining a starting form where some cells are being allowedto divide (ELF is set to off) whereas the others are forebidden to divide (ELF isset to on): see figure 6.
Fig. 6. The initial form with two cells: the dark coloured with an ELF set to on andthe light coloured with an ELF set to off
Then, the possible evolutions of this form until the end of the cycle are ob-tained only by the possible transitions of the first ones with regard to the con-straints making by the second ones (see figure 7). The proliferating cells givebirth to proliferating cells too. At the end of the current cycle, here meaningthat the number of cells = 2n, we invert the ELF for each form of the currentset. The proliferating part of the form becomes locked and the former locked partcan now proliferate (see figure 8). Here, we lift the restrictions on the motherand daughter cells, they can divide as far as possible during the cycle. We thushave implemented an original co-evolution system ensured by commuting partsof the form through phases marked by the end of cycles. The gene expressionmethod doesn’t change for the commutative evolution but since we have onlytwo possible cell state (locked and free), we thus have two color levels. For thecells whose the ELF is set to on, they are dark coloured and those for which theELF is set to off are light coloured.
Fig. 7. The five possible evolutions at 4 cells of the initial form

240 A. Sarr, A. Fronville, and V. Rodin
Fig. 8. Here are some possible evolutions at 8 cells of the initial form. They arethe result of the first ELF’s inversion (done in the 4 cells forms). To seek the nextevolutions, the ELF will be inverted in each of the total of 232 forms.
The co-evolution with a constraint of commutative growth has a significantimpact on forms’ development. Indeed, without this constraint, from that ini-tial form, it would be possible to get 369 forms of 8 cells. Therefore, thecommutative evolution reduces by 37% the possibilities.
5.2 Growth in a Restrictive Morphological Environment
The more relevant and more natural way to highlight the influence of constraintson forms’ development is to consider those arising from their morphological en-vironment. Commonly, we make forms grow in a very basic environment that isa 2D Grid. Here, we propose to change the environment a little quirky. Thenwe place a single cell in this restrictive environment to see how many possiblesforms of 8 cells can we get from it (see figure 9).
Fig. 9. Left : a single cell (blue) is placed in the restrictive environment (pink) to grow.Right : Growth of the single cell at 8 cells achieves only 2 forms.

Systematic Simulation of Forms’ Co-evolution 241
Considering the morphological dynamic defined in section 4, from a singlecell, we had 61 forms of 8 cells. So, the choice of a quirky morphological en-vironment has drastically reduced (by 97%) the potential of forms’ development.Besides, in this morphological environment the biggest forms that we can reachare made by 12 cells and are only 5 (see figure 10), we can’t go beyond.
Fig. 10. Growth of the single cell stops at 12 cells. It achieves 5 forms. We can noticethat within these forms, the proliferating cells (with a intermediate color level) can’tno longer divide due to constraints imposed by the environment. Thus growth stops.
6 Discussion
To study and understand the developmental process at the early stage of theembryo when it contains just a few pairs of cells, constraints play a key role. Insupport of this hypothesis, we have presented a mathematical model based onthe viability theory where the morphological dynamics depend on constraints.All cells’ behaviours are able to be described in the model and according to theconstraints, cells can apply one or another action. Due to constraints and appliedactions, cells have respectively status expression and gene expression leading toa differentiation in colors and color levels. With this model, we developed a pro-gram to identify sets of forms that could be obtained from the growth of an initialform over the time. This is a systematic simulation where we use the existingcomputing powers, particularly with a parallel implementation on multicoresprocessor, to explore the entire space of possibles. We have also highlighted theimportance of constraints in cell development. Indeed, by restricting the mor-phological environment or by imposing a commutative evolution to forms, thepossibilities of growth have been significantly reduced.

242 A. Sarr, A. Fronville, and V. Rodin
The interest is to have a view of all possible growth cases of any form. Havinga starting form and a final form, the implementation of a method determiningthe genetic processes that allow to reach the latter from the first can be usedeither for pattern prediction or for forms’ growth controlling.
We aim now to describe the possible evolutions of a form with another cellulardynamic: the programmed cell death or apoptosis. In pattern prediction, this will,for example, provides controls that make a form decreasing. Besides, describingsets of possible evolutions of a given form, both by mitosis and apoptosis, for anypotential disturbances in it, in terms of necrosis or proliferation, we will be ableto provide suitable regulations that allow to compensate them. This would be anautopoietic system that Varela defined in [9]. However, despite the simplicityof the underlying model, algorithms of systematic determination of the space ofpossibles pose significant challenges on Computing. First, representation of thesets of evolutions over time requires huge memory. In addition, running thesesets is often time consuming.
References
1. Aubin, J.P.: Viability theory. Birkhauser (1991)2. Doursat, R.: Organically grown architectures: Creating decentralized, autonomous
systems by embryomorphic engineering. Understanding Complex Systems, pp.167–200 (2008), organic computing
3. Henderson, J., Carter, D.: Mechanical induction in limb morphogenesis: the role ofgrowth-generated strains and pressures. Bone 31(6) (2002)
4. Kauffman, S.A.: The Origins of Order: self-Organization and Selection in Evolution.Oxford University Press, USA (1993)
5. Melani, C., Peyrieras, N., Mikula, K., Zanella, C., Campana, M., Rizzi, B., Veronesi,F., Sarti, A., Lombardot, B., Bourgine, P.: Cells tracking in the live zebrafish embryo.In: Conf. Proc. IEEE Eng. Med. Biol. Soc., vol. 1, pp. 1631–1634 (2007)
6. Muller, G., Newman, S.: Origination of organismal form: beyond the gene in devel-opmental and evolutionary biology. MIT Press (2003)
7. Peyrieras, N.: Morphogenese animale. Morphogenese: L’origine des formes, pp.179–201 (2006)
8. Varela, F.J.: Principles of Biological Autonomy. Developments in Marine Biology.North-Holland (1979)
9. Varela, F.J.: Autonomie et connaissance: essai sur le vivant. Seuil (1989)





























