Embed Size (px)
Citation preview

A
piet(s©
K
1
tssmnclicdt(tlf
0d
Cell Calcium 42 (2007) 133–144
Molecular basis of the CRAC channel
Michael D. Cahalan a,∗, Shenyuan L. Zhang a, Andriy V. Yeromin a,Kari Ohlsen b, Jack Roos b,1, Kenneth A. Stauderman b,1
a Department of Physiology and Biophysics and Center for Immunology, University of California, Irvine, CA 92697, United Statesb TorreyPines Therapeutics Inc., La Jolla, CA 92037, United States
Received 1 March 2007; received in revised form 13 March 2007; accepted 14 March 2007Available online 7 May 2007
bstract
Ca2+ release-activated Ca2+ (CRAC) channels, located in the plasma membrane, are opened upon release of Ca2+ from intracellular stores,ermitting Ca2+ entry and sustained [Ca2+]i signaling that replenishes the store in numerous cell types. This mechanism is particularly importantn T lymphocytes of the immune system, providing the missing link in the signal transduction cascade that is initiated by T cell receptorngagement and leads to altered expression of genes that results ultimately in the production of cytokines and cell proliferation. In the past
hree years, RNA interference screens together with over-expression and site-directed mutagenesis have identified the triggering moleculeStim) that links store depletion to CRAC channel-mediated Ca2+ influx and the pore subunit (Orai) of the CRAC channel that allows highlyelective entry of Ca2+ ions into cells.2007 Published by Elsevier Ltd.
rai
bgaacatc
1i
pa
eywords: CRAC channel; T lymphocyte; Store-operated calcium; Stim; O
. Introduction
This review will cover the discovery of Stim and Orai andheir functional roles in store-operated Ca2+ influx, empha-izing our own contributions. Up front, we recognize thateveral other groups have been engaged in a similar quest;any of their key contributions are represented in accompa-
ying articles in this series. We introduce the topic by firstonsidering background information on calcium signaling inymphocytes, the electrical description of CRAC current, andts physiological role in lymphocytes. Then we discuss how aandidate-based RNA interference (RNAi) screen led to theiscovery of Drosophila Stim. In further work, we identifiedwo functions of Stim and the mammalian homologue STIM1collectively referred to here as Stim1): the sensor function
hat initially detects the reduction of Ca2+ content in theumen of the endoplasmic reticulum (ER); and the messengerunction provided by Stim1 translocation to the plasma mem-∗ Corresponding author. Tel.: +1 949 824 7776; fax: +1 949 824 3143.E-mail address: [email protected] (M.D. Cahalan).
1 Present address: CalciMedica Inc., La Jolla CA 92037, United States.
Chantor
143-4160/$ – see front matter © 2007 Published by Elsevier Ltd.oi:10.1016/j.ceca.2007.03.002
rane to activate the CRAC channel. We further discuss ourenome-wide RNAi screen that identified Drosophila Oraind additional genes that are required for CRAC channelctivity; the recipe for greatly amplified CRAC current byo-expression of Stim + Orai; and the identification of aminocids that regulate Ca2+ selectivity, clinching the identifica-ion of Orai as the pore-forming component of the CRAChannel.
.1. Lymphocytes and the early activation pathway of anmmune response
T lymphocytes require direct contact with antigen-resenting cells (APC) such as dendritic cells to becomectivated. In parallel with work on the CRAC channel, theahalan lab group, in collaboration with Ian Parker at UCI,as used two-photon microscopy to image living T cellsnd dendritic cells interacting within intact mouse lymph
odes [1,2]. Under basal conditions without antigen, con-acts between dendritic cells and T cells are initiated by virtuef robust T cell motility in combination with extension andetraction of dendritic cell dendrites, leading to a very high
1 ll Calcium 42 (2007) 133–144
fDtictclioa
1
r(eatptodtidptaTsabrowbaaiststuiiaaf
etTf
m[pttober[acmtrC[pApimimaiai
1a
34 M.D. Cahalan et al. / Ce
requency of transient T cell/dendritic cell interactions [3].endritic cells capture antigens in the periphery and bring
hem into the lymph node, serving as APC to initiate themmune response. When antigen is present, T cell–dendriticell contact durations are prolonged as Ca2+ signaling is ini-iated; and after several hours, T cells are stably associated inlusters with individual dendritic cells [4]. Finally, the T cellset go and begin to divide about 24 h after initially encounter-ng the antigen. Our story about the CRAC channels focusesn the T cell Ca2+ signal that is initiated by contact with APCnd is crucial for initiation of an adaptive immune response.
.2. Ion channels in human T cells
Ca2+ influx across the plasma membrane is primarilyesponsible for elevation of cytosolic free Ca2+ concentration[Ca2+]i) following TCR engagement. But how does calciumnter the cell? Even earlier, nearly 25 years ago and shortlyfter the patch-clamp method was developed, researchers inhe Cahalan lab initiated the study of ion channels in lym-hocytes [5]. At that time, the importance of Ca2+ signalingo lymphocyte activation had already been proposed [6], andur initial expectation was that calcium channels might beetected electrically. At first, we investigated K+ channelshat regulate the membrane potential of T cells (and therebyndirectly modulate Ca2+ signaling), but no Ca2+ current wasetected. By combining single-cell Ca2+ monitoring withatch recording, Richard Lewis in the Cahalan lab found ainy Ca2+-selective current with an inwardly rectifying I–Vnd a very positive reversal potential in Jurkat T cells [7].he Ca2+ current, only a few picoamperes per cell, activatedpontaneously during whole-cell recording with dialysis ofCa2+ chelator. The same current could also be activated
y T cell receptor stimulation during perforated-patchecording in a close temporal relationship with the upstrokef the cytosolic Ca2+ signal monitored simultaneouslyith a fluorescent Ca2+-indicator. The current could belocked by external application of Ni2+ or Cd2+, agents thatlso blocked the mitogen-evoked Ca2+ signaling. Duringctivation of the current, single-channel currents and evenncreased current noise were not evident, indicating a lowingle-channel conductance. At the time, a possible link ofhe Ca2+ current across the surface membrane to the internaltores was discussed, but the relation to store depletion ando models for “capacitative calcium entry” (CCE) [8] wasnclear. The current was further studied by other groups,ncluding Richard Lewis at Stanford and Reinhold Pennern Gottingen, and was shown to be activated by thapsigarginnd by intracellular IP3 [9–11], strengthening its identity asstore-operated Ca2+ channel. It was given the name CRAC,
or Ca2+ release-activated Ca2+ current, by Penner.Fig. 1 highlights the function of ion channels during an
ncounter between a T cell and an APC that has processedhe antigens specific for the T cell receptor expressed on the
cell. Very few channels are open initially; in fact only aew of the voltage-gated Kv1.3 channels in the cell suffice to
ssml
Fig. 1. Ion channels and signaling pathways in T cells.
aintain the resting membrane potential of about −50 mV12]. Contact with peptide-MHC on the APC initiates theroximal signaling cascade involving adapter molecules andyrosine kinases, resulting in phospholipase C activation andhe formation of the two messengers, IP3 and DAG. IP3pens the IP3 receptor-channels localized in the ER mem-rane, which in turn releases Ca2+ from the ER and begins tolevate cytosolic Ca2+. Depletion of the ER Ca2+ stores indi-ectly leads to Ca2+ influx through CRAC channels, drivingCa2+]i into the micromolar range. As [Ca2+]i rises, Ca2+-ctivated K+ channels are opened and K+ leaves the cell,ausing the membrane potential to become more negative toaintain the driving force for Ca2+ influx. The relative con-
ribution of voltage-gated and Ca2+-activated K+ channels toegulate membrane potential and counterbalance the influx ofa2+ depends upon the T cell subset and their activation state
13]. Ca2+ binds to calmodulin and activates calcineurin, ahosphatase that dephosphorylates a transcription factor NF-T, which then translocates to the nucleus where it binds toromoter regions to turn on expression of numerous genes,ncluding the key cytokine, Interleukin-2 [14,15]. The phar-acological agent thapsigargin bypasses the upstream events
n T cell signaling by inhibiting the SERCA pump in the ERembrane, which leads to depletion of the ER Ca2+ store and
ctivation of CRAC channels that trigger subsequent eventsncluding gene expression. Thapsigargin was thus employeds a very useful tool in our RNAi screening effort, describedn Sections 2 and 3 below.
.3. Ca2+ signaling and gene expression during T cellctivation
By the mid-1990s, the calcium dependence of gene expres-
ion for T cell activation was determined quantitatively iningle-cell Ca2+ assays combined with NFAT reporter geneonitoring [15,16]. To activate this key transcriptional regu-atory pathway, at least 1–2 h of elevated [Ca2+]i is required.

ll Calc
TtaociorCcgbCd
1
ripndboate
2Sc
2b
KIievtnctaTacflRs
TC
R
M.D. Cahalan et al. / Ce
he Ca2+ signal may be oscillatory or sustained, and it ishought that oscillations in [Ca2+]i may provide a means toctivate transcription of different genes selectively and with-ut the requirement for continuous Ca2+ elevation, whichould activate cell death pathways [17,18]. The content of thenternal Ca2+ store is limited in T cells, and Ca2+ influx fromutside of the cell is required to elevate Ca2+ to a level that isequired for specific gene expression during T cell activation.RAC channel activity and downstream Ca2+ effects via thealcineurin/NFAT pathway are thus critical for changes inene expression elicited by TCR stimulation, as evidencedy altered gene expression in T cells that lack functionalRAC channels from patients with severe combined immuneeficiency (SCID) disorder [14].
.4. CRAC channels at the start of the millennium
As of 2003, then, the biophysical properties of CRAC cur-ent had been characterized, as summarized in Table 1. Themmunological importance of the CRAC current to the NFATathway and to T cell activation was also clear. However,either the channel itself nor the mechanism that links storeepletion to CRAC channel opening were known. The majorreakthroughs finally came in 2005 and 2006 when we and
ther groups used RNAi to identify genes that were requirednd site-directed mutagenesis to define the functional role ofwo proteins, Stim1 and Orai1, in turning on and physicallymbodying the CRAC channel, respectively.n
ia
able 1omparison of CRAC current biophysical properties
Property Native T cell CRAC[7,9–11,53–55]
Native S2 CRAC [2
Store dependence Yes Yes
Inactivation by:Store refilling Yes YesLocal [Ca2+]i Yes YesRun-down Yes Yes
Fast inactivation (100 mstime scale)
Yes No
Calcium selectivity Yes YesPermeant divalent cations Ca2+ > Ba2+ > Sr2+» Mg2+ Ba2+ > Ca2+ > Sr2+
Unitary channelconductance in 20 mMCa2+
21 fS 36 fS
Monovalent current inzero divalent
Yes Yes
Monovalent cationselectivity, PCs/PNa
Na+ > Cs+, 0.09–0.13 Na+ > Cs+, 0.17
Depotentiation Yes Yes
Pharmacology:Gd3+ or La3+ nM (Gd3+), reversible nM (Gd3+), reversiSKF 96365 �M, partially reversible �M, partially rever2-APB Low �M – potentiation;
high �M – inhibitionLow �M – potentiahigh �M – inhibitio
esults are summarized from the indicated references.
ium 42 (2007) 133–144 135
. RNAi candidate screen leads to identification oftim, the Ca2+ sensor and messenger for CRAChannel activation
.1. Choice of S2 cells for RNAi that led to the screeningreakthrough
In the late summer of 2000, three of us (JR, KO andS) joined a new company (now TorreyPines Therapeutics
nc.) and as part of an early project sought a novel way todentify the molecular components of store-operated Ca2+
ntry (SOCE), especially the elusive CRAC channel. Pre-ious work had suggested certain TRP channels might behe molecular counterparts of both store-operated Ca2+ chan-els and CRAC channels, but biophysical properties of TRPhannels and CRAC channels did not match, and it seemedhe field was nibbling around the edges because it lacked
more systematic and direct way to attack the problem.hen a colleague, Craig Montell, brought to our attentionnewly developed technique for suppressing gene function
alled RNAi, which had shown utility in plants, worms, andies. In these cases, introduction to cells of a double-strandedNA (dsRNA) mimicking a specific mRNA sequence causes
equence-specific degradation of the corresponding endoge-
ous mRNA, leading to the suppression of gene expression.The Drosophila S2 cell line attracted our attention becausets utility for studying Ca2+ signaling mechanisms waslready known [19–24]. S2 cells were an attractive target for
6,33] Stim + Orai [33,48] Stim + E180D Orai [48]
Yes Yes
Not tested Not testedYes YesYes Yes
No No
Yes No» Mg2+ Ca2+ ∼ Sr2+
∼Ba2+ » Mg2+Negligible
Not tested N/A
Yes Yes
Na+ > Cs+, 0.13 Na+ ∼ Cs+, 0.71
Yes No
ble nM (Gd3+), reversible nM (Gd3+), reversiblesible Not tested Not testedtion;n
Low �M – potentiation;high �M – inhibition
Low and high �M – complexbehavior (potentiation andinhibition phases)

1 ll Calc
ptcgiisdCc(oastot>ilaNimb2adccScacioer
2
fuibtatt[nNdtp
iRmpbiaScs
2
wagwssDniPtpwvg(i∼gpStSttiocsatgh
2
c
36 M.D. Cahalan et al. / Ce
erforming an RNAi-based screen to identify genes that con-rols SOCE because most fundamental cellular processes areonserved from Drosophila to mammals and the Drosophilaenome had just been published earlier in 2000 [25]. Of greatmportance in the interpretation of such a screen, the TorreyP-nes group teamed-up with the Cahalan group at UC Irvine tohow that S2 cells possess a store-operated Ca2+ current. Aetailed patch-clamp analysis showed that the store-operateda2+ current in S2 cells was remarkably similar in biophysicalharacteristics to the CRAC current in human lymphocytesTable 1) [26], so the choice of S2 cells turned out to be a goodne. Both currents are highly Ca2+ selective (PCa/PNa > 1000)nd can be activated by maneuvers that deplete the ER Ca2+
tore: cell dialysis with a Ca2+ chelator (passive store deple-ion); internal application of IP3; or external applicationf thapsigargin or ionomycin. Both exhibit the characteris-ic inwardly rectifying I–V shape with a reversal potential50 mV and an unusually low single-channel conductance
n the femtosiemen range. In the absence of external diva-ent cations, both currents are carried by monovalent cationsnd sodium is more permeant than cesium. The monovalenta+ current declines (depotentiation) over tens of seconds
n the continued absence of external divalents. The phar-acological properties are also quite similar: currents can
e suppressed by lanthanides (La3+, Gd3+), SKF96365 or-APB (high �M range); while the application of 2-APBt lower concentration potentiates both currents. A moreetailed comparison of the biophysical properties of CRACurrent in S2 and T cells is presented in Table 1, along withharacteristics of CRAC current induced by expression oftim + Orai. These initial findings validated the use of S2ells for identifying SOC/CRAC channel components for usnd other investigators. That S2 cells possess a CRAC-likehannel may not be surprising, as the S2 cell line was orig-nally derived from a primary culture of late stage (20–24 hld) Drosophila melanogaster embryos and, based upon genexpression patterns and phagocytic activity, is thought toepresent a haemocyte or macrophage-like cell [27,28].
.2. How RNAi works in S2 cells
The Drosophila S2 cell culture system is ideally suitedor moderate- to high-throughput assays of gene functionsing RNAi-mediated gene silencing. This is because RNAin S2 cells is simple, efficient, and long-lived. S2 cells cane treated with long, ∼500 base-pair dsRNAs, which areaken up easily from the medium by the cells [29]. Andvantage of S2 cells is that they do not require transfec-ion reagents for uptake of dsRNAs. The dsRNAs appearo enter the cells through receptor-mediated endocytosis30]. Once inside the cells, dsRNAs are processed to 21–23ucleotide fragments called small interfering RNAs (siR-
As) by the action of the enzyme Dicer. The use of longsRNA probes increases the likelihood of producing siRNAshat efficiently bind to the RNA-inducing silencing com-lex (RISC), thereby reducing optimization steps necessarySfltt
ium 42 (2007) 133–144
n designing siRNA probes for RNAi. Once bound to theISC complex, siRNAs initiate degradation of the comple-entary mRNA sequences and subsequent suppression of
rotein translation. These considerations enhance the feasi-ility of conducting a high-throughput RNAi-based screenn S2 cells. Since the Drosophila genome is much smallernd was better examined at the time than the human genome,2 cells provided the ideal model system to test the role ofandidate genes in CRAC channel function by systematicallyuppressing functional expression through RNAi.
.3. Choice of candidates by bioinformatics
At the time we initiated our first RNAi screen, genome-ide dsRNA libraries were not available, so we chosebioinformatics approach to identify potential candidate
enes. At the beginning, we tested about 20 genes thatere either known TRP family members or TRP-related
equences. When suppression of these genes failed to affecttore-operated Ca2+ entry in S2 cells, we searched therosophila genome for all genes annotated as ion chan-els in the GadFly database (release 2). In addition, wedentified genes having an InterPro channel-like motif (e.g.,ROSITE, PRINTS, PRODOM) and genes that shared a Mul-
iple Expectation maximum for Motif Elicitation (MEME)rofile built from members of the TRP family. From this list,e excluded genes we had already tested, well-characterizedoltage-regulated channels, non-calcium channels, and otherenes that had functional data published in the literaturee.g., cacophony, eag, Glu-RIIA, Glu-RIIB, para, Sh) retain-ng ∼80% of the candidates from the Gadfly-derived list and
90% of the motif-derived list. Finally, we selected otherenes containing Ca2+ binding motifs, genes annotated aslasma membrane ATPases, or genes with putative roles inOC influx, arriving at a final list of 150 genes that were then
argeted by RNAi individually and tested. As it turned out,tim was included in the list not only because of its puta-ive Ca2+ binding domain, but also because a transient bug inhe genome annotation identified Stim as having a potassiumon-channel motif. This highlights the fact that at the time ofur initial screen the annotation of the fly genome was stillhanging. In addition, novel classes of genes with undefinedequence motifs and architectures could not be discovered bybioinformatics approach. Thus, another key component of
he CRAC channel was only discovered later using unbiasedenome-wide RNAi screens (i.e., olf186-F, the Drosophilaomolog of human Orai1 or CRACM1) [31–33].
.4. How the initial candidate screen worked
The initial screen (Fig. 2) utilized a FluoroSkan single-hannel fluorimeter to measure baseline and SOCE signals.
2 cells were treated with the appropriate dsRNAs in T-75asks for 5 days, harvested and plated in 96-well plates, andhen loaded with the Ca2+ sensitive dye fluo-4 in buffer con-aining calcium. Cells were washed in calcium-free buffer and

M.D. Cahalan et al. / Cell Calc
flIbtvCioanftroSaui
2i
sS[aFCtpvCif
npttDbiiipoTtrt
2m
difeStaifsoasnwS
rtrbB(aSorp(WaS
Fig. 2. Candidate RNAi screening protocol.
uorescence levels were then monitored by the FluoroSkan.nitial fluorescence was recorded for a brief period to estimateasal Ca2+ levels, followed by addition of either vehicle orhapsigargin to discharge intracellular Ca2+ stores and acti-ate store-operated Ca2+ channels. After a short incubation,aCl2 was added to the cells and a single fluorescence read-
ng was recorded three min after Ca2+ addition as an indexf store-operated Ca2+ influx in each well. This single-pointssay was possible because in our hands the Ca2+ entry sig-al became stable 2–4 min after Ca2+ addition, and it greatlyacilitated the throughput of the RNAi screen. It was withhis assay that we identified the activity of Stim [34] (firstevealed in a patent application published in 2004 with pri-rity to 2003 [35], and presented publicly at the Biophysicalociety meeting in February 2005), a finding consistent withsubsequent, independent report by Liou et al. [36]. We latersed this basic paradigm in a genome-wide RNAi screen todentify the role of olf186-F (Orai) [33].
.5. Validation of Stim and STIM1 by single-cell Ca2+
maging and patch-clamp
Our initial study validated the requirement for Stim byingle-cell Ca2+ monitoring and by whole-cell recording in2 cells pretreated with dsRNAs for Stim or control genes34]. In Ca2+ imaging experiments, we used a thapsigarginnd Ca2+ readdition protocol to assess SOCE in single cells.ollowing addition of thapsigargin in the absence of externala2+, a small transient rise in cytosolic free Ca2+ represents
he release transient due to leak from the store into the cyto-lasm. Subsequent readdition of external Ca2+ produces a
ery large Ca2+ signal that is due to Ca2+ influx throughRAC channels that are activated as a result of thagpsigargin-nduced Ca2+ store depletion. In cells pretreated with dsRNAsor Stim, resting Ca2+ was unaltered and store content was
msno
ium 42 (2007) 133–144 137
ormal, but Ca2+ influx upon readdition of external Ca2+ wasrofoundly inhibited. Patch-clamp experiments confirmedhat knockdown of Stim by RNAi very effectively suppressedhe CRAC current. There are two mammalian homologs ofrosophila Stim, STIM1 and STIM2. Suppression of STIM1y treatment of HEK293 cells with STIM1-specific siRNAsnhibited thapsigargin-evoked Ca2+ entry and Ca2+ influxnduced by muscarinic receptor stimulation, without alter-ng resting Ca2+ levels, Ca2+ release transients, or membraneotentials [34]. In our hands, suppression of STIM2 had lessf an effect on SOCE than did STIM1 knockdown. In Jurkatcells, we again confirmed that suppression of STIM1 led
o a profound reduction of the Ca2+ signal and CRAC cur-ent, this time using a short hairpin loop (shRNA) constructo reduce STIM1 expression selectively.
.6. Dual functional role of Stim1: ER Ca2+ sensor andessenger to the plasma membrane
In the experiments described above, RNAi knockdownemonstrated a functional requirement for Stim1 expressionn supporting CRAC channel activity. But how does Stim1unction, as a single span transmembrane protein that does notven remotely resemble any known ion channel? Drosophilatim and its mammalian homologs, STIM1 and STIM2, are
ype 1 membrane proteins. The signal peptide sequence at themino terminus and one predicted transmembrane segmentndicated that the amino (N-) terminal portion of the proteinaces the extracellular fluid or the lumen of the ER. The con-erved N-terminal EF-hand motif suggested a possible rolef Stim1 in Ca2+ signaling, and since Stim1 did not resembleny known ion channel we formulated the idea that it mayerve as a Ca2+ sensor to trigger the process of CRAC chan-el activation. The approach we took was to overexpress theild-type (WT) Stim1 protein or Stim1 mutant proteins in2 and Jurkat T cells [37].
Our strategy for a series of point mutations was to dis-upt Ca2+ binding to the EF-hand domain while preservinghe overall protein structure, targeting different conservedesidues that had been shown in other EF-hand proteins andy crystal structures to be essential for Ca2+ binding (Fig. 3).y making several different mutations on the same sites
D to N or D to A, and E to Q or E to A; Fig. 3 ovals)nd corresponding mutations in different species (Stim andTIM1), we tried to avoid false-positive results. The mutantsf Stim or STIM1 were expressed in S2 or Jurkat T cells,espectively, by transfection, and calcium signaling was com-ared in native cells and cells overexpressing either wild-typeWT) or mutant Stim/STIM1. Expression was confirmed by
estern blot which showed similar expression levels of WTnd mutants proteins. The S2 cells overexpressing the WTtim had no phenotype, whereas expression of the EF-hand
utants produced an elevation of resting [Ca2+]i that wehowed was due to constitutive activation of CRAC chan-els, since it was reduced to control levels by three inhibitorsf CRAC current: 2-aminoethyldiphenyl borate (2-APB),

138 M.D. Cahalan et al. / Cell Calcium 42 (2007) 133–144
equenc
SmohccstrnfitdChlm
tiSWWpbptpia
fmodlbSismCatrwctt
tasrat[
Fig. 3. Stim1 s
KF96365, and Gd3+. In Jurkat T cells, a modest enhance-ent of thapsigargin-evoked Ca2+ influx was produced by
verexpressing the WT protein, and expression of the EF-and mutant elevated cytosolic Ca2+ levels, as seen with S2ells. This rise in Ca2+ was also completely blocked by CRAChannel inhibitors. Importantly, there was no change in ERtore content or release kinetics, leading to the conclusionhat expression of the EF-hand mutants bypassed the normalequirement for store depletion and activated CRAC chan-els even though the store was still replete with Ca2+. Thesendings suggested that Stim1 serves as an ER Ca2+ sensor
hat can trigger subsequent events when the Ca2+ store isepleted. According to this idea, Stim1 changes its state toa2+-unbound upon store depletion or by crippling the EF-and and is the effector that initiates a sequence of eventseading to activation of the CRAC channel in the plasma
embrane.In parallel with these functional studies, we determined
he distribution of the overexpressed STIM1 proteins bymmunofluorescence using polyclonal antibodies specific toTIM1 and found a very different pattern of expression forT and EF-hand mutant STIM1 [37]. The overexpressedT STIM1 proteins were primarily localized in an ER-like
attern with some expression also visible at the plasma mem-rane, whereas the EF-hand mutant STIM1 proteins wereredominantly seen at the plasma membrane in aggregations
hat we termed hotspots. Importantly, similar hotspots, oruncta, of native STIM1 were observed in several cell types,ncluding Jurkat T cells, primary human T cells, RBL cells,nd PC12 cells, using the same polyclonal antibodies shortlysSor
es and motifs.
ollowing store depletion. We used SERCA as a bona fidearker of the ER for comparison to the native STIM1 pattern
f subcellular localization which changed following storeepletion. In normal Ringer solution, there was extensive co-ocalization of both proteins. But when the store was depletedy thapsigargin, co-localization was reduced, because theTIM1 protein had migrated and was predominantly seen
n hotspots on the cell surface. In addition, a time-coursetudy showed that this redistribution of STIM1 to the plasmaembrane was rapid, overlapping with the time-course ofRAC channel activation. After presenting these data at thennual Biophysical Society meeting (Feb. 2005), we decidedo explore the subcellular localization of STIM1 with higheresolution by electron microscopy. The polyclonal antibodiesere conjugated to quantum dots that are both highly fluores-
ent and visible as particles by electron microscopy. By thisechnique, native STIM1 in T cells was shown to redistributeo the plasma membrane following Ca2+ store depletion.
In summary, our results indicate that Stim1 has two func-ions: using the EF-hand to sense luminal ER Ca2+ content;nd migrating to the plasma membrane when the EF-handenses lowered Ca2+ in the ER. A similar conclusion waseached using an EF-hand YFP-tagged STIM1 mutant toctivate SOCE and YFP-tagged STIM1 to track the migra-ion of STIM1 to the cell surface following store depletion36]. The Ca2+ sensor hypothesis was strengthened by a sub-
equent in vitro biochemical study using a core N-terminusTIM1 construct [38]. An apparent dissociation constant (Kd)f 200–600 �M was measured, consistent with the functionalange of Ca2+ levels within the ER upon store depletion.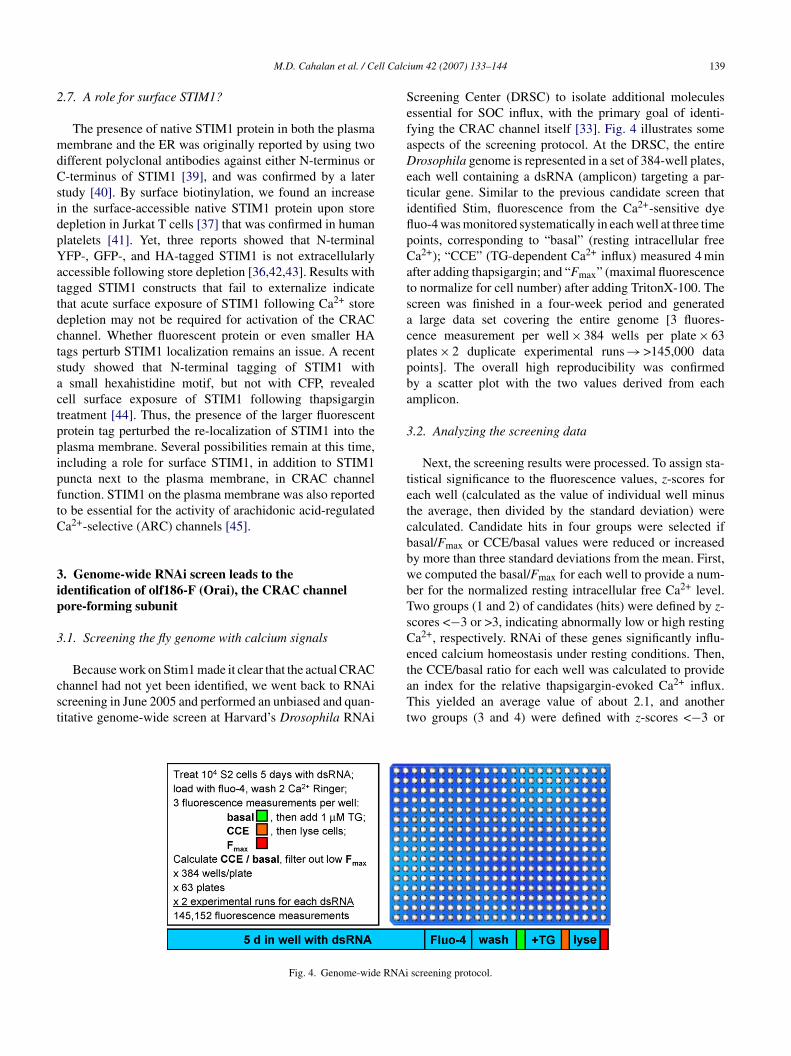
ll Calc
2
mdCsidpYattdctsactppipftC
3ip
3
cst
SefaDetiflpCatsacppba
3
tetcbbwbTsCe
M.D. Cahalan et al. / Ce
.7. A role for surface STIM1?
The presence of native STIM1 protein in both the plasmaembrane and the ER was originally reported by using two
ifferent polyclonal antibodies against either N-terminus or-terminus of STIM1 [39], and was confirmed by a later
tudy [40]. By surface biotinylation, we found an increasen the surface-accessible native STIM1 protein upon storeepletion in Jurkat T cells [37] that was confirmed in humanlatelets [41]. Yet, three reports showed that N-terminalFP-, GFP-, and HA-tagged STIM1 is not extracellularly
ccessible following store depletion [36,42,43]. Results withagged STIM1 constructs that fail to externalize indicatehat acute surface exposure of STIM1 following Ca2+ storeepletion may not be required for activation of the CRAChannel. Whether fluorescent protein or even smaller HAags perturb STIM1 localization remains an issue. A recenttudy showed that N-terminal tagging of STIM1 with
small hexahistidine motif, but not with CFP, revealedell surface exposure of STIM1 following thapsigarginreatment [44]. Thus, the presence of the larger fluorescentrotein tag perturbed the re-localization of STIM1 into thelasma membrane. Several possibilities remain at this time,ncluding a role for surface STIM1, in addition to STIM1uncta next to the plasma membrane, in CRAC channelunction. STIM1 on the plasma membrane was also reportedo be essential for the activity of arachidonic acid-regulateda2+-selective (ARC) channels [45].
. Genome-wide RNAi screen leads to thedentification of olf186-F (Orai), the CRAC channelore-forming subunit
.1. Screening the fly genome with calcium signals
Because work on Stim1 made it clear that the actual CRAChannel had not yet been identified, we went back to RNAicreening in June 2005 and performed an unbiased and quan-itative genome-wide screen at Harvard’s Drosophila RNAi
taTt
Fig. 4. Genome-wide RNAi
ium 42 (2007) 133–144 139
creening Center (DRSC) to isolate additional moleculesssential for SOC influx, with the primary goal of identi-ying the CRAC channel itself [33]. Fig. 4 illustrates somespects of the screening protocol. At the DRSC, the entirerosophila genome is represented in a set of 384-well plates,
ach well containing a dsRNA (amplicon) targeting a par-icular gene. Similar to the previous candidate screen thatdentified Stim, fluorescence from the Ca2+-sensitive dyeuo-4 was monitored systematically in each well at three timeoints, corresponding to “basal” (resting intracellular freea2+); “CCE” (TG-dependent Ca2+ influx) measured 4 minfter adding thapsigargin; and “Fmax” (maximal fluorescenceo normalize for cell number) after adding TritonX-100. Thecreen was finished in a four-week period and generated
large data set covering the entire genome [3 fluores-ence measurement per well × 384 wells per plate × 63lates × 2 duplicate experimental runs → >145,000 dataoints]. The overall high reproducibility was confirmedy a scatter plot with the two values derived from eachmplicon.
.2. Analyzing the screening data
Next, the screening results were processed. To assign sta-istical significance to the fluorescence values, z-scores forach well (calculated as the value of individual well minushe average, then divided by the standard deviation) werealculated. Candidate hits in four groups were selected ifasal/Fmax or CCE/basal values were reduced or increasedy more than three standard deviations from the mean. First,e computed the basal/Fmax for each well to provide a num-er for the normalized resting intracellular free Ca2+ level.wo groups (1 and 2) of candidates (hits) were defined by z-cores <−3 or >3, indicating abnormally low or high restinga2+, respectively. RNAi of these genes significantly influ-nced calcium homeostasis under resting conditions. Then,
he CCE/basal ratio for each well was calculated to providen index for the relative thapsigargin-evoked Ca2+ influx.his yielded an average value of about 2.1, and anotherwo groups (3 and 4) were defined with z-scores <−3 or
screening protocol.

1 ll Calc
>etef3coo(werCtatt
So
3w
o1kimgh
40 M.D. Cahalan et al. / Ce
3, indicating abnormally low or high store-operated Ca2+
ntry, respectively. Assuming that RNAi of genes (e.g. Stim)hat control SOCE do not influence the resting Ca2+ lev-ls, overlapping candidates in groups 1 and 4 were removedrom group 4, and overlapping candidates in groups 2 and
were removed from group 3 to avoid false-positive out-omes resulting from altered resting Ca2+ levels. The valuef Fmax was used to filter out wells in which cells did not growr failed to adhere, using a z-score of Fmax <−2 as a cutoffconfirmed by visual observation). In addition, we filtered outells having more than five off-targets (non-specific RNAi
ffect). We then focused on filtered hits in group 3 with 75emaining candidates, which should include the long-soughtRAC channel pore gene. The list was further narrowed down
o those candidates with predicted transmembrane segmentsnd with mammalian homologs; both are essential condi-ions for the CRAC pore subunit. In the end, a final list of 11ransmembrane protein hits was generated [33]. Gratifyingly,
apwm
Fig. 5. Fly and human Orai-relat
ium 42 (2007) 133–144
tim was one of the strongest hits with a CCE/basal valuef 1.26.
.3. Olf186-F (Orai) as a conserved membrane proteinith three human homologs
Uniquely among the filtered transmembrane protein hits,lf186-F (now renamed Orai), with a CCE/basal value of.31 and four predicted transmembrane segments, had nonown function. Although only one member is representedn fly (dOrai, Fig. 5) and worm genomes, a conserved three-
ember gene family is present in all sequenced mammalianenomes. These are now renamed Orai1, Orai2, and Orai3 onuman chromosomes 12, 7, and 16, respectively. Sequences
nd other key features are shown in Fig. 5. There are fourredicted transmembrane segments in each Orai homologhich are all highly conserved, especially the first two trans-embrane segments and the loop region in between. Theseed sequences and motifs.

ll Calc
pb
3p
ldFtdwswwwktdtmsvCSc
ttweda1tcsg1top
3c
rcTcoasS
gaCatcf
3s
datiOrtfamfonttntoowtSdnrfctsrsloOcobiTi
M.D. Cahalan et al. / Ce
rovided hints for our follow-up mutagenesis strategy (seeelow).
.4. Validation of hits by single-cell Ca2+ imaging andatch-clamp
Several final candidates were validated at the single-cellevel by both Ca2+ imaging and patch-clamp analysis forirect RNAi effects on CRAC activity [33]. As for olf186-(Orai), the vast majority of cells pretreated with dsRNA
argeting olf186-F did not exhibit Ca2+ influx upon storeepletion and had greatly reduced CRAC current densities,hereas both resting Ca2+ level and store content were the
ame as in control cells. We also validated the SERCA pump,hich could have been a false-positive hit since the screenas based on thapsigargin. In single-cell Ca2+ assays andhole-cell recording from cells pretreated with dsRNA tonock down SERCA, there was a highly significant reduc-ion in thapsigargin-evoked Ca2+ entry and in CRAC currentensities recorded during passive store depletion. It remainso be determined whether this reduction resulted from abnor-
ally low Ca2+ store content (assayed by ionomycin-evokedtore release) or elevated [Ca2+]i levels. Syntaxin 5 was alsoalidated as a genuine hit. Thus, by RNAi we confirmed witha2+ imaging and patch-clamp recording that Stim, Orai,ERCA, and Syntaxin 5 are each required for normal CRAChannel function.
An RNAi screen based on NF-AT translocation as a func-ional measurement was performed in DRSC at the sameime [31]. Combined with genetic mapping of SCID patientshich was initiated in 1996, Orai was also identified as an
ssential component for store-operated Ca2+ influx and theownstream NF-AT relocation into the nucleus. Moreover,recessive point mutation in human Orai1 on chromosome2 was isolated from SCID patients [31]. This channelopa-hy further reinforces the functional significance of CRAChannels (and Orai1) for the immune system. A third RNAicreen initiated several months later also identified the sameene (which was renamed as CRACM1 for CRAC modulator), together with another candidate (dpr3) called CRACM2 inhe fly genome [32]. Remarkably, all three screens identifiedlf186-F (Orai, CRACM1) as an essential transmembranerotein.
.5. Overexpression of Orai and Stim: amplified CRACurrent
RNAi can test whether a particular molecule may beequired for normal function. But in a complex multi-omponent system, several required elements may be present.hus, we sought to investigate effects of overexpression. Weloned the cDNA of Orai from S2 cells and expressed it with
r without co-transfected Stim. When Orai was expressedlone, CRAC current density increased three-fold, a highlyignificant increase and in contrast to the lack of effect whentim was overexpressed by itself. Importantly, when bothsiim
ium 42 (2007) 133–144 141
enes were transfected, a huge CRAC current was recorded,nd the kinetics of activation were accelerated compared toRAC current induced by overexpression of Orai alone. Themplified CRAC current exhibited all the biophysical charac-eristics of native CRAC current in S2 cells (Table 1). Theseoexpression results provided further evidence that Orai mayorm the CRAC channel pore [33].
.6. Mutations in Orai that affect CRAC channel Ca2+
electivity
The ability of ion channels to select among various ionsepends on interactions between amino acid side chains, ionsnd water molecules. Therefore, we sought to identify par-icular amino acids that, when mutated, resulted in alteredon selectivity of CRAC current following expression ofrai + Stim. Ion selectivity is usually accomplished by loop
egions between transmembrane segments. It has been shownhat negatively charged amino acids are the key elementsor the Ca2+-selectivity filter of voltage-gated Ca2+ channelsnd TRPV6 [46,47]. The loop between transmembrane seg-ents 1 and 2 of the fly Orai gene and mammalian Orai
amily members is the most conserved (Fig. 5). We focusedn negatively charged residues and prepared a series of ala-ine substitutions. Mutation of the conservative glutamateo alanine at position 180 of Orai resulted in inhibition ofhe background S2 CRAC current, indicating a dominant-egative action. A conservative point mutation at this positiono aspartate dramatically altered the ion selectivity propertiesf CRAC current. Whereas a large inward current devel-ped during passive store depletion when WT Orai + Stimere co-transfected, an outward current developed when
he point mutant E180D Orai was expressed together withtim. This was because the CRAC channel I–V characteristicisplayed pronounced outward rectification, rather than theormal inward rectification of native or amplified CRAC cur-ent. Ion substitution experiments clearly revealed a changerom Ca2+ selective permeation to monovalent cation currentsarried by Na+ or Cs+. Furthermore, several other proper-ies related to ion selectivity were affected. In divalent-freeolution, the relative Na+ to Cs+ permeability, PNa/PCs, waseduced. The increase in Cs+ permeability is consistent with alightly enlarged selectivity filter. Furthermore, the monova-ent currents failed to depotentiate. The dramatic alterationsf ion selectivity by a point mutation at position 180 ofrai indicate that Orai is the pore-forming subunit of the
hannel, responsible for Ca2+ selectivity. Interestingly, threether mutants near 180 had reduced sensitivity to the channellocker Gd3+ possibly by reduced charge screening result-ng in a reduced local concentration of the trivalent blocker.aken together, these results lead to the conclusion that Orai
s a bona fide CRAC channel pore subunit [48]. A corre-
ponding point mutation in Orai1 at position 106 resultsn similar changes [49,50], although with some differencesn rectification between the corresponding Orai and Orai1utants.

1 ll Calc
3
etOm[tbip(offaaifraamatandda
4
sviEaiOtig2s
uaTsvws
tndOghst
R
[
[
[
[
[
[
[
42 M.D. Cahalan et al. / Ce
.7. Interaction with Stim
So far, the CRAC channel gating mechanism has not beenlucidated. The molecular events between Stim1 transloca-ion and CRAC activation remain undefined. Both STIM1 andrai1 accumulate at junctional sites where the ER and plasmaembrane are closely apposed following store depletion
43,51]. In S2 cells, we found a greatly enhanced interac-ion between Orai and Stim upon store depletion, assessedy co-immunoprecipitation (co-IP), reflecting a dynamicnteraction model for CRAC channel activation [48]. Oneossibility is that Stim delivers information about the storeempty) to Orai by direct physical contact, which in turnpens the channels. However, amino acid sequences requiredor the interaction and a possible requirement for other scaf-olding proteins need to be further tested. As Drosophila Stimnd Orai are both required to reconstitute CRAC channelctivity in HEK cells (unpublished data), it will be interest-ng to see whether these two proteins alone form a minimalunctional unit of CRAC channels. On the other hand, theedistribution of both STIM1 and Orai1 hint that there may befinely tuned recruitment mechanism to generate a polarizedctivated CRAC channel distribution for Ca2+ signaling inicrodomains. An interaction between mammalian STIM1
nd Orai1 was also reported via co-IP assay without TG-reatment [50]. The interaction may be indirect or of lowffinity as another co-IP based study showed that Orai1 didot associate with STIM1 in several detergent and salt con-itions tested [52]. Further work is needed to reconcile theseata and to determine how store depletion affects the inter-ction.
. Overview and prospects
RNAi screening [31–34,36] and subsequent functionaltudies have clarified the molecular requirements and acti-ation mechanisms of the CRAC channel, a sort of holy grailn the field of Ca2+ signaling. In human T cells, STIM1 sensesR Ca2+ content using an EF-hand motif near the N-terminusnd translocates to the plasma membrane when the Ca2+ stores depleted [36,37]. The Ca2+ channel itself is formed fromrai1 in T cells, since a naturally occurring mutation within
he first transmembrane segment abrogates channel activityn SCID patients [31], and a point mutant of a conservedlutamate residue between transmembrane segments 1 andin Drosophila Orai or human Orai1 alters the Ca2+ ion
electivity of the channel [48–50].The next challenge is to understand the detailed molec-
lar composition and organization of CRAC channels, asunique Ca2+ channel unrelated to other channel families.he subunit organization needs to be defined, and further
ite-directed mutagenesis with functional assays will pro-ide more information. Ultimately a crystal or NMR structureill be essential to complete this mission. High-throughputcreening may provide useful CRAC-specific blockers. Fur-
[
ium 42 (2007) 133–144
hermore, the functional role of Stim and Orai homologs willeed to be defined. The three human Orai homologs exhibitifferent tissue expression patterns. The fact that normalrai2 and Orai3 genes cannot compensate the mutated Orai1ene in SCID patients indicates that the three members mayave different physiological roles [52]. Genetic targeting isure to provide fresh insights, as will in vivo pharmacologicalargeting.
eferences
[1] M.D. Cahalan, I. Parker, S.H. Wei, M.J. Miller, Two-photon tissueimaging: seeing the immune system in a fresh light, Nat. Rev. Immunol.2 (2002) 872–880.
[2] M.J. Miller, S.H. Wei, I. Parker, M.D. Cahalan, Two-photon imaging oflymphocyte motility and antigen response in intact lymph node, Science296 (2002) 1869–1873.
[3] M.J. Miller, A.S. Hejazi, S.H. Wei, M.D. Cahalan, I. Parker, T cellrepertoire scanning is promoted by dynamic dendritic cell behaviorand random T cell motility in the lymph node, Proc. Natl. Acad. Sci.USA 101 (2004) 998–1003.
[4] M.J. Miller, O. Safrina, I. Parker, M.D. Cahalan, Imaging the single celldynamics of CD4+ T cell activation by dendritic cells in lymph nodes,J. Exp. Med. 200 (2004) 847–856.
[5] T.E. DeCoursey, K.G. Chandy, S. Gupta, M.D. Cahalan, Voltage-gatedK+ channels in human T lymphocytes: a role in mitogenesis? Nature307 (1984) 465–468.
[6] J.C. Metcalfe, T. Pozzan, G.A. Smith, T.R. Hesketh, A calcium hypoth-esis for the control of cell growth, Biochem. Soc. Symp. 45 (1980)1–26.
[7] R.S. Lewis, M.D. Cahalan, Mitogen-induced oscillations of cytosolicCa2+ and transmembrane Ca2+ current in human leukemic T cells, CellRegul. 1 (1989) 99–112.
[8] J.W. Putney Jr., A model for receptor-regulated calcium entry, CellCalcium 7 (1986) 1–12.
[9] M. Hoth, R. Penner, Calcium release-activated calcium current in ratmast cells, J. Physiol. 465 (1993) 359–386.
10] M. Hoth, R. Penner, Depletion of intracellular calcium stores acti-vates a calcium current in mast cells, Nature 355 (1992) 353–356.
11] A. Zweifach, R.S. Lewis, Mitogen-regulated Ca2+ current of T lym-phocytes is activated by depletion of intracellular Ca2+ stores, Proc.Natl. Acad. Sci. USA 90 (1993) 6295–6299.
12] M.D. Cahalan, K.G. Chandy, T.E. DeCoursey, S. Gupta, A voltage-gated potassium channel in human T lymphocytes, J. Physiol. 358(1985) 197–237.
13] C.M. Fanger, H. Rauer, A.L. Neben, M.J. Miller, H. Wulff, J.C. Rosa,C.R. Ganellin, K.G. Chandy, M.D. Cahalan, Calcium-activated potas-sium channels sustain calcium signaling in T lymphocytes. Selectiveblockers and manipulated channel expression levels, J. Biol. Chem. 276(2001) 12249–12256.
14] S. Feske, J. Giltnane, R. Dolmetsch, L.M. Staudt, A. Rao, Gene regu-lation mediated by calcium signals in T lymphocytes, Nat. Immunol. 2(2001) 316–324.
15] P.A. Negulescu, N. Shastri, M.D. Cahalan, Intracellular calcium depen-dence of gene expression in single T lymphocytes, Proc. Natl. Acad.Sci. USA 91 (1994) 2873–2877.
16] C.M. Fanger, M. Hoth, G.R. Crabtree, R.S. Lewis, Characterizationof T cell mutants with defects in capacitative calcium entry: genetic
evidence for the physiological roles of CRAC channels, J. Cell Biol.131 (1995) 655–667.17] R.E. Dolmetsch, R.S. Lewis, C.C. Goodnow, J.I. Healy, Differentialactivation of transcription factors induced by Ca2+ response amplitudeand duration, Nature 386 (1997) 855–858.

ll Calc
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
M.D. Cahalan et al. / Ce
18] R.E. Dolmetsch, K. Xu, R.S. Lewis, Calcium oscillations increasethe efficiency and specificity of gene expression, Nature 392 (1998)933–936.
19] R.C. Hardie, P. Raghu, Activation of heterologously expressedDrosophila TRPL channels: Ca2+ is not required and InsP3 is notsufficient, Cell Calcium 24 (1998) 153–163.
20] D. Cordova, V.R. Delpech, D.B. Sattelle, J.J. Rauh, Spatiotemporalcalcium signaling in a Drosophila melanogaster cell line stably express-ing a Drosophila muscarinic acetylcholine receptor, Invert. Neurosci.5 (2003) 19–28.
21] J.E. Swatton, S.A. Morris, F. Wissing, C.W. Taylor, Functional prop-erties of Drosophila inositol trisphosphate receptors, Biochem. J. 359(2001) 435–441.
22] P.R. Towers, D.B. Sattelle, A Drosophila melanogaster cell line (S2)facilitates post-genome functional analysis of receptors and ion chan-nels, Bioessays 24 (2002) 1066–1073.
23] S. Yagodin, R.C. Hardie, S.J. Lansdell, N.S. Millar, W.T. Mason, D.B.Sattelle, Thapsigargin and receptor-mediated activation of DrosophilaTRPL channels stably expressed in a Drosophila S2 cell line, CellCalcium 23 (1998) 219–228.
24] S. Yagodin, N.B. Pivovarova, S.B. Andrews, D.B. Sattelle, Functionalcharacterization of thapsigargin and agonist-insensitive acidic Ca2+
stores in Drosophila melanogaster S2 cell lines, Cell Calcium 25 (1999)429–438.
25] M.D. Adams, S.E. Celniker, R.A. Holt, C.A. Evans, J.D. Gocayne,P.G. Amanatides, S.E. Scherer, P.W. Li, R.A. Hoskins, R.F. Galle, R.A.George, S.E. Lewis, S. Richards, M. Ashburner, S.N. Henderson, G.G.Sutton, J.R. Wortman, M.D. Yandell, Q. Zhang, L.X. Chen, R.C. Bran-don, Y.H. Rogers, R.G. Blazej, M. Champe, B.D. Pfeiffer, K.H. Wan,C. Doyle, E.G. Baxter, G. Helt, C.R. Nelson, G.L. Gabor, J.F. Abril, A.Agbayani, H.J. An, C. Andrews-Pfannkoch, D. Baldwin, R.M. Ballew,A. Basu, J. Baxendale, L. Bayraktaroglu, E.M. Beasley, K.Y. Beeson,P.V. Benos, B.P. Berman, D. Bhandari, S. Bolshakov, D. Borkova, M.R.Botchan, J. Bouck, P. Brokstein, P. Brottier, K.C. Burtis, D.A. Busam,H. Butler, E. Cadieu, A. Center, I. Chandra, J.M. Cherry, S. Cawley, C.Dahlke, L.B. Davenport, P. Davies, B. de Pablos, A. Delcher, Z. Deng,A.D. Mays, I. Dew, S.M. Dietz, K. Dodson, L.E. Doup, M. Downes, S.Dugan-Rocha, B.C. Dunkov, P. Dunn, K.J. Durbin, C.C. Evangelista,C. Ferraz, S. Ferriera, W. Fleischmann, C. Fosler, A.E. Gabrielian, N.S.Garg, W.M. Gelbart, K. Glasser, A. Glodek, F. Gong, J.H. Gorrell, Z.Gu, P. Guan, M. Harris, N.L. Harris, D. Harvey, T.J. Heiman, J.R. Her-nandez, J. Houck, D. Hostin, K.A. Houston, T.J. Howland, M.H. Wei,C. Ibegwam, et al., The genome sequence of Drosophila melanogaster,Science 287 (2000) 2185–2195.
26] A.V. Yeromin, J. Roos, K.A. Stauderman, M.D. Cahalan, A store-operated calcium channel in Drosophila S2 cells, J. Gen. Physiol. 123(2004) 167–182.
27] M. Ramet, P. Manfruelli, A. Pearson, B. Mathey-Prevot, R.A.Ezekowitz, Functional genomic analysis of phagocytosis and identi-fication of a Drosophila receptor for E. coli, Nature 416 (2002) 644–648.
28] I. Schneider, Cell lines derived from late embryonic stages ofDrosophila melanogaster, J. Embryol. Exp. Morphol. 27 (1972)353–365.
29] C.A. Worby, N. Simonson-Leff, J.E. Dixon, RNA interference of geneexpression (RNAi) in cultured Drosophila cells, Sci. STKE (2001),PL1.
30] J. Ulvila, M. Parikka, A. Kleino, R. Sormunen, R.A. Ezekowitz, C.Kocks, M. Ramet, Double-stranded RNA is internalized by scavengerreceptor-mediated endocytosis in Drosophila S2 cells, J. Biol. Chem.281 (2006) 14370–14375.
31] S. Feske, Y. Gwack, M. Prakriya, S. Srikanth, S.H. Puppel, B. Tanasa,
P.G. Hogan, R.S. Lewis, M. Daly, A. Rao, A mutation in Orai1 causesimmune deficiency by abrogating CRAC channel function, Nature 441(2006) 179–185.32] M. Vig, C. Peinelt, A. Beck, D.L. Koomoa, D. Rabah, M. Koblan-Huberson, S. Kraft, H. Turner, A. Fleig, R. Penner, J.P. Kinet, CRACM1
[
ium 42 (2007) 133–144 143
is a plasma membrane protein essential for store-operated Ca2+ entry,Science 312 (2006) 1220–1223.
33] S.L. Zhang, A.V. Yeromin, X.H. Zhang, Y. Yu, O. Safrina, A. Penna, J.Roos, K.A. Stauderman, M.D. Cahalan, Genome-wide RNAi screen ofCa2+ influx identifies genes that regulate Ca2+ release-activated Ca2+
channel activity., Proc. Natl. Acad. Sci. USA 103 (2006) 9357–9362.34] J. Roos, P.J. DiGregorio, A.V. Yeromin, K. Ohlsen, M. Lioudyno,
S. Zhang, O. Safrina, J.A. Kozak, S.L. Wagner, M.D. Cahalan, G.Velicelebi, K.A. Stauderman, STIM1, an essential and conserved com-ponent of store-operated Ca2+ channel function, J. Cell Biol. 169 (2005)435–445.
35] J. Roos, K.A. Stauderman, G. Velicelebi, K. Ohlsen, P.J. DiGrego-rio, Methods of modulating and of identifying agents that modulateintracellular calcium, in: WO2004/078995, 2003.
36] J. Liou, M.L. Kim, W.D. Heo, J.T. Jones, J.W. Myers, J.E. Ferrell Jr.,T. Meyer, STIM is a Ca2+ sensor essential for Ca2+-store-depletion-triggered Ca2+ influx, Curr. Biol. 15 (2005) 1235–1241.
37] S.L. Zhang, Y. Yu, J. Roos, J.A. Kozak, T.J. Deerinck, M.H. Ellisman,K.A. Stauderman, M.D. Cahalan, STIM1 is a Ca2+ sensor that acti-vates CRAC channels and migrates from the Ca2+ store to the plasmamembrane, Nature 437 (2005) 902–905.
38] P.B. Stathopulos, G.Y. Li, M.J. Plevin, J.B. Ames, M. Ikura, Stored Ca2+
depletion-induced oligomerization of stromal interaction molecule 1(STIM1) via the EF-SAM region: An initiation mechanism for capac-itive Ca2+ entry, J. Biol. Chem. 281 (2006) 35855–35862.
39] S.S. Manji, N.J. Parker, R.T. Williams, L. Van Stekelenburg, R.B. Pear-son, M. Dziadek, P.J. Smith, STIM1: a novel phosphoprotein locatedat the cell surface, Biochim. Biophys. Acta 1481 (2000) 147–155.
40] M.A. Spassova, J. Soboloff, L.P. He, W. Xu, M.A. Dziadek, D.L.Gill, STIM1 has a plasma membrane role in the activation of store-operated Ca2+ channels, Proc. Natl. Acad. Sci. USA 103 (2006) 4040–4045.
41] J.J. Lopez, G.M. Salido, J.A. Pariente, J.A. Rosado, Interaction ofSTIM1 with endogenously expressed human canonical TRP1 upondepletion of intracellular Ca2+stores, J. Biol. Chem. 281 (2006)28254–28264.
42] J.C. Mercer, W.I. Dehaven, J.T. Smyth, B. Wedel, R.R. Boyles, G.S.Bird, J.W. Putney Jr., Large store-operated calcium selective currentsdue to co-expression of Orai1 or Orai2 with the intracellular calciumsensor, Stim1, J. Biol. Chem. 281 (2006) 24979–24990.
43] M.M. Wu, J. Buchanan, R.M. Luik, R.S. Lewis, Ca2+ store depletioncauses STIM1 to accumulate in ER regions closely associated with theplasma membrane, J. Cell Biol. 174 (2006) 803–813.
44] C.T. Hauser, R.Y. Tsien, A hexahistidine-Zn2+-dye label reveals STIM1surface exposure, Proc. Natl. Acad. Sci. USA 104 (2007) 3693–3697.
45] O. Mignen, J.L. Thompson, T.J. Shuttleworth, STIM1 regulates Ca2+
entry via arachidonate-regulated Ca2+-selective (ARC) channels with-out store-depletion or translocation to the plasma membrane, J. Physiol.(2006).
46] P.T. Ellinor, J. Yang, W.A. Sather, J.F. Zhang, R.W. Tsien, Ca2+ channelselectivity at a single locus for high-affinity Ca2+ interactions, Neuron.15 (1995) 1121–1132.
47] T. Voets, A. Janssens, G. Droogmans, B. Nilius, Outer pore archi-tecture of a Ca2+-selective TRP channel, J. Biol. Chem. 279 (2004)15223–15230.
48] A.V. Yeromin, S.L. Zhang, W. Jiang, Y. Yu, O. Safrina, M.D. Cahalan,Molecular identification of the CRAC channel by altered ion selectivityin a mutant of Orai, Nature 443 (2006) 226–229.
49] M. Prakriya, S. Feske, Y. Gwack, S. Srikanth, A. Rao, P.G. Hogan,Orai1 is an essential pore subunit of the CRAC channel, Nature 443(2006) 230–233.
50] M. Vig, A. Beck, J.M. Billingsley, A. Lis, S. Parvez, C. Peinelt,
D.L. Koomoa, J. Soboloff, D.L. Gill, A. Fleig, J.P. Kinet, R. Penner,CRACM1 multimers form the ion-selective pore of the CRAC channel,Curr. Biol. 16 (2006) 2073–2079.51] R.M. Luik, M.M. Wu, J. Buchanan, R.S. Lewis, The elementary unitof store-operated Ca2+ entry: local activation of CRAC channels by

1 ll Calc
[
[
[operated calcium channels in T lymphocytes, J. Gen. Physiol. 107(1996) 597–610.
44 M.D. Cahalan et al. / Ce
STIM1 at ER-plasma membrane junctions, J. Cell. Biol. 174 (2006)815–825.
52] Y. Gwack, S. Srikanth, S. Feske, F. Cruz-Guilloty, M. Oh-Hora, D.S.
Neems, P.G. Hogan, A. Rao, Biochemical and functional characteriza-tion of Orai family proteins, J. Biol. Chem. (2007).53] A. Lepple-Wienhues, M.D. Cahalan, Conductance and permeationof monovalent cations through depletion-activated Ca2+ channels(ICRAC) in Jurkat T cells, Biophys. J. 71 (1996) 787–794.
[
ium 42 (2007) 133–144
54] A. Zweifach, R.S. Lewis, Calcium-dependent potentiation of store-
55] A. Zweifach, R.S. Lewis, Rapid inactivation of depletion-activated cal-cium current (ICRAC) due to local calcium feedback, J. Gen. Physiol.105 (1995) 209–226.





























