Embed Size (px)
Citation preview

Molecular Diversity Analysis of Whitefly(Bemisia tabaci) Collected from Different
Regions of Madhya Pradesh
THESIS
Submitted to the
Jawaharlal Nehru Krishi Vishwa Vidyalaya, Jabalpur
In partial fulfillment of the requirement forthe Degree of
MASTER OF SCIENCE
In
AGRICULTURE(MOLECULAR BIOLOGY AND BIOTECHNOLOGY)
By
SUMIT PRAKASHRAO KALEDivision of Plant Molecular Biology
Biotechnology CentreJawaharlal Nehru Krishi Vishwa Vidyalaya, Jabalpur
Jabalpur (MP)
2013

CERTIFICATE- I
This is to certify that the thesis entitled, “Molecular Diversity Analysisof Whitefly (Bemisia tabaci) Collected from Different Regions of MadhyaPradesh” submitted in partial fulfillment of the requirement for the degree of
MASTER OF SCIENCE in Agriculture (Molecular Biology andBiotechnology) of Jawaharlal Nehru Krishi Vishwa Vidyalaya, Jabalpur is a
record of the bonafide research work carried out by Mr. SUMITPRAKASHRAO KALE under my guidance and supervision. The subject of
the thesis has been approved by the Student’s Advisory Committee and the
Director of Instruction.
No part of the thesis has been submitted for any other degree or
diploma (Certificate awarded etc.) or has been published / published part has
been fully acknowledged. All the assistance and help received during the
course of the investigation has been acknowledged by him.
(Dr. S. Tiwari)Chairman of the Advisory Committee
THESIS APPROVED BY THE STUDENT’S ADVISORY COMMITTEE
Chairman (Dr. S. Tiwari) ……………………………………………..
Member (Dr. L. P. S. Rajput) ……………………………………………..
Member (Dr. U. K. Khare) …. ……………………………………….


CERTIFICATE - II
This is to certify that the thesis entitled, “Molecular Diversity Analysisof Whitefly (Bemisia tabaci) Collected from Different Regions of MadhyaPradesh” submitted by Mr. SUMIT PRAKASHRAO KALE to the Jawaharlal
Nehru Krishi Vishwa Vidyalaya, Jabalpur, in partial fulfillment of the
requirement for the degree of MASTER OF SCIENCE in AGRICULTURE((Molecular Biology and Biotechnology) JNKVV, Jabalpur, after evaluation
has been approved by the External Examiner and Student’s Advisory
Committee after an oral examination on the same.
Place : Jabalpur (Dr. S. Tiwari)Date: ………… Chairman of the Advisory Committee
MEMBER OF THE STUDENT’S ADVISORY COMMITTEE
Chairman : Dr. S. Tiwari .…………………
Member : Dr. L. P. S. Rajput .…………………
Member : Dr. U. K. Khare .…………………
Director, Biotechnology Centre : Dr. S. Tiwari .…………………
Director of Instructions : Dr. P. K. Mishra .…………………

i
List of Contents
ChapterNo.
Title Page
I Introduction 1 – 4
II Review of Literature 5 – 20
III Materials and Methods 21 – 30
IV Results 31 – 36
V Discussion 37 – 41
VI Summery, Conclusions and Suggestionsfor further work 42 – 43
6.1 Summary 42
6.2 Conclusions 43
6.3 Suggestions for further work 43
References 44 – 53
Vita

ii
List of Tables
TableNo. Title Page
3.1.1 Bemisia tabaci samples used for present studywith their location of collection.
21
3.2.1 Composition of DNA extraction solution. 24
3.3.1a Mitochondrial Cytochrome Oxidase I (mtCOI)primers and their sequences used in the study.
28
3.3.1b List of components with their concentrations usedfor mtCOI-PCR.
28
3.3.1c Temperature profile used in PCR Amplification forCI-J-2195 and L2-N-3014.
29
3.4 List of components with their concentrationsused for RAPD-PCR.
29
3.5.1 Temperature profile used in PCR Amplification forRAPD.
30
3.5.2 RAPD primers and their sequences used in thestudy.
30
4.1 Populations of whitefly, Bemisia tabaci collectedfrom various host plants in different geographicalregions of East Madhya Pradesh.
32
4.1.2 Efficiency of the whitefly DNA extraction method. 33
4.1.3 Spectrophotometric data of whitefly DNA extractedthrough standardized protocol.
34
4.5 RAPD primers analysis. 36
iii
List of Figures
SI. Title Page(between pages)
3.1.1a Map of B. tabaci collection site in MadhyaPradesh.
22
3.1.1b Images of Bemisia tabaci. 22
4.1 DNA isolation of whitefly population by CTAB method. 36 – 37
4.2 DNA isolation of whitefly population by Nonidet NP-40method.
36 – 37
4.3 DNA isolated from 17 populations of whitefly (B.tabaci) collected from different geographical regions ofMadhya Pradesh.
36 – 37
4.4 mtDNA analysis generated among the 17 samples ofwhitefly (B. tabaci) (Lane 1-17) using primer
36 – 37
4.5 RAPD analysis generated among the 17 samples ofwhitefly (B. tabaci) (Lane 1-17) using primer OPF-2
36 – 37
4.6 RAPD analysis generated among the 17 samples ofwhitefly (B. tabaci) (Lane 1-17) using primer OPH-9
36 – 37
4.7 RAPD analysis generated among the 17 samples ofwhitefly (B. tabaci) (Lane 1-17) using primer OPH-11
36 – 37
4.8 RAPD analysis generated among the 17 samples ofwhitefly (B. tabaci) (Lane 1-17) using primer OPH-16
36 – 37

iv
ABBREVIATIONS
% : Per centl : Micro literM : Micro molarAFLP : Amplified fragment length polymorphismBp : Base pairCTAB : Cetyl trimethyl ammonium bromideDNA : Deoxyribose nucleic acidEDTA : Ethylene diamine tetra-acetate
ISSR : Inter simple sequence repeatITSI : Internally transcribed spacer 1 sequenceFW : ForwardM : MolarmA : Mili ampereMbp : Million base pairMg : Mili gramMl : Mili litermM : Mili molarmt COI : Mitochondrial cytochrome oxidase IMin : MinuteNg : Neno gramNaOH : Sodium hydroxideNaCl : Sodium chlorideNonidet NP-40 : 4-Nonylphenyl-polyethylene glycolHCL : Hydrochloric acidO.D. : Optical densityPAGE : Polyacrylamide gel electrophoresisPCR : Polymerase chain reactionPVP : Poly-vinyl poly propilidinepH : Power of hydrogenpg : PicogramRAPD : Random amplified polymorphic DNARFLP : Restriction fragment length polymorphismRW : ReverseRpm : Revolution per minuteSCAR : Sequence characterized amplified regionSSR : Simple sequences repeatSDS : Sodium-dodocel sulphateKb : Killo base pairTAE : Tris base acetic acid glacial EDTATris : Trisma baseTE : TrisUV : Ultra violet

v
v/v : Volume by volumew/v : Weight by volume

1
INTRODUCTION
The whitefly Bemisia tabaci (Genn.) (Hemiptera: Aleyrodidae) is
one of the most important crop pest in the world and it serves as for
vector of more than 100 plant viruses (Jones, 2003). Most of these
viruses belong to the genus Begomovirus, one of the most important
emerging groups of plant viruses (Brown, 2000). B. tabaci has a high
intraspecific biological and genetic variability (De Barro et al., 2000).
On the basis of morphological characters it is very difficult to
distinguish different juvenile and adult B. tabaci (Rosell et al., 1997;
Calvert et al., 2001).
B. tabaci comprises biotypes which are difficult to differentiate
through conventional taxonomy due to morphological similarity (Brown
et al., 1995), whereas such biotypes of whitefly can be distinguished at
molecular level because of finding more genetic complexity in
whiteflies (Boykin et al., 2007). The genetics complexity within B.
tabaci was first recognized in the 1950’s when morphologically
indistinguishable populations were reported to differ in host range,
host-plant adaptability, and plant virus-transmission capabilities (Bird
and Maramorosch, 1978). B. tabaci was composed of a series of
biotypes (Costa and Brown, 1991; Bedford et al., 1994) that are
complex of highly cryptic sibling species, which lack morphological
traits for differention of behavioral and or genetic variants which proved
by biochemical and molecular analysis on B. tabaci collected from
different geographical regions of the world.
Whitefly (Bemisia tabaci) is a noxious insect which attack several
field and greenhouse cultivated crops in India. It belongs to a group of
insects named whiteflies that is commonly known as different names
based on crops on which it attack e.g. tobacco, cotton or sweet potato
whitefly (Rekha et al., 2005). It (Gennadius) (Hemiptera: Aleyrodidae)
is one of the most devasting tropical and sub-tropical agricultural pests
(Byrne and Bellows, 1991) affecting the yield of a broad range of

2
agricultural, fiber, vegetable and ornamental crops (Cahill et al., 1996)
and is considered one of the world’s top invasive species (Boykin et
al., 2007). B. tabaci became an important globally as a serious pest
because of the polyphagous nature of some biotype and the diverse
ways that it damage crops (Rekha et al., 2005). Both nymphs and
adults feed on plant sap and cause yellowing, molting and leaf fall. The
excretion of a substance known as honeydew facilitates fungus (sooty
mold) to colonize the leaf surface, which may affect the development of
the plant (Byrne and Bellows, 1991). Dinsdale et al., (2010) have
recently proposed that B. tabaci should be considered as a cryptic
species complex comprising 11 groups containing 24 species. The
more reproducible and informative method available to determine the
genetic affiliation of a B. tabaci population is sequencing the
mitochondrial cytochrome oxidase I (mtCOI) gene (De Barro et al.,
2005) and use consensus sequences to assign group affiliation
(Dinsdale et al., 2010). The Middle East-Asia Minor 1 group contains
the B and B2 biotypes and the Mediterranean the Q, J, L, Sub Saharan
Africa Silver leaf biotypes (Dinsdale et al., 2010). The biotype B was
introduced in Brazil in the beginning of the 1990’s (Lourencao and
Nagai, 1994) and in the last few years has spread to several states
causing losses in many crops (Lima et al., 2002).
Accurate identification of insects that are pests and or virus
vectors is a prerequisite for their effective management to reduce crop
damage (Brown, 2000). Analysis of the mitochondrial cytochrome
oxidase I (mtCOI) gene of worldwide collections of the whitefly have
greatly improved the understanding of the genetic diversity of the B.
tabaci species complex (Bosco et al., 2006; Dinsdale et al., 2010). On
the basis of microsatellite, mitochondrial and ribosomal markers
analysis, molecular phylogeny of B. tabaci specimens has identified a
large number of races and genetic groups that has established in
different parts of the world with each one represented by a large
number of genetic population and sub populations with little or no gene
flow between them. In a specific geographic region, native whiteflies

3
remain active on many cultivated plant as hosts and on weeds as
collateral hosts throughout the year. These host plant serving as
alternate hosts to whiteflies are known to exert a strong influence on
selection of specific whitefly genotypes out of the existing genotypes in
a particular agro-climatic region (Gupta et al., 2010).
The analysis of genetic variation using DNA fingerprinting
techniques has become an important approach in taxonomic,
population genetic and evolutionary studies of a variety of insect
species. Ambiguity in the identification of B. tabaci biotypes can be
eliminated using molecular markers. A major advantage of employing
DNA markers is that they are not influenced by environmental effects
or epistatic interactions. Molecular markers detect variation at the level
of the DNA sequence and have proved to be an extremely effective
tool for distinguishing between closely related genotypes.
Molecular markers have become available for identifying the
different genetic groups of B. tabaci that are mitochondrial COI or
ribosomal ITS1 DNA sequences. Molecular based assays have been
used to distinguish various populations of B. tabaci. Among the
different molecular markers, RAPD-PCR (Lima et al., 2000; Lima et al.,
2002; Fontes et al., 2010), Mitochondrial Cytochrome Oxidase I
(mtCOI), Microsatellite markers, Internally transcribed spacer 1
sequences (ITS1) (Boykin et al., 2007) and AFLP markers (Cervera et
al., 2000) have been used to analyze genetic diversity and amplify
various specific fragments linked to genes in the whitefly genome
(Boykin et al., 2007). Molecular technology has provided valuable
molecular marker investigational tools. The genetic differentiation of B.
tabaci populations may lead to important implications for the control of
this pest in India and worldwide, since it includes populations of
different host plants, from different geographic regions, and genetic
variations within these populations (Valle et al., 2011). For most
molecular studies, the isolation of high quality DNA is an essential
prerequisite. Isolation of DNA is difficult when insects are too small to
dissect them. The extraction and purification of intact DNA is often

4
extremely difficult due to two major problems: the minute size of the
individuals and the low amount of total DNA in insects (Redi and
Garagna 1987; Bertolani et al., 1994; Garagna et al., 1996). Additional
problems are encountered with the step of preservation, the handling
of specimens in the field or laboratory, the choice of the extraction
protocol or commercial kit, or the polymerase enzymes and the
sequencing reactions. Additionally, some DNA isolation methods and
commercially available kits tend to produce low DNA yields with short
storage life (Zidani et al., 2005), which make them unsuitable for some
molecular applications. Due to these facts, standardization of DNA
extraction protocol from whitefly for molecular analysis is needed.
This research is based on the identification of species in the
genus Bemisia, but the taxonomy of whiteflies has long been
problematic because of similarities in the morphology of pupae and
adults. The aim of this study was to characterize B. tabaci populations
collected on host plants from different geographical regions of East
Madhya Pradesh.
In the light of above facts, the present experiments were
envisaged to fulfill following objectives:
• To standardize protocol for DNA isolation from whitefly.
• To analyze the genetic diversity of whitefly using DNA fingerprinting.

5
REVIEW OF LITERATURE
Whiteflies are small, often inconspicuous insects that are
globally distributed as agricultural pests of both greenhouse and field
crops. Although > 1,500 species of whiteflies exist, only a few cause
serious economic losses (Bink-Moenen, 1990; Martin, 2004). Their
common name, whitefly, is due to the presence of white wax and lipid
particles that cover the body and wings of most adult species (Byrne
and Hadley, 1988; Buckner et al., 1994). Whiteflies are polyphagous
herbivores that reduce crop yields by extracting water, carbohydrates
and amino acids from plant phloem (Lioyd, 1922). Whitefly can
transmit plant viruses. However, serious viral diseases are more
commonly associated with B. tabaci (Cohen, 1990). As phloem-feeding
insects, whiteflies excrete sticky honeydew that can cover fruit and
foliage of crops. Honeydew fosters the growth of sooty mold
(Cladosporium) on plants and reduces plant photosynthesis (Lioyd,
1922; Hoddle et al., 1998; Smith et al., 2001)
2.1 Host Plant and Natural Enemies
B. tabaci has been recorded from more than 600 different plant
species (Mound and Halsey, 1978; Greathead, 1986; Secker et al.,
1998) and its polyphagous nature has been document worldwide (Bird,
1957; Costa and Russell, 1975; Bird and Maramorosch, 1978; Butler et
al., 1986, Costa and Brown, 1990, 1991; Costa et al., 1991; Burban et
al., 1992). B. tabaci also is a major pest of greenhouse crops,
particularly ornamentals, although major crops under greenhouse
production such as tomato, pepper, beans eggplant, and cucumber are
also attacked (Cock, 1986).
Since whiteflies are minute insects, many of their predators in
particular the more prey specific ones are small arthropods. The
location of whiteflies, especially the immature stages that are fixed
upon plant leaves also limits the kind of predators to those that occur
on foliage. The natural enemies of whitefly are Encarsia sp., Chrysopa

6
basalis Walker, Chrysopa sp., Lycoza sp., Oxypes sp., and
Coccinellidae (Kheinmeesook, 1997). In addition to parasitoids and
predators, pathogens are also natural enemies of whiteflies. Several of
them were found to cause fatal disease to whitefly especially the
entomophathogenic fungi. It is well known that fungi invade insect via
external cuticle and not be ingested to initiate disease. This makes
them prime candidates for use against plant sucking insects including
whitefly. The fungi, Aschersonia aleyrodis and Paecilomyces
fumosoroseus were commercially produced and used effectively for
controlling whitefly (Lacey et al., 1996). In greenhouse, the fungi,
Verticillium lecanii and A. aleyrodis were used to control whitefly and
aphids (Dowell, 1990). It is believed that there are still numerous other
species of fungi that can serve as potential biological agents for
controlling whitefly as well as for other plant sucking insect species.
2.2 Whiteflies as Agricultural Pests
Whiteflies are major pests of many agricultural crops worldwide.
They belong to the family Aleyrodidae of the order Hemiptera. More
than 1450 species have been described as belonging to two
subfamilies and the most economically important pest species are
members of the Aleyrodinae (Bink-Moenen and Mound, 1990; Martin
et al., 2000).
Whiteflies that are commonly known as different names based
on crops on which it attacks e.g., cotton, tobacco or sweet-potato
whitefly, is a diverse species found outdoors in tropical and sub-
tropical regions. In temperate countries it can be found only in crops
grown under protection (Martin et al., 2000). It affects hundreds of
different plant species (Byrne et al., 1990; Martin et al., 2000) and
transmitting more than 110 plant viruses, belonging to the
Begomovirus, Crinivirus and Carlavirus genera (Jones, 2003). Most of
those viruses can cause severe damage to crops and so even very low
populations of this pest can result in major crop failures. Several
biotypes of B. tabaci have been identified; the two currently of most

7
importance to agricultural environments are termed biotypes B and Q
(Perring, 2001).
2.3 Molecular Markers for the identification of species/biotype
Markers are characters whose inheritance pattern can be
followed at morphological (e.g. flower colour), biochemical (proteins
and isozymes) and DNA levels (molecular markers). These characters
are called markers as they are used to obtain indirect information
about the genetics of other traits of interest in the organism under
investigation (Jena, 2000). Variation in DNA sequences have been
extensively explored as genetic markers for genome mapping. DNA
based markers have several advantages over morphological and
biochemical markers such as pleiotropic or epistatic effects, enabling
the use of non-destructive method and use less amount of tissue,
independent to environmental stresses or management practices and
provide easy access, high reproducibility and high genetic resolution.
Perhaps the most widespread application of DNA markers is in the
construction of genetic maps, which can be used to determine the
chromosomal location of genes affecting either simple or complex traits
(Lander and Botstein, 1989; Dudley et al., 1992; Lee et al., 1996).
Genetic markers were being used in biology, well before it was
known that DNA was the hereditary material. Visible markers,
mutations in genes with visible consequences such as dwarfism or
colours, have been used in genetic studies since early in the twentieth
century (Morgan, 1911). In recent years the burgeoning field of
molecular biology has provided tools suitable for rapid and detailed
genetic analysis of higher organisms, including agricultural species.
Information from DNA markers serves many diverse purposes, such as
forensic science, paternity testing, identifying genes and inferring
evolutionary relationships among organisms (Paterson et al., 1993).
The advent of molecular markers revolutionized the entire
scenario of biological sciences. DNA-based molecular markers have
acted as versatile tools and have found their own position in various

8
fields like taxonomy, physiology, embryology, genetic engineering, etc.
(Joshi et al., 1999). They are no longer looked upon as simple DNA
fingerprinting markers in variability studies or as forensic tools. Since
their development, they are constantly being modified to enhance their
utility and to bring about automation in the process of genome
analysis. Random Amplified Polymorphic DNA (RAPD) markers are
one of the most commonly used markers as they offer easy handling
and produce quick results. RAPD markers have been extensively used
in many crops as genetic markers for assessment of genetic variability
and have proved highly efficient for assessment of genetic diversity.
These markers are mostly dominant and detect variations in both
coding as well as non-coding regions of the genome.
About 1,200 whitefly species have been described (Mound and
Halsey, 1978), although the real number in existence is probably much
higher. Most whitefly species are insect pests in the tropic. Variation in
host plant preference, life cycle and even disease transmission
capability can be expected between populations in different regions
and habitats. Whiteflies were commonly described only from their
pupal case (Bink-Moenen, 1983; Martin, 1985). At the same level
distinction such as host race is biotype, a term used to designate
populations that lack morphological distinction, but that possess other
characteristics which serve to separate them from other populations
(Claridge et al., 1997).
Recently, molecular based assays have been used to distinguish
various populations of B. tabaci. A popular technique has been the use
of Mitochondrial Cytochrome Oxidase I (mtCOI), Microsatellite
markers, internally transcribed spacer 1 sequences (ITS1) and AFLP
markers to amplify various fragments in the whitefly genome. These
fragments are easily seen as variable sized bands using standard
agarose gel electrophoresis.
Campbell (1993) and Campbell et al. (1994, 1996) performed
sequence analyses of the 18S rRNA genes from biotype A and B and

9
showed 2bp substitutions out of the 1039bp fragments analysed. Other
genes have been used as well for proper identification.
Frohlich et al. (1999) performed Phylogenetic analyses on 10 B.
tabaci populations from various locations using 550bp region of the
mitochondrial 16S rDNA and 700bp region of the mitochondrial
cytochrome oxidase I (COI) gene. They reported that the 16S
fragments were useful in separating New World and Old World whitefly
biotypes, and they placed esterase biotype B within Old World Israel-
Yemen clade. Analysis of CO1 sequences supported 5 distinct clades;
Benin (esterase biotype B), Sudan (esterase biotype L), India
(esterase biotype G), Israel-Yemen-USA (esterase biotype B), and
Costa Rica-Mexico-Puerto Rico-USA (esterase biotype A).
Cervera et al. (2000) used AFLP markers for showing the
genetic similarities between B. tabaci with B. medinae Gomez-Memor
and another of B. afer Priesner and Hosny and analysis showed that B.
tabaci a minimum similarity coefficient of 0.32 and separated from the
two other species with a similarity coefficient of 0.07.
Li et al. (2005) used internally transcribed spacer 1 sequences
(ITS1) of ribosomal DNA of B biotype and other biotypes which were
collected from different location in China, were sequenced and
analysed. The B biotype-specific primers were then designed for rapid
identification of B biotype of B. tabaci and result showed that the
diagnostic primer only gave a positive result with B biotype
Delatte et al. (2005) showed two genetic types of B. tabaci were
distinguished using RAPD-PCR and Cytochrome Oxidase I (COI) gene
sequence comparison. One type was assigned to B biotype and other
was genetically dissimilar to the population named Ms after the
Mascarenes Archipelago. Both B and Ms Population of B. tabaci
induced silverleaf symptoms on cucurbita spp, and were able to
acquire and transmit TYLCV. Taken together these results indicate that
Ms Genetics type should be considering a new biotype of B. tabaci.

10
Bosco et al. (2006) used PCR-RFLP identification of B. tabaci
biotype in the Mediterranean Basin. Developed a method for rapid
biotyping of B. tabaci populations for five biotypes such as B, Q, M, S
and T reported until now in Mediterranean have been tested by PCR
amplification of the cytochrome oxidase I mitochondrial gene followed
by restriction with enzyme True91.
Ueda and Brown (2006) showed first report of the Q biotype of
B. tabaci in Japan by mitochondrial cytochrome oxidase I sequence
analysis. The molecular genetics identification and phylogenetic
relationships of 12 B. tabaci populations collected from representative
location in Japan. Phylogenetic analysis of the whitefly mtCOI
sequence indicated that both the invasive B and Q biotype was found
at four locations Mihara in Hiroshima, Nishigoshi in Kumamoto,
Miyanojo and Okuchi in Kagoshima, the remaining eight collections
were identified as B biotype. This is the first report of the introduction of
Q biotype in Japan.
Boykin et al. (2007) used a Bayesian phylogenetic technique for
understanding of global B. tabaci phylogenetic relationships to
elucidate the relations among all COI DNA sequence data available in
Genbank for B. tabaci worldwide and analyze 12 major well-resolved
genetic groups showing a close relationship with Asian biotype.
Hsieh et al. (2007) used mitochondrial cytochrome oxidase I
(mtCOI) gene was to reconstruct a phylogenetic tree for identification
of biotype B and Q and to study the relationships between invasive
events and ornamental plants.
Scott et al. (2007) showed first record of B. tabaci biotype Q in
New Zealand. The whitefly was identified morphologically as B. tabaci
and capsicum represents a new host. Biotypes B and Q are
morphologically indistinguishable. Molecular diagnosis (Cytochrome
oxidase subunit I) of samples collected in a survey in 2006 from
capsicums and poinsettias revealed the presence of B. tabaci biotype
Q in New Zealand.

11
Tsagkarakoua (2007) showed biotype status and genetic
polymorphism of the whitefly (B. tabaci) (Hemiptera: Aleyrodidae) in
Greece by using sequencing of the mitochondrial cytochrome oxidase I
(mtCOI) gene and genotyping using microsatellite markers. Analyses
of the mtCOI sequences revealed a high homogeneity between the
Greek samples which clustered together with Q biotype samples that
had been collected from other countries. When genetic polymorphism
was examined using six microsatellite markers, the Greek samples,
which were all characterized as Q biotype were significantly
differentiated from each other and clustered into at least two distinct
genetic populations.
Bel-Kadhi et al. (2008) used Bem-23 microsatellite marker for
molecular characterization of B. tabaci Biotypes in Southern Tunisia.
The samples were collected from 8 geothermal areas and one from
Douz Oasis. Which showed that presence of biotype “B” 7 of the 8
geothermal sites having 220 base pair band for this biotypes while the
geothermal sites of B. tabaci population is present as a mixture of
biotype “B” and “Q” as revealed by the 410 base pair bands.
Rabello (2008) diversity analysis of B. tabaci biotypes by using
RAPD, PCR-RFLP and sequencing of the ITS1 rDNA region. He
analysed B. tabaci biotypes B, BR, Q and Cassava, which showed a
high similarity between the individuals of the B and Q biotypes, which
could be distinguished from the BR individuals. A phylogenetic tree
based on ITS1 rDNA sequence was constructed. This is the first report
of the ITS1 rDNA sequence of B. tuberculata and of the BR biotype of
B. tabaci.
Bethke et al. (2009) showed first record of the Q biotype of the
sweet potato whitefly (B. tabaci) in Guatemala, using esterase isozyme
patterns and mtCOI sequencing. Adult whitefly and immature were
collected from commercially grown poinsettia plants and only Q biotype
was found on commercially grown poinsettia plants. This is the first
reports of the Q biotype in Guatemala.

12
Fujiie et al. (2009) used mitochondrial cytochrome oxidase I
(mtCOI) sequence for identification of the whitefly biotype collected
from different geographical location of autumn-cultured potato fields in
Syria (Japan) and confirmed that they were the Q, B and non-B (B2)
biotype of B. tabaci.
Mckenzie (2009) showed distribution of B. tabaci (Hemiptera:
Aleyrodidae) biotypes in Florida-Investigating the Q Invasion by using
mitochondrial cytochrome oxidase I subunit and microsatellite markers.
The biotype status of submitted B. tabaci samples was determined by
polymerase chain reaction (PCR) amplification and sequencing of a
700-800bp small subunit (mtCOI) gene fragment, PCR amplification,
and size determination of two unique microsatellite markers and
esterase zymogram analysis. 180 collections were sampled from 23
counties. Of these samples, 58% were from vegetables, 37% were
from ornamentals, and 5% were from peanuts, alfalfa, and weeds. 18%
of all collections were found to be the Q biotype that came from
greenhouse grown ornamental and herbs located in six counties.
Sequence comparison of the mtCOI gene identified three separate
haplotypes within Florida that were defined as Q1, Q2, and Q3.
Shatters (2009) showed improved DNA barcoding method for B.
tabaci and related Aleyrodidae: development of universal and B. tabaci
biotype-specific mitochondrial cytochrome oxidase I polymerase chain
reaction primers. These primers produce a 478bp, 405bp, and 303bp
mtCOI fragment for the B, New World, and Q biotypes, respectively in
their study.
Chu et al. (2010) applied mitochondrial cytochrome oxidase I
(mtCOI) specific marker for phylogenetic analysis among whitefly from
china and grouped into different cluster.
Fontes et al., (2010) used microsatellite marker for diversity
analysis among whitefly isolate collected from different regions of
Brazil and found geographical grouping among them.

13
Chu et al. (2011) showed investigation of the genetic diversity of
an invasive whitefly (Bemisia tabaci) in China using both mitochondrial
and nuclear DNA markers. He explains the changes in genetic
variation between its home range in the Mediterranean region and its
invasion range we show that Q in Shandong likely originated in the
western Mediterranean. His studied also found that the haplotype
diversity was low compared with its presumed geographic origin,
whereas microsatellite allele diversity showed no such decline.
Ma et al. (2011) used microsatellite markers for the identification
of the spiralling whitefly (Aleurodicus dispersus) species examined in
individual whitefly from Hainnan Island and Canary Islands (China) and
observed expected heterozygosity values were 0.773 and 0.585,
respectively. Conclude that microsatellite markers provide powerful
tools for ecological, epidemiological and population genetics studies on
this highly invasive insect.
Rocha et al. (2011) found mitochondrial cytochrome oxidize I
(mtCOI) marker for the identification of B-Biotype of B. tabaci is
presents on vegetable in Sao Paul State, Brazil.
Valley et al. (2011) used COI gene marker for the mtDNA
variability in whitefly populations in Brazil. He collected three individual
samples from host plant species, such as soybean, pumpkin and
tomato. He found that all haplotypes belong to biotype B, which is
confirmed by the haplotype network as well as genetic structure
analysis, showed that the host plant species do not influence
population structuring of this pest, only the geographical location
mattered.
Mugerw et al. (2012) showed genetic diversity and geographic
distribution of B. tabaci (Gennadius) (Hemiptera: Aleyrodidae)
genotypes associated with cassava in East Africa by using partial
sequences of the mitochondria cytochrome oxidase I (mtCOI) DNA, in
cassava growing areas of Kenya, Tanzania and Uganda. Two distinct
species were obtained including sub-Saharan Africa 1 (SSA1),

14
comprising of two sub-clades (I and II), and a South West Indian
Ocean Islands (SWIO) species. Showed the SSA1, sub-clade I
sequences shared similarity of 97.8-99.7% with the published Uganda
1 genotypes, and diverged by 0.3-2.2%. A pairwise comparison of
SSA1 sub-clade II sequences revealed a similarity of 97.2-99.5% with
reference southern Africa genotypes, and diverged by 0.5-2.8%. The
SSA1 sub-clade I whiteflies were widely distributed in East Africa (EA).
The SWIO species had a sequence similarity of 97.2-97.7% with the
published Reunion sequence and diverged by 2.3-2.8%.
Xiao-Jun (2012) showed diversity and genetic differentiation of
the whitefly B. tabaci species complex in China based on mtCOI and
cDNA-AFLP analysis. In an extensive field survey of the B. tabaci
complex present throughout part of China from 2004 to 2007. He
obtained 93 samples of B. tabaci from 22 provinces. He determined
that these Chinese haplotypes included 2 invasive species (Middle
East-Asia Minor 1 and Mediterranean), and 4 indigenous cryptic
species (Asia II 1, Asia II 3, China 3 and Asia II 7) by sequencing
mitochondrial cytochrome oxidase I gene (mtCOI). The diversity and
genetic differentiation of a subset of 19 populations of B. tabaci were
studied using cDNA amplified fragment length polymorphism (AFLP).
Mediterranean showed the lowest degree of similarity than the other
species. The data indicate that both Middle East-Asia Minor 1 and
Mediterranean were rapidly established in China.
2.3.1 Random Amplified Polymorphic DNA (RAPD)
RAPD primer was first developed in 1990 by Welsh and
McClelland using PCR to randomly amplified anonymous segments of
nuclear DNA with an identical pair of primers 8-10bp in length. RAPD
polymorphism could occur to base substitution at the primer binding
site or due to indels in the regions between the sites.
William et al. (1990) proposed the use of single arbitrary 10 base
oligonucleotide PCR primer for generation of molecular markers called
as RAPD markers. These could be easily developed and since were

15
based on PCR amplification followed by agarose gel electrophoresis,
they were readily detected. As a result, RAPD permit the wider
application of molecular maps in plant and insect science.
RAPD techniques have many other applications, including the
identification of cultivars/varieties, introgression studies, determination
of parentage, phylogenetic analysis and construction of genetic maps.
Polymorphisms were generated by random amplified polymorphic DNA
analysis used for fingerprinting by Connolly et al. (1994) and for
evaluating genetic relationships among diversified cultivars by Stiles et
al. (1993).
2.3.2 Molecular Markers and Diversity Analysis
Markert and Moller (1959) showed that genetic differences in
easily assayed enzymes (isozymes) changed their rate of migration
through gel matrices, in response to an electrical field. Using visible
markers together with isozymes, substantial genetic maps of some
organism had already been assembled by the late 1970s.
Litt and Luty (1989) found that microsatellite as simple sequence
repeats (SSRs) were short DNA sequence stretches consisting arrays
of mono, di, tri, tetra, or penta nucleotide units widely dispersed
throughout the genome.
Bell-Johnson et al. (1998) found the most fundamental of these
tools was DNA markers that detect differences in the genetic
information carried by different individuals. By knowing the location of a
gene, one can determine its presence by using nearby DNA markers,
without actually observing the phenotype, which could save a lot of
time and money, especially for quantitatively inherited traits.
Rafalski (2002) stated that DNA based genetic markers had
forever changed the practice of genetics. In the 20 years since that
discovery many different types of DNA based genetic marker had been
used for the construction of genetic maps, for the analysis of genetic
diversity, trait mapping as well as for applied diagnostic purposes.

16
Dangi et al. (2004) reported that RAPD markers were generated
through PCR amplifications of random genomic DNA segments using
10-15 base pairs of arbitrary sequence.
2.4 Genetic diversity in whitefly (Bemisia tabaci)
Perring et al. (1993) used RAPD-PCR to show differences in
amplification products between biotype A and biotype B. They found
90% similarity in the bands within each biotype and only 10% similarity
in the bands between biotypes.
Gawal and Bartlett (1993) found biotype A and B were easily
distinguished by RAPD-PCR. They noted that the speed, ease, and
ability to quantify differences among RAPD marker make this
technique a desirable tool for genetic studies of B. tabaci.
De Barro and Driver (1997) showed RAPD-PCR similar banding
patterns from populations of the B biotype from Australia, Cook Island,
Israel, Netherlands, New Caledonia, and USA. They also reported
differences in banding patterns among whiteflies from Sudan (esterase
biotype L), India (esterase biotype H, esterase biotype G), Pakistan
(esterase biotype K), New Zealand (esterase biotype unknown), Costa
Rica (esterase biotype C), Hainan (esterase biotype unknown), and
Nauru (esterase biotype unknown).
Guiro et al. (1997) evaluated B biotype from Denmark, France,
Israel, Italy, Japan, Netherlands, Spain and USA. They reported
90%similarity among biotypes in their study. They also found
populations from Spain and Portugal (both designated as RAPD type
II) to be distinct from all other populations tested in their study. RAPD-
PCR also showed distinct banding patterns from between the B
biotype, RAPD type II, and population from India, Pakistan, Turkey,
and biotype A from the USA.
De Barro et al. (1998) found distinct RAPD-PCR banding
patterns between a native population from Australia (Australian biotype
AN), the B biotype, and a unknown population from Nauru (designated
the Nauru biotype NA).

17
Lima et al. (2000) studied on identification of Bemisia tabaci
(Gennadius) (Hemiptera: Aleyrodidae) biotypes in Brazil using RAPD
markers. Whiteflies were collected from cultivated plants and weeds
from 57 different localities and on 27 distinct crops. RAPD analyses
using two selected 10-mer primers reliably identified the BR biotype
and the B biotype of B. tabaci and also differentiated other whitefly
species. The presence of the B biotype was confirmed in 20 Brazilian
states. The BR and B biotypes of B. tabaci were found to coexist in the
whitefly populations of three different localities: Jaboticabal, SP;
Rondonopolis and Cuiaba, MT, and Goiania, GO.
Banks et al. (2001) reported that presence of B-biotype of B.
tabaci in the Kolar district of Karnataka state, India. Adult B. tabaci
collected from tomato plants at nine states within epidemic zone and
analyzed by RAPD-PCR using primers OPB-20 and OPB-11, indicated
that hundred per cent of individuals per site had identical patterns to
those of B-biotype individuals from Israel and Florida.
Lima et al. (2002) showed genetic diversity of B. tabaci (Genn.)
populations in Brazil revealed by RAPD markers. A total of 72 markers
were generated by five RAPD primers and used in the analysis. All
primers produced RAPD patterns that clearly distinguished the Bemisia
biotypes and the two other whitefly species. Results also showed that
populations of the B biotype have considerable genetic variability.
Cluster analysis demonstrated that, in general, Brazilian biotype B
individuals are scattered independently in the localities where samples
were collected. The present study showed that the B biotype is
disseminated throughout the sampled areas, infesting several host
plants and predominates over the A biotype.
Horowitze et al. (2003) showed Q of B. tabaci identified in Israel
by using polyacrylamide gel electrophoresis (PAGE) during 1999-2000.
Whitefly samples were collected from several field as well as
greenhouse sites was determined by PAGE and by RAPD-PCR using
primers of arbitrary sequence. Based on the collected samples, it

18
appeared that both the B and Q biotype were presents in Israel and
that field population consist of a mixture of the two biotypes.
Callejas et al. (2005) used RAPD markers to detect genetic
pattern in spiralling whitefly (Aleurodicus disperses) (Hemiptera:
Aleyrodidae) populations from the Canary Islands. He found that, 68
different band were scored in 7 population using 6 primers for
amplification and no different RAPD pattern were found among
population from different Islands of Canaries. It will indicate that a very
high genetic similarity among populations and low level of genetic
variability.
Delatte et al. (2005) showed two genetic types of B. tabaci were
distinguished using RAPD-PCR and Cytochrome Oxidase I (COI) gene
sequence comparison. One type was assigned to B biotype and other
was genetically dissimilar to the population named Ms after the
Mascarenes Archipelago. Both B and Ms Population of B. tabaci
induced silverleaf symptoms on cucurbita spp, and were able to
acquire and transmit TYLCV. Taken together these results indicate that
Ms Genetics type should be considering a new biotype of B. tabaci.
Hasan (2006) used RAPD markers for diversity analysis of B.
tabaci (Gennadius) (Homoptera: Aleyrodidae) biotypes in Jordan.
Whiteflies were collected from cultivated plants, wild plants and weeds.
RAPD analysis identified B-Biotypes as well as cluster analysis
demonstrated that, biotype B individual which are scattered
independently in the localities according to the host plant and coexists
with cultivated plants. While an intermediate whitefly population BA
having biotype A distinguish band only with ORP-04 according to
Jaccard’s similarity. They have higher genetic distance with B-
Biotypes as well as percentage of biotypes A, B and BA samples.
Sharma et al. (2008) used RAPD markers for identification of
genetic diversity among whitefly collected from different plant hosts
and concluded that RAPD markers are able to differentiate the whitefly
isolates with specific host.

19
Qiu et al. (2009) used morphological characters, RAPD-PCR
and COI gene sequence for identification of three major B. tabaci
biotypes in China. He concluded that B, Q and Cv biotypes are
morphologically different in posterior wax and RAPD-PCR band
showed revealed differences in these biotypes using H16 primer as
well as based on COI gene.
Perumal et al. (2009) used RAPD markers for host plant
mediated population variation of cotton whitefly B. tabaci Gennadius
(Aleyrodidae; Homoptera) characterized with random DNA markers
and found that variation based on host plants being utilized by the
whitefly population.
Helmi (2010) used RAPD markers for identification of whitefly on
different host plant species in Egypt. Six host plant populations were
screened with seven RAPD primers and these populations were
clustered into two main groups with higher similarity matrix
percentages.
Gupta et al. (2010) used RAPD markers for identification of
specificity in B. tabaci for host plants were sequenced and developed
SCAR markers from B. tabaci genotype holding specificity to a specific
host plant.
Hameed et al. (2012) showed genetic diversity analysis of B.
tabaci population in Pakistan using RAPD markers. A total 80 samples
of B. tabaci collected from 14 district of the Punjab province and 7
district of the Sindh province were included. B. tabaci populations were
grouped into three main clusters and clearly distinguished the non B
biotype from the B biotype. This analysis showed that non B biotype is
prevalent in both provinces however B biotype is restricted to few
locations in Sindh.
Abdullahi (2013) showed molecular characterization of whitefly,
B. tabaci (Hemiptera: Aleyrodidae) populations infecting cassava. B.
tabaci collected from cassava and other plants in major cassava-
cultivation area of sub-Sahara Africa and from around the world. The

20
RAPD-PCR marker used to examine the genetic structure of the
population. Analysis of the internally transcribed spacer regions I (ITS
1) of the ribosomal DNA confirmed that the cassava population of B.
tabaci population were distinct from non-cassava population.
21
MATERIALS AND METHODS
The present study entitles “Molecular Diversity Analysis ofWhitefly (Bemisia tabaci) Collected from Different Regions ofMadhya Pradesh” was conducted at Plant Molecular Biology
Laboratory, Biotechnology Centre, Jawaharlal Nehru Agriculture
University, Jabalpur, India. This chapter deals with the materials and
methods used for molecular diversity analysis using different RAPD
molecular markers.
3.1 Materials
3.1.1 Source of Biological Material
About 17 populations of whiteflies (Table 3.1.1) were collected
from different geographical regions of East Madhya Pradesh.
Table 3.1.1: Bemisia tabaci samples use for present study withtheir location of collection.
SI. Collection site Latitude (N) Longitude (E)
1 Seoni 22º06”N 79º35”E2 Chhindwada 22º03”N 78º59”E3 Betul 21º88”N 77º98”E4 Hoshangabad 22º46”N 77º45”E5 Piparia 22º45”N 78º21”E6 Narsinghpure 22º57”N 79º15”E7 Sagar 23º5”N 78º5”E8 Tikamgarh 24º45”N 78º53”E9 Chhatarpure 24º54”N 79º38”E10 Katni 23º47”N 80º27”E11 Rewa 24º32”N 81º18”E12 Sidhi 24º2”N 82º39”E13 Satna 24º34”N 80º55”E14 Mandla 22º43”N 80º35”E15 Dindori 22º57”N 81º41”E16 Shahdol 23º00”N 81º3”E17 Jabalpur 23º10”N 79º59”
22
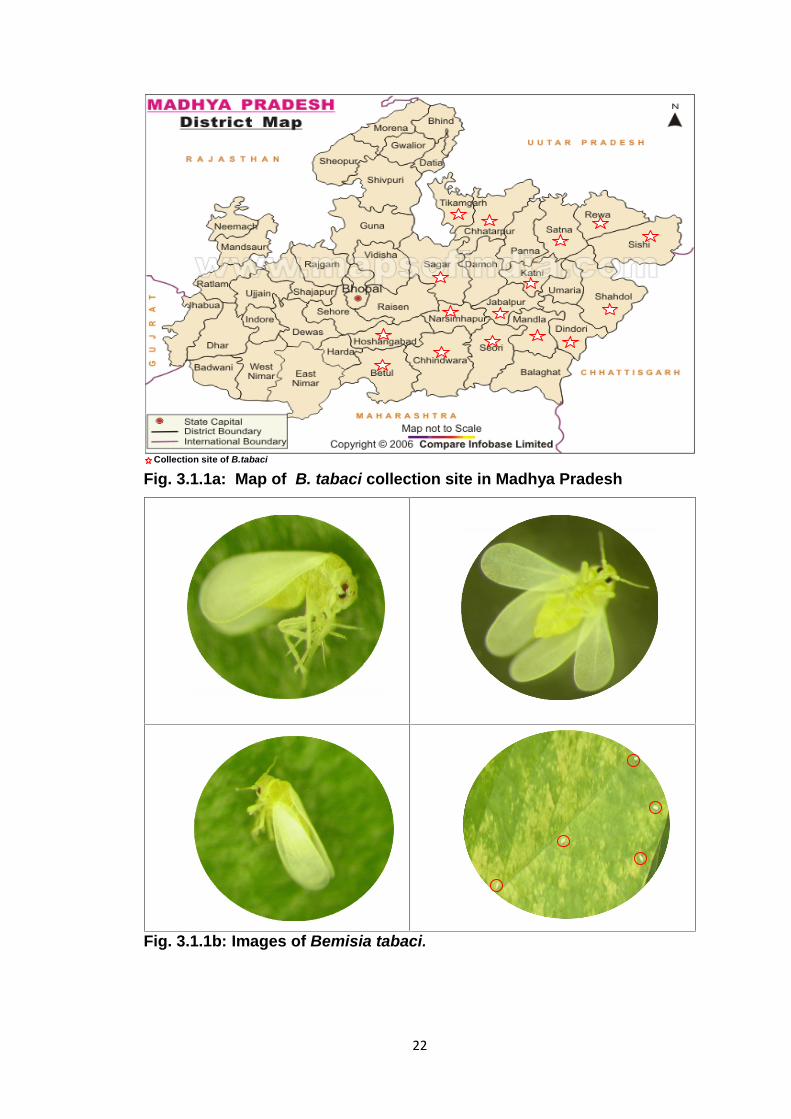
Collection site of B.tabaci
Fig. 3.1.1a: Map of B. tabaci collection site in Madhya Pradesh
Fig. 3.1.1b: Images of Bemisia tabaci.

23
3.1.2 Collection of Samples
Different host crops were selected to collect whitefly in morning
hours from the field during the kharif season 2012 for extraction of
genomic DNA. The collected samples were placed in centrifuge tubes
to transfer and then stored at -20°C.
3.1.3 Chemicals
Chemicals used for DNA extraction were purchased from Sigma
Aldrich (USA) and for PCR from Bangalore, Genie (India). DNA
ladders were purchased from Bangalore Genie (100bp) and Fermentas
Pvt. Ltd. India (1kb).
3.1.4 Molecular Markers
3.1.4.1 Mitochondrial Cytochrome Oxidase I (mtCOI)
mtCOI gene used as specific molecular marker to identify B
biotype of B. tabaci variant that exhibit rich biological differences, but
lack in morphological features.
3.1.4.2 Random Amplification Polymorphic (RAPD)
Random amplification polymorphic (RAPD) molecular markers
were used for diversity analysis in B. tabaci (Whitefly) collected from
different regions of East Madhya Pradesh. Random Amplified
Polymorphic DNA analysis of B. tabaci was done by using decamer
nucleotide primer obtained from R. Admin.
3.2 Methods
3.2.1 DNA isolation
Genomic DNA was isolated using standardized protocol. The
method described below gave a good quality and quantity of DNA.
Whiteflies are the most preferred material as it is difficult to collect at
any time and few amount of inhibitors and pigments that may hamper
the quality of DNA. The primary requirement for DNA isolation is to
extract DNA from cells in solution. Physical grinding results in physical
breakage of cells. Subsequently, addition of extraction solution
24
containing detergent like sodium-dodocel sulphate (SDS) assist
breakage of cell and nuclear membrane made up of lipids, which aids
in cell lysis and the components of cell are dispersed in the solution.
The DNA extraction solution was prepared as per details given
below.
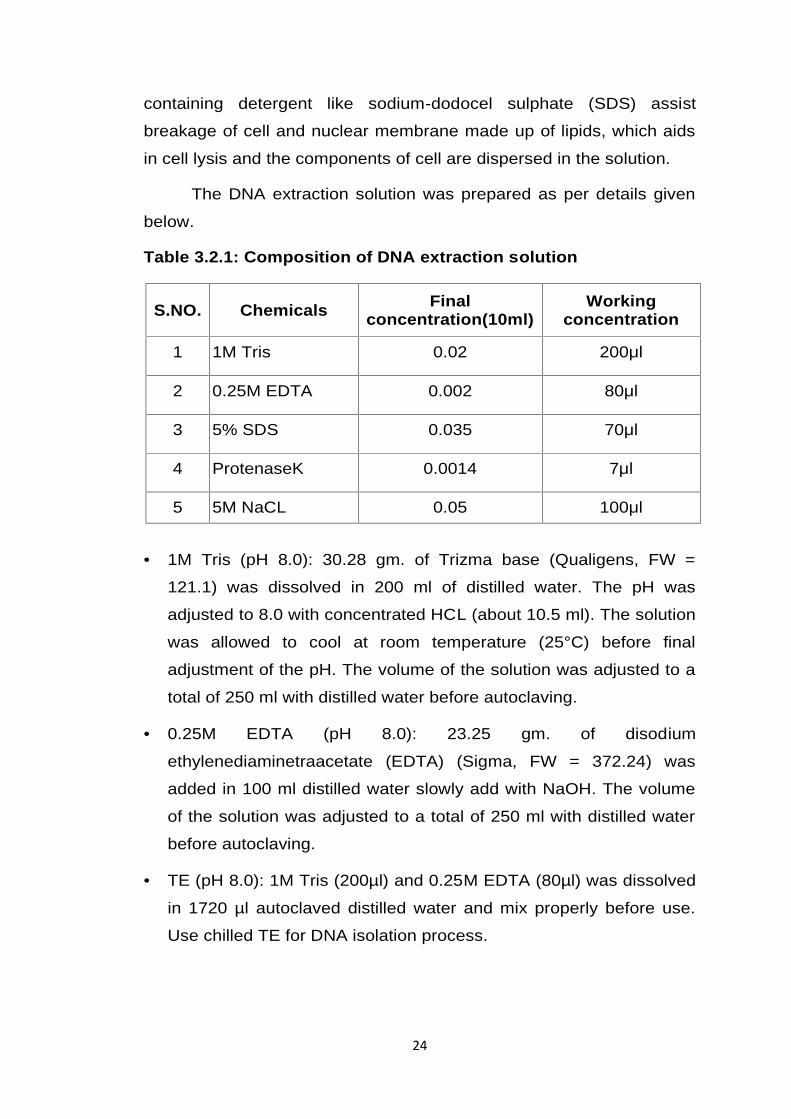
Table 3.2.1: Composition of DNA extraction solution
S.NO. Chemicals Finalconcentration(10ml)
Workingconcentration
1 1M Tris 0.02 200μl
2 0.25M EDTA 0.002 80μl
3 5% SDS 0.035 70μl
4 ProtenaseK 0.0014 7μl
5 5M NaCL 0.05 100μl
1M Tris (pH 8.0): 30.28 gm. of Trizma base (Qualigens, FW =
121.1) was dissolved in 200 ml of distilled water. The pH was
adjusted to 8.0 with concentrated HCL (about 10.5 ml). The solution
was allowed to cool at room temperature (25°C) before final
adjustment of the pH. The volume of the solution was adjusted to a
total of 250 ml with distilled water before autoclaving.
0.25M EDTA (pH 8.0): 23.25 gm. of disodium
ethylenediaminetraacetate (EDTA) (Sigma, FW = 372.24) was
added in 100 ml distilled water slowly add with NaOH. The volume
of the solution was adjusted to a total of 250 ml with distilled water
before autoclaving.
TE (pH 8.0): 1M Tris (200µl) and 0.25M EDTA (80µl) was dissolved
in 1720 µl autoclaved distilled water and mix properly before use.
Use chilled TE for DNA isolation process.

25
5% SDS: 5 gm. of Sodium-dodocel sulphate (SDS) (Himedia, Mol.
Wt. = 288.38) was added in autoclaved distilled water and makeup
volume 100 ml. It was mix properly in distilled water before use.
ProtinaseK: 40mg (Thermo Scientific, ~20mg/ml) dissolved in 2 ml
nuclease free water and mix properly before use. Keep in deep
freeze for further use.
5M NaCl: 73.05 gm. Sodium Chloride pellets (Merck, FW = 58.44)
was dissolved in 200 ml distilled water. The total volume was
adjusted to 250 ml with distilled water before autoclave.
3.2.2 The steps for DNA isolation are:
1. 30 whiteflies were thoroughly macerated with a micropestle in a
1.5 ml microcentrifuge tube.
2. Add 500 µl chilled TE, 70µl 5% SDS and 6µl proteinase-K was
added and mixed thoroughly.
3. The samples were incubated in a water bath at 55°C for 30 min.
Meanwhile, the samples were frequently mixed every 10 min to
ensure complete and even extraction.
4. The samples then removed from water bath, add chilled 100µl
(5M) NaCl and invert two times (slowly) properly. Keep the
sample in -20°C deep freeze or on ice cube for 30 min.
5. The samples were then cooling centrifuged for about 10 min at
10,000 rpm.
6. Supernatant so obtained was transferred to a fresh tube.
7. Then an equal (to supernatant) volume of chloroform: isoamyl
alcohol (24:1) v/v was added and mixed thoroughly but gently for
not less than 5 min.
8. The mixture was then centrifuged for about 10 min at 10,000
rpm.
9. Supernatant so obtained was transferred to a fresh tube.

26
10. An equal (to supernatant) volume of (100%) absolute ethanol
was added, mixed gently by inverting tubes and kept for
overnight undisturbed in normal freeze.
11. The mixture was then centrifuged for about 12 min at 10,000
rpm.
12. The supernatant was now discarded.
13. The tubes were placed at room temperature and allowed the
traces of ethanol to evaporate and dry the pellet. Then pellet was
dissolved in 40 µl TE for further use.
3.2.3 DNA Purification
The purification of DNA was carried out in order to remove the
impurities like RNA, proteins and polysaccharides. These are
considers as inhibitors in DNA amplification during PCR. Following
method was followed.
1. 5µl of RNase (5 mg/ml) was added to DNA extract, mixed well and
incubated at 37°C for 30 min.
2. This was followed by the addition of equal volumes of phenol:
chloroform: isoamyl alcohol (25:24:1) v/v and mixed vigorously.
3. The above mixture was centrifuged at 14,000 rpm for 10 min.
4. Supernatant was transferred to a fresh microcentrifuge tube. Equal
volume of pre-chilled ethanol (100%) was added and mixed gently
for DNA precipitation.
5. The precipitated DNA was pelleted by centrifugation at 12,000 rpm
for 5 min.
6. The pellet was dried at room temperature to completely remove
ethanol and was then dissolved in 40µl TE buffer and stored at -
20°C for further use.
3.2.4 Purity of DNA
Purity of DNA was checked by taking the ratio of Optical Density
(O.D) using UV-Spectrophotometer at 260 nm to that of 280 nm. 1ml

27
TE buffer was taken in a cuvette tube and calibrated the UV-
Spectrophotometer at 260 nm as well as 280 nm wavelength. 2µl of
DNA were added to 998μl of TE buffer, mixed properly and the optical
density (O.D) was measured. The samples which had the O.D ratio
between 1.7-1.9 (Maniatis et al., 1982) were used in subsequent
experiments. The DNA samples showing ratio beyond these values
were purified again.
3.2.5 Quantification of DNA
Isolated DNA was quantified by measuring the absorbance at
260 nm and 280 nm on a UV-spectrophotometer. 50µl/ml
concentration of double stranded DNA showed an absorbance of 1 at
260 nm. Concentration of DNA sample was calculated using following
formula:
O.D. 260 nm × 50µl DNA/ml × Dilution factor1000
3.2.6 Quality of DNA
The quality, quantity and integrity of isolated DNA were also
checked by horizontal submarine gel electrophoresis. 4µl of DNA
samples were loaded in each well in agarose gels along with the 1µl
λHind-III standard digested DNA ladder in 0.8% gel concentration at 60
volts for 90 min. The gel was stained by ethidium bromide and
observed under UV-Trans-illuminator. The amount of fluorescence was
proportional to the total mass of DNA. After quantification the DNA was
diluted by TE. The final concentration of DNA obtained was 25ng/µl.
3.2.7 Dilution of DNA
The quantified DNA was diluted according to the DNA quantity in
each sample for PCR amplified in sterile double distilled water.
Dilutions were carried out according to the following formula:
Dilution =Required concentration of DNA (ng/µl) X Total volume required (µl)
Available concentration of DNA (ng/µl)
28
3.3 DNA analysis of Mitochondrial Cytochrome Oxidase I(mtCOI)
The amplification of genomic DNA was carried out by using
mtCOI gene sequence. The COI region of the mtDNA gene was
partially amplified with the CI-J-2195 and L2-N-3014 pair of primers.
Procedure for amplification of DNA was followed as per protocol
described by Rocha et al. (2011). Primer sequences, the components
and their concentration used in the mtCOI-PCR reaction were
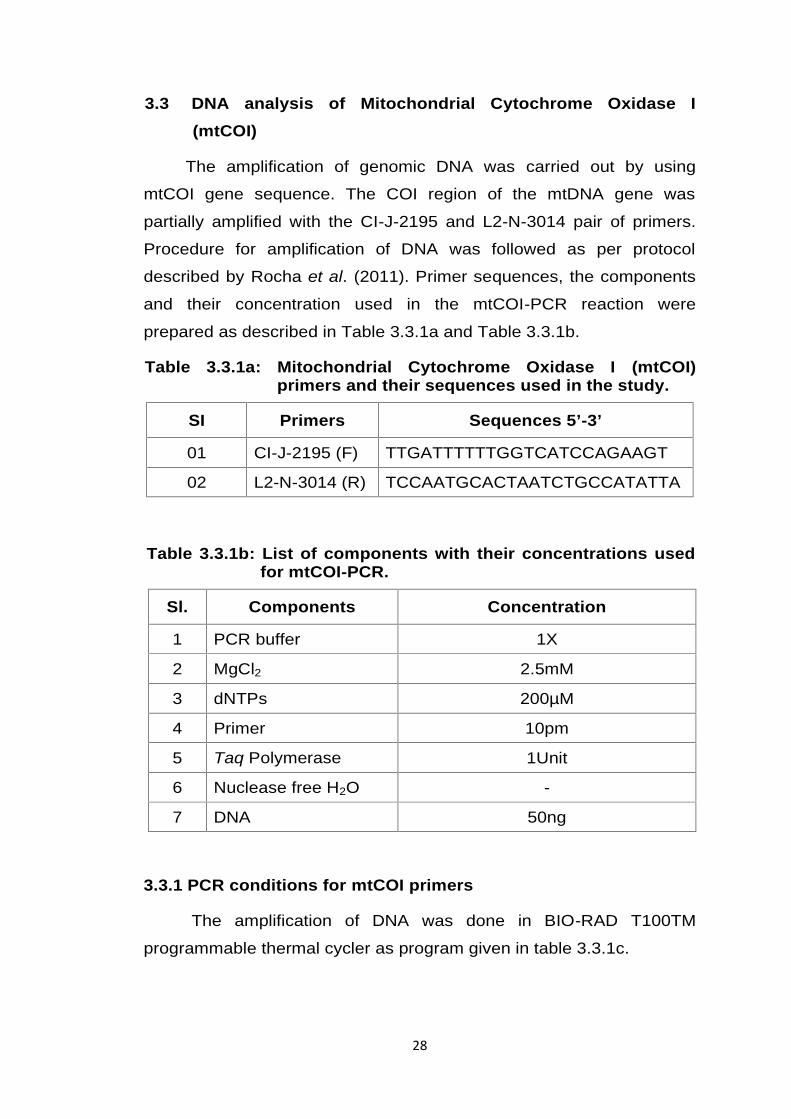
prepared as described in Table 3.3.1a and Table 3.3.1b.
Table 3.3.1a: Mitochondrial Cytochrome Oxidase I (mtCOI)primers and their sequences used in the study.
SI Primers Sequences 5’-3’
01 CI-J-2195 (F) TTGATTTTTTGGTCATCCAGAAGT
02 L2-N-3014 (R) TCCAATGCACTAATCTGCCATATTA
Table 3.3.1b: List of components with their concentrations usedfor mtCOI-PCR.
Sl. Components Concentration
1 PCR buffer 1X
2 MgCl2 2.5mM
3 dNTPs 200µM
4 Primer 10pm
5 Taq Polymerase 1Unit
6 Nuclease free H2O -
7 DNA 50ng
3.3.1 PCR conditions for mtCOI primers
The amplification of DNA was done in BIO-RAD T100TM
programmable thermal cycler as program given in table 3.3.1c.
29
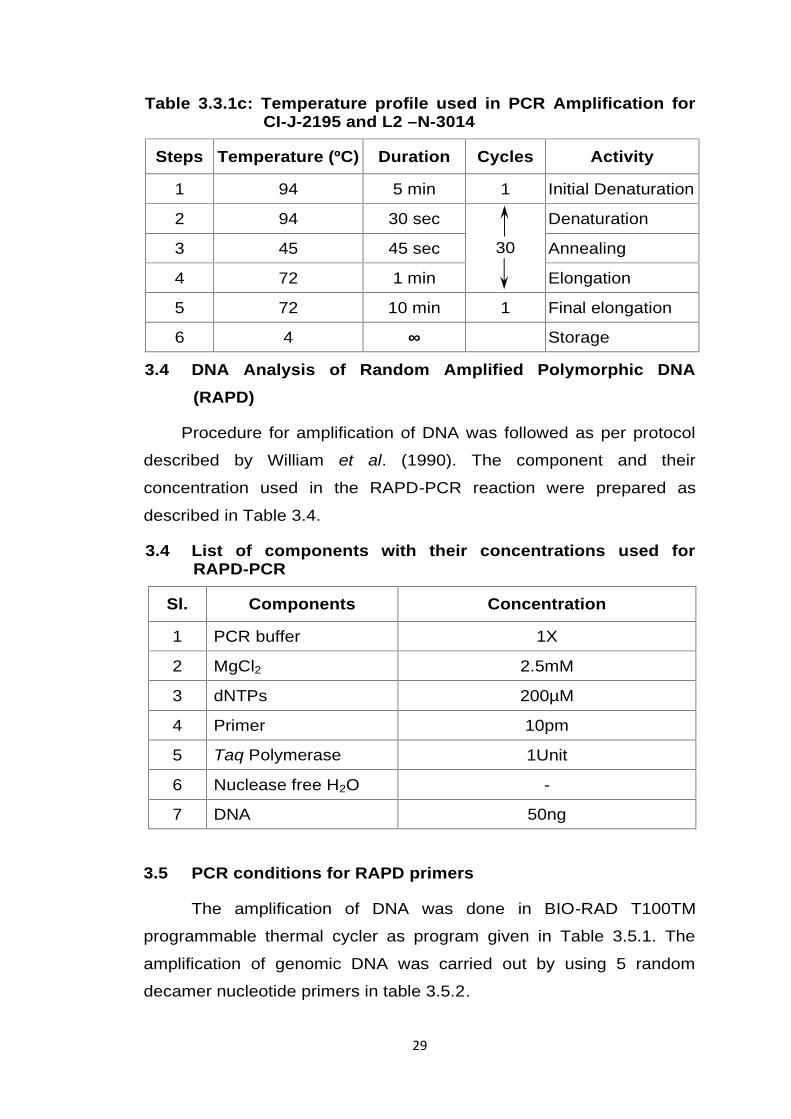
Table 3.3.1c: Temperature profile used in PCR Amplification forCI-J-2195 and L2 –N-3014
Steps Temperature (ºC) Duration Cycles Activity
1 94 5 min 1 Initial Denaturation
2 94 30 sec
30
Denaturation
3 45 45 sec Annealing
4 72 1 min Elongation
5 72 10 min 1 Final elongation
6 4 ∞ Storage
3.4 DNA Analysis of Random Amplified Polymorphic DNA(RAPD)
Procedure for amplification of DNA was followed as per protocol
described by William et al. (1990). The component and their
concentration used in the RAPD-PCR reaction were prepared as
described in Table 3.4.
3.4 List of components with their concentrations used forRAPD-PCR
Sl. Components Concentration
1 PCR buffer 1X
2 MgCl2 2.5mM
3 dNTPs 200µM
4 Primer 10pm
5 Taq Polymerase 1Unit
6 Nuclease free H2O -
7 DNA 50ng
3.5 PCR conditions for RAPD primers
The amplification of DNA was done in BIO-RAD T100TM
programmable thermal cycler as program given in Table 3.5.1. The
amplification of genomic DNA was carried out by using 5 random
decamer nucleotide primers in table 3.5.2.
30
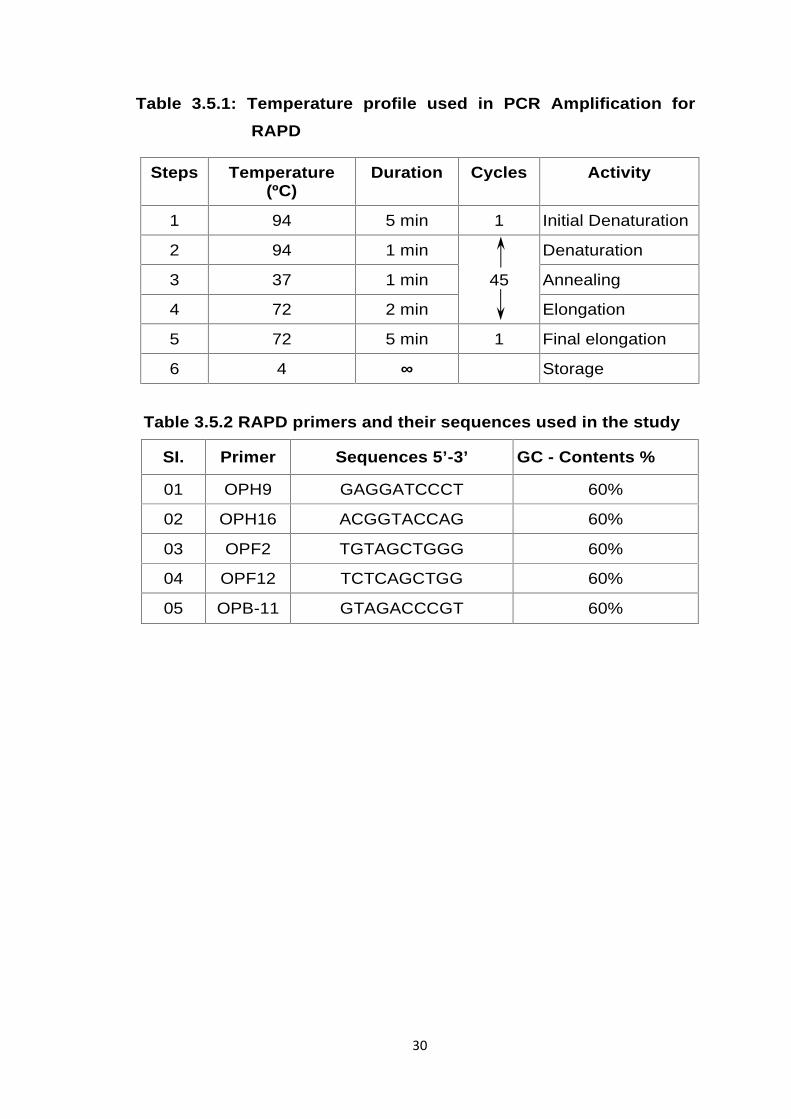
Table 3.5.1: Temperature profile used in PCR Amplification forRAPD
Steps Temperature(ºC)
Duration Cycles Activity
1 94 5 min 1 Initial Denaturation
2 94 1 min
45
Denaturation
3 37 1 min Annealing
4 72 2 min Elongation
5 72 5 min 1 Final elongation
6 4 ∞ Storage
Table 3.5.2 RAPD primers and their sequences used in the study
SI. Primer Sequences 5’-3’ GC - Contents %
01 OPH9 GAGGATCCCT 60%
02 OPH16 ACGGTACCAG 60%
03 OPF2 TGTAGCTGGG 60%
04 OPF12 TCTCAGCTGG 60%
05 OPB-11 GTAGACCCGT 60%

31
RESULTS
Whitefly, Bemisia tabaci ranks among the most noxious insects
attacking several field and greenhouse cultivated crops in India.
Biotypes of whitefly cannot be differentiated morphologically. DNA
markers represent very effective tool for analyzing genetic diversity of
any insect. With the known frequencies of each allele in the population
efficient characterization of whitefly could be achieved through
molecular markers.
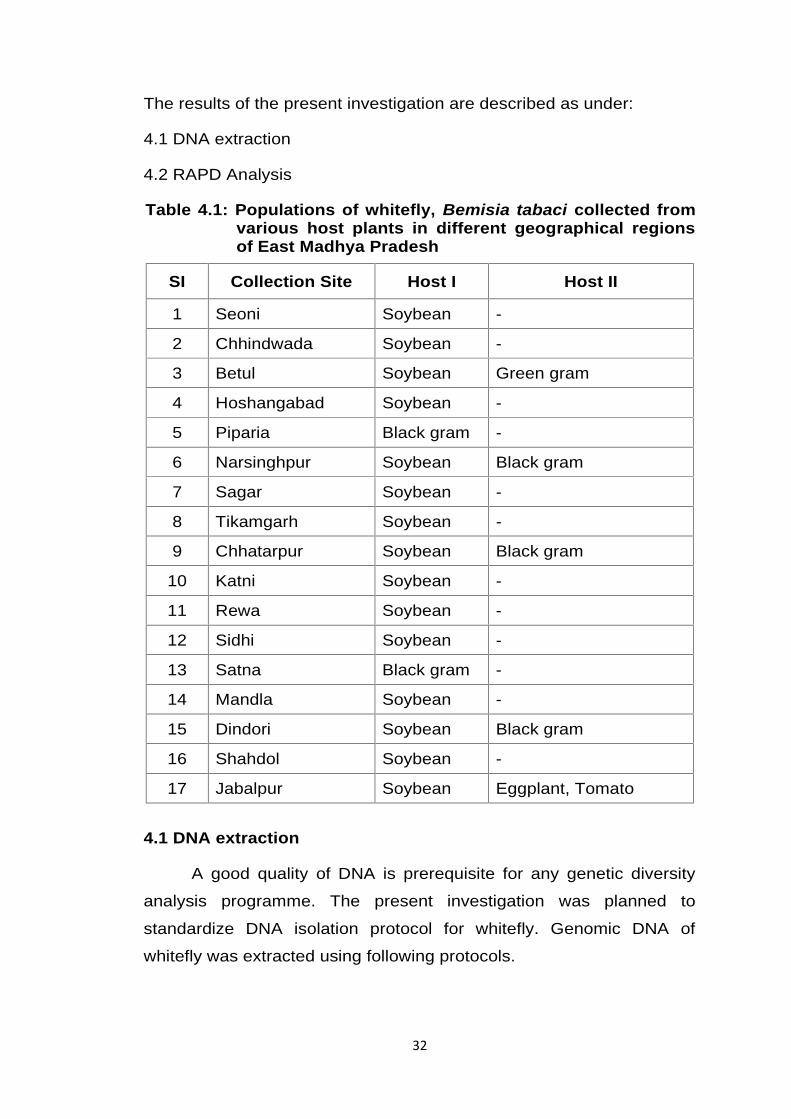
A total of 17 samples (Table 4.1) of whitefly collected from
different hosts growing in open environments areas from 17 different
geographical regions of East Madhya Pradesh. Apart from collection of
samples of whitefly, mainly from soybean crop whitefly populations
were also collected from green gram, black gram, eggplant and tomato
cultivated nearby soybean field. It is expected that whitefly may use
these plants as alternate hosts during harvesting time of soybean. At
least 200 individual adult whiteflies were collected from each location
for DNA analysis.The whiteflies were morphologically indistinguishable
from those collected from other localities. Collected samples were
used for molecular characterization during the present investigation
with the help of RAPD markers.
DNA fingerprinting or molecular characterization approaches
based on polymerase chain reaction have become methods of choice
for molecular diversity studies. It has distinct advantages to
discriminate samples with similar morphological characters. Being the
dominant marker, RAPD is one of the most common markers used in
the study of insect genetic diversity and characterization. These
markers may detect variation in both coding as well as noncoding
regions of the genome.
32
The results of the present investigation are described as under:
4.1 DNA extraction
4.2 RAPD Analysis
Table 4.1: Populations of whitefly, Bemisia tabaci collected fromvarious host plants in different geographical regionsof East Madhya Pradesh
SI Collection Site Host I Host II
1 Seoni Soybean -
2 Chhindwada Soybean -
3 Betul Soybean Green gram
4 Hoshangabad Soybean -
5 Piparia Black gram -
6 Narsinghpur Soybean Black gram
7 Sagar Soybean -
8 Tikamgarh Soybean -
9 Chhatarpur Soybean Black gram
10 Katni Soybean -
11 Rewa Soybean -
12 Sidhi Soybean -
13 Satna Black gram -
14 Mandla Soybean -
15 Dindori Soybean Black gram
16 Shahdol Soybean -
17 Jabalpur Soybean Eggplant, Tomato
4.1 DNA extraction
A good quality of DNA is prerequisite for any genetic diversity
analysis programme. The present investigation was planned to
standardize DNA isolation protocol for whitefly. Genomic DNA of
whitefly was extracted using following protocols.
33
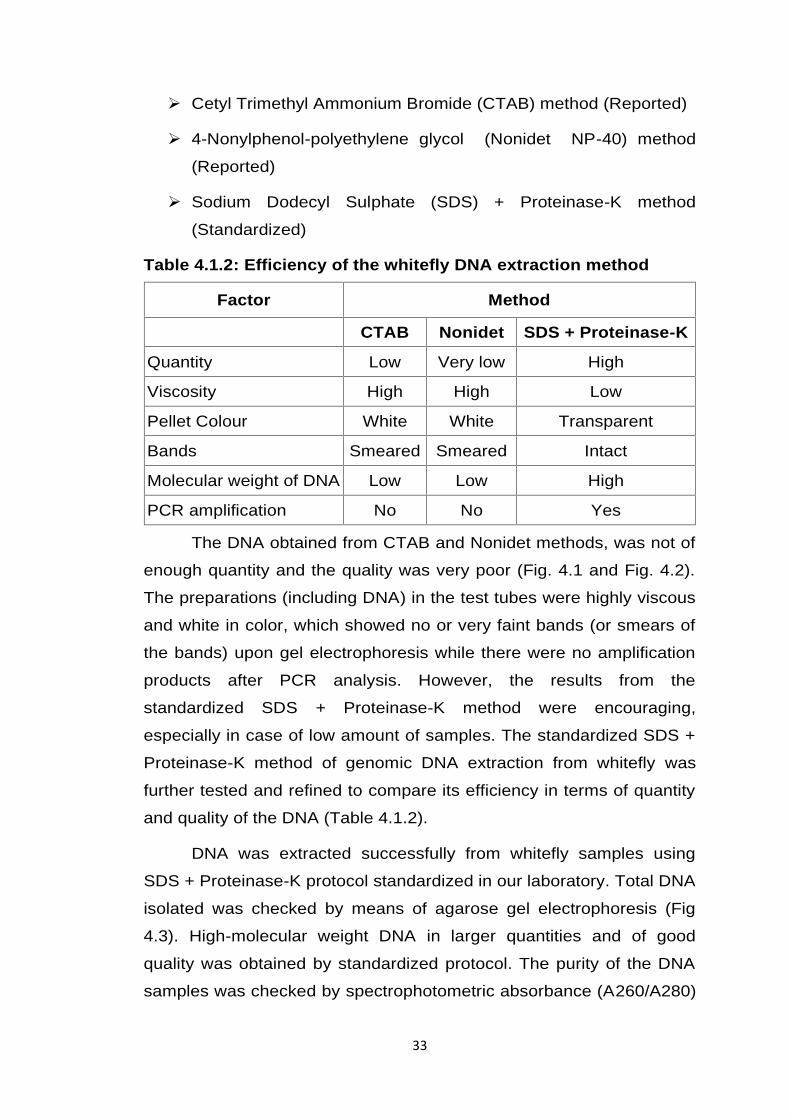
Cetyl Trimethyl Ammonium Bromide (CTAB) method (Reported)
4-Nonylphenol-polyethylene glycol (Nonidet NP-40) method
(Reported)
Sodium Dodecyl Sulphate (SDS) + Proteinase-K method
(Standardized)
Table 4.1.2: Efficiency of the whitefly DNA extraction method
Factor Method
CTAB Nonidet SDS + Proteinase-K
Quantity Low Very low High
Viscosity High High Low
Pellet Colour White White Transparent
Bands Smeared Smeared Intact
Molecular weight of DNA Low Low High
PCR amplification No No Yes
The DNA obtained from CTAB and Nonidet methods, was not of
enough quantity and the quality was very poor (Fig. 4.1 and Fig. 4.2).
The preparations (including DNA) in the test tubes were highly viscous
and white in color, which showed no or very faint bands (or smears of
the bands) upon gel electrophoresis while there were no amplification
products after PCR analysis. However, the results from the
standardized SDS + Proteinase-K method were encouraging,
especially in case of low amount of samples. The standardized SDS +
Proteinase-K method of genomic DNA extraction from whitefly was
further tested and refined to compare its efficiency in terms of quantity
and quality of the DNA (Table 4.1.2).
DNA was extracted successfully from whitefly samples using
SDS + Proteinase-K protocol standardized in our laboratory. Total DNA
isolated was checked by means of agarose gel electrophoresis (Fig
4.3). High-molecular weight DNA in larger quantities and of good
quality was obtained by standardized protocol. The purity of the DNA
samples was checked by spectrophotometric absorbance (A260/A280)
34
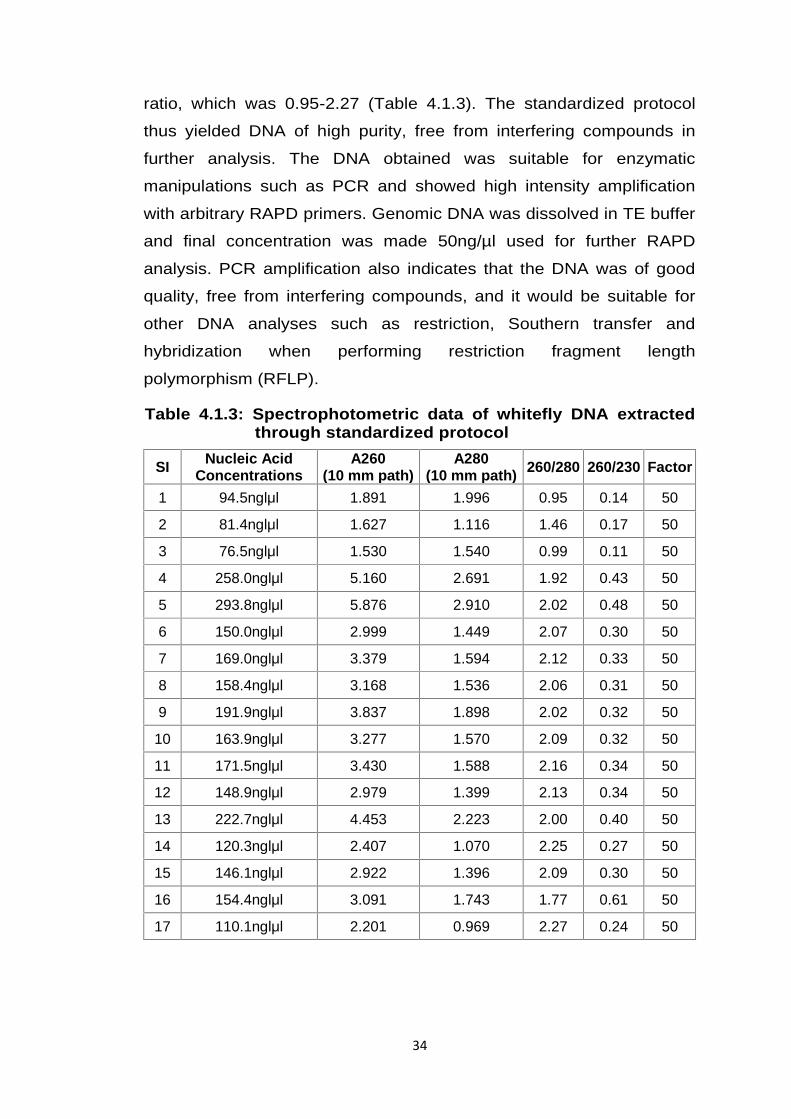
ratio, which was 0.95-2.27 (Table 4.1.3). The standardized protocol
thus yielded DNA of high purity, free from interfering compounds in
further analysis. The DNA obtained was suitable for enzymatic
manipulations such as PCR and showed high intensity amplification
with arbitrary RAPD primers. Genomic DNA was dissolved in TE buffer
and final concentration was made 50ng/µl used for further RAPD
analysis. PCR amplification also indicates that the DNA was of good
quality, free from interfering compounds, and it would be suitable for
other DNA analyses such as restriction, Southern transfer and
hybridization when performing restriction fragment length
polymorphism (RFLP).
Table 4.1.3: Spectrophotometric data of whitefly DNA extractedthrough standardized protocol
SI Nucleic AcidConcentrations
A260(10 mm path)
A280(10 mm path) 260/280 260/230 Factor
1 94.5nglμl 1.891 1.996 0.95 0.14 50
2 81.4nglμl 1.627 1.116 1.46 0.17 50
3 76.5nglμl 1.530 1.540 0.99 0.11 50
4 258.0nglμl 5.160 2.691 1.92 0.43 50
5 293.8nglμl 5.876 2.910 2.02 0.48 50
6 150.0nglμl 2.999 1.449 2.07 0.30 50
7 169.0nglμl 3.379 1.594 2.12 0.33 50
8 158.4nglμl 3.168 1.536 2.06 0.31 50
9 191.9nglμl 3.837 1.898 2.02 0.32 50
10 163.9nglμl 3.277 1.570 2.09 0.32 50
11 171.5nglμl 3.430 1.588 2.16 0.34 50
12 148.9nglμl 2.979 1.399 2.13 0.34 50
13 222.7nglμl 4.453 2.223 2.00 0.40 50
14 120.3nglμl 2.407 1.070 2.25 0.27 50
15 146.1nglμl 2.922 1.396 2.09 0.30 50
16 154.4nglμl 3.091 1.743 1.77 0.61 50
17 110.1nglμl 2.201 0.969 2.27 0.24 50

35
Analysis of mitochondrial cytochrome oxidase I (mtCOI)
Gene mtCOI was used as molecular marker to identify B. tabaci
variant that exhibit rich biological differences. Part of the mitochondrial
cytochrome oxidase I (mtCOI) gene was amplified with pairs of primers
C1-J-2195 (F) and L2-N-3014 (R) specific to COI region of whitefly
were used for PCR amplification. The PCR products derived from all
samples of DNA were of expected size, ~880bp (Fig. 4.4). This
suggested that the primer pair used was specific to COI region of B.
tabaci and could be used to amplify mtDNA region for further analysis.
The presence of mtCOI amplification was visualized by electrophoresis
in 1.2% agarose gel stained with ethidium bromide.
Analysis of RAPD-PCR in all whitefly populations
RAPD analysis is simple technique and can be performed even
in a moderately equipped laboratory. Initially five primers were
screened and a total of three primers were selected on the basis of
sharp and clear banding pattern for final RAPD-PCR analysis. The
PCR reaction was carried out using a single decamer primer at a time.
The sequences of these primers are presented in table 3.5.2.
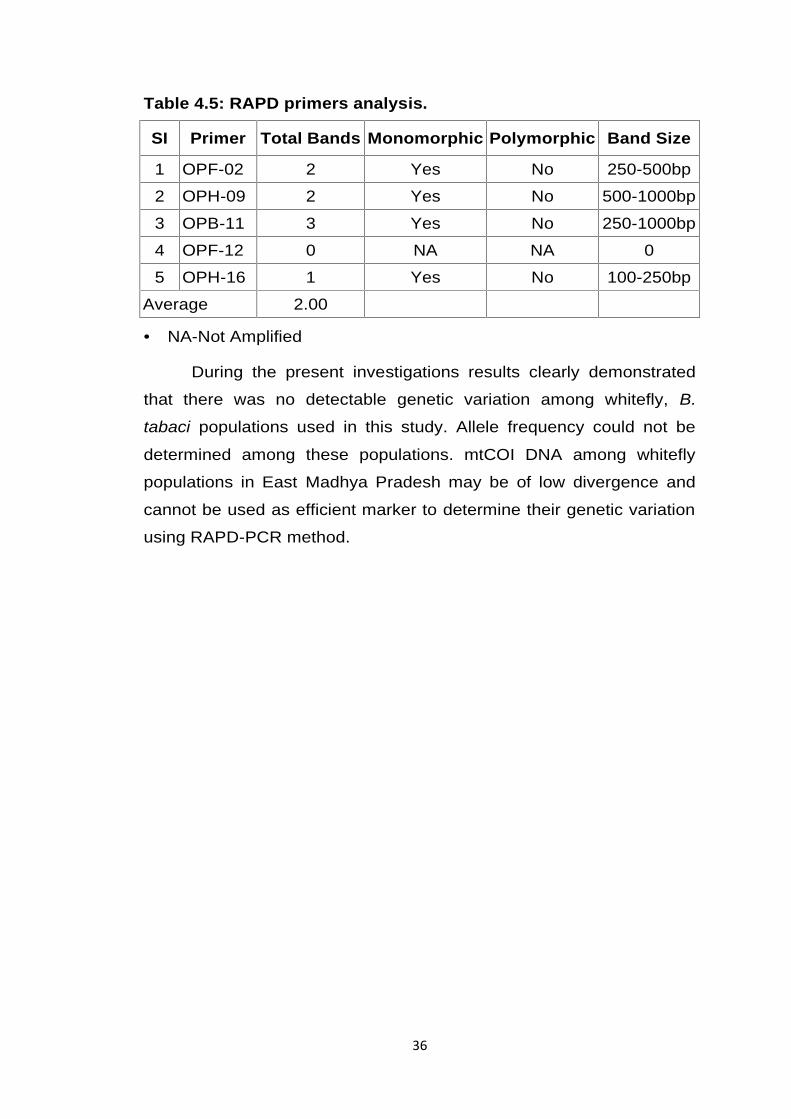
During the present investigation five decamer primers amplified
8 RAPD marker loci. The size of amplified marker ranged from 100bp-
1000bp. Maximum number of bands i.e. three were scored by primers
OPB-11 while minimum numbers of bands i.e. one produced by OPH-
16.
Out of eight bands scored by RAPD markers, all found to be
monomorphic. Average numbers of bands per primers were 2.0. In this
study, used primers for diversity analysis such as, OPF-02, OPH-09,
OPB-11, OPF-12 and OPH-16 all have same GC contents percentage
(60%). We found OPF-2, OPH-9 and OPB-11 with sharp and clear
band, while OPH-16 RAPD amplified DNA only in five samples. OPF-
12 did not amplify DNA among tested population.
36
Table 4.5: RAPD primers analysis.
SI Primer Total Bands Monomorphic Polymorphic Band Size
1 OPF-02 2 Yes No 250-500bp
2 OPH-09 2 Yes No 500-1000bp
3 OPB-11 3 Yes No 250-1000bp
4 OPF-12 0 NA NA 0
5 OPH-16 1 Yes No 100-250bp
Average 2.00
NA-Not Amplified
During the present investigations results clearly demonstrated
that there was no detectable genetic variation among whitefly, B.
tabaci populations used in this study. Allele frequency could not be
determined among these populations. mtCOI DNA among whitefly
populations in East Madhya Pradesh may be of low divergence and
cannot be used as efficient marker to determine their genetic variation
using RAPD-PCR method.

37
DISCUSSION
The whitefly (Hemiptera: Aleyrodidae) is one of the most
important pest of crops in the world and is a vector of more than 100
plant viruses (Jones et al., 2003). Bemisia tabaci belongs to a group of
insects named whiteflies that is commonly known as different names
based on crops on which it attack e.g. tobacco, cotton or sweet potato
whitefly (Rekha et al., 2005). It (Gennadius) (Hemiptera: Aleyrodidae)
is one of the most devasting tropical and sub-tropical agricultural pests
(Byrne and Bellows, 1991) affecting the yield of a broad range of
agricultural, fiber, vegetable and ornamental crops (Cahill et al., 1996)
and is considered one of the world’s top invasive species (Boykin et
al., 2007). B. tabaci has increased in importance globally as a serious
pest, because of the polyphagous nature of some biotype and the
diverse ways that it damage crops (Rekha et al., 2005).
Genetic diversity analysis is the first and foremost step bio-
control agent improvement programmed against strain of pest or
insects. However, to have a reliable estimate of genetic relationship
and genetic diversity, generally a large number of polymorphic markers
are required. DNA markers are very effective tool for analysing genetic
diversity in any organism.
Genetic diversity analysis is possible using various molecular
markers such as ISSR (Blair et al., 1999), RAPD (Dawson et al., 1993)
and AFLP (Negi et al., 2000) etc. It greatly depends upon type of
markers used, their distribution in the genome, loci they amplify, level
of polymorphism and reproducibility. PCR based molecular markers
(RAPDs, ISSRs, STMSs etc.) are preferred over hybridization based
markers like RFLPs for genetic diversity analysis, because they permit
the use of smaller amount of DNA from each plant or insect being
genotype and also reduce the time, labour and operational cost of DNA
extraction (Sant et al., 1999).

38
The whitefly, Bemisia tabaci populations were collected from
plants growing in open environments areas in different geographical
regions of East Madhya Pradesh during kharif season 2012. A total of
17 populations of whiteflies were collected different geographical
regions.
Prerequisite for taking advantage of these techniques is good
quality of genomic DNA. Whitefly contains small fragment size of DNA,
which make the DNA isolation task more difficult. Different published
protocols were used for whitefly genomic DNA extraction. We used
proteinase-K with SDS in our protocol. The ultimate goal of DNA
extraction is to obtain DNA with high molecular weight and without
impurities. The disruption of cellular and nuclear membrane is the first
necessary step to obtained purified DNA. The detergents
(Surfactants), such as the sodium dodecyl sulfate (SDS) and CTAB,
were adopted to accomplish disruptions but unable to stable DNA for
long time in case of whitefly. CTAB can precipitate genomic DNA, and
it is also popular because of its ability to remove polysaccharides from
bacterial and plant preparations (Ausbel et al., 1998). Published
methods to isolate DNA from insects are not always effective, because
they have high concentrations of phenolics and other secondary plant
compounds in their digestive tracts. Phenolics, as powerful oxidizing
agents, can reduce the yield and purity of DNA by binding covalently
with the extracted DNA, thereby inhibiting further enzymatic processes
of the DNA such as restriction digestion and polymerase chain reaction
(PCR) (Juen and Traugott 2006: Serrano et al., 1999).
The present investigation emphasized to know the effect of
proteinase-K on extracted genomic DNA of the whitefly populations
found that significant effect where as CTAB and Nonidet-40 method
did not show any major impact on total DNA concentration. The
combination of SDS and Proteinase-K was better at separating DNA
from almost all the polysaccharides and protein complexes. Residual
proteins and lipids could readily be removed by extraction with
chloroform-isoamyl alcohol (Merante et al., 1998). Proteinase-K is

39
hydrolytic enzyme which acts by hydrolyzing peptide bonds and exhibit
a wide range of cleavage preferences. The Proteinase-K can also be
used as a protective agent during DNA extraction, by inactivating
endogenous nucleases which are responsible for degradation of DNA.
Sodium Dodecyl Sulphate (SDS) with Proteinase-K gave good
quality and quantity of DNA. Whereas, other tried protocols using
different chemicals such as Nonidet NP-40 (Gupta et al., 2010) and
CTAB didn’t produce DNA with enough quantity. The isolated DNA
samples with all protocols were run on 0.8% agarose gel. DNA isolated
by standardized protocol (SDS + Proteinase-K) produce intact band of
high molecular weight whereas, DNA from other protocols produced
smears that was due to low quantity of DNA with contamination of
interfering compounds. Similarly, Helmi (2010) also found good quality
of DNA with SDS method but DNA was not stable for long time.
However DNA isolated from using standardized protocol was of
enough quality 100-150μg/g and good quality. The genomic DNA
isolated by protocol standardized by us was stable for long time.
The biotypes of B. tabaci are important because they act as a
major vector of many plant viruses in many parts of the world.
Therefore, there is a need to understand the prevalence of vector
biotypes under local conditions. For identification of whitefly biotype
used the Mitochondrial Cytochrome Oxidase I (mtCOI) molecular
marker, we used the PCR condition as mentioned by Rocha et al.
(2011). The PCR products which derived from all samples of DNA
were of expected size, ~880bp. Our result was in accordance with
Rocha et al. (2011). They also found ~880bp amplified product in B
biotype when they used mtCOI primers during their study on whitefly.
This suggested that the primer pair used was specific to COI region of
B. tabaci and could be used to amplify COI region for further analysis.
Which indicate the presence of B biotype population in East Madhya
Pradesh.
During this study, RAPD markers were used for diversity
analysis among identified B biotypes. Random amplified polymorphic

40
DNA polymerase chain reaction (RADP-PCR) is a relatively simple,
inexpensive and rapid technique, revealing polymorphisms which are
useful as genetic and taxonomic markers (Welsh and McClelland,
1990). RAPD has been applied to study of insects (Haymer, 1994) and
to differentiate whiteflies, including the identification of different
biotypes of B. tabaci (Guirao et al., 1997; De Barro and Driver, 1997;
Cervera et al., 2000; Moya et al., 2001).
During present investigation, five reported RAPD markers
(Calvert et al., 2001) were screened for PCR amplification, out of
these, 3 RAPD primers were selected, based upon their sharp and
multiple banding pattern using two accessions of whitefly. These three
markers amplified all samples. One RAPD marker OPH-16 amplified
only five samples (1, 2, 3, 4 and 13). Eight marker loci were amplified
by four decamer primers; Average numbers of bands per primers were
2.00. Good and clear banding profile has been obtained by primers
with 60% GC content. Fritsch et al. (1993) also demonstrated the
importance of the GC contain of primers on the yield of PCR amplified
products. All markers amplified monomorphic bands among all B
biotypes.
In the aspect of genetic variability study due to host plants in B.
tabaci populations no variation was found in our study whereas,
Sharma et al. (2008) detected genetic variability due to host plants in
B. tabaci populations that collected from six different host plants using
RAPD-PCR. Perumal et al. (2009) also found differences among B.
tabaci populations collected from four different host plants.
RAPD markers represent an efficient and inexpensive way to
generated molecular data and thus have been used successfully in
various taxonomic polygenetic studies (Aboelwafa et al., 1995; Sharma
et al., 1995; Friesen et al., 1997; Smelcerovic et al., 2006). RAPD
markers are generated by PCR amplification of random genomic DNA
segments with single primer in an arbitrary sequence. They are usually
dominant markers with polymorphism between individual defined by

41
the presence or absence of a particular RAPD band (Staub et al.,
1993).
Results clearly demonstrated that there was no detectable
genetic variation among whitefly, B. tabaci populations used in this
study. Allele frequency could not be determined among these
populations. Whitefly populations of Eastern part of Madhya Pradesh
may be of low divergence or mtCOI DNA cannot be used as efficient
marker to determine their genetic variation using RAPD-PCR method.
It seems that B. tabaci found in the regions of collection are of
conserved type.

42
SUMMARY, CONCLUSIONS AND SUGGESTIONS FORFUTHER WORK
6.1 Summary
The present study entitles “Molecular Diversity Analysis of
Whitefly (Bemisia tabaci) Collected from Different Regions of Madhya
Pradesh” was carried out with the objectives to standardize genomic
DNA isolation method and analyze the genetic diversity of whitefly
using DNA fingerprinting.
This study aimed to employ DNA based method to clarify the
existing of biotype among whitefly populations in East Madhya
Pradesh, India. The genomic DNA of whitefly collected from different
geographical regions of East Madhya Pradesh was isolated by
standardize protocol (SDS + Proteinase-K) produce intact band with
high molecular weight. The mtCOI primer was used for identification of
B biotype. Amplified products were resolved by electrophoresis on
1.2% agarose gel and photographed under gel documentation system.
Specific mtCOI primer CI-J-2195 and L2-N-3014 showed the presence
of ~880bp bands which confirmed the presence of B biotype in
different geographical regions of East Madhya Pradesh.
All identified B biotypes were used for molecular diversity
analysis. The DNA samples were amplified with 5 RAPD primers in
thermal cycler. Amplified products were resolved by electrophoresis on
1.2% agarose gel and photographed under gel documentation system.
Four decamer primers amplified 8 RAPD marker loci. All 8 bands
scored by RAPD markers found to be monomorphic. Average number
of bands per primer was 2.00. One RAPD primer is not amplified in all
population. No variation was found among whitefly samples collected
from different geographical regions of East Madhya Pradesh.

43
6.2 Conclusion
Whitefly, Bemisia tabaci populations genomic DNA extracted
through standardized protocol (SDS + Proteinase-K) at Biotechnology
Centre, JNKVV, Jabalpur, get a good results for further analysis.
These genomic DNA were analyzed by mtCOI and RAPD molecular
markers. This COI gene fragment is sequence marker used for to
identify B. tabaci variant that exhibit rich biological differences, but lack
in morphological features. It is a variable region that serves as fast
clock of evolution. The COI region of the mt DNA gene was partially
amplified with the CI-J-2195 and L2-N-3014 pair of primers. The
analysis of PCR with mtCOI primers provided amplification of the
expected size of ~880bp. This band size showed the presence of “B”
biotype in 17 different geographical regions of East Madhya Pradesh,
India.
A total four RAPD primers were used to show the genetic
diversity between the whiteflies collected from 17 different
geographical regions of East Madhya Pradesh. A total of 8 bands were
generated by 4 RAPD primers. All of them were monomorphic. The
amplification of monomorphic bands showed 100% genetic similarity
among all “B” biotypes in Eastern part of Madhya Pradesh.
6.3 Suggestion
1) An intensive survey is required for the identification of biotype of
Bemisia tabaci.
2) For more reliable data and for assessing genetic diversity, more
number of primers (RAPD) and more stringent molecular markers
such as SSR and AFLP should be applied.
3) More genotypes can be used for diversity analysis using RAPD
markers.

44
REFERENCES
Abdullahi, I., S. Winter, G.I. Atiri and G. Thottappilly (2013). Molecularcharacterization of whitefly, Bemisia tabaci (Hemiptera:Aleyrodidae) populations infesting cassava. Bull. Entomol. Res.,93(2): 97-106.
Abo-elwafa, A.K. Murai and T. Shimada (1995). Intra and interspecificvariations in Lens revealed by RAPD markers. Theor. Appl.Genet., 90: 335-340.
Ausbel, M., R. Brent, R.E. Kingston, D.D Moore, J.G Seidman and K. Struhl(1998). Curr. prot. Mol. Biol, Wiley and Sons, New York, 561-563.
Banks, G.K., J. Colvin, R.K.C. Reddy, N.M. Maruthi, V. Muniyappa, M.HVenkatesh, M.K. kumar, S.A. Padmaja, J.F. Beitia and F.S.Seal (2001). First report of B. tabaci, B-biotype in India and anassociated tomato leaf curl virus disease epidemic. Plant. Dis.,85: 231.
Bedford, I. D., P.G. Markham, J.K. Brown and R.C. Rosell (1994).Geminiviruses transmission and biological characterization ofBemisia tabaci (Gennadius) from different geographic regions.Ann. Appl. Bio., 125: 311-325.
Bel-Kadhi, M.S., J.C. Onillon and J.L. Cenis (2008). Molecularcharacterization of Bemisia tabaci biotypes in Southern Tunisia.Tunisian J Plant Prot., 3: 79-85.
Bell-Johnson, B.B., G.G. Johnson, J. Meksem and D.A. Lightfoot (1998).Methods for high-throughput marker assisted selection forsoybean. Soybean Genet. News., 25: 115-118.
Bertolani, R., S. Garagna, G.C. Manicardi, L. Rebecchi and C.A. Redi (1994).New data on the nuclear DNA content in some species oftardigrades. In: R. Argano, C. Cirotto, E. Grassi Milano and L.Mastrolia (Eds), Contributions to Animal Biology, HalocynthiaAssociation, Palermo. 103-109.
Bethke, J.K., F.J. Byrne, G.S. Hodges, C.L. Mckenzie and R.G. Shatters Jr.(2009). First record of the Q biotype of the sweetpotato whitefly,Bemisia tabaci, in Guatemala. Phytoparasitica, 37: 61-64.
Bink-Moenen, R.M. (1983). Revision of the African whiteflies (Aleyrodidae)mainly based on collection from Tchad. Monografieen van deNederlandse Entomol. Enw., 10: 273-287.
Bink-Moenen, R.M. (1990). Whiteflies: Diversity, Biosystematics andEvolutionary Patterns: Whiteflies: their Bionomics, Pest Statusand Management (ed. by D Gerling) Intercrop Ltd., Andover,Hants, UK., 1-11.
Bink-Moenen, R.M. and L.A. Mound (1990). Whiteflies: Diversity,biosystematics and evolutionary patterns: Whiteflies: theirbionomics, pest status and management (ed. by D. Gerling)Intercept, Andover, UK.1-11.

45
Bird, J. (1957). A whitefly transmitted mosaic of Jatropha gossypifolia. Univ.Puerto Rico, Agric. Exp. Stn., 22: 35.
Bird, J. and K. Maramorosch (1978). Viruses and virus diseases associatedwith whiteflies. Adv. Virus. Res., 71: 233-260.
Blair, M.W., O. Panaud and S.R. McCouch (1999). Inter-simple sequencerepeats (ISSR) amplification for analysis of microsatellite motiffrequency and fingerprinting in rice (Oryza sativa L.). Theor.Appl. Genet., 98: 780-792.
Bosco, D., A. Loria, C. Sartor and J.L. Cenis (2006). PCR-RFLP Identificationof Bemisia tabaci Biotypes in the Mediterranean Basin.Phytoparasitica, 34(3): 243-251.
Boykin, L.M., R.G. Shatters Jr, R.C. Rosell, C.L. Mckenzie, R.A. Bagnall,P.D. Barro and D.R. Frohlich (2007). Global relationships ofBemisia tabaci (Hemiptera: Aleyrodidae) revealed usingBayesian analysis of mitochondrial COI DNA sequences. Mol.Phylogenet. Evol., 44: 1306-1319.
Brown, J.K. (2000). Molecular markers for the identification and globaltracking of whitefly vector-begomovirus complexes. Virus Res.,71: 233-260.
Brown, J.K., D.R. Frohlich and R.C. Rosell (1995). The sweetpotato orsilverleaf whiteflies: biotypes of Bemisia tabaci or a speciescomplex? Ann. Rev. Entomol., 40: 511-534.
Buckner, J.S., D.R. Nelson and M.C. Mardaus (1994). The lipid-compositionof the wax particles from adult whiteflies, Bemisia tabaci andTrialeurodes vaporariorum. Insect. Bioch. Mol. Biol., 24: 977-987.
Burban, C., L.D.C. Fishpool, C. Fauqet, D. Fargette, and J.C. Thouvenel(1992). Host associated biotypes within West Africanpopulations of the whitefly Bemisia tabaci (Genn.), Hom.Aleyrodidae. J. App. Entomol., 113: 416-425.
Butler, G.D. Jr., T.J. Henneberry and W.D. Hutchison (1986). Biology,sampling and population dynamics of Bemisia tabaci., In G.E.Russell, ed. Agric. Zool. Rev., Intercept, UK. 17-28.
Byrne, D.N. and N.F. Hadley (1988). Particulate surface waxes of whitefliesmorphology, composition and waxing behavbiour. Physiol.Entomol., 13: 267-276.
Byrne, D.N. and T.S. Bellows Jr (1991). Whitefly biology. Ann. Rev. Entomol.,36: 431-457.
Byrne, D.N., A.C. Cohen and E.A. Draeger (1990). Water uptake from planttissue by the egg pedicel of the greenhouse whitefly,Trialeurodes vaporariorum (Westwood) (Homopter: Alyrodidae).Can. J. Zool., 68: 1193-1195.
Cahill, M., I. Denholm, G. Rose, K. Gorman and D. Johnston (1996).Relationship between bioassay data and the simulated fieldperformance of insecticides against susceptible and resistantBemisia tabaci. Bull. Entomol. Res., 86: 109-116.
Callejas, C., F.J. Beitia, A. Gobbl, A. Velasco and M.D. Ochando (2005). Theuse of RAPD markers to detect genetic patterns in Aleurodicus

46
disperses (Hemiptera: Aleyrodidae) populations from theCanary Islands. Eur. J. Entomol., 102: 289-291.
Calvert, L.A., M. Cuervo, J.A. Arroyave, L.M. Constantino, A. Bellotti and D.Frohlich (2001). Morphological and mitochondrial DNA markeranalyses of whiteflies (Homoptera: Aleyrodidae) colonizingcassava and beans in Colombia. Ann. Entomol. Soc. Am., 94:512-519.
Cambell, B.C. (1993). Cogurent evolution between whiteflies (Homoptera:Aleyrodidae) and their bacterial endosymbionts based onrespective 18S and 16S rDNAs. Curr. Microbiol., 26: 129-132.
Cambell, B.C., J.D. Seffen-Cambell and R.J. Gill (1994). Evolutionary originof whiteflies (Hemiptera: Sternorrhynchi: Aleyroididae) inferredfrom 18S rDNA sequences. Insect. Mol. Biol., 3(2): 73-88.
Cambell, B.C., J.D. Seffen-Cambell, and R.J. Gill (1996). Origin and radiationof whiteflies: an initial molecular phylogenetic assessment. In D.Gerling and R.T. Mayer, eds. Bemisia: 1995 Taxonomy,Biology, Damage, Control, and Mangement. Intercept Ltd.,Andover, Hants, UK. 29-51.
Cervera, M.T., J.A. Cabezas, B. Simon, J.M.M. Zapater, F. Beitia and J.L.Cenis (2000). Genetic relationships among biotypes of Bemisiatabaci (Hemiptera: Aleyrodidae). Bull. Entomol. Res., 90: 391-396.
Chu, D., C.S. Gao, P.D. Barro, F.H. Wan, Y.J. Zhang (2011). Investigation ofthe genetic diversity of an invasive whitefly (Bemisia tabaci) inChina using both mitochondrial and nuclear DNA markers. Bull.Entomol.Res., 101(4): 467-475.
Chu, D., G. Liu, F. Wan, Y. Tao and R. Gill (2010). Phylogenetic analysis andrapid identification of the whitefly, Bemisia afer, in china. J.Insect. Sci., 10(86): 1-9.
Claridge, M.F., H.A. Dawah and M.R. Wilson (1997). Species in insectherbivores and parasitoids-sibling species, host races andbiotypes. In M.F. Claridge, A. Dawah and M.R. Wilson, eds.Species, The Units of Biodiversity. Chapman and Hall, London.247-272.
Cock, M.J.W. (1986). Bemisia tabaci, a literature survey on the cottonwhitefly with an annotated bibliography. CAB IIBC, SilwoodPark, UK, 121.
Cohen, S. (1990). Epidemiology of Whitefly-Transmitted Viruses: Whiteflies:their Bionomics, Pest Status and Management (ed. by D.Gerling) Intercept Ltd., Andover, Hants, UK., 211-225.
Connoly, A.G., I.D. Godwin, M. Cooper and I.H. Delacy (1994). Interpretationof randomly amplified polymorphic DNA marker data forfingerprinting sweet potato (Ipomoea batatas L.) genotypes.Theor. Appl. Genet., 88: 332-336.
Costa, A.S. and M. Russell (1975). Failure of Bemisia tabaci to breed oncassava plants in Brazil (Homoptera: Aleyrodidae). Ciencia.Cultura., 27: 388-390.

47
Costa, H.S. and J.K. Brown (1990). Variability in biological characteristicisozyme patterns and virus transmission among population ofBemisia tabaci Genn. in Arizona. Phytopathology, 80: 888.
Costa, H.S. and J.K. Brown (1991). Variation in biological characteristics andesterase patterns among populations of Bemisia tabaci, and theassociation of one population with silverleaf symptom induction.Entomol. Exp. Appl., 61: 211-219.
Costa, H.S., J.K. Brown and D.N. Byrne (1991). Life history traits of thewhitefly, Bemisia tabaci (Homoptera:Aleyrodidea) on six virus-infected or healthy plant species. Environ. Entomol., 20: 1102-1107.
Dangi, R.S., L. Meena, L.B. Choudhari, P.K Ranjekar and V.S. Gupta (2004).Assessment of genetic diversity in Trigonella foenum-graecumand Trigonella caerulea using ISSR and RAPD markers. BMCPlant Bio., 47: 4-13.
Dawson, I.K., K.J. Chalmers, R. Waugh and W. Powell (1993). Detection andanalysis of genetic variation in Hordeum spontaneumpopulations from Isreal using RAPD markers. Mol. Ecol., 2: 151-159.
De Barro, P.J. and F. Driver (1997). Use of RAPD-PCR to distuinguish the Bbiotype from other biotypes of Bemisia tabaci (Gennadius)(Hemiptera:Aleyrodidae). Aust. J. Entomol., 36: 149-152.
De Barro, P.J., F. Drive, J.W. Trueman and J. Curran (2000). Phylogeneticrelationships of world populations of Bemisia tabaci(Gennadius) using ribosomal ITS1. Mol. Phylogenet. Evol., 16:29-36.
De Barro, P.J., J.W.H. Trueman and D.R. Frohlich (2005). Bemisia argentifoliiis a race of B. tabaci (Hemiptera: Aleyrodidae): the moleculargenetics differentiation of B. tabaci populations around theworld. Bull. Entomol. Res., 95: (3) 193-203.
De Barro, P.J., W. Liebregts and M. Carver (1998). Distribution and identity ofbiotypes of Bemisia tabaci (Gennadius) (Hemiptera:Aleyrodidae) in member countries of the sacretariat of thePacific Community. Aust. J. Entomol., 37: 214-218.
Delatte, H., B. Reynaud, M. Granier, L. Tharnary, J.M. Lett, R. Goldbach andM. Peterschmitt (2005). A silverleaf-inducing biotype Ms of B.tabaci (Hemiptera: Aleyrodidae) indigenous to the islands of theSouth-West Indian Ocean. Bull. Entomol. Res., 95(1): 29-35.
Dennehy, T.J., B.A. DeGain and V.S. Harpold (2008). Biotype designationsand Insecticide susceptibility of south-western Bemisia tabaci.Veg. Rep., 152.
Dinsdale, A., L. Cook, C. Riginos, Y.M. Buckley, P. De Barro (2010). Refinedglobal analysis of Bemisia tabaci (Hemiptera: Sternorrhyncha:Aleyrodoidea: Aleyrodidae) Mitochondrial cytochrome oxidase 1to identify species level genetic boundaries. Ann. Entomol. Soc.Am., 103: 196-208.
Dowell, R.V. (1990). Integrating biological control of whiteflies into cropmanagement systems. In D. Gerling, ed. Whiteflies, their

48
Bionomics, Pest Status and Management. Intercept, Andover,UK., 315-335.
Dudley, W., A. Sanghai-Maroof and G.K. Rufner (1992). Molecular markerinformation and selection of parents in corn breeding programs.,Crop Sci., 32: 301-304.
Fontes, F.V.H.M., C.A. Colombo and A.L. Lourencao (2010). Molecularcharacterization and genetic divergence of Bemisia tabaci(Genn.) (Hemiptera: Aleyrodidae) on different crops andgrowing areas. Neotrop. Entomol., 39: 221-226.
Friesen, V.L., B.C. Congdon, H.E. Walsh and T.P. Birt (1997). Intron variationin marbled murrelets detected using analysis of single-strandedconformational polymorphism. Mol. Eco., 6: 1047-1058.
Fritsch, P., M.A. Hanson, C.D. Spore, P.E. Pack and L.H. Risebeg (1993).Consistancy of RAPD primer amplification strength amongdistantly related taxon of flowering plants. Plant Mol. Biol Rep.,11: 10-20.
Frohlich, D.R., I. Torres-Jerez, I.D. Bedford, P.G. Markham and J.K. Brown(1999). A phylogeographical analysis of the Bemisia tabacispecies complex based on mitochondrial DNA markers. Mol.Entomol., 8: 1683-1691.
Fujiie, A., A.M.S. Omar, A.B. Sawas, A. Abbas, M.A. Hadi, E.A. Sawas, A.Barakat, S. Ueda and K.T. Natsuaki (2009). Geographicdistribution of Bemisia tabaci biotypes collected from autumn-cultured potato fields in Syria. J. ISSAAS., 15(2): 12-20.
Garagna, S., L. Rebecchi and A. Guidi (1996). Genome size variation inTardigrada. Zool. J. Linn. Soc., 116: 115-121.
Gawal, N.J. and A.C. Bartlett (1993). Characterization of differences betweenwhiteflies using RAPD-PCR. Insect. Mol. Biol., 2(1): 33 – 38.
Greathead, A.H. (1986). Host plants. In M.J.W. Cock, ed. Bemisia tabaci-Aliterature Survey on the Cotton Whitefly with an AnnotatedBibliography. CAB International Institutes, Biological Control.Silwood Park, UK. 17-26
Guirao, P., F. Beitia and J.L. Cenis (1997). Biotype determination of Spanishpopulations of Bemisia tabaci (Hemiptera: Aleyrodidae). Bull.Entomol. Res., 87: 587-593.
Gupta, V.K., R. Sharma, V. Jindal and V.K. Dilawari (2010). SCAR markersfor identification of host plant specificity in whitefly, Bemisiatabaci (Genn). Indian J. of Biotechnol., 9: 360-366.
Hameed, S., S. Hameed, M. Sadia and S.A. Malik (2012). Genetic diversityanalysis of Bemisia tabaci populations in Pakistan using RAPDmarkers. Mol. Biol. Genet., 15(6): 5.
Hasan, H.S. (2006). Survey of Bemisia tabaci (Gennadius) (Homoptera:Aleyrodidae) Biotypes in Japan using RAPD markers. J.Entomol., USA. 3(4): 290-297.
Haymer, D. (1994). Arbitrary (RAPD) primer sequences used in insectstudies. Insect Mol. Bio., 3(3): 191-194.
Helmi, A. (2010). Host plants-associated population variations of Bemisiatabaci (Genn.) (Hemiptera: Sternorrhyncha: Aleyrodidae)

49
characterized random DNA markers. Mun. Entomol. Zool., 5(2):677-683.
Hoddle, M.S., R.G.V. Driesche and J.P. Sanderson (1998). Biology and useof the whitefly parasitoid Encarsia formosa. Ann. Rev. Entomol.,43: 645-669.
Horowitz, A.R., I. Denholm, K. Garman, J.L. Cenis, S. Kontsedalov and I.Ishaaya (2003). Biotype Q of Bemisia tabaci Identified in Israel.Phytoparasitica, 31(1): 94-98.
Hsieh, C.H., C.H. Wang and C.C. Ko (2007). Evidence from molecularmarkers and population genetics analyses suggests recentinvasion of the western north pacific region by biotype B and Qof Bemisia tabaci (Gennadius). Environ. Entolmol., 36(4): 952-961.
Jena, K.K. (2000). DNA markers, polymorphism and applications in cropimprovement. In: Proc. Of delivered lectures during 2nd markersassisted selection MAS training programme. August 30-Sept.14.
Jones, D.R. (2003). Plant viruses transmitted by whiteflies. Eur. J. PlantPath., 109: 195-219.
Joshi, S.P., P.K. Ranjekar and V.S. Gupta (1999). Molecular markers in plantgenome analysis. Plant Molecular biology Group, Division ofBiochemical Sciences, National Chemical Laboratory, Pune.
Juen, A and M. Traugott (2006). Amplification facilitators and multiplex PCR:tools to overcome PCR-inhibition in DNA-gut-content analysis ofsoil-living invertebrates. Soil Biol. Biochem., 38: 1872-1879.
Kheinmeesook, P. (1997). Insect pests of vegetables. Entomology andZoology Division, Ministry of agriculture, Bangkok.
Lacey, L.A., J.J. Fransen, R. Carruthers (1996). Global distribution ofnaturally occurring fungi of Bemisia, their biologies and use asbiological control agents. In: D. Gerling, R.T Mayer, (Eds.),Bemisia 1995-Taxonomy, Biology, Damage, Control andManagement. Intercept, Andover, UK. 401-433.
Lander, E.S. and D. Botstein (1989). Mapping mendelian factors underlyingQTL using RFLP linkage maps. Genetics, 121: 185-199.
Lee, S.H., M.A. Bailey, M.A.R. Mian, T.E. Carter, D.A. Ashley, D.A. Hussey,W.A. Parrot and H.R. Boerma (1996). Molecular markersassociated with soybean lodging. Crop Sci., 36: 728-735.
Li, Z., D. Hu, Y. Song and Z. Shen (2005). Molecular differentiation of the Bbiotypes of Bemisia tabaci (Hemiptera: Aleyrodidae), based oninternally transcribed spacer 1 sequences. Eur. J. Entomol.,102: 293-297.
Lima, L.H.C., D. Navia, P.W. Inglis and M.R.V. de Oliveira (2000). Survey ofBemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biotypesin Brazil using RAPD markers. Genet. Mol. Biol., 23(4): 781-785.
Lima, L.H.C., L. Campos, M.C. Moretzsohn, D. Navia and M.R.V. de Oliveira(2002). Genetic diversity of Bemisia tabaci (Genn.) Populations

50
in Brazil revealed RAPD markers. Genet. Mol. Biol., 25(2): 217-223.
Lioyd, L.L. (1922). The control of the greenhouse whitefly (Asterochitonvaporariorum) with notes on its biology. Ann. Appl. Biol., 9: 1-32.
Litt, M. and A. Luty (1989). A hyper variable microsatellite revealed by in vitroamplification of a dinucleotide repeat within the cardiac muscleactin gene. Am. J. Human Genet., 44: 397-401.
Lourencao, A.L. and H. Nagai (1994). Outbreaks of Bemisia tabaci in the SaoPaulo State, Brazil. Bragantia 53: 53-59. (in Portuguese withabstract in English).
Ma, G.C., X. Wu, C.L. Ma, H.L. Wu, H.Y. Hu, L.M. Niu and Y.G. Fu (2011).Isolation and characterization of polymorphic microsatellite lociin Aleurodicus disperses (Hemiptera, Aleyrodidae). Genet. Mol.Res., 10(4): 2523-2526.
Markert, C.L. and F. Moller (1959). Multiple forms of enzymes: tissue,ontogenetic and species-specific patterns. Proc. Natl. Acad Sci.,45: 753-763.
Martin, J.H. (1985). The Whiteflies of New Guinea. Bull. British Mus. Nat.Hist., 50(3): 303-351.
Martin, J.H. (2004). Whiteflies of Belize (Hemiptera: Aleyrodidae). Part 1-Introduction and account of the subfamily AleurodicinaeQuaintance and Baker. Zootaxa., 3: 119.
Martin, J.H., D. Mifsud and C. Rapisarda (2000). The whiteflies (Hemiptera:Aleyrodidae) of Europe and the Mediterranean Basin. Bull.Entomol. Res., 90: 407-448.
Mckenzie, C.L., G. Hodges, L.S. Osborne, F.J. Byrne and R.G. Shatters(2009). Distribution of Bemisia tabaci (Hemiptera: Aleyrodidae)biotypes in Florida-investigating the Q Invasion. J. Econ.Entomol., 102(2): 670-676.
Merante, F., S. Raha and M. Ling (1998). Isolation of total cellular DNA fromtissues and cultured cells. In: Rapley R, Walker J, editors.Molecular biomethods handbook. Totawa: Human Press Inc.; 9-16.
Morgan, T.H. (1911). Random segregation versus coupling in mendelianInheritance. Sci., 34: 384-397.
Mound, L. and S. Halsey (1978). Whitefly of the World: A SystematicCatalogue of the Aleyrodidae (Homoptera) with Host Plant andNatural Enemy Data. British Museum Natural History, London.
Moya, A., P. Guirao, D. Cifuentes, F. Beitia and J.L. Cenis (2001). Geneticdiversity of Iberian populations of Bemisia tabaci (Hemiptera:Aleyrodidae) based on random amplified polymorphic DNA-polymerase chain reaction. Mol, Ecol, 10: 891-897.
Mugerwa, H., M.E.C. Rey, T. Alicai, E. Ateka, H. Atuncha, J. Ndunguru andP. Sseruwagi (2012). Genetic diversity and geographicdistribution of Bemisia tabaci (Gennadius) (Hemiptera:Aleyrodidae) genotypes associated with cassava in East Africa.Ecol. Evol., 2(11): 2749-2762.

51
Negi, M.S., A. Singh and M. Lakshmikumaran (2000). Genetic variation andrelationship among and within Withania species as revealed byAFLP markers. Genome, 43: 975-980.
Paterson, A.H., C.L. Brubaker and F. Wendel (1993). A rapid method forextraction of cotton (Gossypium spp.) genomic DNA suitable forRFLP or PCR analysis. Plant Mol. Res., 11: 122-127.
Perring, T.M. (2001). The Bemisia tabaci species complex. Crop Prot., 20:725-737.
Perring, T.M., R.J. Rodriguez, C.A. Farrar and T.S. Bellows Jr. (1993).Identification of whitefly species by genomic and behaviouralstudies. Sci., 259: 74-77.
Perumal, Y., M. Marimuthu, A.P. Salim and B. Ponnusamy (2009). Host PlantMediated Population Variations of Cotton Whitefly Bemisiatabaci Gennadius (Aleyrodidae: Homoptera) Characterized withRandom DNA Markers. Am. J. Biochem. Biotechnol., 5(1): 40-46.
Qiu, B., Y. Chen, L. Liu, W. Peng, X. Li, M.Z. Ahmed, V. Mathur, Y. Du and S.Ren. (2009). Identification of three major Bemisia tabacibiotypes in China based on morphological and DNApolymorphisms. Prog. Nat. Sci., 19: 713-718.
Rabello, A.R., P.R. Queiroz, K.C.C. Simoes, C.O. Hiragi, L.H.C. Lima, M.R.V.Oliveira and A. Mehta (2008). Diversity analysis of Bemisiatabaci biotypes: RAPD, PCR-RFLP and sequencing of the ITS1rDNA region. Genet. Mol. Biol., 31(2): 585-590.
Rafalski, A. and S. Tingey (1993). RFLP map of soybean (Glycine max)2N=40. In: O’Brien S. J. (ed.) Genetic maps: Locus maps ofcomplex genomes. Cold Spring Harbor Laboratory Press, NewYork, 6: 149-156.
Redi, C.A. and S. Garagna (1987). Cytochemical evaluation of the nuclearDNA content as a tool for taxonomical studies in eutardigradesIn: R. Bertolani (Ed.), Biology of Tardigrades. SelectedSymposia and Monographs, U.Z.I., 1. Mucchi Editore, Modena,Italy: 73-80.
Rekha, A.R., M.N. Maruthi, V. Muniyappa and J. Colvin (2005). Occurrenceof three genotype clusters of Bemisia tabaci and the rapidspread of the B biotype in South India. Entomol. Exp. Appl.,117: 221-233.
Rocha, K. C.G., J.M. Marubayashi, J.N. Castillo, V.A. Yuki, C.F. Wilcken,M.A. Pavan and R.K. Sakate (2011). Only the B biotype ofBemisia tabaci is present on vegetables in Sao Paulo State,Brazil. Sci. Agric. (Piracicaba, Braz.), 68(1): 120-123.
Rosell, R.C., I.D. Bedford, D.R. Frohlich, R.J. Gill and J.K. Brown (1997).Analysis of morphological variation in distinct populations ofBemisia tabaci (Homoptera: Aleyrodidae). Ann. Entomol. Soc.Am., 90: 575-589.
Sant, V.J., A.G. Patankar, N.D. Sarode, L.B. Mhase, M.N. Sainani, R.B.Deshmukh, P.K. Ranjekar and V.S. Gupta (1999). Potential ofDNA markers in detecting divergence and in analyzing

52
heterosis in Indian elite chickpea cultivars. Theor. Appl. Genet.,98: 1217.
Scott, I.A.W., P.J. Workman, G.M. Drayton, and G.M. Burnip (2007). Firstrecord of bemisia tabaci biotype Q in New Zealand. NewZealand Plant Prot., 60: 264-270.
Secker, A.E., I.A. Bedford, P.G. Markham, and M.E.C. William (1998).Squash, a reliable field indicator for the presence of B biotype oftobacco whitefly, Bemisia tabaci, 837-478.
Serrano, M.G., L.R. Nunes, M. Campaner, G.A. Buck, E.P. Camargo (1999).Trypanosomatidae: Phytomonas detection in plants andphytophagous insects by PCR amplification of a genus-specificsequence of the spliced leader gene. Exp. Parasitol., 91: 268-279.
Sharma, P.C., P. Winter, T. Bunger, B. Huttel, F. Weigand, K. Weising and G.Kahl (1995). Abundance and polymorphism of di-tri- and tetranucleotide tandem repeats in chickpea (Cicer arientinum L.).Theor. Appl. Genet., 90: 90-96.
Sharma, R.K., V.K. Gupta, J. Jindal and V.K. Dilawari (2008). Hostassociation genetic variations in whitefly, Bemisia tabaci(Genn.). Indian J. Biotechnol., 7: 366-370.
Shatters, R.G., C.A. Powell, L.M. Boykin, H. Liansheng and C.L. Mckenzie(2009). Improved DNA Barcoding Method for Bemisia tabaciand Related Aleyrodidae: Development of Universal andBemisia tabaci Biotype-Specific Mitochondrial CytochromeOxidase I Polymerase Chain Reaction Primers. J. Econ.Entomol., 102(2): 750-758.
Smelcerovic, A., V. Verma, M. Spiteller, S.M. Ahad, S.C. Puri, G.N. Qazi(2006). Phytochemical analysis and genetic characterization ofsix Hypericum Species from Serbia, Phytochemistry, 67: 171-178.
Smith, H.A., R. McSorley and J.A.S. Izaguirre (2001). Effect of intercroppingcommon bean with poor hosts and non-hosts on numbers ofimmature whiteflies (Homoptera: Aleyrodidae) in the SalamaValley, Guatemala. Environ. Entomol., 30: 89-100.
Staub, I., C. Lemme, S. Sondur, M.B. Morsidae and R. Manshardt (1993).Using randomly amplified polymorphism DNA for evaluatinggenetic relationship among papaya cultivars. Theor. Appl.Genet., 80: 697-701.
Stiles, J.I., C. Lemme, S. Sondur, M.B. Morsidi and R. Manshardt (1993).Using randomly amplified polymorph DNA for evaluating geneticrelationships among papaya cultivars. Theor. Appl. Genet., 85:697-701.
Tsagkarakoua, A., C.S. Tsigenopoulosa, K. Gormana, J. Lagnela and I.D.Bedforda (2007). Biotype status and genetic polymorphism ofthe whitefly Bemisia tabaci (Hemiptera: Aleyrodidae) in Greece:mitochondrial DNA and microsatellites. Bull. Entomol. Res.,97(1): 29-40.

53
Ueda, S. and J.K. Brown (2006). First report of the Q Biotype of Bemisiatabaci in Japan by Mitochondrial Cytochrome Oxidase Isequence analysis. Phytoparasitica, 34(4): 405-411.
Valley, G.E., A.L. Lourencao, M.I. Zucchi, J.B. Pinheiro and A.G. Abreu(2011). mtDNA variability in whitefly (Bemisia tabaci)populations in Brazil. Genet. Mol. Res., 10(3): 2155-2164.
Welsh and M. McClelland (1990). Fingerprinting genomes using PCR witharbitrary primers. Nucl. Acid Res., 18: 7213-7218.
Williams, J.G.K., A.R. Kubelik, K.J. Livak, J.A. Rafalski, and S.V. Tingey(1990). DNA polymorphisms amplified by arbitrary primers areuseful as genetic markers. Nucl. Acids Res., 18: 6531-6535.
Xiao-Jun, G., R. Qiong, Z. Fan, L. Chen, Z. Hong-yu and Gao Xi-wu (2012).Diversity and Genetic Differentiation of the Whitefly Bemisiatabaci Species Complex in China Based on mtCOI and cDNA-AFLP Analysis. J. Integ. Agric., 11(2): 206-214.
Zidani, S., A. Ferchichi and M. Chaieb (2005). Genomic DNA extractionmethod from pearl millet (Pennisetum glaucum) Leaves. Afr. J.Biotechnol., 4: 862-866.

Abstract
1. Title of the thesis : “Molecular Diversity Analysis ofWhitefly (Bemisia tabaci)Collected from Different Regionsof Madhya Pradesh”.
2. Student Name : Sumit Prakashrao Kale
3. Postal Address : Hatekar layout, Behind Gadgebaba
Vacchanalya, Near Kastkar Sankul,
Mothi Umeri, Akola-444001 (M.S)
4. Advisor Name : Dr. S. Tiwari
5. Address (Office) : Biotechnology Centre
J.N.K.V.V., Jabalpur (M.P.)
6. Degree awarded : M. Sc. (Ag.) Molecular Biology and
Biotechnology
7. Year of award of degree : 2013
8. Major subject : Molecular Biology and Biotechnology
9. Total number of pages in the thesis :
10. Number of words in the abstract :
Signature Signature Signature
Dr. S. Tiwari Dr. S. Tiwari Sumit Prakashrao Kale
Major Advisor Director Biotechnology Centre Student

ABSRACT
The present study entitles “Molecular diversity analysis of whitefly (Bemisia
tabaci) collected from different regions of Madhya Pradesh” was carried out with the
objectives to standardize genomic DNA isolation method and analyze the genetic
diversity of whitefly using DNA fingerprinting.
This study aimed to employ DNA based method to clarify the existing of biotype
among whitefly populations in East Madhya Pradesh, India. The genomic DNA of
whitefly collected from different geographical regions of East Madhya Pradesh was
isolated by standardize protocol (SDS + Proteinase K) produce intact band with
high molecular weight. The mtCOI primer was used for identification of B biotype.
Amplified products were resolved by electrophoresis on 1.2% agarose gel and
photographed under gel documentation system. Specific mtCOI primer CI-J-2195
and L2-N-3014 showed the presence of ~880bp bands which confirmed the
presence of B biotype in different geographical regions of East Madhya Pradesh.
All identified B biotypes were used for molecular diversity analysis. The DNA
samples were amplified with 5 RAPD primers in thermal cycler. Amplified products
were resolved by electrophoresis on 1.2% agarose gel and photographed under gel
documentation system. Four decamer primers amplified 8 RAPD marker loci. All 8
bands scored by RAPD markers found to be monomorphic. Average number of
bands per primer was 2.00. One RAPD primer is not amplified in all population. No
variation was found among whitefly samples collected from different geographical
regions of East Madhya Pradesh. Its seems that B. tabaci found in there region of
collection are of conserved type.

VITA
The author of this manuscript Sumit Kale S/o Shri Prakashrao Gulabrao Kale was
born on 23rd of March 1987 at Akola district of Akola (M.S). He passed him Higher
Secondary School Certificate Examination with 64.83% marks from Amravati Board.
He joined Shri Shivaji College of Agricultural Biotechnology, Amravati (M.S.) in the
year 2006 and successfully completed him B.Sc. (Ag.) Biotechnology degree in the year
2010 with an OGPA 7.2 out of 10.00 scales.
Subsequent to graduation, he joined M.sc (Ag). Molecular Biology and
Biotechnology at Biotechnology Centre, Jawaharlal Nehru Agriculture University,
Jabalpur (M.P.) in the year 2011. In partial fulfillment of Master’s Degree, he was
allotted a research problem entitled “Molecular Diversity Analysis of Whitefly (Bemisia
tabaci) Collected from Different Regions of Madhya Pradesh”. This was successfully
completed by him and presented in the form of this thesis. He completed M.Sc. (Ag.)
Molecular Biology and Biotechnology, Degree with an OGPA of 7.79 out of 10.00 scale.
Merit scholarship was awarded by university post graduation. He actively participated in
various college activities.
Mr. Sumit Prakashrao Kale
Hatekar Layout, Behind Gadgebaba Vachanalya,
Near Kastkar Sankul, Mothi Umeri,
Akola-444001 (M.S).
Email I.D: [email protected]



![UNITED STATES ENVIRONMENTAL PROTECTION AGENCY …Oct 24, 2016 · [Coleoptera: Scarabaeidae Image] [Myzus persicae Sulzer Image] [Bemisia tabaci Gennadius Image] [Hemiptera: Tingidae](https://img.pdfslide.us/doc/110x75/5fac2ba33ce6863dca33c554/united-states-environmental-protection-agency-oct-24-2016-coleoptera-scarabaeidae.jpg)

























