Embed Size (px)
Citation preview

J. Biosci., Vol. 15, Number 4, December 1990, pp. 261-269. © Printed in India. Molecular cloning and restriction enzyme analysis of a long repetitiveDNA sequence in rice
V. S. GUPTA*, M. S. DHAR, B. G. PATIL, G. S. NARVEKAR,S. R. RA WAT and P. K. RANJEKARDivision of Biochemical Sciences, National Chemical Laboratory, Pune 411 008, India MS received 11 November 1989; revised 8 October 1990 Abstract. Rice long repetitive DNA (9-20 kbp) reassociating at Cot 50 M.s was cloned in pBR325. Out of several recombinants (Camr Ampr Tets), only a few were selected randomly for further characterization. The insert size in all these clones was 3-4 kbp. Restriction enzyme analysis showed the absence of EcoRI and BclI sites, presence of a single PstI and PvuII site and multiple sites for AluI in 3 clones namely pRLl, pRL7 and pRL10. The BamHI-PstI fragment of about 0·4 kbp in the pRL7 insert DNA (pRL7-0·4 kbp) was subcloned in M13mpl8 and partially sequenced using Sanger’s dideoxynucleotide chain termination method. Dot matrix comparison of this sequence with rice rDNA sequences revealed low homology with the 25S rDNA sequence of rice, however, hybridisation did not indicate any homology. Keywords. Rice; long repetitive DNA; cloning; restriction enzyme analysis.
Introduction In recent years, a large number of repetitive DNA sequences have been cloned and characterized from a number of plant species. Their species-specific nature has been clearly demonstrated in closely as well as distantly related plants (Jelinek, 1982; Evans et al., 1983; Sorenson, 1984; Pental and Barnes, 1985; Sala et al., 1985; Metzlaff et al., 1986; Sonina et al., 1989). Also, a specific repetitive sequence has been used as marker to study phylogenetic relationships (Hallden et al., 1987). Besides cloning, a few repeat families have also been sequenced (Orgel and Crick, 1980; Deumling, 1981; Kato et αl., 1984), and these studies have revealed conservation/divergence of specific repeat elements.
Earlier, we have described the unique pattern of genome organization in rice wherein the repetitive DNA sequences are arranged as tandem arrays of length as high as 6·5 and 20 kbp (Gupta et al., 1981; Dhar et al., 1988). Furthermore, we have also shown the presence of 4 different types of repeat families in rice repetitive DNA. Out of these 4 families, the long repeats reassociating by Cot 50 M.s showed some homology to the long and short repeat sequences reassociating by Cot 0·1 M.s and to the short repeats of Cot 50 M.s itself. As a continuation of this study, we have carried out cloning and analysis of long repeats reassociating by Cot 50 M.s and these data are included in this report. *To whom all the correspondence should be addressed.
261

262 Gupta et al. Materials and methods Molecular cloning and sequencing of rice repetitive DNA The rice long repetitive DNA sequences reassociating at Cot 50 M.s (9–20 kbp) were isolated according to Dhar et al. (1988). These were then digested with BamHI and cloned in pBR325 (Prentki et al., 1981) and Escherichia coli DHI was used as the host. Furthermore, the 0·4 kbp BamHI-PstI fragment was subcloned in M13mpl8 and E. coli JM101 (Norrander et al., 1983; Yannisch-Perron et al., 1984) was used as the host for transformation.
The procedures followed for plasmid cloning were essentially according to Maniatis et al. (1982) and the steps involved in M13 cloning and sequencing were according to the New England Biolab’s Laboratory Manual. Isolation of recombinant plasmid DNA This was carried out by the rapid isolation method of Barnes (1977) and by alkali lysis method of Birnboim and Doly (1979) and the recombinant DNAs were used for further characterization. Isolation and characterization of template DNA for sequencing The recombinant (white) plaques were used for isolation of template DNA by the method of Schreier and Cortese (1979). The insert size was checked by gel electrophoresis as described by Messing (1983).
Finally, the cloned rice repetitive DNA was sequenced essentially according to Sanger et al. (1977).
The samples were loaded on 8% Polyacrylamide gel of thickness 0·6 mm,Electrophoresis was carried out at 2000 V. After electrophoresis, the gel with the glass plate was exposed to X-ray film for about 16 h at – 70°C.
Sequence analysis was carried out using SEQAID II programme of D. Jones which was a kind gift by Prof. S. Muthukrishnan, Kansas State University, Kansas, USA. Results and discussion Cloning of rice long repetitive DNA in pBR325 The S 1 nuclease resistant long repetitive DNA sequences (9-20 kbp) reassociating at Cot 50 M.s were used for cloning in pBR325. To check the digestibility of the DNA, it was treated with a few restriction enzymes such as EcoRI, SalI, PvuI, PstI and BamHI which have unique cloning sites in pBR325. Out of these enzymes, only BamHI showed the desired digestion in the range of 2-9 kbp and hence was used during cloning. The recombinant clones were selected by their sensitivity towards tetracycline and resistance towards chloramphenicol and ampicillin (Camr Ampr Tets). The presence of rice repetitive sequences in these clones was
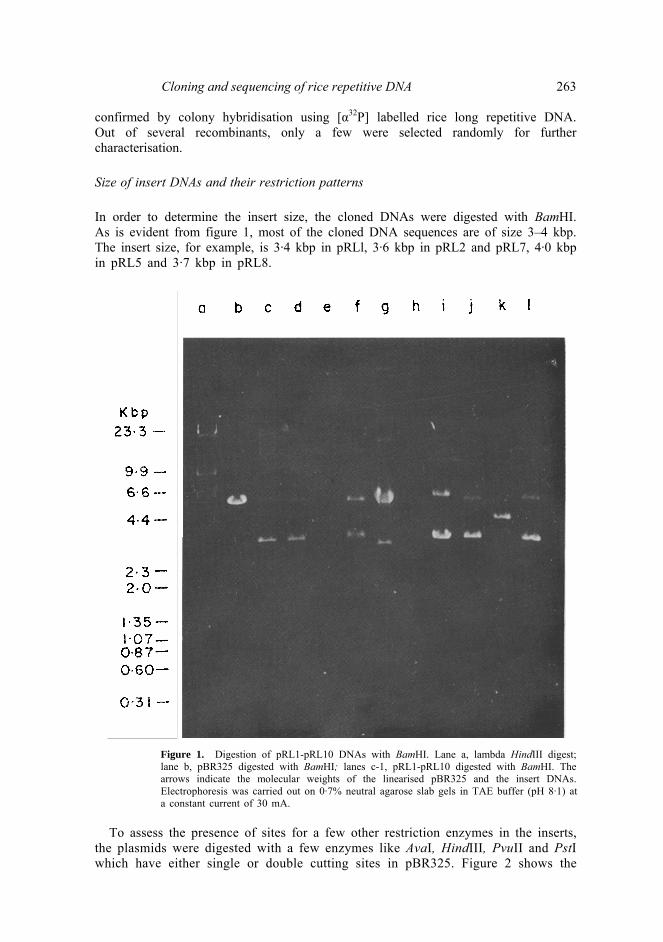
Cloning and sequencing of rice repetitive DNA 263 confirmed by colony hybridisation using [α32P] labelled rice long repetitive DNA. Out of several recombinants, only a few were selected randomly for further characterisation. Size of insert DNAs and their restriction patterns In order to determine the insert size, the cloned DNAs were digested with BamHI. As is evident from figure 1, most of the cloned DNA sequences are of size 3–4 kbp. The insert size, for example, is 3·4 kbp in pRLl, 3·6 kbp in pRL2 and pRL7, 4·0 kbp in pRL5 and 3·7 kbp in pRL8.
Figure 1. Digestion of pRL1-pRL10 DNAs with BamHI. Lane a, lambda HindIII digest; lane b, pBR325 digested with BamHI; lanes c-1, pRL1-pRL10 digested with BamHI. The arrows indicate the molecular weights of the linearised pBR325 and the insert DNAs. Electrophoresis was carried out on 0·7% neutral agarose slab gels in TAE buffer (pH 8·1) ata constant current of 30 mA.
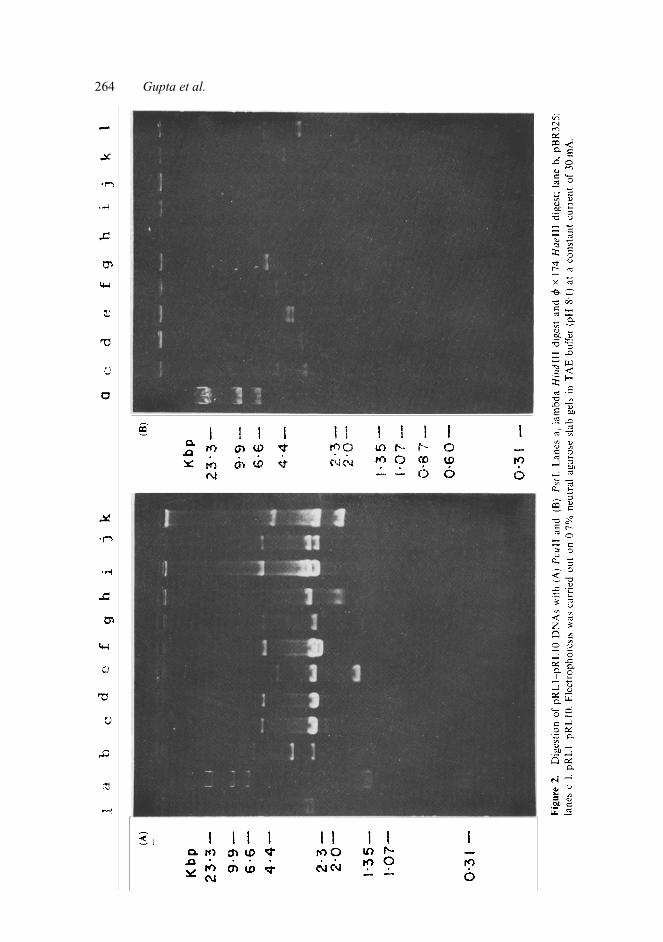
To assess the presence of sites for a few other restriction enzymes in the inserts,
the plasmids were digested with a few enzymes like AvaI, HindIII, PvuII and PstI which have either single or double cutting sites in pBR325. Figure 2 shows the
264 Gupta et al.
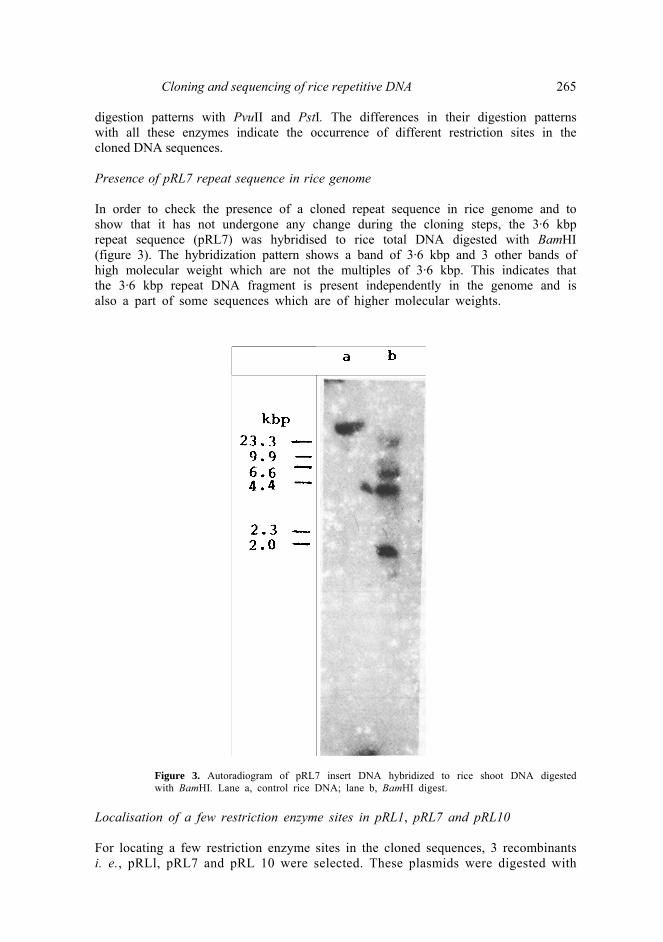
Cloning and sequencing of rice repetitive DNA 265 digestion patterns with PvuII and PstI. The differences in their digestion patterns with all these enzymes indicate the occurrence of different restriction sites in the cloned DNA sequences. Presence of pRL7 repeat sequence in rice genome In order to check the presence of a cloned repeat sequence in rice genome and to show that it has not undergone any change during the cloning steps, the 3·6 kbp repeat sequence (pRL7) was hybridised to rice total DNA digested with BamHI (figure 3). The hybridization pattern shows a band of 3·6 kbp and 3 other bands of high molecular weight which are not the multiples of 3·6 kbp. This indicates that the 3·6 kbp repeat DNA fragment is present independently in the genome and is also a part of some sequences which are of higher molecular weights.
Figure 3. Autoradiogram of pRL7 insert DNA hybridized to rice shoot DNA digested with BamHI. Lane a, control rice DNA; lane b, BamHI digest.
Localisation of a few restriction enzyme sites in pRL1, pRL7 and pRL10 For locating a few restriction enzyme sites in the cloned sequences, 3 recombinants i. e., pRLl, pRL7 and pRL 10 were selected. These plasmids were digested with

266 Gupta et al. BamHI and the corresponding inserts were eluted and used for further characterisation using a few enzymes like PvuII, EcoRI, PstI, BclI and AluI in single and double digestions. Since the strategy followed for the analysis of various restriction enzyme digestions was the same for all the 3 plasmids, a detailed analysis of only pRL7 is described. On digestion of pRL7 plasmid DNA with PvuII, 3 bands of 3·6, 3·4 and 2·6 kbp were observed which clearly revealed a PvuII site in the cloned DNA cutting it into two fragments of molecular weights 1·7 and 1·9 kbp. This was confirmed when the eluted insert DNA was digested with PvuII and two bands of 1·7 and 1·9 kbp were observed. In order to localise these positions, the pRL7 DNA was double digested with PvuII/EcoRI. Since, only the 3·4 kbp band in the PvuII digest of pRL7 was reduced to 3·3 kbp in PvuII/EcoRI double digest, it fixed the position of 1·7 kbp fragment in the insert DNA towards the EcoRI site in pBR325 followed by 1·9 kbp PvuII BamHI fragment. An appearance of 3·2 kbp band on PstI digestion of pRL7 insert DNA indicated the presence of a single PstI site at a distance of 0·4 kbp from the BamHI site in the insert DNA. To determine at which end the PstI site was present in pRL7 insert DNA, double digestion of pRL7 with BamHI/PstI was carried out. It showed two bands of 6·0 and 3·2 kbp and a faint band of 0·4 kbp which reflected the PstI site at a distance of 3·6 kbp from position no. 1 in pBR325. Finally, enzyme digestion of pRL7 insert DNA with AluI was analysed on 10% native, Polyacrylamide gel. AluI digest showed a number of bands in the range of 1·35 kbp to 0·068 kbp indicating a number of AluI sites in the cloned DNA sequence.
Based on these studies, it can be concluded that the rice repetitive DNAs cloned in pRL 1, pRL7 and pRL 10 contain no sites for EcoRI and BclI, single sites for PvuII and PstI and many sites for AluI (figure 4).
Figure 4. Line diagram showing the restriction enzyme sites in the cloned sequences in pRLl, pRL7 and pRL10 DNAs.

Cloning and sequencing of rice repetitive DNA 267 Subcloning of rice long repetitive DNA in M 13 After localising a few restriction enzyme sites in the pRLl, pRL7 and pRL10 insert DNAs, the next step was to determine the nucleotide sequence of these repeats. The 0·4 kbp fragment obtained after digestion of pRL7 insert DNA with ΒαmHI and PstI was selected for this purpose. This fragment was also observed to be present in pRLl and pRL10 insert DNAs. This fragment was then sequenced using Ml3cloning and sequencing system. Sequencing analysis of the 0·4 kbp repeat element (pRL7-0·4 kbp) using dideoxy chain termination method The 0·4 kbp BamHI-PstI fragment of pRL7 was sequenced by Sanger’s dideoxy nucleotide chain termination method. The partial sequence obtained from 5' → 3'end is given below.
The restriction enzyme site analysis of the pRL7-0·4 kbp sequence shows the
presence of single sites for some enzymes like FnudII, ECORII, ScrFI and BinI and many sites for enzymes like AluI, TaqI, MaeI and ΜnlI. In case of AluI, it is interesting to note that most of the fragment sizes indicated by computer analysis agree well with the molecular weights of a few bands observed in pRL7 insert DNA digested with AluI suggesting thereby that bands of 0·068, 0·072, 0·088 and 0·150 kbp in AluI digest may be a part of this BamHI-PstI fragment (pRL7- 0·4 kbp). Further analysis shows that this sequence has a G+C content of about53%.
Finally for comparing the pRL7-0·4 kbp sequence with the rice 25S (figure 5A), 17S rDNA (figure 5C) and the spacer fragment between 25S and 17S rDNAs(figure 5B) (Takaiwa et al., 1984, 1985a, b), dot matrix was generated using the pRL7-0·4 kbp sequence on Y-axis (Y sequence) and the known sequences on X-axis(X sequence) on the principle of Novotny (1982) and Jagadeeswaran and McGuire (1982). To reduce the background noise, it was found useful not to compare small fragments but 48 and 26 bases at a time. This eliminated most of the random fortuitous similarity while preserving the features of true homology. In the construction of dot matrix figure (figure 5), when the segment for the above length from both the sequences is equal to a minimal match of 20 or 15 bases, a dot is placed in the matrix at the coordination corresponding to the location of nucleotide in the sequence. The horizontal continuous dots show the homology between two
5’

268 Gupta et al.
Figure 5. Dot matrix comparison of the rice repetitive DNA sequence. The rice repetitive pRL7– 0·4 kbp sequence (Y-axis) is compared to the rice 25S(A), 17S rDNAs (C) and the spacer fragment between 25S and 17S (B) (X-axis). The window of comparison was 48 and 26 bases at a time with 50% identity required for a positive result to be recorded.
different sequences. Continuous lines parallel to a fixed angle, from top to bottom, suggest homology over a long stretch of sequences, while short lines indicate homology over a shorter stretch in discrete packets of sequences.
From figure 5, it is clear that the sequence pRL.7–0·4 kbp shows more homology with the nucleotide sequence of 25S rDNA fragment than with 17S rDNA and the spacer fragment. However, there are no homologies over a longer stretch of sequences in any of the 3 comparisons. It has already been reported that in rice rDNA fragment of 8·9 kbp, there is a Bam HI site cutting it into two fragments of 3·8 and 5·0 kbp (Reddy and Padayatty, 1987). Furthermore, this 3·8 kbp fragment is present in both the 25S and 17S rDNA genes. However, there was no hybridization of the probes with 3·6/0·4 kbp fragments (data not presented) indicating that this rice repeat unit is different from ribosomal DNA.

Cloning and sequencing of rice repetitive DNA 269 Acknowledgements One of the authors (M.S.D.) acknowledges Council of Scientific and Industrial Research, New Delhi for the award of a fellowship. G. S. N. and S. R. R. are the receipients of the fellowship from Hindustan Lever Research Foundation, Bombay. References Barnes, W. M. (1977) Science, 195, 393. Birnboim, H. C. and Doly, J. (1979) Nucleic Acids Res., 7, 1513. Deumling B, (1981) Proc. Natl. Acad. Sci. USA, 78, 338. Dhar, Μ. S., Dabak, M. M., Gupta, V. S. and Ranjekar, P. K. (1988) Plant Sci, 55, 43. Evans. I. J., James, A. M. and Barnes, S. R. (1983) J. Mol. Biol, 170, 803. Gupta, V. S., Gadre, S. R. and Ranjekar, P. Κ. (1981) Biochim. Biophys. Acta, 656, 147. Hallden, C., Bryngelsson, T., Sall, Τ. and Gustafsson, Μ. (1987) J. Mol. Evol, 25, 318. Jagadeeswaran, P. and McGuire, P. Μ. (1982) Nucleic Acids Res., 10, 433. Jelinek, W. R. (1982) Annu. Rev. Biochem., 51, 813. Kato, Α., Yakura, K. and Tanifuji, S. (1984) Nucleic Acids Res., 12, 6415. Maniatis, T., Fritsch, E. F. and Sambrook, J. (1982) Molecular cloning: A laboratory manual (New York: Cold Spring Harbor University). Messing, J. (1983) Methods Enzymol. 101C, 20. Melziaff, M., Troebner, W., Baidauf, F., Schlegel, R. and Cullum, J. (1986) Theor. Appl. Genet., 72, 207. Norrander, J., Kempe; T. and Messing, J. (1983) Gene, 26, 101. Novotny, J. (1982) Nucleic Acids Res., 10, 127. Orgel, L. E. and Crick, F. H. U. (1980) Nature (London), 284, 604. Pental, D, and Barnes, S. R. (1985) Theor. Appl. Genet., 70, 185. Prentki, P., Karsch, F., Iida, S. and Meyer, J. (1981) Gene, 14, 298. Reddy, P. S. and Padayatty, J. D. (1987) Indian J. Biochem. Biophys., 24, 293. Sala, C., Biasini, M. G., Morandi, C, Nielson, E., Parisi, B. and Sala, F. (1985) J. Plant Physiol, 118, 409. Sanger, F., Nicklen, S. and Coulson, A. R. (1977) Proc. Natl. Acad. Sei. USA, 74, 5463. Schreier, Ρ. Η. and Cortese, R. (1979).J. Mol. Biol., 129, 169. Sonina, Ν. V., Lushnikova, Α. Α., Tihonov, A. P. and Ananiev, E. V. (1989) Theor. Appl. Genet., 78, 589. Sorenson, J. C. (1984) Adv. Genet., 22, 109. Takaiwa, F., Oono, K. and Sugiura, M. (1984) Nucleic Acids Res., 12, 5441. Takaiwa, F., Oono, K., Iida, Y. and Sugiura, Μ. (1985a) Gene, 57, 255. Takaiwa, F., Oono, Κ. and Sugiura, Μ. (1985b) Plant Mol. Biol., 4, 355. Yannisch-Perron, C., Vieira, J. and Messing, J. (1984) Gene, 33, 103.