Embed Size (px)
Citation preview

BRIEF REPORT
Molecular evolution of the VP7 gene of Japanese G2 rotavirusesbefore vaccine introduction
L. P. Do • T. Nakagomi • Y. H. Doan •
Y. Kitahori • O. Nakagomi
Received: 10 May 2013 / Accepted: 20 June 2013
� Springer-Verlag Wien 2013
Abstract Changes in the prevalence of G2 rotavirus after
vaccine introduction are an important issue. However, such
changes in a given country should be interpreted in the
global context over time. We determined 35 Japanese G2
sequences and compared them with 508 globally collected
G2 sequences. The D96N substitution, a substitution
known to be associated with an abrupt increase in G2
strains and antigenic changes, emerged in those strains that
formed a nascent lineage outside of the currently pre-
dominant lineage (sublineage IVa). Further studies are
warranted to monitor the potential of their global spread,
since they also appeared in Europe and Australia.
Rotavirus A is the most important etiological agent of severe
acute gastroenteritis in infants and young children world-
wide. Two outer capsid proteins, VP7 and VP4, are inde-
pendently involved in virus neutralization and define the G
type and the P type, respectively. More than three quarters of
human rotaviruses carry G1P[8], G2P[4], G3P[8], G4P[8]
and G9P[8], of which approximately 12 % are G2P[4] [2].
Currently, two globally licensed rotavirus vaccines are
available: a monovalent G1P[8] human rotavirus vaccine
(Rotarix, GlaxoSmithKline Biologicals, Rixensart, Bel-
gium) and a pentavalent bovine-human reassortant rotavirus
vaccine (RotaTeq, Merck and Co., Whitehouse Station, PA,
USA). A marked increase and the predominance of G2P[4]
rotavirus strains were observed concurrently with the intro-
duction of the universal mass vaccination program with the
monovalent vaccine in Brazil [6, 12, 14]. In Australia, where
two rotavirus vaccines were used in different regions, fol-
low-up studies after vaccination showed that in the states
using RotaTeq, G1P[8] was predominant, followed by
G2P[4], whereas in the states using Rotarix, G2P[4] was
predominant, followed by G1P[8] [9, 17]. On the other hand,
a similarly high prevalence of G2P[4] was observed in other
countries where no rotavirus immunization program was
implemented, such as Argentina and Paraguay, two neigh-
boring countries of Brazil [5, 10].
Thus, it is important to interpret the changes in the
prevalence of G2 strains after vaccine introduction in a
given country from the phylogenetic perspective over time.
In Japan, Rotarix was introduced into the private market as
the first rotavirus vaccine in November 2011, and RotaTeq
followed in July 2012. Because there were only 20 Japa-
nese G2 VP7 gene sequences available in the GenBank
database, this study was undertaken to determine the VP7
sequence of the G2 rotavirus strains collected in Japan over
the last 31 years (1980 to 2011) to understand how G2
rotavirus strains evolved over time under unvaccinated
conditions. In the phylogenetic analysis, we placed a spe-
cial emphasis on the amino acid substitution at position 96
because there was evidence that a D96N substitution in
VP7 was related to an increase or the re-emergence of G2
strains, as reported from Taiwan, Thailand, Bangladesh,
South Africa, the United Kingdom, and Nepal [1, 4, 7, 13,
19, 20]. Furthermore, Doan et al. [4] clarified that the
D96N substitution is the salient feature of virtually all G2
strains circulating in the world for the last decade.
A total of 35 rotavirus strains analyzed in this study
were isolated from archival stool specimens obtained from
L. P. Do � T. Nakagomi � Y. H. Doan � O. Nakagomi (&)
Department of Molecular Microbiology and Immunology,
Graduate School of Biomedical Sciences, Nagasaki University,
1-12-4 Sakamoto, Nagasaki 852-8523, Japan
e-mail: [email protected]
Y. Kitahori
Nara Prefectural Institute for Hygiene and Environment,
Nara, Japan
123
Arch Virol
DOI 10.1007/s00705-013-1804-6

MMA08-4/Myanmar/2008 mani-268/India/2007
Matlab6/Bangladesh/2004 CU110-BK/Thailand/2008
05N011/Nepal/2005 16054 09MA/Brazil/2009 LB2764/USA/2005-06
GER31/Germany/2008 DC2020/Madagascar/2004
Seoul-397/South Korea/2009 B110056/Japan/2011
MR 4717/Mauritius/2000 XJ00-486/China/2000
Bangla66/Bangladesh/1993 NG5113/Negeria/1999
GH1803/Ghana/1999 4476PT/South Africa/1997
80123/Japan/2008 80120/Japan/2008
CH-130/China/2003 90096/Japan/2009
GER84/Germany/2008 26-Palermo/Italy/2003 07Y003/Japan/2007
80004/Japan/2008 Arg2395/Argentina/2007
Seoul-556/South Korea/2009 LB2772/USA/2005-06
14322 07MG/Brazil/2007 01P017/Japan/2001
01P007/Japan/2001 01P012/Japan/2001
01P129/Japan/2001 01P014/Japan/2001 01R137/Japan/2001
KO2/Japan/2001 J-4787/Japan/2001-2003
J-4708/Japan/2001-2003 J-4709/Japan/2001-2003
VN-11/Vietnam/2002 J-4765-Japan-2001-2003
J-4777/Japan/2001-2003 Py40699/Paraquay/1999
KY3103/Kenya/1999 01P008/Japan/2001 CMH277/Thailand/2001
TD64/Taiwan/1992 CIT-220RV/Ireland/2000
Mvd9707/Uruguay/1997 Bangla84/Bangladesh/1994
SL244/Sierra Leone/2005 356PT/South Africa/1996
87H038/Japan/1987 90Y289/Japan/1990
89A049/Japan/1989 90H377/Japan/1990
90H415/Japan/1990 88H449/Japan/1988 89H354/Japan/1989
92H085/Japan/1992 92H102/Japan/1992
JAPAN085/Japan/1993 CHIN8/China/1990
JAPAN076/Japan/1993 86Y1329/Japan/1986
86Y187/Japan/1986 Japan0022/Japan/1983
Japan38/Japan/1983 KY3303/Kenya/1999
PAK426/Pakistan/1993 TB-Chen/China/1996
S2/Japan/1981 TA81/Taiwan/1981
89Y1520/Japan/1989 90Y593/Japan/1990
TE29/Taiwan/1993 90H094/Japan/1990
90H115/Japan/1990 92H006/Japan/1992
87Y003/Japan/1987 88Y1112/Japan/1988
JAPAN137/Japan/1992 JAPAN58/Japan/1992
TWCH105/Thailand/1994 88F007/Japan/1988
85A21410/Japan/1985 86A005/Japan/1986
88Y259/Japan/1988 89H279/Japan/1988
87Y1438/Japan/1987 BE1248/Belgium/2009
GER177/Germany/2008 B110119/Japan/2011
B110005/Japan/2011 CK20047/Australia/2011
KUN/Japan/1980 TMCII/Japan/1980
83A001/Japan/1983 DS-1/USA/1976
TA3/Taiwan/1981 GR410/South Africa/1985
Hu5/Australia/1977 1831GR/South Africa/1993
92A/Australia/1992 Rj5619/Brazil/2002
H105-Palermo/Italy/1996 Nov04-H383/Russia/2004
90
99
99
94
99
85
88
92
91
88
8888
97
70
73
77
9890
84
9793
93
98
99
74
93
99
82
99
84
99
94
0.01
Lineage II
Lineage I
Lineage III
Group IV except IVa
Sub-lineage IVa
Lineage IVIVa-1
IVa-2
IVa-3
Fig. 1 A phylogenetic tree
based on the Japanese G2 VP7
nucleotide sequences
determined in this study
(indicated in boldface with
closed circular dots) along with
other Japanese G2 sequences (in
boldface) and representative
sequences obtained from the
GenBank database. The tree was
constructed using the neighbor-
joining method with bootstrap
probabilities after 1,000
replicate trials. The genetic
distance is indicated at the
bottom. Percent bootstrap
support is indicated by the value
at each node where the value is
70 % or larger. Five porcine G2
strains (accession numbers
DQ534015, AY766085,
JN974821, JX498969, and
JN974817) were used as the
outgroup, which are not shown
in the tree. Only the node where
the root is located is shown
L. P. Do et al.
123

children in Akita or Yamagata Prefecture between 1983
and 2007, and in Nara Prefecture between 2008 and 2011.
The VP7 nucleotide sequence for these 35 G2 strains were
determined essentially as described previously [4] except
that an additional primer pair G2VP7-14F (5’-AGA ATT
TCC GTC TGG CTA G-3’) and G2VP7-998R (5’-AAT
GAT CTT GAC CGT TTG GAC-3’) was used for cDNA
amplification.
We compiled the G2 VP7 nucleotide sequences that
were determined in this study with those available in the
GenBank database by using the BLAST program (http://
blast.ncbi.nlm.nih.gov/), with the DS-1 VP7 sequence as a
query sequence. We retrieved more than 1000 sequences to
include all possible G2 VP7 sequences but used only
sequences that were at least 615 nucleotides in length,
spanning positions from 307 to 921 (corresponding to
amino acid residues 87 to 291). The sequences of reas-
sortant vaccine strains were also excluded. To distinguish
the sequences of G2 strains from those of other genotypes,
a preliminary phylogenetic tree was constructed using the
MEGA 5.0 software package [18]. Porcine G2 strains are
known to have the highest sequence similarities to human
G2 strains among all rotavirus A strains previously repor-
ted [8]. With the porcine G2 strains as an indicator as the
closest outgroup, we detected a large cluster formed by G2
strains from humans in the preliminary phylogenetic tree,
enabling the removal of strains outside of this large cluster
as non-G2 strains. Thus, we obtained 509 nucleotide
sequences of human G2 strains, of which 21 sequences
were from the strains detected in Japan. The AU605
sequence was excluded because the original stool specimen
(86A005) from which AU605 was isolated in cell culture
was used in this study to determine the VP7 sequence.
Thus, a total of 543 human G2 sequences, including 35
sequences determined in this study and 508 sequences
downloaded from the GenBank database were analyzed
together with 20 Japanese G2 sequences.
While the analysis was done by constructing a phylo-
genetic tree containing all 543 human G2 sequences
detected between 1980 and 2011, for the sake of simplicity,
an abridged tree is presented in Fig. 1. All 55 Japanese G2
sequences are included in this tree, but only selected ref-
erence sequences are shown to reflect the basic topology of
the original tree. The Japanese G2 sequences clustered into
three lineages, I, III and IV, but there was no sequence
belonging to lineage II (Fig. 1). Lineage I, to which the
prototype DS-1 strain belonged, contained only one Japa-
nese strain detected in 1983 (83A001) (Figs. 1, 2). The
nucleotide sequence identities between 83A001 and other
G2 VP7 sequences in this lineage including those from
Taiwanese, South African and Australian strains were
[97.9 %. With respect to the four amino acid positions in
the VP7 antigenic regions previously described as potential
epitopes for G2 strains [4], the 83A001 strain had an amino
acid constellation of A-D-N-N at positions 87, 96, 213 and
242, respectively, which was identical to that of the pro-
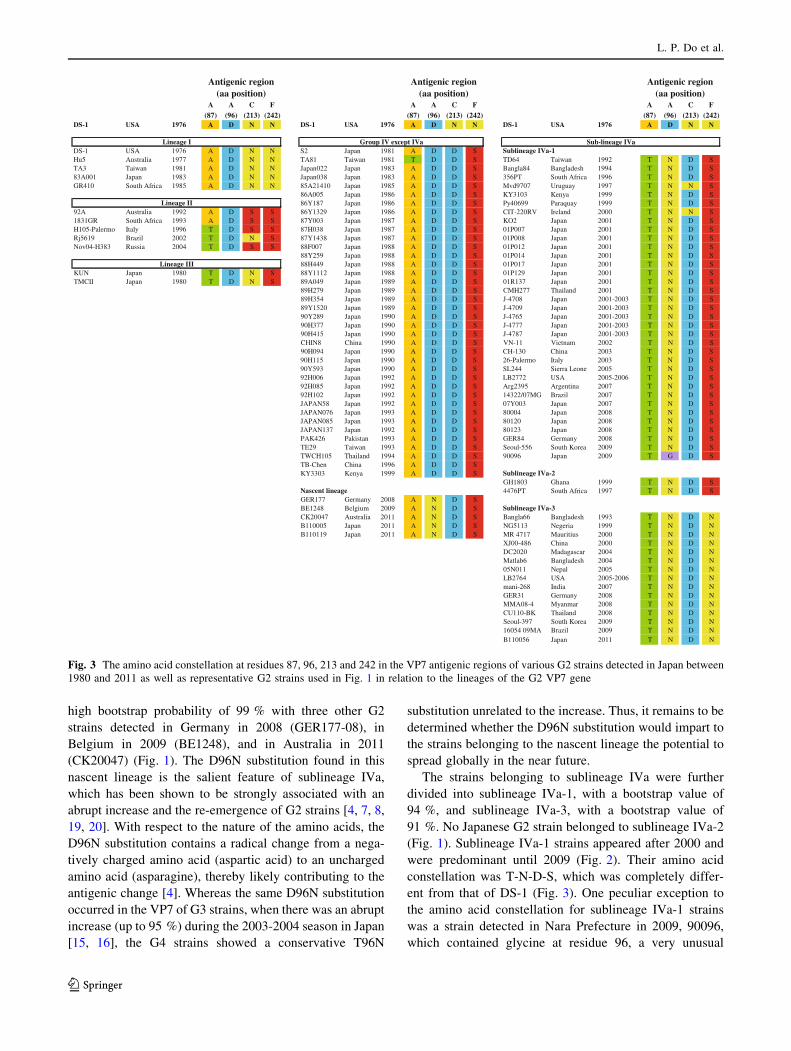
totype DS-1 strain (Fig. 3).
Lineage III contained only two Japanese strains detected
in a single year, 1980 (Fig. 2), and they were the only
strains in the world that belonged to lineage III (Fig. 1).
They had A87T and N242S substitutions from the amino
acid constellation of the lineage I strains; hence, an amino
acid constellation of T-D-N-S (Fig. 3). From the evolu-
tionary perspective, it was noted that G2 strains with lin-
eages I and III never appeared again since the early 1980s,
when they were detected for the first time in Japan (Fig. 2).
Lineage IV contained 52 Japanese strains. Within line-
age IV, however, there was sublineage IVa, with a bootstrap
value of 89 % (Fig. 1), which comprised 90 % of G2 strains
circulating after 2000 in Japan (19/21) (Fig. 2) as well as
[95 % of the global collection. The salient feature of the
strains that belong to this currently and globally predomi-
nant sublineage IVa is the presence of a D96N substitution
[4]. The remaining strains within lineage IV (33) clustered
into a group that was named ‘‘group IV except IVa’’
because of a low bootstrap value (26 % in the original
phylogenetic tree containing 543 G2 sequences). Strains
belonging to ‘‘group IV except IVa’’ had the substitutions
N213D and N242S from the amino acid constellation of the
lineage I strains; hence, an amino acid constellation of A-D-
D-S (Fig. 3). The strains carrying this constellation were
detected between 1981 and 1993 (Fig. 2). However, after an
interval of 18 years of non-detection, two strains (B110005
and B110119) belonging to this group emerged again in
2011, this time with an additional amino acid substitution,
D96N; hence, an amino acid constellation of A-N-D-S
(Figs. 2, 3). More importantly, however, these two strains
(B110005 and B110119) formed a nascent lineage with a
1980
1985
1990
1995
2000
2005
2010
2011
Lineage I
Lineage III
Group IV-except IVa
Sublineage IVa-1
Sublineage IVa-3
Fig. 2 The distribution of G2 strains in Japan by the year of detection
and the lineages to which they belonged. Each dot represents one
strain
Molecular evolution of Japanese G2 rotaviruses
123
high bootstrap probability of 99 % with three other G2
strains detected in Germany in 2008 (GER177-08), in
Belgium in 2009 (BE1248), and in Australia in 2011
(CK20047) (Fig. 1). The D96N substitution found in this
nascent lineage is the salient feature of sublineage IVa,
which has been shown to be strongly associated with an
abrupt increase and the re-emergence of G2 strains [4, 7, 8,
19, 20]. With respect to the nature of the amino acids, the
D96N substitution contains a radical change from a nega-
tively charged amino acid (aspartic acid) to an uncharged
amino acid (asparagine), thereby likely contributing to the
antigenic change [4]. Whereas the same D96N substitution
occurred in the VP7 of G3 strains, when there was an abrupt
increase (up to 95 %) during the 2003-2004 season in Japan
[15, 16], the G4 strains showed a conservative T96N
substitution unrelated to the increase. Thus, it remains to be
determined whether the D96N substitution would impart to
the strains belonging to the nascent lineage the potential to
spread globally in the near future.
The strains belonging to sublineage IVa were further
divided into sublineage IVa-1, with a bootstrap value of
94 %, and sublineage IVa-3, with a bootstrap value of
91 %. No Japanese G2 strain belonged to sublineage IVa-2
(Fig. 1). Sublineage IVa-1 strains appeared after 2000 and
were predominant until 2009 (Fig. 2). Their amino acid
constellation was T-N-D-S, which was completely differ-
ent from that of DS-1 (Fig. 3). One peculiar exception to
the amino acid constellation for sublineage IVa-1 strains
was a strain detected in Nara Prefecture in 2009, 90096,
which contained glycine at residue 96, a very unusual
A A C F A A C F A A C F(87) (96) (213) (242) (87) (96) (213) (242) (87) (96) (213) (242)
DS-1 USA 1976 A D N N DS-1 USA 1976 A D N N DS-1 USA 1976 A D N N
DS-1 USA 1976 A D N N S2 Japan 1981 A D D SHu5 Australia 1977 A D N N TA81 Taiwan 1981 T D D S TD64 Taiwan 1992 T N D STA3 Taiwan 1981 A D N N Japan022 Japan 1983 A D D S Bangla84 Bangladesh 1994 T N D S83A001 Japan 1983 A D N N Japan038 Japan 1983 A D D S 356PT South Africa 1996 T N D SGR410 South Africa 1985 A D N N 85A21410 Japan 1985 A D D S Mvd9707 Uruguay 1997 T N N S
86A005 Japan 1986 A D D S KY3103 Kenya 1999 T N D S86Y187 Japan 1986 A D D S Py40699 Paraquay 1999 T N D S
92A Australia 1992 A D S S 86Y1329 Japan 1986 A D D S CIT-220RV Ireland 2000 T N N S1831GR South Africa 1993 A D S S 87Y003 Japan 1987 A D D S KO2 Japan 2001 T N D SH105-Palermo Italy 1996 T D S S 87H038 Japan 1987 A D D S 01P007 Japan 2001 T N D SRj5619 Brazil 2002 T D N S 87Y1438 Japan 1987 A D D S 01P008 Japan 2001 T N D SNov04-H383 Russia 2004 T D S S 88F007 Japan 1988 A D D S 01P012 Japan 2001 T N D S
88Y259 Japan 1988 A D D S 01P014 Japan 2001 T N D S88H449 Japan 1988 A D D S 01P017 Japan 2001 T N D S
KUN Japan 1980 T D N S 88Y1112 Japan 1988 A D D S 01P129 Japan 2001 T N D STMCII Japan 1980 T D N S 89A049 Japan 1989 A D D S 01R137 Japan 2001 T N D S
89H279 Japan 1989 A D D S CMH277 Thailand 2001 T N D S89H354 Japan 1989 A D D S J-4708 Japan 2001-2003 T N D S89Y1520 Japan 1989 A D D S J-4709 Japan 2001-2003 T N D S90Y289 Japan 1990 A D D S J-4765 Japan 2001-2003 T N D S90H377 Japan 1990 A D D S J-4777 Japan 2001-2003 T N D S90H415 Japan 1990 A D D S J-4787 Japan 2001-2003 T N D SCHIN8 China 1990 A D D S VN-11 Vietnam 2002 T N D S90H094 Japan 1990 A D D S CH-130 China 2003 T N D S90H115 Japan 1990 A D D S 26-Palermo Italy 2003 T N D S90Y593 Japan 1990 A D D S SL244 Sierra Leone 2005 T N D S92H006 Japan 1992 A D D S LB2772 USA 2005-2006 T N D S92H085 Japan 1992 A D D S Arg2395 Argentina 2007 T N D S92H102 Japan 1992 A D D S 14322/07MG Brazil 2007 T N D SJAPAN58 Japan 1992 A D D S 07Y003 Japan 2007 T N D SJAPAN076 Japan 1993 A D D S 80004 Japan 2008 T N D SJAPAN085 Japan 1993 A D D S 80120 Japan 2008 T N D SJAPAN137 Japan 1992 A D D S 80123 Japan 2008 T N D SPAK426 Pakistan 1993 A D D S GER84 Germany 2008 T N D STE29 Taiwan 1993 A D D S Seoul-556 South Korea 2009 T N D STWCH105 Thailand 1994 A D D S 90096 Japan 2009 T G D STB-Chen China 1996 A D D SKY3303 Kenya 1999 A D D S
GH1803 Ghana 1999 T N D S4476PT South Africa 1997 T N D S
GER177 Germany 2008 A N D SBE1248 Belgium 2009 A N D SCK20047 Australia 2011 A N D S Bangla66 Bangladesh 1993 T N D NB110005 Japan 2011 A N D S NG5113 Negeria 1999 T N D NB110119 Japan 2011 A N D S MR 4717 Mauritius 2000 T N D N
XJ00-486 China 2000 T N D NDC2020 Madagascar 2004 T N D NMatlab6 Bangladesh 2004 T N D N05N011 Nepal 2005 T N D NLB2764 USA 2005-2006 T N D Nmani-268 India 2007 T N D NGER31 Germany 2008 T N D NMMA08-4 Myanmar 2008 T N D NCU110-BK Thailand 2008 T N D NSeoul-397 South Korea 2009 T N D N16054 09MA Brazil 2009 T N D NB110056 Japan 2011 T N D N
Sublineage IVa-1
Antigenic region(aa position)
Nascent lineage
Group IV except IVaLineage I
Sublineage IVa-2
Sublineage IVa-3
Antigenic region(aa position)
Antigenic region(aa position)
Lineage II
Lineage III
Sub-lineage IVa
Fig. 3 The amino acid constellation at residues 87, 96, 213 and 242 in the VP7 antigenic regions of various G2 strains detected in Japan between
1980 and 2011 as well as representative G2 strains used in Fig. 1 in relation to the lineages of the G2 VP7 gene
L. P. Do et al.
123

amino acid for a G2 strain at this position. To the best of
our knowledge, this was the only G2 strain that had glycine
at position 96 in the G2 VP7. However, glycine at position
96 is highly conserved in G1 strains except for only two
strains detected in Argentina [3].
Only one strain, B110056, detected for the first time in
Japan in 2011 belonged to sublineage IVa-3 (Figs. 1, 2).
However, this sublineage was shown to be a contemporary,
globally dominant sublineage. In Brazil, there was an
apparent shift of sublineages from IVa-1 to IVa-3 after the
introduction of a rotavirus vaccine [11]. Thus, it is specu-
lated that a shift from sublineage IVa-1 to IVa-3 will be
facilitated by the widespread use of rotavirus vaccines in
Japan.
In conclusion, this study provided the knowledge base
on the molecular evolution of Japanese G2 rotavirus strains
over a 31-year-period under unvaccinated conditions. The
results demonstrated the emergence of a nascent lineage
that had the D96N substitution outside of the currently and
globally predominant lineage. Further studies are therefore
warranted to monitor the potential of the global spread of
the emerging strains that have acquired the D96N substi-
tution, since they also appeared in Europe and Australia.
Acknowledgments We acknowledge Dr, Katsushima for providing
some of the stool specimens containing G2 rotaviruses that were used
in this study.
This study was supported in part by the Global Center of Excel-
lence Program on Integrated Global Control Strategy for Tropical and
Emerging Infectious Diseases from the Ministry of Education, Cul-
ture, Sports, Science and Technology, Japan. This study was also
supported in part by a grant-in-aid for scientific research from the
Ministry of Health, Labor, and Welfare, Japan.
References
1. Ahmed K, Ahmed S, Mitui MT, Rahman A, Kabir L, Hannan A,
Nishizono A, Nakagomi O (2010) Molecular characterization of
VP7 gene of human rotaviruses from Bangladesh. Virus Genes
40:347–356
2. Banyai K, Laszlo B, Duque J, Steele AD, Nelson EA, Gentsch
JR, Parashar UD (2012) Systematic review of regional and
temporal trends in global rotavirus strain diversity in the pre
rotavirus vaccine era: insights for understanding the impact of
rotavirus vaccination programs. Vaccine 30(Suppl 1):A122–
A130
3. Barril P, Martınez L, Giordano M, Masachessi G, Isa M, Pavan J,
Glikmann G, Nates S (2013) Genetic and antigenic evolution
profiles of G1 rotaviruses in Cordoba, Argentina, during a
27-year period (1980-2006). J Med Virol 85:363–369
4. Doan YH, Nakagomi T, Cunliffe NA, Pandey BD, Sherchand JB,
Nakagomi O (2011) The occurrence of amino acid substitutions
D96N and S242N in VP7 of emergent G2P[4] rotaviruses in
Nepal in 2004–2005: a global and evolutionary perspective. Arch
Virol 156:1969–1978
5. Esteban LE, Rota RP, Gentsch JR, Jiang B, Esona M, Glass RI,
Glikmann G, Castello AA (2010) Molecular epidemiology of
group A rotavirus in Buenos Aires, Argentina 2004-2007:
reemergence of G2P[4] and emergence of G9P[8] strains. J Med
Virol 82:1083–1093
6. Gurgel RQ, Cuevas LE, Vieira SC, Barros VC, Fontes PB, Sal-
ustino EF, Nakagomi O, Nakagomi T, Dove W, Cunliffe N, Hart
CA (2007) Predominance of rotavirus P[4]G2 in a vaccinated
population, Brazil. Emerg Infect Dis 13:1571–1573
7. Iturriza-Gomara M, Cubitt D, Desselberger U, Gray J (2001)
Amino acid substitution within the VP7 protein of G2 rotavirus
strains associated with failure to serotype. J Clin Microbiol
39:3796–3798
8. Khamrin P, Peerakome S, Tonusin S, Malasao R, Okitsu S,
Mizuguchi M, Ushijima H, Maneekarn N (2007) Changing pat-
tern of rotavirus G genotype distribution in Chiang Mai, Thailand
from 2002 to 2004: decline of G9 and reemergence of G1 and G2.
J Med Virol 79:1775–1782
9. Kirkwood CD, Boniface K, Barnes GL, Bishop RF (2011) Dis-
tribution of rotavirus genotypes after introduction of rotavirus
vaccines, Rotarix(R) and RotaTeq(R), into the National Immu-
nization Program of Australia. Pediatr Infect Dis J 30(Sup-
pl):S48–S53
10. Martinez M, Amarilla AA, Galeano ME, Aquino VH, Farina N,
Russomando G, Parra GI (2010) Predominance of rotavirus
G2P[4] and emergence of G12P[9] strains in Asuncion, Paraguay,
2006-2007. Arch Virol 155:525–533
11. Mascarenhas JD, Lima CS, de Oliveira DS, Guerra Sde F,
Maestri RP, Gabbay YB, de Lima IC, de Menezes EM, Linhares
Ada C, Bensabath G (2010) Identification of two sublineages of
genotype G2 rotavirus among diarrheic children in Parauapebas,
Southern Para State, Brazil. J Med Virol 82(4):712–719
12. Nakagomi T, Cuevas LE, Gurgel RG, Elrokhsi SH, Belkhir YA,
Abugalia M, Dove W, Montenegro FM, Correia JB, Nakagomi O,
Cunliffe NA, Hart CA (2008) Apparent extinction of non-G2
rotavirus strains from circulation in Recife, Brazil, after the
introduction of rotavirus vaccine. Arch Virol 153:591–593
13. Page NA, Steele AD (2004) Antigenic and genetic characteriza-
tion of serotype G2 human rotavirus strains from South Africa
from 1984 to 1998. J Med Virol. 72:320–327
14. Patel MM, de Oliveira LH, Bispo AM, Gentsch J, Parashar UD
(2008) Rotavirus P[4]G2 in a vaccinated population, Brazil.
Emerg Infect Dis 14:863–865
15. Phan TG, Khamrin P, Quang TD, Dey SK, Takanashi S, Okitsu S,
Maneekarn N, Ushijima H (2007) Detection and genetic char-
acterization of group A rotavirus strains circulating among chil-
dren with acute gastroenteritis in Japan. J Virol 81:4645–4653
16. Phan TG, Trinh QD, Khamrin P, Kaneshi K, Ueda Y, Nakaya S,
Nishimura S, Sugita K, Nishimura T, Yamamoto A, Takanashi S,
Yagyu F, Okitsu S, Ushijima H (2007) Emergence of new variant
rotavirus G3 among infants and children with acute gastroenter-
itis in Japan during 2003-2004. Clin Lab 53:41–48
17. Snelling TL, Andrews RM, Kirkwood CD, Culvenor S, Carapetis
JR (2011) Case-control evaluation of the effectiveness of the
G1P[8] human rotavirus vaccine during an outbreak of rotavirus
G2P[4] infection in central Australia. Clin Infect Dis 15:191–199
18. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S
(2011) MEGA5: molecular evolutionary genetics analysis using
maximum likelihood, evolutionary distance, and maximum par-
simony methods. Mol Biol Evol 28:2731–2739
19. Trinh QD, Pham NT, Nguyen TA, Phan TG, Yan H, le Hoang P,
Khamrin P, Maneekarn N, Li Y, Okitsu S, Mizuguchi M,
Ushijima H (2010) Sequence analysis of the VP7 gene of human
rotaviruses G2 and G4 isolated in Japan, China, Thailand, and
Vietnam during 2001-2003. J Med Virol 82:878–885
20. Zao CL, Yu WN, Kao CL, Taniguchi K, Lee CY, Lee CN (1999)
Sequence analysis of VP1 and VP7 genes suggests occurrence of
a reassortant of G2 rotavirus responsible for an epidemic of
gastroenteritis. J Gen Virol 80:1407–1415
Molecular evolution of Japanese G2 rotaviruses
123








![Emergence of Unusual G6P[6] Rotaviruses in Children ...Emergence of Unusual G6P[6] Rotaviruses in Children, Burkina Faso, 2009–2010 Technical Appendix Technical Appendix Table 1](https://img.pdfslide.us/doc/110x75/5f0ac8c87e708231d42d537a/emergence-of-unusual-g6p6-rotaviruses-in-children-emergence-of-unusual-g6p6.jpg)




![ResearchConcentration of acrylamide in a polyacrylamide ...a PAGE gel affected the relative position of the VP7 gene of G2 rotavirus strains [24], we analyzed the effects of acrylamide](https://img.pdfslide.us/doc/110x75/5e4369d50d65d67dee398521/researchconcentration-of-acrylamide-in-a-polyacrylamide-a-page-gel-affected.jpg)