Embed Size (px)
Citation preview

1040-2519/97/0601-0137$08.00
137
MARKSTRICHOMEDEVELOPMENTIN ARABIDOPSISAnnu.Rev. Plant Physiol. Plant Mol. Biol. 1997. 48:137–63Copyright© 1997by AnnualReviewsInc. All rightsreserved
MOLECULARGENETICANALYSISOF TRICHOME DEVELOPMENT INARABIDOPSIS
M. David MarksDepartment of GeneticsandCell BiologyandDepartmentof PlantBiology,Universityof Minnesota, St.Paul, Minnesota55108
KEY WORDS: cell differentiation, cell fate,lateralinhibition, transcription factor
ABSTRACT
Two basic questionsin developmental biologyare: How doesacell knowwhenit should orshouldnotdifferentiate,andonceacell iscommittedtodifferentiate,how is that process controlled?Thefirst processregulates the arrangement orpattern of thevarious cell types,whereas the second makescells functionallydistinct. Together, thesetwo processes define plant morphogenesis. Trichomedevelopment in Arabidopsis provides an excellent model to analyze thesequestions. First, trichome development in Arabidopsis is a relatively simpleprocess. A single epidermal cell differentiates into a unicellular trichome.Second, thisdifferentiation occursin anonrandom pattern ontheplant surface.Finally, theprocessisamenable togenetic analysisbecausemany mutationsthataffect trichomedifferentiation do not alter other aspects of plant development.Thus far, more than 20 genes affecting trichome development have beenidentified. Thisreviewexaminesthecurrent stateof ourunderstanding of thesegenes.
CONTENTSINTRODUCTION..................................................................................................................... 138TRICHOMEDEVELOPMENTIN ARABIDOPSIS............................................................... 138TRICHOMEMUTANTS.......................................................................................................... 142
Mutations AffectingEarly TrichomeDevelopment............................................................. 143ReducedBranchingMutants ............................................................................................... 146TrichomeExpansion............................................................................................................ 149Maturation Mutants............................................................................................................. 149

Genetic Interactions ............................................................................................................ 151MOLECULAR STUDIES......................................................................................................... 151
GL1...................................................................................................................................... 151TTG...................................................................................................................................... 153Interactions BetweenGL1and TTG.................................................................................... 155GL2...................................................................................................................................... 156ZWI...................................................................................................................................... 157Other Genes......................................................................................................................... 158
CONCLUSIONSAND FUTURE DIRECTIONS.................................................................... 159
INTRODUCTION
Most highereukaryoticorganismsbeginlife asa single-celledzygote.Duringorganismaldevelopmentthis cell divides,andtheresultingprogenydifferenti-ateandacquirespecialfunctions.In manyplants,thefirst division of a zygoteresultsin two cellswith different fates.Onecell will form thesuspensor,andthe othercell will producethe embryoproper.Much later, divisions of pro-todermalcells in leaf primordia generatedaughtercells that differentiateintovastlydifferentcell types.Thus,from thefirst to theterminalcell divisionsofplantdevelopment,controlof cell fate isimportant.
The developmentof plant leaf hairs,trichomes,providesan excellentsys-tem to study the control of cell fate (24, 49, 61–63, 76). First, trichomesdevelopon theepidermalsurface,andall phasesof trichomedevelopmentcanbeobserved.Second,developmentof trichomesis a relativelysimpleprocess.A single epidermalcell differentiatesinto a single-celledtrichome.Third, thedevelopmentof trichomescanbe geneticallydissectedbecausenormalplantgrowth and developmentdo not require the presenceof trichomes.Finally,understandingtrichomedevelopmentmayhavepracticalimplicationsbecausethere is a correlationbetweenthe presenceof trichomes and resistancetoherbivoryby certain insect pests (1,27, 37, 39, 44–46, 53, 68). Thegenetics oftrichomeformationhasbeenstudiedin otherplant species(4, 19, 28, 40, 41,52, 81),but this review focuses onArabidopsis.
TRICHOMEDEVELOPMENT IN ARABIDOPSIS
Trichomesarenormallypresenton theleaves,stems,andsepalsof Arabidop-sis (Figure 1A, B, C). They are normally absentfrom the roots, hypocotyl,cotyledons,petals,stamens,andcarpels.Themorphology of trichomesvariesfrom unbranchedspikes,which aremostcommonly found on the stemsandsepals,to structurescontainingtwo to five branches,which arefound on theleaves.Most trichomemutationsaffect all of the trichomeson a plant. This
138 MARKS

Fig
ure
1T
rich
omes
onA
rabi
dops
isse
edlin
gs.(
A)
Lea
ftr
icho
mes
.(B
)S
tem
tric
hom
es.(
C)
Sepa
ltri
chom
es.A
rrow
sde
note
tric
hom
es.
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 139

suggeststhatwhile differenttrichomesmayhavedifferentmorphologies,theirdevelopmentis controlledby the samegenes.
Trichomedevelopmentproceedsasa waveacrosstheepidermalsurfaceontheleaf (31,60). The firsttrichomeinitiates onthe tipof theadaxial surface ofthe first primordiumafter it achievesa lengthof approximately 100 µm (47).As trichomesmature at theleaf tip, new trichomes emerge progressivelytoward the base(Figure 2A). In addition,new trichomesinitiate in betweendevelopingtrichomesthat havebeenseparatedfrom oneanotherby dividing
Figure2 Trichomedevelopment onArabidopsisleaves.(A) Scanningelectronmicrograph(SEM)of ayoung leaf with mature and developing trichomes.Thick arrow denotesdeveloping trichome;thin arrow denotes mature trichome. (B) Section through emerging trichome. Arrow denotesenlargednucleus. (C) SEM of developing trichomes.Arrows denote expanding branches.Whitebarsindicateapproximately 12µm.
140 MARKS

epidermalcells.Trichomeinitiation is foundonly in regionswhereepidermalcell division isoccurring.
The first detectablestepin the commitment to the trichomecell fate is acessationof cell division; however,nuclearDNA synthesis continues,andthecommittedcell undergoesat leasttwo roundsof endoreduplication,reachingatleast8N (Figure2B) (31, 69). Cellssurroundinga committedcell continuetodividenormally. After thecommittedcell radiallyexpandsto adiameterthatisapproximatelytwofold greaterthanthesurrounding cells, it beginsto expandpreferentiallyon its outersurfaceto form a stalk (Figure2C) (60, 64). As thestalk forms, diffuse growth throughoutthe cell results in continued radialexpansion.The nucleusmigratesinto the aerial portion of the stalk shortlybefore secondary protuberances(31), which subsequently expand intothebranches,emergefrom theaerialtip (Figure2C). Duringbranchformation, thenucleusundergoesanotherround of endoreduplication and migratesto thebaseof the last branchthat forms (31). Expansionceaseswhen a trichomereachesa heightof 200–300µm anda basediameterof approximately50 µm.During trichomematurationthecell wall thickensto approximately5 µm, andthetrichomesurfacebecomescoveredwith papillae.In addition,theepidermalcells aroundthe baseof a trichomeacquirea distinct rectangularshape.Itappearsasthoughthe trichomebaseoften pushesunderthe surroundingepi-dermal cells to createa socket.Thus, the surrounding cells are sometimesreferredto as socket cells(31).
Detailedcellular analysisof Arabidopsistrichomedevelopmenthasyet tobe completed.Thus, little is known about the roleof the endomembranesystemor cytoskeleton in trichomemorphogenesis.
Trichome development hasbeenusedasamarkerfor leafheteroblasty(85).Differencesin trichomeshapeandposition canbeobservedin acomparisonoftheinitial andlaterleaves.Thefirst two to three leaves have adaxialtrichomesbut lack trichomeson their abaxial surfaces.Later rosetteleaveshave anincreasingnumberof abaxial trichomes. This progressioncontinueson thebract-likeleaveson thestem.The first bractshavetrichomeson both adaxialand abaxial surfaces; however, later bracts have diminished numbers ofadaxial trichomes, whilemaintaining theirabaxial trichomes.
Trichomesare evenly distributedacrossthe leaf surfaces,but contiguoustrichomesarerarelyobserved.This typeof arrangementhasbeendescribedasan isotropicpattern(25). A statistical analysisof trichomespacinghasshownthatit is nonrandom(47), that is, it is statistically significant thatno trichomesare contiguous. The parameterR was set as a ratio betweenthe measuredaveragedistancesbetweennearestneighbortrichomeson the leaf surfaceandtheaveragenearestneighbordistanceexpectedfor a randompatternwith the
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 141

samedensity(5). A randompatternwould havea valueof R = 1, whereasamaximum spacingarrangement(i.e. all trichomesequal distancefrom oneanother)would result in R = 2.15.A valueof R = 1.40,which representsasignificant deviation fromarandomdistribution,wasobserved[P < 0.01(47)].This indicatesthatthere isa minimum distancebetween trichomes.
To studythedevelopmentof thetrichome pattern,the frequencyandspac-ing of initiatingtrichomes werestatistically analyzed(47). In asampleof 2120epidermalcellson youngleaf primordia, it wasfoundthatthefractionof cellsthat commit to the trichomepathwaywas0.041.With this populationsize,iftrichomeinitiationwasstochastic,then16 neighboring trichomesshouldhavebeenobserved. Becausenone was observed,the probability that trichomeinitiation was a randomeventin this trichomesamplewas lessthan 10−8.
Becauseplant cells do not migrate,thereare two main ways in which anonrandompatterncan be generated.First, it is possible a trichomeand thecells thatsurroundit arederivedfrom thesamecell lineage,andonly onecellin thegroupbecomesatrichome;afterthiscell differentiatescontinuedepider-mal cell divisions would always separatedevelopingtrichomes.A secondpatterningmechanismrequirescell-to-cell communication. A sectorialanaly-sis was usedto analyzethe mechanismcontrolling trichome spacing(47).Plants containing a GUS reporter genethat had beeninactivatedby amaizeActransposonwereused.TheGUScodingsequencewasunderthecontrolof theCaMV 35SRNA promoter;Ac transposition during early plant developmentwould result in a largeclonal sectorof GUS positive cells (51). To test thehypothesisthat trichomespacingis controlledby cell lineage,trichomesthatdevelopedalong the borderof GUS positive sectorswereanalyzed.No evi-dence for a cell lineageassociatedwith trichomeswas found. Thus, apparentlytrichomespacingis controlledby a mechanisminvolving cell-to-cellcommu-nication.
TRICHOME MUTANTS
Mutationsaffectingtrichomeinitiation,spacing,density, andshapehavebeenrecovered.Someof the mutationsaffect nontrichome developmental proc-esses.Trichomemutants were first usedasconvenientgeneticmarkers.Theglabrous1(gl1) mutant,which lackstrichomeson mostsurfaces,wasusedinearly genemapping studies(67). In 1978 distorted1 (dis1) and distorted2(dis2) mutants, which havedefectsin trichomecell expansion,wereusedtomap genesto chromosome1 (13). In 1982 trichomemutants were usedtocalculatemutation frequenciesgeneratedusing severaldifferent mutagens(43). In 1988,a review by Haughn& Somerville (24) first documentedthe
142 MARKS

possibleuseof trichomemutantsasa modelto addressquestionsconcerningcell fate and differentiation. In 1994 Hülskampet al describedmany newtrichomemutantsthat were recoveredfrom a saturationscreen(31). Severalother recentreportsalso describethe characterizationof new trichome mu-tants.
MutationsAffecting Early Trichome Development
The recessivegl1 and transparent testaglabra (ttg) mutationshavethe mostdramaticaffecton trichomeformation(42, 43).Strongloss-of-functionmuta-tions in eithergeneresultsin a completelossof trichomeformationon mostaerialsurfaces(Figure3A). The gl1 mutation only appearsto affect trichomedevelopment;however, the ttg mutation has severaldevelopmental conse-quences.ttg plantslack anthocyaninpigments, which causesttg seedlingstolack redpigmentsandseedsto beyellow insteadof reddishbrown.ttg mutantseedsalso lack thepolysaccharidemucilagethataccumulatesin theouterlayerof thetesta.Asidefrom thelack of mucilageandnormalpigmentation,the ttgseedcoatdevelopsnormally. Finally, ttg mutants produceectopicroot hairs(18). The root epidermisof Arabidopsisnormally containstwo typesof cellfiles. In onefile all cellsareroot hairs,andthecellsareslightly lesselongatedthanthecells in theotherfile type,which containsonly nonhaircells.In a ttgroot, the cellfiles thatnormallyproduce roothairs are unaltered.However, thecells in files thatarenormallyhairlessassumethelesselongatedshapeof haircells andmost,butnotall, of the cellsin these filesform hairs(18).
The lossof trichome initiation is not complete in eithermutant.Both gl1andttg mutants havea few trichomeson themarginof therosetteandcaulineleaves(Figure3B). In addition, ttg plantsoften havetrichomesnearthe leafmarginof theadaxialsurface(Figure3C). Apparentlythemarginaltrichomesarecontrolledby genesotherthanGL1 or TTG (74).Mutationsthatresultin alossof the marginaltrichomeshavenot beendescribed.Othermutations thataffect trichome morphology also affect the morphology of the margin tri-chomes(MD Marks, unpublisheddata).Therefore,many genesasidefromGL1andTTGareactivein bothmarginandnonmargintrichomes.
Weakallelesof bothgl1 andttg havebeenidentified.gl1-2 plantsexhibit apartial lossof trichomes,with a markedreductionof trichomesin themidveinregion(12). Trichomedifferentiationis alteredbecausemanyof the leaf tri-chomesonly form rudimentaryspikes.In addition,the trichomespacingpat-tern isaltered inthatside-by-siderudimentarytrichomesare notuncommon.
Severalweakallelesof ttg havebeenidentified (48). ttg-10 plantsexhibitclustersof normalandrudimentary trichomesalongthe leaf margin.Interest-ingly, the ttg-10mutation doesnot affectall thedevelopmental processesthat
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 143
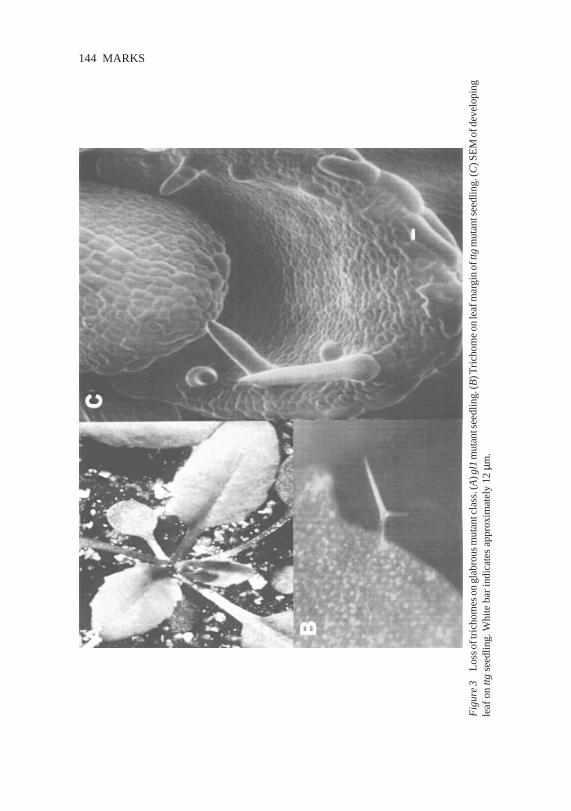
Fig
ure
3L
osso
ftri
chom
eson
glab
rous
mut
antc
lass
.(A
)gl1
mut
ants
eedl
ing.
(B)T
rich
ome
onle
afm
argi
nof
ttgm
utan
tsee
dlin
g.(C
)SE
Mof
deve
lopi
ngle
afon
ttgse
edlin
g.W
hite
bar
indi
cate
sap
prox
imat
ely
12µm
.
144 MARKS

normally arealteredin ttg mutants.ttg-10 plantslack testapigmentation, butthey appearto exhibit normalseedlingpigmentation, andtheir seedsdevelopmucilage.
Mosaicanalysishasbeenusedto determinethat the GL1 geneappearstoact cell autonomously. Rédei exposedseedsfrom a GL1/gl1 heterozygousplantto X rays(80).He subsequently identifiedglabroussectorson theresult-ing plants.More recently,heterozygousseedsweretreatedwith EMS,andtheresulting plants producedglabroussectorsthat were not seenon wild-typeplants(31). Theglabrouspatchesarethoughtto resultfrom GL1 markerloss,uncovering thegl1 mutant allelein heterozygousplants. Thepresenceofsectorsindicatesthat cellsoutside thesectorcannotprovidea diffusablesub-stance or signalto overcomethe effect of thegl1 mutation.
It was noted that the leavesof Landsbergerecta (Ler) plants containedfewertrichomesthantheleavesof Columbia (Col) plants[Ler ∼10 vs Col ∼30on the first leaf (63)]. Using the Lister & Deanrecombinantinbredmappinglines (55), which weregeneratedwith Ler andCol parents,it waspossibletousequantitative trait analysisto maptheloci responsible for thedifferencesintrichomedensity(47). Onemajor locusaccountsfor 73%of thevarianceandmapsin the regionbetweenthe markerserectaandm220on chromosome2(LOD score:27.54). This locus has beennamedREDUCED TRICHOMENUMBER(RTN). The RTNCol allele exhibits incompletedominanceover theRTNLer allele,becauseF1 hybridshavean intermediatenumberof trichomeson first leaves.
Apparently RTN controls thepersistenceof trichome initiation (47). Acomparisonof trichomeinitiation on plantscarryingtheCol or theLer allelesrevealedthattrichomeinitiationbeginsonthefirst leafprimordiaaftera lengthof ∼100µm is reached.As theleafprimordiaincreasedfrom 100µm in lengthto 300 µm, equalnumbersof trichomeswere initiated in eachbackground.However, Ler leaf primordia longer than 300 µm rarely initiated new tri-chomes,whereasCol leaf primordia over 800 µm long continuedto initiatenew trichomes.
There areseveralmodes bywhich RTN could influence thetiming oftrichomeinitiation (47). It is possible that RTN increasesthe areaof lateralinhibition aroundinitiating trichomes.This seemsunlikely becausethe dis-tancebetweenthe early initiating trichomeson both Col and Ler leavesareroughly the same.More likely is the possibility that RTN is involved incontrollingeitherthe lengthof time during which epidermalcellsarecompe-tent to respondto a trichome-inductivesignalor the lengthof time they areableto producetheinductive signal.For example,if TTGand/orGL1 expres-sion resultsin an inductive signal for trichomeinitiation, thenRTNcould be
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 145

involved in the transductionof thesignal orin positively or negatively regulat-ing the timing of TTG and/orGL1 expression.The incompletedominanceoftheCol alleleover the Ler allelesuggeststhat the level of RTNexpressionisimportant. It also is possiblethat severalclosely linked genesinfluencingtrichomedensityare locatedin theRTNregion.
Recessivemutationsin theGLABRA2(GL2) genedisruptnormaltrichomemorphogenesis(43). Two classesof trichomesdevelopon gl2 mutants. Oneclassof trichomeshasa rudimentaryspikethat projectsupward(Figure4A).Toward the marginof gl2 leaves,trichomesaremorenormal in appearance;however, thesetrichomesare less branchedthan normal. Mosaic analysessuggestthat GL2 actscell autonomously (31). Mutations in GL2 also affectotherdevelopmentalprocesses.Like ttg, gl2 mutantslack seedcoatmucilageandproduce ectopic hairsin the root (43,66). However,thereis amorphologi-cal differencein the root hair phenotypebetweenthe two mutants. Normally,hair-bearingcells are shorterthan nonhaircells. In ttg mutants,all the rootepidermalcells assumethe morphologyof the hair cells and many producehairs. In contrast,while all the epidermalcells in a gl2 mutantcan producehairs,the files of cells thatnormallywould not becomehairsmaintaina moreexpanded shape.
Recessivemutations in the GLABRA3(GL3) genehave two effects ontrichomedevelopment(43). On early leavesthereis a decreasein trichomeinitiation(Figure3B). As in gl2 mutants andweakgl1 andttg mutants,thelossof trichome initiation is most striking in the midvein region of the leaves.However,on later leavestrichomeinitiation is moreuniform. The trichomesthatdo developtendto belessbranchedandaremoreslenderthannormal.Inaddition, it hasbeenobservedthat trichomesin gl3 mutantsundergofewerroundsof endoreduplicationthan wild-type trichomes(31).
Mutations in either the TRYPTYCHON(TRY) or KAKTUS (KAK) genesresultin larger thanwild-type trichomeswith increasedbranchformation(31).Trichomeson thesemutantsalsoexhibit an increasein endoreduplication.Ithas beenreportedthat try mutants have a greaternumberof clusteredtri-chomesthan are found on wild-type plants (31). Thus, TRY also may beinvolvedin controlling the proposedlateralinhibition pathway.
ReducedBranchingMutants
Severalrecessivemutations appearto reducetrichomebranchingwithout af-fecting trichomeinitiation. Hülskampet al divided branchinginto a primaryphasethat generatesthe first branchand a secondaryphasethat generatessubsequentbranches(31). The stichel (sti) mutationeliminatesboth of these
146 MARKS
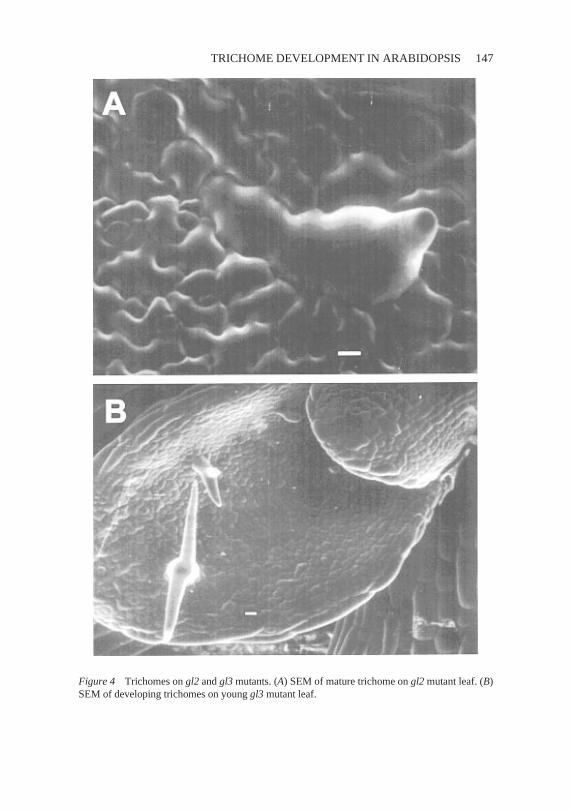
Figure 4 Trichomesongl2 andgl3 mutants. (A) SEMof mature trichomeongl2 mutantleaf.(B)SEMof developing trichomesonyoung gl3 mutantleaf.
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 147
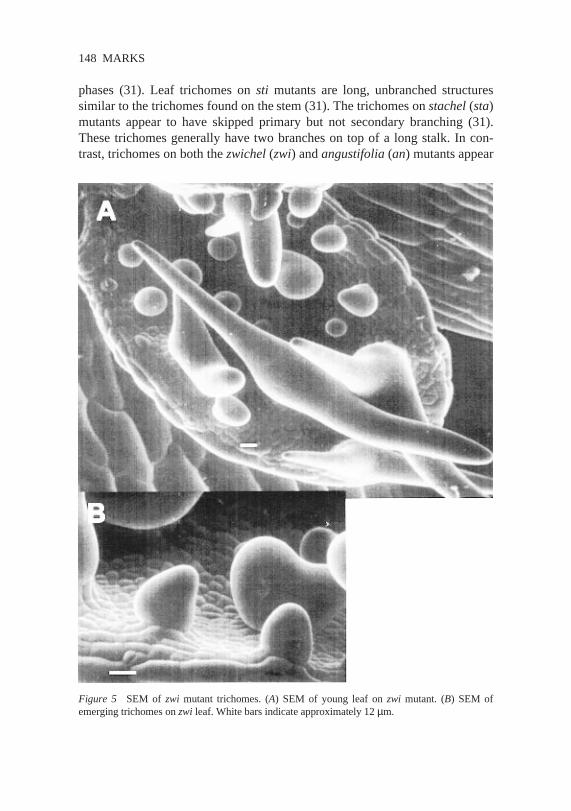
phases(31). Leaf trichomes on sti mutants are long, unbranchedstructuressimilar to the trichomes found on thestem(31).The trichomes onstachel(sta)mutantsappearto haveskippedprimary but not secondarybranching(31).Thesetrichomesgenerallyhavetwo brancheson top of a long stalk. In con-trast,trichomeson boththezwichel(zwi) andangustifolia (an) mutantsappear
Figure 5 SEM of zwi mutant trichomes.(A) SEM of young leaf on zwi mutant. (B) SEM ofemerging trichomes onzwi leaf.Whitebarsindicateapproximately 12µm.
148 MARKS

to undergoonly primary branching(31, 43). Trichomeson both of thesemutantsgenerallyhavetwo branchesthatoriginatefrom a regioncloseto thetrichomebase(Figure 5).A mosaicanalysissuggeststhatAN actscell autono-mously(31). In an plantsthe leavesarenarrowandthestemsandsiliquesaretwisted.
TrichomeExpansion
Generaltrichomeexpansionis affectedby mutations in eightdifferentgenes.Theserecessivemutations include dis1 and dis2 as well as gnarled (grl),klunker(klk), spirrig (spi), wurm(wrm), crooked(crk), andalien (ali) (13,31).Trichomeson thesemutantsexhibit irregular expansion(illustrated by thedevelopingtrichomeson the dis2 mutantshownin Figure6). Becausethesemutantsdo not displayotherobviousphenotypes,the genesdefinedby thesemutations may be involved in expansionprocessesthat are unique to tri-chomes.Alternately,thesegenesmay encodeproductsimportant for generalcell expansion,but may be membersof genefamilies that are expressedinvariouscell types.As might be expectedfor this class,mosaicanalysisindi-catesthat DIS2 appearsto act cell autonomously (31). In contrast,the sameanalysisindicatesthat DIS1 may act througha noncell-autonomousmecha-nism becauseEMS-treatedplantslackeddis1 trichomesectors.As a control,theheterozygousdis1plantswerealsoheterozygousfor thean mutation, andan trichomesectorswere identified.
The singed(sne) mutation also resultsin a generalalterationof trichomecell expansion(62). The trichomesappearto developnormally, but the tri-chomestalk andbranchesareslightly twisted.In addition, this mutation alsocauses a shorteningof the roothairs.
MaturationMutants
Severalrecessivemutationsappearto alter the final stagesof trichomedevel-opment.Theunderdevelopedtrichome(udt) mutationresults intrichomesthatare slightly more slenderthan wild-type and that produceunderdevelopedpapillaetoward the tips of the branches(24). Threeothermutations,chablis(cha), chardonnay(cdo), and retsina (rts), result in trichomesthat lack therough papillate surfaceof wild-typematuretrichomes(31).
Potikha& Delmerusedan elegantscreento isolatea mutantdeficient insecondarycellulosedepositionin trichomecell walls (77). Trichomesexhibitstrongbirefringenceunderpolarizedlight, a characteristicof cell walls con-taininglargeamounts of highly orderedcellulosemicrofibrils.Theyidentifiedthe recessivetrichomebirefringence(tbr) mutant that lacks birefringenttri-
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 149
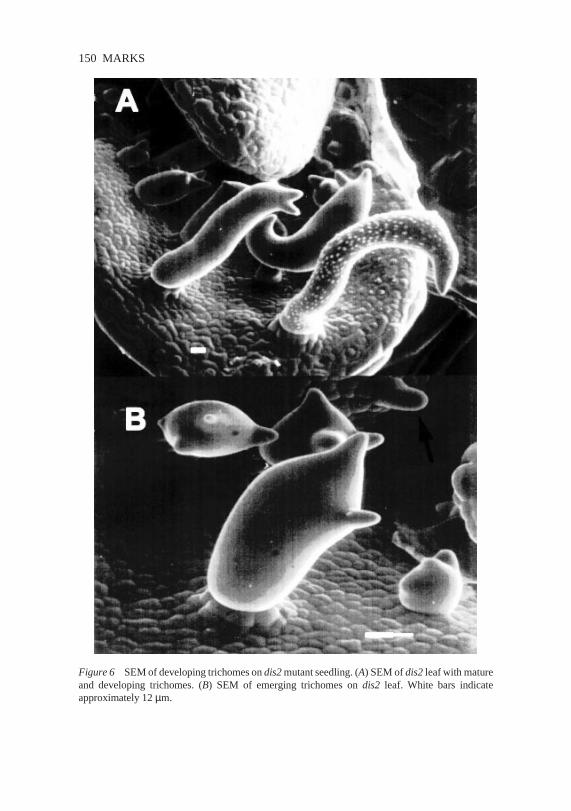
Figure6 SEMof developingtrichomesondis2mutantseedling. (A) SEMof dis2leafwith matureand developing trichomes. (B) SEM of emerging trichomes on dis2 leaf. White bars indicateapproximately12µm.
150 MARKS

chomes.Mature tbr trichomeswereessentially wild-type in shape,but hadasmoothsurfaceinsteadof the wild-type rough papillate surface.Quantificationof cellulosein trichomesisolatedfrom wild-type andtbr plantsindicatedthattbr trichomes have 18% of the cellulosenormally found in wild-type tri-chomes.The tbr mutation also may result in a reductionof cellulosein thexylem.
Genetic Interactions
Classicalgeneticanalysesindicatethat thegl1 and ttg mutationsareepistaticto all otherknowntrichome mutations.Plantsdoublymutantfor gl1 or ttg andother trichomemutantsare glabrous.The geneticrelationship betweengl2,gl3, and try hasalsobeenanalyzed(MD Marks, unpublisheddata;31). Thegl3/try doublemutanthasgl3-like trichomes,but retainstheclusteringpheno-typeof try. This suggeststhatGL3 functionsdownstreamof TRYfor branch-ing, andthatthewild-type functionof TRYmaybe toinhibit branch formationby directlyor indirectly inhibiting GL3 activity. It alsohas been foundthatthetry mutationsuppressesthegl2 phenotype;this suggeststhatTRYactsdown-streamof GL2. In this case,the function of GL2 could be to inhibit TRYactivity, resulting ina promotion of branch formation. The lack of aerialexpansionof gl2 mutant trichomescould result from unrestrictedwild-typeTRYactivity. The phenotype of thegl2/try and try/gl3 doublemutants wouldindicatethatthese three genesact in a simple linear pathway:
GL2—-| TRY—-| GL3.
Thephenotypeof thegl2/gl3 doublemutantsuggeststhat this is not thecase.With a simple linear relationship, one would predict that a gl2/gl3 doublemutant phenotypewould more closely resemblethat of gl3. Instead,thegl2/gl3 phenotypeis more extremethan either single mutant;even less tri-chomeoutgrowthis observedthanongl2 plants.While this resultdoesnot ruleout the possibility that a linear pathwayexists,it doessuggestthat GL2 andGL3have someseparate functionsactingthroughparallelpathways.
MOLECULAR STUDIES
GL1
GL1 wasoneof the first Arabidopsisgenesisolatedby T-DNA tagging.Thegl1-43 allele was identified in a population of plantsderivedfrom the 43rdtransformantgeneratedby the Agrobacterium-mediatedseedtransformationprocedure(15). Unlike strong loss of GL1 function mutationswith no tri-
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 151

chomeson either the stemsor leaves,gl1-43 lackedtrichomes only on thestems(16). An 8-kb fragmentthathadtheability to molecularly complementthegl1 mutation wasidentified(26). Sequencingthat theGL1 geneencodesamemberof themybclassof transcriptionfactors (74).
Mybgeneshavebeenfoundin all highereukaryoticorganismsandyeast(3,36,83,84,86).Theycontainoneto threemybdomains,eachof which appearto form a helix-turn-helix structure(17, 22, 59). Themybdomainsparticipatein DNA binding andarelocatedat the aminoterminus. The myb family wasfirst identifiedasanoncogeneassociatedwith theavianmyeloblastomavirus(83). The cellular c-mybgenewassubsequentlyidentified in vertebratesandwas found to play an important role in controlling the maturationof whiteblood cells (54). In animals,mybgenefamilies arecomposedof only a fewmembers.
The first isolated plant geneencodinga transcription factor was the mybgene,C1 (8, 75). The maizeC1 generegulatesanthocyaninsynthesis in thealeurone(6). Subsequentlyit was found that plantscontain large myb genefamilies (33, 65, 74). Membersof the family typically havetwo myb repeatstoward their amino termini anda divergentaminoacid sequencetoward thecarboxy-terminal(49). Most memberssharevery littl e sequenceidentity intheir carboxy-terminals. A few membershave onlyonemybdomain(2).
Severalplant myb genesother than GL1 and C1 havea known function.The maizePl genecontrolsseedlinganthocyaninsynthesis(7). The maizePgenecontrolsstepsin the synthesisof the red phlobaphenepigment in eartissue(23). The MIXTA geneof Antirrhinum participatesin controlling theshape of epidermalpetalcells(72).
TheGL1geneencodesaproteinwith two mybrepeatsandacarboxy-termi-nal domainof approximately 120aminoacidsthat is not significantly similarto othersequencesin thedatabases(49,74).Thecarboxy-terminalregiondoescontainseveralclustersof acidic aminoacidsthat may function astranscrip-tional activators(74). In theweakallelegl1-2 themolecularlesionis a smalldeletion that results in the loss of the terminal 27 amino acids (12). Themissingregioncontainsoneof the acidic clusters.It hasbeenfound that thegl1-1 allele containsa deletionremovingthe completeGL1 codingregionaswell as flankingpromoter elements,and theonly phenotypeis a loss oftrichomes(74). Thus, it is likely that the function of GL1 is restrictedtocontrollingtrichomedevelopment.
In situ hybridizationto localizeGL1 mRNA showedthatGL1 is expressedin fields of cells from which trichomesareinitiating (50). Cellscommitted tothe trichome pathwayhavemore GL1 mRNA activity than the surroundingcells,which might indicatethatautocatalytic up-regulationof GL1 expression
152 MARKS

occursoncea cell commits to the trichome cell fate. To examinethe DNAelementsresponsiblefor regulatingGL1 transcription, a GUS constructcon-tainingpromotersequencesfrom the5′ noncodingregionof GL1wasanalyzed(74). This constructdirectedGUSexpressionto thestipules,which arefoundat the baseof the leaves.Later studiesindicatedthat this expressionpatternappearsto inconsequentialand is not requiredfor the initiation of leaf tri-chomes(MD Marks,unpublisheddata).It is possible thatthestipule enhancerelementis requiredfor the expressionof a genelocated nearGL1.
The molecularlesion in the gl1-43 allele suggestedthat 3′ noncodingse-quencescouldcontainimportantregulatorysequences.This T-DNA–inducedallele containsan insertionthat is over 1000 basesdownstreamof the tran-scribed3′ noncodingsequences(74).At first, it appearedthatthis regiononlywasimportant for controlling GL1 expressionin thestembecausethepheno-type of the gl1-43 mutantis a loss of stemtrichomesbut not leaf trichomes(64).However,it wasfoundthatfusing5′ and3′ GL1 noncodingsequencestothe GUS constructresultedin a reconstructionof the patternof expressionfound by in situ hybridization (50). It also was found that the 3′ sequenceswereessentialfor GL1 function, becauseremovalof thesesequencesfrom aDNA fragmentcontaining theGL1 generesultedin a lossof molecularcom-plementation of both leaf and stemtrichomes on gl1 mutants(50). Sinceasequencedownstreamof theinsertion in gl1-43wasrequiredfor GL1 function,the reason theinsertionresultsonly in a lossof stemtrichomesis notknown.
The level of GL1 expressionis critical in controlling trichomeinitiation.Plantscontainingaconstructwith theGL1geneunderthecontrolof theCaMV35SRNA promoter(35SGL1)accumulategreaterthannormal levelsof GL1mRNA (48) andGL1 protein(D Szymanski& MD Marks,unpublisheddata).In either wild-type orgl1 mutants, thisoverexpressionof GL1 resultsin bothareductionof leaf trichomesandin aninduction of ectopictrichomeson organsthatarenormallyglabrous(48). The induction of ectopictrichomesis signifi-cant,becauseit indicatesthatexpressionof GL1 cancausetrichomeinitiation.Since TTG may be involved in causingthe reductionin leaf trichomeson35SGL1plants,a discussion of this phenomenonwill follow a review of thepossiblemolecularidentity of TTG.
TTG
TheTTGgenehasnot beenisolated.However,anongoingchromosomewalkhasnarroweddownthelocationof thegeneto a small regionon chromosome5 (A Walker,personalcommunication).TTGmaybeahomologof themaizeRgene(57). In maize,productionof theanthocyaninpigmentin thealeuroneis
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 153

controlledby the regulatorygenesC1 and R (6). R encodesa protein withhelix-loop-helix DNA binding and proteindimerization motifs characteristicof membersof the L-myc family of transcription factors(58, 78). Lloyd et al(57) demonstratedthat ArabidopsisplantsexpressingR underthe control oftheCaMV35Spromoter(35SR)synthesizedgreaterthannormalquantitiesofanthocyanin andhad extra trichomes. Thefirst two leaveshad a two- tofive-fold increasein trichomesover nontransformedcontrols.R expressionalsoresultedin ectopictrichomeson thepetals,stamens,andpistils. ttg plantstransformedwith R producedboth trichomesandanthocyanin.It wasshownsubsequentlythat R inducedthe synthesisof seedcoatmucilagein ttg plants(MD Marks,unpublisheddata)andaffectedtheexpressionof root hairs(18).While ttg mutantsnormally produceroot hairs in all files of the epidermis,35SRttg plantsproducedfew hairs,aswould bepredictedif TTG inhibits hairformation in the nonhairfiles of the root (18). The correctionof the diversedefectsin ttg plantsby R suggeststhatTTGmayencodeanR homolog.It alsois possible thatTTGregulatesanRhomolog (57).
A searchfor Arabidopsis cloneswith homologyto R hasnot resultedin theisolation of TTG. Furthermore,severalexpressedtags (cDNA clones)withsequencesimilarity to R havebeenidentified,but theydo not mapto TTG (AWalker, personalcommunication).Attempts to usethe R homolog in Antir-rhinum delila to complementttg mutantshavefailed (21, 71). Nevertheless,theR genehasproveda valuabletool to studyTTGfunctionandthe controloftrichomeinitiation.
Lloyd et al (56) useda novel approachto induceR activity in ttg plants.Theyattachedthesteroid-binding domainof therat glucocorticoid receptortothe carboxy-terminalof the R gene(RGR). Fusionof the binding domaintoother transcription factors previously had beenshown to imposehormonedependenceon the activity of the fusedfactor (11, 29). ttg plantscontainingRGR failed to producetrichomes.RGR ttg plants grown on agar mediumcontainingaslittl e as1 nM of dexamethasone produceda few trichomes(56).Plantsgrown on mediumcontaining10 nM dex had wild-type numbersoftrichomeson their first leaf pair. Nontransformedplants or plants carryingunmodifiedRdid not respond tothe dextreatment.
RGR plantswere usedto study trichomedevelopment (56). A plant wasgrown for eight dayson soil and then immersedin one µm dex. Sequentialscanningelectronmicroscopy(SEM) imageswereobtainedby makingcastsofmolds producedwith a dental impression polymer. Twenty-four hoursafterdex treatment,the surfaceof the youngestleaf was coveredwith emergingtrichomes.To define the spatialand temporalpatternof trichomeinitiation,plantswereeithergerminatedon dexandthenremovedfrom dexor theywere
154 MARKS

startedon medium without dex and then transferredto dex-containingme-dium. Leavesof plantsremovedfrom dex hadtrichomesonly at the tips andwere glabrousat the base.On plantstransferredto dex, the first leavesthatformedhadtrichomesonly at thebase,whereasthe later leaveswerecoveredwith trichomes.Theseresultsmirror thegeneralimpression that is formedbyexaminingwild-type leaves(Figure 1A): Trichomeformation proceedsas awavefrom thetip (oldesttissue)of the leaf to thebase(youngesttissue).Theresultsof Lloyd et al (56) extendthe analysisto indicatethat the patternoftrichomeinitiation is the resultof the timing of the competenceof epidermalcells torespondto TTG.
InteractionsBetween GL1and TTG
Plantsconstitutively expressingeitherGL1 or R wereusedto testhypothesesaboutthe rolesof GL1 andTTG in trichomedevelopment(48). To determinewhetherconstitutively expressedGL1 couldbypasstheneedfor TTG,crosseswere madebetween35SGL1 and ttg plants.Becauseglabrous35SGL1 ttgplantswereidentified in theF2 generation,it wasconcludedthatconstitutiveGL1 cannotbypassTTG. When 35SRplantswere crossedto a gl1 mutant,glabrous35SRgl1 plantswerefoundin theF2.Thus,it is likely thatconstitu-tive TTG cannotbypassthe requirementfor GL1. To determinethe effect ofconstitutively expressingboth GL1 and R, 35SGL1 and 35SR plants werecrossed.The constitutive expressionof both genesin the sameplant had adramatic impact on trichome initiation. The F1 plants exhibited abundanttrichomeson the hypocotyls andon both the adaxialandabaxialsurfacesofthecotyledons.Thefirst andlaterleavesweredenselycoveredwith trichomeson bothsurfaces,andleaf expansionwasseverelylimi ted.TheresultssuggestthatTTGandGL1 mayinteractto promotetrichomeinitiation (48). In supportof direct physical interaction,it hasbeenshownthat antibodies to GL1 canco-precipitateGL1 and Rproteins(Marks& Jilk, unpublisheddate).
The results also have implications for interpreting the phenotype of35SGL1 plants (48). Theseplants make fewer than normal leaf trichomes.However,this phenotypeis suppressedin 35SGL1/35SRplants.Two modelshavebeenproposedto explainthe reductionin leaf trichomes.1. ExcessfreeGL1 protein(not boundto targetDNA sequences)in thenucleustitratesTTGfrom complexeswith GL1 boundto targetgenes.If transcriptionalactivationrequiresthe interactionof TTG andGL1 proteinat the targetpromoter,thenthe titration of TTG by free GL1 could lower the expressionof targetgenesand,in turn, resultin fewer trichomes.ExcessTTG protein(in theform of R)couldpreventthe titration.2. ExcessGL1 activatesgenesthatparticipatein a
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 155

lateralinhibition of trichomeinitiation, resultingin fewer trichomes.The twomodelsarenot mutually exclusive;it is possiblethat the lateral inhibition iscontrolledin part bythe ratioof a TTG-GL1complexvs free GL1.
The hypothesis that GL1 and TTG interactto control lateral inhibition issupportedby theinteractionbetweenweak alleles ofGL1andTTG(JA Larkin& MD Marks,unpublisheddata).As describedabove,it hasbeenfound thattheplantsdoubly heterozygousfor both weakttg andgl1 mutantalleleshavegreaterthannormalnumbersof clusteredtrichomes;that is, lateralinhibitionappearsto be reduced.Larkin et al (48) found that plantswith one or twocopies of the 35SGL1 (35SGL1/-) construct and heterozygous for TTG(TTG/ttg) havea greaternumberof leaf trichomesthanplantsthathaveoneortwo copiesof 35SGL1 in a homozygous TTG background.Approximately30% of the trichomes on 35SGL1/-TTG/ttg were presentin clusters.Thissuggeststhat the levelsof GL1 andTTG expressionareimportant in control-ling lateral inhibition.
GL2
TheGL2 gene,like GL1, wasisolatedby T-DNA tagging(82). In a screenof10,000transformedlines, sevenindependentgl2 mutants were isolatedbutonly one of these,gl2-2, had a T-DNA insert linked to the GL2 locus. Awild-type GL2 fragmentwas definedby complementation of transgenicgl2(82). GL2 encodesa 744aaproteinthathasanamino-terminalacidicdomainfollowed by thehomeodomain (82). Directly downstreamof thehomeoboxisa motif that could encodean antipathic helix that could promote proteindimerization.The aminoacid sequencedownstreamof the putativehelix se-quencedoesnot showsignificantsimilarity to anyproteinin thedatabases.Acomparisonof planthomeodomain genesindicatesthatGL2 is mostsimilar toa classthathasa leucinezipperdomainon thecarboxylsideof thehomeodo-main(38).
In situ hybridization analysis indicates that GL2 mRNA is expressedstronglyin developingtrichomes(82). Immunolocalizationof theGL2 proteinand the analysisof plants containinga GL2 promoterGUS reportergeneconstruct(GL2GUS) indicate a more complex patternof expression(MDMarks & D Szymanski,unpublisheddata).Approximately 2000 basesof 5′noncodingsequencewere fused to the coding sequenceof GUS to createGL2GUS. The staining patternof GL2GUS plants indicatesthat GL2, likeGL1, is expressedin fields of epidermalcells beforetrichome initiation. Thelevel ofGL2expressionincreases incells committing to the trichome cell fate.GL2 also is expressedin mesophyll cells; GL2 proteinsappearin both the
156 MARKS

cytoplasmand nucleusof nontrichomecells. In developingtrichomesGL2proteinis primarily localizedto thenucleus.
By geneticanalysis,GL2 functionsdownstreamof GL1, but GL1 doesnotcontrol the nontrichomeexpressionpatternof GL2. Furthermore,the non-trichomeexpressionpatternis observedin ttg mutants.Consequently, genesotherthanGL1 or TTGcontrol theearlyexpressionpatternof GL2.GL2GUSis stronglyexpressedin themalformedtrichomespresenton gl2 mutants.ThisresultsuggeststhatGL2 proteindoesnot regulateits own expressionin devel-opingtrichomes.
The analysisof GL2 promoterdeletionshas identified a region that isimportantfor controlling GL2 expressionin trichomes.TheentireGL2 codingsequenceandapproximately1.5 kb of 5′ noncodingsequencecould molecu-larly complementgl2. Removalof approximately 125basesfrom the 5′ end ofthis fragmentyielded partial complementation (82). This region containsamyb consensusbindingsite.Thus,althoughtheGL1 proteindoesnot regulatetheearlyexpressionpatternof GL2, it or anothermyb proteincould influencetheexpressionof GL2 in developingtrichomesby bindingto themybbindingsite.
ZWI
Wild-type leaf trichomesnormally havea stalk and threeto four branches,whereaszwi mutanttrichomeshavea shortenedstalk andonly two branches.Threeindependentzwi mutantswereisolatedfrom Feldmann’s T-DNA lines.Two of thesehadinsertsthat co-segregatedwith the mutantphenotype(73).Plasmidrescuewas usedto isolateplant DNA flanking the insertions(14).Sequenceanalysisof the region flanking oneof the insertsrevealedthat theZWI encodesa large kenesin-likeprotein (73). Thesegenefragmentswereusedto isolate an intact geneand flanking DNA. A 27-kb fragmentcouldmolecularlycomplement thezwi mutant.
Kinesinsaremicrotubule motorproteinscharacterizedby a conservedheaddomain that comprisesthe motor domain and a nonconservedtail region,which is thoughtto participatein binding cargo(20). ZWI encodesa kinesinwith themotordomainlocatedtowardthecarboxyterminus (73). In addition,the 5′ portion of ZWI encodesa regionwith similarity to a classIV myosinfoundin Acanthamoeba(30).Thefunctionof class IV myosins and therole ofthe regionwith similarity are unknown.
While the characterizationof ZWI wasbeingcompleted,the sequencebe-cameavailable forakinesin-like proteinfrom Arabidopsis, isolated becauseofits ability to bindcalmodulin (79). Sequence comparisonbetweenZWIandthe
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 157

kinesin-like calmodulin-binding protein (KCBP) indicateonly a few nucleo-tide differences,and Southernhybridization analysisindicated that only asinglecopy of a genewith high similarity to eitherZWI or KCBP exists(73,79).Thus,ZWIandKCBPare thesamegene.Thecalmodulin-bindingdomainis composedof a 21-aminoacid sequencelocatedvery closeto the carboxyterminus oftheprotein(79).Binding studies indicateZWI bindsto calmodulinwith high affinity in the presenceof Ca2+ (1 µm). ThesestudiesstronglysuggestthatCa2+ and calmodulin are involvedin regulatingZWIactivity.
Northernhybridization analysisshowedthat ZWI is expressedin flowers,leaves,roots,andculturedcallustissues(73,79).Theonly discerniblepheno-typeof thezwi mutantis theabnormalbranchingandcell expansionpatternoftrichomes.It is possible that only partial lossof function allelesof zwi havebeenidentified. However,one of the T-DNA insertionsdisruptsthe regionencodingtheATP-bindingandmicrotubule-bindingdomains(73). The inser-tion producesan in-framestopcodonthat would most likely result in a ZWIprotein lacking motor function.Thereareat leastthreeexplanationsthat canaccountfor the lack of phenotype,asidefrom alteredtrichomeformation,inplantshomozygousfor this insertion.1. It is possible that ZWI expressionisrequiredonly in trichomes. 2.Anothergenecanfunctionin placeof ZWI in thenontrichomecells,but not in trichomes.3. Only theaminoterminalof ZWI isrequired innontrichomecells.
The ZWI promotersequenceshavenot yet beenidentified, but it appearsthat like GL1, sequencesin the3′ OTR of ZWI arerequiredfor its regulation(50, 73). The insertionin oneof the taggedmutantsis locatedtwo kb down-streamof thepolyadenylation site.This insertion maydisrupta trichome-spe-cific transcriptional enhancer.
OtherGenes
Two othergeneshavebeenclonedthatappearto haveimportantfunctionsincontrolling trichome initiation.Thefirst is theCONSTITUTIVEPHOTOMOR-PHOGENIC(COP1) gene(9). COP1wasshownto encodea proteinwith azinc finger domainin theaminoterminal,followed by a coiled-coiledzipper-like domain,and endswith WD-40 repeats(10). All thesedomainscouldparticipatein protein-proteininteractions.ApparentlyCOP1 functionsto re-pressphotomorphic genesin roots and in shootsof dark-grownplants(87).COP1may actat the level of transcriptionbecauseCOP1proteinis found inthe nucleusin dark-grownplants. In light-grown shoots,COP1 protein islocalizedto the cytoplasm. Miséraet al characterizedplantscarryinga lethalallele of COP1called fusca1(fus1) (70). Plantshomozygous for fus1die as
158 MARKS

young seedlings.Furthermore,theseseedlingsaccumulatehigh amountsofanthocyanin.To study the function of COP1 in the adult seedling,heterozy-gous fus1 seedswere treatedwith EMS to induce fus1/fus1 sectorsin theresultingseedlings.Mesophyllsectorshadunderexpandedcells that accumu-lated anthocyanin(70). Epidermalsectors,in contrast,exhibitedectopictri-chomes.A modelhasbeenproposedin which COP1 is anupstreaminhibitorof TTG(70).
Recentlyanotherttg-like mutant,ttg2, hasbeenisolated(34). This mutanthas a reducednumberof trichomesthat are less branchedthan normal. Inaddition, like ttg1, ttg2 mutant seedslack seedcoat mucilage and are lesspigmented.The genemutation mapsto the bottom of chromosome2. Themutant was isolated from a population of plants carrying the maize Actransposonthat mobilized the endogenoustag elementinto the gene(34).Isolationandcharacterizationof thegenehasshownthat it encodesa productwith similaritiesto theSPF1proteinof sweetpotatothatbindsSP8sequencesin thepromoterof beta-amylaseand sporamingenes (32).
CONCLUSIONSAND FUTUREDIRECTIONS
Many mutations definegenesthataffectdiscreteaspectsof trichomedevelop-ment.Geneticanalysesindicatethat someof thesegenesact in linear path-ways,whereasotherspossibly work throughunrelatedparallelpathways(31,49). Molecularanalysesof severalclonedgenesled to proposalsfor specificfunctionsfor GL1, GL2, TTG (if TTG is anR homolog),TTG2,andZWI.Thefirst four genesencodetranscription factors.For thesetranscriptionfactors,thelay questionis how theyareregulatedandwith which factorsdo theyinteract.How do theseinteractionsinfluencetheir function?Whatgenesdo theyregu-late?The answersto thesequestions should identify genesacting both up-streamanddownstreamof thesefactors.The characterizationof manyof theregulatedgeneswill extendtheanalysisof trichomedevelopmentto thecellu-lar level. However,understanding how the expressionof variousstructuralproteins and enzymes, whicharelikely targets oftheregulatorygenes,leadstotrichomedifferentiationwill be challenging.
A geneticanalysis will beessential. Oftenthe sequencesof the genesgeneticallyidentified will be informative.For example,ZWI encodesa genewith a microtubule motordomain. This indicatesthat thecytoskeleton playsarole in trichome morphogenesis, andZWInow becomesa reagent toprobe thisrole.Some “trichome genes”will turn out tobegenespreviously characterizedin a different context.Again ZWI providesan example;KCBP(=ZWI) wasisolatedbecauseof its calmodulin-bindingdomain. The analysisof ZWI can
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 159

define how Ca2+ and calmodulin regulatea developmentalprogram.In thenearfuture, many geneticallyidentified trichomegeneswill be isolatedandcharacterized.The analysisof thesegeneslikely will result in the charac-terizationof many different aspectsof trichomecell type determination anddifferentiation.
Visit the Annual Reviews home pageat http://www.annurev.org
LiteratureCited
1. ÅgrenJ, SchemskeD. 1994. Evolution oftrichome number in a naturalized popula-tionof Brassica rapa. Am.Nat. 143:1–13
2. Baranowskij N, Frohberg C, Prat S,Wil lmitzer L. 1994. A novel DNA bindingprotein with homology to Myb oncoprote-inscontainingonly onerepeatcanfunctionasa transcriptional activator. EMBOJ. 13:5283–392
3. BiesalskiHK, DoepnerG, TzimasG, Ga-mulin V, SchroderHC, et al. 1992. Modu-lation of myb gene expressionin spongesby retinoic acid. Oncogene7:1765–74
4. Bowley SR,Lackle SM. 1989. Geneticsofnonglandular stemtrichomesin RedCover(TrifoliumpratensL.).J.Hered.80:472–74
5. ClarkPJ,EvansFC.1954.Distancetonear-estneighbor asa measureof spatial rela-tionships in populations. Ecology 35:445–53
6. Coe EH, Neuffer MG, Hoisington DA.1988. The genetics of corn. In Corn andCorn Improvement. Agron. Monogr. No.18, ed. GF Sprague, JW Dudley, pp. 81–236. Madison, WI: Am. Soc.Agron.
7. Cone KC, CoccioloneSM, MoehlenkampCA, WeberT, Drummond BJ, et al. 1993.Role of theregulatory genepl in thephoto-control of maize anthocyanin pigmenta-tion. Plant Cell 5:1807–16
8. ConeKC, Burr FA, Burr B. 1986. Molecu-lar analysisof themaizeanthocyanin regu-latory locusC1.Proc.Natl. Acad. Sci. USA83:9631–35
9. DengX-W, CasparT, Quail P. 1991. cop1:aregulatory locusinvolvedin light-control-led development and gene expression inArabidopsis.GenesDev. 5:1172–82
10. DengX-W, Matsui M, Wei N, Wagner D,Chu A, et al. 1992. COP1, anArabidopsisregulatory gene, encodes a protein withboth azinc-binding motif andGβ homolo-gousdomain. Cell 71:791–801
11. EilersM, PicardD, Yamamoto K, BishopJ.1989. Chimaerasof myconcoproteinandsteroid receptors causehormone-depend-ent transformation of cells. Nature 340:66–68
12. Esch JJ, Oppenheimer DG, Marks MD.1994. Characterization of a weakalleleofthe GL1 gene of Arabidopsis thaliana.Plant Mol. Biol. 24:203–7
13. Feenstra WJ. 1978. Contiguity of linkagegroups I and IV as revealed by linkagerelationship of two newlyisolatedmarkersdis-1anddis-2.Arab. Inf. Serv. 15:35–38
14. FeldmannK. 1992. T-DNA insertion mu-tagenesis in Arabidopsis: seedinfection/transformation. In Methodsin ArabidopsisResearch, ed. C Koncz, N-H Chua, JSchell, pp. 274–89. Singapore:World Sci.
15. FeldmannKA, MarksMD. 1987.Agrobac-terium-mediatedtransformation of germi-nating seedsof Arabidopsis thaliana: anontissue culture approach. Mol. Gen.Genet.208:1–9
16. Feldmann KA, Marks MD, ChristiansonML, QuatranoRS.1989.A dwarfmutantofArabidopsisgeneratedbyT-DNA insertionmutagenesis.Science243:1351–54
17. Frampton J, Leutz A, Gibson T, Graf T.1989. DNA-binding domainancestry. Na-ture342:134
18. GalwayME, MasucciJD,LloydAM, Wal-bot V, Davis RW, Schiefelbein JW. 1994.TheTTGgeneisrequiredtospecify epider-mal cell fate and cell patterning in theArabidopsisroot. Dev. Biol. 166:740–54
19. GoffredaJC,Szymkowiak EJ,SussexIM,Mutschler MA. 1990. Chimeric tomatoplantsshow that aphid resistance andtri-acylglucose productionare epidermalautonomous characters.Plant Cell 2:643–49
20. Goldstein LSB. 1993. With apologies toScheherazade:tailsof 1001kinesinmotors.Annu. Rev. Genet. 27:319–51
160 MARKS

21. GoodrichJ, CarpenterR, Coen E.1992. Acommon gene regulatespigmentation pat-tern in diverse plant species. Cell 68:955–64
22. GrafT. 1992. Myb:atranscriptionalactiva-tor linking proliferation anddifferentiationin hematopoietic cells. Curr. Opin. Genet.Dev. 2:249–55
23. Grotewold E,Drummond B, BowenB, Pe-terson T. 1994. The myb-homologous Pgenecontrolsphlobaphenepigmentationinmaizefloral organsby directly activating aflavonoid biosynthetic gene subset. Cell76:543–53
24. Haughn GW, Somerville CR. 1988. Ge-netic control of morphogenesisin Arabi-dopsis. Dev. Genet. 9:73–89
25. HeldLI. 1991. Bristlepatterning in Droso-phia. BioEssays13:633–40
26. HermanPL, Marks MD. 1989. Trichomedevelopment in Arabidopsis thaliana. II.Isolation and complementation of theGLABROUS1gene.Plant Cell 1:1051–55
27. Holt J,BirchN.1984.Taxonomy,evolutionand domestication of Vicia in relation toaphid resistance. Ann. Appl. Biol. 105:547–56
28. HombergenE-J,BachmanK. 1995. RAPDmapping of three QTLs determing tri-chomeformationin MicroserishybridH27(Asteraceae: Lactuceae). Theor. Appl.Genet. 90:853–58
29. HopeTJ,HuangXJ,McDonaldD, ParslowT. 1990. Steroid-receptor fusion of the hu-man immunodeficiency virus type 1 Revtransactivator: mapping cryptic functionsof thearginine-richmotif .Proc.Natl.Acad.Sci. USA87:7787–91
30. HorowitzJ,HammerJI.1990.A newAcan-thamoeba myosin heavy chain. J. Biol.Chem.265:20646–52
31. HülskampM, MiséraS, Jürgens G. 1994.Genetic dissection of trichome cell devel-opmentin Arabidopsis.Cell 76:555–66
32. Ishiguro S, Nakamura K. 1994. Charac-terization of a cDNA encoding a novelDNA-binding protein, SPF1, that recog-nizes SP8 sequences in the 5’ upstreamregions of genescoding for sporaminandbeta-amylasefromsweetpotato.Mol. Gen.Genet. 244:563–71
33. JacksonD,Culianez-MaciaF,Prescott AG,RobertsK, MartinC.1991.Expressionpat-ternsof mybgenesfrom Antirr hinum flow-ers.Plant Cell 3:115–25
34. JohnsonC, Symth D. 1996. A gene fromArabidopsis that regulatestrichome devel-opment, seedpigmentation and mucilageproduction. Proc.Aust. Soc.Biochem.Mol.Biol. 28:In press
35. Johnson HB. 1975. Plant pubescence:anecological perspective. Bot. Rev. 41:233–58
36. Katzen AL, Kornberg TB, Bishop JM.1985. Isolation of the proto-oncogene c-myb from D. melanogaster. Cell 41:449–456
37. Kennedy G, Sorenson C. 1985. Role ofglandular trichomes in the resistance ofLycopersion hirsutum f. glabratum toColorado potato beetle (Coleoptera:Chrysomelidae). J. Econ. Entomol.78:547–51
38. KerstetterR,VollbrechtE,LoweB, VeitB,Yamaguchi J, Hake S. 1994. Sequenceanalysisandexpressionpatternsdividethemaizeknotted1-like homeobox genesintotwo classes.Plant Cell 6:1877–87
39. KhanZR,WardJT, Norris DM. 1986. Roleof trichomesin soybeanresistanceto cab-bagelooper, Trichoplusiani. Entomol. Exp.Appl. 42:109–17
40. KlothRH.1995. Interactionof twoloci thataffecttrichomedensity in upland cotton. J.Hered.86:78–80
41. KlothRH.1993. Newevidencerelating thepilosealleleandmicronairereadingin cot-ton. CropSci.33:683–87
42. Koornneef M. 1981. The complex syn-drome of ttg mutants. Arab. Inf. Serv. 18:45–51
43. KoornneefM, DellaertSWM,vanderVeenJH.1982. EMS- andradiation-inducedmu-tation frequencies at individual loci inArabidopsis thaliana (L) Heynh. Mutat.Res.93:109–23
44. Lamb R. 1980. Hairs protectpods ofmus-tard(Brassicahirta‘gisilba’) fromfleabee-tle feeding damage.Can. J. Plant. Sci. 60:1439–40
45. Lamb R. 1982. Economics of insecticidalcontrol of flea beetles (Coleoptera:Chrysomelidae)attacking rapein Canada.Can. Entomol. 114:827–40
46. LambR.1984.Effectsof fleabeetles,Phyl-lotreta ssp. (Chrysomelidae:Coleoptera),on the survival, growth, seedyield andquality of canola,rapeandyellowmustard.Can. Entomol. 116:269–80
47. Larkin JC,Young N, Prigge M, Marks M.1996. Thecontrol of trichomespacing andnumber inArabidopsis. Development 122:997–1005
48. LarkinJC,OppenheimerDG,LloydA, Pa-parozzi ET, MarksMD. 1994. TherolesofGLABROUS1 andTRANSPARENT TESTAGLABRAgenesin Arabidopistrichomede-velopment.Plant Cell 6:1065–76
49. Larkin JC,OppenheimerDG, Marks MD.1994. The GL1 gene andthe trichomede-
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 161

velopmental pathway in Arabidopsisthaliana. In Results and Problemsin CellDifferentiation 20: Plant Promoters andTranscription Factors, ed. L Nover, pp.259–75. Berlin: Springer-Verlag
50. Larkin JC, Oppenheimer DG, Pollock S,Marks MD. 1993. ArabidopsisGLABROUS1 gene requires downstreamsequences for function. Plant Cell5:1739–48
51. LawsonEJR,Scofield SR,Sjodin C,JonesJDG,DeanC.1994. Modification of the5’untranslatedleaderregionof themaizeAc-tivator elementleads to increasedactivityin Arabidopsis. Mol. Gen. Genet. 245:608–15
52. Lee JA. 1985. Revision of the genetics ofthe hairiness-smoothness system ofGossypium. J. Hered.76:123–26
53. Levin DA. 1973. Therole of trichomesinplantdefense.Q. Rev. Biol. 48:3–15
54. Lipsick JS, Baluda MA. 1986. The myboncogene. In Gene Amplification andAnalysis,Vol. 4,Oncogenes,ed.TSPapas,GF Vande Woude, pp. 73–98. New York:Elsevier
55. Lister C, DeanC. 1993. Recombinant in-bred linesfor mapping RFLP andpheno-typic markers in Arabidopsis thaliana.Plant J. 4:745–50
56. LloydAM, SchenaM, WalbotV,DavisRW.1994. Epidermalcell fatedetermination inArabidopsis:patterns definedby a steroid-inducible regulator. Science266:436–39
57. Lloyd AM, Walbot V, Davis RW. 1992.Anthocyanin productionin dicotsactivatedby maizeanthocyanin-specific regulators,RandC1. Science258:1773–75
58. Ludwig SR, Habera LF, Dellaporta SL,WesslerSR. 1989. Lc, a memberof themaizeRgenefamily responsiblefor tissue-specific anthocyaninproduction, encodesaprotein similar to transcription factors andcontains the Myc homology region. Proc.Natl. Acad. Sci. USA86:7092–96
59. LüscherB, EisenmanRN.1990. NewlightonMyc andMyb. PartII. Myb.GenesDev.4:2235–41
60. Marks MD. 1994. The making of a planthair. Curr. Biol. 4:621–23
61. Marks MD, EschJ, HermanP, Sivakuma-ranS,OppenheimerD. 1991. A model forcell-typedetermination anddifferentiationin plants. In Molecular Biology of PlantDevelopment,ed.GJenkins,WSchuch,pp.77–87. Cambridge:Co. Biol.
62. Marks MD, EschJJ.1992. Trichome for-mation in Arabidopsis asa genetic modelfor studying cell expansion. Curr. Top.Plant Biochem.Physiol. 11:131–42
63. Marks MD, Esch JJ. 1994. Morphologyanddevelopment of mutantandwild typetrichomes on the leavesof Arabidopsisthaliana. In Arabidopsis:AnAtlasof Mor-phology and Development, ed.J Bowman,pp. 56–73. NewYork: Springer-Verlag
64. MarksMD, FeldmannKA. 1989.Trichomedevelopmentin Arabidopsisthaliana. I. T-DNA tagging of the GLABROUS1gene.Plant Cell 1:1043–50
65. MaroccoA, WissenbachM, BeckerD,Paz-AresJ, Saedler H, Salamini F. 1989. Mul-tiplegenesaretranscribedin Hordeumvul-gare and Zea mays that carry the DNAbinding domain of the myb oncoproteins.Mol. Gen.Genet. 210:183–87
66. MasucciJ,RerieW, ForemanD, ZhangM,GalwayM, etal.1996.ThehomeoboxgeneGLABRA2 is requiredfor position-depend-entcelldifferentiationin theroot epidermisof Arabidopsisthaliana.Development122:1253–60
67. McKelvie A. 1965. Preliminary data onlinkagegroups in Arabidopsis. Arab. Inf.Serv. 1S:79–84
68. Meisner J, Mitchell B. 1983. Phagodeter-rencyinducedby two cruciferousplants inadults of the fleabeetle Phyllotreta strio-lata (Coleoptera: Chrysomelidae). Can.Entomol. 115:1209–14
69. MelaragnoJ,MehrotaB,ColemanA. 1993.Relationship betweenendoploidy andcellsize in epidemal tissue of Arabidopsis.Plant Cell 5:1661–68
70. MiséraS,MüllerA, Weiland-HeideckerU,Jürgens G. 1994. The FUSCA genes ofArabidopsis: negative regulators of lightresponses.Mol. Gen.Genet. 244:242–52
71. MooneyM, DesnosT, Harrison K, JonesJ,CarpenterR,CoenE.1995. Alteredregula-tion of tomato and tobaccopigmentationgenescausedby the delila gene of Antir -rhinum.Plant J. 7:333–39
72. Noda K, Glover B, LinsteadP, Martin C.1994. Flower colour intensity depends onspecializedcell shapecontrolledbyaMyb-related transcription factor. Nature 369:661–64
73. Oppenheimer DG, Esch J, Marks MD.1992. Molecular genetics of Arabidopsistrichomedevelopment. In Control of PlantGene Expression, ed. DPS Verma, pp.275–86. BocaRaton, FL: CRC
74. OppenheimerDG, HermanPL, EschJ,Si-vakumaranS,MarksMD. 1991. A myb-re-latedgenerequiredfor leaftrichomediffer-entiation in Arabidopsis is expressedinstipules.Cell 67:483–93
75. Oppenheimer DG, Pollock M, Vacik J,Ericson B, FeldmannK, Marks M. 1996.
162 MARKS

Essential role of a novel kinesin-like pro-tein in Arabidopsis trichome morphogene-sis.Submitted
76. Paz-AresJ, Ghosal D, WienandU, Peter-sonPA, SaedlerH. 1987. Theregulatory c1locusof Zeamaysencodesa protein withhomology tomybproto-oncogeneproductsandwithstructural similaritiesto transcrip-tional activators.EMBOJ. 6:3553–58
77. Potikha T, DelmerD. 1995. A mutant ofArabidopsis thaliana displaying alteredpatterns of cellulose deposition. Plant J.7:453–60
78. PuruggananMD, WesslerS.1994.Molecu-lar evolutionof theplantR regulatory genefamily. Genetics138:849–54
79. Reddy A, Safadi F, Narasimholu S,Golovkin M, Hu X. 1996. A novel plantcalmodulin-binding protein with a kinesinheavy chain motor domain. J. Biol. Chem.271:7052–60
80. RédeiGP. 1967. Genetic estimateof cellu-lar autarky. Experientia 23:584
81. ReevesAF Jr. 1977. Tomato trichomesandmutationsaffecting theirdevelopment.Am.J. Bot. 64:186–89
82. Rerie WG, Feldmann KA, Marks MD.
1994. The GLABRA2 gene encodesaho-meo domain protein required for normaltrichome development in Arabidopsis.Genes Dev. 8:1388–99
83. RousselM, Saule S, Lagrou C, RommensC, Beug H, etal. 1979. Threenewtypesofviral oncogeneof cellular origin specificfor haematopoieticcelltransformation.Na-ture281:452–55
84. Stober-Grasser U, Brydolf B, Bin X,Grasser F, Firtel RA, Lipsick JS. 1992.Dictyostelium MYB: evolution ofa DNA-binding domain. Oncogene7:589–96
85. Telfer A, Poethig A. 1994. Leaf develop-mentin Arabidopis. In Arabidopsis,ed.EMeyerowitz,C Somerville.Plainview, NY:Cold SpringHarbor
86. Tice-Baldwin K, Fink GR,Arndt KT.1989.BAS1hasanMybmotif andactivatesHIS4transcription only in combination withBAS2. Science246:931–35
87. vonArnim A, DengX-W.1994.Light inac-tivationof Arabidopsis photomorphogenicrepressorCOP1 involves a cell-specificregulation of its nucleocytoplasmic parti-tioning. Cell 79:1035–45
TRICHOMEDEVELOPMENTIN ARABIDOPSIS 163









![The m6A Reader ECT2 Controls Trichome Morphology by ... · The m6A Reader ECT2 Controls Trichome Morphology by Affecting mRNA Stability in Arabidopsis[OPEN] Lian-Huan Wei,a Peizhe](https://img.pdfslide.us/doc/110x75/5f076dc27e708231d41ceff2/the-m6a-reader-ect2-controls-trichome-morphology-by-the-m6a-reader-ect2-controls.jpg)
![A Large-Scale Genetic Screen in Arabidopsis to …A Large-Scale Genetic Screen in Arabidopsis to Identify Genes Involved in Pollen Exine Production1[C][W][OA] Anna A. Dobritsa*, Aliza](https://img.pdfslide.us/doc/110x75/5f7232dfe2cc56738026b15f/a-large-scale-genetic-screen-in-arabidopsis-to-a-large-scale-genetic-screen-in-arabidopsis.jpg)

![The YTH Domain Protein ECT2 Is an m6A Reader Required for ...Nov 06, 2017 · The YTH Domain Protein ECT2 Is an m6A Reader Required for Normal Trichome Branching in Arabidopsis[OPEN]](https://img.pdfslide.us/doc/110x75/6038cfad82c8025670713575/the-yth-domain-protein-ect2-is-an-m6a-reader-required-for-nov-06-2017-the.jpg)
















