Embed Size (px)
Citation preview

Rädler / Mathias SS2011
1
Monomer/ Oligomere Polymer/Aggregat
Zucker Glucose, Energie, Zellmarker
PolysaccharideStrukturbausteine
Fettsäuren Lipide, FettEnergiespeicher
MembranenKompartmentisierung
Nukleinsäuren ATP, cAMP
Energie, Signalsubstanzen
RNA, DNAInformationsspeicher
Aminosäuren PeptideHormone
Proteine, Enzyme. Motoren, molekulare
Erkennung

2
Ribosomale Proteinsynthese
Rädler / Mathias SS2011
•mehrere rRNA Stränge•> 50 Proteine•Durchmesser ~21nm

Rädler / Mathias SS2011
3
in vitro:

Rädler / Mathias SS2011
4
Length CouplingEfficiency
Coupling Efficiency
Coupling Efficiency
Coupling Efficiency
0 0.995 0.99 0.98 0.97 0.965 0.98 0.95 0.92 0.89 0.8510 0.96 0.91 0.83 0.76 0.6915 0.93 0.87 0.75 0.65 0.5620 0.91 0.83 0.68 0.56 0.4625 0.89 0.79 0.62 0.48 0.3830 0.86 0.75 0.56 0.41 0.3135 0.84 0.71 0.50 0.36 0.2540 0.82 0.67 0.45 0.30 0.2045 0.80 0.63 0.41 0.26 0.1750 0.78 0.60 0.37 0.22 0.1455 0.76 0.58 0.34 0.19 0.1160 0.74 0.55 0.30 0.17 0.0965 0.73 0.53 0.27 0.14 0.0770 0.71 0.50 0.25 0.12 0.06
Coupling Efficiency Vs. Peptide LengthPeptide

Rädler / Mathias SS2011
5

Rädler / Mathias SS2011
6

7
Different graphical representations of the same protein
Rädler / Mathias SS2011

Rädler / Mathias SS2011
8
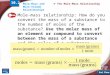
Faltungsproblem
Konformation eines Proteins als Random Walk:
Gitter-Modell: Kleines Protein mit 100 Aminosäuren
=> Mögliche Konformationen: 3100≈ 1030
Interne Dynamik typ ns
⇒Zeit, um alle möglichen
Kombinationen durchzuspielen ≈ 1021
sVergleiche: Alter des Universums ≈ 1020 s !
Mother nature has no folding probem,but we do!
Rädler / Mathias SS2011
9
Rädler / Mathias SS2011
1010
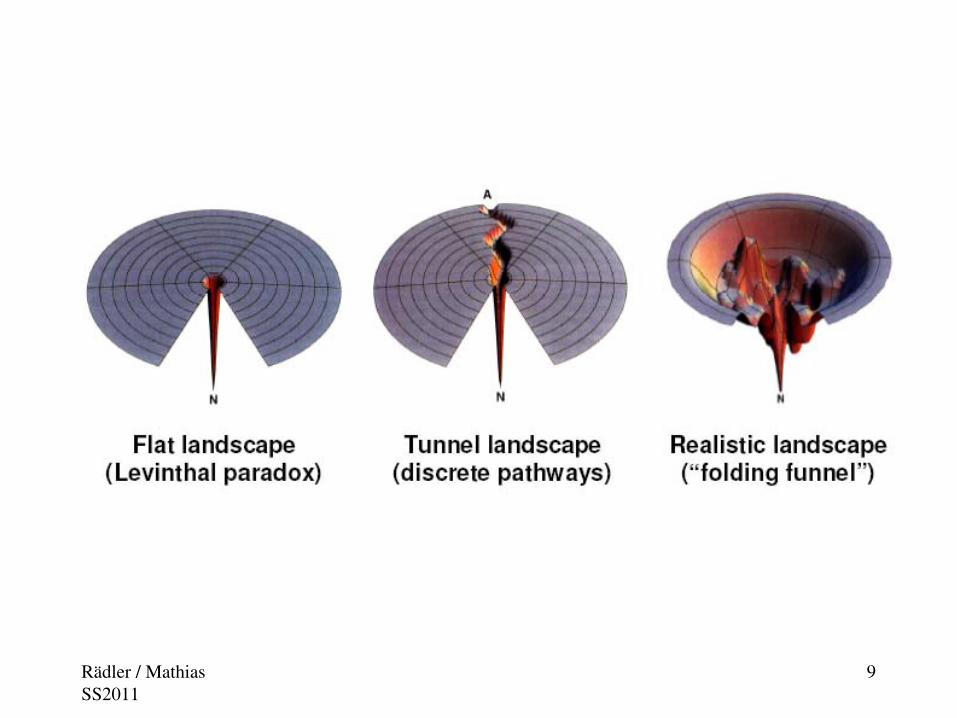
Protein Folding Related Diseases

1111Rädler / Mathias SS2011

Proteinfunktionen
• Zellgerüst• Motoren• Sensoren• Photosynthese• Enzymatische Katalyse• Ionenkanäle• .....Rädler / Mathias SS2011
12

Stäbchenzelle
Beispiel: Die Sehkaskade
Rädler / Mathias SS2011
13

Rädler / Mathias SS2011
14Proc Natl Acad Sci U S A. 2002 April 30; 99(9): 5982–5987.

Rädler / Mathias SS2011
15

Rädler / Mathias SS2011
16

Photozyklus
Rädler / Mathias SS2011
17

Rädler / Mathias SS2011
18

Rädler / Mathias SS2011
19
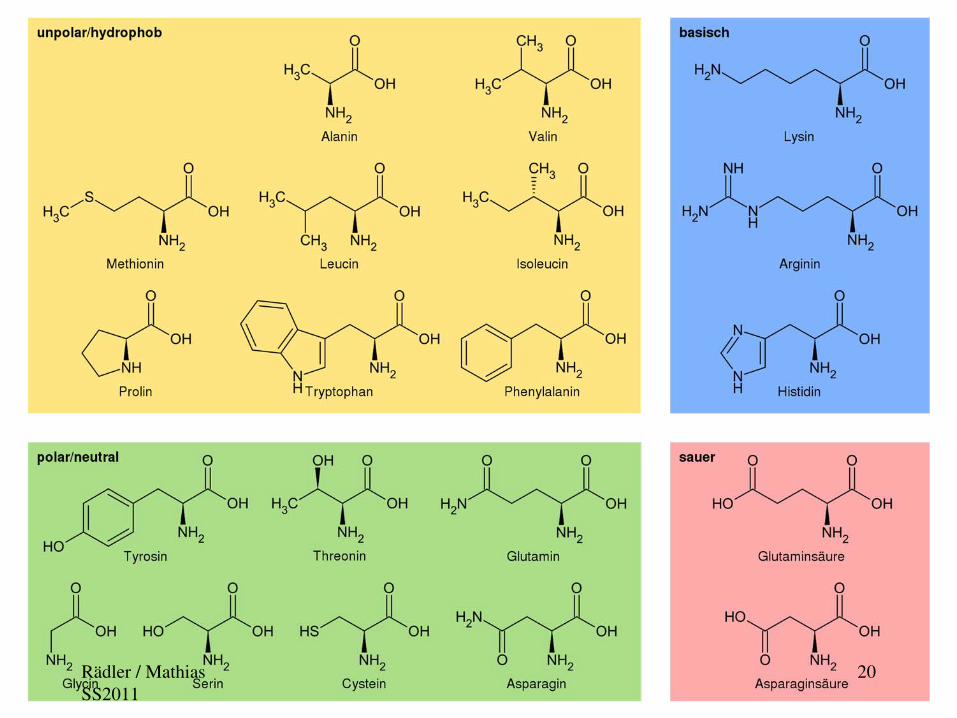
Rädler / Mathias SS2011
20

Chemical modifications and processing alter the biological activity of proteins
Rädler / Mathias SS2011
21

Rädler / Mathias SS2011
22Lö 27, 28, 31M 567/ 568

Lö 28
Rädler / Mathias SS2011
23

Rädler / Mathias SS2011
24

Rädler / Mathias SS2011
25

Disulfidbrücken stabilisieren Proteine (können in seltenen Fällen auch Knoten bilden)
HUMAN AGOUTI RELATED PROTEIN
Rädler / Mathias SS2011
26

Rädler / Mathias SS2011
27Lö 38

Rädler / Mathias SS2011
28Lö 28

Rädler / Mathias SS2011
29

Rädler / Mathias SS2011
30

Rädler / Mathias SS2011
31

Table 3-2. Covalent and Noncovalent Chemical Bonds
Strength (kcal/mole)*
Bond Type Length (nm) In Vacuum In Water
Covalent 0.15 90 90Ionic 0.25 80 3Hydrogen 0.30 4 1van der Waals attraction 0.35 0.1 0.1(per atom)
*. The strength of a bond can be measured by the energy required to break it, here given in kilocalories per mole (kcal/mole). ( One kilocalorie is the quantity of energy needed to raise the temperature of 1000 g of water by 1C. An alternative unit in wide use is the kilojoule, kJ, equal to 0.24 kcal.) Individual bonds vary a great deal in strength, depending on the atoms involved and their precise environment, so that the above values are only a rough guide. Note that the aqueous environment in a cell will greatly weaken both the ionic and the hydrogen bonds between nonwater molecules (Panel 3-1, pp. 92-93). The bond length is the center-to-center distance between the two interacting atoms; the length given here for a hydrogen bond is that between its two nonhydrogen atoms.
Rädler / Mathias SS2011
32

Rädler / Mathias SS2011
33
Steric limitations on the bondangles in a polypeptide chain. (A) Each amino acid contributesthree bonds (colored red) to itspolypeptide chain. The peptide bondis planar ( gray shading) and doesnot permit rotation. By contrast, rotation can occur about the C α-C bond, whose angle of rotation iscalled psi (ψ), and about the N-C α bond, whose angle of rotation iscalled phi (). The R group denotesan amino acid side chain. (B) The conformation of the main-chainatoms in a protein is determined byone pair of phi and psi angles foreach amino acid; because of stericcollisions within each amino acid, most pairs of phi and psi angles do not occur. In this so-calledRamachandran plot, each dotrepresents an observed pair of anglesin a protein. (B, from J. Richardson, Adv. Prot. Chem. 34:174-175, 1981.)

Rädler / Mathias SS2011
34Lovell et al. 2003 Proteins 50:437

Glyzin, Pre-Prolin, Prolin
Rädler / Mathias SS2011
35BMC Struct Biol. 2005; 5: 14. doi: 0.1186/1472-6807-5-14.

36
A β−sheet is a commonstructure formed byparts of the polypeptidechain in globularproteins. At the top, a domain of 115 aminoacids from an immunoglobulin moleculeis shown; it consists of a sandwich-like structure oftwo β− sheets, one ofwhich is drawn in color. At the bottom, a perfectantiparallel β− sheet isshown in detail, with theamino acid side chainsdenoted R. Note thatevery peptide bond ishydrogen-bonded to a neighboring peptide bond. The actual sheet structuresin globular proteins areusually less regular thanthe β− sheet shown here, and most sheets areslightly twistedRädler / Mathias
SS2011
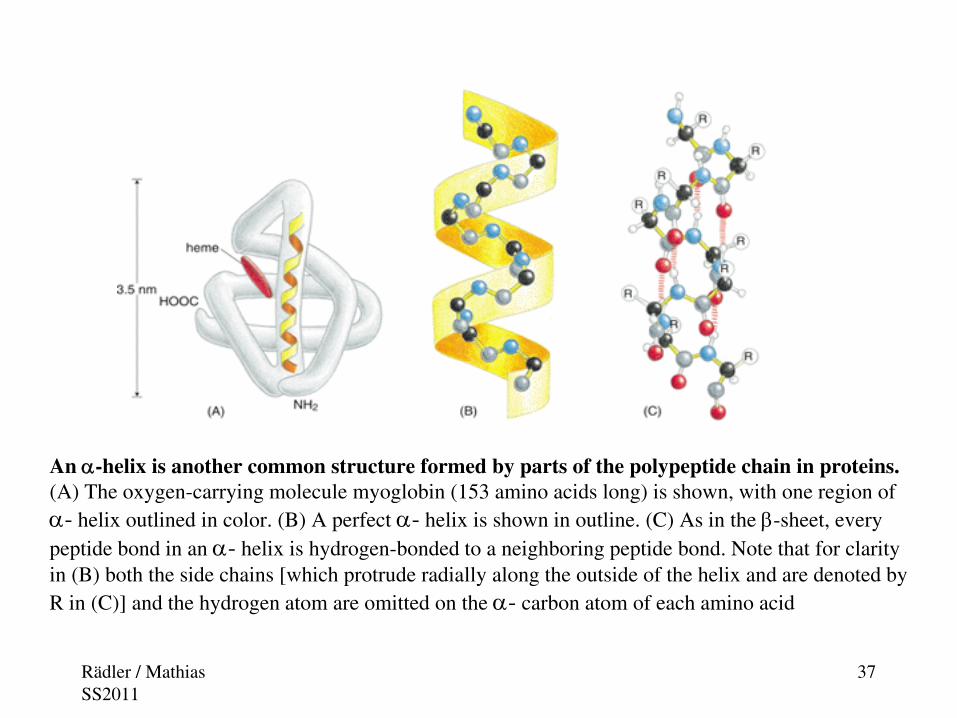
An α-helix is another common structure formed by parts of the polypeptide chain in proteins. (A) The oxygen-carrying molecule myoglobin (153 amino acids long) is shown, with one region of α- helix outlined in color. (B) A perfect α- helix is shown in outline. (C) As in the β-sheet, every peptide bond in an α- helix is hydrogen-bonded to a neighboring peptide bond. Note that for clarity in (B) both the side chains [which protrude radially along the outside of the helix and are denoted by R in (C)] and the hydrogen atom are omitted on the α- carbon atom of each amino acid
Rädler / Mathias SS2011
37

Rädler / Mathias SS2011
38
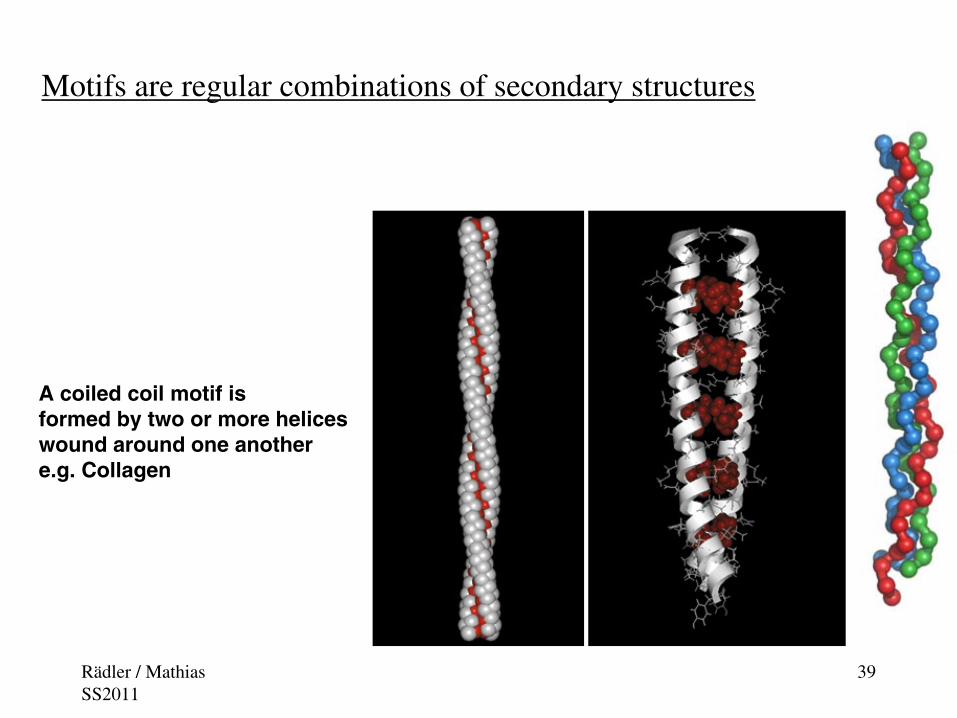
Motifs are regular combinations of secondary structures
A coiled coil motif is formed by two or more heliceswound around one anothere.g. Collagen
Rädler / Mathias SS2011
39

Three levels of organization of a protein. The three-dimensional structure of a protein can be described in terms of different levels of folding, each of which is constructed from the preceding one in hierarchical fashion. These levels are illustrated here using the catabolite activator protein (CAP), a bacterial gene regulatory protein with two domains. When the large domain binds cyclic AMP, it causes a conformational change in the protein that enables the small domain to bind to a specific DNA sequence. The amino acid sequence is termed the primary structure and the first folding level the secondary structure. As indicated under the brackets at the bottom of this figure, the combination of the second and third folding levels shown here is commonly termed the tertiary structure, and the fourth level (the assembly of subunits) the quaternary structure of a protein
Rädler / Mathias SS2011
40

The information for protein folding is encoded in the sequence
Rädler / Mathias SS2011
41

Primary and secondary structure in hemagglutinin
Rädler / Mathias SS2011
42

Tertiary and quaternary structure in hemagglutinin
Rädler / Mathias SS2011
43

Up To Date No Unified Folding Theory
Rädler / Mathias SS2011
44

Rädler / Mathias SS2011
45
Folding Funnel

Folding of proteins in vivo is promoted by chaperones
Rädler / Mathias SS2011
46