Embed Size (px)
Citation preview

Vol. 181. No. 2. 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
December 16, 1991 Pages 707-713
MOLECULARCLONINGANDNU~TIDESEQ~CEOF SmmCES GRISEUS TRYPSIN GENE
Jee Cheon Kim, Seung Hee Cha*, Seong Tae Jeong, San Kon Oh* and Si Myung Byun’
Department of Life Science, Korea Advanced Institute of Science and Technology 373-l Kuseong-Dong, Yuseong-Gu, Taejon, Korea
*Agency for Defense Development, P.O. Box 35 Yuseong, Taejon, Korea
Received November 5, 1991
SUMMARY Streptomymgriseus trypsin (E.G. 3.4.21.4) isoneof themajorextra&l~protei.nase, which is secreki by S. @cur. The gene encoding S. grirem trypsin was isolated from a S. grisew genomic library by using a synthetic oligonuckotide probe. Fragments containing the gene for S. griseus ttypsin were characterized by hybridization and demonstration of proteolytic activity in S. lividan. Deduced amino acid sequence from the nucleotide sequence suggests that S . griseus hypsin is produced as a precursor, consisting of three portions; an aminoterminal pre sequence (32 amino acid residues), a pro sequence (4 residues), and the matute trypsin. wrth a computed molecular weight of 23,112.
The S. griseus trypsin consists of 223 amino acids The existence of proline at the pro and mature junction
suggests that the processing of S. grirezkr trypsin is non-autocatalytic. 0 1991 Academic Press, Inc.
Stmplmnyces griseus, an organism used for the commercial production of pronase, sect&s many extracellular proteins (14). Srregzovnyces grireus trypsin (SGT) is a bacterial serine proteinase that, curiously, is more similar to mammalian proteinase than to S. griseus protease A and B, two serine proteinases from the same bacterium (25). The general similarity of the substrate binding regions of SGT andbovinetrypsin(BT)isconsistentwi~Ihe~~oftheitinteractionswithsubstratesandinfiibitors. SGT cleaves the oxidized B chain of insulin in the same manner as BT, as well as the synthetic substrate N-&enzoyl-kuginmeethylester(20). It~bepurifisdonthesameaffulitycolumnasBT(28). Also, it is inhiiited by the same pmkinase inhibitors (19). Although the structure of SGT has been extensively
studied, the gene encoding SGT has not been characterized. This report describes the structure of a S. griseus gene which is responsible for the exptession of
SGT. The nucleotide sequence suggests that proteinase is initially secmted as a pmcursor which is p~toremoveaveryshort~~peptide@ropeptide)fromthematureproteinase. We propose the genetic designation SprT for the unmapped gene for SGT.
‘To whom unrespondence should be addressed. . . Abbrevta&ns used are : SOT, Sfnzptmyces g&em trypti, BT, bovine trypsin; kbp, kilobase
pair(s); SDS, sodium dodecyl sulfate, PAGE, polyacrylamide electrophoresis.
OCO6-291x/91 $1.50
707 Copyright 0 1991 by Academic Press, inc.
All rights of reproduction in any form reserved.

Vol. 181, No. 2, 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
Strains andPlasm& S. grkeur A’ICC 10137, S. lividans TK24, andpIJ702 (12) wereobtained from theKoreanCollech’onfortheTypeCultures,GeneticEngineesingCenter,~~Insti~~ofScienceand Tcchnolog , Taejon, Korea. EFcherichiacoli JM107orIM109 wereused foralltransformation. pUC18 and pUC1 B werepurchased from Bethesda Besearch Labomtories, Inc. Media, Growth, and Transformation: Growth of Smpmnyces mycelium for the isolation of DNA or the preparation of protoplast has been described previously (12). Transformants were screened for proteol 2 P %(w
‘c activity on YD plates (5) containing 30 pg of thiostrepton per ml, 0.5 %(w/v) glycine, and v) skim nnlk. Competent cells of E, coli strains for tmnsformation were prepared as described
by Hanahan (7). E. coli transfommnts were grown on LB plates (26) containing 100 pg ampicillin. Materiakz Oligonucleotides were synthesized by the solid phase phosphoramidate methods with a Beckman system I plus DNA syntheskr. Enzymes for digesting and modifying DNA were purchased from KGSCO Biotech (Seongnam, Korea) and used in W with the recommendations of the supplier. The radioisotopes [&%i]dATP and [T-~*P]ATP were from Amersham International. Consauction of Genomic Library: Chromosomal DNA of S. g&em ATCC 10137, prepared as descrii previously (12), was digested to completion with ECORI-Hind III and tiactionated by electrophoresis on a 0.8 96 agarose gel. DNA fragments in sire 6.8 kilobase pairs (kbp) were isolated from the agarose gel. The plasmid vector pUC18 and pUC19 were digested with &oRI-Hind III and tmated with calf intestinal alkaline phosphatase @o&ringer Mannheim Biochemicals). The S. grisem DNA fragments (0.5 pg) and vectors (0.2 pg) were seqmtially ligated in a tinal volume of 50 d as described previously (26). Approximately 7,500 transformants of E. coli IMlCt7 were obtained from each ligation reaction. Subcloniug of Proteinase Gene Fragments: A hybrid Smptomyms - E. coli vector was constructed by ligating pII702, which had been linearized with Sac I, into the Sue I site of pUCl8. The resultant vector, pUI718-2, was capable of replicating in S. liridan and E. coli. The multiple cloning sites of this vector, originated from pUC18, were used for subchming DNA fragments of the proteinase gene. Gther fragments were adapted parually with ECORI linker to tkcilitate ligahon into the ECORI site. Hybridization: A 23mer oligonucleotide (5’AAC GC(GC) GAC GAG TGG ATC CAG GT- 3’) was designed from an amino sequence (NADEWIQV) of SGT. For use as a hybridization probe, the oligonucleotide was end-labeled by T4 polynuclcotide kinase and [-$*P]ATP. Digested genomic or plasmid DNA was elecuophorescd and directly hybridized (24). The S. griseur genomic library was screened by colony hybridization as described previously (26). DNA Sequencing: DNA sequencing was performed by the dideoxy-chain termination method (27) using Sequenase(U.S.BiochemicaIs). SubclonesforsequencingwerepreparedinthepUC19orpUC18,and the dideoxy sequencing reaction were run by 40 forward or reverse primer (New England Biolabs). To reduce compressions, sequencing reactions were carried out 42 “C with dlTP substituted for dGTP or C residues were modified with methoxylamine and sodium bisulfite after the sequencing reaction (1). Also, in some experiments, 10 % formamide (10) was added to 6 96 polyacrylamide7M urea gel.
!Screening for Trypsii Gene by Hybridization: An oligonucleotide probe was designed from one of the SGT amino acid sequences by using the known codon bias for Streptmycces species (9). ‘Ihe utility of the nucleotide probe was demonstrated by hybridization to genomic DNA of S. griseus. As anti- cipated, the probe hybridized to a fragment generated by EkoRI-Hind IlI (6.8 kbp), but no such hybridization was observed with DNA from S. Zitidans. By using the oligonucleotide probe, plasmids containing spfl were isolated from agenomic DNA librsry prepamd from S. gliseur DNA. Gf4,500 E. coli transformants that were screened by colony blot hybridization, 9 were detected by oligonucleotide probe and isolated for tkther characterization. These colonies contained single class of plasmids, based on restriction analysis. As expected, based on the hybridizationofgenomic DNA, the plasmids contained a 6.8 kbp EcoRI-Hind III fmgment. CharacterizationofTkypsinGene: TheDNAf@mentsisolatedbyhybridirationscreeningweretested forexpressionofproteolyticactivity. The 6.8kbp EcoRI-Hind IIIliagmentswereligatedintothe ECORI- HindID site of thevector pUI718-2, to allow transformation of S. hi&m, with selection forthiostrepton

Vol. 181, No. 2, 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
I J
Restriction endonuclease maps and sequencing strategy for the 1.8 kbp BglI fragment Fig. 1. contamlng spfl. (A) The thick line indicates the minimum restriction fragment capable of hybridizing to the oligonucleotide probe. (B) The arrows indicate the direction and length of the sequence determmed by the dideoxy chain termination methods. ding region.
The thick line indicates the spfl The organization of the structural gene, with pre-pro peptide (36 amino acid,-)
and mature proteinase (223 amino acids, q ), is shown below map. Abbreviations: Bm, BomKI; BgI, BglI; BgII, Bgm, EC, EcoRI; Hd, HWIII; Kp, &@I; Pv, ⅈ Sa, SolI; SC, SacI; Sm, SmuI.
red.ance. Transformants containing this construction were then tested on a milk plate containing 0.5 5% glycine for secretion of protekse. A clear zone, although of very small size, representing the degmdation of milk proteins, surrounded each transformant that contained &oRI-Hind III fragment. The clear zones were not found around S. fividans coIonies which contained either pIJ7U2 only or no plasmid. The intact proteinase gene could be delimited to a 1.8 kbp Bgl I fragment. This functionally active subclone contained the region which hybridized to the nuckotide probe (Fig. 1.).
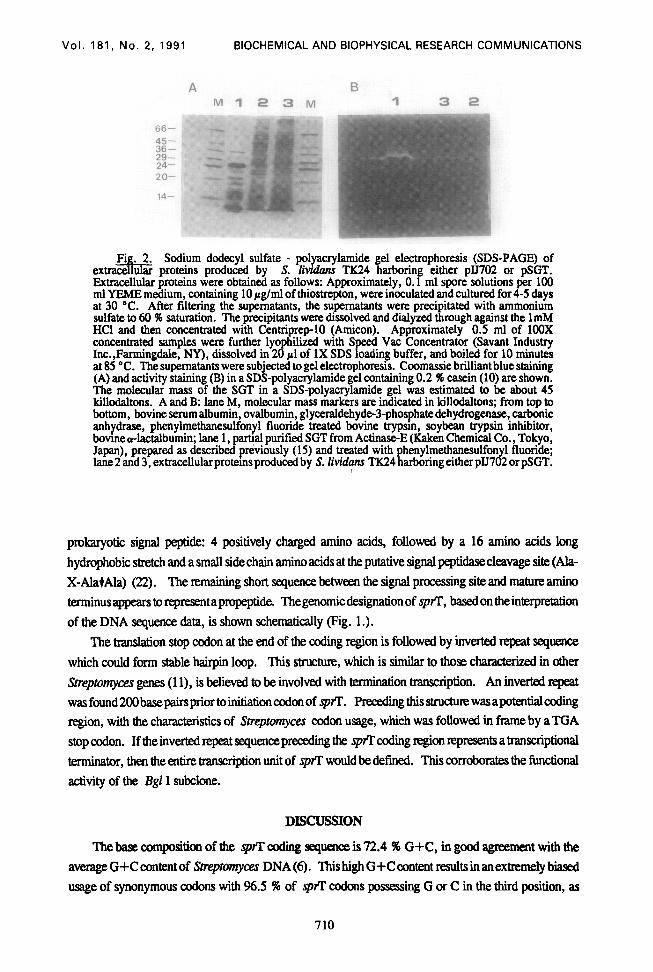
To identify the protein correqonding to SGT in 5. lividan, the activity staining of proteinase was pedormed (Fig. 2.). The proteinase activity was found only in culture supematant of S. Zivih TIC24
harboringpSGTwhichispUJ718-2derivativescontaininga1.8kbpBgZIfkagment. Thesodiumdodecyl sulMepolyaqlamidegelelectrophoresis (SDS-PAGE) of extmceklarproteins produced by 5. lividans TIC24 harboring pSGT showed that the protein responsible for the activity had the same molecular weight as the reference SGT purified from ActinaseE. This result was consistent with the result of SDS-PAGE of immunopmcipitants between rabbit anti-SGT immunoglobulin G and extracellular proteins (Data not shown). DNA !kquence of Trypsin Gene: The sequencing strategy for the spfl is shown (Fig. 1.). Analysis of the sequence revealed an open reading frame (nucleotide 525 - 1304), encoding 259 amino acids, that is preceded by a ribosomal binding site (GAAAGAAGG), which is complementary to a squence close to th~3’ end of the 16s rRNA of S. grireur (17), assumed to be the spfl coding sequence (Fig. 3.). The predicted sequence of SGT differed from the published amino acid sequence (21). It should be corrected by the insertion of two serine residues near position 76 (Ser) according to the numbering ofa- chymohypsin (25). Four residues, Gln74-Ser75-Ser76-Ser77 in the current sequence, were proposed initially as Gln75-Ser76Gly77Ala79 (25). The N-termimd 32 amino acids resemble a typical
709
Vol. 181, No. 2, 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
A B M123M 1 3 2
Fig. 2. Sodium dodecyl sulfate - polyacrylamide gel electrophoresis (SDS-PAGE) of extracellular proteins produced by 5. Zivrdans TX24 harboring either pJJ702 or pSGT. Extracellular proteins were obtained as follows: Approximately, 0.1 ml spore solutions per 100 ml YEME medium, containing 10 pglml of thiostrepton, were inoculated and cultured for 4-5 days at 30 “C. After filtering the supematants, the supematants were precipitated with ammonium sulfate to 60 96 saturation. The precipitants were dissolved and diiyxed through against the 1mM HCl and then concentrated with Centriprep-10 (Amicon). Approximately 0.5 ml of 100X concentrated samples were further lyophilixed with Speed Vat Concentrator (Savant Industry Inc.,Farmingdale, NY), dissolved in 20 pl of 1X SDS loading buffer, and boiled for 10 minutes at 85 “C. The supematants were subjected to gel electrophoresis. Coomassie brilliant blue staining (A) and activity staining (B) in a SDS-polyacrylamide gel containing 0.2 A casein (10) are shown. The molecular mass of the SGT in a SDS-polyacrylamide gel was estimated to be about 45 killodaltons. A and B: lane M, molecular mass markers are indicated in killodaltons; from top to bottom, bovine serum albumin, ovalbumm, glyceraldehyde-3-phos@ate dehydrogenase,. carbonic anhydrase, phenylmethanesulfonyl fluonde treated bovme trypsm, soybean trypsm mmbitor, bovmec&ctalbumin; lane 1, partial purified SGT from Actinase-E (Kalcen Chemical Co., Tokyo, Japan), prepared as described previously (15) and treated with phenylmethanesulfonyl fluoride; lane2 and 3, extracellularprotemsproduced by S. lividu~ TK24 harboring eitherplJ702 orpSGT.
prolcqotic signal peptkk 4 positively charged amino acids, followed by a 16 amino acids long hydrophobic stretch and a small side chain amino acids at the putative signal @&se cleavage site (Ala-
X-Al&Q) (22). The remaining short sequence between the signal prwzssing site and mature amino tertninusappearstorepresentapqeptide. Thegenomicdesignationof spfl, basedontheinterpretation
of the DNA sequence data, is shown schematically (Fig. 1.).
The translation stop uxion at the end of the coding region is followed by inverted repeat sequence
which could form stable hairpin loop. This structure, which is similar to those characterized in other Streptomyces genes (1 l), is believed to be involved with termination transcription. An inverted repeat
wasfound2oObasepairspriortoinitiationcodonof SprT. Pre&ingt.hisstructtnewasapotentialcoding
region, with the characteristics of Sfrepmnyces codon usage, which was followed in frame by a TGA
stop coclon. If the inverted repeat sequence preceding the SpJT coding region represents a transcriptionaI
terminator, then the entire transcription unit of spr’I would be defined. This corroborates the functional
activity of the Bgl I subclone.
DISCUSSION
The base composition of the qfl coding sequence is 72.4 96 G+C, in good agreement with the avemgeG+Ccontentof Sfrepfonyces DNA@). ThishighG+Ccontentresultsinanextremelybiased usage of synonymous codons with 96.5 46 of spfl codons possessing G or C in the third position, as
710

Vol. 181, No. 2, 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
1 CCCCOOCATCOCCCCCCCCACCCOCAACATCTACCCCCCCT 91 COCCCCCCCOOCCCGCCGCGAACTCCGCCCCCCCCCCACC 181 CTGCAGCTCCTACCGGOCCTCCCG~C~CCCTCCGGCA 271 -20.0 GccccccTCCccC~TAC~ACA~A~TCC~GCCGTA~T~TTCCG~C~~CGTCCC~GTCCG~TGACC~ 361 CGTCCTGGACGGGCCGTOGTOOCCCOTACCACCTTGGCTT~CCG~TGTGCGCCTTGT~CAG~C~CC~TTCCCGA~T~~G~TC 451
TGCATGACCATOCCGTCGCGCCCCGTCGGGOTTCCCACAGCGAC~CCC~C~~GM~CMTC~TGMG~~TC~C -10 RBS MKHFLR
541 GTGCGCTOAAGAGATGCTCCCTCGCCGT~C~CCGTffiC~T~~GT~T~CCTC~GCC~T~C~CCTC~C~CCCC~CC
ALKRCSVAVATVAIAVVGLQPVTASA+APNPt 631 CCGTCGTCCCCOCMCCCCCCCCGCCCACGGCGAGTTCCCCTT~TGGTC~GCTCTC~T~CTG~CGG~CCCTCTA~CC~~
VVGGTRAAQGEFPPHVRLSHG CGGALYAQD 721 ACATCGTCCTCACCGCGGCCCACTGCGTGAGCGGATCGGC
IVLTAAHCVSGSGNNTSITATGGVVDLQSS 811 CCAGCGCCGTCAAGGTCCGCTCCACCAAGGTCCTCCAGGCCCC
SAVKVRSTKVLQAPGYNGTGKDWALIKLAQ 901 AGCCCATCMCCAGCCCACGCTGMGATCGC~C~C~CCGC~A~C~~~~TTCACCGT~C~~~~C~C~~
PINQPTLKIATTTAYNQGTFTVAGWGANRE 991 AGGGCGGCAGCCAOCAGCGCTACCTGCTCAAGGCCMCGTCCCGTTCGTCTCCGA~CCGC~GC~CTC~CGTAC~CMCGAG~~
GGSQQRYLLKANVPPVSDAACRSAYGNELV 1081 TGGCCMCGAGGAGATCTGCGC~GACACCGG
ANEEICAGYPDTGGVDTCQGD SGGPMFRKD 1171 AC~CGCCGACGAGTGGATCC~TCCTCACCTCGC
NADEWIQVGIVSWGYGCARPGYPGVYTEVS 1261 CGACCTTCCCTTCCGCCATCGCCTCGCCCGCCGCCCGCA~GCTC~A~CA~TAC~CACCC~CCGCTCC~GGTGAT~CCCCTGATC
TFASAIASAARTL* 135j -32.0 ACCGCCGCCOCCCGTCCCTCTTCTCCGGCCCCCCCCCGCCTGCT 1441 GCGCOTGGACCGGGTGGTAGTCACCGAACTCGACGOATG 1531 AGCCGGGGCCGACCCCGCCCGCCATGGCCCCCTCGTCGCCG~TCGT~C~AC~ACT~CTG~CGTGAGCGT~TGACGCCCTCGTGCT~A~AG~ 1621 TGAGGGTCATGTGCCAGGCGMCTCCCCGGGCC~GGCACC~TGA~TGA~ACGAGACGACGAGGGGCCCGGCA~CCTCGA~TTG~ 1711 GCTCGGCCTGGTGCTCGTCGCCCTCGTAGAGCATCCGGAAC 1801 TTCCGGGCCOCCATCGAGCACGTCTGOCCGGCGGTCACCG
1892
Fig. Nucleotide sequence of sprT. (GenBanlcaccession No. M64471) Thededuced amino acid sequence, which is numbered from the termini of the mature proteinase, is shown under the nucleotide sequence. Stop codon is indicated by asterisk. The processing sites for the signal peptide (downward arrow) and the mature proteinase (upward arrow) are shown. Bases involved with the putative +osome binding site (RF%) are indicated by underline. are overhned.
The inverted repeat sequences The free.energy changes (kilocalories per mole) of the hypothetical hair@ loops
were caktdated as desertbed previously (3 1). otide probe is double underlined.
DNA sequence which hybridized to the ohgonucle- The putative -35 and -10 regions, which show similarity to the
consensus sequence of Streptontyces species (13), are indicated by underline.
has been cbervd in other Sfnzpturnyca genes (10). Also noticeable is a geneml and pronounced
preference (129:28) for a third position C over G where synonymous axtons allow such a choice. In
comparison,thebasecompositionoutddeoftheproposedopenreadingfrsunesisuniformlydistributed
with each tmckotide triplet. ‘IIre maintermnce of a biased codon usage throughout each coding region
711

Vol. 181, No. 2, 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
and -02 of the triplet perk&city outside of the reading frames strengthens the assignment of initiation and termination codons (3).
Recent studies with secreted proteinazs from both gram-positive and gmm-negative bacteria, in&ding Bacilhs species (16,32), Lysobactor enzymogens (29), Neisseria gonorrhoeae (23), Sermtia (18),Strepcomycesspe~ies(4,9),Vibriocholera(8), and ZJre~wagu#i~(3O),haveshownthatallof
these bacterial proteinase are synthesized as precursors, although the pro region varies in its amino or carboxyl terminal location. All of these precursors contain extra signal sequences and pro sequences, except Semua protease (18) which lacks such signal sequence. The processing of each precursor proteinase is probably at&catalytic, since the amino acid sequence near the scissile bond should provide agoodsubstrate. TheputativeprosequenceofspflisAla-Pro-Asn-Pro,whichisveryshortascompared
to other bacterial prosequences. The p mcessing of SGT precursor is probably non-autocatalytic, since the amino acid sequence near the pro and mature junction is not a good substrate of SGT itself. Another proteinase, one of the component of pronasc, is probably involved in proessing of SGT. In practice, pronase contains the component that hydrolyzes an artificial substrate, L-plhe-~nitroanilide (0.6
increase of optical density at 420 nm per mg of pronase within 30 minutes).
The function of the propeptide is still unclear. It may play an essential role in guiding the folding of proproteinase molecule into the proper conformation necessary for activity such as has been observed with subtilisin (32) and +tic protease (29). In case of spfl, the pro sequence may not play a role in guiding the folding of pro enzyme because of its small size, but probably play a function to keep the
precursor inactive, as is the case with bovine trypsinogen whose pro sequence is Val-Asp-AspAsp-Asp Lys. The non-autocatalytic junction of the precursor and Longisporus trypsii inhibitor that is secreted by S. Zivihns 66 (2) maybe responsible for the low proteinase activity in culture supernatant of S. lividanr TK24 harboring pSGT. Further investigation will be directed toward unraveling the activation mechanism of the proteinase precursor and elucidating the exact function of the pro sequence.
REFERENCES
1. Ambartsumyan, N.S. and Mazo, A.M. (1980) FEBS L&t. 114,265-268. 2. Be&a, T.R., Fomwald, J.A., Gorniak, J.G., Rosenberg, M., Stickler, J.E., and Taylor, D.P.
(1988) PCl- Int. Appl. WO 88 01,278. 3. Bibb, M.J., Fmdlay, P.R., and Johnson, M.W. (1984) Gene 30, 157-166. 4. Chang, P.C., Kuo, T.-C., Tsugita, A., and Lee, Y .-H.W. (1990) Gene 88,87-95 5. Foor, F., Roberts, G.R., Morin, N., Snyder, L., Hwang., M., Gibbons, P.M., Pamdiso,
M.J., Stotish, R.L., Ruby, C.L., Wolanski, B., and Suer&r, S.L. (1985) Gene 39, 11-16. 6. Gladek, A. and Zakrzewska, J. (1984) FEMS Micmbiol. Lett. 24,73-76. 7. Hanahan, D. (1983) J. Mol. Biol. 166, 557-580. 8. H&e, C.C. andFinkelstein, R.A. (1991) J. Bacterial. 173,3311-3317. 9. Henderson. G.. Krvgsman, P., Liu, C.J., Darvey, C.C., and Malek, L.T. (1987) J. Bacterial.
12.
13.
169,3778-3784. -- Hensen, C. and Dowdle, E.B. (1980) Anal. B&hem. 102,196202. Hopwood, D.A., Bibb, M.J., Charter, K.F,, Janseen, G.R., Ma&u%%, F., and Smith, C.P. (1986) In Regulation of Gene Expression - 25 Years Gn (Booth, L.R. and Higgins, C.F. Eds.), pp. 251-276. Cambridge University Press, Cambridge. H wood, D.A.,Bibb, M.J., Charter, K.F.,Kieser,T.,Bruton, C.J., Kieser, H.M., L . & , D.J., Smith, C.P., Ward, J.M., and Schrempf, H. (1985) Genetic Manipulation of Strepwmym: A Laboratory Manual. The John Innes Foundation, Norwrch. Horii, H.. Ishizaki, T., Pa&, S.-Y., Manome, T., and Murooka, Y. (1990) J. Bact&i01: 172,36443653.
14. Jur&k. L,, Jhonson, P., Olafson, R.W., and Smillie, L.B. (1971) Can. J. Biochem. 49,548-56%
712

Vol. 181, No. 2, 1991 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
15. Ju&ek, L., Jhonson, P., Olafson, R.W., and Smillie, L.B. (1971) Can. J. B&hem. 49, 1195-1201.
16. Kaneko, R., Koyama, N., Tsai, Y.C., Juang, R.-Y., Yoda, K., and Yamasaki, M. (1989) J. Bacterial. 171,5232-5236.
17. Kim,E.,Kim,H.,Kang,K.H,,Kho, Y.H.,andPark,Y.-H. (1991) NucleicAcidsRes. D, 1149. 18. Nakahama, K., Yoshimum, K., Marumoto, R., Kikuchi, M., Lee, L.S., Has, T., and
Mat&ma, H. (1986) Nucleic Acids Res. 14,5843-5855. 19. Nagata, K. and Yoshida, N. (1983) J. Biochem. (Tokyo) 93,909-919. 20. Olafson, R.W., Jurdsek, L., Carpenter, M.R., and Smillie, L.B. (1975) Biochemistry 14,
1168- 1177. 21. Olafson, R.W. and Smillie, L.B. (1975) Biochemistry 14,1161-1167. 22. PerlmaqD. and Halvorson, H.O. (1983) J. Mol. Biol. 200, 523-551. 23. Pokier, J., Halter, R., Beyreuther, K., and Meyer, T.F. (1987) Nature 325,458-462. 24. Purrcllo, M. and Balaq L. (1983) Anal. Biochem. l28,393-397. 25. Read, R.J. and James, M.N.G. (1988) J. Mol. Biol. 200, 523-551. 26. Sambrook, J., Fritsch, E.F., and Ma&is, T. (1989) Molecular Cloning: A Laboratory
Mammal (2nd Edition). Cold Spring Harbor Labomtory, Cold Spring Harbor, NY. 27. Sanger, F., Nicklen, S., and Co&on, A.R. (1977) Proc. Natl. Acad. Sci. USA
74,5463-5467. 28. Shimma, K. and Kasai, K.-I. (1984) J. Chromatogr. 315, 161-166. 29. S&m, J.L. and Agard, D.A. (1989) Nature 341,462-464. 30. Terada, I., Kwon, S.-T., Miyata, Y., Matsuzawa, H., and Ohta, T. (1990) J. Biol. Chem.
265,6576-6581. 31. Turner, D.H., Sugimoto, N., Saeger, J.A., Longfellow, C.E., Freier, SM., and Kierzek, R.
(1987) Cold Spring Harbor Symp. Quant. Biol. 52, 123-133. 32. Zhu, X., Ohta, Y., Jordan, F., and Inouye, M. (1989) Nature 339,483-484.
713





























