Embed Size (px)
Citation preview

1 This article is protected by copyright. All rights reserved.
Molecular and physiological responses of trees to waterlogging stress1 1
2
Jürgen Kreuzwieser1*
, Heinz Rennenberg1 3
4
1Institute of Forest Science, Chair of Tree Physiology, Albert-Ludwigs-Universität 5
Freiburg; Georges-Köhler-Allee 53; 79110 Freiburg, Germany 6
7
8
9
10
*corresponding author and present address: 11
Institute of Forest Science, Chair of Tree Physiology, Albert-Ludwigs-Universität 12
Freiburg; Georges-Köhler-Allee 53; 79110 Freiburg, Germany 13
Email : [email protected] 14
15
16
17
18
19
20
21
This article has been accepted for publication and undergone full peer review but has not
been through the copyediting, typesetting, pagination and proofreading process, which
may lead to differences between this version and the Version of Record. Please cite this
article as doi: 10.1111/pce.12310
Acc
epte
d A
rticl
e

2 This article is protected by copyright. All rights reserved.
22
Abstract 23
One major effect of global climate change will be altered precipitation patterns in many 24
regions of the world. This will cause a higher probability of long-term waterlogging in 25
winter/spring and flash floods in summer due to extreme rainfall events. Particularly trees 26
not adapted at their natural site to such waterlogging stress can be impaired. Despite the 27
enormous economic, ecological and social importance of forest ecosystems, the effect of 28
waterlogging on trees is far less understood than the effect on many crops or the model 29
plant Arabidopsis. There is only a handful of studies available investigating the 30
transcriptome and metabolome of waterlogged trees. Main physiological responses of trees 31
to waterlogging include the stimulation of fermentative pathways and an accelerated 32
glycolytic flux. Many energy consuming, anabolic processes are slowed down to 33
overcome the energy crisis mediated by waterlogging. A crucial feature of waterlogging 34
tolerance is the steady supply of glycolysis with carbohydrates, particularly in the roots; 35
stress sensitive trees fail to maintain sufficient carbohydrate availability resulting in the 36
dieback of the stressed tissues. The present review summarizes physiological and 37
molecular features of waterlogging tolerance of trees; the focus is on carbon metabolism in 38
both, leaves and roots of trees. 39
40
Keywords: waterlogging, hypoxia, trees, carbon metabolism, nitrogen metabolism, 41
transcriptome, metabolome. 42
43
44
Acc
epte
d A
rticl
e

3 This article is protected by copyright. All rights reserved.
45
Introduction 46
47
The concentrations of the trace gases carbon dioxide, methane and nitrous oxide in the 48
atmosphere are continuously rising due to anthropogenic activity. Between the pre-49
industrial era and 2005 they increased by 36% (CO2; from 280 to 379 ppm), 148% (CH4; 50
from 715 to 1774 ppb) and 18% (N2O; from 270 to 319 ppb) (Forster et al., 2007). 51
Consequently, the global air temperatures considerably increased between 1850 and 2007 52
and will further increase in the future (Christensen et al., 2007). In Central Europe, for 53
example, a temperature rise in the range of 1.9-7.0°C is expected in the next 50 years 54
(Frei, 2004). Such temperature elevation will strongly affect the global hydrological cycle. 55
Like in North and Central Europe, annual precipitation in East Africa, Northern, East, 56
South and Southeast Asia, Canada and Northeast USA is likely to increase whereas it will 57
decrease in Central America, Southwest USA, Mediterranean Europe, and Central Asia 58
(Christensen et al., 2007). However, precipitation will not change equally over the year; in 59
Central Europe, winter precipitation is predicted to increase in the future, but summer 60
precipitation will be considerably lower causing a higher possibility of drought periods 61
during the summer months (Frei et al., 2006). On the other hand, most model projections 62
forecast increased extreme precipitation events despite decreased mean summer 63
precipitation (Palmer and Räisänen, 2002; Christensen and Christensen, 2003). Because of 64
such intense rainfall events, terrestrial ecosystems in the concerned regions will 65
experience more and probably longer waterlogging periods during winter and spring and 66
more extreme short-term flooding events during summer (Christensen and Christensen, 67
2003; Kundzewicz et al., 2005; Kundzewicz, 2006) particularly on compacted and / or 68
heavy, clay-rich soils where drainage is inhibited (Dennis and Grindley, 1983; Kozlowski, 69
1984). 70 Acc
epte
d A
rticl
e

4 This article is protected by copyright. All rights reserved.
71
Soil processes 72
Flooding (i.e. water standing above soil level) and waterlogging (i.e. only the soil is 73
flooded) lead to a deprivation of oxygen in the soil because the floodwater entering the 74
soil removes oxygen rich air from soil pores. In addition, the ca. 10.000 times higher 75
diffusion resistance of oxygen in water than in air leads to an inhibited supply of the soil 76
with oxygen in water saturated soils (Armstrong, 1979). Microbial and plant activities 77
quickly consume the remaining oxygen leading either to hypoxic (low oxygen 78
concentrations: mitochondrial respiration reduced; fermentation takes place) or anoxic 79
(oxygen absent: energy gain by fermentation only) conditions (Pradet and Bomsel, 1978). 80
Due to the lack of oxygen, soil physico-chemical properties such as pH and redox 81
potential strongly change during waterlogging (Pezeshki and Chambers, 1985a; 1985b). 82
The diminished gas diffusion velocity between the soil and the atmosphere causes an 83
accumulation of gaseous compounds in the waterlogged soil such as the plant hormone 84
ethylene or other metabolic products like carbon dioxide (Jackson, 1982). Oxygen 85
shortage further affects microbial communities in the soil (Unger et al., 2009) and 86
numerous microbial processes, which eventually cause changes in soil chemical 87
composition. This can lead to a reduced abundance of oxidised nutrients (e.g. NO3-, SO4
2-, 88
Fe3+
) and elevated levels of reduced compounds such as Mn2+
, Fe2+
, H2S, NH4+, and 89
organic compounds (alkanes, acids, carbonyls, etc) which can be toxic for plants 90
(Ponnamperuma, 1972; 1984; McKee and McKevlin, 1993; Snowden and Wheeler, 1993; 91
Lucassen et al., 2000; 2002; Jackson and Colmer, 2005). The velocity and extent of the 92
changes of soil physico-chemical properties depends on soil type, the duration of the 93
waterlogging event, prevailing environmental conditions (such as temperature) and the 94
type of flooding or waterlogging (Drew, 1997; Kozlowski, 1997). Stagnant conditions 95
reduce the oxygen availability in the soil much faster leading to stronger stress conditions 96
Acc
epte
d A
rticl
e

5 This article is protected by copyright. All rights reserved.
than moving floodwater where turbulences facilitate oxygen solubilisation in the 97
floodwater. 98
99
Effects on trees 100
About 31% of the terrestrial earth surface, i.e. around 4 billion hectares, is covered by 101
forest ecosystems with Latin America (sharing 24% of the world’s forest), former Soviet 102
Union (21%), Africa (20%), and North-America (16%) as the regions with the largest 103
forested areas (FAO, 2012; The Columbia Electronic Encyclopedia, 2012). It is obvious 104
that forests fulfil a plethora of essential ecological (e.g. maintenance of biodiversity, 105
involvement in biogeochemical cycles of water, carbon and nitrogen), economic (e.g. 106
timber and other natural resources, energy source) and social (e.g. recreation) functions. In 107
addition, trees are of particular importance in urban environments as street and park 108
vegetation (Dwyer et al., 1991) or in orchards and as bioenergy source from fast growing 109
plantations, the latter covering 187 million hectares in 2000, with a strongly increasing 110
trend (Carnus et al., 2006). 111
112
Because major parts of the landscapes covered by trees/forests are assumed to be more 113
strongly exposed to waterlogging and flooding in the future, plant mechanisms to cope 114
with such stress conditions are of particular interest. However, today the knowledge on 115
physiological and molecular aspects of flooding/waterlogging tolerance in trees is far 116
behind that of herbaceous species (Kreuzwieser et al., 2009). This is due to the fact that 117
studying trees provides particular challenges. Trees are characterised by longevity, making 118
it difficult to work with adult trees under environmentally controlled conditions. The 119
combination of long lifetime and seasonality complicates many plant internal processes: 120
alternating phases of dormancy and growth over the growing season need well-adjusted 121
storage and remobilization processes in order to support meristematic tissues with 122
Acc
epte
d A
rticl
e

6 This article is protected by copyright. All rights reserved.
nutrients (Tuskan et al., 2003). Supply of growing tissues with nutrients depends on highly 123
orchestrated and regulated long-distance transport processes in phloem and xylem. 124
Storage, mobilization, and long-distance transport can all be affected in different ways by 125
environmental factors such as soil oxygen deficiency. The lack of knowledge on 126
mechanisms of environmental control of these processes in trees compared to crops and 127
model plants such as Arabidopsis or rice becomes particularly evident at the molecular 128
level (Kreuzwieser et al., 2009; Mustroph et al., 2010; Christianson et al., 2010; Narsai et 129
al., 2011). This is partially due to the limited availability of relevant tools and techniques. 130
For example, the first commercially distributed microarray for a tree species became 131
available for poplar only in 2006, after the genome of Populus trichocarpa has been 132
sequenced (Tuskan et al., 2006). As nicely reviewed by Neale and Kremer (2011), forest 133
tree genomics made great progress in recent years as modern sequencing technologies 134
(next generation sequencing, NGS) considerably facilitate tree genome studies and 135
transcriptome profiling (RNA-seq). This certainly will allow faster progress of research on 136
trees in near future. 137
138
As aerobic organisms, trees depend on a steady supply with oxygen to all living cells, and 139
interruption from oxygen availability therefore causes disturbance of plant metabolism 140
(Drew, 1997; Bailey-Serres and Voesenek, 2008). Depending on the tolerance of soil 141
oxygen depletion, this can cause dysfunction of processes at the cellular level, eventually 142
leading to visible damages. Flooding and waterlogging tolerance and the occurrence of 143
injuries is strongly species-dependent (Table 1; Gill, 1970; Kozlowski, 1982; 1997; 144
McClean, 2000; Glenz et al., 2006; Niinemets and Valladares, 2006; Kramer et al., 2008; 145
Parolin et al., 2004; 2010; Ferner et al., 2012). Highly adapted species survive 146
waterlogging periods up to some months without any injuries (Table 1), but less tolerant 147
or sensitive species can develop damages already after a few hours of oxygen deprivation 148
Acc
epte
d A
rticl
e

7 This article is protected by copyright. All rights reserved.
(see Kozlowski, 1997; Glenz et al., 2006). Besides a reduction of root (see refs in 149
Kozlowski, 1997) and shoot growth (e.g. Colin-Belgrand et al., 1990; Pezeshki et al., 150
1996; Parolin, 2001; Ye et al., 2003; Alaoui-Sossé et al., 2005; Mielke et al., 2005; Parelle 151
et al., 2006; Neatrour et al., 2007; Ferreira et al., 2007; de Oliveira and Joly, 2010; Ferry et 152
al., 2010; Guo et al., 2011), typical symptoms of soil oxygen shortage in sensitive trees are 153
leaf necrosis and shedding, bark damages, elevated susceptibility to fungal and insect 154
pathogens, or dieback of the whole tree (Kozlowski, 1997; Parolin, 2001; Kreuzwieser et 155
al., 2004; Parolin and Wittmann, 2010). The extent of damages depends on the type 156
(stagnant or moving water), duration and height of flooding, the environmental conditions 157
during the stress event (e.g. air, water and soil temperature, solar radiation), the season, 158
but also on a wide range of plant specific features (Kozlowski, 1997; Vreugdenhil et al., 159
2006). The development of damages depends to a high degree on the species considered 160
but also on the ecotype of a given species (Jaeger et al., 2009; Guo et al. 2011), the tree’s 161
age, size and developmental stage (Kozlowski, 1997; Siebel and Blom, 1998; Glenz et al., 162
2006). It is generally observed that adult trees tolerate waterlogging and flooding better 163
than seedlings of the same species (Table 2) (Gill, 1970; Siebel and Blom, 1998). Recent 164
studies even indicated that the sex of a tree plays a role in flooding tolerance of dioecious 165
species such as willow and poplar. There is clear evidence that female willow and poplar 166
trees are more tolerant against the stress than male trees explaining the spatial segregation 167
of the sexes with higher abundance of females in low-elevation zones of riparian forests 168
(Hultine et al. 2007; Nielsen et al. 2010). Nielsen et al. (2010) therefore proposed the 169
concept of “strategic positioning” where the seed-producing female trees are better 170
adapted to sites which are more often flooded and where seedling recruitment usually 171
occurs. 172
173
Tree adaptation to flooding 174
Acc
epte
d A
rticl
e

8 This article is protected by copyright. All rights reserved.
Tree species inhabiting ecosystems, which are regularly exposed to flooding, evolved a 175
broad range of adaptive strategies to cope with the stress mediated by this exposure. Most 176
wetland species apply avoidance strategies based on morphological-anatomical features. 177
Such adaptations have been reviewed for woody species by Kozlowski (1997) and Glenz 178
et al. (2006), and are therefore only briefly mentioned here. Many flood tolerant species 179
develop hypertrophied lenticels at the stem base in response to flooding (compilations of 180
tree species are given by Kozlowski (1997) and Glenz et al. (2006)). These organs 181
penetrate the relatively strongly gas resistant phellogen layer of the trees, enabling gas 182
exchange between stem and environment. Thus, hypertrophied lenticels allow oxygen 183
uptake into the plant, but they are also assumed to contribute to the release of gaseous 184
compounds (carbon dioxide, acetaldehyde, ethanol) out of the stem into the atmosphere 185
(Li et al., 2006; Shimamura et al., 2010). Another feature often associated with the 186
appearance of hypertrophied lenticels is the formation of adventitious roots (Glenz et al., 187
2006) (Fig. 1). Such roots are produced when the primary root system of the tree is 188
impaired because of soil oxygen deficiency. Adventitious roots possess a high portion of 189
intercellular spaces facilitating longitudinal oxygen transport. In a studies with Central 190
Amazonian trees such as Salix martiana and Tabernaemontana juruana, it was 191
demonstrated that the main entry point of atmospheric oxygen were gas-permeable pores 192
in the stem near the origin of the adventitious roots (Haase et al., 2003; Haase & Rätsch, 193
2010). Uptake of oxygen seems also to be possible along the root if it is growing at the 194
water surface being in contact with the atmosphere (Haase et al., 2003). Such oxygen 195
uptake is required for the maintenance of mitochondrial respiration; it further allows radial 196
oxygen loss (ROL) from the roots which contributes to the oxidation of the rhizosphere 197
(Kludze et al., 1994; Li et al., 2006). Further important functions of adventitious roots are 198
absorption of water and nutrients as a replacement of the damaged primary root system 199
(Barlow, 1986; Calvo-Polanco et al., 2012). Impressive examples for adventitious roots 200
Acc
epte
d A
rticl
e

9 This article is protected by copyright. All rights reserved.
are the prop roots of Rhizophora and the pneumatophores of Avicennia, both inhabitants of 201
mangrove ecosystems (Baylis, 1950; Allaway et al., 2001; Aziz and Khan, 2001) (Fig. 1). 202
The formation of aerenchyma is a third adaptation of plants to cope with oxygen 203
deficiency in the soil (Kozlowski, 1997; Kludze et al., 2004). The formation of lacunae air 204
spaces enhances the porosity of the root tissue facilitating oxygen diffusion within roots 205
and they are likely to contribute to the export of phytotoxic volatile metabolites 206
(acetaldehyde, ethanol) from the plant (Visser et al., 1997). The formation of 207
hypertrophied lenticels, adventitious roots and aerenchyma depends on the accumulation 208
of ethylene in plant tissue (Steffens et al., 2006; Bailey-Serres et al., 2012). Plant internal 209
concentrations of ethylene increase, if floodwater surrounding the plant inhibits the 210
diffusive loss of this volatile plant hormone into the atmosphere, and if – at the same time 211
– ethylene biosynthesis proceeds (Bailey-Serres and Voesenek, 2008). In addition to 212
ethylene, reactive oxygen species (ROS) are involved as signalling intermediates in this 213
ethylene-controlled adaptation (Steffens et al., 2013). Other components also known to 214
play a role in adventitious root formation are the auxin indole acetic acid (IAA) and NO 215
(see Bailey-Serres et al., 2012). IAA abundance induces a transient accumulation of NO 216
(Pagnussat et al., 2002) which in turn activates a MAPK signalling cascade eventually 217
leading to adventitious root formation (Pagnussat et al. 2004). Such knowledge has been 218
gained mainly by studies with herbaceous plants; the mechanisms of hypoxia induced 219
adventitious root formation in trees and particularly the interplay of the different 220
components are still widely unknown. 221
222
223
Physiological effects of waterlogging on trees 224
225
Net CO2 assimilation and stomatal conductance of waterlogged trees 226
Acc
epte
d A
rticl
e

10 This article is protected by copyright. All rights reserved.
One of the most often studied physiological processes during waterlogging of trees is leaf 227
gas exchange, particularly net CO2 assimilation. It is a general phenomenon that 228
assimilation rates tend to decrease during periods of waterlogging stress as observed in 229
trees of tropical (Nunez-Elisea et al., 1999; Fernandez et al., 1999; Ojeda et al., 2004; 230
Fernandez, 2006; Herrera, 2013) and temperate (Pezeshki and Chambers 1985a; 1985b; 231
1986; Pezeshki, 1994; Pezeshki et al., 1996, Dreyer et al., 1991; Reece and Riha, 1991; 232
Beckman et al., 1992; Ewing, 1996; Gravatt and Kirkby, 1998; Jaeger et al., 2009; Ferner 233
et al., 2012) ecosystems. The extent of this decrease depends on the species’ tolerance to 234
soil oxygen deficiency. Highly tolerant trees maintain rates of photosynthesis at a 235
relatively high level or are even unaffected by the stress, whereas net CO2 assimilation of 236
less tolerant or sensitive species is strongly reduced (Dreyer, 1994; Wagner and Dreyer, 237
1997; Graves et al., 2002; Vu and Yelenosky, 2006; Jaeger et al., 2009; Parent et al., 2011; 238
Ferner et al., 2012). In addition, as seen in flood tolerant tree species of the Amazonian 239
floodplain, rates of photosynthesis can completely recover or even increase during long-240
term periods of soil oxygen deficiency; such recovery often coincides with morpho-241
anatomical changes like the appearance of hypertrophied lenticels and/or adventitious 242
roots (Herrera, 2013). 243
244
The reasons for inhibited photosynthesis of waterlogged trees are still not completely 245
understood. There are strong hints that both, non-stomatal and stomatal limitations are 246
involved. Non-stomatal limitation is associated with lowered pigment concentrations in 247
leaves of waterlogged trees (Kreuzwieser et al., 2002; Ojeda et al., 2004), decreased 248
activity (Vu and Yelenosky, 2006) and abundance (Herrera, 2013) of ribulose-1,5-249
bisphosphate carboxylase/oxygenase (Rubisco) and accumulation of soluble carbohydrates 250
which might cause feedback inhibition of photosynthesis (Iglesias et al., 2002; Islam and 251
MacDonald, 2004; Rengifo et al., 2005; Vu and Yelenosky, 2006; Jaeger et al., 2009; 252
Acc
epte
d A
rticl
e

11 This article is protected by copyright. All rights reserved.
Ferner et al., 2012). On the other hand, waterlogging causes stomatal closure, which has 253
been discussed as the main reason for reduced photosynthesis in numerous trees (Reece 254
and Riha, 1991; Gravatt and Kirkby, 1998, Pezeshki et al., 1996; Schmull and Thomas, 255
2000; Rodríguez-Gamir et al., 2011; Ferner et al., 2012). Although intensively studied, the 256
mechanisms leading to reduced stomatal conductance during periods of waterlogging is far 257
from being understood. It is assumed to be connected to reduced root hydraulic 258
conductivity and subsequently lowered water absorption by the roots and/or caused by 259
chemical signals, which are transported from waterlogged roots to the shoot via the 260
transpiration stream. The nature of potential signalling compounds responsible for 261
stomatal closure is still not known (Else et al., 1996; 2006; Aroca et al., 2011). The 262
involvement of abscisic acid (ABA) transport in the xylem has been proposed (Jackson et 263
al., 2003; Herrera, 2013) but is still a matter of debate, particularly since some reports 264
clearly excluded root-to-shoot ABA transport (Else et al., 2006; Rodríguez-Gamir et al., 265
2011). Waterlogging induced changes in the pH of the xylem sap have also been proposed 266
as a long-distance signal. However, there does not seem to be a consistent plant response 267
as both alkalisation (Else et al., 2006) and acidification (Rodríguez-Gamir et al., 2011) of 268
the xylem sap was observed in waterlogged plants. Recently, mobilization of root stored 269
sulphate and its root-to-shoot transport have been proposed to mediate stomatal closure in 270
response to drought (Ernst et al., 2010); whether these processes are also involved in 271
stomatal closure in response to waterlogging remains to be elucidated. 272
273
Reduced root hydraulic conductance 274
Waterlogging induced reduction of stomatal conductance is often associated with 275
diminished root hydraulic conductance as demonstrated in many flood tolerant and 276
sensitive tree species (Syvertsen et al., 1983; Dreyer, 1994; Schmull and Thomas, 2000; 277
Islam et al., 2003; Nicolás et al., 2005; Rodríguez-Gamir et al., 2011). The phenomenon 278
Acc
epte
d A
rticl
e

12 This article is protected by copyright. All rights reserved.
seems to be more pronounced in the latter group of trees and can be due to a higher degree 279
of damage of their root system and/or lower root biomass due to impaired growth 280
(Schmull and Thomas, 2000). Water absorption by roots is at least partially (ca. 50%) 281
mediated by root water channels (aquaporins) of the plasma membrane intrinsic protein 282
(PIP) family (Tournaire-Roux et al., 2003; McElrone et al., 2007). It can therefore be 283
assumed that modifications of the channel protein or down-regulation of aquaporin 284
expression and subsequent reduced protein abundance might contribute to reduced water 285
permeability of waterlogged roots. The water transport across the plasmalemma can be 286
slowed-down efficiently if a histidine residue of PIPs is protonated, a process depending 287
on acidification of the cytosol (Tournaire-Roux et al., 2003). This cytosolic decrease of pH 288
is a common phenomenon in roots of waterlogged plants that occurs within minutes (Gout 289
et al., 2001). It is assumed to be caused by (i) a passive influx of protons from the external 290
solution or from the vacuole, (ii) the hydrolysis of nucleoside triphosphates and sugar 291
phosphates, and (iii) the accumulation of organic acids including the biosynthesis of lactic 292
acid (Davies et al., 1974; Felle, 2001; Gout et al., 2001). Besides modulation of the 293
aquaporin protein, down-regulation of aquaporin gene expression has also been reported in 294
some tree species (Fig. 2) (Kreuzwieser et al., 2009; LeProvost et al., 2011; Rodríguez-295
Gamir et al., 2011). The question arises, how changes in root hydraulic conductance can 296
be translated into altered stomatal conductance in the leaves. As mentioned above, root-to-297
shoot transport of signalling compounds might be one option. Another widely ignored 298
possibility of root-to-shoot communication is hydraulic signalling. This mechanism of 299
signalling is supposed to maintain water homeostasis in drought stressed plants as recently 300
reviewed by Christmann et al. (2013). It depends on the transfer of a hydraulic signal, i.e. 301
a change in plant water potential, in the xylem and conversion of this signal into a 302
biochemical signal in target cells. A hydraulic – so far unidentified - sensor in leaf cells 303
could initiate a signaling cascade eventually causing ABA biosynthesis leading to the 304
Acc
epte
d A
rticl
e

13 This article is protected by copyright. All rights reserved.
closure of stomata (Christmann et al., 2013). The idea of hydraulic signaling is supported 305
by reduced water potential in a variety of waterlogged trees that usually is much stronger 306
in flood sensitive than in flood tolerant species (Ruiz-Sánchez et al., 1996; Nicolás et al., 307
2005; Ortuno et al., 2007; Parent et al., 2011; Striker, 2012; Herrera, 2013). 308
Figure 3 summarizes some aspects of plant water relations in waterlogged trees; the switch 309
from normoxia to hypoxia/anoxia by soil waterlogging, results in major changes of root 310
metabolism (see chapters below). This metabolic adjustment is associated with a drop of 311
cytoplasmic pH in root cells (Davies et al., 1974; Roberts et al., 1984; Felle, 2001; Gout et 312
al., 2001), which causes protonation of the PIPs responsible for water absorption by roots. 313
Consequently, root hydraulic resistance increases thereby inhibiting water uptake and 314
affecting plant water status. A hydraulic signal or a chemical signal of yet unknown nature, 315
which is transferred / transported in the xylem, communicates apparent water limitation to 316
the shoot, thereby initiating stomatal closure. 317
318
319
Molecular and physiological changes in roots of waterlogged trees 320
321
Waterlogging causes an enhanced glycolytic flux and fermentative processes 322
If stress avoidance strategies such as hypertrophied lenticels, adventitious roots or 323
aerenchyma are not yet formed or are overburden during a stress period, waterlogged roots 324
may become oxygen deficient and molecular and physiological tolerance mechanisms are 325
essential for plant survival. Several studies indicated that under such conditions major 326
changes occur in the metabolism of roots of trees. In flood tolerant poplar trees, for 327
example, over 2,000 differentially expressed genes were detected after 5 hours of hypoxic 328
treatment compared to roots under normoxic conditions. Less dramatic changes (ca. 1,000 329
differentially expressed genes) were observed in cotton roots, a flood sensitive woody 330
Acc
epte
d A
rticl
e

14 This article is protected by copyright. All rights reserved.
species, hypoxically treated for 4 hours (see Christianson et al., 2010). Such effects on the 331
trees’ transcriptome were accompanied by strong changes in the root metabolite profile 332
(Kreuzwieser et al., 2009). Because oxygen is the final electron acceptor of mitochondrial 333
respiration, this crucial process of plant energy metabolism is slowed down under hypoxia 334
or even completely inhibited under anoxia. Thus, ATP cannot be produced any longer by 335
aerobic respiration and the cells affected suffer from an energy crisis (Bailey-Serres and 336
Voesenek, 2008). Numerous molecular and physiological studies with flood tolerant and 337
sensitive tree species have demonstrated that fermentative pathways are stimulated under 338
such conditions. Consistent with observations in herbaceous plants (Davies et al., 1974; 339
Roberts et al., 1984), also trees seem to switch in response to hypoxia initially from 340
mitochondrial respiration to lactic acid fermentation. This switch has been documented at 341
both, the metabolite level and the level of lactate dehydrogenase (LDH) gene expression 342
(Joly and Crawford, 1982; Gout et al., 2001; Kolb et al., 2002; Kreuzwieser et al., 2009). 343
Because the accumulation of lactic acid leads to an acidification of the cytosolic pH 344
(Davies et al., 1974), LDH activity is lowered and alcoholic fermentation is stimulated. 345
This change in fermentation processes is suggested from increased pyruvate decarboxylase 346
(PDC) and alcohol dehydrogenase (ADH) activities in waterlogged roots, accompanied by 347
the formation of the intermediate acetaldehyde (Atkinson et al., 2008) and the end-product 348
ethanol (Joly and Crawford, 1982; Kreuzwieser et al., 1999; 2002; Kolb et al., 2002; 349
Jaeger et al., 2009; Ferner et al., 2012). Consistently, elevated transcript levels of PDC and 350
ADH are observed in a great variety of tree species (e.g. Christianson et al., 2010; 351
LeProvost et al., 2012); in poplar, elevated ADH and PDC transcript abundance appeared 352
in less than one hour after the change from normoxia to hypoxia (Kreuzwieser et al., 353
2009). There are hints that flood tolerant species exhibit higher rates of alcoholic 354
fermentation than flood sensitive trees (Porth et al. 2005; Parelle et al. 2006; LeProvost et 355 Acc
epte
d A
rticl
e

15 This article is protected by copyright. All rights reserved.
al., 2012), but such patterns are not consistent (Yamanoshita et al., 2005; Ferner et al., 356
2012). 357
358
The few studies on changes in the transcriptome of waterlogged woody species (poplar: 359
Kreuzwieser et al., 2009; cotton: Christianson et al., 2010; pedunculate and sessile oak: 360
LeProvost et al., 2012) suggest that together with an up-regulation of the genes of 361
fermentative pathways (mainly LDH, ADH and PDC), also the glycolytic flux is 362
enhanced, most likely in order to maintain ATP production under conditions of inhibited 363
mitochondrial respiration. In poplar and cotton, enhanced glycolytic flux is suggested from 364
an up-regulation of key enzymes of this pathway, i.e. phosphofructokinase and pyruvate 365
kinase (Kreuzwieser et al., 2009; Christianson et al., 2010). Accelerated glycolysis has 366
also been suggested from a transcriptome approach in which two oak species of different 367
flood tolerance were compared (LeProvost et al., 2012). Maintaining enhanced glycolysis 368
by a steady and sufficient supply with carbohydrates seems to be crucial for tree survival 369
under hypoxia. There are clear experimental evidences that flood tolerant and sensitive 370
species differ in their ability to maintain adequate carbohydrate supply over prolonged 371
periods of waterlogging (Vu and Yelenoski, 1991; Ferner et al., 2012; LeProvost et al., 372
2012). Whereas more sensitive trees deplete of soluble sugars already after some days of 373
waterlogging, more tolerant species maintain carbohydrate concentrations at a high level 374
(Fig. 4) (Herschbach et al., 2005; Jaeger et al., 2009; Martínez-Alcántara et al., 2012; 375
Ferner et al., 2012). The increased demand for soluble carbohydrates in roots of 376
waterlogged trees can at least partially be satisfied by degradation of starch reserves in 377
flood tolerant species (Kreuzwieser et al., 2009; LeProvost et al., 2012). Studies with 378
hypoxically treated poplar indicated another possibility likely to contribute to alleviate the 379
enhanced sugar demand in roots. Elevated levels of succinate together with an up-380
regulation of genes encoding for lipases and enzymes involved in fatty acid degradation, 381
Acc
epte
d A
rticl
e

16 This article is protected by copyright. All rights reserved.
as well as up-regulation of the isocitrate lyase and the malate synthase genes suggest that 382
the glyoxylate cycle is induced under hypoxia. This pathway is assumed to link lipid 383
degradation with carbohydrate biosynthesis (Eastmond et al., 2000); in waterlogged 384
poplar, it could channel acetyl-CoA formed from β-oxidation of fatty acids into sugar 385
biosynthesis (Kreuzwieser et al., 2009). Degradation of fatty acids has also been observed 386
in some neo-tropical tree species due to prolonged waterlogging (Kolb et al., 2002). 387
Elevated sugar consumption in hypoxia stressed roots could also be compensated by 388
enhanced shoot-to-root transport of photo-assimilates in the phloem (Kreuzwieser et al., 389
2009; Merchant et al., 2010). Moreover, some studies underlined the importance of altered 390
sucrose degradation during hypoxia stress. Changes in root transcript levels indicate that 391
sucrose cleavage via invertases is replaced by phosphorolytic degradation via sucrose 392
synthase (Kreuzwieser et al., 2009; Christianson et al., 2010a; 2010b; LeProvost et al., 393
2012). From an energetic point of view, this switch makes sense, because sucrose synthase 394
uses only one molecule of pyrophosphate as a substrate during sucrose cleavage, whereas 395
invertases use two ATP molecules. The question arises why normoxic roots do not use this 396
pathway as well. Several studies indicated that under non-stress conditions sucrose 397
synthase delivers UDP-glucose mainly for the well-regulated process of cellulose 398
biosynthesis (Albrecht and Mustroph, 2003). Up-regulated UDP-glucose delivery could 399
impair this process. However, under hypoxic conditions cellulose biosynthetic is strongly 400
down-regulated (Kogawara et al., 2006; Kreuzwieser et al., 2009) and it seem 401
energetically advantageous if UDP-glucose is channelled into glycolysis. 402
It seems to be paradox that - particularly in flood sensitive but to a minor extent also in 403
flood tolerant species - root flooding leads to an accumulation of carbohydrate 404
concentrations in the leaves (Vu and Yelenosky, 1991; Gravatt and Kirby, 1998; Merchant 405
et al., 2010; Martínez-Alcántara et al., 2012; Ferner et al., 2012; Herrera, 2013). The even 406
more massive accumulation of soluble carbohydrates in the phloem of some waterlogged, 407
Acc
epte
d A
rticl
e

17 This article is protected by copyright. All rights reserved.
stress sensitive trees together with considerably reduced carbohydrate concentrations in 408
the roots, suggests that in flooding sensitive species the transport of sugars from the 409
phloem into root cells is dramatically impaired during flooding (Fig. 4) (Jaeger et al., 410
2009; Merchant et al., 2010; Ferner et al., 2012). Alternatively, the conversion of sucrose 411
into monosaccharides may severely be affected. Impaired shoot-to-root transport of photo-412
assimilates has been demonstrated directly from slowed down transport of the 13
C tracer 413
after 13
CO2 fumigation of leaves of flooded citrus trees (Martínez-Alcántara et al., 2012). 414
However, the exact mechanisms causing impaired supply of hypoxia treated roots with 415
photo-assimilates is still not fully understood and needs further investigations. 416
417
Trees become energy safers during periods of waterlogging 418
From the massive changes in transcript abundance observed in roots of poplar, cotton and 419
oak trees, it can be concluded that – in accordance with observations in herbaceous species 420
– numerous energy intensive processes are slowed down under hypoxic conditions 421
(Kreuzwieser et al., 2009; Christianson et al., 2010; LeProvost et al., 2012). For example, 422
the transcript levels of many genes encoding transporters are decreased suggesting 423
diminished nutrient uptake in waterlogged trees (Kreuzwieser and Gessler, 2010). This 424
assumption is supported by reduced rates of N uptake and strongly affected nitrate and 425
amino acid concentrations in roots of hypoxically treated trees (Kreuzwieser et al., 2002; 426
Alaoui-Sossé et al., 2005). Other energy consuming processes strongly affected in 427
waterlogged trees are the biosynthesis of cell wall components such as cellulose, 428
hemicellulose, and cell wall proteins, as well as lignin (Kreuzwieser et al., 2009; 429
Christianson et al., 2010a; b; LeProvost et al., 2012). Consistent with slowed down 430
anabolic processes, reduced growth is often observed in waterlogged trees; growth of 431
flooding sensitive trees is usually stronger impaired than that of more tolerant species (see 432
Kreuzwieser et al., 2004; Herrera 2013). 433
Acc
epte
d A
rticl
e

18 This article is protected by copyright. All rights reserved.
434
Changes in N metabolism of waterlogged roots 435
As mentioned above, root nitrate and ammonium uptake is often strongly impaired by 436
waterlogging. Such effects were much more pronounced in flood sensitive compared to 437
more tolerant species (Kreuzwieser et al., 2002). Transcript abundance profiles indicate 438
that many genes involved in amino acid biosynthesis and degradation differed in hypoxia 439
treated trees (Kreuzwieser et al., 2009; Christianson et al., 2010; LeProvost et al., 2012). 440
Consistent with gene expression patterns, many amino acids with increased abundance in 441
hypoxia treated trees were derived from pyruvate (e.g. alanine, valine, leucine) or other 442
intermediates of glycolysis (glycine, serine, tyrosine). In contrast, amino acids formed 443
from TCA cycle components (glutamine, glutamate, aspartate, asparagine) generally 444
showed lower levels in hypoxic than in normoxic roots (Kreuzwieser et al., 2002; 2009; 445
Jaeger et al., 2009). The latter finding suggests that the metabolic flux of organic acids 446
into the TCA cycle is inhibited under hypoxic conditions; thus, lower concentrations of 447
these amino acids seem to be caused by limited availability of carbon compounds from the 448
TCA cycle. 449
450
Kreuzwieser et al. (2002) observed a significant accumulation of γ-aminobutyrate 451
(GABA) and alanine but decreased glutamate levels in roots of waterlogged European 452
beech, Pedunculate oak and Grey poplar. Accumulation of GABA and alanine was 453
strongest in the roots of flooding sensitive beech. This metabolite pattern indicates that the 454
GABA shunt was induced by waterlogging; this assumption is further supported by a 455
strongly elevated transcript abundance of the glutamate decarboxylase gene in 456
waterlogged poplar roots (Kreuzwieser et al., 2009). The GABA shunt is a proton 457
consuming pathway and is thought to contribute to pH stabilisation during periods of 458
oxygen deprivation (Crawford et al., 1994). The metabolite GABA has also been 459
Acc
epte
d A
rticl
e

19 This article is protected by copyright. All rights reserved.
discussed as a signalling compound, likely to contribute to ethylene biosynthesis and, 460
therefore, being involved in morphological adaptations of waterlogged plant (Kreuzwieser 461
et al., 2009). 462
463
It has been hypothesized that nitrate contributes to the maintenance of redox and energy 464
homeostasis of hypoxia treated cells (Igamberdiev and Hill, 2004). The responsible 465
mechanism proposed depends on the contribution of class 1 non-symbiotic haemoglobin, 466
NO and the enzyme nitrate reductase (NR). In several trees oxygen deprivation causes an 467
up-regulation of the gene encoding for non-symbiotic haemoglobin (Parent et al., 2008; 468
Kreuzwieser et al., 2009; Parent et al., 2011; LeProvost et al., 2012). This protein is 469
directly involved in the oxidation of NO thereby forming nitrate. Nitrate can again be 470
reduced by the action of NR to yield NO. The reduced haemoglobin molecule is oxidized 471
by interaction with molecular oxygen; this step also involves oxidation of NADH. The 472
haemoglobin/NO cycle therefore contributes to the maintenance of glycolysis and 473
consequently to ATP production during hypoxia (Igamberdiev et al., 2005). As a 474
signalling compound, NO generated by NR activity has a regulatory function for 475
morphological adaptations to flooding such as the formation of aerenchyma, adventitious 476
roots and hypertrophied lenticels (Parent et al., 2011). Importantly, the higher flooding 477
tolerance of Pedunculate oak (moderately tolerant) compared Sessile oak (flooding 478
sensitive) has been linked to the abundance of the non-symbiotic haemoglobin that was 479
found to be considerably higher in the more flooding tolerant genotype (Parent et al., 480
2011). 481
482
The root-to-shoot transport of metabolites is affected by flooding 483
Metabolite profiling indicates massive changes in the content of soluble carbohydrates, 484
amino acids and organic acids in roots and leaves of waterlogged trees (Kreuzwieser et al., 485
Acc
epte
d A
rticl
e

20 This article is protected by copyright. All rights reserved.
2002; Kreuzwieser et al., 2009; Jaeger et al., 2009). Despite its importance for 486
communication between distant organs, long-distance transport of metabolites from shoot 487
to roots and vice versa has not been studied intensively in waterlogged plants. Hypoxia 488
induced altered metabolite and nutrient levels have been observed in the phloem of 489
Eucalyptus globulus suggesting that the transport between shoot and roots is impaired by 490
waterlogging (Merchant et al., 2010). Figure 5 indicates that hypoxia also considerably 491
affects the transport of metabolites from roots to the shoot. In consistence with elevated 492
levels of GABA, alanine, glycine in the roots of waterlogged poplar (Kreuzwieser et al., 493
2009), these amino acids were present in higher concentrations in the xylem sap of 494
hypoxia stressed trees (Fig. 5; Fig. 6). Similarly, the concentrations of the product of 495
alcoholic fermentation accumulated in roots and the xylem sap (Kreuzwieser et al., 1999). 496
On the other hand, metabolites with lower abundance in waterlogged roots (glutamate, 497
glutamine) tended to show decreased xylem sap concentrations under the same conditions. 498
Similar effects were found in flooding sensitive Fagus sylvatica and Quercus robur 499
seedlings (Kreuzwieser, unpublished data). Future studies should aim to get a more 500
detailed picture on hypoxia caused effects on long-distance transport of metabolites 501
between roots and the shoot. Such changes could contribute to signalling between below- 502
and above-ground plant parts. 503
504
Several studies have demonstrated that waterlogging strongly affects the exchange of trace 505
gases between leaves and the atmosphere. A common phenomenon observed in 506
hypoxically treated trees are the strongly induced emissions of acetaldehyde and ethanol 507
by the leaves (Fig. 6) (Kreuzwieser et al., 1999; Holzinger et al., 2000; Parolin et al., 2004; 508
Rottenberger et al., 2008; Copolovici and Niinemets, 2010; Bracho-Nunez et al., 2012). In 509
Grey poplar, for example, ca. 75% of the ethanol formed in the roots via alcoholic 510
fermentation is transported to the leaves with the transpiration stream (Kreuzwieser et al., 511
Acc
epte
d A
rticl
e

21 This article is protected by copyright. All rights reserved.
1999). Considering the high membrane permeability of ethanol, it is assumed that the 512
xylem loading of ethanol occurs passively by diffusion. In the leaves ethanol is oxidised 513
by ADH thereby forming acetaldehyde which is further converted into acetate 514
(Kreuzwieser et al., 2001). Acetate can enter primary carbon metabolism after activation 515
to acetyl-CoA (Ferner et al., 2012). Small portions of the ethanol delivered to the leaves is 516
released as volatile compounds ethanol, acetaldehyde and acetate into the atmosphere via 517
the stomata (Kreuzwieser et al., 1999; Ferner et al., 2012). The transport of ethanol from 518
roots to the shoot and its conversion to metabolites used in primary carbon metabolism can 519
be seen as a physiological adaptation to waterlogging, since the energy rich carbon 520
skeletons of these compounds can be re-used in the leaves. In addition, an accumulation of 521
phytotoxic acetaldehyde is avoided in the roots. However, a clear correlation of this 522
mechanism to a species’ flooding tolerance has not been observed, as both, highly tolerant 523
and less tolerant species show this ethanol cycling (Kreuzwieser and Rennenberg, 2013). 524
525
Interestingly, not only metabolites linked to fermentative pathways show altered emission 526
due to waterlogging but also several other trace gases (Fig. 6) (Copolovici and Niinemets, 527
2010; Holzinger et al., 2010). These volatiles are typically stress induced like ethylene, 528
NO or wound induced VOC. The latter compounds are products of the lipoxygenase 529
reaction such as hexenal or hexenol (Copolovici and Niinemets, 2010). The emission of 530
most of these compounds correlates with the flooding tolerance of the tree investigated: 531
flooding tolerant trees show lower emission rates than more sensitive species. In addition, 532
the emission of NO correlates with the trees’ flooding tolerance, probably its role during 533
oxidative stress scavenging (Copolovici and Niinemets, 2010). Another compound, which 534
is emitted at higher rates in waterlogged trees, is methanol. This alcohol is formed during 535
cell wall modifications by pectin methylesterases; these enzymes catalyse the 536
demethylation of pectins during leaf expansion and cell wall degradation (Hüve et al., 537
Acc
epte
d A
rticl
e

22 This article is protected by copyright. All rights reserved.
2007; Copolovici and Niinemets, 2010). The increased emission rates of methanol from 538
leaves of waterlogged trees might therefore be a result of stress-induced cell wall 539
degradation in the leaves or a product of aerenchyma and adventitious root formation in 540
the roots. These mechanisms would depend on the transport of methanol from roots to the 541
shoot in the xylem sap of the trees. Future studies should therefore include an analysis of 542
methanol in the xylem sap of waterlogged trees. 543
544
Conclusions and perspectives 545
Despite their great economic, ecological and social significance, the response of trees to 546
waterlogging is far from being understood. This is due to a lack of studies at both, the 547
physiological and the molecular level. So far, there are only three publications on four 548
woody species providing data on changes of the transcriptome of trees in response to 549
waterlogging. In these studies, two flooding tolerant (poplar, Pedunculate oak) and two 550
sensitive species (cotton, Sessile oak) were investigated (Kreuzwieser et al., 2009; 551
Christianson et al., 2010; LeProvost et al., 2012). Even less data are available for 552
waterlogging effects on the metabolome of trees. However, such information is urgently 553
needed for a better understanding of physiological adaptations of woody species to 554
hypoxia. Surprisingly, although the xylem sap of trees can be collected relatively easily, 555
studies on hypoxia effects on the composition of the xylem sap are scarce and metabolite 556
profiling has not been reported. Future studies, investigating the effect of waterlogging 557
stress on trees, should include such approaches in order to elucidate which processes are 558
decisive for flooding tolerance of trees. The few –omics studies performed indicate 559
similarities between herbaceous plants and trees. It seems to be common that hypoxia 560
causes an energy crisis in plants leading to down-regulation of energy consuming 561
processes including shoot and root growth. Apparently, the initiation of fermentative 562
pathways together with enhanced glycolytic flux is of greatest importance for survival of 563
Acc
epte
d A
rticl
e

23 This article is protected by copyright. All rights reserved.
waterlogging periods (Fig. 6). In trees, steady carbohydrate supply for maintenance of 564
glycolysis seems to be crucial and flooding sensitive and tolerant species display large 565
differences in this capability. The reasons for such differences are, however, not 566
understood and should be in the focus of future research. 567
568
569 570
References 571
572
Alaoui-Sossé B., Gérard B., Binet P., Toussaint M.-L. & Badot P.-M. (2005) Influence of 573
flooding on growth, nitrogen availability in soil, and nitrate reduction of young oak 574
seedlings (Quercus robur L.). Annals of Forest Science 62, 593-600. 575
Albrecht G. & Mustroph A. (2003) Localization of sucrose synthase in wheat roots: 576
increased in situ activity of sucrose synthase correlates with cell wall thickening by 577
cellulose deposition under hypoxia. Planta 217, 252–260. 578
Allaway W.G., Curran M., Hollington L.M., Ricketts M.C. & Skelton N.J. (2001) Gas 579
space and oxygen exchange in roots of Avicennia marina (Forssk.) Vierh. var. 580
australasica (Walp.) Moldenke ex N. C. Duke, the Grey Mangrove. Wetlands 581
Ecology and Management 9, 211–218. 582
Armbruster J., Muley-Fritze A., Pfarr U., Rhodius R., Siepmann-Schinker D., Sittler B., 583
Späth V., Trémolières M., Rennenberg H. & Kreuzwieser J. (2006) FOWARA - 584
Forested water retention areas – guideline for decision makers, forest managers and 585
land owners. Albert-Ludwigs-Universität Freiburg. pp. 84. 586
Armstrong W. (1979) Aeration in higher plants. Advances in Botanical Research 7:225-587
332. 588
Aroca R., Porcel R. & Ruis-Lozano J.M. (2011) Regulation of root water uptake under 589
abiotic stress conditions. Journal of Experimental Botany 63: 43–57. 590
Atkinson C.J., Harrison-Murray R.S. & Taylor J.M. (2008) Rapid flood-induced stomatal 591
closure accompanies xylem sap transportation of root-derived acetaldehyde and 592
ethanol in Forsythia. Environmental and Experimental Botany 64, 196–205. 593 Acc
epte
d A
rticl
e

24 This article is protected by copyright. All rights reserved.
Aziz I. & Khan M.A. (2001) Effect of Seawater on the Growth, Ion Content and Water 594
Potential of Rhizophora mucronata Lam. Journal of Plant Research 114, 369-373. 595
Bailey-Serres J. & Voesenek L.A.C.J. (2008) Flooding stress: acclimations and genetic 596
diversity. Annual Reviews of Plant Biology 59, 313–339. 597
Bailey-Serres J., Cho Lee S. & Brinton E. (2012) Waterproofing crops: effective flooding 598
survival strategies. Plant Physiology 160, 1698-1709. 599
Barlow P.W. (1986) Adventitious roots of whole plants: Their forms, functions, and 600
evolution. In New Root Formation in Plants and Cuttings, Jackson M.B. (Ed.). 601
Martinus Nijhoff Publishers, Dordrecht, The Netherlands. 602
Baylis G.T.S. (1950) Root system of the New Zealand mangrove. Transactions of the 603
Royal Society of New Zealand 78: 509–514. 604
Beckman T.G., Perry R.L. & Flore J.A. (1992) Short-term flooding affects gas exchange 605
characteristics of containerized sour cherry trees. Hortscience 27, 1297-1301. 606
Bracho-Nunez A., Knothe N.M., Costa W.R., Liberato M.A.R., Kleiss B., Rottenberger S., 607
Piedade M.T.F. & Kesselmeier J. (2012) Root anoxia effects on physiology and 608
emissions of volatile organic compounds (VOC) under short- and long-term 609
inundation of trees from Amazonian floodplains. SpringerPlus 1:9 doi:10.1186/2193-610
1801-1-9. 611
Calvo-Polanco M., Jorge Señorans J. & Zwiazek J.J. (2012) Role of adventitious roots in 612
water relations of tamarack (Larix laricina) seedlings exposed to flooding. BMC 613
Plant Biology 12:99. http://www.biomedcentral.com/1471-2229/12/99. 614
Carnus J.-M., Parrotta J., Brockerhoff E., Arbez M., Jactel H., Kremer A., Lamb D., 615
O’Hara K.& Walters B. (2006) Planted forests and biodiversity. Journal of Forestry 616
104, 65–77. 617
Christensen J.H., Christensen O.B. (2003) Severe summertime flooding in Europe. 618
Nature, 421, 805 619
Christensen J.H., Hewitson B., Busuioc A., Chen A., Gao X., Held I., Jones R., Kolli 620
R.K., Kwon W.-T., Laprise R., Magaña Rueda V., Mearns L.,. Menéndez C.G, 621
Räisänen J.,. Rinke A., Sarr A. & Whetton P. (2007) Regional Climate Projections. 622
In: Climate Change 2007: The Physical Science Basis. Contribution of Working 623
Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate 624
Acc
epte
d A
rticl
e

25 This article is protected by copyright. All rights reserved.
Change [Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, M. 625
Tignor and H.L. Miller (eds.)]. Cambridge University Press, Cambridge, United 626
Kingdom and New York, NY, USA. 627
Christianson J.A., Llewellyn D.J., Dennis E.S. & Wilson I.W. (2010) Comparisons of 628
early transcriptome responses to low-oxygen environments in three dicotyledonous 629
plant species. Plant Signalling and Behaviour 30, 1006–1009. 630
Christmann A., Grill E. & Huang J. (2013) Hydraulic signals in long-distance signalling. 631
Current Opinion in Plant Biology 16, 293–300. 632
Colin-Belgrand M., Dreyer E. & Biron P. (1990) Sensitivity of seedlings from different 633
oak species to waterlogging: effects on root growth and mineral nutrition. Annals of 634
Forest Science 48, 193–204. 635
Copolovici L. & Niinemets U. (2010) Flooding induced emissions of volatile signalling 636
compounds in three tree species with differing waterlogging tolerance. Plant, Cell 637
and Environment 33, 1582–1594. 638
Crawford L.A., Bown A.W., Breitkreuz K.E. & Guinel F.C. (1994) The synthesis of 639
gamma-aminobutyric acid in response to treatments reducing cytosolic pH. Plant 640
Physiology 104, 865–871. 641
Davies D.D., Grego S. & Kenworthy P. (1974) The control of the production of lactate 642
and ethanol by higher plants. Planta 118, 297–310. 643
de Oliveira V.C. & Joly C.A. (2010) Flooding tolerance of Calophyllum brasiliense 644
Camb. (Clusiaceae): morphological, physiological and growth responses. Trees 24, 645
185–193. 646
Dennis C.W. & Grindley J. (1984) Probability of waterlogging estimated from historical 647
rainfall records. Agricultural Water Management 6, 397–401. 648
Dister E. (1983) Zur Hochwassertoleranz von Auenwaldbäumen an lehmigen Standorten. 649
Verhandlungen der Gesellschaft für Ökologie, Mainz Band 10, 325-336. 650
Drew M.C. (1997) Oxygen deficiency and root metabolism: Injury and acclimation under 651
hypoxia and anoxia. Annual Reviews of Plant Molecular Biology 48, 223-250. 652
Dreyer E. (1994) Compared sensitivity of seedlings from 3 woody species (Quercus robur 653
L., Quercus rubra L. and Fagus sylvatica L.) to waterlogging and associated root 654 Acc
epte
d A
rticl
e

26 This article is protected by copyright. All rights reserved.
hypoxia: effects on water relations and photosynthesis. Annals of Forest Science 51, 655
417-429. 656
Dreyer E., Colin-Belgrand M. & Biron P. (1991) Photosynthesis and shoot water status of 657
seedlings from different oak species submitted to waterlogging. Annals of Forest 658
Science 48, 205-214. 659
Dwyer J.F., Schroeder H.W. & Gobster P.H. (1991) The significance of urban trees and 660
forests: towards a deeper understanding of values. Journal of Arboriculture 17, 276-661
284. 662
Eastmond P.J., Germain V., Lange P.R., Bryce J.H., Smith S.M. & Graham I.A. (2000) 663
Postgerminative growth and lipid catabolism in oilseeds lacking the glyoxylate cycle. 664
Proceedings of the National Academy of Science USA 97, 5669–5674. 665
Else M.A., Taylor J.M. & Atkinson C.J. (2006) Anti-transpirant activity in xylem sap from 666
flooded tomato (Lycopersicon esculetum Mill.) plants is not due to pH-mediated 667
redistributions of root- or shoot-sourced ABA. Journal of Experimental Botany 57, 668
3349–3357. 669
Else M.A., Tiekstra A.E., Croker S.J., Davies W.J. & Jackson M.B. (1996) Stomatal 670
closure in flooded tomato plants involves abscisic acid and a chemical unidentified 671
anti-transpirant in xylem sap. Plant Physiology 112, 239-247. 672
Ernst L., Goodger J.Q.D., Alvarez S., Marsh E.L., Berla B., Lockhart E., Jung J., Li P.H., 673
Bohnert H.J. & Schachtman D.P. (2010) Sulphate as a xylem-borne chemical signal 674
precedes the expression of ABA biosynthetic genes in maize roots. Journal of 675
Experimental Botany 61, 3395-3405. 676
Felle H.H. (2001) pH: signal and messenger in plant cells. Plant Biology 3, 577–591. 677
Fernandez M. D., Pieters A., Donoso C., Herrera C., Tezara W., Rengifo E. & Herrera A. 678
(1999) Seasonal changes in photosynthesis of trees in the flooded forest of the Mapire 679
River. Tree Physiology 19, 79-85. 680
Fernandez M.D. (2006) Changes in photosynthesis and fluorescence in response to 681
flooding in emerged and submerged leaves of Pouteria orinocoensis. Photosynthetica 682
44, 32-38. 683 Acc
epte
d A
rticl
e

27 This article is protected by copyright. All rights reserved.
Ferner E., Rennenberg H. & Kreuzwieser J. (2012) Effect of flooding on C metabolism of 684
flood-tolerant (Quercus robur) and non-tolerant (Fagus sylvatica) tree species. Tree 685
Physiology. 32, 135-145. 686
Ferreira C.S., Piedade M.T.F., Junk W.J. & Parolin P. (2007) Floodplain and upland 687
populations of Amazonian Himatanthus sucuuba: effects of flooding on germination, 688
seedling growth and mortality. Environmental and Experimental Botany 60, 477–483. 689
Ferry B., Morneau F., Bontemps J.D., Blanc L. & Freycon V. (2010) Higher treefall rates 690
on slopes and waterlogged soils result in lower stand biomass and productivity in a 691
tropical rain forest. Journal of Ecology 98, 106–116. 692
Food and Agriculture Organization (FAO) (2012) State of the World’s Forests 2012. 693
Rome. FAO of the United Nations. 60 p. 694
Forster P., Ramaswamy V., Artaxo P., Berntsen T., Betts R., Fahey D.W., Haywood J., 695
Lean J., Lowe D.C., Myhre G., Nganga J., Prinn R., Raga G., Schulz M. & Van 696
Dorland R. (2007) Changes in Atmospheric Constituents and in Radiative Forcing. 697
In: Climate Change 2007: The Physical Science Basis. Contribution of Working 698
Group I to the Fourth Assessment Report of the Intergovernmental Panel on Climate 699
Change [Solomon, S., D. Qin, M. Manning, Z. Chen, M. Marquis, K.B. Averyt, 700
M.Tignor and H.L. Miller (eds.)]. Cambridge University Press, Cambridge, United 701
Kingdom and New York, NY, USA. 702
Frei C. (2004) Eidgenössische Technische Hochschule (Zürich) Institut für Atmosphäre 703
und Klima, and MeteoSchweiz, 2004. Klimazukunft der Schweiz-Eine 704
probabilistische Projektion. MeteoSchweiz, Zürich:8. 705
Frei, C., Regina Schöll R., Fukutome S., Schmidli J. & Vidale P.L. (2006) Future change 706
of precipitation extremes in Europe: Intercomparison of scenarios from regional 707
climate models. Journal of Geophysical Research 111, D06105, 708
doi:10.1029/2005JD005965. 709
Gill C. (1970) The flooding tolerance of woody species – a review. Forestry Abstracts 31, 710
671-688. 711
Glenz C., Schlaepfer R., Iorgulescu I. & Kienast F. (2006) Flooding tolerance of Central 712
European tree and shrub species. Forest Ecology and Management 235, 1-13. 713 Acc
epte
d A
rticl
e

28 This article is protected by copyright. All rights reserved.
Gout E., Boisson A.-M., Aubert S., Douce R. & Bligny, R. (2001) Origin of the 714
cytoplasmic pH changes during anaerobic stress in higher plant cells. Carbon-13 and 715
phosphorous-31 nuclear magnetic resonance studies. Plant Physiology 125, 912–925. 716
Gravatt D.A. & Kirby C.J. (1998) Patterns of photosynthesis and starch allocation in 717
seedlings of four bottomland hardwood tree species subjected to flooding. Tree 718
Physiology 18, 411-417. 719
Guo X-Y, Huang Z-Y, Xu A-C & Zhang X-S. (2011) A comparison of physiological, 720
morphological and growth responses of 13 hybrid poplar clones to flooding. Forestry 721
84, doi:10.1093/forestry/cpq037. 722
Haase K., De Simone O., Junk W.J. & Schmidt W. (2003) Internal oxygen transport in 723
cutting from flood-adapted várzea tree species. Tree Physiology 23, 1069-1076. 724
Haase K. & Rätsch G. (2010) The morphology and anatomy of tree roots and their 725
aeration strategies. In: Junk W.J., Piedade M.T.F., Wittmann F., Schöngart J. & 726
Parolin P. (eds.) Central Amazonian floodplain forests: ecophysiology, biodiversity 727
and sustainable management. Springer, Berlin/Heidelberg/New York. pp. 141-162. 728
Herrera A. (2013). Responses to flooding of plant water relations and leaf gas exchange in 729
tropical tolerant trees of a black-water wetland. Frontiers in Plant Science 4. doi: 730
10.3389/fpls.2013.00106 731
Herschbach C., Mult S., Kreuzwieser J. & Kopriva S. (2005) Influence of anoxia on whole 732
plant sulphur nutrition of flooding tolerant poplar (Populus tremula x P. alba). Plant, 733
Cell and Environment 28, 167-175. 734
Holzinger R., Sandoval-Soto L., Rottenberger S., Crutzen P.J. & Kesselmeier J. (2000) 735
Emissions of volatile organic compounds from Quercus ilex L. measured by Proton 736
Transfer Reaction Mass Spectrometry under different environmental conditions. 737
Journal of Geophysical Research 105, 20573–20579. 738
Hultine K.R., Bush S.E., West A.G. & Ehleringer J.R.. (2007) Population structure, 739
physiology and ecohydrological impacts of dioecious riparian tree species of western 740
North America. Oecologia 154:85–93. 741
Hüve K., Christ M.M., Kleist E., Uerlings R., Niinemets Ü., Walter A. & Wildt J. (2007) 742
Simultaneous growth and emission measurements demonstrate an interactive control 743 Acc
epte
d A
rticl
e

29 This article is protected by copyright. All rights reserved.
of methanol release by leaf expansion and stomata. Journal of Experimental Botany 744
58, 1783–1793. 745
Igamberdiev A.U. & Hill R.D. (2004) Nitrate, NO and haemoglobin in plant adaptation to 746
hypoxia: an alternative to classic fermentation pathways. Journal of Experimental 747
Botany 55, 2473-2482. 748
Igamberdiev A.U., Baron K., Manac’h-Little N., Stoimenova M. & Hill R.D. (2005) The 749
haemoglobin/nitric oxide cycle: involvement in flooding stress and effects on 750
hormone signaling. Annals of Botany 96, 557-564. 751
Iglesias D.J., Lliso I., Tadeo F.R. & Talón M. (2002) Regulation of photosynthesis 752
through source: sink imbalance in citrus is mediated by carbohydrate content in 753
leaves. Physiologia Plantarum 116, 563–572. 754
Islam M.A. & MacDonald S.E. (2004) Ecophysiological adaptations of black spruce 755
(Picea marinana) and tamarack (Larix laricina) seedlings to flood. Trees 18, 35-42. 756
Islam M.A., MacDonald S.E. & Zwiazek J.J. (2003) Responses of black spruce (Picea 757
mariana) and tamarack (Larix laricina) to flooding and ethylene. Tree Physiology 23, 758
545–552. 759
Jackson M.B. & Colmer T.D. (2005) Response and Adaptation by Plants to Flooding 760
Stress. Annals of Botany 96, 501–505. 761
Jackson M.B. (1982) Ethylene as a growth promoting hormone under flooded conditions. 762
In: Wareing PF, ed. Plant growth regulators. London: Academic Press, 291–301. 763
Jaeger C., Gessler A., Biller S., Rennenberg H. & Kreuzwieser J. (2009) Differences in C 764
metabolism of ash species and provenances as a consequence of root oxygen 765
deprivation by waterlogging. Journal of Experimental Botany 60: 4335-4345. 766
Joly C.A. & Crawford R.M.M. (1982) Variation in tolerance and metabolic responses to 767
flooding in some tropical trees. Journal of Experimental Botany 33, 799-809. 768
Kludze H.K., Pezeshki S.R. & DeLaune R.D. (1994) Evaluation of root oxygenation and 769
growth in baldcypress in response to short-term soil hypoxia. Canadian Journal of 770
Forest Research 24, 804–809. 771
Kolb R.M., Rawyler A. & Braendle R. (2002) Parameters affecting the early seedling 772
development of four neotropical trees under oxygen deprivation stress. Annals of 773
Botany 89, 551-558. 774
Acc
epte
d A
rticl
e

30 This article is protected by copyright. All rights reserved.
Kozlowski T.T. (1984a) Extent, causes and impacts of flooding. In Flooding and plant 775
growth. Kozlowski T.T., ed., Academic Press, Orlando, FL, pp 1-7. 776
Kozlowski T.T. (1984b) Plant responses to flooding of soil. Bio Science 34, 162–167. 777
Kozlowski T.T. (1997) Responses of woody plants to flooding and salinity. Tree 778
Physiology Monograph No.1. 29 pages. 779
Kramer K., Vreugdenhil S.J. & van der Werf D.C. (2008) Effects of flooding on the 780
recruitment, damage and mortality of riparian tree species: A field and simulation 781
study on the Rhine floodplain. Forest Ecology and Management 255, 3893–3903. 782
Kreuzwieser J. & Gessler A. (2010) Global climate change and tree nutrition: influence of 783
water availability. Tree Physiology 30, 1221-1234. 784
Kreuzwieser J. & Rennenberg H. (2013) Flooding-driven emissions from trees. In: 785
Biology, controls and models of tree volatile organic compound emissions 786
(Niinemets Ü. & Monson R.K., eds.) Springer series Tree Physiology volume 5, pp. 787
237-252. 788
Kreuzwieser J., Fürniss S. & Rennenberg H. (2002) The effects of flooding on the N 789
metabolism of flood tolerant and sensitive tree species. Plant, Cell and Environment 790
25, 1039-1050. 791
Kreuzwieser J., Harren F.J.M., Laarhoven L.J.J., Boamfa I., te Lintel-Hekkert S., 792
Scheerer U., Hüglin C. & Rennenberg H. (2001) Acetaldehyde emission by the 793
leaves of trees - correlation with physiological and environmental parameters. 794
Physiologia Plantarum 113, 41–49. 795
Kreuzwieser J., Hauberg J., Howell K.A., Carroll A., Rennenberg H., Millar A.H. & 796
Whelan J. (2009) Differential response of Grey poplar leaves and roots underpins 797
stress adaptation during hypoxia. Plant Physiology, 149, 461-473. 798
Kreuzwieser J., Scheerer U. & Rennenberg H. (1999) Metabolic origin of acetaldehyde 799
emitted by poplar (Populus tremula x P. alba) trees. Journal of Experimental Botany 800
50, 757-765. 801
Kundzewicz Z.W. (2005) Climate change and floods. WMO Bulletin 55, 170-173. 802
Kundzewicz Z.W., Ulbrich U., Brücher T., Gracyk D., Krüger A., Leckebusch G., Menzel 803
L., Pinskwar I., Radziejewski M. & Szwed M. (2005) Summer floods in central 804
Europe – climate change track? Natural Hazards 36, 165-189. 805
Acc
epte
d A
rticl
e

31 This article is protected by copyright. All rights reserved.
Laur J. & Hacke U.G. (2013) Transpirational demand affects aquaporin expression in 806
poplar roots. Journal of Experimental Botany 64, 2283-2293. 807
LeProvost G., Sulmon C., Frigerio J.M., Bodénès C., Kremer A. & Plomion C. (2012) 808
Role of waterlogging-responsive genes in shaping interspecific differentiation 809
between two sympatric oak species. Tree Physiology 32, 119-134. 810
Li S., Pezeshki S.R. & Shields F.D. (2006) Partial flooding enhances aeration in 811
adventitious roots of black willow (Salix nigra) cuttings. Journal of Plant Physiology 812
163, 619-628. 813
Lucassen E.C.H.E.T., Bobbink R., Smolders A.J.P., van der Ven P.J.M., Lamers L.P.M. & 814
Roelofs J.G.M. (2002) Interactive effects of low pH and high ammonium levels 815
responsible for the decline of Cirsium dissectum (L.) Hill. Plant Ecology 165, 45 – 52. 816
Lucassen E.C.H.E.T., Smolders A.J.P. & Roelofs J.G.M. (2000) Increased ground-water 817
levels cause iron toxicity in Glyceria fluitans (L.). Aquatic Botany 66, 321 – 327. 818
Martínez-Alcántara1 B., Jover S., Quinones A., Forner-Giner M.A., Rodríguez-Gamir J., 819
Legaz F., Primo-Millo E. & Iglesias D.J. (2012) Flooding affects uptake and 820
distribution of carbon and nitrogen in citrus seedlings. Journal of Plant Physiology 821
169, 1150-1157. 822
McClean J. (2000) Wetter is not always better - Flood Tolerance of Woody Species. 823
Technical Note #52 from Watershed Protection Techniques. 1, 208-210. 824
McKee W.H. & McKevlin M.R. (1993) Geochemical processes and nutrient uptake by 825
plants in hydric soils. Environmental Toxicology and Chemistry 12, 2197–2207. 826
Merchant A., Peuke A.D., Keitel C., Macfarlane C., Warren C.R. & Adams M.A. (2010) 827
Phloem sap and leaf δ13C, carbohydrates, and amino acid concentrations in 828
Eucalyptus globulus change systematically according to flooding and water deficit 829
treatment. Journal of Experimental Botany 61, 1785-1793. 830
Mielke M.S., De Almeida A.A.F., Gomes F.P., Mangabeira P.A.O. & Silva D.D.C. (2005) 831
Effects of soil flooding on leaf gas exchange and growth of two neotropical pioneer 832
tree species. New Forests 29, 161–168. 833
Mustroph A., Lee S.C., Oosumi T., Zanetti M.E., Yang H., Ma K., Yaghoubi-Masihi A., 834
Fukao T. & Bailey-Serres J. (2010) Cross-kingdom comparison of transcriptomic 835 Acc
epte
d A
rticl
e

32 This article is protected by copyright. All rights reserved.
adjustments to low-oxygen stress highlights conserved and plant-specific responses. 836
Plant Physiology 152: 1484–1500. 837
Narsai R., Rocha M., Peter Geigenberger P., James Whelan J. & van Dongen J.T. (2011) 838
Comparative analysis between plant species of transcriptional and metabolic 839
responses to hypoxia. New Phytologist 190, 472-487. 840
Nealle D.B. & Kremer A. (2011) Forest tree genomics: growing resources and 841
applications. Nature Reviews Genetics 12, 111-122. 842
Neatrour M.A., Jones R.H. & Golladay S.W. (2007) Response of three floodplain tree 843
species to spatial heterogeneity in soil oxygen and nutrients. Journal of Ecology 95, 844
1274–1283. 845
Nicolás E., Torrecillas A., Dell’Amico J., Alarcón J. (2005) The effect of short-term 846
flooding on the sap flow, gas exchange and hydraulic conductivity of young apricot 847
trees. Trees-Structure and Function 19, 51–57. 848
Nielsen J.L., Rood S.B., Pearce D.W., Letts M.G. & Jiskoot H. (2010) Streamside trees: 849
responses of male, female and hybrid cottonwoods to flooding. Tree Physiology 30, 850
1479-1488. 851
Niinemets Ü. & Valladares F. (2006) Tolerance to shade, drought, and waterlogging of 852
temperate northern hemisphere trees and shrubs. Ecological Monographs 76, 521-853
547. 854
Nunez-Elisea R., Schaffer B., Fisher J.B., Colls A.M. & Crane J.H. (1999) Influence of 855
flooding on net CO2 assimilation, growth and stem anatomy of Annona species. 856
Annals of Botany 84, 771-80. 857
Ojeda M., Schaffer B. & Davies F.S. (2004) Flooding, root temperature, physiology and 858
growth of two Annona species. Tree Physiology 24, 1019 –1025. 859
Ortuño M.F., Alarcón J.J., Nicolás E. & Torrecillas A. (2007) Water status indicators of 860
lemon trees in response to flooding and recovery. Biologia Plantarum 51, 292-296. 861
Pagnussat G.C., Simontacchi M., Puntarulo S. & Lamattina L. (2002) Nitric oxide is 862
required for root organogenesis. Plant Physiology 129, 954–956. 863
Pagnussat G.C., Lanteri M.L., Lombardo M.C. & Lamattina L. (2004) Nitric oxide 864
mediates the indole acetic acid induction activation of a mitogen-activated protein 865 Acc
epte
d A
rticl
e

33 This article is protected by copyright. All rights reserved.
kinase cascade involved in adventitious root development. Plant Physiology 135, 279-866
286. 867
Palmer T.N. & Räisänen J. (2002) Quantifying the risk of extreme seasonal precipitation 868
events in a changing climate. Nature 415, 512-514. 869
Parelle J., Brendel O., Bodenes C., Berveiller D., Dizengremel P., Jolivet Y. & Dreyer E. 870
(2006) Differences in morphological and physiological responses to water-logging 871
between two sympatric oak species (Quercus petraea [Matt.] Liebl., Quercus robur 872
L.). Annals of Forest Science 63, 849–859. 873
Parent C., Berger A., Folzer H., Dat J.F., Crèvecoeur M., Badot P.-M. & Capelli N. (2008) 874
A novel nonsymbiotic hemoglobin from oak: cellular and tissue specificity of gene 875
expression. New Phytologist 177, 142-154. 876
Parent C., Crèvecoeur M., Capelli N. & Dat J.F. (2011) Contrasting growth and adaptive 877
responses of two oak species to flooding stress: role of non-symbiotic haemoglobin. 878
Plant, Cell and Environment 34, 1113-1126. 879
Parent C., Crèvecoeur M., Capelli N. & Dat J.F. (2011) Contrasting growth and adaptive 880
responses of two oak species to flooding stress: role of non-symbiotic haemoglobin. 881
Plant, Cell and Environment 34, 1113–1126. 882
Parolin P. & Wittmann F. (2010) Struggle in the flood: tree responses to flooding stress in 883
four tropical floodplain systems. AoB Plants 2010: doi: 10.1093/aobpla/plq003. 884
Parolin P. (2001) Morphological and physiological adjustments to waterlogging and 885
drought in seedlings of Amazonian floodplain trees. Oecologia 128, 326–335. 886
Parolin P., De Simone O., Haase K., Waldhoff D., Rottenberger S., Kuhn U., Kesselmeier 887
J., Kleiss B., Schmidt W., Piedade M.T.F. & Junk W.J. (2004) Central Amazonian 888
Floodplain Forest: Tree Adaptations in a Pulsing System. The Botanical Review 70, 889
357–380. 890
Parolin P., Lucas C., Piedade M. T. F. & Wittmann F. (2010) Drought responses of 891
extremely flood-tolerant trees of Amazonian floodplains. Annals of Botany 105, 129–892
139. 893
Pezeshki S.R. & Chambers J.L. (1985a) Stomatal and photosynthetic response of sweet 894
gum (Liquidambar styraciflua) to flooding. Canadian Journal of Forest Research 15, 895
371--375. 896
Acc
epte
d A
rticl
e

34 This article is protected by copyright. All rights reserved.
Pezeshki S.R. & Chambers J.L. (1985b) Responses of cherry bark oak (Quercus falcata 897
var. pagodaefolia) seedlings to short-term flooding. Forest Sciences 31, 760--771. 898
Pezeshki S.R. (1994) Responses of baldcypress (Taxodium distichum) seedlings to 899
hypoxia: Leaf protein content, ribulose-1,5-bisphosphate carboxylase/oxygenase 900
activity and photosynthesis. Photosynthetica 30, 59-68. 901
Pezeshki S.R., & Chambers J.C. (1986) Variation in flood-induced stomatal and 902
photosynthetic responses of three bottomland tree species. Forest Sciences 32, 914-903
923. 904
Pezeshki S.R., Pardue J.H. & DeLaune R.D. (1996) Leaf gas exchange and growth of 905
flood-tolerant and flood-sensitive tree species under low soil redox conditions. Tree 906
Physiology 16, 452-458. 907
Pezeshki, S.R., Pardue, J.H. & DeLaune, R.D. (1996) Leaf gas exchange and growth of 908
flood-tolerant and flood-sensitive tree species under low soil redox conditions. Tree 909
Physiology 16, 453-458. 910
Ponnamperuma F.N. (1972) The chemistry of submerged soils. Advances in Agronomy 24, 911
29-95. 912
Ponnamperuma F.N. (1984) Effects of flooding on soils. Chapter 2. In Flooding and plant 913
growth. (Kozlowski, T.T., ed.), Orlando: Academic Press, pp. 9-193. 914
Porth I., Koch M., Berenyi M., Burg A. & Burg K. (2005) Identification of adaptation-915
specific differences in mRNA expression of sessile and pedunculate oak based on 916
osmotic-stress-induced genes. Tree Physiology 25, 1317–1329. 917
Pradet A. & Bomsel J.L. (1978) Energy metabolism in plants under hypoxia and anoxia. 918
In Plant life in anaerobic environments. Hook D.D. & Crawford R.M.M., eds. Ann 919
Arbor Science Publishers Inc., Ann Arbor, pp. 89-118. 920
Reece C.F. & Riha S.J. (1991) Role of root systems of Eastern larch and White spruce in 921
response to flooding. Plant, Cell and Environment 14, 229-234. 922
Rengifo E., Tezara W. & Herrera A. (2005) Water relations, chlorophyll a fluorescence 923
and carbohydrate contents in trees of a tropical forest in response to flood. 924
Photosynthetica 43, 203-210. 925 Acc
epte
d A
rticl
e

35 This article is protected by copyright. All rights reserved.
Roberts J.K.M., Callis J., Wemmer D., Walbot V. & Jardetzky O. (1984) Mechanism of 926
cytoplasmic pH regulation in hypoxic maize root tips and its role in survival under 927
hypoxia. Proceeding of the National Academy of Science USA 81, 3379–3383. 928
Rodríguez-Gamir J., Ancillo G., González-Mas M.C., Primo-Millo E., Iglesias D.J. & 929
Forner-Giner M.A. (2011) Root signalling and modulation of stomatal closure in 930
flooded citrus seedlings. Plant Physiology and Biochemistry 49, 636–645. 931
Ruiz-Sánchez M.C., Domingo R., Morales D. & Torrecillas A. (1996).Water relations of 932
Fino lemon plants on two rootstocks under flooded conditions. Plant Sciences 120, 933
119–125. 934
Schaffrath J. (2000) Auswirkungen des extremen Sommerhochwassers des Jahres 1997 935
auf die Gehölzvegetation in der Oderaue bei Frankfurt (O). Naturschutz und 936
Landschaftspflege in Brandenburg 9, 4-13. 937
Schmull M. & Thomas F.M. (2000) Morphological and physiological reactions of young 938
deciduous trees (Quercus robur L., Q. petraea [Matt.] Liebl., Fagus sylvatica L.) to 939
waterlogging. Plant and Soil 225, 227-242. 940
Shimamura S., Yamamoto R., Nakamura T., Shimada S. & Komatsu S. (2010) Stem 941
hypertrophic lenticels and secondary aerenchyma enable oxygen transport to roots of 942
soybean in flooded soil. Annals of Botany 106, 277-284. 943
Siebel H.N. & Blom C.W.P.M. (1998) Effects of irregular flooding on the establishment 944
of tree species. Acta Botanica Neerlandica 47, 231–240. 945
Snowden R.E. &Wheeler B.D. (1993) Iron toxicity to fen plant species. Journal of 946
Ecology 81, 35 – 46. 947
Späth V. (1988) Zur Hochwassertoleranz von Auenwaldbäumen. Natur und Landschaft 948
H.7/8, S.312-315. 949
Späth V. (2002) Hochwassertoleranz von Waldbäumen in der Rheinaue. Allgemeine 950
Forstzeitschrift/ Der Wald 15, 807-810. 951
Steffens B., Wang J. & Sauter M. (2006) Interactions between ethylene, gibberelin and 952
abscisic acid regulate emergence and growth rate of adventitious roots in deepwater 953
rice. Planta 223, 604-612. 954 Acc
epte
d A
rticl
e

36 This article is protected by copyright. All rights reserved.
Steffens B., Steffen-Heins A. & Sauter M. (2013) Reactive oxygen species mediate 955
growth and death in submerged plants. Frontiers in Plant Science 4, doi: 956
10.3389/fpls.2013.00179. 957
Striker G.G. (2012) Flooding stress on plants: anatomical, morphological and 958
physiological responses. Mworia J. (ed.), ISBN 978-953-51-0355-4. 959
Syvertsen J.P., Zablotowicz R.M. & Smith Jr M.L. (1983) Soil temperature and flooding 960
effects on two species of citrus. I. Plant growth and hydraulic conductivity. Plant 961
and Soil 72:3–12. 962
The Columbia Electronic Encyclopedia, 6th ed. Copyright © 2012, Columbia University 963
Press. http://www.infoplease.com/encyclopedia/science/forest-the-importance-964
forests.html#ixzz2hryOK2Ua 965
Tournaire-Roux C., Sutka M., Javot H., Gout E., Gerbeau P., Luu D.T., Bligny R. & 966
Maurel C. (2003) Cytosolic pH regulates root water transport during anoxic stress 967
through gating of aquaporins. Nature 425, 393–397. 968
Tuskan G.A., DiFazio S. & Teichmann T. (2003) Poplar genomics is getting popular: The 969
impact of the poplar genome project on tree research. Plant Biology 5, 1-3. 970
Tuskan G.A., DiFazio S. Jansson S., Bohlmann J., Grigoriev I., Hellsten U., ... Rokhsar D. 971
(2006). The genome of black cottonwood, Populus trichocarpa (Torr. & Gray). 972
Science 313, 1596–1604. 973
Unger I.M., Kennedy A.C. & Muzika R.-M. (2009) Flooding effects on soil microbial 974
communities. Applied Soil Ecology 42, 1–8. 975
Visser E., Nabben R., Blom C. & Voesenek L. (1997) Elongation by primary lateral roots 976
and adventitious roots during conditions of hypoxia and high ethylene 977
concentrations. Plant, Cell and Environment 20, 647-653. 978
Vreugdenhil S.J., Kramer K. & Pelsma T. (2006) Effects of flooding duration, -frequency 979
and -depth on the presence of saplings of six woody species in north-west Europe. 980
Forest Ecology and Management 236, 47-55. 981
Vu J. & Yelenosky G. (1991) Photosynthetic responses of citrus trees to soil flooding. 982
Physiologia Plantarum 81, 7–14. 983
Vu J.C.V. & Yelenoski G. (2006) Photosynthetic response of citrus trees to soil flooding. 984
Physiologia Plantarum 81, 7-14. 985
Acc
epte
d A
rticl
e

37 This article is protected by copyright. All rights reserved.
Yamanoshita T., Masumori M., Yagi H. & Kojima K. (2005) Effects of flooding on 986
downstream processes of glycolysis and fermentation in roots of Melaleuca cajuputi 987
seedlings. Journal of Forest Research 10, 199-204. 988
Ye Y., Tam N.F.Y., Wong Y.S. & Lu C.Y. (2003) Growth and physiological responses of 989
two mangrove species (Bruguiera gmynorrhiza and Kandelia candel) to 990
waterlogging. Environmental and Experimental Botany 49, 209-221. 991
992
993
Acc
epte
d A
rticl
e

38 This article is protected by copyright. All rights reserved.
994 Figure legends 995
Fig. 1: dventitious roots of different tree species as adaptive responses to flooding. A, 996
the mangrove species Rhizophora; B, close-up of Rhizophora prop roots; C, 997
pneumatophores of Avicennia; D, adventitious roots of Grey poplar (P. x canescens). 998
999
Fig. 2: Aquaporin gene expression in poplar roots as affected by waterlogging. Leafs of P. 1000
x canescens trees were harvested after 168 hours, roots after 5, 24 and 168 hours of 1001
waterlogging and transcript abundances were determined by microarray analysis. Shown 1002
are the log2 values of fold changes of flooded compared to control trees by using the 1003
colour code indicated. Relative abundance indicates the raw signal value of normoxic 1004
controls on the microarray. Data are from Kreuzwieser et al. (2009). 1005
1006
Fig. 3: Processes assumed to be involved in the reduction of hydraulic conductance of 1007
flooded roots. A change from normoxia to flooding induced hypoxia or anoxia causes 1008
several adjustments in root metabolism (Kreuzwieser et al., 2009). Cytosolic pH decrease 1009
may result from lactic acid fermentation, hydrolysis of nucleoside triphosphates (NTP), 1010
proton influx from vacuoles or external medium, and the biosynthesis of organic acids 1011
other than lactic acid (Gout et al., 2001). Subsequent protonation of PIPs reduces the water 1012
absorption by roots, thereby decreasing the root water potential (Tournaire et al., 2003). 1013
Root-to-shoot signals of unknown nature (hydraulic signal or chemical signal) lead to the 1014
closure of the stomata. 1015
1016
Fig. 4. Effects of waterlogging on whole plant carbon cycling of flood tolerant and flood 1017
sensitive tree species. Net photosynthesis and soluble carbohydrate contents in leaves, 1018
roots, phloem and xylem sap were determined in plants waterlogged for 7 days and in non-1019 Acc
epte
d A
rticl
e

39 This article is protected by copyright. All rights reserved.
waterlogged control plants (data from Ferner et al., 2012). Starch content of leaves and 1020
roots were not affected by waterlogging (Ferner et al., 2012) (data not shown;). Blue / red 1021
/green colours indicate values significantly higher / lower / unaffected compared to 1022
controls; the numbers give percent differences relative to controls. Grey arrows and areas 1023
indicate the 100 % levels of control plants. 1024
1025
Fig. 5. Waterlogging induced changes in the xylem sap composition of poplar trees. 1026
Xylem sap of 3 months old Populus x canescens trees normally watered or waterlogged 1027
for 2 days was collected and analysed for metabolites via GC/MS (Kreuzwieser et al., 1028
2009). Log2 of fold changes (FC) is displayed by the colour code shown. P-values of a 1029
Student’s t’test are indicated (n=4). 1030
1031
Fig. 6. Simplified scheme of the temporal response of trees to waterlogging. Processes and 1032
metabolites indicated in blue are usually up-regulated and increased in abundance, 1033
respectively, due to the stress. Red colour indicated processes usually down-regulated or 1034
metabolites with reduced abundance. 1035
1036
Acc
epte
d A
rticl
e
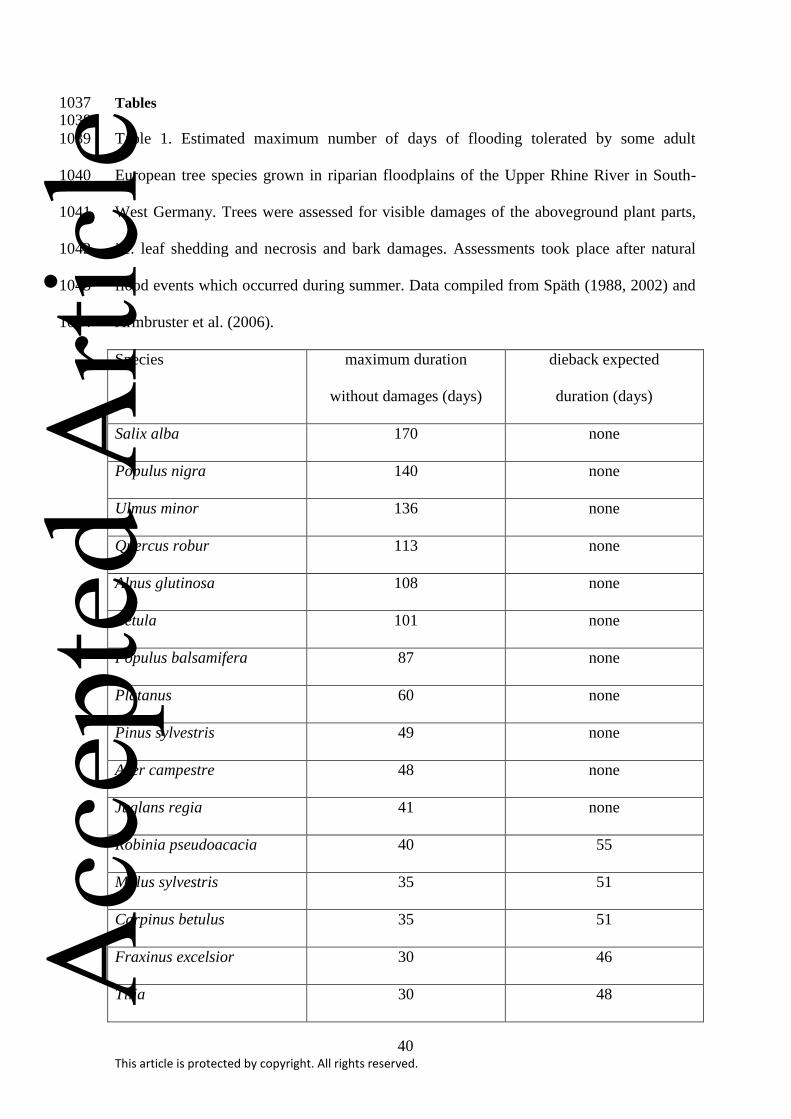
40 This article is protected by copyright. All rights reserved.
Tables 1037 1038
Table 1. Estimated maximum number of days of flooding tolerated by some adult 1039
European tree species grown in riparian floodplains of the Upper Rhine River in South-1040
West Germany. Trees were assessed for visible damages of the aboveground plant parts, 1041
i.e. leaf shedding and necrosis and bark damages. Assessments took place after natural 1042
flood events which occurred during summer. Data compiled from Späth (1988, 2002) and 1043
Armbruster et al. (2006). 1044
Species maximum duration
without damages (days)
dieback expected
duration (days)
Salix alba 170 none
Populus nigra 140 none
Ulmus minor 136 none
Quercus robur 113 none
Alnus glutinosa 108 none
Betula 101 none
Populus balsamifera 87 none
Platanus 60 none
Pinus sylvestris 49 none
Acer campestre 48 none
Juglans regia 41 none
Robinia pseudoacacia 40 55
Malus sylvestris 35 51
Carpinus betulus 35 51
Fraxinus excelsior 30 46
Tilia 30 48 Acc
epte
d A
rticl
e

41 This article is protected by copyright. All rights reserved.
Acer pseudoplatanus <30 43
Acer platanoides <12 43
Fagus sylvatica 9 37
Prunus avium 10 12
1045
1046
Acc
epte
d A
rticl
e

42 This article is protected by copyright. All rights reserved.
1047
Table 2: Some examples of waterlogging tolerances of field grown European tree species 1048
of different age. Trees were grown under natural conditions in riparian forests; they were 1049
assessed for stress symptoms of the aboveground parts (leaf necrosis, leaf shedding, bark 1050
damages) after flood events during the vegetation period. Data compiled from Dister 1051
(1983), Späth (1988, 2002), Schaffrath (2000), Armbruster et al. (2006). 1052
Species Maximal waterlogging period without stress symptoms
[number of days]
Seedling Young stand Adult tree
Quercus robur
Fraxinus excelsior
30-46
14-35
25
48
113-150
30-72
Acer pseudoplatanus 16-20 17-46 17-30
Fagus sylvatica 9 16 9-35
1053
Acc
epte
d A
rticl
e

pce_12310_f1.tif
Acc
epte
d A
rticl
e
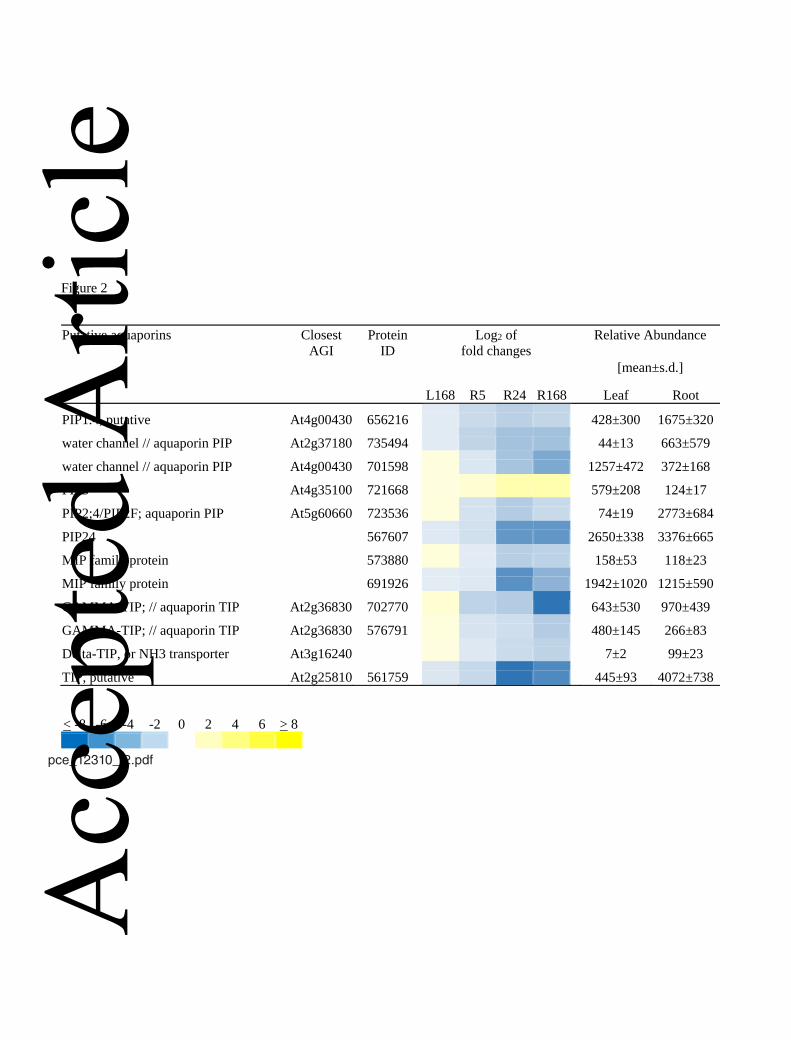
Figure 2
Putative aquaporins Closest AGI
ProteinID
Log2 of fold changes
Relative Abundance
[mean±s.d.]
L168 R5 R24 R168 Leaf Root
PIP1.4, putative At4g00430 656216 428±300 1675±320
water channel // aquaporin PIP At2g37180 735494 44±13 663±579
water channel // aquaporin PIP At4g00430 701598 1257±472 372±168
PIP3 At4g35100 721668 579±208 124±17
PIP2;4/PIP2F; aquaporin PIP At5g60660 723536 74±19 2773±684
PIP24 567607 2650±338 3376±665
MIP family protein 573880 158±53 118±23
MIP family protein 691926 1942±1020 1215±590
GAMMA-TIP; // aquaporin TIP At2g36830 702770 643±530 970±439
GAMMA-TIP; // aquaporin TIP At2g36830 576791 480±145 266±83
Delta-TIP, or NH3 transporter At3g16240 7±2 99±23
TIP, putative At2g25810 561759 445±93 4072±738
< -8 -6 -4 -2 0 2 4 6 > 8
pce_12310_f2.pdf
Acc
epte
d A
rticl
e
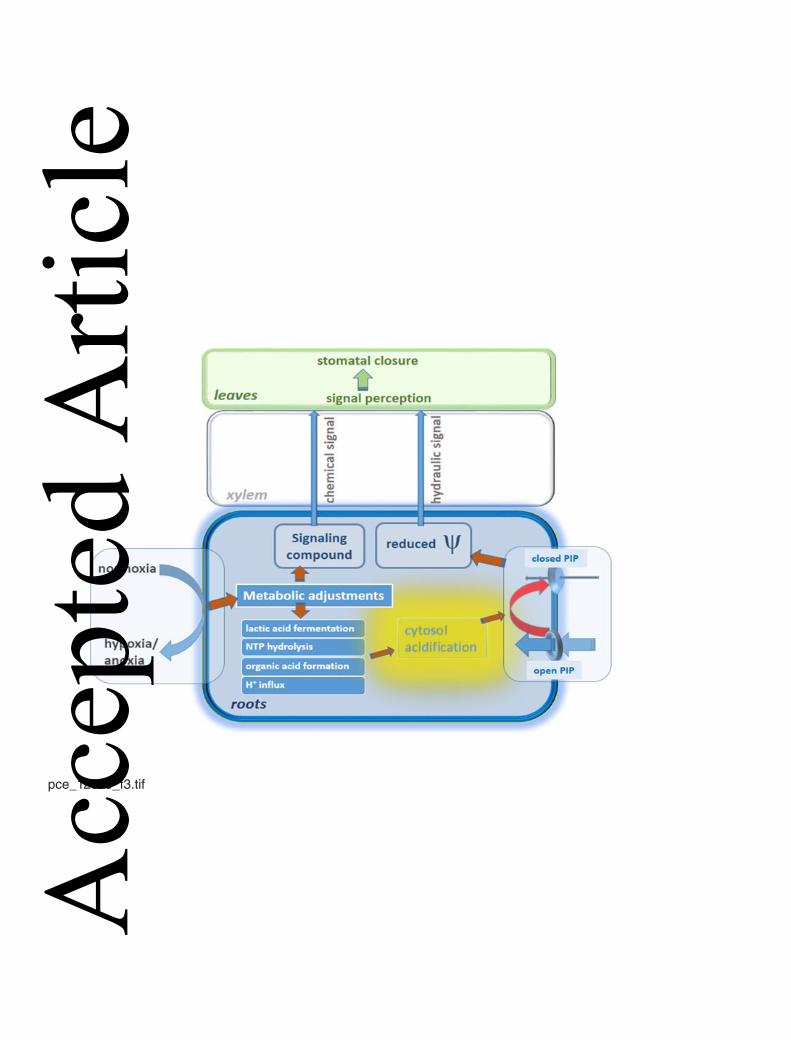
pce_12310_f3.tif
Acc
epte
d A
rticl
e
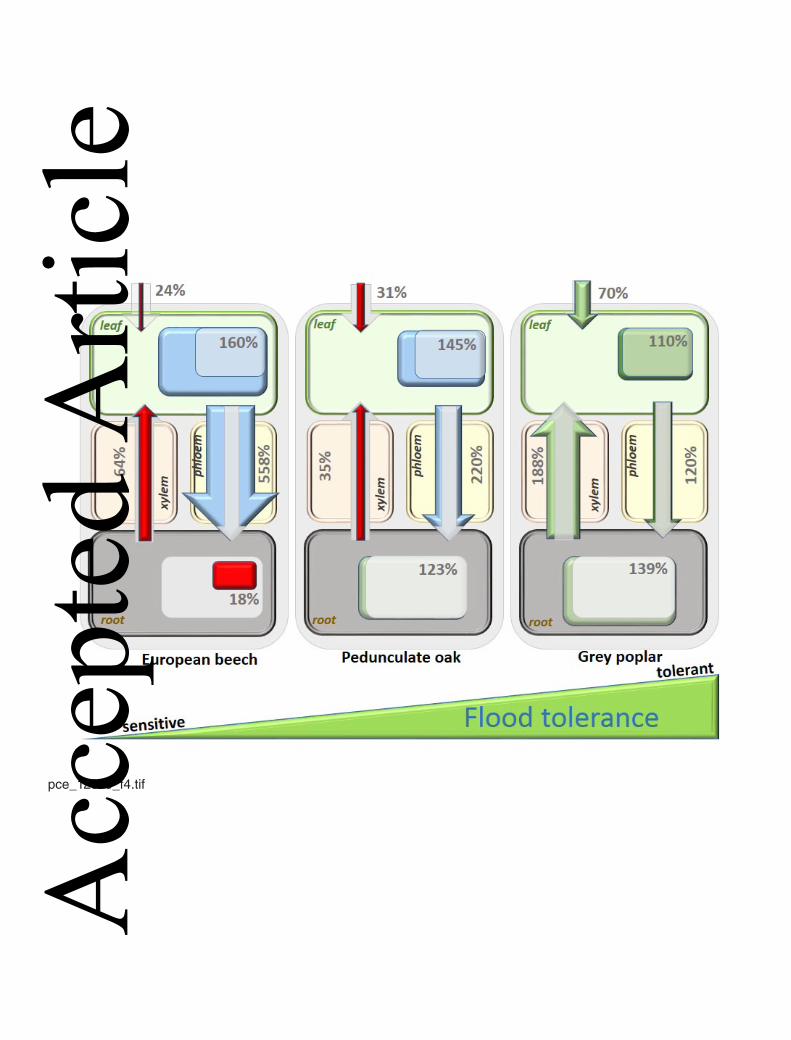
pce_12310_f4.tif
Acc
epte
d A
rticl
e
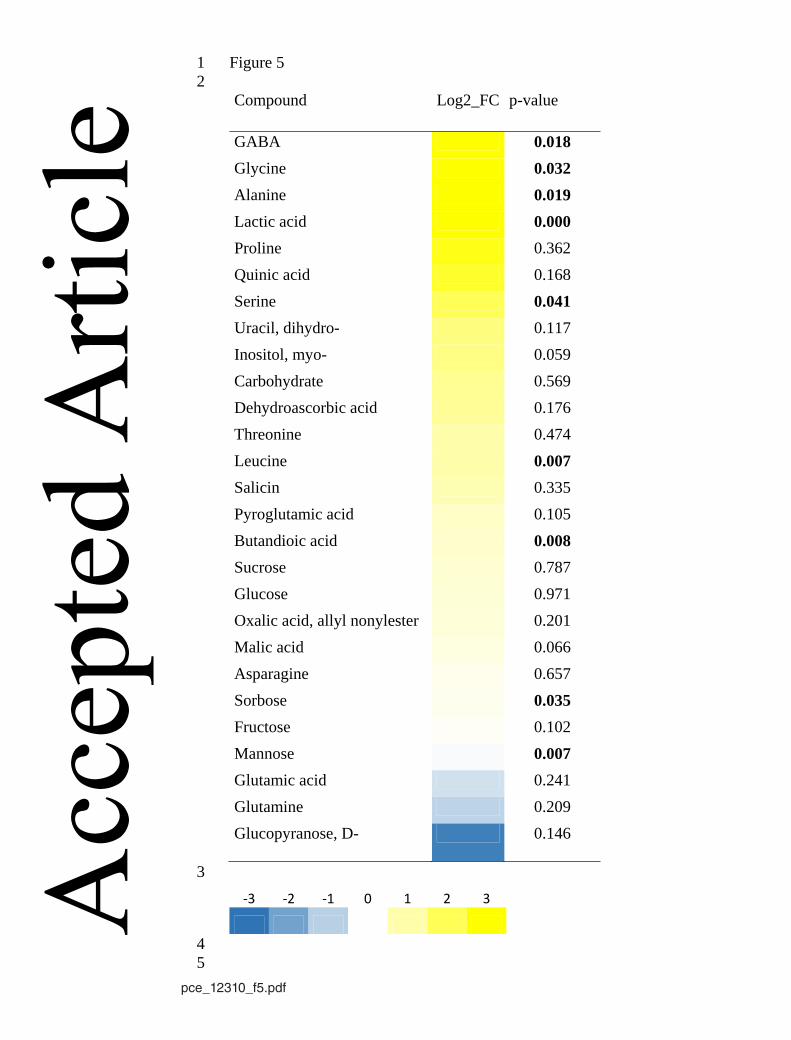
Figure 5 12
Compound Log2_FC p-value GABA 0.018Glycine 0.032Alanine 0.019Lactic acid 0.000Proline 0.362 Quinic acid 0.168 Serine 0.041Uracil, dihydro- 0.117 Inositol, myo- 0.059 Carbohydrate 0.569 Dehydroascorbic acid 0.176 Threonine 0.474 Leucine 0.007Salicin 0.335 Pyroglutamic acid 0.105 Butandioic acid 0.008Sucrose 0.787 Glucose 0.971 Oxalic acid, allyl nonylester 0.201 Malic acid 0.066 Asparagine 0.657 Sorbose 0.035Fructose 0.102 Mannose 0.007Glutamic acid 0.241 Glutamine 0.209 Glucopyranose, D- 0.146
3‐3 ‐2 ‐1 0 1 2 3
45
pce_12310_f5.pdf
Acc
epte
d A
rticl
e
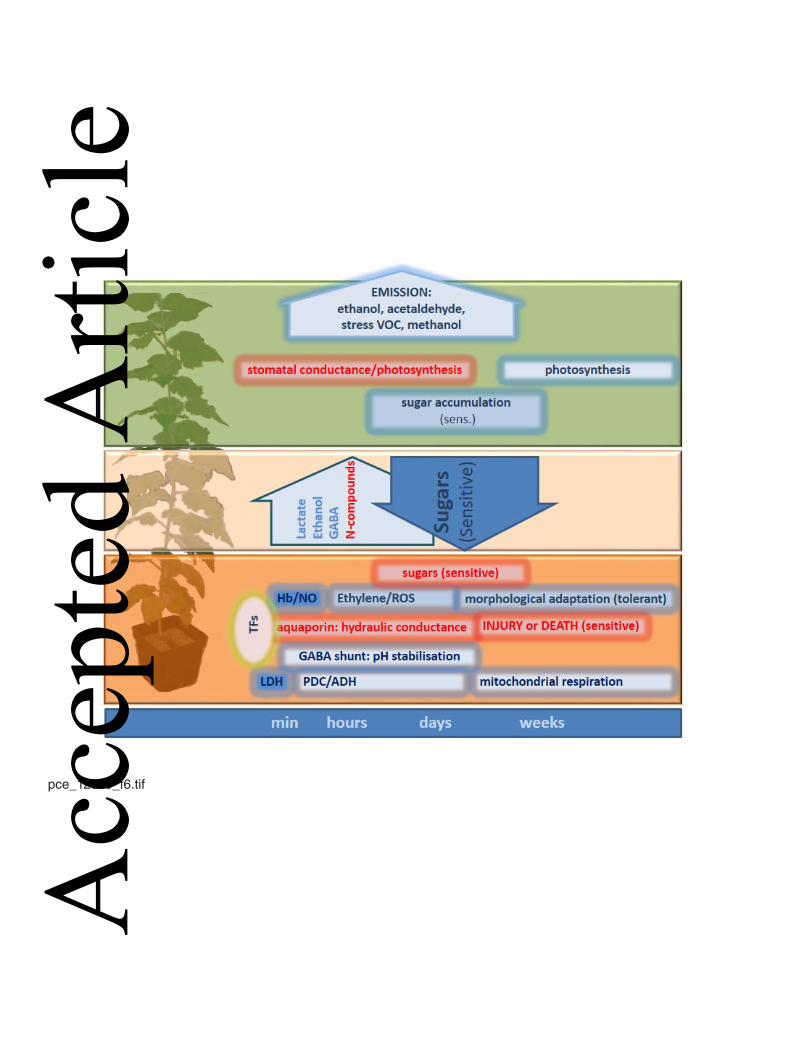
pce_12310_f6.tif
Acc
epte
d A
rticl
e





























