Embed Size (px)
Citation preview

REVIEWSA PEER REVIEWED FORUM
Morphogen Gradients, Positional Information, andXenopus: Interplay of Theory and ExperimentJEREMY GREEN*Dana Farber Cancer Institute, Harvard Medical School Department of Genetics, Boston, Massachusetts
ABSTRACT The idea of morphogen gradi-ents has long been an important one in develop-mental biology. Studies with amphibians and withXenopus in particular have made significant con-tributions to demonstrating the existence, iden-tity, and mechanisms of action of morphogens. Me-soderm induction and patterning by activin,nodals, bone morphogenetic proteins, and fibro-blast growth factors have been analyzed thor-oughly and reveal recurrent and combinatorialroles for these protein growth factor morphogensand their antagonists. The dynamics of nodal-typesignaling and the intersection of VegT and �-cate-nin intracellular gradients reveal detailed steps inearly long-range patterning. Interpretation of gra-dients requires sophisticated mechanisms forsharpening thresholds, and the activin-Xbra-Gscsystem provides an example of this. The under-standing of growth factor signal transduction haselucidated growth factor morphogen action andprovided tools for dissecting their direct long-range action and distribution. The physical mech-anisms of morphogen gradient establishment arethe focus of new interest at both the experimentaland theoretical level. General themes and emerg-ing trends in morphogen gradient studies are dis-cussed. © 2002 Wiley-Liss, Inc.
Key words: nodal; activin; embryo; patterning
INTRODUCTION
A morphogen may be defined as a substance whosenonuniform distribution in a field of cells differentiallydetermines the fate and phenotype of those cells. Gen-erally, morphogens are thought of as being distributedin a gradient and as carrying positional informationrather than just an “on” or “off” signal to cells. A gradeddistribution of a morphogen provides both polarity (theslope of the gradient) and a scalar value (the concen-tration itself) that can be interpreted by cells accordingto threshold values to provide a coordinate system. Theidea that morphogen gradients provide a coordinatesystem for guiding cell fate during embryonic develop-ment is over 100 years old (Wolpert, 1996; Lawrenceand Struhl, 1996). It was first proposed by Thomas
Hunt Morgan (later famous for pioneering Drosophilagenetics) during his early studies on amphibian andplanarian development and regeneration (quoted inLawrence, 1988). Until relatively recently, morphogenswere entirely theoretical entities: nice ideas to explainsome features of development and regeneration butstill in the realm of mathematical theory. How couldthe morphogen concept survive so long without directexperimental verification? Part of the answer is surelyits formal beauty and explanatory power. For example,when a morphogen provides positional information tocells, it provides both robustness and flexibility duringdevelopment and in evolution. Robustness comes fromthe ability of a morphogen gradient to coordinate long-range patterning and establish itself, through feedbackmechanisms, despite perturbations. Flexibility is a con-sequence of the separation between positional informa-tion (the morphogen gradient itself) and the interpre-tation of that information by cells. For example, cellscan use the same molecular coordinate system (BMPsand their inhibitors—see below) to determine their dor-soventral position and fate in both flies and verte-brates, even though the structures that emerge arevery different. Many of these aspects of morphogenaction were described in detail by Wolpert (1969, 1971).
Thus, the morphogen gradient idea was (and is) abeautiful and neat solution to the problem of embryonicpatterning in theory, but the existence and identity ofmorphogens had to be proved in vivo. As has emergedover the past 15 years of developmental biology, em-bryos of diverse species do in fact use morphogen gra-dients repeatedly in the development of the individualembryo. There have been several reviews that providean overview of known morphogen gradients, especiallyin Drosophila development (Lawrence and Struhl,
Grant sponsor: National Institutes of Health; Grant sponsor: Na-tional Science Foundation; Grant sponsor: Claudia Adams Barr Pro-gram.
*Correspondence to: Jeremy Green, Dana Farber Cancer Institute,Harvard Medical School Department of Genetics, 44 Binney Street,Boston, MA 02115. E-mail: [email protected]
Received 5 August 2002; Accepted 21 August 2002DOI 10.1002/dvdy.10170
DEVELOPMENTAL DYNAMICS 225:392–408 (2002)
© 2002 WILEY-LISS, INC.

1996; Dale, 1997; Strigini and Cohen, 1999; Tickle,1999; Podos and Ferguson, 1999; Day and Lawrence,2000; Tabata, 2001; Gurdon and Bourillot, 2001; Law-rence, 2001a). This review will focus on morphogengradients in vertebrates with an emphasis on howstudies using Xenopus have provided insight into boththe existence of morphogens and their detailed mech-anisms of action. The diversity of contexts in whichmorphogens have been shown to operate is now solarge that, inevitably, some will be omitted from thisreview, and for that the author apologizes.
XENOPUS AND GROWTH FACTORMORPHOGENS
Between its earliest formulation by Morgan and re-cent times, the idea of morphogens in development hashad a roller-coaster history (see Historical Note). Bythe mid-1980s, Drosophila genetics and molecular bi-ology was demonstrating that intracellular proteins—specifically transcription factors Bicoid and Hunch-back—were bona fide morphogens. In vertebrates,retinoic acid appeared to act as a morphogen in thelimb and the hindbrain. However, most of the evidencefor morphogen action by retinoic acid was indirect andis now known not to be true, at least for the limb (seebelow). Thus, the evidence for the existence and themolecular nature of morphogens in vertebrates was notclear-cut. A breakthrough came by means of Xenopusand the study of a process simpler than morphogenaction, namely mesoderm induction.
Spemann had originally developed the concept ofinduction to describe the process whereby the develop-ing eye caused overlying ectoderm to become a lens(Spemann, 1938). Induction is a simple process in thesense that there is a signaling tissue and a respondingtissue, and the latter has only two developmental choices,i.e., to be induced or not (Wolpert, 1989). Nieuwkoop,a pioneer in Xenopus embryology, conducted an impor-tant series of experiments in the early 1970s in whichhe showed that mesoderm (prospective notochord,muscle, and blood) could arise by means of inductivesignals. He made “sandwiches” of vegetal pole explants(prospective endoderm) and animal caps (prospectiveectoderm). Explanted alone, animal caps become epi-dermis and express all the molecular as well as mor-phologic features of skin. Nieuwkoop showed that thevegetal poles induced the animal caps to become mus-cle instead of skin (Nieuwkoop, 1969). The induction ofthe animal cap to become mesoderm (the “animal capassay”; see Green, 1999) became the basis for assayingcompounds for mesoderm-inducing activity. Proteingrowth factors in the fibroblast growth factor (FGF)family and transforming growth factor-beta (TGF-�)superfamily were identified as potent mesoderm-induc-ing factors in Xenopus (Slack et al., 1987; Smith, 1987;Kimelman and Kirschner, 1987; Smith et al., 1988). Amessenger RNA encoding Vg1, a member of the TGF-�superfamily, was the first example of an asymmetri-cally localized embryonic RNA (Weeks and Melton,1987), adding momentum to the idea that growth fac-
tors might be morphogens. (Ironically, despite manyefforts, no morphogen function for Vg1 has yet beenfound in the frog, although it may have a role in left–right patterning [Hyatt et al., 1996] and an apparentorthologue may play a role in axial patterning in thechick [Seleiro et al., 1996; Shah et al., 1997]). TheTGF-� superfamily member that emerged first as amesoderm-inducing factor par excellence was activin(Smith et al., 1990; although this is probably not theendogenous inducer-morphogen; see below). WhereasFGF mimicked the ventroposterior-inducing activity,activin could induce dorsal mesoderm (Symes andSmith, 1987; Green et al., 1990). Strikingly, activincould induce not just dorsal mesodermal tissue typesbut also characteristic cell movements and gross elon-gation of explants mirroring the convergent extensionof endogenous dorsal mesoderm (Symes and Smith,1987).
Dose–response experiments with FGF and activinindicated that these might be morphogens in that dif-ferent doses induced different cell types (Slack et al.,1987; Green and Smith, 1990). The malleability of theXenopus embryo provided the means to test this possi-bility more precisely. Blastula cells can be dissociatedby incubating explants in calcium-free medium. Bydissociating cells, precise control of activin concentra-tion was achieved and this control revealed exactly themultiple threshold-delimited responses predicted bymorphogen theory (Green and Smith, 1990, 1991;Green et al., 1992). Furthermore, the low-to-high se-quence of induced markers corresponded to the ventro-posterior-to-dorsoanterior sequence of tissues observedin vivo (Green et al., 1992).
Two types of questions came out of the early studiesin Xenopus with activin and FGF. One type of questionasked whether growth factors in general and activinand FGF in particular really act in the embryo, and, ifso, when and where? The other type of question askedhow could such morphogens work with respect to theiractions in tissues and their interpretation by respond-ing cells. The progress made in answering these twotypes of question is the subject of the rest of this re-view.
EARLY XENOPUS MORPHOGENS IN VIVO:NOT ACTIVIN BUT NODALS?
It turns out that, although activin was and still is anexcellent model for morphogen action, it may not beimportant in early in vertebrate patterning. However,at least two related growth factors, the Nodals and thebone morphogenetic proteins (BMPs), certainly are.Evidence for this finding emerged gradually, and thisgradualness illustrates some of the difficulties of get-ting definitive answers to the questions of where andwhen morphogens act in vivo. One of the first steps wasthe cloning of “activin receptors”—receptors that cer-tainly bind and allow activin signaling but may have abroader range of TGF-� superfamily ligands. Thesewere found to be transmembrane serine-threonine ki-nases and, like TGF-� receptors, function as het-
393MORPHOGEN GRADIENTS AND XENOPUS

erodimers of a Type I and Type II receptor protein(reviewed in Whitman, 2001; Attisano and Wrana,2002). Kinase-dead mutant versions were made andused as dominant negative reagents. The loss-of-func-tion phenotypes produced by expression of activinreceptor mutants included lack of mesoderm and con-version of prospective endodermal cells to a neurecto-dermal fate (Hemmati-Brivanlou and Melton, 1992,1994). This finding was among the first indications thatactivin-like activity is not just sufficient but also nec-essary for mesodermal and endodermal (“mesendoder-mal”) development and that, apart from a potential rolein dorsoventrolateral patterning, activin-like activity isnecessary for germ-layer patterning of the animal–vegetal axis. However, the interpretation of such ex-periments was muddied by the apparent cross-specific-ity of mutant receptors and the ability of different TypeI and Type II receptors to hetrodimerize with one an-other (Chang et al., 1997; New et al., 1997). Thus, itwas not possible to say that the mutants blocked en-dogenous activin signaling as such, but only some ac-tivin-like signal. Furthermore, the absence of activinRNA in the early embryo and the failure of specificactivin-blocking reagents to affect normal development(Thomsen et al., 1990; Schulte-Merker et al., 1994)indicated that the endogenous player might not beactivin itself.
Mouse and zebrafish genetics have given the bestevidence for the existence in vivo of an essential dorsal-specific activin-like activity, and these genes have beenidentified as the nodal gene in mouse and the nodal-related genes cyclops and squint in zebrafish (reviewedby Whitman, 2001). The nodal proteins are, like ac-tivin, members of the TGF-� superfamily. Mouse nodalmutants lack a primitive streak and all axial struc-tures, whereas zebrafish mutants have severely dis-rupted axes but retain overall axial organization andsome somites. Xenopus phenocopies of nodal mutantshave been achieved by expression of nodal antagonists.These phenocopies include Lefty2/antivin (Tane-gashima et al., 2000), Cerberus-short (Agius et al.,2000), and cm-Xnr2, a dominant inhibitory form ofXenopus nodal-related-2 (Osada and Wright, 1999). Aswith zebrafish loss of nodal function, the treated Xeno-pus embryos have disrupted but not abolished axes.The situation is complicated in Xenopus by the exis-tence of several nodal-related genes with the appropri-ate sites of expression and inducing activity, namelyXnrs 1, 2, 4, 5, and 6, and the protein Derriere (Jones etal., 1995; Smith et al., 1995; Lustig et al., 1996; Josephand Melton, 1997; Osada and Wright, 1999; Agius etal., 2000; Takahashi et al., 2000; Onuma et al., 2002).The differences between the mouse on the one handand the fish and frog embryos on the other could also beto do with embryonic-extraembryonic signaling in themouse or, perhaps, to maternally stored factors thatallow residual signaling. It may be relevant that, inXenopus, the nodals 1, 2, and 4, and Derriere are directearly targets of the maternally encoded T-box tran-scription factor VegT (Kofron et al., 1999) and that
VegT (also known as BraT, Xombi) and the closelyrelated Antipodean protein are distributed in a vege-tal-to-animal gradient in the egg (see Fig. 3; reviewedin Kavka and Green, 1997). VegT depletion experi-ments have shown that it is required for the formationof endoderm and the positioning of the other germlayers in the animal-vegetal axis (Zhang et al., 1998).Because there is evidence for both indirect non–cell-autonomous action of VegT by means of nodals anddirect, cell autonomous action on mesodermal markergenes and that this direct action is dose-dependent(Clements et al., 1999; Kavka and Green, 2000), it maybe that VegT serves as an intracellular morphogen, notunlike bicoid in Drosophila (discussed in Kavka andGreen, 1997).
If nodals are required for normal axial patterning,can it be said that they are acting in a graded morpho-gen-like way in vivo and, if so, when and where? Doseresponsiveness was demonstrated for activin in ex-planted cells, as described above, but there is compa-rable evidence for endogenously expressed gradednodal signals in vivo. Ectopic expression studies inzebrafish show that Squint (though not Cyclops) caninduce different cell types in a dose-responsive mannerin a dorsal-to-ventrolateral sequence (Gritsman et al.,2000). Furthermore, progressive attenuation of thissignal in mutants lacking a nodal coreceptor (One-Eyed Pinhead/Cripto) progressively abolishes tissuesin the predicted order (Gritsman et al., 2000). In Xeno-pus, titrating expression of exogenous Cerberus-short,a nodal antagonist, produces progressive ventrolateral-to-dorsal attenuation of the mesodermal genebrachyury (Agius et al., 2000), and this graded attenu-ation does suggest a functional dorsoventral gradientof nodal signaling.
Despite the above results, evidence against a strictlydorsoventral gradient comes from experiments by Choand colleagues in which they identified an element inthe promoter of the Organizer (dorsal) -specific genegoosecoid that is responsive to activin-like (or nodal-like) stimuli. When this element was coupled to a re-porter gene and injected into embryos, no difference inreporter activity between dorsal and ventral sides ofthe embryo could be observed (Watabe et al., 1995).There was, however, a vegetal-to-animal difference,consistent with a gradient of induction by maternalVegT acting by means of nodals. One explanation forthis finding might be that the dorsoventral difference istoo subtle or too dynamic to be detectable by the re-porter assay. Evidence supporting the dynamic natureof nodal signaling and perhaps the clearest demonstra-tion of direct, graded action of an endogenous nodal-type signal in vivo comes from analysis of the intracel-lular transduction of nodal signals. Like other TGF-�superfamily factors, nodal signals by means of recep-tors that phosphorylate intracellular Smad proteins(Smad2 in the case of both activin and nodal). High-resolution in situ analysis of nodal signaling was facil-itated by the availability of antibodies that specificallydetect the activated (phosphorylated) form of Smad2
394 GREEN

(Faure et al., 2000; Lee et al., 2001) (Fig. 1). Thisanalysis showed that there was indeed a vegetal-to-animal gradation of phospho-Smad2. In the dorsoven-tral axis, a dynamic activation pattern was observedsuch that signaling was initiated dorsally and pro-gressed in a dorsal-to-ventral sequence, ultimately fad-ing in the same sequence as though a wave of activa-tion progresses across the embryo followed by a wave ofdown-regulation or negative feedback. In zebrafish em-bryos, a similar dynamic wave of signaling was ob-served in the expression of the nodal-responsivebikhari gene (Vogel and Gerster, 1999). These dataconstitute good evidence for at least one “well-behaved”vegetal-to-animal nodal gradient and possibly a seconddorsal-to-ventral functional gradient or traveling frontacting a little later and by means of some more dy-namic mechanism. Note that even a traveling front ofactivation could, if cells can integrate activation overtime, be interpreted as a graded signal (see Fig. 2 andGradient Formation below). However, it should benoted that nodals are eventually expressed throughoutthe dorsoventral axis, with the expression (at the RNAlevel) progressing from the dorsal to the ventral side ofthe embryo (Jones et al., 1995; Joseph and Melton,1997; Osada and Wright, 1999; Agius et al., 2000; Ta-kahashi et al., 2000; Onuma et al., 2002). Thus, theywould only need to act over very few cell diameters topattern the entire mesoderm and endoderm. It is,therefore, possible that nodals really act virtually cell-autonomously and that their graded action—as mea-sured by progressive expression and progressive Smadphosphorylation—is a function of a prior gradient ofsomething else. However, at the limits of its expressiondomain, it seems likely that nodal protein would act asa morphogen in vivo even if it is over a short rangeencompassing a few cell diameters. Short range of ac-tion does not preclude morphogen activity. For exam-ple, a Wingless (Wnt) morphogen gradient is required
for the fine-scale patterning of Drosophila segments(Lawrence and Struhl, 1996). Detection of such a gra-dient in Xenopus or zebrafish is simply beyond theresolution of analyses performed to date. In mouseembryos, the situation may be even more complicatedby the fact that Organizer functions are split betweenthe embryonic node and the anterior visceral endodermand that both of these operate in a rapidly growing andfluid cellular context (discussed in Whitman, 2001).
Although nodals may be more important than activinas dorsalizing factors in early development, activin isstill used as a model morphogen for experiments ana-lyzing diffusion of morphogens through tissues. Ironi-cally, this is despite evidence that activin and nodalsdiffer greatly in this respect (see Establishment of Mor-phogen Gradients below) and perhaps in other ways.
MORPHOGENS IN VIVO. PART II:FGFS AND BMPS
Compared with the TGF-� superfamily members,the FGFs have not been as intensively tested as classicmorphogens. For example, in Xenopus, it is more oftenconsidered a mesodermal inducer or competence factorthat provides a positive feedback loop for maintenanceof the nodal-induced mesodermal gene brachyury (Cor-nell and Kimelman, 1994a; LaBonne and Whitman,1994; Cornell et al., 1995; Schulte-Merker and Smith,1995). However, FGF may also be a morphogen. Dose–response experiments together with activin suggesteda model in which FGF acts in an animal-to-vegetal (i.e.,
Fig. 1. In situ detection of Nodal/activin-type signals as phospho-Smad2 (based on Lee et al., 2001). Sagittal sections of late blastula andearly gastrula Xenopus embryos are depicted. Black dots representnuclei stained for phospho-Smad2–specific antibodies. Below each dia-gram is an impression of the phospho-Smad2 distribution based on theoriginal figures. The staining progresses in a somewhat graded travelingfront from the dorsoanterior side (D, right) to the ventroposterior side (V,left) and then disappears in the same sequence. At stages 9.5 and 9.75,there is a vegetal-to-animal staining gradient. Staining of horizontal sec-tions (not shown) indicates that between stages 9.5 and 10�, the stainingappears to be graded. Weak staining of the deep yolky vegetal core maybe a staining artifact. Note that Smad1 phosphorylation, indicative ofBMP signaling, is initially fairly uniform but clears progressively from thedorsal side (Faure et al., 2000; Lee et al., 2001), presumably because ofnodal induction of BMP inhibitors (see text).
Fig. 2. Static gradients and traveling fronts of morphogen both canprovide positional information. A field of cells can gradually fill withmorphogen (M) over time (time points t1, t2, and t3) but in different ways.a: With an appropriate sink for morphogen, a gradient can form and bemaintained. A sink can be destruction of the morphogen or its seques-tration, for example by endocytosis. b: However, if the sink can be easilysaturated, morphogen builds up and progresses through the field of cellsas a “traveling front.” c: However, if cells can integrate the morphogensignal over time, for example by accumulating some intracellular productsuch as a gene transcript, then a gradient of positional value can still beestablished. If the traveling front of morphogen is followed by a travelingfront of its removal or inhibition, as may be the case with Xenopusnodal-type signaling (Fig. 1), then the positional value will become pro-gressively leveled off (c, t3).
395MORPHOGEN GRADIENTS AND XENOPUS

approximately posterior-to-anterior) gradient in themesoderm (Green et al., 1992). FGF does seem to beable to diffuse over several cell diameters (Christenand Slack, 1999), and it has been proposed to be amorphogen in early posterior patterning of the nervoussystem in Xenopus: embryonic FGF (eFGF) may actduring gastrulation stages to induce expression of pos-terior Hox genes and posterior neural markers (Xu etal., 1997; Isaacs et al., 1998; Pownall et al., 1998;Holowacz and Sokol, 1999). However, more detailedanalysis of FGF’s potential morphogen activity is diffi-cult because the rapid increase in anatomic complexitymakes quantitation problematic. It would be interest-ing to see an attempt to quantify FGF activation inmore detail, perhaps by higher-resolution in situ stud-ies with the anti–phospho-ERK antibodies that revealFGF signal transduction (Christen and Slack, 1999),just as the phospho-Smad2 antibodies have done fornodal signaling.
In the vertebrate limb, there is has been some sug-gestion that FGF might act as a proximodistal morpho-gen (Papageorgiou and Almirantis, 1996; Vargesson etal., 2001). Hitherto, proximodistal patterning has beenexplained by the so-called progress zone model. Thismodel is a non–morphogen-based model in which FGFmerely maintains cellular proliferation in the distalmesenchyme, and cells are patterned according to thetime they leave this proliferative zone (Wolpert, 1999;Ng et al., 1999). However, recently this model has beenchallenged by evidence that pattern specification oc-curs through a more static mechanism in the early limbbud and that FGF in the apical ectodermal ridge playsthe role of anti-apoptosis factor (Dudley et al., 2002).This model poses two gradient-related questions: whatis the molecule that specifies the domains of the earlylimb bud, and what mechanism provides the progres-sion of stable fate determination that is observed?
Xenopus has played a major role in the characteriza-tion of another morphogen gradient system, namelythat of BMPs. BMPs (with the exception of BMP1; seebelow) are members of the TGF-� superfamily. BMPswere first implicated in morphogenetic gradients bystudies of a Drosophila BMP orthologue, Decapentaple-gic (Dpp), in which a dorsal-to-ventral dose-dependentcell type specification was demonstrated (Ferguson andAnderson, 1992). When BMP4 was overexpressed inXenopus embryos, the result was a striking ventraliza-tion and abolition of the Organizer (Jones et al., 1992;Dale et al., 1992). Expression of a dominant-negativemutated form of BMP receptor correspondingly blockedventral development and induced partial ectopic axeswhen the RNA was injected on the ventral side (Graffet al., 1994). Thus, it was clear that BMPs act duringgastrula stages to limit the dorsalizing effects of theOrganizer. BMP2, BMP4, and BMP7 are all expressedappropriately in Xenopus (Hemmati-Brivanlou andThomsen, 1995; Nishimatsu and Thomsen, 1998) andBMP2 (swirl) and BMP7 (snailhouse) mutants of ze-brafish are dorsalized (Dick et al., 2000; Schmid et al.,2000).
When the dorsalizing molecules noggin and chordinwere cloned, it soon became clear that these were BMPantagonists. The noggin and chordin stories each re-veal some of the great strengths of Xenopus as anexperimental system. Noggin was cloned by injectingpools of in vitro–transcribed RNAs into early embryoand simply observing them for signs of ectopic axisformation (Smith and Harland, 1992). Positive poolswere then subdivided and re-assayed until single RNAscontaining the axis-inducing activity were isolated.This approach has led to the cloning of several inter-esting genes (e.g., Smith et al., 1995; Lemaire et al.,1995; Lustig et al., 1997; Laurent et al., 1997), and it ishard to imagine a similar approach being so effective inanother species. Chordin was cloned as a dorsally en-riched gene, one of a series of genes cloned by theDeRobertis group based on their anatomic restrictionof their enrichment in embryos manipulated to be dor-salized (Sasai et al., 1994). Isolation of sufficient ma-terial from specific anatomic regions was facilitated bythe large size of Xenopus embryos, whereas the dorsal-izing manipulations were only possible because of theaccessibility of the early embryo and the large back-ground knowledge of how such manipulations are car-ried out (Blumberg et al., 1991). Further biochemicalstudies that showed that both noggin and chordin pro-teins could bind BMP4 directly to inhibit its activity(Zimmerman et al., 1996; Piccolo et al., 1996).
The antagonism between BMP and chordin is a re-markable example of conservation of embryonic pat-terning mechanisms. In Drosophila, the BMP homo-logue Dpp is antagonized by a chordin homologue,short gastrulation (sog; Marques et al., 1997). The pro-teins are sufficiently conserved that they retain dorsal-izing and ventralizing activities in cross-species ex-pression experiments (Francois and Bier, 1995; Sasaiet al., 1995; Schmidt et al., 1995; Holley et al., 1995,1996). Most exciting, from an evolutionary point ofview, is that, whereas BMP is ventral and chordindorsal, in flies, it is the other way around: Dpp is dorsaland sog is ventral. This finding coincides with severalanatomic features, most prominently the nerve cordthat is ventral in flies and dorsal in vertebrates. Thisevidence confirms an old idea that vertebrates andarthropods share a common “bilaterian” ancestor thatwas inverted in one of these during evolution (seeJones and Smith, 1995; DeRobertis and Sasai, 1996;Ferguson, 1996). The conservation of mechanism goesseveral steps further (reviewed in Dale and Jones,1999; Niehrs et al., 2000; Ray and Wharton, 2001). Forexample, BMP1 and its Drosophila counterpart,Tolloid, unlike the other BMPs, are proteases that bothfunction to cleave and thereby inactivate the BMP/Dppantagonists chordin/sog (Marques et al., 1997; Bladeret al., 1997; Goodman et al., 1998; Wardle et al., 1999).Twisted gastrulation is a direct-binding BMP antago-nist/modulator that is also conserved between flies andvertebrates (Oelgeschlager et al., 2000; Chang et al.,2001; Ross et al., 2001; Scott et al., 2001). Thus, thedorsoventral antagonism of early Drosophila and Xe-
396 GREEN

nopus embryos is a sophisticated balancing of forces. Itis worth noting that the robustness and resistance toperturbation of gradients must depend on feedback andantagonistic mechanisms, and it should, therefore, beno surprise that the early embryo regulates these po-tent morphogens by means of multiple levels of antag-onism. In addition, antagonistic morphogens are ablein principle to provide sharper response thresholds aspart of a “winner-takes-all” mechanism (see Interpre-tation of Gradients, below). The existence of additionalinhibitors, namely the proteins follistatin, cerberus,and gremlin, reinforces this notion.
What is the specific evidence that the BMP system isa morphogen in vivo? The best evidence that BMP4itself fits the morphogen definition is the dose–re-sponse experiments carried out by using Xenopus me-soderm and ectodermal markers as assays (Wilson andHemmati-Brivanlou, 1995; Dosch et al., 1997; Wilsonet al., 1997; Nishimatsu and Thomsen, 1998; Marom etal., 1999). A slightly later role in neuronal patterninghas also been proposed (Chitnis, 1999). However, thereis some evidence that a BMP4 activity gradient isachieved not by graded distribution of BMP4 as suchbut by an inversely graded distribution of its inhibitors,noggin and chordin (Jones and Smith, 1998; Blitz et al.,2000; and see Establishment of Gradients, below).
MORPHOGENS IN VIVO. PART III:INTERACTING MORPHOGENS
It is worth re-emphasizing that the TGF-� superfam-ily morphogens are not the only ones. The reader of thisreview is urged to refer to excellent accounts of someother morphogen systems, particularly in the Drosoph-ila segmentation and imaginal disc patterning pro-cesses (Lawrence and Struhl, 1996; Day and Lawrence,2000; Vervoort, 2000; Lawrence, 2001b) and in thedorsoventral patterning of the vertebrate neural tube(Lee and Jessell, 1999; Tabata, 2001; Marti and Bovo-lenta, 2002). However, there are perhaps two simplify-ing generalizations that can help in appreciating thecomplexity of morphogen-dependent patterning. One isthat, although there are many proven or likely morpho-gen molecules, in fact there is a relatively small cast of“usual suspects,” and the other is that these few oftenact in combination, either orthogonally, in opposition,or quick succession. Thus, Gerhart (1999) has writtenof five major pathways that are highly conserved inevolution and repeatedly used in early developmentand another five common in later development andorganogenesis. The early pathways are the Wnt-�-cate-nin pathway, receptor serine-threonine kinase (TGF-�superfamily) pathways, receptor tyrosine kinase (epi-dermal growth factor [EGF], FGF, etc.) pathways, theHedgehog pathway, and the Notch/Delta pathway.With the possible exception of Notch/Delta signaling,which generally governs neighboring cell inhibitorysignaling, each of these pathways can operate bymeans of a distributed ligand acting as a morphogen.Furthermore, these pathways often operate in pairs(Cornell and Kimelman, 1994b), some of which may be
acting as orthogonal gradients. For example, the inter-section (i.e., spatial overlap) of Wnt-�-catenin–relatedsignals and TGF-� (nodal) signals triggers the localinitiation of the Xenopus Organizer (Watabe et al.,1995; Crease et al., 1998) (Fig. 3). Specifically, theOrganizer is generated by the interaction of at leastone prior “south–north” gradient (that of VegT) withlocal concentration of (as yet unidentified) “dorsal de-terminants” on one side of the early embryo (Fig. 3).The VegT gradient induces nodals, whereas the dorsaldeterminants act like a Wnt signal in that they lead tolocal enrichment of intracellular �-catenin. An Orga-nizer forms where an intermediate concentration ofnodal activity coincides with high levels of �-catenin. Itis possible to interpret this spatial restriction as a pureand simple combinatorial mechanism without a role forgradients. However, there are some indications that amore sophisticated mechanism of orthogonal gradientsmay be at work. The Organizer is quite structured evenat early stages (Vodicka and Gerhart, 1995; Zoltewiczand Gerhart, 1997), and its outside-to-inside structurehas functional significance (Shih and Keller, 1992).One might, therefore, speculate that the dorsal deter-minants (or the �-catenin that they enrich) constituteor initiate an outside-to-inside morphogen gradient(Fig. 3). The gradients could be purely intracellular(i.e., VegT and �-catenin transcription factors) and/orbe expressed as gradients of extracellular growth factor(nodal) concentration. Such orthogonal gradientsmight consist of a TGF-� (nodal) and FGF (Green et al.,1992). Of course, orthogonality is not a rigid rule. Forexample, Wnt and TGF-� (i.e., BMP) act in opposinggradients to pattern the neural tube (Lee and Jessell,1999; Tabata, 2001; Marti and Bovolenta, 2002). None-theless, the idea of orthogonal gradients providingtruly Cartesian coordinates to cells is a very attractiveone. A particularly beautiful example of the successionand combinatorial action of orthogonally acting pat-terning molecules is that of mesodermal patterning inDrosophila (Carmena et al., 1998). Loss-of-functionand targeted misexpression studies with Wingless(Wnt) and Dpp (BMP) show that orthogonal stripes ofthese factors determine muscle cell progenitors. Withinthese progenitor clusters, further patterning by FGFand EGF takes place. The precise muscle types inducedare dependent on the local concentration of EGF (Buffet al., 1998), suggesting that gradients and not justbinary combinations of growth factors may be impor-tant. Much effort is now being made to establish mo-lecular anatomy of embryos at the level of transcrip-tional targets of these pathways and the role anddistributions of these ligands.
The validity of the morphogen concept leads directlyto two further big questions. One is how morphogengradients are interpreted and the other is how morpho-gens get to be properly distributed in the first place.These are the subjects of the following two sections ofthis review.
397MORPHOGEN GRADIENTS AND XENOPUS
INTERPRETATION OF GRADIENTS. PART I:THRESHOLD RESPONSES AND
SHARPENING MECHANISMS
The conversion of an analogue input (a morphogenconcentration) into a digital output (i.e., one cell fatevs. another) is, intuitively, a rather unusual process inthe sense that, as biologists, we are more used to rela-tively gradual changes in space and time. In practice, ofcourse, threshold responses are common (Gurdon et al.,1998; Gurdon and Bourillot, 2001). To give distinct celltypes, morphogen interpretation relies on the sharp-ness of threshold responses to cellular stimuli. Thisfinding suggests some kind of cooperativity leading to asteep sigmoid response in the interpreting cell (Lewiset al., 1977; Slack, 1993). Over time, an initial steepsigmoid response can be sharpened still further bypositive feedback, cross-inhibition (Fig. 4), and physi-cal mechanisms such as cell sorting.
An elegant example of simple cooperativity in devel-opment is the hunchback promoter in Drosophila.Here, the direct cooperative binding of multiple Bicoidmolecules to the target hunchback promoter occurs inthe early embryonic syncytium (Driever et al., 1989;Driever and Nusslein-Volhard, 1989). The morphogengradient, thus, is unencumbered by the need for cellsurface receptors and signal transduction molecules.Another example, which works orthogonally to Bicoidand hunchback, is the regulation of dorsal by basic
helix-loop-helix transcription factors (Jiang and Le-vine, 1993). An important factor in the cooperativity ofsuch promoter-based switches seems to be the presenceof competing inhibitors (Rossi et al., 2000). Detailedappreciation of this type of molecular switch comesfrom the much earlier studies of lambda phage promot-ers and switching of transcription by protein action onthe chromosome is well understood (Ptashne, 1992). Atthe other extreme are cooperative interactions occur-ring with receptors and associated cofactors and trans-duction molecules at the cell surface. Examples of this
Fig. 3. Intersecting morphogens that pattern the early Xenopus em-bryo. The Xenopus egg contains dorsal determinants at the vegetal pole(V) and a vegetal-to-animal (A) gradient of the VegT transcription factor.Sperm entry (Sp) triggers cortical rotation (CR) and active transport thatspreads the dorsal determinants (Dd) from the vegetal pole to the sideopposite the sperm entry point. The VegT gradient specifies the threegerm layers, acting as an intracellular morphogen, whereas the dorsaldeterminants trigger local accumulation of �-catenin. The Organizerarises at the intersection of the dorsal determinants and the mesodermaland endomesodermal levels of VegT. Speculatively, different parts of theOrganizer may arise from different combinations of VegT and Dd. Thus,{Dd-high � VegT-medium}� trunk Organizer, {Dd-high � VegT-medium-high}� bottle cells, and {Dd-medium � VegT-medium-high} � headmesendoderm/Nieuwkoop center. Note that the idea of threshold levelsfor Dd and of a vegetal threshold of VegT delimiting the vegetal extent ofthe Organizer are this author’s speculation (denoted by “?”) based on thefinding that Dd at the vegetal pole induces Organizer genes only tran-siently (Brannon and Kimelman, 1996; Medina et al., 1997). For othersources, see text. VP, ventroposterior; DA, dorsoposterior.
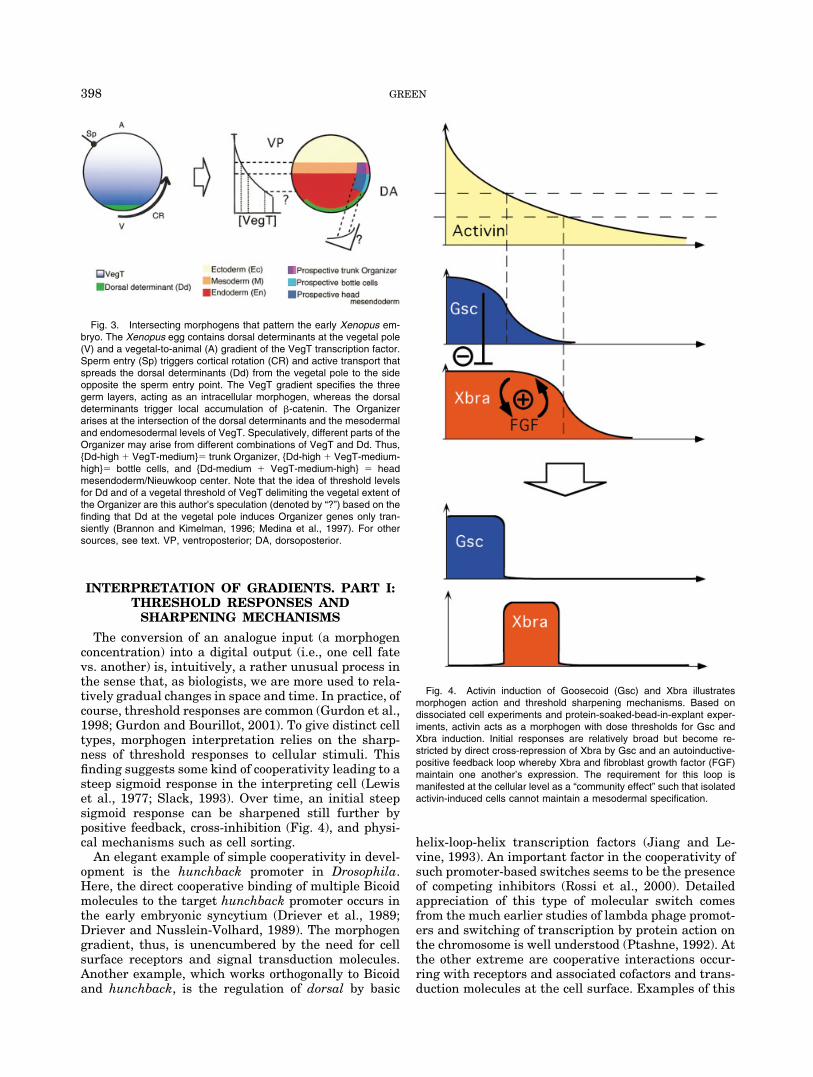
Fig. 4. Activin induction of Goosecoid (Gsc) and Xbra illustratesmorphogen action and threshold sharpening mechanisms. Based ondissociated cell experiments and protein-soaked-bead-in-explant exper-iments, activin acts as a morphogen with dose thresholds for Gsc andXbra induction. Initial responses are relatively broad but become re-stricted by direct cross-repression of Xbra by Gsc and an autoinductive-positive feedback loop whereby Xbra and fibroblast growth factor (FGF)maintain one another’s expression. The requirement for this loop ismanifested at the cellular level as a “community effect” such that isolatedactivin-induced cells cannot maintain a mesodermal specification.
398 GREEN

are known in the responses of immune cells to differentlevels of antigen (Malissen, 1998). It is unclear whetherthe lack of examples of such sophisticated cell-surfaceco-cooperativity in embryology is because embryosrarely use this mechanism or because it is more com-plex and, thus, harder to study. In between, an appar-ently simple positive feedback mechanism within asignal transduction kinase cascade is sufficient to pro-vide a nearly all-or-none switch, as in the case ofMAPK activation by progesterone in the Xenopus oo-cyte (Ferrell and Machleder, 1998). In that case, thefeedback is provided by MAPK phosphorylation of themore upstream transducer, Mos, which leads to in-creased Mos protein accumulation.
In Xenopus, the detailed analysis of the regulation ofthe Xenopus brachyury (Xbra) mesoderm gene has re-vealed that both positive and negative feedback con-tribute to its threshold response (Fig. 4). With increas-ing doses of activin, transcription of first Xbra and thengoosecoid are induced. The initial dose window of Xbrainduction is bounded by initially relatively “fuzzy” dosethresholds, but it progressively sharpens with timeover a few hours (Green et al., 1994; Gurdon et al.,1999). Two mechanisms have been identified that con-tribute to this sharpening. One of these mechanisms iscell contact-dependent and consists of a positive feed-back loop between Xbra expression and FGF signaling.Thus FGF induces Xbra gene transcription, and, inturn, the Xbra protein is a transcription factor of theFGF gene (Isaacs et al., 1994; Schulte-Merker andSmith, 1995). This autocatalytic loop tends to amplify alow level of Xbra expression but have no role in theabsence of Xbra; hence, it sharpens the expression/no-expression threshold. A manifestation of this is the“community effect” whereby a critical mass (i.e., num-ber) of clustered Xbra-positive cells is required formaintenance of a muscle cell fate (Gurdon et al., 1993).This effect is assayed by sandwiching activin-inducedcells either singly or in clumps between uninducedanimal caps. Activin-treated cells surrounded by un-treated tissue revert to an epidermal fate while clumpsof induced cells differentiate as muscle (Gurdon, 1988).This effect is mediated by eFGF (Standley et al., 2001,2002) and presumably results from a process wherebyeFGF in isolated induced cells dissipates too rapidly tomaintain the induced state.
The second Xbra dose–response threshold sharpen-ing mechanism that has been described is the cross-inhibition of Xbra by goosecoid. Specifically, goosecoidis induced only at higher activin/nodal doses and itrepresses Xbra expression (Artinger et al., 1997; Lau-rent et al., 1997; Latinkic et al., 1997; Danilov et al.,1998). Goosecoid is itself a transcriptional repressorand probably represses Xbra directly (Smith andJaynes, 1996). Xbra, on the other hand, does not di-rectly repress Goosecoid expression (Artinger et al.,1997) but perhaps reduces its maintenance, either bymeans of FGF or intracellularly, perhaps by means ofsuppression of Xlim-1 and Otx1 action (Mochizuki etal., 2000). This kind of cross-repression is likely to be a
highly general feature of morphogen interpretation bycells, because active repression seems to be as impor-tant or more important compared with activation indetermining spatially restricted expression of severalgenes (e.g., Yuh et al., 1998; Lerchner et al., 2000).
A recent article analyzing the precision of morpho-gen gradients in early Drosophila embryos illustratesthat there is still much to learn about gradient sharp-ening mechanisms and developmental precision.Houchmandzadeh et al. (2002) used quantitative fluo-rescence staining to examine the distributions of Bicoidand Hunchback proteins in the early embryo. Theyshowed that, although the Bicoid distribution wasquite variable, the distribution of Hunchback (which isprimarily regulated by Bicoid) is extremely tight atcertain critical positions in the embryo (viz. variationin the middle of the embryo of 30% and 4% egg lengthfor Bicoid and Hunchback, respectively). Furthermore,the Hunchback distribution scaled well to overall em-bryo length, whereas the Bicoid distribution did not.Further genetic analysis showed that, among all thecandidate genes and a large proportion of the entiregenome, the only gene that affected the precision of theHunchback gradient was the maternal gene staufen.The mechanism whereby the staufen gene product pro-duces such precision is as yet mysterious.
INTERPRETATION OF GRADIENTS. PART II:FROM RECEPTORS TO NUCLEUS
The thresholding mechanisms just described pre-sume a simple quantitative correlation between extra-cellular morphogen concentration and intracellular ac-tivation of the relevant transducing factor(s). As itturns out, activin does indeed work this way (see be-low). However, it is not known whether this is the casefor any other morphogen, and certainly, other mecha-nisms can be envisaged. In Xenopus, a detailed analy-sis of how cells “sense” applied growth factor concen-tration has been achieved in a series of elegant studiesby Gurdon and colleagues by using activin as a modelmorphogen (reviewed in McDowell and Gurdon, 1999).These studies exemplify some of the best aspects ofworking with Xenopus. Gurdon showed that the dose–response thresholds seen in Xenopus cells could beconverted into positional information. This was done byconstructing artificial activin sources (beads soaked inthe growth factor) and placing them next to responsiveanimal cap tissue (Gurdon et al., 1994, 1995). It wasobserved that the activin-soaked bead provokes a waveor ripple of Xbra expression followed by appearance ofa domain of goosecoid expression close to the beaditself. Thus, Xbra and goosecoid transcriptional re-sponses were detected further from and closer to thebeads, respectively, as would be expected from a high-to-low gradient of activin in the tissue. Tissue accumu-lates and “remembers” the concentrations of activinthat it receives, a phenomenon described as a “ratchet-like” mechanism (Green et al., 1990; Gurdon et al.,1995). This is in contrast to many biological systemswhere a stimulus is followed by damping or habitua-
399MORPHOGEN GRADIENTS AND XENOPUS

tion, resulting in a transient rather than cumulativeeffect. By using radioactively labeled activin protein,Dyson and Gurdon (1998) showed that the responses ofcells depend on the absolute number of receptors occu-pied (rather than, say, the ratio of occupied to unoccu-pied receptors) and that this number is relativelysmall: per cell, approximately 100 for Xbra and 300 forGsc transcription. Importantly, activin binds with highaffinity and long occupancy (ligand off-time), providinga basis for the cumulative memory of activin concen-tration.
The detailed understanding of the TGF-� superfam-ily signal transduction mechanism provides essentialinformation for the understanding of how activin andpresumably nodal signaling works (reviewed in Atti-sano and Wrana, 2002). The transduction pathway isshort: ligand-dimerized receptor kinases phosphorylateintracellular Smad proteins (predominantly Smad2 foractivin and nodal; Smad1, Smad5, and Smad8 forBMPs) that then, helped by cofactors, enter the nucleusand act as positive transcription factors. Given thecatalytic nature of the receptor kinases, this processcould, in theory, be highly amplifying (as it is withprogesterone and the MAPK cascade mentionedabove). However, a threefold difference in the nuclearconcentration of phosphorylated Smad2 is sufficient todifferentiate Xbra and Gsc responses, just as with theexternal concentration of activin, and that changingapplied activin changes the amount of nuclear Smad2in a faithful 1:1 ratio (Shimizu and Gurdon, 1999). Thisfinding shows that no amplification of the activin signaloccurs. Pulse-chase experiments showed that Smad2turns over relatively rapidly and that ongoing serine-threonine kinase activity, presumably from activatedreceptors, is necessary for maintenance of nuclearSmad2 concentrations (Bourillot et al., 2002). Thus, wehave a picture whereby activin accumulates on thereceptors and stays there, providing an ongoing phos-phorylation of Smad 2 and, hence, an ongoing flux ofSmad2 from cytoplasm to nucleus. This mechanismprovides an explanation of the ratchet effect and cellu-lar memory of applied activin dose. It would be inter-esting to learn whether nodal or any of the other TGF-�superfamily members operates in exactly the sameway.
As development proceeds, cells lose their ability torespond to specific inducing signals and, in the case ofactivin, this occurs during gastrulation stages. Recentevidence is that this loss of competence is mediated byexclusion of Smad2 from the nucleus (Grimm and Gur-don, 2002). Loss of competence is important as a way oflimiting the progression of a signal and for allowingmultiple rounds of patterning that subdivide embryosinto finer and finer territories.
ESTABLISHMENT OF MORPHOGENGRADIENTS. PART I: EVIDENCE FOR
DIRECT, LONG-RANGE ACTION
Although it is possible to detect the intracellularBicoid gradient in situ at the level of RNA and as a
protein distribution, for a long time, there was pessi-mism about the possibility of detecting the graded dis-tribution of growth factors in vivo. This view arosebecause growth factors are extremely potent, oftenhave biological effects at picomolar concentrations, andconsequently might never be detectable by typical an-tibody techniques. However, recently there has beensome success in detecting morphogen gradients in situin vertebrate tissues by using antibodies. For example,improved antibodies allowed the demonstration of gra-dients of Sonic Hedgehog and Indian Hedgehog mor-phogens in several structures (e.g., tooth, neural tube,somites) of the mouse embryo (Gritli-Linde et al.,2001). In Xenopus, direct visualization of early endog-enous secreted morphogens has not been achieved.However, growth factor-soaked beads have been usedto visualize artificial gradients of activin and TGF-�1at physiologically functional levels that produce theappropriate dose-dependent effects (McDowell et al.,2001). In Drosophila, use of green fluorescent protein(GFP) -tagged Decapentaplegic (Dpp) has allowed thevisualization of a gradient of that morphogen (Entchevet al., 2000; Teleman and Cohen, 2000).
Even without direct visualization, there is now abody of evidence for direct action of several morpho-gens over many cell diameters. Before detailing some ofthis evidence, it is worth mentioning an example of acandidate morphogen that turned out not to be the realthing. It was thought for a long time that retinoic acidwas the morphogen that patterned the anteroposterioraxis of the vertebrate limb, determining the numberand type of digits (Tickle, 1999). This reasoning wasbased on dose–response experiments not unlike someof those described above for activin in the Xenopusmesoderm. However, retinoic acid reporter gene exper-iments, among others, indicated that retinoic acid doesnot act as a morphogen (Noji et al., 1991) but insteadlocally induces a source of a real morphogen with di-rect, long-range action, namely Sonic Hedgehog protein(Riddle et al., 1993).
Evidence for the possibility of direct long-range ac-tion of TGF-� superfamily morphogen signaling hasbeen found in Xenopus. Directness is important, be-cause it distinguishes morphogens from inducers ofmorphogens (as in the Shh/retinoic acid example justcited). First, activated mutants of activin/nodal recep-tors act cell-autonomously, ruling out a mechanismwhereby activin or nodal induce a secondary long-range factor (“relay mechanism;” Jones et al., 1996).Second, activin and TGF-�1 are able to signal acrosslayers of cells lacking receptors (Gurdon et al., 1994),indicating that these factors were not inducers of othermorphogens and, therefore, were acting directly oncells at some distance from their source. Elegant ge-netic studies using receptorless cells had previously ledto similar conclusions for Dpp (BMP4) and Wingless(Wnt) signaling in Drosophila (Nellen et al., 1996;Zecca et al., 1996; Lecuit et al., 1996). A weakness ofthe Xenopus studies is that they relied on the use ofligands and receptors (activin and TGF-�1 and a
400 GREEN

TGF-� receptor) that are not expressed in the embryo.Evidence for direct signaling by an endogenous TGF-�superfamily morphogen in a vertebrate has come fromzebrafish. Schier and colleagues showed that Squint(nodal) could have long-range effects that are depen-dent on the nodal coreceptor One-Eyed Pinhead/Cripto,and yet these signals could traverse cells lacking thiscoreceptor (Chen and Schier, 2001). However, eventhough Squint is an endogenous nodal, several caveatsto this experiment should be noted. First, althoughusing an “endogenous” ligand, it was ectopically ex-pressed in this experiment. Second, the read-out forthis experiment was transcription of nodal-responsivemarkers (albeit directly responsive ones), and therewas no direct visualization of a gradient. Third, butperhaps most interesting, the mutant cells lacking thecoreceptors may have provided an abnormally permis-sive tissue for the long-range diffusion of the ligand(see next section). Thus, although evidence that activinand nodal are capable of long-range action throughtissues under the right circumstances, the specific ev-idence for direct long-range action in vivo is tantaliz-ingly incomplete.
Similar experiments have been used to show thatSonic Hedgehog (Shh) acts in a direct, dose-dependentlong-range way in the chick neural tube. Thus, chang-ing the strength of an Shh signal from the notochordcan shift the positions of various positional markers inthe overlying neural tube, but when cells at variousdistances from the source are made Shh-insensitive (byexpressing a mutant receptor), their fates are shifted(Briscoe et al., 2001). In addition to the Hedgehog mor-phogens and the TGF-�–related activin and nodal pro-teins, there is evidence in the Xenopus blastula andgastrula for long-range action of BMP inhibitors nogginand chordin (Jones and Smith, 1998; Blitz et al., 2000).Given that there is no evidence for a gradient of BMP4itself and that BMP4 has only a very short range ofaction (Jones et al., 1996), the apparently graded ef-fects of BMP4 signaling in the embryo, thus, may bedue to an oppositely directed gradient of these BMPinhibitors in vivo (see Dale and Jones, 1999).
During the exploration of the range of TGF-� super-family morphogens in Xenopus, it has emerged thattheir range of action is affected by variations in boththe nature of the morphogen and the tissue throughwhich it acts. For example, TGF-�1 itself has a rela-tively short range when introduced into tissues bymeans of expression from injected RNA but is able todiffuse over many cell diameters when introduced asactive protein loaded onto beads (Reilly and Melton,1996; McDowell et al., 2001). One possible explanationfor this finding is that the processing of TGF-�1 fromits inactive precursor may be limiting (McDowell et al.,2001). Processing of TGF-� superfamily members iscomplex but clearly very important (Gleizes et al.,1997). They are synthesized as zymogen-like pre-pro-teins whose carboxyl-terminal one third is the actualligand, whereas the amino-terminal two thirds is a“latency peptide” or “pro-domain.” The pro-domain is
cleaved from, but can remain noncovalently associatedwith, the ligand peptide. In some cases, a second pro-teolytic cleavage releases the pro-domain. The struc-ture and processing of the pro-domain of a given TGF-�superfamily member can regulate its activity. For ex-ample, the BMP-4 requires two successive proteolyticcleavages, and modification of either of the cleavedsites significantly alters its range of action and stabil-ity (Cui et al., 2001). A similar role for the pro-domainof mammalian nodal has been demonstrated (Constamand Robertson, 1999). In Xenopus, it was shown thatunmodified Xnr2 acts only over a short range but achimeric protein consisting of the N-terminal pro-do-main of activin fused to the ligand region of Xnr2 hasthe long-range transmission properties of activin(Jones et al., 1996). Thus, the type of precursor fromwhich the TGF-� ligands are produced, and specificallythe N-terminal pro-domain, can determine the range ofaction. These findings should serve as a caution thatthe rich experimental literature on the long-range mor-phogen actions of activin should not be applied uncriti-cally to the likely endogenous actions of nodals. Recentstudies (Yeo and Whitman, 2001; Eimon and Harland,2002) have revealed yet another layer of regulatorycomplexity: they show that many TGF-� superfamilymembers can form heterodimers, including those be-tween BMPs, nodals, and derriere, and that the het-erodimers have distinct inducing properties that con-found interpretations of which signaling molecules areacting when and where in vivo.
The importance of the state of the tissue throughwhich morphogens is transmitted is revealed by exper-iments in which disaggregation and reaggregation ofanimal cap cells has the effect of permitting long-rangeaction of the otherwise short-range Xnr2 (Jones et al.,1996). Of course, such an artificially perturbed tissueonly hints that an intact extracellular matrix of normaltissue is important. Clearer evidence for this comesfrom Drosophila genetics. Drosophila mutants tout-velu and dally prevent normal Wingless morphogendiffusion and are defective in the synthesis extracellu-lar matrix–associated heparan sulfate proteoglycans(HSPGs; Lin and Perrimon, 2000). The role of suchmatrix factors in morphogen diffusion is still poorlyunderstood, although they can certainly act as corecep-tors in target cells (Lin and Perrimon, 1999). It isperhaps significant that HSPGs are important in me-soderm formation (Itoh and Sokol, 1994; Brickman andGerhart, 1994) and that the restriction of the range-of-action of BMP4 depends substantially on an HSPG-binding three amino acid motif in its N-terminal region(Ohkawara et al., 2002).
ESTABLISHMENT OF MORPHOGENGRADIENTS. PART II: PHYSICALMECHANISMS OF MORPHOGEN
DISTRIBUTION
The word “diffusion” is used above in the rather loosesense of “spread through tissues.” However, there hasbeen a great deal of interest lately in the actual phys-
401MORPHOGEN GRADIENTS AND XENOPUS

ical mechanisms by which morphogens come to be dis-tributed in gradients (Strigini and Cohen, 1999; Nara-yanan and Ramaswami, 2001; Teleman et al., 2001;Freeman, 2002; Christian, 2002; Entchev and Gonza-lez-Gaitan, 2002; Cadigan, 2002). Passive diffusion isbut one of the possible mechanisms. Understandingthese mechanisms is important for the integration intodetailed models of the phenomena of cell polarity es-tablishment and tissue and organ regeneration as wellas the many phases of normal development.
The relative simplicity of passive diffusion has madeit the basis for a great number of theoretical mathe-matical models of morphogen action. Francis Crick, forexample, argued that, by making reasonable estimatesof the effective diffusion constants and taking into ac-count the known times of patterning events (5–10 hr)and distances over which patterning occurs (less than 1mm; Wolpert, 1969), a simple model could be con-structed to make a gradient with passive diffusion of asmall (300–500 Da) molecule morphogen (Crick, 1970).A more detailed model using passive diffusion has evenbeen made for the specific case of mesodermal pattern-ing in Xenopus embryos (Weyer et al., 1977). In thesemodels, a steady-state gradient is generated by assum-ing a morphogen source and a distal sink or a distrib-uted decay of the morphogen. We now know that mor-phogens can be much larger that Crick’s postulated“small molecule,” but to retain the idea of a passivediffusion mechanisms, this merely requires that mod-els assume an effective diffusion constant for tissuesthat is lower. Simple models are sufficiently elastic toaccommodate this discovery.
More sophisticated models recently have given thenotion of passive morphogen diffusion both a challengeand a rejoinder. Kerzberg and Wolpert (1998) showedthat inclusion of receptor kinetics in diffusion modelsmakes a big difference to the modeled progression ofmorphogens through tissues. They found that recep-tors tend to trap morphogen molecules, so that insteadof formation of a gradient, morphogen progressesacross a field of cells as a traveling front of receptorsaturation. Such a traveling front could in principleresult in a graded response if cells can integrate theirexposure over time, because proximal cells are satu-rated for proportionately longer than more distal ones(Fig. 2). Such integration (dose � concentration �time) is an established property of, for example, theactivin response of Xenopus cells (Green et al., 1990)and is likely a function of slow dissociation of activinfrom its receptor (Dyson and Gurdon, 1998) and sus-tained receptor kinase activity (Bourillot et al., 2002).A traveling front is observed in the form of Smad2activation (phosphorylation) progressing from the Or-ganizer to the other side of the Xenopus and zebrafishlate blastula/early gastrula (Faure et al., 2000; Lee etal., 2001; Fig. 1). Of interest, the traveling front ofSmad2 activation is followed by a traveling front ofinactivation. Thus, any gradient-like interpretationmust take place in a tightly restricted time window.
To account for actual gradient formation—observed,for example, for Dpp gradients in Drosophila, Kerzbergand Wolpert suggested a “bucket brigade” model inwhich morphogen molecules bound to receptors on onecell could be handed off to receptors on an adjacent cell.This process restores robust gradient formation with-out requiring passive diffusion but has the disadvan-tage that there is no experimental evidence for such amechanism. Recently, Lander et al. (2002) have pro-duced a simple alternative model that invokes passivediffusion and receptor kinetics but does not lead tomorphogen trapping. This model instead includes thewell-known processes of ligand-receptor complex inter-nalization (endocytosis) and ligand degradation. Evenif the internalization is made reversible, the modelrobustly predicts gradient formation with plausible ki-netics and physical range.
The model of Lander et al. (2002) importantly under-cuts the assumption that, if endocytosis is important ingradient formation, then it must be as a part of trans-cytosis, i.e., the endocytic uptake, transit though thecytoplasm, and exocytic secretion of ligands. The trans-cytosis model has arisen from several observations ofDpp and Wingless gradient formation in Drosophila.Direct visualization of morphogen gradients (using ei-ther antibodies or biologically active GFP-conjugatedmorphogens) has shown a significant fraction of themolecules to be localized to intracellular vesicles (En-tchev et al., 2000). In the case of Dpp, its spread andconsequent gradient formation is dependent on dynamin,a GTPase that mediates clathrin-coated endocytosisthat is mutated in shibire mutant embryos (see En-tchev and Gonzalez-Gaitan, 2002). This endocytosis-dependent morphogen movement is observed both forDpp and for Wingless (Moline et al., 1999; Strigini andCohen, 2000; Dubois et al., 2001). This observation hasled to models in which morphogen molecules are trans-ported through cells by endocytosis on one side, vesic-ular transport across the cytoplasm and exocytosis onthe other. This transcytosis would allow for sophisti-cated regulation of morphogen movement. It also couldbe a mechanism for preventing a morphogen from es-caping from a tissue. Such escape could be problematicin cases such as the Drosophila imaginal disc wherethe epithelial sheet being patterned is highly foldedand such escape could allow inappropriate hopping ofthe morphogen from one fold to another (Entchev et al.,2000). However, the mathematical modeling describedabove shows that endocytosis is just as important fordiffusion-based gradient formation as it is for transcy-tosis. Lander et al. argue that the kinetics of morpho-gen gradient formation in vivo is too fast comparedwith the estimated rates of transcytosis. Similarly, abucket brigade relay a la Kerzberg-Wolpert would betoo slow—at least in the case of TGF-�-type morpho-gens with their slow dissociation from receptors. Itshould be added that dynamin (shibire) mutations inendocytosis could also affect the levels of expression ofreceptors and other molecules on the cell surface and,thus, inhibit Dpp movement by mechanisms other than
402 GREEN

either transcytosis or ligand-mediated receptor endo-cytosis.
Drosophila Wingless, unlike Dpp, gradient formationdoes not depend on dynamin function, and a Winglessprotein gradient is mostly observed extracellularly (seeTeleman et al., 2001). This finding could be taken as anargument that it does not require transcytosis or endo-cytosis to form a gradient. However, there is someevidence in favor of Wingless transcytosis. Winglesshas been observed in a population of actively trans-ported vesicles dubbed “argosomes” (Greco et al., 2001).These argosomes move very rapidly across cells, con-sistent with active molecular motors, and can containWingless protein but are regulated and selective in thecargoes that they carry (Greco et al., 2001). Much is yetto be learned about argosomes, but their rapid move-ment at least overcomes some theoretical objections tothe slowness of transcytosis in morphogen gradientformation.
Cytonemes are another type of cellular organellethat could contribute to morphogen distribution. Cy-tonemes are many cell-diameters–long, actin-contain-ing filamentous cellular extensions or filopodia thatcould, in principle, receive morphogens from nonadja-cent cells (Ramirez-Weber and Kornberg, 1999; Korn-berg, 1999). Cytonemes are not well characterized, andat present, it is difficult to say whether they do indeedplay a role in morphogen gradients. However, they doserve as a caution that models in which cells are merelyclose-packed membranous bags are likely to be wide ofthe mark.
It seems likely that different morphogens, or eventhe same morphogen, may form a gradient by usingdifferent mechanisms in different species and tissues.However, that morphogens themselves are evolution-arily conserved opens the possibility that there are onlya few favored mechanisms. The transmission andspread of morphogens has been one area where theoryhas had a fruitful dialogue with experiment, and thisdialogue will no doubt continue. One experimentshould be mentioned that seems to defy the currentconsensus between theory and experiment on the im-portance of endocytosis. The spread of TGF-�1 throughanimal cap tissues expressing high-affinity receptors isindependent of its temperature-sensitive internaliza-tion (McDowell et al., 2001). Even considering that thisis in an artificial situation using possibly traumatizedexplanted tissues, it still allows for the possibility that,despite the theory and the Drosophila examples, thistype of gradient is entirely diffusion-based. It should beadded that most studies on morphogen function havebeen on epithelia (e.g., the Drosophila imaginal disc) orthe tightly packed tissue of the Xenopus blastula. How-ever, much signaling occurs in mesenchyme or betweenmesenchyme and epithelium (e.g., the limb, the tooth).It is an open question whether and how gradientsmight be formed in the loose structure of mesenchyme,or whether, as seems to be the case in the proximodis-tal patterning of the limb, gradient patterning takes
place when cells are tightly packed (Dudley et al.,2002).
MORPHOGENS NOW: GENERAL PRINCIPLESAND EMERGING THEMES
Not so long ago, Lewis Wolpert wrote a review enti-tled “100 Years of Positional Information” (Wolpert,1996). He could have written “100 years of morpho-gens” except that, until relatively recently, morpho-gens were theoretical rather than established physicalentities. Although physical identification of morpho-gens has been accumulating since the mid-1980s, theirexistence and the free use of the word morphogen hasbecome noticeably more current over the past 3 or 4years.
As complete genome sequences are being revealed, itis becoming possible to think of finite numbers of typesof genes and to classify morphogens into relatively fewclasses. TGF-betas, Wingless/Wnts, and Hedgehogs areso frequently used in early embryos that one can hopethat a detailed understanding of these three will ex-plain a significant majority of early patterning pro-cesses. It is also worth pointing out that there aremany gradients, but each has relatively few thresholds.For example, activin can induce many responses incells, but of these responses, perhaps only two aredirect (exemplified by Xbra and Goosecoid), and theothers arise from secondary interactions between in-duced cells (Green et al., 1994; Wilson and Melton,1994). Based on most mathematical models of gradientpatterning, this reasoning makes sense: it is easier toachieve precision throughout a given territory by usingmany sequential rounds of patterning and requiringless of each than using a single gradient with morethresholds (Green and Cooke, 1991; see also Yates andPate, 1989).
Another theme that has emerged over the past fewyears is that the detailed understanding of the bio-chemistry of signal transduction—a study once alien toanatomically oriented developmental biologists—hasbecome central to unraveling the intricacies of spatialregulation and morphogen gradient action. In this re-gard, Xenopus has provided a bridge between biochem-istry and patterning studies as a leading system in theelucidation of the TGF-� and Wnt transduction path-ways, among others.
Three areas are likely to be of crucial importance inunderstanding morphogen action in the near future.First, as is clear from the analysis of morphogen trans-mission, much more will come out of subcellular anat-omy and classic cell biology. Thus, to complement thedetailed knowledge of the kinases and phosphatasesfamiliar to the “signal transductionologists,” we willhave to study molecules that regulate endocytosis, ar-gosomes, cytonemes, and vesicular (and other intracel-lular) trafficking. Second, the role of feedback regula-tion will have to be further analyzed. The studies onthreshold sharpening mentioned above certainly onlyscratch the surface of the problem of how scaling andregeneration are achieved. Third, the processes that
403MORPHOGEN GRADIENTS AND XENOPUS

precede and follow morphogen gradient establishment,namely boundary establishment and cell polarity, willbecome integrated into the developmental narrative.Boundary establishment has obvious intuitive mecha-nistic significance, but our knowledge of the physicalmechanisms that govern it is still in its infancy (Irvineand Rauskolb, 2001). As for polarity, the way in whichmorphogen gradients provide both scalar and vectorialinformation to cells is almost definitive for true mor-phogens (Wolpert, 1969; Lawrence, 2001a). Recentprogress has been made in understanding the detailedmolecular mechanisms of planar polarity (e.g., Law-rence et al., 2002; Yang et al., 2002; Tree et al., 2002).The picture that emerges is that graded extracellularsignals provide a bias for movement of specific mole-cules to one end of a cell or the other and that theseintracellular distributions are reinforced by very localpositive and negative feedback loops. So far, this workhas come almost exclusively from the study of Drosoph-ila, but despite that Xenopus laevis genetics are tech-nically intractable, its deep familiarity, large size, ac-cessibility, and ease of manipulation will no doubtcontinue to place it centrally in these ongoing studies.
MORPHOGEN GRADIENTS:HISTORICAL NOTE
Before he did the Organizer experiment, Spemannshowed that bisected amphibian embryos give smallbut anatomically normal embryos (see Hamburger,1988), whereas Waddington showed that adding extratissue to a frog embryo modifies the size but not theanatomy of the resulting tadpole (Waddington, 1938).These are but two amphibian examples among manyother experiments from the late 1800s through to the1930s on diverse species, demonstrating long-range co-ordinated regulation of patterning in embryos. To ex-plain such phenomena, the concept of the “morphoge-netic field” was developed and elaborated on in the1920s and 1930s (reviewed in Gilbert, 2000) to describethe tissue or region in which such long-range patternregeneration or stability could be observed. This ideabecame the “gradient field,” essentially identical to thecurrent concept of morphogen gradients. Many articlesand books were written on the subject (see Gilbert,2000, for references). A particularly intriguing examplewas Dalcq and Pasteels, who proposed a two-coordinategradient system for early amphibian patterning (Dalcqand Pasteels, 1937; Dalcq, 1938) that fits well withsome of our current ideas (see Fig. 3).
Although it was at one time popular, the idea ofgradient fields fell out of favor in the late 1930s andearly 1940s. This trend was partly due the decline ofGerman embryology under the Nazis, but there weretwo scientific reasons. One was the rise of genetics:ironically, T.H. Morgan, who had been among the firstto think about gradients, became actively hostile to thisidea from “old style” experimental embryology, whichchallenged the new focus on genes. A second reasonwas the spectacular failure of biochemists to identify
morphogens, in particular the Organizer substance(Witkowski, 1985, 1987).
Interest in morphogens was revived when mathema-ticians and physicists brought a theoretical inclinationto problems in biology after WWII. One of the first ofthese investigators was Alan Turing, who (after devel-oping the mathematics that underlies all digital com-puting) devised a “reaction-diffusion” model for regu-lating morphogens and by the way coined the term“morphogen” (Turing, 1952). However, Turing’s was alonely voice: as late as 1970, Crick (another physicistwho went into biology) could write that “gradients havebecome a dirty word” (Crick, 1970). In the 1970s, led byWolpert (1969), Crick, Meinhardt, and Gierer, therewas a surge in gradient theories informed by a newconfidence in the power of molecular biology, althoughwith little molecular support.
The 1980s brought the power of Drosophila geneticsto the problem of developmental patterning with thefruition of the work of Lewis, Nusslein-Volhard, andWieschaus, among others. Meanwhile, the growth ofmolecular biology and the characterization of proteingrowth factors in cell culture led to the discovery of realmorphogens in vertebrate embryos (see main text).This might be described as the end of “Morgan’s De-tour,” from amphibian gradients by means of Drosoph-ila genetics and back again.
ACKNOWLEDGMENTS
I thank John Fallon, Qiufu Ma, Melissa Rubock, andthe anonymous referees for critical reading of the manu-script and helpful suggestions. J.B.A.G. is supported bygrants from the NIH and National Science Foundation.
REFERENCES
Agius E, Oelgeschlager M, Wessely O, Kemp C, De Robertis EM. 2000.Endodermal nodal-related signals and mesoderm induction in Xe-nopus. Development 127:1173–1183.
Artinger M, Blitz I, Inoue K, Tran U, Cho KW. 1997. Interaction ofgoosecoid and brachyury in Xenopus mesoderm patterning. MechDev 65:187–196.
Attisano L, Wrana JL. 2002. Signal transduction by the TGF-betasuperfamily. Science 296:1646–1647.
Blader P, Rastegar S, Fischer N, Strahle U. 1997. Cleavage of theBMP-4 antagonist chordin by zebrafish tolloid. Science 278:1937–1940.
Blitz IL, Shimmi O, Wunnenberg-Stapleton K, O’Connor MB, ChoKW. 2000. Is chordin a long-range- or short-range-acting factor?Roles for BMP1- related metalloproteases in chordin and BMP4autofeedback loop regulation. Dev Biol 223:120–138.
Blumberg B, Wright CV, De Robertis EM, Cho KW. 1991. Organizer-specific homeobox genes in Xenopus laevis embryos. Science 253:194–196.
Bourillot PY, Garrett N, Gurdon JB. 2002. A changing morphogengradient is interpreted by continuous transduction flow. Develop-ment 129:2167–2180.
Brannon M, Kimelman D. 1996. Activation of Siamois by the Wntpathway. Dev Biol 180:344–347.
Brickman MC, Gerhart JC. 1994. Heparitinase inhibition of meso-derm induction and gastrulation in Xenopus laevis embryos. DevBiol 164:484–501.
Briscoe J, Chen Y, Jessell TM, Struhl G. 2001. A hedgehog-insensitiveform of patched provides evidence for direct long-range morphogenactivity of sonic hedgehog in the neural tube. Mol Cell 7:1279–1291.
404 GREEN

Buff E, Carmena A, Gisselbrecht S, Jimenez F, Michelson AM. 1998.Signalling by the Drosophila epidermal growth factor receptor isrequired for the specification and diversification of embryonic mus-cle progenitors. Development 125:2075–2086.
Cadigan KM. 2002. Regulating morphogen gradients in the Drosoph-ila wing. Semin Cell Dev Biol 13:83–90.
Carmena A, Gisselbrecht S, Harrison J, Jimenez F, Michelson AM.1998. Combinatorial signaling codes for the progressive determina-tion of cell fates in the Drosophila embryonic mesoderm. Genes Dev12:3910–3922.
Chang C, Wilson PA, Mathews LS, Hemmati-Brivanlou A. 1997. AXenopus type I activin receptor mediates mesodermal but not neu-ral specification during embryogenesis. Development 124:827–837.
Chang C, Holtzman DA, Chau S, Chickering T, Woolf EA, HolmgrenLM, Bodorova J, Gearing DP, Holmes WE, Brivanlou AH. 2001.Twisted gastrulation can function as a BMP antagonist. Nature410:483–487.
Chen Y, Schier AF. 2001. The zebrafish Nodal signal Squint functionsas a morphogen. Nature 411:607–610.
Chitnis AB. 1999. Control of neurogenesis—lessons from frogs, fishand flies. Curr Opin Neurobiol 9:18–25.
Christen B, Slack JM. 1999. Spatial response to fibroblast growthfactor signalling in Xenopus embryos. Development 126:119–125.
Christian JL. 2002. Argosomes: intracellular transport vehicles forintercellular signals? Sci STKE 2002:E13.
Clements D, Friday RV, Woodland HR. 1999. Mode of action of VegTin mesoderm and endoderm formation. Development 126:4903–4911.
Constam DB, Robertson EJ. 1999. Regulation of bone morphogeneticprotein activity by pro domains and proprotein convertases. J CellBiol 144:139–149.
Cornell RA, Kimelman D. 1994a. Activin-mediated mesoderm induc-tion requires FGF. Development 120:453–462.
Cornell RA, Kimelman D. 1994b. Combinatorial signaling in develop-ment. Bioessays 16:577–581.
Cornell RA, Musci TJ, Kimelman D. 1995. FGF is a prospectivecompetence factor for early activin-type signals in Xenopus meso-derm induction. Development 121:2429–2437.
Crease DJ, Dyson S, Gurdon JB. 1998. Cooperation between theactivin and Wnt pathways in the spatial control of organizer geneexpression. Proc Natl Acad Sci U S A 95:4398–4403.
Crick F. 1970. Diffusion in embryogenesis. Nature 5231:420–422.Cui Y, Hackenmiller R, Berg L, Jean F, Nakayama T, Thomas G,
Christian JL. 2001. The activity and signaling range of matureBMP-4 is regulated by sequential cleavage at two sites within theprodomain of the precursor. Genes Dev 15:2797–2802.
Dalcq AM. 1938. Form and causality in early development. London:Cambridge University Press.
Dalcq A, Pasteels JJ. 1937. Une conception nouvelle des bases physi-ologiques de la morphogenese. Arch Biol 48:669–710.
Dale L. 1997. Development: morphogen gradients and mesodermalpatterning. Curr Biol 7:R698–R700.
Dale L, Jones CM. 1999. BMP signalling in early Xenopus develop-ment. Bioessays 21:751–760.
Dale L, Howes G, Price BM, Smith JC. 1992. Bone morphogeneticprotein 4: a ventralizing factor in early Xenopus development. De-velopment 115:573–585.
Danilov V, Blum M, Schweickert A, Campione M, Steinbeisser H.1998. Negative autoregulation of the organizer-specific homeoboxgene goosecoid. J Biol Chem 273:627–635.
Day SJ, Lawrence PA. 2000. Measuring dimensions: the regulation ofsize and shape. Development 127:2977–2987.
DeRobertis EM, Sasai Y. 1996. A common plan for dorsoventral pat-terning in Bilateria. Nature 380:37–40.
Dick A, Hild M, Bauer H, Imai Y, Maifeld H, Schier AF, Talbot WS,Bouwmeester T, Hammerschmidt M. 2000. Essential role of Bmp7(snailhouse) and its prodomain in dorsoventral patterning of thezebrafish embryo. Development 127:343–354.
Dosch R, Gawantka V, Delius H, Blumenstock C, Niehrs C. 1997.Bmp-4 acts as a morphogen in dorsoventral mesoderm patterningin Xenopus. Development 124:2325–2334.
Driever W, Nusslein-Volhard C. 1989. The bicoid protein is a positiveregulator of hunchback transcription in the early Drosophila em-bryo. Nature 337:138–143.
Driever W, Thoma G, Nusslein-Volhard C. 1989. Determination ofspatial domains of zygotic gene expression in the Drosophila em-bryo by the affinity of binding sites for the bicoid morphogen.Nature 340:363–367.
Dubois L, Lecourtois M, Alexandre C, Hirst E, Vincent JP. 2001.Regulated endocytic routing modulates wingless signaling in Dro-sophila embryos. Cell 105:613–624.
Dudley AT, Ros MA, Tabin CJ. 2002. A re-examination of the ProgressZone Model: distinguishing specification and determination duringlimb development. Nature (in press).
Dyson S, Gurdon JB. 1998. The interpretation of position in a mor-phogen gradient as revealed by occupancy of activin receptors. Cell93:557–568.
Eimon PM, Harland RM. 2002. Effects of heterodimerization andproteolytic processing on Derriere and Nodal activity: implicationsfor mesoderm induction in Xenopus. Development 129:3089–3103.
Entchev EV, Gonzalez-Gaitan MA. 2002. Morphogen gradient forma-tion and vesicular trafficking. Traffic 3:98–109.
Entchev EV, Schwabedissen A, Gonzalez-Gaitan M. 2000. Gradientformation of the TGF-beta homolog Dpp. Cell 103:981–991.
Faure S, Lee MA, Keller T, ten Dijke P, Whitman M. 2000. Endoge-nous patterns of TGFbeta superfamily signaling during early Xe-nopus development. Development 127:2917–2931.
Ferguson EL. 1996. Conservation of dorsal-ventral patterning in ar-thropods and chordates. Curr Opin Genet Dev 6:424–431.
Ferguson EL, Anderson KV. 1992. Decapentaplegic acts as a morpho-gen to organize dorsal-ventral pattern in the Drosophila embryo.Cell 71:451–461.
Ferrell JE Jr, Machleder EM. 1998. The biochemical basis of anall-or-none cell fate switch in Xenopus oocytes. Science 280:895–898.
Francois V, Bier E. 1995. Xenopus chordin and Drosophila shortgastrulation genes encode homologous proteins functioning in dor-sal-ventral axis formation. Cell 80:19–20.
Freeman M. 2002. Morphogen gradients, in theory. Dev Cell 2:689–690.
Gerhart J. 1999. 1998 Warkany lecture: signaling pathways in devel-opment. Teratology 60:226–239.
Gilbert S. 2000. The “re-discovery” of morphogenetic fields. Availableat: http://www.devbio.com/chap03/link0305.shtml
Gleizes PE, Munger JS, Nunes I, Harpel JG, Mazzieri R, Noguera I,Rifkin DB. 1997. TGF-beta latency: biological significance andmechanisms of activation. Stem Cells 15:190–197.
Goodman SA, Albano R, Wardle FC, Matthews G, Tannahill D, DaleL. 1998. BMP1-related metalloproteinases promote the develop-ment of ventral mesoderm in early Xenopus embryos. Dev Biol195:144–157.
Graff JM, Thies RS, Song JJ, Celeste AJ, Melton DA. 1994. Studieswith a Xenopus BMP receptor suggest that ventral mesoderm-inducing signals override dorsal signals in vivo. Cell 79:169–179.
Greco V, Hannus M, Eaton S. 2001. Argosomes: a potential vehicle forthe spread of morphogens through epithelia. Cell 106:633–645.
Green JBA. 1999. The animal cap assay. Methods Mol Biol 127:1–13.Green JBA, Cooke J. 1991. Induction, gradient models and the role of
negative feedback in body pattern formation in the amphibianembryo. Semin Dev Biol 2:95–106.
Green JBA, Smith JC. 1990. Graded changes in dose of a Xenopusactivin A homologue elicit stepwise transitions in embryonic cellfate. Nature 347:391–394.
Green JBA, Smith JC. 1991. Growth factors as morphogens: do gra-dients and thresholds establish body plan? Trends Genet 7:245–250.
Green JBA, Howes G, Symes K, Cooke J, Smith JC. 1990. The bio-logical effects of XTC-MIF: quantitative comparison with XenopusbFGF. Development 108:173–183.
Green JBA, New HV, Smith JC. 1992. Responses of embryonic Xeno-pus cells to activin and FGF are separated by multiple dose thresh-olds and correspond to distinct axes of the mesoderm. Cell 71:731–739.
405MORPHOGEN GRADIENTS AND XENOPUS

Green JBA, Smith JC, Gerhart JC. 1994. Slow emergence of a mul-tithreshold response to activin requires cell-contact-dependentsharpening but not prepattern. Development 120:2271–2278.
Grimm OH, Gurdon JB. 2002. Nuclear exclusion of Smad2 is a mech-anism leading to loss of competence. Nat Cell Biol 4:519–522.
Gritli-Linde A, Lewis P, McMahon AP, Linde A. 2001. The where-abouts of a morphogen: direct evidence for short- and graded long-range activity of hedgehog signaling peptides. Dev Biol 236:364–386.
Gritsman K, Talbot WS, Schier AF. 2000. Nodal signaling patternsthe organizer. Development 127:921–932.
Gurdon JB. 1988. A community effect in animal development. Nature336:772–774.
Gurdon JB, Bourillot PY. 2001. Morphogen gradient interpretation.Nature 413:797–803.
Gurdon JB, Kato K, Lemaire P. 1993. The community effect, dorsal-ization and mesoderm induction. Curr Opin Genet Dev 3:662–667.
Gurdon JB, Harger P, Mitchell A, Lemaire P. 1994. Activin signallingand response to a morphogen gradient. Nature 371:487–492.
Gurdon JB, Mitchell A, Mahony D. 1995. Direct and continuous as-sessment by cells of their position in a morphogen gradient. Nature376:520–521.
Gurdon JB, Dyson S, St Johnston D. 1998. Cells’ perception of positionin a concentration gradient. Cell 95:159–162.
Gurdon JB, Standley H, Dyson S, Butler K, Langon T, Ryan K,Stennard F, Shimizu K, Zorn A. 1999. Single cells can sense theirposition in a morphogen gradient. Development 126:5309–5317.
Hamburger V. 1988. The heritage of experimental embryology: HansSpemann and the organizer. New York: Oxford University Press.
Hemmati-Brivanlou A, Melton DA. 1992. A truncated activin receptorinhibits mesoderm induction and formation of axial structures inXenopus embryos. Nature 359:609–614.
Hemmati-Brivanlou A, Melton DA. 1994. Inhibition of activin recep-tor signaling promotes neuralization in Xenopus. Cell 77:273–281.
Hemmati-Brivanlou A, Thomsen GH. 1995. Ventral mesodermal pat-terning in Xenopus embryos: expression patterns and activities ofBMP-2 and BMP-4. Dev Genet 17:78–89.
Holley SA, Jackson PD, Sasai Y, Lu B, De Robertis EM, HoffmannFM, Ferguson EL. 1995. A conserved system for dorsal-ventralpatterning in insects and vertebrates involving sog and chordin.Nature 376:249–253.
Holley SA, Neul JL, Attisano L, Wrana JL, Sasai Y, O’Connor MB, DeRobertis EM, Ferguson EL. 1996. The Xenopus dorsalizing factornoggin ventralizes Drosophila embryos by preventing DPP fromactivating its receptor. Cell 86:607–617.
Holowacz T, Sokol S. 1999. FGF is required for posterior neuralpatterning but not for neural induction. Dev Biol 205:296–308.
Houchmandzadeh B, Wieschaus E, Leibler S. 2002. Establishment ofdevelopmental precision and proportions in the early Drosophilaembryo. Nature 415:798–802.
Hyatt BA, Lohr JL, Yost HJ. 1996. Initiation of vertebrate left-rightaxis formation by maternal Vg1. Nature 384:62–65.
Irvine KD, Rauskolb C. 2001. Boundaries in development: formationand function. Annu Rev Cell Dev Biol 17:189–214.
Isaacs HV, Pownall, ME, Slack JM. 1994. eFGF regulates Xbra ex-pression during Xenopus gastrulation. EMBO J 13:4469–4481.
Isaacs HV, Pownall ME, Slack JM. 1998. Regulation of Hox geneexpression and posterior development by the Xenopus caudal ho-mologue Xcad3. EMBO J 17:3413–3427.
Itoh K, Sokol SY. 1994. Heparan sulfate proteoglycans are requiredfor mesoderm formation in Xenopus embryos. Development 120:2703–2711.
Jiang J, Levine M. 1993. Binding affinities and cooperative interac-tions with bHLH activators delimit threshold responses to the dor-sal gradient morphogen. Cell 72:741–752.
Jones CM, Smith JC. 1995. Inductive signals. Revolving vertebrates.Curr Biol 5:574–576.
Jones CM, Smith JC. 1998. Establishment of a BMP-4 morphogengradient by long-range inhibition. Dev Biol 194:12–17.
Jones CM, Lyons KM, Lapan PM, Wright CV, Hogan BL. 1992. DVR-4(bone morphogenetic protein-4) as a posterior-ventralizing factor inXenopus mesoderm induction. Development 115:639–647.
Jones CM, Kuehn MR, Hogan BL, Smith JC, Wright CV. 1995. Nodal-related signals induce axial mesoderm and dorsalize mesodermduring gastrulation. Development 121:3651–3662.
Jones CM, Armes N, Smith JC. 1996. Signalling by TGF-beta familymembers: short-range effects of Xnr-2 and BMP-4 contrast with thelong-range effects of activin. Curr Biol 6:1468–1475.
Joseph EM, Melton DA. 1997. Xnr4: a Xenopus nodal-related geneexpressed in the Spemann organizer. Dev Biol 184:367–372.
Kavka AI, Green JB. 1997. Tales of tails: Brachyury and the T-boxgenes. Biochim Biophys Acta 1333:F73–F84.
Kavka AI, Green JB. 2000. Evidence for dual mechanisms of meso-derm establishment in Xenopus embryos. Dev Dyn 219:77–83.
Kerszberg M, Wolpert L. 1998. Mechanisms for positional signallingby morphogen transport: a theoretical study. J Theor Biol 191:103–114.
Kimelman D, Kirschner M. 1987. Synergistic induction of mesodermby FGF and TGF-beta and the identification of an mRNA coding forFGF in the early Xenopus embryo. Cell 51:869–877.
Kofron M, Demel T, Xanthos J, Lohr J, Sun B, Sive H, Osada S,Wright C, Wylie C, Heasman J. 1999. Mesoderm induction in Xe-nopus is a zygotic event regulated by maternal VegT via TGFbetagrowth factors. Development 126:5759–5770.
Kornberg T. 1999. Pictures in cell biology. Cytonemes. Trends CellBiol 9:434.
LaBonne C, Whitman M. 1994. Mesoderm induction by activin re-quires FGF-mediated intracellular signals. Development 120:463–472.
Lander AD, Nie Q, Wan FY. 2002. Do morphogen gradients arise bydiffusion? Dev Cell 2:785–796.
Latinkic BV, Umbhauer M, Neal KA, Lerchner W, Smith JC, CunliffeV. 1997. The Xenopus Brachyury promoter is activated by FGF andlow concentrations of activin and suppressed by high concentrationsof activin and by paired-type homeodomain proteins. Genes Dev11:3265–3276.
Laurent MN, Blitz IL, Hashimoto C, Rothbacher U, Cho KW. 1997.The Xenopus homeobox gene twin mediates Wnt induction ofgoosecoid in establishment of Spemann’s organizer. Development124:4905–4916.
Lawrence PA. 1988. Background to bicoid. Cell 54:1–2.Lawrence PA. 2001a. Morphogens: how big is the big picture? Nat Cell
Biol 3:E151–E154.Lawrence PA. 2001b. Wingless signalling: more about the Wingless
morphogen. Curr Biol 11:R638–R639.Lawrence PA, Struhl G. 1996. Morphogens, compartments, and pat-
tern: lessons from Drosophila? Cell 85:951–961.Lawrence PA, Casal J, Struhl G. 2002. Towards a model of the
organisation of planar polarity and pattern in the Drosophila ab-domen. Development 129:2749–2760.
Lecuit T, Brook WJ, Ng M, Calleja M, Sun H, Cohen SM. 1996. Twodistinct mechanisms for long-range patterning by Decapentaplegicin the Drosophila wing. Nature 381:387–393.
Lee KJ, Jessell TM. 1999. The specification of dorsal cell fates in thevertebrate central nervous system. Annu Rev Neurosci 22:261–294.
Lee MA, Heasman J, Whitman M. 2001. Timing of endogenous ac-tivin-like signals and regional specification of the Xenopus embryo.Development 128:2939–2952.
Lemaire P, Garrett N, Gurdon JB. 1995. Expression cloning of Siam-ois, a Xenopus homeobox gene expressed in dorsal-vegetal cells ofblastulae and able to induce a complete secondary axis. Cell 81:85–94.
Lerchner W, Latinkic BV, Remacle JE, Huylebroeck D, Smith JC.2000. Region-specific activation of the Xenopus brachyury promoterinvolves active repression in ectoderm and endoderm: a study usingtransgenic frog embryos. Development 127:2729–2739.
Lewis J, Slack JM, Wolpert L. 1977. Thresholds in development. JTheor Biol 65:579–590.
Lin X, Perrimon N. 1999. Dally cooperates with Drosophila Frizzled 2to transduce Wingless signalling. Nature 400:281–284.
Lin X, Perrimon N. 2000. Role of heparan sulfate proteoglycans incell–cell signaling in Drosophila. Matrix Biol 19:303–307.
Lustig KD, Kroll K, Sun E, Ramos R, Elmendorf H, Kirschner MW.1996. A Xenopus nodal-related gene that acts in synergy with nog-
406 GREEN

gin to induce complete secondary axis and notochord formation.Development 122:3275–3282.
Lustig KD, Stukenberg PT, McGarry TJ, King RW, Cryns VL, MeadPE, Zon LI, Yuan J, Kirschner MW. 1997. Small pool expressionscreening: identification of genes involved in cell cycle control, ap-optosis, and early development. Methods Enzymol 283:83–99.
Malissen B. 1998. Translating affinity into response. Science 281:528–529.
Marom K, Fainsod A, Steinbeisser H. 1999. Patterning of the meso-derm involves several threshold responses to BMP- 4 and Xwnt-8.Mech Dev 87:33–44.
Marques G, Musacchio M, Shimell MJ, Wunnenberg-Stapleton K, ChoKW, O’Connor MB. 1997. Production of a DPP activity gradient inthe early Drosophila embryo through the opposing actions of theSOG and TLD proteins. Cell 91:417–426.
Marti E, Bovolenta P. 2002. Sonic hedgehog in CNS development: onesignal, multiple outputs. Trends Neurosci 25:89–96.
McDowell N, Gurdon JB. 1999. Activin as a morphogen in Xenopusmesoderm induction. Semin Cell Dev Biol 10:311–317.
McDowell N, Gurdon JB, Grainger DJ. 2001. Formation of a func-tional morphogen gradient by a passive process in tissue from theearly Xenopus embryo. Int J Dev Biol 45:199–207.
Medina A, Wendler SR, Steinbeisser H. 1997. Cortical rotation isrequired for the correct spatial expression of nr3, sia and gsc inXenopus embryos. Int J Dev Biol 41:741–745.
Mochizuki T, Karavanov AA, Curtiss PE, Ault KT, Sugimoto N,Watabe T, Shiokawa K, Jamrich M, Cho KW, Dawid IB. 2000.Xlim-1 and LIM domain binding protein 1 cooperate with varioustranscription factors in the regulation of the goosecoid promoter.Dev Biol 224:470–485.
Moline MM, Southern C, Bejsovec A. 1999. Directionality of winglessprotein transport influences epidermal patterning in the Drosoph-ila embryo. Development 126:4375–4384.
Narayanan R, Ramaswami M. 2001. Endocytosis in Drosophila:progress, possibilities, prognostications. Exp Cell Res 271:28–35.
Nellen D, Burke R, Struhl G, Basler K. 1996. Direct and long-rangeaction of a DPP morphogen gradient. Cell 85:357–368.
New HV, Kavka AI, Smith JC, Green JBA. 1997. Differential effectson Xenopus development of interference with type IIA and type IIBactivin receptors. Mech Dev 61:175–186.
Ng JK, Tamura K, Buscher D, Izpisua-Belmonte JC. 1999. Molecularand cellular basis of pattern formation during vertebrate limb de-velopment. Curr Top Dev Biol 41:37–66.
Niehrs C, Dosch R, Onichtchouk D. 2000. Embryonic patterning ofXenopus mesoderm by Bmp-4. Ernst Schering Res Found Workshop29:165–190.
Nishimatsu S, Thomsen GH. 1998. Ventral mesoderm induction andpatterning by bone morphogenetic protein heterodimers in Xenopusembryos. Mech Dev 74:75–88.
Nieuwkoop PD. 1969. The formation of the mesoderm in urodeleanamphibians. I. Induction by the endoderm. Wilhelm Roux’ ArchEntwicklungsmech Org 162:341–373.
Noji S, Nohno T, Koyama E, Muto K, Ohyama K, Aoki Y, Tamura K,Ohsugi K, Ide H, Taniguchi S. 1991. Retinoic acid induces polariz-ing activity but is unlikely to be a morphogen in the chick limb bud.Nature 350:83–86.
Oelgeschlager M, Larrain J, Geissert D, De Robertis EM. 2000. Theevolutionarily conserved BMP-binding protein Twisted gastrulationpromotes BMP signalling. Nature 405:757–763.
Ohkawara B, Iemura S, ten Dijke P, Ueno N. 2002. Action range ofBMP is defined by its N-terminal basic amino acid core. Curr Biol12:205–209.
Onuma Y, Takahashi S, Yokota C, Asashima M. 2002. Multiple nodal-related genes act coordinately in Xenopus embryogenesis. Dev Biol241:94–105.
Osada SI, Wright CV. 1999. Xenopus nodal-related signaling is essen-tial for mesendodermal patterning during early embryogenesis.Development 126:3229–3240.
Papageorgiou S, Almirantis Y. 1996. Gradient model describes thespatial-temporal expression pattern of Hoxa genes in the develop-ing vertebrate limb. Dev Dyn 207:461–469.
Piccolo S, Sasai Y, Lu B, De Robertis E. 1996. Dorsoventral patterningin Xenopus: inhibition of ventral signals by direct binding of chordinto BMP-4. Cell 86:589–598.
Podos SD, Ferguson EL. 1999. Morphogen gradients: new insightsfrom DPP. Trends Genet 15:396–402.
Pownall ME, Isaacs HV, Slack JM. 1998. Two phases of Hox generegulation during early Xenopus development. Curr Biol 8:673–676.
Ptashne M. 1992. A genetic switch: phage lambda and higher organ-isms. Cambridge, MA: Cell Press, Blackweel Scientific.
Ramirez-Weber FA, Kornberg TB. 1999. Cytonemes: cellular pro-cesses that project to the principal signaling center in Drosophilaimaginal discs. Cell 97:599–607.
Ray RP, Wharton KA. 2001. Twisted perspective: new insights intoextracellular modulation of BMP signaling during development.Cell 104:801–804.
Reilly KM, Melton DA. 1996. Short-range signaling by candidatemorphogens of the TGF beta family and evidence for a relay mech-anism of induction. Cell 86:743–754.
Riddle RD, Johnson RL, Laufer E, Tabin C. 1993. Sonic hedgehogmediates the polarizing activity of the ZPA. Cell 75:1401–1416.
Ross JJ, Shimmi O, Vilmos P, Petryk A, Kim H, Gaudenz K, Herman-son S, Ekker SC, O’Connor MB, Marsh JL. 2001. Twisted gastru-lation is a conserved extracellular BMP antagonist. Nature 410:479–483.
Rossi FM, Kringstein AM, Spicher A, Guicherit OM, Blau HM. 2000.Transcriptional control: rheostat converted to on/off switch. MolCell 6:723–728.
Sasai Y, Lu B, Steinbeisser H, Geissert D, Gont LK, De Robertis EM.1994. Xenopus chordin: a novel dorsalizing factor activated by or-ganizer-specific homeobox genes. Cell 79:779–790.
Sasai Y, Lu B, Steinbeisser H, De Robertis EM. 1995. Regulation ofneural induction by the Chd and Bmp-4 antagonistic patterningsignals in Xenopus. Nature 376:333–336.
Schmid B, Furthauer M, Connors SA, Trout J, Thisse B, Thisse C,Mullins MC. 2000. Equivalent genetic roles for bmp7/snailhouseand bmp2b/swirl in dorsoventral pattern formation. Development127:957–967.
Schmidt J, Francois V, Bier E, Kimelman D. 1995. Drosophila shortgastrulation induces an ectopic axis in Xenopus: evidence for con-served mechanisms of dorsal-ventral patterning. Development 121:4319–4328.
Schulte-Merker S, Smith JC. 1995. Mesoderm formation in responseto Brachyury requires FGF signalling. Curr Biol 5:62–67.
Schulte-Merker S, Smith JC, Dale L. 1994. Effects of truncated ac-tivin and FGF receptors and of follistatin on the inducing activitiesof BVg1 and activin: does activin play a role in mesoderm induction?EMBO J 13:3533–3541.
Scott IC, Blitz IL, Pappano WN, Maas SA, Cho KW, Greenspan DS.2001. Homologues of Twisted gastrulation are extracellular cofac-tors in antagonism of BMP signalling. Nature 410:475–478.
Seleiro EA, Connolly DJ, Cooke J. 1996. Early developmental expres-sion and experimental axis determination by the chicken Vg1 gene.Curr Biol 6:1476–1486.
Shah SB, Skromne I, Hume CR, Kessler DS, Lee KJ, Stern CD, DoddJ. 1997. Misexpression of chick Vg1 in the marginal zone inducesprimitive streak formation. Development 124:5127–5138.
Shih J, Keller R. 1992. The epithelium of the dorsal marginal zone ofXenopus has organizer properties. Development 116:887–899.
Shimizu K, Gurdon JB. 1999. A quantitative analysis of signal transduc-tion from activin receptor to nucleus and its relevance to morphogengradient interpretation. Proc Natl Acad Sci U S A 96:6791–6796.
Slack JM. 1993. Embryonic induction. Mech Dev 41:91–107.Slack JM, Darlington BG, Heath JK, Godsave SF. 1987. Mesoderm
induction in early Xenopus embryos by heparin-binding growthfactors. Nature 326:197–200.
Smith JC. 1987. A mesoderm inducing factor is produced by a Xeno-pus cell line. Development 99:3–14.
Smith WC, Harland RM. 1992. Expression cloning of noggin, a newdorsalizing factor localized to the Spemann organizer in Xenopusembryos. Cell 70:829–840.
Smith ST, Jaynes JB. 1996. A conserved region of engrailed, sharedamong all en-, gsc-, Nk1-, Nk2- and msh-class homeoproteins, me-
407MORPHOGEN GRADIENTS AND XENOPUS

diates active transcriptional repression in vivo. Development 122:3141–3150.
Smith JC, Yaqoob M, Symes K. 1988. Purification, partial character-ization and biological effects of the XTC mesoderm-inducing factor.Development 103:591–600.
Smith JC, Price BM, Van Nimmen K, Huylebroeck D. 1990. Identifi-cation of a potent Xenopus mesoderm-inducing factor as a homo-logue of activin A. Nature 345:729–731.
Smith WC, McKendry R, Ribisi S Jr, Harland RM. 1995. A nodal-related gene defines a physical and functional domain within theSpemann organizer. Cell 82:37–46.
Spemann H. 1938. Embryonic development and induction. New Ha-ven: Yale University Press.
Standley HJ, Zorn AM, Gurdon JB. 2001. eFGF and its mode of actionin the community effect during Xenopus myogenesis. Development128:1347–1357.
Standley HJ, Zorn AM, Gurdon JB. 2002. A dynamic requirement forcommunity interactions during Xenopus myogenesis. Int J Dev Biol46:279–283.
Strigini M, Cohen SM. 1999. Formation of morphogen gradients in theDrosophila wing. Semin Cell Dev Biol 10:335–344.
Strigini M, Cohen SM. 2000. Wingless gradient formation in theDrosophila wing. Curr Biol 10:293–300.
Symes K, Smith JC. 1987. Gastrulation movements provide an earlymarker of mesoderm induction in Xenopus laevis. Development101:339–349.
Tabata T. 2001. Genetics of morphogen gradients. Nat Rev Genet2:620–630.
Takahashi S, Yokota C, Takano K, Tanegashima K, Onuma Y, Goto J,Asashima M. 2000. Two novel nodal-related genes initiate earlyinductive events in Xenopus Nieuwkoop center. Development 127:5319–5329.
Tanegashima K, Yokota C, Takahashi S, Asashima M. 2000. Expres-sion cloning of Xantivin, a Xenopus lefty/antivin-related gene, in-volved in the regulation of activin signaling during mesoderm in-duction. Mech Dev 99:3–14.
Teleman AA, Cohen SM. 2000. Dpp gradient formation in the Dro-sophila wing imaginal disc. Cell 103:971–980.
Teleman AA, Strigini M, Cohen SM. 2001. Shaping morphogen gra-dients. Cell 105:559–562.
Thomsen G, Woolf T, Whitman M, Sokol S, Vaughan J, Vale W,Melton DA. 1990. Activins are expressed early in Xenopus embry-ogenesis and can induce axia mesoderm and anterior structures.Cell 63:485–493.
Tickle C. 1999. Morphogen gradients in vertebrate limb development.Semin Cell Dev Biol 10:345–351.
Tree DR, Shulman JM, Rousset R, Scott MP, Gubb D, Axelrod JD.2002. Prickle mediates feedback amplification to generate asym-metric planar cell polarity signaling. Cell 109:371–381.
Turing AM. 1952. The chemical basis of morphogenesis: a reaction-diffusion model for development. Philos Trans R Soc Lond B Biol Sci237:37–72.
Vargesson N, Kostakopoulou K, Drossopoulou G, Papageorgiou S,Tickle C. 2001. Characterisation of hoxa gene expression in thechick limb bud in response to FGF. Dev Dyn 220:87–90.
Vervoort M. 2000. hedgehog and wing development in Drosophila: amorphogen at work? Bioessays 22:460–468.
Vodicka MA, Gerhart JC. 1995. Blastomere derivation and domains ofgene expression in the Spemann Organizer of Xenopus laevis. De-velopment 121:3505–3518.
Vogel AM, Gerster T. 1999. Promoter activity of the zebrafishbhikhari retroelement requires an intact activin signaling pathway.Mech Dev 85:133–146.
Waddington CH. 1938. Regulation of amphibian gastrulae with addedectoderm. J Exp Biol 15:377–381.
Wardle FC, Angerer LM, Angerer RC, Dale L. 1999. Regulation ofBMP signaling by the BMP1/TLD-related metalloprotease, SpAN.Dev Biol 206:63–72.
Watabe T, Kim S, Candia A, Rothbacher U, Hashimoto C, Inoue K,Cho KW. 1995. Molecular mechanisms of Spemann’s organizer for-mation: conserved growth factor synergy between Xenopus andmouse. Genes Dev 9:3038–3050.
Weeks DL, Melton DA. 1987. A maternal mRNA localized to thevegetal hemisphere in Xenopus eggs codes for a growth factor re-lated to TGF-beta. Cell 51:861–867.
Weyer CJ, Nieuwkoop PD, Lindenmayer A. 1977. A diffusion modelfor mesoderm induction in amphibian embryos. Acta Biotheor 26:164–180.
Whitman M. 2001. Nodal signaling in early vertebrate embryos:themes and variations. Dev Cell 1:605–617.
Wilson PA, Hemmati-Brivanlou A. 1995. Induction of epidermis andinhibition of neural fate by Bmp-4. Nature 376:331–333.
Wilson PA, Lagna G, Suzuki A, Hemmati-Brivanlou A. 1997. Concen-tration-dependent patterning of the Xenopus ectoderm by BMP4and its signal transducer Smad1. Development 124:3177–3184.
Wilson PA, Melton DA. 1994. Mesodermal patterning by an inducergradient depends on secondary cell-cell communication. Curr Biol4:676–686.
Witkowski J. 1985. The hunting of the organizer: an episode in bio-chemical embryology. TIBS 10:379–381.
Witkowski JA. 1987. Optimistic analysis—chemical embryology inCambridge 1920–42. Med Hist 31:247–268.
Wolpert L. 1969. Positional information and the spatial pattern ofcellular differentiation. J Theor Biol 25:1–47.
Wolpert L. 1971. Positional information and pattern formation. CurrTop Dev Biol 6:183–224.
Wolpert L. 1989. Positional information revisited. Development 107:3–12.
Wolpert L. 1996. One hundred years of positional information. TrendsGenet 12:359–364.
Wolpert L. 1999. Vertebrate limb development and malformations.Pediatr Res 46:247–254.
Xu RH, Kim J, Taira M, Sredni D, Kung H. 1997. Studies on the roleof fibroblast growth factor signaling in neurogenesis using conju-gated/aged animal caps and dorsal ectoderm-grafted embryos.J Neurosci 17:6892–6898.
Yang CH, Axelrod JD, Simon MA. 2002. Regulation of Frizzled byfat-like cadherins during planar polarity signaling in the Drosoph-ila compound eye. Cell 108:675–688.
Yates K, Pate E. 1989. A cascading development model for amphibianembryos. Bull Math Biol 51:549–578.
Yeo C, Whitman M. 2001. Nodal signals to Smads through Cripto-dependent and Cripto-independent mechanisms. Mol Cell 7:949–957.
Yuh CH, Bolouri H, Davidson EH. 1998. Genomic cis-regulatory logic:experimental and computational analysis of a sea urchin gene.Science 279:1896–1902.
Zecca M, Basler K, Struhl G. 1996. Direct and long-range action of awingless morphogen gradient. Cell 87:833–844.
Zhang J, Houston DW, King ML, Payne C, Wylie C, Heasman J. 1998.The role of maternal VegT in establishing the primary germ layersin Xenopus embryos. Cell 94:515–524.
Zimmerman LB, DeJesus-Escobar JM, Harland RM. 1996. The Spe-mann Organizer signal noggin binds and inactivates bone morpho-genetic protein 4 Cell 86:599–606.
Zoltewicz JS, Gerhart JC. 1997. The Spemann organizer of Xenopus ispatterned along its anteroposterior axis at the earliest gastrulastage. Dev Biol 192:482–491.
408 GREEN





























