Embed Size (px)
Citation preview

Cancer Therapy: Preclinical
Monoclonal Antibody against Cell Surface GRP78 as a NovelAgent in Suppressing PI3K/AKT Signaling, Tumor Growth,and Metastasis
Ren Liu1, Xiuqing Li1, Wenming Gao1, Yue Zhou1, Shiuan Wey2, Satyajit K. Mitra5, Valery Krasnoperov5,Dezheng Dong4, Shuanglong Liu3, Dan Li3, Genyuan Zhu2, Stan Louie4, Peter S. Conti3, Zibo Li3,Amy S. Lee2, and Parkash S. Gill1
AbstractPurpose: The ER chaperone GRP78 translocates to the surface of tumor cells and promotes survival,
metastasis, and resistance to therapy. An oncogenic function of cell surface GRP78 has been attributed
to the activation of the phosphoinositide 3-kinase (PI3K) pathway. We intend to use a novel anti-
GRP78 monoclonal antibody (MAb159) to attenuate PI3K signaling and inhibit tumor growth and
metastasis.
ExperimentalDesign:MAb159was characterized biochemically. Antitumor activity was tested in cancer
cell culture, tumor xenograft models, tumor metastasis models, and spontaneous tumor models. Cancer
cells and tumor tissueswere analyzed for PI3K activity.MAb159washumanized and validated for diagnostic
and therapeutic application.
Results:MAb159 specifically recognized surface GRP78, triggered GRP78 endocytosis, and localized
to tumors but not to normal organs in vivo. MAb159 inhibited tumor cell proliferation and enhanced
tumor cell death both in vitro and in vivo. In MAb159-treated tumors, PI3K signaling was inhibited
without compensatory MAPK pathway activation. Furthermore, MAb159 halted or reversed tumor
progression in the spontaneous PTEN–loss-driven prostate and leukemia tumor models, and inhibited
tumor growth and metastasis in xenograft models. Humanized MAb159, which retains high affinity,
tumor specific localization, and the antitumor activity, was nontoxic in mice, and had desirable
pharmacokinetics.
Conclusions: GRP78-specific antibody MAb159 modulates the PI3K pathway and inhibits tumor
growth and metastasis. Humanized MAb159 will enter human trials shortly. Clin Cancer Res; 1–10.
�2013 AACR.
IntroductionCancer cells are characterized by metabolic alterations
and the tumor microenvironment is often marked withimpaired blood flow and hypoxia, all of which can elicitendoplasmic reticulum (ER) stress. Tumor cells adapt tothese adverse conditions by activating the unfolded protein
response (UPR), with induction of GRP78 as a majorprosurvival arm of the UPR signaling pathways (1, 2).GRP78, also referred to as BiP/HSPA5, is a 78-kD glu-cose–regulated protein with potent antiapoptotic proper-ties that plays critical roles in cancer cell survival, tumorprogression, metastasis, and resistance to therapy (3–5).Knockdown of GRP78 by siRNA in cancer cells as well astumor-associated endothelial cells reduced their prolifer-ative rate and sensitized them to chemotherapeutic treat-ment (6, 7). Genetically altered GRP78 mouse modelsfurther demonstrated the critical role of GRP78 in cancerin vivo. For instance, GRP78 haploinsufficiency delayedtumor development, growth, and inhibited metastasis (8,9). Inmiceharboringbiallelic conditional knockout of bothGRP78 and tumor suppressor PTEN in the prostate epithe-lium, prostate tumorigenesis was potently arrested (10). Inaddition, inducible heterozygous knockout ofGRP78 in thehematopoietic system suppresses PTEN-null leukemogen-esis with no harmful effect on hematopoiesis (11). Strik-ingly, in both the prostate and leukemiamodels, phosphoi-nositide 3-kinase (PI3K)/AKT signaling resulting from the
Authors' Affiliations: Departments of 1Pathology, 2Biochemistry andMolecular Biology, and 3Radiology; 4School of Pharmacy, University ofSouthern California Keck School of Medicine, USC Norris ComprehensiveCancer Center; and 5VasGene Therapeutics Inc., Los Angeles, California
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clincancerres.aacrjournals.org/).
R. Liu and X. Li contributed equally to this work.
Both A.S. Lee and P.S. Gill are co-senior authors.
Corresponding Author: Parkash S. Gill, University of Southern California,1441 Eastlake Ave, RM NOR 6332, Los Angeles, CA 90033. Phone: 323-865-3909; Fax: 323-865-0092; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-13-1106
�2013 American Association for Cancer Research.
ClinicalCancer
Research
www.aacrjournals.org OF1
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

loss of PTEN was greatly impaired with only partial loss ofGRP78 (10, 11). Collectively, these studies establish GRP78as a novel regulator of the PI3K/AKT oncogenic signalingand a target for anticancer therapy.
Although traditionallyGRP78 has been regarded as an ERlumenal protein, evidence has accumulated that a fractionof GRP78 can exist on the plasmamembrane of specific celltypes (12–14) and that ER stress actively promotes cellsurface localization of GRP78 (15). Whereas the physio-logic function of GRP78 on the cell surface is still emerging,recent studies show that cell surface GRP78 forms com-plexes with specific protein partners, regulating both pro-liferation and viability (14, 16), suggesting that cell surfaceGRP78 presents an opportunity for therapeutic targeting(17–19).
Here we report the generation of a monoclonal antibody(mAb; MAb159) against human GRP78 that shows antitu-mor activity with no toxicity and also inhibits PI3K signal-ing. This antibody opens up a unique opportunity to studyboth the biology of cell surface GRP78 and its therapeuticpotential.
Materials and MethodsAntibodies, reagents, and cell lines
A549, HT29, Colo205, MCF7, 4T1, and 293T cell lineswere obtained from the American Type Culture Collec-tion. C4-2B cell was kindly provided by Michael Stallcup(University of South California), and H249 was kindlyprovided by Dr. Ravi Salgia of the University of Chicago.The generation of the B16-Fluc-A1 melanoma cell line hasbeen described previously (9). All these cells were prop-agated in RPMI-1640 supplemented with 10% FBS, 100units/mL of penicillin, and 100 mg/mL streptomycin fromCellgro. The CE1 cell line was kindly provided by Dr.Pradip Roy-Burman and cultured as previously described(20). These cell lines have been validated by HLA typingand molecular phenotyping relative to the respective
primary tumors. Detailed information about antibodiesand reagents used in this study can be found in Supple-mentary Materials and Methods.
Generation of mAbs and affinity analysisThe procedure for generation of mAbs is described in
Supplementary Materials and Methods. The affinity ofmAbs to antigen was determined by Scatchard assay asdescribed previously (21).
In vivo and ex vivo near-infrared fluorescence imagingH249 tumor-bearing mice were injected with Cy5.5-
labeled humanized MAb159 or control antibody. In vivofluorescence imaging was performed using the XenogenLumina XR Imaging System and analyzed using the IVISLiving Imaging 3.0 software. Twenty-eight hours after injec-tion, the tumors and organs were harvested for ex vivofluorescence imaging. Please see Supplementary Materialsand Methods for detailed procedure.
PTEN-null modelsThe prostate-specific PTEN knockout model has been
described previously (22). Mice were treated with MAb159,and prostate tumors were monitored by luminescenceimaging (xenogen). The Pten (floxed/floxed); Mx-1 cre leu-kemic model and the protocols for flow cytometry foranalysis of leukemic blasts and peripheral blood countshave been described previously (11). Please see Supple-mentary Materials and Methods for detailed procedure.
Murine tumor xenograft modelsThe procedures for murine tumor xenograft studies and
immunohistochemical analysis were as described previous-ly (21) and are further described in Supplementary Materi-als and Methods.
Statistical AnalysisThe statistical significance of differences in different
samples or groups was determined using an unpairedtwo-tailed Student t test. Results were considered signifi-cantly different if the P value was less than 0.05.
ResultsGeneration of mAbs specific to surface GRP78
We immunized mice with a hexahistidine-tagged andsecreted form of human GRP78 (Supplementary Fig.S1A), and screened a panel of mAbs capable of bindingthe native form of GRP78. The criteria for the desirableantibody were high-affinity binding to native cell surfaceGRP78 and the ability to endocytose. MAb159 fulfilledthese requirements. MAb159 is highly specific for GRP78,as the binding of MAb159 to GRP78 was completelyabolished by purified soluble GRP78 protein (Supple-mentary Fig. S1B). In addition, we performed an immu-noprecipitation study with MAb159 and confirmed thatGRP78 can be pulled down from cell lysate using massspectrometric analysis (data not shown). MAb159 hasno cross-reactivity to GRP78’s closest paralog HSP70
Translational RelevanceA major obstacle in cancer therapy is the damage to
normal organs by conventional chemotherapy andradiotherapy. This highlights the need for therapy thatspecifically targets and kills cancer cells while sparingnormal cells. One emerging target is GRP78, which ispreferentially expressed on cancer cell surface and pro-motes tumor cell survival and metastasis. We havedeveloped a monoclonal antibody MAb159 againstGRP78 to target surface GRP78 and block its oncogenicfunctions. MAb159 effectively images tumors in vivo,suppresses PI3K/AKT signaling, induces apoptosis, andinduces tumor regression in xenografts and spontaneoustumor models. The humanized MAb159 retains itsGRP78-binding affinity and efficacy and is nontoxic tonormal organs. We plan to initiate human clinical trialshortly.
Liu et al.
Clin Cancer Res; 2013 Clinical Cancer ResearchOF2
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

(Supplementary Fig. S1C), but it recognizes mouseGRP78, which is 99% conserved in amino acids withhuman GRP78 (Supplementary Fig. S1D). Using scatch-ard analysis, we have determined that MAb159 has highaffinity to human GRP78 (Kd ¼ 1.7 nmol/L; Supplemen-tary Fig. S1E), hence suitable for therapeutic develop-ment. When incubated with cultured cells at 4�C,MAb159 bound to the cell surface of cancer cells but notto normal human dermal fibroblasts (Fig. 1A, left). Underglucose-starvation conditions, which mimicked nutrientdeprivation in the tumor microenvironment, greateramount of MAb159 was recruited to cancer cell surface.This is consistent with the previous findings that surfaceGRP78 significantly increases when the cell is under stress(14, 15). In addition, we determined whether MAb159preferably localizes to tumor and not normal organs bytracking biotin-labeled MAb159 in HT29 xenografttumor–bearing mice in vivo. Biotin-MAb159 was onlyseen in the tumor, not in the normal organs, includingheart, liver, and kidney (Fig. 1A, right).When incubated with cells at 37�C, MAb159 underwent
endocytosis and was localized to the intracellular clathrin-coated endosomes compared with a fine ring-like appear-ance at the cell surface when incubated at 4�C (Fig. 1B, top;Supplementary Fig. S2A). We next examined the surfaceGRP78 level with a second antibody and found that surfaceGRP78wasmarkedly reduced afterMAb159 treatment (Fig.1B, bottom). Moreover, this reduction of GRP78 can berevertedwith chlorpromazine (Supplementary Fig. S2B), aninhibitor specific for clathrin-mediated endocytosis (23).These data implicate that MAb159 led to internalization ofsurface GRP78.
MAb159 induces tumor cell apoptosis and inhibitsPI3K signaling
The effect of MAb159 on tumor cells was first tested invitro. Before incubation with MAb159, cells from the breastcarcinoma cell line MCF7 and colon cancer cell line HT29were cultured in glucose-free medium to enhance surfaceGRP78 level and thus the efficacy of the antibody. MAb159significantly reduced the cell viability in bothMTT (Fig. 1C,top) and clonogenic assay (Supplementary Fig. S3A), cor-relating with increased apoptosis as determined by terminaldeoxynucleotidyl transferase–mediated dUTP nick endlabeling (TUNEL; Fig. 1C, bottom) and M30 apoptosisassay (Supplementary Fig. S3B). In addition, we have mea-sured activated caspase levels and showed that MAb159treatment activates caspases 8 and 9 (Supplementary Fig.S3C), which represent the activation of extrinsic and intrin-sic apoptotic pathways, respectively. This suggests that, atleast in part, the loss of cell viability was initiated at the cellmembrane.
The PI3K signaling pathway regulates many biologicevents in the cells, including cell survival, and GRP78 haspreviously been shown to modulate its activity. Therefore,we examined whether MAb159-treated cell–surface-GRP78-expressing tumor cells have alterations in PI3Kactivity as measured by the changes in phosphorylated AKTand S6 levels. We found, indeed, that both phosphorylatedAKT and S6 levels were reduced in antibody-treated cellscompared with the controls (Fig. 1D). Because MAb159-induced GRP78 endocytosis led to the loss of cell surfaceGRP78 and inhibition of pAKT signaling, we anticipate thatblocking GRP78 endocytosis would antagonize the activityof MAb159. We, however, were unable to conduct this
Figure 1. Characterization ofmAb targeting specifically to surfaceGRP78. A, specificity of GRP78 antibodyMAb159. Left, live cells were stainedwithMAb159at 4�C. Surface GRP78 staining (green) presents on cancer cells C4-2B and MCF7, but not normal human dermal fibroblast (NHFD). Glucose starvationfor 2 days significantly increased surface GRP78 on MCF7 cells (bottom right). Right, biotinylated MAb159 (50 mg) was administered intravenously totwo HT29 tumor-bearing mice, mice were sacrificed after 6 hours, tissues were harvested, and MAb159 was localized with fluorochrome-conjugatedstreptavidin (green). MAb159 was only detected in tumor, not in normal organs. Nuclei were counterstained with 40, 6-diamidino-2-phenylindole(DAPI; blue). Scale bar, 20 mm.B, top, biotinylatedMAb159was incubatedwith glucose-starvedMCF7 cells for 1 hour at 4�Cor 37�C. Localization ofMAb159is shown in green (confocal image). Endocytosis was observed only at 37�C. Bottom, MAb159 or control IgG was incubated with glucose-starvedMCF7 cells for 48 hours at 37�C. Surface GRP78 was detected using a polyclonal antibody (Clone H129; Santa Cruz Biotechnology) without fixation andpermeabilization at 4�C. Scale bar, 20 mm. C, MCF7 and HT29 cells were glucose starved and treated with control IgG or MAb159 for 5 days. Top,cell viability was determined with MTT assay. Data are presented as mean � SE (n ¼ 4). Bottom, MAb159 triggered apoptosis in glucose-starved MCF7cells determined by TUNEL assay (green). Nuclei were counterstained with DAPI (blue). Scale bar, 100 mm. D, glucose-starved HT29 cells were treatedwith 50 mg/mL MAb159 for 3 days. Whole-cell lysate immunoblotting shows that MAb159 decreased phosphorylated AKT (pAKT) and S6 (pS6) levels. Threeindependent experiments were performed. ImageJ software (NIH) was used for quantification of relative pAKT and pS6 levels. �,P < 0.02, as determined by anunpaired two-tailed Student t test.
GRP78 Antibody Inhibits PI3K and Tumor Growth
www.aacrjournals.org Clin Cancer Res; 2013 OF3
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

experiment because chlorpromazine by itself reduces pAKTlevels and causes cellular toxicity (data not shown; ref. 24).
The above results indicate that MAb159 inhibits the PI3Kpathway and induces cell apoptosis, which is consistentwith the recent indication that cell surfaceGRP78may be anupstream regulator of PI3K/AKT but not mitogen-activatedprotein kinase (MAPK) signaling (14). We, thus, wished todetermine if there is a direct interaction between cell surfaceGRP78 and PI3K components.We expressed a FLAG-taggedGRP78 with KDEL deletion for enhanced translocation tothe cell surface (15), labeled the cell surface proteins withbiotin, and purified the biotin-labeled proteins on mono-meric avid beads. GRP78 was immunoprecipitated fromthis pool of surface proteins, and its interacting partnerswere detected by Western blot analysis. As seen in Supple-mentary Fig. S4A, GRP78 formed a complex with p85, theregulatory subunit of PI3K, but not with ERK1/2 or a cellsurface–specific protein EphB2. We further confirmed thisinteraction under endogenous expression conditions. Sur-face GRP78 was induced with thapsigargin (15), and cellsurface proteins were enriched using the biotin–avidinsystem. Coimmunoprecipitation of surface GRP78 andp85 was achieved with either GRP78 or p85 antibody(Supplementary Fig. S4B). These results provide the firstevidence that surface GRP78 binds to the PI3K componentand suggest that surface GRP78 regulates PI3K signalingthrough direct complex formation with the PI3K subunits.
MAb159 inhibits tumor growth and causes tumorregression in various xenograft tumor models
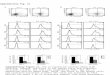
The efficacy of the GRP78 antibody MAb159 in vivo wasexamined in various tumor xenograft models includingHT29 (colon cancer), H249 (small cell lung carcinoma),and A549 (lung adenocarcinoma). These cells have rela-tively high (4.6%–9.4%; Supplementary Fig. S5) surfaceGRP78 expression compared with normal cells (15).MAb159 treatment led to 50%, 58%, and 78% tumorgrowth inhibition in these models, respectively (Fig. 2A).
To further test if the combination of GRP78-targetedtherapy and conventional chemotherapy leads to greaterefficacy, we combined MAb159 and irinotecan in a coloncancer xenograft model. The colon cancer model was cho-sen because GRP78 overexpression was reported to beassociated with colorectal carcinogenesis (25). Whenadministered alone, MAb159 inhibited tumor growth by54% compared with the control group. Irinotecan mono-therapy inhibited tumor growth by 85%. The combinationtherapy caused tumor regression to 47% of the startingtumor volume (Fig. 2A).
MAb159 reduces proliferation, induces apoptosis,impairs tumor vasculature, and inhibits PI3K signalingin tumor xenografts
At the end of the xenograft experiments, tumorswere harvested for analysis. In the MAb159-treated group,the proliferation index (Ki67 staining) was markedlyreduced, apoptosis (TUNEL assay) was significantlyincreased (Fig. 2B), and vessel density (CD31 staining) had
a modest decrease (Supplementary Fig. S6A). In addition,MAb159 treatment led to a marked reduction in phosphor-ylated AKT, mTOR, and S6 (Fig. 2B–D; Supplementary Fig.S6B), indicating inhibition of PI3K signaling. Meanwhile,there was no increase in phosphorylated ERK1/2 and Src(Fig. 2C and D), indicating that these compensatory path-ways, often induced with PI3K inhibitors and resistance totherapy,were not activated. In addition, it is notable that thesystemic administration of the antibody was well tolerated,as measured by the animal food intake, body weight (datanot shown), and microscopic examination of vital organs(Supplementary Fig. S7).
MAb159 inhibits tumor metastasisGRP78 promotes growth of blood vessels in the tumor
accompanied by tumor growth and metastasis (9). Target-ing surface GRP78 with a peptide conjugated to proapop-totic molecules inhibits metastases (19). Therefore, wetested MAb159 in an orthotopic tumor model using mousebreast adenocarcinoma cell 4T1 (26, 27). Here, 4T1 cellswere implanted into the #4 fat pads of isogenic BALB/cmice,whichwere treatedbiweeklywithMAb159 at a dose of10 mg/kg or with saline. Within 7 days, MAb159 inhibitedprimary tumor growth and secondary metastasis to #9 fatpads (n ¼ 2; Supplementary Fig. S8A). After 13 days,metastatic tumors were present on the liver surface ofcontrol animals (n ¼ 3) but not in MAb159-treated mice(n¼ 3; Fig. 3A, left). Further focal liver necrosis was seen incontrol but not in MAb159-treated animals (data notshown;Fig. 3A). By day 14 in all control mice, both primarytumor and #9 fat pad secondary tumors had become largeand invaded through the body wall into the underlyingperitoneal cavity. In comparison, MAb159-treated miceexhibited complete or near-complete primary tumor regres-sion (data not shown; Supplementary Fig. S8B). There wasno visible contralateral metastasis in any of the MAb159-treated animals. Histologic evaluation of lung metastasisshowed that the majority of the lung volume of controlanimals was occupied by metastatic breast cancer thatresulted in internal hemorrhaging in 50% of control lungs(Fig. 3A, right). Lung metastasis was significantly inhibitedby MAb159 treatment (Fig. 3B), and there was no evidenceof internal hemorrhaging in any of the M159-treated ani-mals (Fig. 3A, right). Analysis of breast tumor tissues showsthat MAb159 significantly reduced the pS6 level, indicativeof inhibition of PI3K signaling (Supplementary Fig. S8C).
We also examined the effect ofMAb159 on themetastaticgrowth of a syngeneic melanoma cancer cell line (9). B16-Fluc-A1 melanoma cells stably expressing luciferase wereinjected intravenously. Tumor metastasis and progressionin the lungs was monitored live with a whole-animalluminescence imaging system. MAb159 treatment signifi-cantly reduced the formation of lung tumors (Fig. 3C, left).At the end of the experiment, lungs were harvested andpigmented tumors were observed on the lung surface.Compared with the control group, the lungs fromMAb159-treated mice had significantly fewer tumors (Fig.3C, right).
Liu et al.
Clin Cancer Res; 2013 Clinical Cancer ResearchOF4
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

MAb159 suppresses PTEN deletion–induced prostatecancer progression and leukemogenesisWe tested MAb159 in the setting of constitutively active
PI3K in the PTEN knockout spontaneous tumor models,including inducible PTEN knockout in the prostate andhematopoietic system. In the PTEN knockout spontaneousprostate cancer model, PTEN deletion is achieved with theinduction of Cre under probasin promoter. In addition,
luciferase expression is induced by the same Cre; therefore,PTEN-deficient prostate cells can be imaged with a lumi-nescence imaging system (22). Tumor develops in theprostate in 2 to 3 months. Control immunoglobulin G(IgG) or MAb159 was given to 2-month-old PTEN nullmice 2 times a week at a dose of 10 mg/kg. Tumor progres-sion was monitored with live-animal luminescence imag-ing. In the MAb159-treated group, there was marked tumor
Figure 2. MAb159 inhibits xenograft tumor growth and attenuates PI3K signaling. A, the efficacy ofMAb159was examined in HT29, H249, and A549 xenograftmodels. The tumors were treated with 10mg/kg antibodies, twice a week. On the right, MAb159 enhanced the efficacy of irinotecan (18mg/kg, twice a week)in the colon cancer colo205 xenograft model. B, representative immunofluorescence staining of MAb159-treated A549 tumors. MAb159 treatment reducedcell proliferation (Ki67), induced apoptosis (TUNEL), and reduced phosphorylated S6. Nuclei were counterstained with DAPI (blue). C, similar to B,immunohistochemical staining shows that MAb159 reduced phosphorylated AKT level, but not phosphorylated MAPK (ERK1/2). Scale bar, 100 mm.All quantification data here are presented as mean � SEM. �, P < 0.05; ��, P < 0.02; ���, P < 0.001, as determined by an unpaired two-tailed Student t test.NS, not significant. D, the frozen sections of A549 tumors (four tumors from each group) were lysed, and the combined lysates were subjected to Westernblot analysis. Quantification of relative phosphorylated AKT, S6, Erk1/2, and Src was performed with Odyssey (Li-Cor).
GRP78 Antibody Inhibits PI3K and Tumor Growth
www.aacrjournals.org Clin Cancer Res; 2013 OF5
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

regression (Fig. 4A). In contrast, mice in the control IgG-treated group uniformly progressed. Histologic analysis ofdorsolateral prostate indicates that the control IgG-treatedprostate had extensive adenocarcinoma (Fig. 4B). In con-trast, the MAb159-treated prostate only had mild prostateintraepithelial neoplasia (Fig. 4B). Further immunohisto-chemical analysis shows that MAb159 significantly reducedpAKT and pS6 levels, suggesting inhibition of PI3K signal-ing (Fig. 4B). There was an insignificant decrease in phos-phorylated ERK in the MAb159-treated group (Fig. 4B). Inaddition,we studied the efficacy ofMAb159 in the xenograftmodel of a hormone refractory-mouse prostate cancer cellline CE1 that is established from a postcastration-recurrent
tumor of PTEN-deficient mouse (20). More than 50% CE1tumor growth inhibition was achieved with MAb159 treat-ment (Supplementary Fig. S9).
As reported previously, inducible knockout of PTEN inthe hematopoietic system leads to the development ofmyeloproliferative disorders and eventual leukemia, whichis suppressed by GRP78 haploinsufficiency (11, 28). ThesePTEN knockout mice when treated with control IgG hadmorbid hunched posture, whereas those treated withMAb159 appeared normal (Fig. 5A). PTEN deficiency ledto significant increase in leukemic blast cells in the bonemarrow, aswell as increase in spleenweight. PTEN-deficientmice treated with MAb159 showed a prominent reduction
Figure 3. MAb159 inhibits xenografttumor Metastasis. A, MAb159inhibited lung and liver metastasisin a 4T1 orthotopic breast cancermodel. Pictures of hematoxylin andeosin (H&E) staining of lung andliver from normal mice with noinjection of 4T1 cells are shown ontop. On day 13, no tumor was seenon the liver of MAb159-treatedmice, whereas large tumors wereobserved on the liver of controlgroup mice (left, demarcated withblack dotted line). Quantification oftumor nodules per liver is shown onthe bottom. On day 14, large areasof lung were infiltrated with tumorcells in the control group(demarcated with black dottedline), whereas only few tumor cellscould be observed in MAb159-treated lungs (right). B,representative pictures of lungmetastasis on day 14. MAb159treatment led to significantly lessmetastasis compared with thecontrol IgG group. Tumor isdemarcated by the dashed whiteline. C, MAb159 inhibits lungmetastasis of B16 melanoma cells.Mice were treated with 10 mg/kgMAb159, twice a week, right afterthe injection of B16 cellsexpressing luciferase. Live mouseimages taken 3 and 4 weeks afterB16 cell injection are shown on theleft. The representative pictures ofharvested lungs at the end of theexperiment (4 weeks) are shown onthe right. The number of pigmentedtumors on the lung surface wasmuch less in the MAb159-treatedgroup than the control group.Quantification data are presentedas mean � SEM (n ¼ 3). Signalswere quantified with ImageJsoftware. �, P < 0.05; ��, P < 0.002,as determined by an unpaired two-tailed Student t test.
Liu et al.
Clin Cancer Res; 2013 Clinical Cancer ResearchOF6
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

in spleen size and percentage of blast cells in the bonemarrow, compared with those injected with control IgG(Fig. 5B), and restoration ofwhite blood cells, lymphocytes,monocytes, and granulocytes similar to the level of wild-type mice (Fig. 5C). The effect on PI3K signaling wasmeasured by the level of phosphorylated AKT withWesternblot analysis (Fig. 5D). Consistent with suppression of AKTactivation following inducible heterozygous ablation ofGRP78 in the same PTEN null model (11), MAb159decreased phosphorylated AKT to normal levels in PTEN-deficient mice.
Affinity, activity, and specificity of humanized MAb159We selected the antibody with the desired features suit-
able for therapeutic development. Next, we preparedhumanized MAb159 to avoid potential immunogenicityin humans. This antibody has affinity to GRP78 (Fig. 6A)and efficacy in tumor growth inhibition (Fig. 6B) compa-rable with the parental murine antibody.To determine the tumor-specific localization of human-
ized MAb159, we imaged tumor-bearing live mice injectedwith Cy5.5-labeled humanized MAb159. We found that28 hours after injection, humanized MAb159 preferen-tially localized to H249 tumors but not to mouse organs(Fig. 6C, left). At the end of the study, the normal organs
together with tumors were harvested for an ex vivo imag-ing. There was a dramatic difference in the signal intensityin the tumor between control IgG and humanizedMAb159 (Fig. 6C, right). The tissues were further sub-jected to immunohistochemical analysis using a humanFc-specific antibody, confirming the specific localizationof humanized MAb159, but not control IgG, to the tumor(Fig. 6D).
Pharmacokinetics and toxicology studies ofhumanized MAb159
We performed pharmacokinetics and toxicology stud-ies of humanized MAb159 in the mouse. A single 10 mg/kg dose of humanized Mab159 was administered intra-venously, and serum levels of the antibody were mea-sured at designated time points. A mean maximum serumconcentration (Cmax) of 61.5 mg/mL was achieved. Themean serum half-life was more than 3 days (Supplemen-tary Table S1). At 10 mg/kg of humanized MAb159, anarea under curve (AUC) of 4045 � 1026 mg�h/mL wasachieved. Toxicology study was conducted in C57BL/7mice treated 2 times a week for 5 weeks with 10 mg/kghumanized MAb159. Overall, there was no significanttoxicity found in either the blood or vital organs ofhumanized MAb159-treated mice (Supplementary Table
Figure 4. MAb159 inhibits PTEN loss-induced spontaneous prostate cancer. A, live imaging pictures of prostate-specific PTEN knockout mice treated withcontrol IgG or MAb159. MAb159 significantly inhibited prostate tumor growth. Fixed imaging setting was used throughout the study. One MAb159-treatedmouse died because of anesthesia at week 4. B, H&E staining of dorsolateral prostate harvested from (A) is shown on the top. Control IgG-treatedprostate shows large area of prostatic adenocarcinoma, whereas MAb159-treated mice only show mild intraepithelial neoplasia of the prostate.Immunohistochemical staining of pS6, pAKT, and pERK are shown on the bottom and corresponding signal quantification is shown on the right. Signal wasquantified with ImageJ software. Quantification data are presented as mean � SEM (n ¼ 4). ��, P < 0.002, as determined by an unpaired two-tailed Studentt test. Scale bar, 100 mm.
GRP78 Antibody Inhibits PI3K and Tumor Growth
www.aacrjournals.org Clin Cancer Res; 2013 OF7
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

S2). No histologic abnormalities were seen except formild inflammation in the pancreas of a control andhumanized MAb159-treated mouse. These results providegood safety and pharmacokinetics data to proceed withclinical trials.
DiscussionCell surface GRP78 is a multifunctional receptor and a
potential target for cancer therapy. For example, proapop-totic moieties or cytotoxic agents were conjugated ontopeptides with high affinity for GRP78 to successfully targetand kill cancer cells (12, 19). Recently, an unconjugatedpeptidic GRP78 ligand also demonstrated toxicity to pros-tate cancer cell (29). A human monoclonal IgM antibodyagainst cell surface GRP78 isolated from a cancer patient iscapable of inducing lipid accumulation and apoptosis incancer cells (30). Here, we screened and identified a novelmAb that recognizes both human and mouse GRP78 withhigh specificity and potently inhibits tumor growth andcauses tumor regression in xenograft tumor models andspontaneous tumor models. In addition, MAb159 sup-presses tumor metastasis in multiple models. Furthermore,the humanized form of this antibody is efficacious andwithout toxicity.
Cell surface GRP78 is an upstream regulator of PI3K/AKTsignaling. GRP78 interacts with Cripto and a2-macroglob-ulin on the cell surface and is required for these factors toactivate downstream PI3K/AKT signaling (16, 31, 32).
Recently, a murine mAb against GRP78 was reported tosuppress AKT activation in a melanoma model (33). Here,we show that surface GRP78 is in the same complex as thep85 subunit of PI3K, and treatment of cells with theGRP78-specific antibody MAb159 led to marked reduction in PI3Ksignaling in both cultured cells andmultiple tumormodels.The spontaneous tumor models include PTEN deletion–induced spontaneous prostate cancer and leukemiamodels.Furthermore, in tumor xenografts subjected to long-termtreatment with MAb159 for up to 30 days, inhibition of thePI3K pathway was sustained, and there was no evidence forinduction of escape mechanisms of resistance to PI3Kinhibition such as MAPK (34). Altogether, these highlightthe importance of surfaceGRP78 inPI3K/AKT signaling andsupport the clinical investigation of MAb159 for PI3K-driven tumors. However, cell surface GRP78 may alsoregulate other critical oncogenic pathways. For example,MAb159 is active to cell lines with Kras mutation (A549)and Braf mutation (HT29 and Colo205). In the light ofproapoptotic activity of MAb159, we are also investigatingthe role of surface GRP78 in the extrinsic apoptoticpathway.
Recent studies reported that surface GRP78 is highlyelevated in tumor-associated vasculature and is requiredfor endothelial cell proliferation and survival (7, 9). Thesefindings indicate that cell surface GRP78-targeting agentswill have dual function: targeting tumor cell and tumorvasculature. Consistently, we have observed that MAb159
Figure 5. MAb159 suppresses PTEN loss-induced spontaneous leukemia. A, hunched posture of PTENf/f;MX1-Cre mice treated with IgG compared withnormal posture of those treated with MAb159. B, quantification of the spleen weight and leukemic blast cell percentages of wild-type (WT; n ¼ 4), cPf/f
treated with IgG (n ¼ 5), and cPf/f treated with MAb159 (n ¼ 8). All data are presented as mean � SE C, complete blood count with tail peripheralblood from WT (n ¼ 4), cPf/f treated with IgG (n ¼ 5), and cPf/f treated with MAb159 (n ¼ 8). All data are presented as mean � SE �, P < 0.05; ��, P < 0.01,as determined by an unpaired two-tailed Student t test. D, results of representative Western blot analysis using bone marrow cell lysates for detectionof the indicated protein levels. For each group, samples from two mice were used for analysis.
Liu et al.
Clin Cancer Res; 2013 Clinical Cancer ResearchOF8
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

reduced the density of tumor endothelial cell and angio-genesis-dependent tumor metastasis, supporting dualtargeting.Antibody tracking and in vivo imaging studies showed
that MAb159 is strictly localized to the tumor and notnormal organs, indicating that antibody targeting is highlyspecific. In agreement with this, formal toxicology study inmice showed that the humanized antibody was well toler-ated and did not induce any noticeable organ toxicity orchanges in blood counts or blood chemistry.Another potential application of MAb159 lies in its
ability to be used for in vivo imaging. MAb159 specifi-cally recognizes surface GRP78 and, thus, can be usedto image the tumor for personalized medicine and deter-mine whether the amount of surface GRP78 in the tumorpredicts disease progression and response to therapy.Clinical trials will incorporate patient imaging as ascreening process for inclusion of study subjects. This isparticularly important in cell surface–GRP78-targetedtherapy: when analyzing archival tumor samples withimmunostaining, intracellular GRP78 will interfere withsuch analysis.In conclusion, we have developed a novel antibody
MAb159 targeting surface GRP78 expressed on tumor cells
and tumor endothelial cells. This antibody disrupts thePI3K signaling pathway and induces apoptosis in tumorcells, while sparing normal cells. It has the potential to beboth a therapeutic and diagnostic agent in cancer.
Disclosure of Potential Conflicts of InterestNo potential conflicts of interest were disclosed.
Authors' ContributionsConception and design: R. Liu, S.K. Mitra, A.S. Lee, P. GillDevelopmentofmethodology:W.Gao, Y. Zhou, V. Krasnoperov, D.Dong,D. LiAcquisitionofdata (provided animals, acquired andmanagedpatients,provided facilities, etc.): R. Liu, W. Gao, Y. Zhou, S. Wey, S. Liu, G. Zhu, S.Louie, P.S. Conti, Z. Li, A.S. Lee, P. GillAnalysis and interpretation of data (e.g., statistical analysis, biosta-tistics, computational analysis):R. Liu, X. Li,W. Gao, Y. Zhou, S.Wey, S.K.Mitra, D. Dong, S. Louie, P.S. Conti, Z. Li, A.S. Lee, P. GillWriting, review, and/or revision of the manuscript: R. Liu, V. Krasno-perov, P.S. Conti, Z. Li, A.S. Lee, P. GillAdministrative, technical, or material support (i.e., reporting or orga-nizing data, constructing databases): R. Liu, X. Li, W. Gao, Y. Zhou, S.Wey, V. Krasnoperov, D. Dong, P. GillStudy supervision: S. Louie, P.S. Conti, Z. Li, A.S. Lee
AcknowledgmentsThe authors thank Drs. Rui Yan, Zhi Sun, Ram Subramanyan, and Darryl
Shibata for their technical assistance and scientific discussions with regard tothis study.
Figure 6. Characterization of humanized MAb159. A, comparison of the affinity and antitumor activity of MAb159 and humanized MAb159 using competitionELISA. Various amounts of murine and humanized MAb159 were tested for competition against a fixed concentration of biotinylated murine MAb159 forbinding to GRP78-His. The binding of biotinylatedMAb159 decreaseswith increasing amounts of bothmurine and humanizedMAb159. HumanizedMAb159shows slightly higher affinity for GRP78 than murine MAb159. B, A549 xenograft were treated with normal mouse IgG, murine MAb159, and humanizedMAb159 (n ¼ 8 each). Antibodies were administered at 10 mg/kg, 2 times a week. Data are presented as mean � SEM. C, tumor-bearing mice wereadministeredCy5.5-conjugated humanizedMAb159 or normal human IgG, and fluorescentwhole-mouse imageswere taken 28hours after injection (left). Themice were subsequently perfused with PBS and formalin, and the ex vivo images of the major organs were taken (right). MAb159, but not control IgG, showsspecific localization to tumor. Quantification of the fluorescent intensity is shown on the bottom. D, the organs in (C) were immunostained with a human Fc-specific antibody, showing the preferential localization of humanizedMAb159 to the tumor. Normal IgG localized to both the liver and the tumor. Scale bar, 100mm.Quantification data are presented asmean�SEM (n¼ 4). Signalswere quantifiedwith ImageJ software. ��,P < 0.005, as determined by an unpaired two-tailed Student t test.
GRP78 Antibody Inhibits PI3K and Tumor Growth
www.aacrjournals.org Clin Cancer Res; 2013 OF9
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

Grant SupportThis work was financially supported by the L.K. Whittier Foundation
(grant to A.S. Lee and P. Gill) and the National Cancer Institute of theNational Institute ofHealth (grant no. R01CA027607 to A.S. Lee andP.Gill).
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby marked
advertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received April 30, 2013; revised August 28, 2013; accepted September 4,2013; published OnlineFirst September 18, 2013.
References1. Ma Y, Hendershot LM. The role of the unfolded protein response in
tumour development: friend or foe? Nat Rev Cancer 2004;4:966–77.2. Luo B, Lee AS. The critical roles of endoplasmic reticulum chaperones
and unfolded protein response in tumorigenesis and anticancer ther-apies. Oncogene 2013;32:805–18.
3. Li J, Lee AS. Stress induction of GRP78/BiP and its role in cancer. CurrMol Med 2006;6:45–54.
4. Lee AS. GRP78 induction in cancer: therapeutic and prognosticimplications. Cancer Res 2007;67:3496–9.
5. Li Z, Li Z. Glucose regulated protein 78: a critical link between tumormicroenvironment and cancer hallmarks. Biochim Biophys Acta 2012;1826:13–22.
6. Pyrko P, Schonthal AH, Hofman FM, Chen TC, Lee AS. The unfoldedprotein response regulator GRP78/BiP as a novel target for increasingchemosensitivity in malignant gliomas. Cancer Res 2007;67:9809–16.
7. Virrey JJ, Dong D, Stiles C, Patterson JB, Pen L, Ni M, et al. Stresschaperone GRP78/BiP confers chemoresistance to tumor-associatedendothelial cells. Mol Cancer Res 2008;6:1268–75.
8. Dong D, Ni M, Li J, Xiong S, Ye W, Virrey JJ, et al. Critical role of thestress chaperone GRP78/BiP in tumor proliferation, survival, andtumor angiogenesis in transgene-induced mammary tumor develop-ment. Cancer Res 2008;68:498–505.
9. DongD,StapletonC, LuoB, XiongS,YeW,ZhangY, et al. A critical rolefor GRP78/BiP in the tumor microenvironment for neovascularizationduring tumor growth and metastasis. Cancer Res 2011;71:2848–57.
10. Fu Y, Wey S, Wang M, Ye R, Liao CP, Roy-Burman P, et al. Pten nullprostate tumorigenesis and AKT activation are blocked by targetedknockout of ER chaperone GRP78/BiP in prostate epithelium. ProcNatl Acad Sci U S A 2008;105:19444–9.
11. Wey S, Luo B, Tseng CC, Ni M, Zhou H, Fu Y, et al. Inducible knockoutof GRP78/BiP in the hematopoietic system suppresses pten-nullleukemogenesis and AKT oncogenic signaling. Blood 2012;119:817–25.
12. Arap MA, Lahdenranta J, Mintz PJ, Hajitou A, Sarkis AS, Arap W, et al.Cell surface expression of the stress response chaperone GRP78enables tumor targeting by circulating ligands. Cancer Cell 2004;6:275–84.
13. Gonzalez-Gronow M, Selim MA, Papalas J, Pizzo SV. GRP78: amultifunctional receptor on the cell surface. Antioxid Redox Signal2009;11:2299–306.
14. Ni M, Zhang Y, Lee AS. Beyond the endoplasmic reticulum: atypicalGRP78 in cell viability, signalling and therapeutic targeting. Biochem J2011;434:181–8.
15. Zhang Y, Liu R, Ni M, Gill P, Lee AS. Cell surface relocalization of theendoplasmic reticulum chaperone and unfolded protein responseregulator GRP78/BiP. J Biol Chem 2010;285:15065–75.
16. Shani G, Fischer WH, Justice NJ, Kelber JA, Vale W, Gray PC. GRP78and cripto form a complex at the cell surface and collaborate to inhibittransforming growth factor beta signaling and enhance cell growth.Mol Cell Biol 2008;28:666–77.
17. Liu Y, Steiniger SC, Kim Y, Kaufmann GF, Felding-Habermann B,Janda KD. Mechanistic studies of a peptidic GRP78 ligand for cancercell-specific drug delivery. Mol Pharm 2007;4:435–47.
18. SatoM, Yao VJ, ArapW, Pasqualini R. GRP78 signaling hub a receptorfor targeted tumor therapy. Adv Genet 2010;69:97–114.
19. Miao YR, Eckhardt BL, Cao Y, Pasqualini R, Argani P, Arap W, et al.Inhibition of establishedmicrometastases by targeted drugdelivery viacell surface-associated GRP78. Clin Cancer Res 2013;19:2107–16.
20. Liao CP, Liang M, Cohen MB, Flesken-Nikitin A, Jeong JH, Nikitin AY,et al. Mouse prostate cancer cell lines established from primary andpostcastration recurrent tumors. Horm Cancer 2010;1:44–54.
21. Krasnoperov V, Kumar SR, Ley E, Li X, Scehnet J, Liu R, et al. NovelEphB4 monoclonal antibodies modulate angiogenesis and inhibittumor growth. Am J Pathol 2010;176:2029–38.
22. Liao CP, Zhong C, Saribekyan G, Bading J, Park R, Conti PS, et al.Mouse models of prostate adenocarcinoma with the capacity tomonitor spontaneous carcinogenesis by bioluminescence or fluores-cence. Cancer Res 2007;67:7525–33.
23. Vercauteren D, Vandenbroucke RE, Jones AT, Rejman J, Demeester J,De Smedt SC, et al. The use of inhibitors to study endocytic pathwaysof gene carriers: optimization and pitfalls. Mol Ther 2010;18:561–9.
24. Shin SY, Lee KS, Choi YK, Lim HJ, Lee HG, Lim Y, et al. Theantipsychotic agent chlorpromazine induces autophagic cell deathby inhibiting the Akt/mTOR pathway in human U-87MG glioma cells.Carcinogenesis 2013;34:2080–9.
25. Takahashi H, Wang JP, Zheng HC, Masuda S, Takano Y. Overexpres-sion of GRP78 and GRP94 is involved in colorectal carcinogenesis.Histol Histopathol 2011;26:663–71.
26. Aslakson CJ, Miller FR. Selective events in the metastatic processdefined by analysis of the sequential dissemination of subpopulationsof a mouse mammary tumor. Cancer Res 1992;52:1399–405.
27. Bao L, Haque A, Jackson K, Hazari S, Moroz K, Jetly R, et al. Increasedexpression of P-glycoprotein is associatedwith doxorubicin chemore-sistance in the metastatic 4T1 breast cancer model. Am J Pathol2011;178:838–52.
28. Yilmaz OH, Valdez R, Theisen BK, Guo W, Ferguson DO, Wu H, et al.Pten dependence distinguishes haematopoietic stem cells from leu-kaemia-initiating cells. Nature 2006;441:475–82.
29. MaddaloD,NeebA, JehleK,SchmitzK,Muhle-Goll C, Shatkina L, et al.A peptidic unconjugated GRP78/BiP ligand modulates the unfoldedprotein response and induces prostate cancer cell death. PLoS One2012;7:e45690.
30. Rauschert N, Brandlein S, Holzinger E, Hensel F,Muller-Hermelink HK,Vollmers HP. A new tumor-specific variant of GRP78 as target forantibody-based therapy. Lab Invest 2008;88:375–86.
31. Misra UK, Gonzalez-GronowM, Gawdi G, Hart JP, Johnson CE, PizzoSV. The role of grp 78 in alpha 2-macroglobulin-induced signal trans-duction. evidence from RNA interference that the low density lipopro-tein receptor-related protein is associated with, but not necessaryfor, GRP 78-mediated signal transduction. J Biol Chem 2002;277:42082–7.
32. Misra UK, Pizzo SV. Receptor-recognized alpha(2)-macroglobulinbinds to cell surface-associated GRP78 and activates mTORC1 andmTORC2 signaling in prostate cancer cells. PLoSOne 2012;7:e51735.
33. de Ridder GG, Ray R, Pizzo SV. A murine monoclonal antibodydirected against the carboxyl-terminal domain of GRP78 suppressesmelanoma growth in mice. Melanoma Res 2012;22:225–35.
34. Camp ER, Summy J, Bauer TW, Liu W, Gallick GE, Ellis LM. Molecularmechanisms of resistance to therapies targeting the epidermal growthfactor receptor. Clin Cancer Res 2005;11:397–405.
Liu et al.
Clin Cancer Res; 2013 Clinical Cancer ResearchOF10
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106

Published OnlineFirst September 18, 2013.Clin Cancer Res Ren Liu, Xiuqing Li, Wenming Gao, et al. and MetastasisAgent in Suppressing PI3K/AKT Signaling, Tumor Growth, Monoclonal Antibody against Cell Surface GRP78 as a Novel
Updated version
10.1158/1078-0432.CCR-13-1106doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2013/09/18/1078-0432.CCR-13-1106.DC1Access the most recent supplemental material at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://clincancerres.aacrjournals.org/content/early/2013/11/15/1078-0432.CCR-13-1106To request permission to re-use all or part of this article, use this link
Research. on March 28, 2020. © 2013 American Association for Cancerclincancerres.aacrjournals.org Downloaded from
Published OnlineFirst September 18, 2013; DOI: 10.1158/1078-0432.CCR-13-1106