Embed Size (px)
Citation preview

Cd
ED
a
ARRAA
KAECAM
1
meociiotim(r[
fitaTo
p
vK
0h
Molecular & Biochemical Parasitology 185 (2012) 121– 126
Contents lists available at SciVerse ScienceDirect
Molecular & Biochemical Parasitology
ysteine protease involving in autophagosomal degradation of mitochondriauring encystation of Acanthamoeba
un-Kyung Moon, Yeonchul Hong, Dong-Il Chung, Hyun-Hee Kong ∗
epartment of Parasitology, Kyungpook National University School of Medicine, Taegu, Republic of Korea
r t i c l e i n f o
rticle history:eceived 12 June 2012eceived in revised form 24 July 2012ccepted 30 July 2012vailable online 14 August 2012
a b s t r a c t
Using the microarray to identify encystation mediating factors, significantly higher expression of a cys-teine protease gene was observed in cysts, compared with trophozoites. Results of real-time PCR analysisalso showed a magnificent increase of cysteine protease levels during encystation of Acanthamoeba. Wenamed the gene cyst specific cysteine protease (cscp) of Acanthamoeba. The purified recombinant pro-tein of CSCP showed activities of papain and cathepsin B against the substrates. During encystation, EGFP
eywords:canthamoebancystationysteine protease
fused CSCP showed colocalization with LysoTracker, an autophagosome marker, in transiently transfectedamoeba. Amoeba transfected with siRNA against cscp was unable to form mature cysts. Undigested mito-chondria in vacuole like structures were observed in cscp siRNA treated cells by transmission electronmicroscopy. These results provide evidence of the important role of CSCP in autophagosomal degradation
cularl
utophagosomeitochondriaof cell constituents, parti
. Introduction
Encystation is one of the most important processes to be deter-ined in Acanthamoeba infection, such as granulomatous amoebic
ncephalitis (GAE) and amebic keratitis (AK) [19]. Transformationf Acanthamoeba trophozoites to cysts occurs under unfavorableonditions, conveying resistance to pH, high temperature, and var-ous biocides [3,18]. Due to the resistant nature of the cyst, theres hardly an effective treatment against Acanthamoeba infection. Inrder to achieve a better understanding of the mechanism of encys-ation, characterization of various encystation mediating factors,ncluding the cyst specific protein 21 (CSP21) gene, encystation
ediating serine protease (EMSP), autophagy related protein 8AcAtg8), autophagy related protein 3 (AcAtg3), and autophagyelated protein 16 (AcAtg16L) of Acanthamoeba was performed2,20,23,24,32].
Results of an analysis of the differential gene expression pro-le between cysts and trophozoites of Acanthamoeba showed thathe “O” article (posttranslational modification, protein turnover,nd chaperones) was highly populated in cyst specific ESTs [21].his result suggested that various proteases mediate encystation
f Acanthamoeba.Acanthamoeba includes four major classes of proteases; asparticrotease, cysteine protease, serine protease, and metalloprotease.
∗ Corresponding author at: Department of Parasitology, Kyungpook National Uni-ersity School of Medicine, 101 Dongin-dong, Joong-gu, Taegu 700-422, Republic oforea. Tel.: +82 53 420 4882; fax: +82 53 422 9330.
E-mail address: [email protected] (H.-H. Kong).
166-6851/$ – see front matter © 2012 Elsevier B.V. All rights reserved.ttp://dx.doi.org/10.1016/j.molbiopara.2012.07.008
y mitochondria, during encystation of Acanthamoeba.© 2012 Elsevier B.V. All rights reserved.
In almost genotypes, serine protease is the most abundant andwell reported for degradation of ECM (extracellular matrix) com-ponents in Acanthamoeba infection [8,11,13]. Metalloproteasehas been reported to exhibit properties of ECM degradation[31]. However, data on cysteine protease are limited, andonly the possibility that they may play a role in intracel-lular protein degradation or phagocytosis has been reported[9]. The role of cysteine protease during the early phaseof encystment in Acanthamoeba has recently been suggested[15].
Roles of cysteine protease in cell differentiation of severalother protozoan parasites have been reported. Specific expres-sion of cysteine peptidase was observed during encystation inEntamoeba invadens [4]. Production of oocysts by Plasmodiumfalciparum was inhibited by cysteine protease inhibitor E64d[5], and the role of cysteine proteases of Trypanosoma bruceibloodstream form in surface coat exchange during differenti-ation has been reported [29]. Necessity of lysosomal cysteinepeptidases for autophagy and differentiation in Leishmania mex-icana has been reported [35]. Findings from these reportsdemonstrated the critical roles of cysteine protease in cell differ-entiation.
Of particular interest, results of microarray analysis betweencysts and trophozoites of Acanthamoeba showed 282-fold expres-sion of a type of cysteine protease in cysts [22]. Cyst specific ESTby KOG (eukaryotic orthologous groups) analysis also revealed a
similar result, where 11 copies of the cysteine protease were iden-tified only in cyst ESTs [21]. These results suggest that the cysteineprotease may play an important role during encystation in Acan-thamoeba.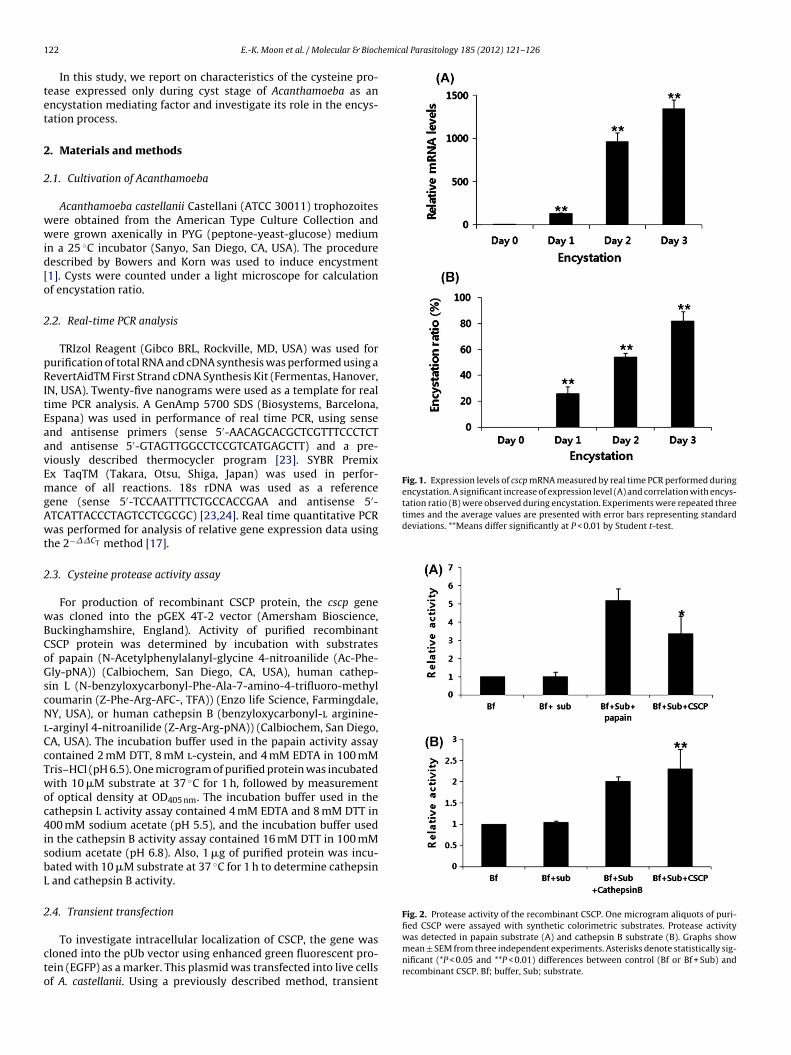
1 emical Parasitology 185 (2012) 121– 126
tet
2
2
wwid[o
2
pRItEaavEmgAwt
2
wBCoGscNlCcTwoc4isbL
2
cto
Fig. 1. Expression levels of cscp mRNA measured by real time PCR performed duringencystation. A significant increase of expression level (A) and correlation with encys-tation ratio (B) were observed during encystation. Experiments were repeated threetimes and the average values are presented with error bars representing standarddeviations. **Means differ significantly at P < 0.01 by Student t-test.
Fig. 2. Protease activity of the recombinant CSCP. One microgram aliquots of puri-fied CSCP were assayed with synthetic colorimetric substrates. Protease activity
22 E.-K. Moon et al. / Molecular & Bioch
In this study, we report on characteristics of the cysteine pro-ease expressed only during cyst stage of Acanthamoeba as anncystation mediating factor and investigate its role in the encys-ation process.
. Materials and methods
.1. Cultivation of Acanthamoeba
Acanthamoeba castellanii Castellani (ATCC 30011) trophozoitesere obtained from the American Type Culture Collection andere grown axenically in PYG (peptone-yeast-glucose) medium
n a 25 ◦C incubator (Sanyo, San Diego, CA, USA). The procedureescribed by Bowers and Korn was used to induce encystment1]. Cysts were counted under a light microscope for calculationf encystation ratio.
.2. Real-time PCR analysis
TRIzol Reagent (Gibco BRL, Rockville, MD, USA) was used forurification of total RNA and cDNA synthesis was performed using aevertAidTM First Strand cDNA Synthesis Kit (Fermentas, Hanover,
N, USA). Twenty-five nanograms were used as a template for realime PCR analysis. A GenAmp 5700 SDS (Biosystems, Barcelona,spana) was used in performance of real time PCR, using sensend antisense primers (sense 5′-AACAGCACGCTCGTTTCCCTCTnd antisense 5′-GTAGTTGGCCTCCGTCATGAGCTT) and a pre-iously described thermocycler program [23]. SYBR Premixx TaqTM (Takara, Otsu, Shiga, Japan) was used in perfor-ance of all reactions. 18s rDNA was used as a reference
ene (sense 5′-TCCAATTTTCTGCCACCGAA and antisense 5′-TCATTACCCTAGTCCTCGCGC) [23,24]. Real time quantitative PCRas performed for analysis of relative gene expression data using
he 2−��CT method [17].
.3. Cysteine protease activity assay
For production of recombinant CSCP protein, the cscp geneas cloned into the pGEX 4T-2 vector (Amersham Bioscience,uckinghamshire, England). Activity of purified recombinantSCP protein was determined by incubation with substratesf papain (N-Acetylphenylalanyl-glycine 4-nitroanilide (Ac-Phe-ly-pNA)) (Calbiochem, San Diego, CA, USA), human cathep-in L (N-benzyloxycarbonyl-Phe-Ala-7-amino-4-trifluoro-methyloumarin (Z-Phe-Arg-AFC-, TFA)) (Enzo life Science, Farmingdale,Y, USA), or human cathepsin B (benzyloxycarbonyl-l arginine-
-arginyl 4-nitroanilide (Z-Arg-Arg-pNA)) (Calbiochem, San Diego,A, USA). The incubation buffer used in the papain activity assayontained 2 mM DTT, 8 mM l-cystein, and 4 mM EDTA in 100 mMris–HCl (pH 6.5). One microgram of purified protein was incubatedith 10 �M substrate at 37 ◦C for 1 h, followed by measurement
f optical density at OD405 nm. The incubation buffer used in theathepsin L activity assay contained 4 mM EDTA and 8 mM DTT in00 mM sodium acetate (pH 5.5), and the incubation buffer used
n the cathepsin B activity assay contained 16 mM DTT in 100 mModium acetate (pH 6.8). Also, 1 �g of purified protein was incu-ated with 10 �M substrate at 37 ◦C for 1 h to determine cathepsin
and cathepsin B activity.
.4. Transient transfection
To investigate intracellular localization of CSCP, the gene wasloned into the pUb vector using enhanced green fluorescent pro-ein (EGFP) as a marker. This plasmid was transfected into live cellsf A. castellanii. Using a previously described method, transient
was detected in papain substrate (A) and cathepsin B substrate (B). Graphs showmean ± SEM from three independent experiments. Asterisks denote statistically sig-nificant (*P < 0.05 and **P < 0.01) differences between control (Bf or Bf + Sub) andrecombinant CSCP. Bf; buffer, Sub; substrate.
E.-K. Moon et al. / Molecular & Biochemical Parasitology 185 (2012) 121– 126 123
F in cyo whichs
t(
2
lb(Gami
2
bempo
2
loawt
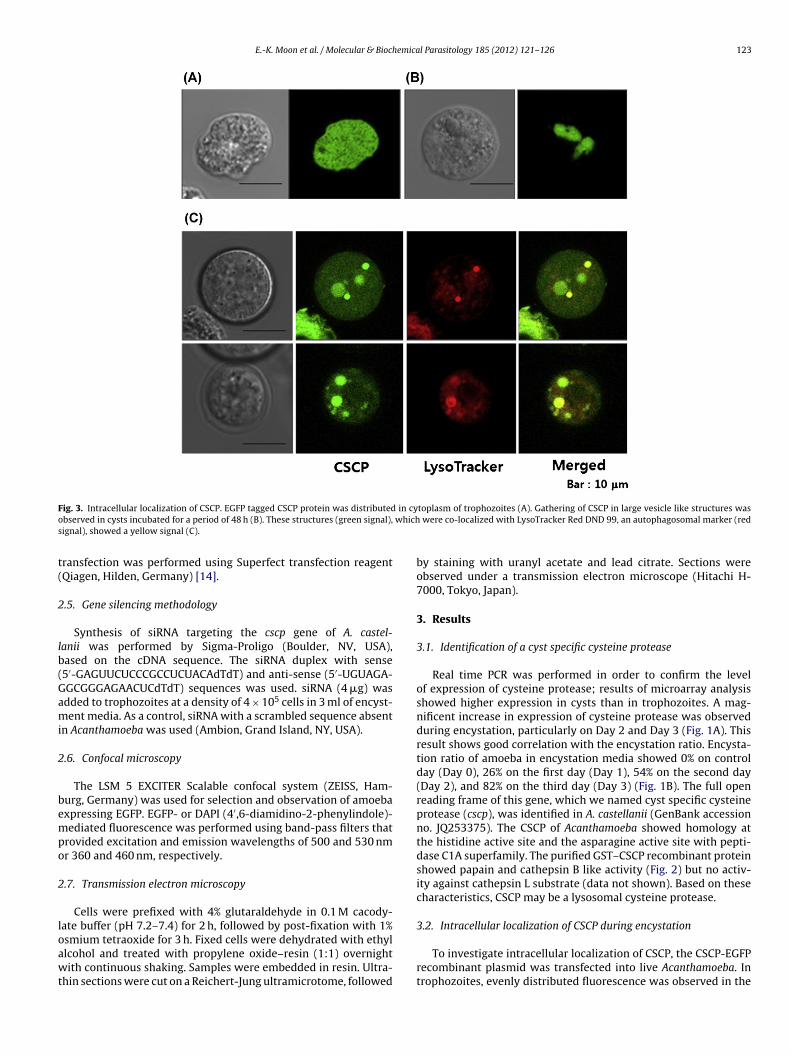
ig. 3. Intracellular localization of CSCP. EGFP tagged CSCP protein was distributedbserved in cysts incubated for a period of 48 h (B). These structures (green signal),ignal), showed a yellow signal (C).
ransfection was performed using Superfect transfection reagentQiagen, Hilden, Germany) [14].
.5. Gene silencing methodology
Synthesis of siRNA targeting the cscp gene of A. castel-anii was performed by Sigma-Proligo (Boulder, NV, USA),ased on the cDNA sequence. The siRNA duplex with sense5′-GAGUUCUCCCGCCUCUACAdTdT) and anti-sense (5′-UGUAGA-GCGGGAGAACUCdTdT) sequences was used. siRNA (4 �g) wasdded to trophozoites at a density of 4 × 105 cells in 3 ml of encyst-ent media. As a control, siRNA with a scrambled sequence absent
n Acanthamoeba was used (Ambion, Grand Island, NY, USA).
.6. Confocal microscopy
The LSM 5 EXCITER Scalable confocal system (ZEISS, Ham-urg, Germany) was used for selection and observation of amoebaxpressing EGFP. EGFP- or DAPI (4′,6-diamidino-2-phenylindole)-ediated fluorescence was performed using band-pass filters that
rovided excitation and emission wavelengths of 500 and 530 nmr 360 and 460 nm, respectively.
.7. Transmission electron microscopy
Cells were prefixed with 4% glutaraldehyde in 0.1 M cacody-ate buffer (pH 7.2–7.4) for 2 h, followed by post-fixation with 1%
smium tetraoxide for 3 h. Fixed cells were dehydrated with ethyllcohol and treated with propylene oxide–resin (1:1) overnightith continuous shaking. Samples were embedded in resin. Ultra-hin sections were cut on a Reichert-Jung ultramicrotome, followed
toplasm of trophozoites (A). Gathering of CSCP in large vesicle like structures was were co-localized with LysoTracker Red DND 99, an autophagosomal marker (red
by staining with uranyl acetate and lead citrate. Sections wereobserved under a transmission electron microscope (Hitachi H-7000, Tokyo, Japan).
3. Results
3.1. Identification of a cyst specific cysteine protease
Real time PCR was performed in order to confirm the levelof expression of cysteine protease; results of microarray analysisshowed higher expression in cysts than in trophozoites. A mag-nificent increase in expression of cysteine protease was observedduring encystation, particularly on Day 2 and Day 3 (Fig. 1A). Thisresult shows good correlation with the encystation ratio. Encysta-tion ratio of amoeba in encystation media showed 0% on controlday (Day 0), 26% on the first day (Day 1), 54% on the second day(Day 2), and 82% on the third day (Day 3) (Fig. 1B). The full openreading frame of this gene, which we named cyst specific cysteineprotease (cscp), was identified in A. castellanii (GenBank accessionno. JQ253375). The CSCP of Acanthamoeba showed homology atthe histidine active site and the asparagine active site with pepti-dase C1A superfamily. The purified GST–CSCP recombinant proteinshowed papain and cathepsin B like activity (Fig. 2) but no activ-ity against cathepsin L substrate (data not shown). Based on thesecharacteristics, CSCP may be a lysosomal cysteine protease.
3.2. Intracellular localization of CSCP during encystation
To investigate intracellular localization of CSCP, the CSCP-EGFPrecombinant plasmid was transfected into live Acanthamoeba. Introphozoites, evenly distributed fluorescence was observed in the
1 emical Parasitology 185 (2012) 121– 126
ctfl4ca
3
tsstetc
3
ttcoimwbimccegmuf(fpA
4
crmtcoAdptcsz
t2
pc
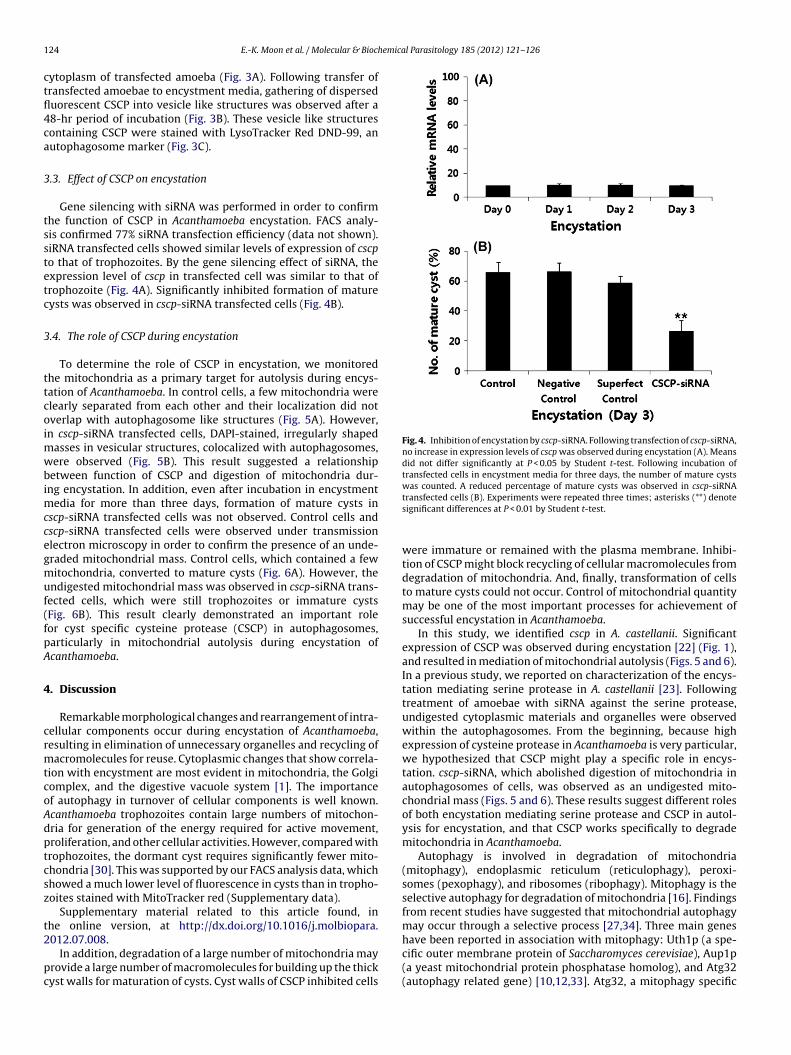
Fig. 4. Inhibition of encystation by cscp-siRNA. Following transfection of cscp-siRNA,no increase in expression levels of cscp was observed during encystation (A). Meansdid not differ significantly at P < 0.05 by Student t-test. Following incubation oftransfected cells in encystment media for three days, the number of mature cystswas counted. A reduced percentage of mature cysts was observed in cscp-siRNA
24 E.-K. Moon et al. / Molecular & Bioch
ytoplasm of transfected amoeba (Fig. 3A). Following transfer ofransfected amoebae to encystment media, gathering of disperseduorescent CSCP into vesicle like structures was observed after a8-hr period of incubation (Fig. 3B). These vesicle like structuresontaining CSCP were stained with LysoTracker Red DND-99, anutophagosome marker (Fig. 3C).
.3. Effect of CSCP on encystation
Gene silencing with siRNA was performed in order to confirmhe function of CSCP in Acanthamoeba encystation. FACS analy-is confirmed 77% siRNA transfection efficiency (data not shown).iRNA transfected cells showed similar levels of expression of cscpo that of trophozoites. By the gene silencing effect of siRNA, thexpression level of cscp in transfected cell was similar to that ofrophozoite (Fig. 4A). Significantly inhibited formation of matureysts was observed in cscp-siRNA transfected cells (Fig. 4B).
.4. The role of CSCP during encystation
To determine the role of CSCP in encystation, we monitoredhe mitochondria as a primary target for autolysis during encys-ation of Acanthamoeba. In control cells, a few mitochondria werelearly separated from each other and their localization did notverlap with autophagosome like structures (Fig. 5A). However,n cscp-siRNA transfected cells, DAPI-stained, irregularly shaped
asses in vesicular structures, colocalized with autophagosomes,ere observed (Fig. 5B). This result suggested a relationship
etween function of CSCP and digestion of mitochondria dur-ng encystation. In addition, even after incubation in encystment
edia for more than three days, formation of mature cysts inscp-siRNA transfected cells was not observed. Control cells andscp-siRNA transfected cells were observed under transmissionlectron microscopy in order to confirm the presence of an unde-raded mitochondrial mass. Control cells, which contained a fewitochondria, converted to mature cysts (Fig. 6A). However, the
ndigested mitochondrial mass was observed in cscp-siRNA trans-ected cells, which were still trophozoites or immature cystsFig. 6B). This result clearly demonstrated an important roleor cyst specific cysteine protease (CSCP) in autophagosomes,articularly in mitochondrial autolysis during encystation ofcanthamoeba.
. Discussion
Remarkable morphological changes and rearrangement of intra-ellular components occur during encystation of Acanthamoeba,esulting in elimination of unnecessary organelles and recycling ofacromolecules for reuse. Cytoplasmic changes that show correla-
ion with encystment are most evident in mitochondria, the Golgiomplex, and the digestive vacuole system [1]. The importancef autophagy in turnover of cellular components is well known.canthamoeba trophozoites contain large numbers of mitochon-ria for generation of the energy required for active movement,roliferation, and other cellular activities. However, compared withrophozoites, the dormant cyst requires significantly fewer mito-hondria [30]. This was supported by our FACS analysis data, whichhowed a much lower level of fluorescence in cysts than in tropho-oites stained with MitoTracker red (Supplementary data).
Supplementary material related to this article found, inhe online version, at http://dx.doi.org/10.1016/j.molbiopara.
012.07.008.In addition, degradation of a large number of mitochondria mayrovide a large number of macromolecules for building up the thickyst walls for maturation of cysts. Cyst walls of CSCP inhibited cells
transfected cells (B). Experiments were repeated three times; asterisks (**) denotesignificant differences at P < 0.01 by Student t-test.
were immature or remained with the plasma membrane. Inhibi-tion of CSCP might block recycling of cellular macromolecules fromdegradation of mitochondria. And, finally, transformation of cellsto mature cysts could not occur. Control of mitochondrial quantitymay be one of the most important processes for achievement ofsuccessful encystation in Acanthamoeba.
In this study, we identified cscp in A. castellanii. Significantexpression of CSCP was observed during encystation [22] (Fig. 1),and resulted in mediation of mitochondrial autolysis (Figs. 5 and 6).In a previous study, we reported on characterization of the encys-tation mediating serine protease in A. castellanii [23]. Followingtreatment of amoebae with siRNA against the serine protease,undigested cytoplasmic materials and organelles were observedwithin the autophagosomes. From the beginning, because highexpression of cysteine protease in Acanthamoeba is very particular,we hypothesized that CSCP might play a specific role in encys-tation. cscp-siRNA, which abolished digestion of mitochondria inautophagosomes of cells, was observed as an undigested mito-chondrial mass (Figs. 5 and 6). These results suggest different rolesof both encystation mediating serine protease and CSCP in autol-ysis for encystation, and that CSCP works specifically to degrademitochondria in Acanthamoeba.
Autophagy is involved in degradation of mitochondria(mitophagy), endoplasmic reticulum (reticulophagy), peroxi-somes (pexophagy), and ribosomes (ribophagy). Mitophagy is theselective autophagy for degradation of mitochondria [16]. Findingsfrom recent studies have suggested that mitochondrial autophagymay occur through a selective process [27,34]. Three main geneshave been reported in association with mitophagy: Uth1p (a spe-
cific outer membrane protein of Saccharomyces cerevisiae), Aup1p(a yeast mitochondrial protein phosphatase homolog), and Atg32(autophagy related gene) [10,12,33]. Atg32, a mitophagy specific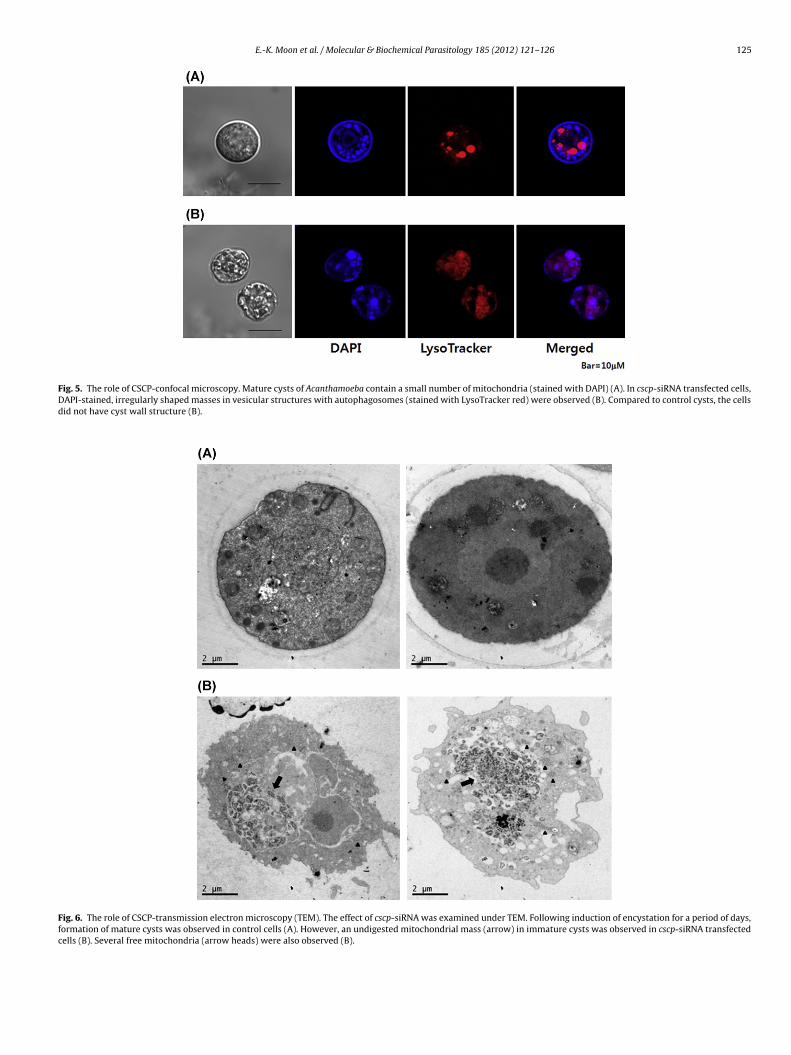
E.-K. Moon et al. / Molecular & Biochemical Parasitology 185 (2012) 121– 126 125
Fig. 5. The role of CSCP-confocal microscopy. Mature cysts of Acanthamoeba contain a small number of mitochondria (stained with DAPI) (A). In cscp-siRNA transfected cells,DAPI-stained, irregularly shaped masses in vesicular structures with autophagosomes (stained with LysoTracker red) were observed (B). Compared to control cysts, the cellsdid not have cyst wall structure (B).
Fig. 6. The role of CSCP-transmission electron microscopy (TEM). The effect of cscp-siRNA was examined under TEM. Following induction of encystation for a period of days,formation of mature cysts was observed in control cells (A). However, an undigested mitochondrial mass (arrow) in immature cysts was observed in cscp-siRNA transfectedcells (B). Several free mitochondria (arrow heads) were also observed (B).

1 emica
rbpmcdTkaeom
ublblmbac
ittc[AtpthpeoApmb
A
fm2
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[sion and selective fusion govern mitochondrial segregation and elimination byautophagy. EMBO Journal 2008;27:433–46.
26 E.-K. Moon et al. / Molecular & Bioch
eceptor in yeast, is important for recruitment of mitochondriay macroautophagy. PINK1 (a mitochondrial serine/threoninerotein kinase) is associated with specific recruitment of Parkin (aultiprotein E3 ubiquitin ligase complex), which promotes mito-
hondrial autophagy [25,26]. However, an enzyme for selectiveegradation of substrate in mitophagy has yet to be discovered.he mechanism of selective degradation of mitochondria is notnown in Acanthamoeba; however, data on siRNA against cscp inssociation with mitochondrial degradation provide importantvidence of mitophagy in Acanthamoeba. Findings from our previ-us report on the Atg16L system also provide strong evidence ofitophagy in encysting Acanthamoeba [32].Intracellular trafficking of autophagosomal enzymes remains
nknown. In general, mediation of transport of lysosomal proteinsy mannose 6-phosphate receptors has been reported [7]. Regu-
ated transport of cysteine protease to lysosomes in E. histolyticay Rab7 isotype has been reported [28]. Mediated transport of
ysozymes and �-hexosaminidase of E. histolytica by a novel trans-embrane receptor in phagosome transport, cysteine protease
inding protein family 8 (CPBF8), has been reported [6]. The mech-nism of intracellular trafficking to autophagosomes of amoebicysteine protease, involving CSCP, should be demonstrated.
The prominent role of proteolytic activity during encystations in accordance with autophagic processes in encysting Acan-hamoeba [1]. Results of cyst specific ESTs analysis also showed thathe ‘O’ article (posttranslational modification, protein turnover, andhaperones) possessed the highest percentage of cyst specific ESTs21]. These results suggested the necessity of various proteases ofcanthamoeba in regulation of autophagic mechanisms in forma-ion of a mature cyst. Although the majority of research on therotease in Acanthamoeba has focused on pathogenesis, interest inhe role of protease in association with encystation or excystationas increased in recent years. We supposed that various proteaseslay important roles in regulation of autophagic processes. Knowl-dge of the role of CSCP could lead to an expanded understandingf autophagic mechanisms in cyst forming protozoa, includingcanthamoeba, during encystation. In order to understand the com-lete mechanism of Acanthamoeba encystation, investigation ofitophagy related genes and genes controlling CSCP activity will
e needed.
cknowledgments
This work was supported by a Korea Research Foundation Grantunded by the Korean Government (MOEHRD, Basic Research Pro-
otion Fund) (2010-0003606) and the Brain Korea 21 Project in011.
eferences
[1] Bowers B, Korn ED. The fine structure of Acanthamoeba castellanii (Neff strain).II. Encystment. Journal of Cell Biology 1969;41:786–805.
[2] Chen L, Orfeo T, Gilmartin G, Bateman E. Mechanism of cyst specific protein 21mRNA induction during Acanthamoeba differentiation. Biochimica et Biophys-ica Acta 2004;1691:23–31.
[3] Coulon C, Collignon A, McDonnell G, Thomas V. Resistance of Acanthamoebacysts to disinfection treatments used in health care settings. Journal of ClinicalMicrobiology 2010;48:2689–97.
[4] Ebert F, Bachmann A, Nakada-Tsukui K, Hennings I, Drescher B, Nozaki T, et al.An Entamoeba cysteine peptidase specifically expressed during encystation.Parasitology International 2008;57:521–4.
[5] Eksi S, Czesny B, van Gemert GJ, Sauerwein RW, Eling W, Williamson KC. Inhi-bition of Plasmodium falciparum oocyst production by membrane-permeantcysteine protease inhibitor E64d. Antimicrobial Agents and Chemotherapy2007;51:1064–70.
[6] Furukawa A, Nakada-Tsukui K, Nozaki T. Novel transmembrane receptorinvolved in phagosome transport of lysozymes and �-hexosaminidase in theenteric protozoan Entamoeba histolytica. PLoS Pathogens 2012;8:e1002539.
[7] Ghosh P, Dahms NM, Kornfeld S. Mannose 6-phosphate receptors: new twistsin the tale. Nature Reviews Molecular Cell Biology 2003;4:202–12 [review].
[
l Parasitology 185 (2012) 121– 126
[8] Hong YC, Kong HH, Ock MS, Kim IS, Chung DI. Isolation and characterization of acDNA encoding a subtilisin-like serine proteinase (ahSUB) from Acanthamoebahealyi. Molecular and Biochemical Parasitology 2000;111:441–6.
[9] Hong YC, Hwang MY, Yun HC, Yu HS, Kong HH, Yong TS, et al. Isolationand characterization of a cDNA encoding a mammalian cathepsin L-like cys-teine proteinase from Acanthamoeba healyi. Korean Journal of Parasitology2002;40:17–24.
10] Kanki T, Wang K, Cao Y, Baba M, Klionsky DJ. Atg32 is a mitochon-drial protein that confers selectivity during mitophagy. Developmental Cell2009;17:98–109.
11] Khan NA. Acanthamoeba: biology and increasing importance in human health.FEMS Microbiology Review 2006;30:564–95 [review].
12] Kissová I, Deffieu M, Manon S, Camougrand N. Uth1p is involved in theautophagic degradation of mitochondria. Journal of Biological Chemistry2004;279:39068–74.
13] Kong HH, Kim TH, Chung DI. Purification and characterization of a secretoryserine proteinase of Acanthamoeba healyi isolated from GAE. Journal of Para-sitology 2000;86:12–7.
14] Kong HH, Pollard TD. Intracellular localization and dynamics of myosin-II andmyosin-IC in live Acanthamoeba by transient transfection of EGFP fusion pro-teins. Journal of Cell Science 2002;115:4993–5002.
15] Leitsch D, Köhsler M, Marchetti-Deschmann M, Deutsch A, Allmaier G, DuchêneM, et al. Major role for cysteine proteases during the early phase of Acan-thamoeba castellanii encystment. Eukaryot Cell 2010;9:611–8.
16] Lemasters JJ. Selective mitochondrial autophagy, or mitophagy, as a targeteddefense against oxidative stress, mitochondrial dysfunction, and aging. Reju-venation Research 2005;8:3–5.
17] Livak KJ, Schmittgen TD. Analysis of relative gene expression data usingreal-time quantitative PCR and the 2(−Delta Delta C(T)) Method. Methods2001;25:402–8.
18] Lloyd D, Turner NA, Khunkitti W, Hann AC, Furr JR, Russell AD. Encystation inAcanthamoeba castellanii: development of biocide resistance. Journal of Eukary-otic Microbiology 2001;48:11–6 [review].
19] Marciano-Cabral F, Cabral G. Acanthamoeba spp. as agents of disease in humans.Clinical Microbiology Reviews 2003;16:273–307 [review].
20] Moon EK, Chung DI, Hong Y, Kong HH. Atg3-mediated lipidation of Atg8is involved in encystation of Acanthamoeba. Korean Journal of Parasitology2011;49:103–8.
21] Moon EK, Chung DI, Hong Y, Kong HH. Expression levels of encystation medi-ating factors in fresh strain of Acanthamoeba castellanii cyst ESTs. ExperimentalParasitology 2011;127:811–6.
22] Moon EK, Xuan YH, Chung DI, Hong Y, Kong HH. Microarray analysis of dif-ferentially expressed genes between cysts and trophozoites of Acanthamoebacastellanii. Korean Journal of Parasitology 2011;49:341–7.
23] Moon EK, Chung DI, Hong YC, Kong HH. Characterization of a serine proteinasemediating encystation of Acanthamoeba. Eukaryotic Cell 2008;7:1513–7.
24] Moon EK, Chung DI, Hong YC, Kong HH. Autophagy protein 8 mediatingautophagosome in encysting Acanthamoeba. Molecular and Biochemical Par-asitology 2009;168:43–8.
25] Narendra D, Tanaka A, Suen DF, Youle RJ. Parkin is recruited selectively toimpaired mitochondria and promotes their autophagy. Journal of Cell Biology2008;183:795–803.
26] Narendra DP, Jin SM, Tanaka A, Suen DF, Gautier CA, Shen J, et al. PINK1 isselectively stabilized on impaired mitochondria to activate Parkin. PLoS Biology2010;8:e1000298.
27] Nowikovsky K, Reipert S, Devenish RJ, Schweyen RJ. Mdm38 protein depletioncauses loss of mitochondrial K+/H+ exchange activity, osmotic swelling andmitophagy. Cell Death and Differentiation 2007;14:1647–56.
28] Saito-Nakano Y, Mitra BN, Nakada-Tsukui K, Sato D, Nozaki T. Two Rab7 iso-types, EhRab7A and EhRab7B, play distinct roles in biogenesis of lysosomes andphagosomes in the enteric protozoan parasite Entamoeba histolytica. CellularMicrobiology 2007;9:1796–808.
29] Santos CC, Coombs GH, Lima AP, Mottram JC. Role of the Trypanosoma bru-cei natural cysteine peptidase inhibitor ICP in differentiation and virulence.Molecular Microbiology 2007;66:991–1002.
30] Siddiqui R, Khan NA. Acanthamoeba is an evolutionary ancestor ofmacrophages: a myth or reality? Experimental Parasitology 2012;130:95–7[review].
31] Sissons J, Kim KS, Stins M, Jayasekera S, Alsam S, Khan NA. Acanthamoeba castel-lanii induces host cell death via a phosphatidylinositol 3-kinase-dependentmchanism. Infection and Immunity 2005;73:2704–8.
32] Song SM, Han BI, Moon EK, Lee YR, Yu HS, Jha BK, et al. Autophagy protein 16-mediated autophagy is required for the encystation of Acanthamoeba castellanii.Molecular and Biochemical Parasitology 2012;183:158–65.
33] Tal R, Winter G, Ecker N, Klionsky DJ, Abeliovich H. Aup1p, a yeastmitochondrial protein phosphatase homolog, is required for efficient sta-tionary phase mitophagy and cell survival. Journal of Biological Chemistry2007;282:5617–24.
34] Twig G, Elorza A, Molina AJ, Mohamed H, Wikstrom JD, Walzer G, et al. Fis-
35] Williams RA, Tetley L, Mottram JC, Coombs GH. Cysteine peptidases CPA andCPB are vital for autophagy and differentiation in Leishmania mexicana. Molec-ular Microbiology 2006;61:655–74.