Embed Size (px)
Citation preview

Molecular Cell
Article
OTULIN Restricts Met1-Linked Ubiquitinationto Control Innate Immune SignalingBerthe Katrine Fiil,1,4 Rune Busk Damgaard,1,4 Sebastian Alexander Wagner,2 Kirstin Keusekotten,3 Melanie Fritsch,1
Simon Bekker-Jensen,1 Niels Mailand,1 Chunaram Choudhary,2 David Komander,3 and Mads Gyrd-Hansen1,*1Department of Disease Biology2Department of ProteomicsNovo Nordisk Foundation Center for Protein Research, Faculty of Health and Medical Sciences, University of Copenhagen,
DK-2200 Copenhagen, Denmark3Medical Research Council Laboratory of Molecular Biology, Cambridge Biomedical Campus, Francis Crick Avenue, Cambridge,
CB2 0QH, UK4These authors contributed equally to this work
*Correspondence: [email protected]
http://dx.doi.org/10.1016/j.molcel.2013.06.004
SUMMARY
Conjugation of Met1-linked polyubiquitin (Met1-Ub)by the linear ubiquitin chain assembly complex(LUBAC) is an important regulatory modification ininnate immune signaling. So far, only few Met1-Ubsubstrates have been described, and the regulatorymechanisms have remained elusive. We recentlyidentified that the ovarian tumor (OTU) family deubi-quitinase OTULIN specifically disassembles Met1-Ub. Here, we report that OTULIN is critical for limitingMet1-Ub accumulation after nucleotide-oligomeriza-tion domain-containing protein 2 (NOD2) stimulation,and that OTULIN depletion augments signalingdownstream of NOD2. Affinity purification of Met1-Ub followed by quantitative proteomics uncoveredRIPK2 as the predominant NOD2-regulated sub-strate. Accordingly, Met1-Ub on RIPK2 was largelyinhibited by overexpressing OTULIN and was in-creased by OTULIN depletion. Intriguingly, OTULIN-depleted cells spontaneously accumulated Met1-Ub on LUBAC components, and NOD2 or TNFR1stimulation led to extensive Met1-Ub accumulationon receptor complex components. We propose thatOTULIN restricts Met1-Ub formation after immunereceptor stimulation to prevent unwarranted proin-flammatory signaling.
INTRODUCTION
An effective immunological barrier between the organism and
the surrounding environment is critical for human health, partic-
ularly at the mucosal surface of the gastrointestinal tract, which
constitutes the body’s largest surface (Chen et al., 2009; Maloy
and Powrie, 2011). Pattern recognition receptors (PRRs) present
on the cell membrane and in the cytoplasm collectively provide
our cells with the capability to recognize molecular patterns on
818 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.
highly diverse pathogens (Takeuchi and Akira, 2010). In
response, PRRs elicit a rapid and efficient immune response,
in part mediated by proinflammatory cytokines such as tumor
necrosis factor (TNF) and interleukins (Baud and Karin, 2009;
Takeuchi and Akira, 2010).
Stimulation of PRRs and cytokine receptors leads to assembly
of signaling complexes where ubiquitin (Ub) ligases conjugate
polyubiquitin (polyUb) on selected substrates to facilitate activa-
tion of mitogen-activated protein (MAP) kinases and the inhibitor
of kappa-B (IkB) kinase (IKK) complex consisting of IKKa, IKKb,
and NEMO (also termed IKKg) (Beug et al., 2012; Jiang and
Chen, 2012). IKK facilitates the degradation of IkBa, leading to
nuclear translocation of nuclear factor kB (NF-kB) transcription
factors. Together with transcription factors activated by MAP
kinases, NF-kB promotes expression of genes that orchestrate
the inflammatory response (Baud and Karin, 2009).
The intracellular PRR nucleotide-oligomerization domain-
containing protein 2 (NOD2) recognizes muramyl dipeptide
(MDP) constituents of bacterial peptidoglycan and plays a
critical role in gastrointestinal immunity (Chen et al., 2009).
Upon stimulation, NOD2 binds the proximal adaptor receptor-
interacting protein kinase 2 (RIPK2), which recruits Ub ligases
of the Inhibitor of Apoptosis (IAP) family (Beug et al., 2012). In
turn, XIAP and cIAPs facilitate nondegradative ubiquitination of
RIPK2 where polyUb formed by XIAP promotes recruitment of
the linear ubiquitin chain assembly complex (LUBAC) composed
of HOIL-1, HOIP, and SHARPIN (Bertrand et al., 2009; Damgaard
et al., 2012). LUBAC conjugates Met1-linked polyUb (Met1-Ub)
to facilitate efficient NF-kB activation and transcription of inflam-
matory mediators. A central regulatory point for this is the
activation of the IKK complex. IKK activation is dependent on
phosphorylation by the K63-Ub-activated TAB/TAK1 complex
as well as the conjugation of Met1-Ub, bound by the IKK subunit
NEMO (Jiang and Chen, 2012; Walczak et al., 2012).
For controlled and beneficial proinflammatory signaling,
conjugation of polyUb must be counterbalanced by deubiquiti-
nases such as CYLD and A20 that regulate different aspects of
proinflammatory signaling (Harhaj and Dixit, 2012). We and
others recently identified the ovarian tumor (OTU) domain family
deubiquitinase OTULIN (also termed FAM105B or Gumby) as a

Molecular Cell
OTULIN Regulates Innate Immune Signaling
Met1-Ub-specific deubiquitinase (Keusekotten et al., 2013;
Rivkin et al., 2013). OTULIN antagonizes LUBAC-mediated
Met1-Ub assembly and NF-kB activation upon TNF and poly(I:C)
treatment, and regulates TNF-induced proinflammatory signal-
ing and cell death (Keusekotten et al., 2013). LUBAC regulates
many aspects of cellular signaling and innate immune signaling
(Tokunaga and Iwai, 2012), and deregulation results in severe
immune dysfunction (Boisson et al., 2012; Gerlach et al., 2011;
Ikeda et al., 2011; Tokunaga et al., 2011). Indeed, we recently re-
ported that LUBAC activity is particularly important for signaling
triggered by the PRRs NOD1 and NOD2 (Damgaard et al., 2012).
Here, we investigated the role of OTULIN in NOD2-mediated
signaling. We find that OTULIN restricts Met1-Ub formation
and that this is important for limiting proinflammatory signaling
in response to NOD2 stimulation. SILAC-based proteomics
identify RIPK2, an essential protein for NOD2 signaling, as a
target for Met1-Ub conjugation after receptor stimulation, and
we find that OTULIN regulates its ubiquitination. Our data
suggest that OTULIN is a new regulator of NOD2 signaling and
is involved in innate immune signaling.
RESULTS
OTULIN Regulates NOD2 Signaling and Is Partof the NOD2 ComplexThe role of OTULIN in LUBAC signaling prompted us to
investigate its cellular function in context of NOD2 signaling.
Stimulation of NF-kB activity by ectopic NOD2 was inhibited
by overexpression of wild-type OTULIN (OTULINWT) but was
alleviated by mutating the catalytic cysteine (C129A), the trypto-
phan residue involved in Met1-Ub binding (W96A) (Keusekotten
et al., 2013), or both (WA/CA) (Figures 1A and 1B; see Figure S1A
online). Accordingly, OTULINWT but not OTULINWA/CA inhibited
nuclear translocation of the NF-kB subunit RelA/p65 after stimu-
lation of U2OS/FlpIn/TRex/HA-NOD2 (U2OS/NOD2) cells with
the NOD2 ligand L18-MDP (Figure 1C, quantified in Figure 1D).
Of note, the U2OS/NOD2 cells responded to L18-MDP without
addition of doxycycline (DOX). Under these conditions HA-NOD2
was expressed at low levels and was not detectable by immuno-
fluorescence staining (Figure S1B), and was detected by
western blotting only after immunoprecipitation with anti-HA
resin (Figure S1C).
RNAi-mediated depletion of OTULIN increased L18-MDP-
induced transcription of NF-kB-responsive genes (Figure 1E;
Figures S1E and S1F) and led to more pronounced degradation
of IkBa (Figure 1F) compared with cells transfected with
mismatch control siRNAs (siMM). Activation of the MAP kinases
p38 and JNK1/2 was only slightly increased by OTULIN deple-
tion (Figure 1F), consistent with the notion that OTULIN disas-
sembles Met1-Ub primarily involved in IKK activation. Of note,
OTULIN migrated as two distinct bands in the Tris-Glycine-
SDS buffer system (Figure 1E), but only as a single species in
MOPS (Figure S1D). The nature of the second band is currently
unknown but might represent an alternative variant or a modified
form of OTULIN.
Immunoprecipitation of HA-tagged NOD2 from DOX-treated
U2OS/NOD2 cells copurified RIPK2, XIAP, and the three LUBAC
components, HOIP, HOIL-1, and SHARPIN. Intriguingly, endog-
M
enous OTULIN also copurified with NOD2 (Figure 2A), suggest-
ing that OTULIN could regulate signaling at the NOD2 signaling
complex. XIAP enhances recruitment of LUBAC to the NOD2
signaling complex, where LUBAC conjugates Met1-Ub to facili-
tate downstream signaling (Damgaard et al., 2012). Overex-
pressed OTULIN efficiently blocked LUBAC-induced NF-kB
activation, suggesting that OTULIN functions downstream of
LUBAC by disassembling Met1-Ub (Figure 2B; Figure S2A).
Accordingly, OTULIN impaired XIAP-mediated NF-kB activity
to an extent similar to that of overexpression of RING-mutated
HOIP (Haas et al., 2009) and HOIL-1 (termed dominant-negative
[DN]-LUBAC) (Figures 2C and 2D; Figures S2B and S2C).
To directly address if OTULIN antagonizes NF-kB activation
by disassembling Met1-Ub, we devised a system whereby a
noncleavable (Gly76 / Ser76; GS) Met1-Ub was docked to
XIAP via an IAP-binding motif (IBM). The engineered protein
comprised Ub fused to the N-terminal Ala-Val-Pro-Ile (AVPI)
IAP-binding motif (IBM) of second mitochondria-derived acti-
vator of caspase (Smac), followed by a FLAG epitope and four
Ub(GS) moieties in tandem (Figure 2E). The N-terminal Ub is
rapidly removed by deubiquitinases to expose the Smac IBM
(Hunter et al., 2003), which enables binding of AVPI-Ub4GS to
XIAP in a manner dependent on an N-terminal alanine (Fig-
ure S2D). AVPI-Ub4GS did not increase NF-kB activity when ex-
pressed alone, but facilitated potent NF-kB activation when
coexpressed with the Ub-ligase-deficient XIAPF495A (XIAPF/A)
(Gyrd-Hansen et al., 2008), which also failed to activate NF-kB
when expressed alone (Figure 2F; Figure S2C). Substituting
Ala1 with Leu prevented NF-kB activation, confirming that
signaling by AVPI-Ub4GS depended on docking to XIAP
(Figure S2E). NF-kB activation induced by XIAPF/A and AVPI-
Ub4GS was not inhibited by overexpression of DN-LUBAC,
showing that the ectopic noncleavable Met1-Ub efficiently
bypassed the requirement for LUBAC activity (Figure 2F; Fig-
ure S2C). Importantly, under these conditions overexpressed
OTULIN also failed to inhibit NF-kB activation (Figure 2G; Fig-
ure S2B), establishing that the inhibitory effect of OTULIN on
NF-kB activation is dependent on its ability to disassemble
Met1-Ub.
RIPK2 Is a Substrate for Met1-Ub in Response to NOD2StimulationNext, we sought to uncover potential OTULIN substrates by
identifying proteins modified by Met1-Ub in response to NOD2
stimulation. For this, we employed a Met1-linkage-specific Ub
binder (M1-SUB) based on NEMO’s UBAN region (Rahighi
et al., 2009). Ectopic expression of Ub-linkage-selective binders
can inhibit cellular signaling processes dependent on theUb link-
age to which it binds (vanWijk et al., 2012). Indeed, expression of
the GFP-coupled M1-SUB (GFP-M1-SUB) inhibited nuclear
localization of RelA/p65 after NOD2 stimulation (Figures 3A
and 3B; Figure S3A). Mutation of residues in the M1-SUB
required for Ub binding (GFP-M1-SUBmut) (Rahighi et al.,
2009; Wu et al., 2006) reversed the inhibitory effect on RelA/
p65 translocation, showing that the M1-SUB inhibited NOD2
signaling by binding to Ub (Figures 3A and 3B; Figure S3A).
Next, we purified Met1-Ub-modified proteins using recombinant
GST-coupled M1-SUB (for brevity henceforth referred to as
olecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc. 819
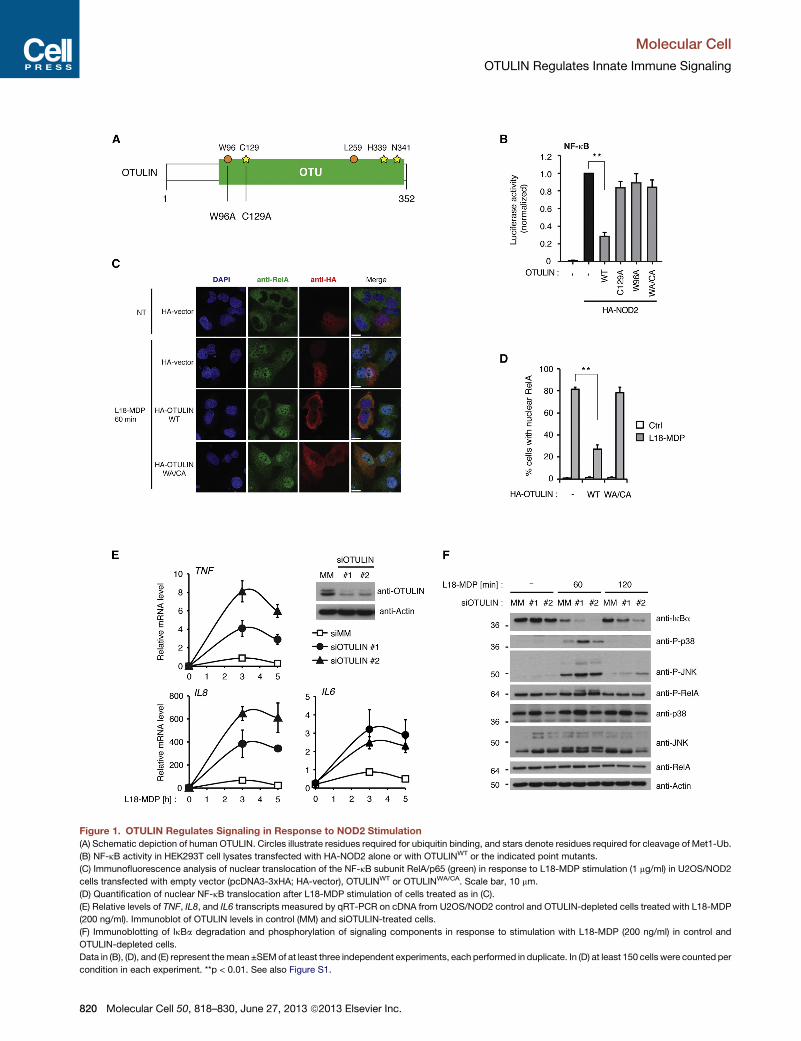
Figure 1. OTULIN Regulates Signaling in Response to NOD2 Stimulation
(A) Schematic depiction of human OTULIN. Circles illustrate residues required for ubiquitin binding, and stars denote residues required for cleavage of Met1-Ub.
(B) NF-kB activity in HEK293T cell lysates transfected with HA-NOD2 alone or with OTULINWT or the indicated point mutants.
(C) Immunofluorescence analysis of nuclear translocation of the NF-kB subunit RelA/p65 (green) in response to L18-MDP stimulation (1 mg/ml) in U2OS/NOD2
cells transfected with empty vector (pcDNA3-3xHA; HA-vector), OTULINWT or OTULINWA/CA. Scale bar, 10 mm.
(D) Quantification of nuclear NF-kB translocation after L18-MDP stimulation of cells treated as in (C).
(E) Relative levels of TNF, IL8, and IL6 transcripts measured by qRT-PCR on cDNA from U2OS/NOD2 control and OTULIN-depleted cells treated with L18-MDP
(200 ng/ml). Immunoblot of OTULIN levels in control (MM) and siOTULIN-treated cells.
(F) Immunoblotting of IkBa degradation and phosphorylation of signaling components in response to stimulation with L18-MDP (200 ng/ml) in control and
OTULIN-depleted cells.
Data in (B), (D), and (E) represent themean ±SEMof at least three independent experiments, each performed in duplicate. In (D) at least 150 cells were counted per
condition in each experiment. **p < 0.01. See also Figure S1.
Molecular Cell
OTULIN Regulates Innate Immune Signaling
820 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.
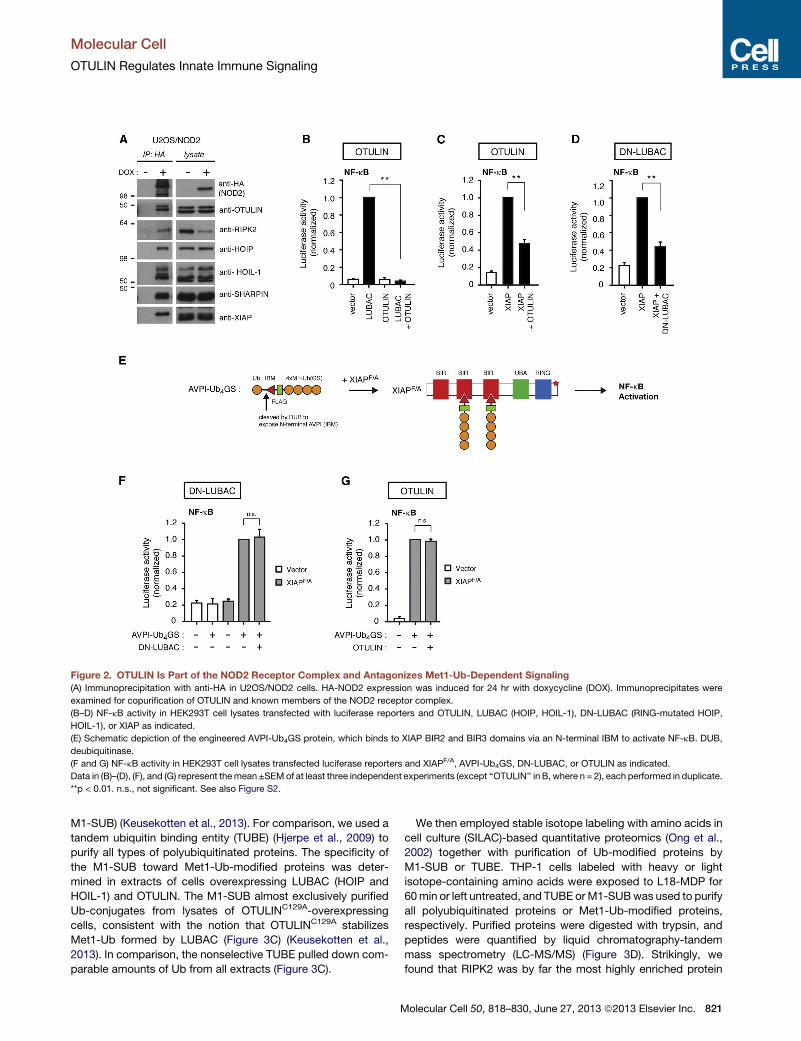
Figure 2. OTULIN Is Part of the NOD2 Receptor Complex and Antagonizes Met1-Ub-Dependent Signaling(A) Immunoprecipitation with anti-HA in U2OS/NOD2 cells. HA-NOD2 expression was induced for 24 hr with doxycycline (DOX). Immunoprecipitates were
examined for copurification of OTULIN and known members of the NOD2 receptor complex.
(B–D) NF-kB activity in HEK293T cell lysates transfected with luciferase reporters and OTULIN, LUBAC (HOIP, HOIL-1), DN-LUBAC (RING-mutated HOIP,
HOIL-1), or XIAP as indicated.
(E) Schematic depiction of the engineered AVPI-Ub4GS protein, which binds to XIAP BIR2 and BIR3 domains via an N-terminal IBM to activate NF-kB. DUB,
deubiquitinase.
(F and G) NF-kB activity in HEK293T cell lysates transfected luciferase reporters and XIAPF/A, AVPI-Ub4GS, DN-LUBAC, or OTULIN as indicated.
Data in (B)–(D), (F), and (G) represent themean ±SEMof at least three independent experiments (except ‘‘OTULIN’’ in B, where n = 2), each performed in duplicate.
**p < 0.01. n.s., not significant. See also Figure S2.
Molecular Cell
OTULIN Regulates Innate Immune Signaling
M1-SUB) (Keusekotten et al., 2013). For comparison, we used a
tandem ubiquitin binding entity (TUBE) (Hjerpe et al., 2009) to
purify all types of polyubiquitinated proteins. The specificity of
the M1-SUB toward Met1-Ub-modified proteins was deter-
mined in extracts of cells overexpressing LUBAC (HOIP and
HOIL-1) and OTULIN. The M1-SUB almost exclusively purified
Ub-conjugates from lysates of OTULINC129A-overexpressing
cells, consistent with the notion that OTULINC129A stabilizes
Met1-Ub formed by LUBAC (Figure 3C) (Keusekotten et al.,
2013). In comparison, the nonselective TUBE pulled down com-
parable amounts of Ub from all extracts (Figure 3C).
M
We then employed stable isotope labeling with amino acids in
cell culture (SILAC)-based quantitative proteomics (Ong et al.,
2002) together with purification of Ub-modified proteins by
M1-SUB or TUBE. THP-1 cells labeled with heavy or light
isotope-containing amino acids were exposed to L18-MDP for
60min or left untreated, and TUBE orM1-SUBwas used to purify
all polyubiquitinated proteins or Met1-Ub-modified proteins,
respectively. Purified proteins were digested with trypsin, and
peptides were quantified by liquid chromatography-tandem
mass spectrometry (LC-MS/MS) (Figure 3D). Strikingly, we
found that RIPK2 was by far the most highly enriched protein
olecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc. 821

Figure 3. Proteome-wide Quantification of NOD2-Regulated Met1-Ub
(A) Immunofluorescence analysis of nuclear translocation of the NF-kB subunit RelA/p65 (red) in response to L18-MDP stimulation (1 mg/ml) in U2OS/NOD2 cells
transfected with GFP vector, GFP-M1-SUB, or a Ub-binding mutant GFP-M1-SUBmut. Scale bar, 10 mm.
(B) Quantification of nuclear NF-kB translocation after L18-MDP stimulation of cells treated as in (A).
(C) Purification of endogenous Ub conjugates using GST-coupledM1-SUB (Met1-Ub) or TUBE (all Ub-linkages). Pull-down with glutathione Sepharose beads on
HEK293Tcell lysates transfectedwith LUBAC (HOIP andHOIL) and the indicatedOTULINmutants. Purifiedmaterial and lysatewere examinedby immunoblotting.
(D) Outline of the SILAC-based proteomics approach for quantification of protein ubiquitination in response to NOD2 stimulation.
(E) MS data from TUBE purification of ubiquitinated proteins from THP-1 cells treated or not with L18-MDP (200 ng/ml) for 60min. The circle representing RIPK2 is
marked in red. Themass spectrum shows the relative abundance of the RIPK2 peptide TQNILLDNEFHVK in L18-MDP-treated (SILAC heavy) cells comparedwith
untreated (SILAC light).
(legend continued on next page)
Molecular Cell
OTULIN Regulates Innate Immune Signaling
822 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.

Molecular Cell
OTULIN Regulates Innate Immune Signaling
(SILAC H/L ratio >20) after L18-MDP stimulation in the TUBE
purification (Figures 3E and 3G; Table S1). This is consistent
with previous reports showing that RIPK2 is the predominant
target for ubiquitination in the NOD2 signaling complex (Dam-
gaard et al., 2012; Hasegawa et al., 2008). Remarkably, in the
M1-SUB purification RIPK2 was enriched >12-fold after NOD2
stimulation and was the only detected protein with a SILAC H/L
ratio >2.5 (Figures 3F and 3G; Table S1). NEMO, XIAP, LUBAC
subunits, and other NOD2 signaling complex components
were not enriched after L18-MDP stimulation, suggesting that
RIPK2 is a major target of Met1-linked ubiquitination in response
to NOD2 stimulation (Figure 3G; Table S1). Time course analysis
confirmed the mass spectrometry data and showed that L18-
MDP-induced ubiquitination of RIPK2 occurred at 60 min, was
detectable both by TUBE andM1-SUB, andwas decreased after
120 and 240 min. Notably, the ubiquitination of RIPK2 coincided
with IkBa phosphorylation and degradation and phosphorylation
of RelA/p65 and MAP kinases (Figure 3H).
OTULIN Limits Met1-Ub Accumulation on RIPK2to Control NOD2 SignalingTo test if OTULIN regulated Met1-linked ubiquitination of RIPK2,
Ub conjugates were isolated from L18-MDP-treated U2OS/
NOD2 cells depleted of OTULIN. Indeed, RNAi-mediated
OTULIN knockdown led to extensive accumulation of Met1-
ubiquitinated RIPK2 that correlated with enhanced degradation
of IkBa in response to NOD2 stimulation with 200 ng/ml L18-
MDP (Figure 4A, lanes 1–6). Conversely, OTULIN overexpression
suppressed Met1-Ub of RIPK2 in a manner dependent on its
catalytic activity (Figure 4B).
These data strongly suggest that RIPK2 is a physiological sub-
strate for Met1-linked polyubiquitination and is regulated by
OTULIN. RIPK2 is also modified by Ub linkages other than
Met1, and genetic deletion of XIAP or pharmacological inhibition
of XIAP dramatically reduces RIPK2 ubiquitination (Damgaard
et al., 2012, 2013). To directly address the contribution of Met1
linkages to Ub-modified RIPK2, we subjected M1-SUB-purified
Ub-RIPK2 to in vitro Ub chain restriction analysis by OTULIN
and other linkage-selective or promiscuous deubiquitinases
(Hospenthal et al., 2013; Mevissen et al., 2013) (Figures 4C and
4D). The promiscuous deubiquitinase USP21 removed virtually
all Ub moieties from RIPK2, leading to a collapse of the RIPK2
‘‘smear’’ into a single band representing unmodified RIPK2 (Fig-
ure 4C, see lanes 1–3 and lanes 8–10). In contrast, incubation
with the K48-linkage-selective OTUB1 did not alter the migration
pattern of Ub-RIPK2 on SDS-PAGE (Figure 4C, compare lanes 2
and 7, and lanes 9 and 14). Importantly, incubation with
OTULINWT but not catalytic inactive OTULINC129A resulted in a
downshift in the migration pattern of Ub-RIPK2, which was
particularly evident for Ub-RIPK2 isolated from OTULIN-
depleted cells (Figure 4C, compare lanes 2, 4, and 5, and lanes
(F) Same as in (E) but after purification of ubiquitinated proteins with M1-SUB. R
(G) SILAC H/L ratios of proteins identified in (E) and (F) with reported function in
(H) Time course analysis of RIPK2 ubiquitination with TUBE and M1-SUB in lysate
lysate were examined by immunoblotting.
Data in (B) represent the mean ±SEM of three independent experiments. At least
Figure S3 and Table S1.
M
9, 11, and 12). Interestingly, OTULIN did not give rise to an
increased level of unmodified RIPK2, suggesting that the Met1
linkages are formed on pre-existing polyubiquitin on RIPK2.
Accordingly, the viral deubiquitinase vOTU that disassembles
all Ub linkages except Met1 (Mevissen et al., 2013) removed
most Ub from RIPK2 and increased the amount of unmodified
and monoubiquitinated RIPK2 (Figure 4C, see lanes 6 and 13).
NEMO facilitates IKK activation through interaction with
Met1-Ub-modified proteins (Rahighi et al., 2009). Accordingly,
depletion of OTULIN enhanced the recruitment of NEMO to
Met1-ubiquitinated RIPK2 in response to L18-MDP, whereas
NEMO recruitment was inhibited by OTULIN overexpression
(Figures 4E and 4F). Notably, ubiquitinated RIPK2 was readily
detected in the lysates from OTULIN-depleted cells treated
with L18-MDP, illustrating extensive ubiquitination of RIPK2
when OTULIN is depleted (Figure 4E).
This finding, together with the observation that OTULIN was
not transcriptionally induced by L18-MDP (Figure 5A), led us to
speculate that OTULIN might regulate the initial response to
the NOD2 ligand rather than function as part of a negative feed-
back mechanism. This notion was supported by an extended
time course analysis of NOD2 signaling, which showed that
Met1-linked ubiquitination of RIPK2 in OTULIN-depleted cells
decreased with kinetics similar to that of the control cells (Fig-
ure 5B). Also, RelA/p65 phosphorylation and IkBa degradation
in OTULIN-depleted cells was temporally comparable to the
L18-MDP response observed in cells transfected with control
siRNA (Figure 5B and Figure 1F). To test if OTULIN depletion
sensitized cells to NOD2 stimulation, we treated U2OS/NOD2
cells with lower concentrations of L18-MDP (Figure 5C; Fig-
ure S4A). Interestingly, treatment of U2OS/NOD2 cells with
L18-MDP diluted 40-fold (5 ng/ml) induced only marginal degra-
dation of IkBa and RelA/p65 phosphorylation in control cells,
whereas signaling was readily detected OTULIN-depleted cells,
albeit with slower kinetics than after treatment with higher ligand
concentrations (Figures 5C and 1F). In line with this, Met1-Ub on
RIPK2 was barely detectable in control cells subjected to the low
concentration of L18-MDP but accumulated in the OTULIN-
depleted cells to levels comparable with those in control cells
treated with 200 ng/ml of L18-MDP (Figure 4A, compare lane 4
with lanes 7–9). Accordingly, the transcriptional response to
5 ng/ml L18-MDP was up to 20-fold higher in OTULIN-depleted
cells than in control siRNA-transfected cells (Figure 5D).
Decreased OTULIN Function Leads to PromiscuousMet1-Ub AccumulationIn nonstimulated cells, Met1-Ub assembled by LUBAC does not
accumulate to detectable levels, suggesting that it is rapidly
turned over. Met1-Ub can, however, be stabilized by exogenous
OTULINC129A (Figure 3A) (Keusekotten et al., 2013). Moreover,
Met1-Ub accumulates on overexpressed HOIP when OTULIN
IPK2 is marked in red.
the NOD2 signaling complex. NA, not available.
s of U2OS/NOD2 cells treated with L18-MDP (200 ng/ml). Purified material and
140 cells were counted per condition in each experiment. *p < 0.05. See also
olecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc. 823
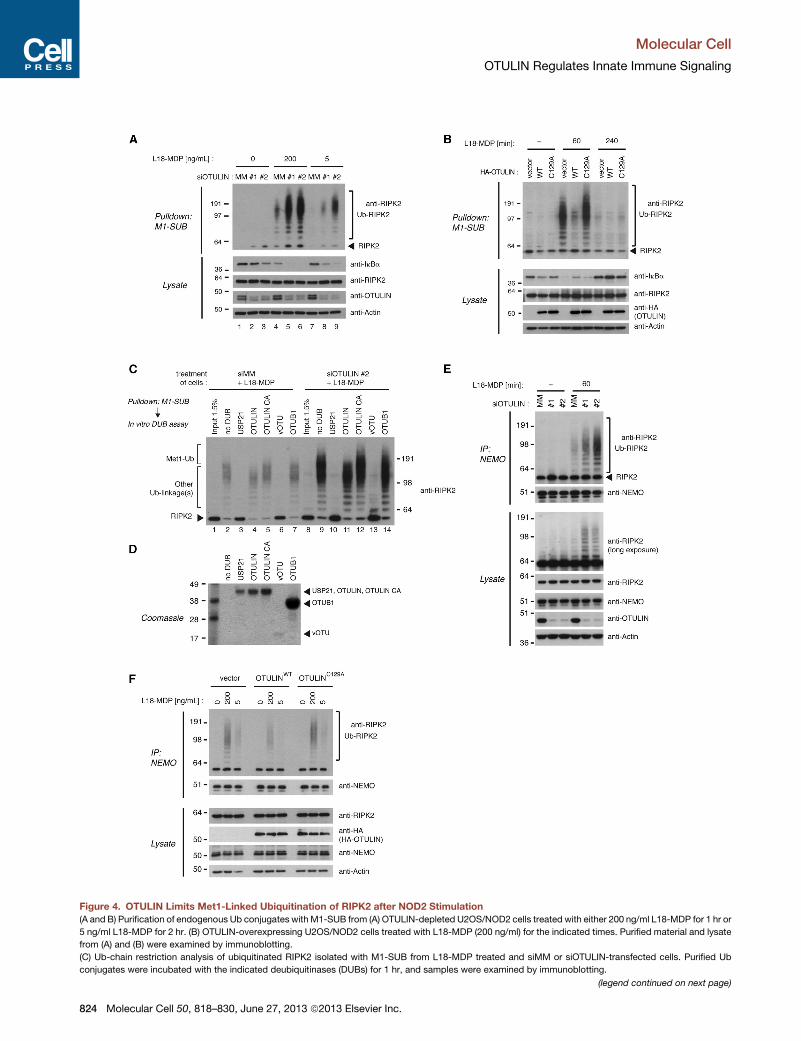
Figure 4. OTULIN Limits Met1-Linked Ubiquitination of RIPK2 after NOD2 Stimulation
(A and B) Purification of endogenous Ub conjugates with M1-SUB from (A) OTULIN-depleted U2OS/NOD2 cells treated with either 200 ng/ml L18-MDP for 1 hr or
5 ng/ml L18-MDP for 2 hr. (B) OTULIN-overexpressing U2OS/NOD2 cells treated with L18-MDP (200 ng/ml) for the indicated times. Purified material and lysate
from (A) and (B) were examined by immunoblotting.
(C) Ub-chain restriction analysis of ubiquitinated RIPK2 isolated with M1-SUB from L18-MDP treated and siMM or siOTULIN-transfected cells. Purified Ub
conjugates were incubated with the indicated deubiquitinases (DUBs) for 1 hr, and samples were examined by immunoblotting.
(legend continued on next page)
Molecular Cell
OTULIN Regulates Innate Immune Signaling
824 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.
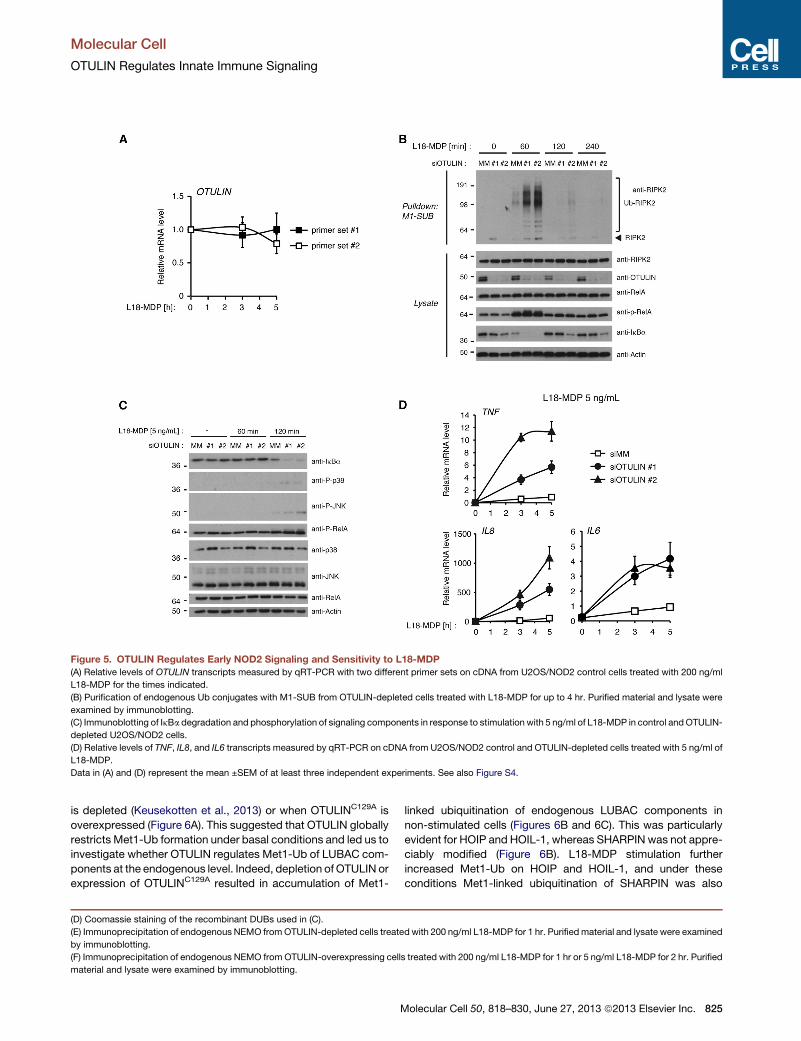
Figure 5. OTULIN Regulates Early NOD2 Signaling and Sensitivity to L18-MDP
(A) Relative levels of OTULIN transcripts measured by qRT-PCR with two different primer sets on cDNA from U2OS/NOD2 control cells treated with 200 ng/ml
L18-MDP for the times indicated.
(B) Purification of endogenous Ub conjugates with M1-SUB from OTULIN-depleted cells treated with L18-MDP for up to 4 hr. Purified material and lysate were
examined by immunoblotting.
(C) Immunoblotting of IkBa degradation and phosphorylation of signaling components in response to stimulation with 5 ng/ml of L18-MDP in control andOTULIN-
depleted U2OS/NOD2 cells.
(D) Relative levels of TNF, IL8, and IL6 transcripts measured by qRT-PCR on cDNA from U2OS/NOD2 control and OTULIN-depleted cells treated with 5 ng/ml of
L18-MDP.
Data in (A) and (D) represent the mean ±SEM of at least three independent experiments. See also Figure S4.
Molecular Cell
OTULIN Regulates Innate Immune Signaling
is depleted (Keusekotten et al., 2013) or when OTULINC129A is
overexpressed (Figure 6A). This suggested that OTULIN globally
restricts Met1-Ub formation under basal conditions and led us to
investigate whether OTULIN regulates Met1-Ub of LUBAC com-
ponents at the endogenous level. Indeed, depletion of OTULIN or
expression of OTULINC129A resulted in accumulation of Met1-
(D) Coomassie staining of the recombinant DUBs used in (C).
(E) Immunoprecipitation of endogenous NEMO fromOTULIN-depleted cells treate
by immunoblotting.
(F) Immunoprecipitation of endogenous NEMO from OTULIN-overexpressing cell
material and lysate were examined by immunoblotting.
M
linked ubiquitination of endogenous LUBAC components in
non-stimulated cells (Figures 6B and 6C). This was particularly
evident for HOIP and HOIL-1, whereas SHARPIN was not appre-
ciably modified (Figure 6B). L18-MDP stimulation further
increased Met1-Ub on HOIP and HOIL-1, and under these
conditions Met1-linked ubiquitination of SHARPIN was also
d with 200 ng/ml L18-MDP for 1 hr. Purifiedmaterial and lysate were examined
s treated with 200 ng/ml L18-MDP for 1 hr or 5 ng/ml L18-MDP for 2 hr. Purified
olecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc. 825
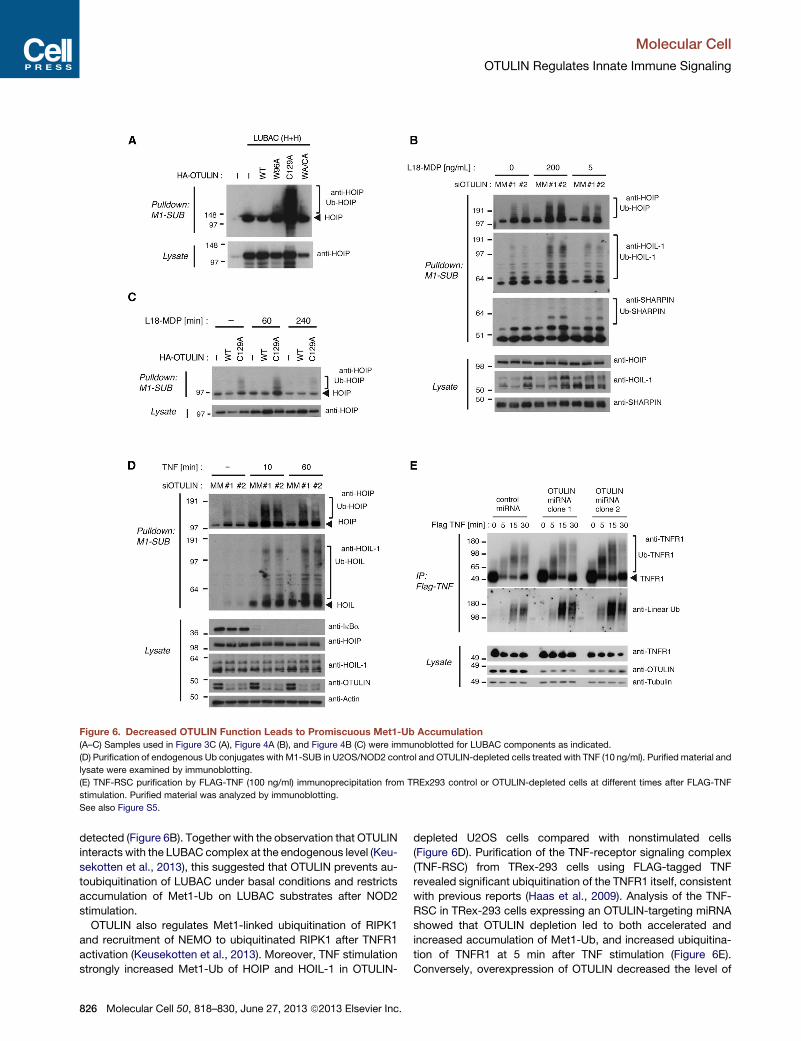
Figure 6. Decreased OTULIN Function Leads to Promiscuous Met1-Ub Accumulation
(A–C) Samples used in Figure 3C (A), Figure 4A (B), and Figure 4B (C) were immunoblotted for LUBAC components as indicated.
(D) Purification of endogenous Ub conjugates with M1-SUB in U2OS/NOD2 control and OTULIN-depleted cells treated with TNF (10 ng/ml). Purified material and
lysate were examined by immunoblotting.
(E) TNF-RSC purification by FLAG-TNF (100 ng/ml) immunoprecipitation from TREx293 control or OTULIN-depleted cells at different times after FLAG-TNF
stimulation. Purified material was analyzed by immunoblotting.
See also Figure S5.
Molecular Cell
OTULIN Regulates Innate Immune Signaling
detected (Figure 6B). Together with the observation that OTULIN
interacts with the LUBAC complex at the endogenous level (Keu-
sekotten et al., 2013), this suggested that OTULIN prevents au-
toubiquitination of LUBAC under basal conditions and restricts
accumulation of Met1-Ub on LUBAC substrates after NOD2
stimulation.
OTULIN also regulates Met1-linked ubiquitination of RIPK1
and recruitment of NEMO to ubiquitinated RIPK1 after TNFR1
activation (Keusekotten et al., 2013). Moreover, TNF stimulation
strongly increased Met1-Ub of HOIP and HOIL-1 in OTULIN-
826 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.
depleted U2OS cells compared with nonstimulated cells
(Figure 6D). Purification of the TNF-receptor signaling complex
(TNF-RSC) from TRex-293 cells using FLAG-tagged TNF
revealed significant ubiquitination of the TNFR1 itself, consistent
with previous reports (Haas et al., 2009). Analysis of the TNF-
RSC in TRex-293 cells expressing an OTULIN-targeting miRNA
showed that OTULIN depletion led to both accelerated and
increased accumulation of Met1-Ub, and increased ubiquitina-
tion of TNFR1 at 5 min after TNF stimulation (Figure 6E).
Conversely, overexpression of OTULIN decreased the level of

Molecular Cell
OTULIN Regulates Innate Immune Signaling
Met1-Ub in the TNF-RSC and reduced the apparent molecular
weight of ubiquitinated TNFR1 (Figure S5).
Together, our data suggest that OTULIN is important for re-
stricting the accumulation of Met1-Ub in innate immune receptor
signaling complexes, and that this is essential for preventing
excessive proinflammatory signaling in response to NOD2
stimulation.
DISCUSSION
Met1-Ub assembled by LUBAC has emerged as a versatile pro-
tein modification that regulates several cellular functions, in
particular proinflammatory signaling (Tokunaga and Iwai,
2012). We and others recently reported that NOD2-mediated
transcription of NF-kB-responsive genes is largely dependent
on LUBAC function (Damgaard et al., 2012; Warner et al.,
2013). In agreement with this, we show here that OTULIN has
an important role in regulating NOD2-mediated signaling and
that reducing OTULIN levels leads to a striking increase in tran-
scription of inflammatory mediators after NOD2 stimulation. In
comparison, OTULIN and LUBAC seem to have more subdued
functions in regulating NF-kB activation in response to TNFR1
stimulation. Instead, deregulation of Met1-Ub formation after
TNF treatment leads to excessive programmed cell death (Bois-
son et al., 2012; Gerlach et al., 2011; Ikeda et al., 2011; Tokunaga
et al., 2009). The underlying mechanisms dictating these differ-
ences between NOD2- and TNFR1-dependent signaling are
not well understood but will be important to dissect in future
studies.
Identifying physiological substrates for ubiquitination by
LUBAC has proven exceptionally difficult, and only few protein
substrates (NEMO and RIPK1) have been reported so far (Bois-
son et al., 2012; Gerlach et al., 2011; Keusekotten et al., 2013;
Tokunaga et al., 2009). One reason for this is that Met1-Ub
assembly is incompatible with N-terminally modified Ub such
as epitope-tagged exogenous ubiquitin (Gerlach et al., 2011;
Kirisako et al., 2006), a strategy widely used to study Lys-linked
ubiquitination. We therefore used a proteomics approach that
combined affinity purification of endogenous Met1-Ub-modified
proteins using a Met1-linkage-selective affinity reagent with
SILAC-based quantitative proteomics. This revealed a remark-
able specificity in ubiquitination after NOD2 stimulation and
identified RIPK2 as the predominant Met1-Ub target. Mass
spectrometry analysis of Ub-modified proteins purified by the
nonselective TUBE and our previous analysis of L18-MDP-
induced ubiquitination (Damgaard et al., 2012) further supported
that RIPK2 is the major substrate for ubiquitination of the NOD2-
signaling complex. We believe that this methodology might be
applicable for analysis of a wide range of ubiquitin-regulated
processes, and for the detection of other Ub linkages in innate
immune signaling.
Using recombinant deubiquitinases for Ub chain restriction
analysis of Ub-modified RIPK2, we provide compelling evidence
that the polyubiquitin conjugated on RIPK2 after NOD2 stimula-
tion comprises at least two distinct Ub linkages. OTULIN
trimmed away the slowest-migrating Ub-RIPK2 species without
increasing the amount of unmodified RIPK2, suggesting that
Met1-Ub is conjugated predominantly on existing polyubiquitin,
M
conjugated on RIPK2 by other Ub ligase(s). Ectopically
expressed XIAP conjugates polyubiquitin on RIPK2 linked
through lysines other than K48 and K63 (Damgaard et al.,
2012), implying that atypical polyubiquitin might play a role in
NOD2 signaling. Detailed Ub chain restriction analysis using a
panel of linkage-selective deubiquitinases (Mevissen et al.,
2013) might thus uncover unappreciated roles for atypical
polyubiquitin in regulation of NOD2 signaling.
Our analysis of Met1-Ub in NOD2 signaling and how it is regu-
lated by OTULIN led to several interesting observations. First,
OTULIN was rate limiting for the accumulation of Met1-Ub on
RIPK2, which was particularly evident when NOD2 was stimu-
lated with low amounts of ligand. Second, OTULIN continuously
disassembled Met1-Ub that, in the absence of OTULIN activity,
accumulated on LUBAC components. Third, stimulation of
NOD2 and TNFR1 resulted in promiscuous Met1-Ub of receptor
components unless counterbalanced by endogenous OTULIN.
These data imply that LUBAC is active under basal conditions
and continuously ubiquitinates itself and possibly other sub-
strates in a not strictly selective manner, and that the Met1-Ub
chains are rapidly disassembled by OTULIN. This is supported
by the observation that OTULIN binds to the LUBAC complex
(Keusekotten et al., 2013), by the strong increase in steady-state
levels of Met1-Ub and HOIP ubiquitination in cells overexpress-
ing LUBAC together with catalytically inactive OTULIN, as well as
by the accumulation of Met1-Ub on endogenous HOIP and
HOIL-1 in cells with decreased OTULIN activity. The balanced
activity of LUBAC and OTULIN may explain why Met1-Ub is pre-
sent at very low levels in cells, and why its abundance is not
significantly increased by overexpression of LUBAC, despite
the fact that LUBAC readily assembles Ub chains in vitro
(Kirisako et al., 2006; Stieglitz et al., 2012).
While LUBAC appears to be active under basal conditions,
Met1-Ub formation on LUBAC components was increased by
NOD2 and TNFR1 stimulation in OTULIN-depleted cells, sug-
gesting that the propensity of LUBAC for assembling Met1-Ub
is regulated by environmental cues such as microbial products
and cytokines. This might occur through direct activation of the
ligase complex or as a consequence of induced proximity of
LUBAC within the formed receptor complexes. LUBAC activity
was recently reported to be regulated by the Ub ligase Parkin
in response to cellular stress signals, exemplifying that LUBAC
activity may indeed be regulated (Muller-Rischart et al., 2013).
Further investigation of LUBAC function in innate immune
receptor signaling may thus reveal additional mechanisms for
modulating formation of productive Met1-Ub.
Other deubiquitinases such as A20 and CYLD function as
negative regulators of immune signaling (Harhaj and Dixit,
2012). A20 contains a K48-Ub-specific OTU domain (Mevissen
et al., 2013) but is thought to disassemble K63-Ub chains in cells
(Wertz et al., 2004), and A20/TNFAIP3 is a transcriptional target
of NF-kB transcription factors. Consequently, A20 is essential for
termination of proinflammatory signaling (Boone et al., 2004; Lee
et al., 2000). In contrast, our data suggest that OTULIN functions
to restrict Met1-Ub accumulation and regulate early signaling
processes. This was supported by the observation that OTULIN
was not transcriptionally activated by NOD2 stimulation and that
OTULIN levels were not elevated, consistent with data obtained
olecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc. 827

Molecular Cell
OTULIN Regulates Innate Immune Signaling
using TNF (Keusekotten et al., 2013). Also, the temporal profile of
RIPK2 ubiquitination, degradation of IkBa, or transcription of
NF-kB-responsive genes after NOD2 stimulation was not
markedly prolonged by OTULIN depletion. Instead, OTULIN
knockdown rendered cells hyperresponsive to NOD2 stimula-
tion, which was particularly evident when ligand concentration
was decreased. We speculate that OTULIN might be involved
in determining the threshold for proinflammatory signaling in
response to NOD2 ligands.
In conclusion, we have shown that OTULIN is essential for
restricting accumulation of Met1-Ub in cells and for limiting
NOD2-dependent proinflammatory signaling. Our data thus pro-
vide evidence that OTULIN is a physiological regulator of innate
immune responses and inflammation. The analysis of OTULIN/
FAM105B-ablated mice will be important for revealing the in vivo
function of OTULIN in innate immunity.
EXPERIMENTAL PROCEDURES
Plasmids and Cloning, Cell Lines, RNA Interference, Antibodies, and
Affinity Resin
See the Supplemental Experimental Procedures.
Receptor Stimulation
THP-1 and U2OS/NOD2 cells were treated with the NOD2 ligand L18-MDP
(InvivoGen, San Diego, CA) for the indicated times with 5–1,000 ng/ml or
TNF 1–100 ng/ml (R&D Systems, Minneapolis, MN); both were added directly
to the culture medium (see the Supplemental Experimental Procedures).
Luciferase Reporter Assays
HEK293T cells were cotransfected with the NF-kB luciferase reporter
construct pBIIX-luc and a thymidine kinase-renilla luciferase construct for
normalization of transfection efficiency. Cells were either cotransfected with
additional plasmids or treated with compounds as indicated elsewhere, and
luciferase assays were performed as previously described (Damgaard et al.,
2012). Individual experiments were performed in duplicate.
Quantitative RT-PCR
Total RNA was isolated from U2OS/NOD2 using RNeasy Mini Kit (QIAGEN,
Hilden, Germany), and DNase digestion was performed on column with the
RNase-Free DNase Set (QIAGEN) according to the manufacturer’s protocol.
Total RNA was reverse transcribed with SuperScript III Reverse Transcriptase
(Invitrogen, Carlsbad, CA) and oligo(d)T primers in the presence of RNasin
(Promega, Madison, WI). QPCR was performed using Brilliant III Ultra-Fast
SYBR Green QPCR Master (Agilent Technologies, Santa Clara, CA). Gene-
specific primers were used to amplify cDNA (see the Supplemental Experi-
mental Procedures).
Purification of Endogenous Ubiquitin Conjugates
Ubiquitin conjugates from cell lysates were pulled down in THP-1 and U2OS/
NOD2 cells using affinity reagents. For isolation of Met1-Ub, recombinant
protein containing one copy of the UBAN region from human NEMO
(residues 257–346) fused to Glutatione-S-transferase (GST) was used (M1-
SUB; full sequence will be made available upon request). TUBE (TUBE1;
Lifesensors, Malvern, PA, was used for the mass spectrometry analysis)
consists of four UBA domains in tandem fused to GST and was used to
purify all polyubiquitin linkages (full sequence will be made available upon
request). Purified material was analyzed by immunoblotting or mass spec-
trometry analysis (see the Supplemental Experimental Procedures).
Deubiquitinase Assays
Ubiquitin conjugates from L18-MDP-treated siMM or siOTULIN#2 were iso-
lated byM1-SUB onGST-beads, washed in PBS Tween-20 (0.1%), and resus-
pended in deubiquitinase-buffer (50 mMHEPES [pH 7.5], 100 mMNaCl, 2 mM
828 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.
DTT, 1 mM MnCl2, 0.01% Brij-35) without or with deubiquitinases (USP21
[0.5 mM], OTULIN [1 mM], OTULIN CA [1 mM], vOTU [0.4 mM], OTUB1
[15 mM]). Samples incubated for 1 hr at 30�C, and LSB buffer was added to
end the reaction. Cloning, expression, and purification of the deubiquitinases
used here is described elsewhere (Mevissen et al., 2013).
Immunoprecipitation
Endogenous NEMO was immunopurified as previously described (Keuse-
kotten et al., 2013) from U2OS/NOD2 cells transfected and treated as indi-
cated. HA-NOD2 from U2OS/NOD2 cells treated with doxycycline (2 mg/ml)
for 24 hr was immunopurified as described for NEMO, except that anti-HA-
agarose resin (Sigma-Aldrich, Gillingham, UK) was used as affinity reagent.
TNF-RSC purification was performed as previously described but using
100 ng/ml FLAG-TNF (Haas et al., 2009; Keusekotten et al., 2013) (see the Sup-
plemental Experimental Procedures).
Immunofluorescence Staining and Microscopy
U2OS/NOD2 cells were fixed in 4% formaldehyde, permeabilized with PBS
containing 0.2% Triton X-100 for 5 min, and incubated with antibodies. Cover-
slips were mounted in Vectashield mounting medium (Vector Laboratories,
Burlingame, CA) containing the DNA stain DAPI. Images were acquired with
an LSM 780 confocal microscope (Carl Zeiss Microimaging, Jena, Germany).
For data quantification, at least 100 HA (OTULIN)- or GFP (M1-SUB)-positive
cells per condition were counted in each experiment (see the Supplemental
Experimental Procedures).
Mass Spectrometry-Based Analysis of L18-MDP-Induced
Ubiquitination
For SILAC labeling, THP-1 cells were cultured in media containing either
L-arginine and L-lysine or L-arginine-U-13C6-15N4 and L-lysine-U-13C6-
15N2 (Cambridge Isotope Laboratories, Andover, MA) as described previously
(Ong et al., 2002). Ubiquitinated proteins from cells treated for 60min with L18-
MDP were purified using TUBE1 or M1-SUB. The enriched proteins were
resolved by SDS-PAGE and digested in-gel with trypsin. Peptide fractions
were analyzed on a quadrupole Orbitrap (Q-Exactive, Thermo Scientific)
mass spectrometer equipped with a nanoflow HPLC system (Thermo Scienti-
fic, Rockford, IL) as described (Michalski et al., 2011). Raw data files were
analyzed using MaxQuant (Cox and Mann, 2008) (see the Supplemental
Experimental Procedures).
Statistical Analysis
The two-tailed Student’s t test was used to determine statistical significance.
Error bars represent SEM.
SUPPLEMENTAL INFORMATION
Supplemental Information includes five figures, one table, Supplemental
Experimental Procedures, and Supplemental References and can be found
with this article at http://dx.doi.org/10.1016/j.molcel.2013.06.004.
ACKNOWLEDGMENTS
We thank Dr. Henning Walczak for reagents and members of the Ubiquitin
Signaling group for helpful suggestions and reading the manuscript. We thank
the Protein Production Facility at the Novo Nordisk Foundation Center for
Protein Research for production of TUBE and M1-SUB. This work was sup-
ported by a Steno Fellowship from the Danish Council for Independent
Research–Natural Sciences (M.G.-H.), the Lundbeck Foundation (M.G.-H.),
the Medical Research Council (U105192732, D.K.), the European Research
Council (D.K.), the EMBO Young Investigator Programme (D.K.), and the Lister
Institute for Preventive Medicine (to D.K.). The Center for Protein Research is
supported by a grant from the Novo Nordisk Foundation.
Received: May 4, 2013
Revised: June 5, 2013
Accepted: June 6, 2013
Published: June 27, 2013

Molecular Cell
OTULIN Regulates Innate Immune Signaling
REFERENCES
Baud, V., and Karin, M. (2009). Is NF-kappaB a good target for cancer therapy?
Hopes and pitfalls. Nat. Rev. Drug Discov. 8, 33–40.
Bertrand, M.J., Doiron, K., Labbe, K., Korneluk, R.G., Barker, P.A., and Saleh,
M. (2009). Cellular inhibitors of apoptosis cIAP1 and cIAP2 are required for
innate immunity signaling by the pattern recognition receptors NOD1 and
NOD2. Immunity 30, 789–801.
Beug, S.T., Cheung, H.H., LaCasse, E.C., and Korneluk, R.G. (2012).
Modulation of immune signalling by inhibitors of apoptosis. Trends Immunol.
33, 535–545.
Boisson, B., Laplantine, E., Prando, C., Giliani, S., Israelsson, E., Xu, Z.,
Abhyankar, A., Israel, L., Trevejo-Nunez, G., Bogunovic, D., et al. (2012).
Immunodeficiency, autoinflammation and amylopectinosis in humans with
inherited HOIL-1 and LUBAC deficiency. Nat. Immunol. 13, 1178–1186.
Boone, D.L., Turer, E.E., Lee, E.G., Ahmad, R.C., Wheeler, M.T., Tsui, C.,
Hurley, P., Chien, M., Chai, S., Hitotsumatsu, O., et al. (2004). The ubiquitin-
modifying enzyme A20 is required for termination of Toll-like receptor
responses. Nat. Immunol. 5, 1052–1060.
Chen, G., Shaw, M.H., Kim, Y.G., and Nunez, G. (2009). NOD-like receptors:
role in innate immunity and inflammatory disease. Annu. Rev. Pathol. 4,
365–398.
Cox, J., and Mann, M. (2008). MaxQuant enables high peptide identification
rates, individualized p.p.b.-range mass accuracies and proteome-wide
protein quantification. Nat. Biotechnol. 26, 1367–1372.
Damgaard, R.B., Nachbur, U., Yabal, M., Wong, W.W., Fiil, B.K., Kastirr, M.,
Rieser, E., Rickard, J.A., Bankovacki, A., Peschel, C., et al. (2012). The ubiqui-
tin ligase XIAP recruits LUBAC for NOD2 signaling in inflammation and innate
immunity. Mol. Cell 46, 746–758.
Damgaard, R.B., Fiil, B.K., Speckmann, C., Yabal, M., zur Stadt, U., Bekker-
Jensen, S., Jost, P.J., Ehl, S., Mailand, N., and Gyrd-Hansen, M. (2013).
Disease-causing mutations in the XIAP BIR2 domain impair NOD2-dependent
immune signaling. EMBO Mol. Med. http://dx.doi.org/10.1002/emmm.
201303090.
Gerlach, B., Cordier, S.M., Schmukle, A.C., Emmerich, C.H., Rieser, E., Haas,
T.L., Webb, A.I., Rickard, J.A., Anderton, H., Wong, W.W., et al. (2011). Linear
ubiquitination prevents inflammation and regulates immune signalling. Nature
471, 591–596.
Gyrd-Hansen, M., Darding, M., Miasari, M., Santoro, M.M., Zender, L., Xue,
W., Tenev, T., da Fonseca, P.C., Zvelebil, M., Bujnicki, J.M., et al. (2008).
IAPs contain an evolutionarily conserved ubiquitin-binding domain that regu-
lates NF-kappaB as well as cell survival and oncogenesis. Nat. Cell Biol. 10,
1309–1317.
Haas, T.L., Emmerich, C.H., Gerlach, B., Schmukle, A.C., Cordier, S.M.,
Rieser, E., Feltham, R., Vince, J., Warnken, U., Wenger, T., et al. (2009).
Recruitment of the linear ubiquitin chain assembly complex stabilizes the
TNF-R1 signaling complex and is required for TNF-mediated gene induction.
Mol. Cell 36, 831–844.
Harhaj, E.W., and Dixit, V.M. (2012). Regulation of NF-kB by deubiquitinases.
Immunol. Rev. 246, 107–124.
Hasegawa, M., Fujimoto, Y., Lucas, P.C., Nakano, H., Fukase, K., Nunez, G.,
and Inohara, N. (2008). A critical role of RICK/RIP2 polyubiquitination in Nod-
induced NF-kappaB activation. EMBO J. 27, 373–383.
Hjerpe, R., Aillet, F., Lopitz-Otsoa, F., Lang, V., England, P., and Rodriguez,
M.S. (2009). Efficient protection and isolation of ubiquitylated proteins using
tandem ubiquitin-binding entities. EMBO Rep. 10, 1250–1258.
Hospenthal, M.K., Freund, S.M., and Komander, D. (2013). Assembly, analysis
and architecture of atypical ubiquitin chains. Nat. Struct. Mol. Biol. 20,
555–565.
Hunter, A.M., Kottachchi, D., Lewis, J., Duckett, C.S., Korneluk, R.G., and
Liston, P. (2003). A novel ubiquitin fusion system bypasses the mitochondria
and generates biologically active Smac/DIABLO. J. Biol. Chem. 278, 7494–
7499.
M
Ikeda, F., Deribe, Y.L., Skanland, S.S., Stieglitz, B., Grabbe, C., Franz-
Wachtel, M., van Wijk, S.J., Goswami, P., Nagy, V., Terzic, J., et al. (2011).
SHARPIN forms a linear ubiquitin ligase complex regulating NF-kB activity
and apoptosis. Nature 471, 637–641.
Jiang, X., and Chen, Z.J. (2012). The role of ubiquitylation in immune defence
and pathogen evasion. Nat. Rev. Immunol. 12, 35–48.
Keusekotten, K., Elliott, P., Glockner, L., Fiil, B., Damgaard, R., Kulathu, Y.,
Wauer, T., Hospenthal, M., Gyrd-Hansen, M., Krappmann, D., et al. (2013).
OTULIN antagonizes LUBAC signaling by specifically hydrolyzing Met1-linked
polyubiquitin. Cell. http://dx.doi.org/10.1016/j.cell.2013.05.014.
Kirisako, T., Kamei, K.,Murata, S., Kato,M., Fukumoto, H., Kanie, M., Sano, S.,
Tokunaga, F., Tanaka, K., and Iwai, K. (2006). A ubiquitin ligase complex
assembles linear polyubiquitin chains. EMBO J. 25, 4877–4887.
Lee, E.G., Boone, D.L., Chai, S., Libby, S.L., Chien, M., Lodolce, J.P., and Ma,
A. (2000). Failure to regulate TNF-induced NF-kappaB and cell death
responses in A20-deficient mice. Science 289, 2350–2354.
Maloy, K.J., and Powrie, F. (2011). Intestinal homeostasis and its breakdown in
inflammatory bowel disease. Nature 474, 298–306.
Mevissen, T.E.T., Hospenthal, M.K., Geurink, P.P., Elliott, P.R., Akutsu, M.,
Arnaudo, N., Ekkebus, R., Kulathu, Y., Wauer, T., Oualid, F.E., et al. (2013).
OTU deubiquitinases reveal mechanisms of linkage-specificity and enable
ubiquitin chain restriction analysis. Cell. http://dx.doi.org/10.1016/j.cell.2013.
05.046.
Michalski, A., Damoc, E., Hauschild, J.P., Lange, O., Wieghaus, A., Makarov,
A., Nagaraj, N., Cox, J., Mann, M., and Horning, S. (2011). Mass spectrometry-
based proteomics using Q Exactive, a high-performance benchtop quadru-
pole Orbitrap mass spectrometer. Mol. Cell. Proteom. 10. http://dx.doi.org/
10.1074/mcp.M111.011015, M111 011015.
Muller-Rischart, A.K., Pilsl, A., Beaudette, P., Patra, M., Hadian, K., Funke, M.,
Peis, R., Deinlein, A., Schweimer, C., Kuhn, P.H., et al. (2013). The E3 ligase
parkin maintains mitochondrial integrity by increasing linear ubiquitination of
NEMO. Mol. Cell 49, 908–921.
Ong, S.E., Blagoev, B., Kratchmarova, I., Kristensen, D.B., Steen, H., Pandey,
A., and Mann, M. (2002). Stable isotope labeling by amino acids in cell culture,
SILAC, as a simple and accurate approach to expression proteomics. Mol.
Cell. Proteomics 1, 376–386.
Rahighi, S., Ikeda, F., Kawasaki, M., Akutsu, M., Suzuki, N., Kato, R., Kensche,
T., Uejima, T., Bloor, S., Komander, D., et al. (2009). Specific recognition of
linear ubiquitin chains by NEMO is important for NF-kappaB activation. Cell
136, 1098–1109.
Rivkin, E., Almeida, S.M., Ceccarelli, D.F., Juang, Y., MacLean, T.A., Srikumar,
T., Huang, H., Dunham, W.H., Fukumura, R., Xie, G., et al. (2013). The linear
ubiquitin-specific deubiquitinase gumby regulates angiogenesis. Nature.
http://dx.doi.org/10.1038/nature12296.
Stieglitz, B., Morris-Davies, A.C., Koliopoulos, M.G., Christodoulou, E., and
Rittinger, K. (2012). LUBAC synthesizes linear ubiquitin chains via a thioester
intermediate. EMBO Rep. 13, 840–846.
Takeuchi, O., and Akira, S. (2010). Pattern recognition receptors and inflam-
mation. Cell 140, 805–820.
Tokunaga, F., and Iwai, K. (2012). LUBAC, a novel ubiquitin ligase for linear
ubiquitination, is crucial for inflammation and immune responses. Microbes
Infect. 14, 563–572.
Tokunaga, F., Sakata, S., Saeki, Y., Satomi, Y., Kirisako, T., Kamei, K.,
Nakagawa, T., Kato, M., Murata, S., Yamaoka, S., et al. (2009). Involvement
of linear polyubiquitylation of NEMO in NF-kappaB activation. Nat. Cell Biol.
11, 123–132.
Tokunaga, F., Nakagawa, T., Nakahara, M., Saeki, Y., Taniguchi, M., Sakata,
S., Tanaka, K., Nakano, H., and Iwai, K. (2011). SHARPIN is a component of
the NF-kB-activating linear ubiquitin chain assembly complex. Nature 471,
633–636.
Walczak, H., Iwai, K., and Dikic, I. (2012). Generation and physiological roles of
linear ubiquitin chains. BMC Biol. 10, 23.
olecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc. 829

Molecular Cell
OTULIN Regulates Innate Immune Signaling
van Wijk, S.J., Fiskin, E., Putyrski, M., Pampaloni, F., Hou, J., Wild, P.,
Kensche, T., Grecco, H.E., Bastiaens, P., and Dikic, I. (2012). Fluorescence-
based sensors to monitor localization and functions of linear and K63-linked
ubiquitin chains in cells. Mol. Cell 47, 797–809.
Warner, N., Burberry, A., Franchi, L., Kim, Y.G., McDonald, C., Sartor, M.A.,
and Nunez, G. (2013). A genome-wide siRNA screen reveals positive and
negative regulators of the NOD2 and NF-kB signaling pathways. Sci. Signal.
6. http://dx.doi.org/10.1126/scisignal.2003305, rs3.
830 Molecular Cell 50, 818–830, June 27, 2013 ª2013 Elsevier Inc.
Wertz, I.E., O’Rourke, K.M., Zhou, H., Eby, M., Aravind, L., Seshagiri, S., Wu,
P., Wiesmann, C., Baker, R., Boone, D.L., et al. (2004). De-ubiquitination and
ubiquitin ligase domains of A20 downregulate NF-kappaB signalling. Nature
430, 694–699.
Wu, C.J., Conze, D.B., Li, T., Srinivasula, S.M., and Ashwell, J.D. (2006).
Sensing of Lys 63-linked polyubiquitination by NEMO is a key event in
NF-kappaB activation [corrected]. Nat. Cell Biol. 8, 398–406.





























