Embed Size (px)
Citation preview

1
Short title: UGT76D1 modulates immune response by DHBA glycosylation 1
2
Corresponding author details: Dr. Bing-kai Hou 3
School of Life Sciences, Shandong University, China 4
5
Article title: 6
Modulation of plant salicylic acid-associated immune responses 7
via glycosylation of dihydroxybenzoic acids 8
9
10
11
Xu-xu Huang, Guo-qing Zhu, Qian Liu, Lu Chen, Yan-jie Li, Bing-kai Hou 12
13
The Key Laboratory of Plant Development and Environment Adaptation Biology, 14
Ministry of Education; School of Life Science, Shandong University, Jinan 250100, 15
China. 16
17
18
One sentence summary: The glycosyltransferase UGT76D1 catalyzes the 19
glycosylation of dihydroxybenzoic acids and modulates plant salicylic acid 20
homeostasis and immune responses. 21
22
23
24
25
List of author contributions: B.K.H conceived the original research plans; X.X.H 26
performed the majority of the experiments; G.Q.Z, Q.L and L.C provided assistance 27
for some experiments; Y.J.L contributed to data analyses and interpretation; B.K.H. 28
and X.X.H wrote the article with input from all the authors. 29
30
31
32
Funding information:This research was supported by the National Natural Science 33
Foundation of China (no. 91217301, 31570299, 31770313 to B.K.H) 34
35
36
37
Corresponding author email:[email protected] 38
Plant Physiology Preview. Published on February 26, 2018, as DOI:10.1104/pp.17.01530
Copyright 2018 by the American Society of Plant Biologists
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
Abstract 39
Salicylic acid (SA) plays a crucial role in plant innate immunity. The deployment of 40
SA-associated immune responses is primarily affected by SA concentration, which is 41
determined by a balance between SA biosynthesis and catabolism. However, the 42
mechanisms regulating SA homeostasis are poorly understood. In this study, we 43
characterized a unique UDP-glycosyltransferase, UGT76D1, which plays an 44
important role in SA homeostasis and associated immune responses in Arabidopsis 45
thaliana. Expression of UGT76D1 was induced by treatment with both the pathogen 46
Pseudomonas syringae pv. tomato (Pst) DC3000 and SA. Overexpression of 47
UGT76D1 resulted in high SA accumulation, significant up-regulation of 48
pathogen-related genes, and a hypersensitive response (HR)-like lesion mimic 49
phenotype. This HR-like phenotype was not observed following UGT76D1 50
overexpression in SA-deficient NahG transgenic or sid2 plants, suggesting that the 51
phenotype is SA dependent. Biochemical assays showed that UGT76D1 glycosylated 52
2,3-dihydroxybenzoic acid (2,3-DHBA) and 2,5-dihydroxybenzoic acid (2,5-DHBA), 53
the major catabolic forms of SA, to their glucose and xylose conjugates in vitro and in 54
vivo. Moreover, in a mutant background blocked in the formation of 2,3-DHBA and 55
2,5-DHBA, UGT76D1 overexpression did not cause a HR-like lesion mimic 56
phenotype. Following infection with Pst DC3000, UGT76D1 knockout mutants 57
displayed a delayed immune response, with reduced levels of DHBA glycosides and 58
SA, and down-regulated SA synthase expression. By contrast, UGT76D1 59
overexpression lines showed an enhanced immune response and increased SA 60
biosynthesis before and after pathogen infection. Thus, we propose that UGT76D1 61
plays an important role in SA homeostasis and plant immune responses by facilitating 62
glycosylation of dihydroxybenzoic acids. 63
64
Key words: Arabidopsis, Glycosyltransferase, Salicylic acid, Dihydroxybenzoic acid, 65
Immune response 66
67
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

3
INTRODUCTION 68
Plants can initiate the specific hypersensitive response (HR), a localized 69
programmed cell death (PCD) response, at infection sites following challenge by 70
biotrophic pathogens (Heath, 2000; Van Doorn, 2011). This response is likely to be a 71
consequence of the progress of pathogen growth restriction (Coll et al., 2010; Coll et 72
al., 2011). The expression and regulation of HR relies on many components, such as 73
salicylic acid (SA), reactive oxygen species (ROS), jasmonic acid (JA), and ethylene 74
(ETH) (Coll et al., 2011). However, we currently have only a fragmented 75
understanding of the roles of the components leading to HR. 76
The identification of a number of lesion mimic mutants (LMMs), which display 77
spontaneous HR-like cell death, has increased our understanding of plant HR (Walbot 78
et al., 1983; Lorrain et al., 2003; Bruggeman et al., 2015). For instance, the LMM ssi4 79
from Arabidopsis, with a gain-of-function mutation of R-protein SSI4, constitutively 80
activates the ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1)-dependent SA 81
signaling pathways, and displays a conditional spontaneous lesion formation, 82
demonstrating the function of the R-protein in triggering HR (Shirano et al., 2002; 83
Zhou et al., 2004). One of the constitutive expresser of pathogenesis-related proteins 84
(cpr) mutants, cpr22, accumulates high levels of SA and displays a spontaneous lesion 85
phenotype (Yoshioka et al., 2001). Reducing the SA level in the accelerated cell death 86
(acd6) mutant or blocking SA signaling in the acd5 mutant attenuates the cell death 87
phenotype (Rate et al., 1999; Greenberg et al., 2000). Transgenic plants expressing 88
NahG (bacterial salicylate hydroxylase), which confers low SA concentration, are also 89
capable of inhibiting lesion formation, providing evidence that SA participates in the 90
regulation of cell death (Rate et al., 1999; Shah et al., 2001; Yoshioka et al., 2001). SA 91
can induce the expression of some pathogenesis-related (PR) genes, and promote the 92
production of ROS, and together they regulate cell death (Jabs, 1999; Alvarez, 2000; 93
Straus et al., 2010). In addition to disease resistance, SA also participates in multiple 94
processes in plants including senescence, which is a further kind of PCD. It has been 95
reported that leaf senescence and disease resistance use the same SA signaling 96
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

4
pathway (Vlot et al., 2009). 97
The biosynthesis of SA in plants has been proposed to occur through two 98
alternative pathways, namely the phenylalanine ammonia-lyase (PAL) and the 99
isochorismatesynthase (ICS) pathways. In the PAL pathway, benzoic acid and 100
ortho-coumaric acid are likely to be the two intermediates for SA biosynthesis 101
(Yalpani et al., 1993; Ryals et al., 1994; Chong et al., 2001). It is believed that the ICS 102
pathway produces most of the SA generated in plants (Wildermuth et al., 2001; 103
Dempsey et al., 2011). A past study showed that the SA level in sid2 (an ICS mutant) 104
was reduced to only 5–10% of the wild-type (WT) level after pathogen infection 105
(Wildermuth et al., 2001). 106
SA biosynthesis is regulated by multiple and complex factors. For instance, 107
ENHANCED DISEASE SUSCEPTIBILITY 1 (EDS1) and NON-RACE-SPECIFIC 108
DISEASE RESISTANCE 1 (NDR1) have been identified as upstream regulators of 109
SA biosynthesis in the effector-triggered immune response (ETI) (Aarts et al., 1998). 110
PHYTOALEXIN DEFICIENT 4 (PAD4) and SENESCENCE-ASSOCIATED 111
CARBOXYLESTERASE 101 (SAG101) are two components of EDS1-mediated 112
immunity (Feys et al., 2001; Feys et al., 2005). In addition, SYSTEMIC ACQUIRED 113
RESISTANCE DEFICIENT1 (SARD1) and its homolog CALMODULIN-BINDING 114
PROTEIN 60g (CBP60g) have been shown to be transcription activators of ICS1 115
expression (Zhang et al., 2010). 116
In addition to upstream regulation, active SA level is also modulated by 117
downstream metabolic modifications, including glycosylation, methylation, amino 118
acid conjugation, and hydroxylation. Different modifications of SA are likely to have 119
different functions in plant defense. Two main glycosyltransferases, UDP-dependent 120
glycosyltransferase 74F1 (UGT74F1) and UGT74F2, can transform SA into its 121
glucoside (SAG) or its glucose ester (SGE) (Lim et al., 2002; Dean and Delaney, 122
2008). SAG and SGE are the inactive forms of SA, and can be transformed back into 123
active SA when plants are challenged by pathogens (Dean and Delaney, 2008). Amino 124
acid-conjugated SA has been reported to be a potential activator of plant immunity 125
(Chen et al., 2013). Hydroxylated SA, including 2,3-dihydroxybenzoic acid 126
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

5
(2,3-DHBA) and 2,5-dihydroxybenzoic acid (2,5-DHBA), is the major metabolic 127
form of SA. Recently, the enzymes catalyzing SA to 2,3-DHBA (S3H) and to 128
2,5-DHBA (S5H) have been identified, and SA was found to accumulate in s3h or 129
s5h mutants (Zhang et al., 2013; Zhang et al., 2017). 130
The roles of 2,3-DHBA and 2,5-DHBA in Arabidopsis are not known; however, 131
2,5-DHBA in tomato is thought to play a signaling role in the activation of inducible 132
defenses. The signaling role of 2,5-DHBA is thought to be complementary to SA 133
because 2,5-DHBA induces the formation of a different set of pathogenesis-related 134
(PR) proteins that are not induced by SA (Bellés et al. 1999). Because of the 135
cytotoxicity of DHBA, 2,3-DHBA and 2,5-DHBA always exist as sugar conjugates in 136
plants (Bartsch et al., 2010). However, it appears that sugar conjugation of DHBA has 137
other physiological significance besides detoxification. 2,5-DHBA 138
5-O-β-D-xylopyranoside has been found to accumulate to high levels in cucumber 139
and tomato after inoculation with different pathogens (Fayos et al., 2006). It was also 140
reported that the sugar conjugates of 2,3-DHBA and 2,5-DHBA increased in 141
Arabidopsis during expression of pathogen resistance (Bartsch et al., 2010). Research 142
suggested that glycosylation of DHBA may be involved in plant immunity. Some 143
glycosyltransferases have been identified to be capable of catalyzing the 144
transformation of 2,3-DHBA and 2,5-DHBA in vitro to form their glucosyl or xylosyl 145
conjugates, although very little has been carried out towards understanding the 146
physiological relevance of these conjugates (Lim et al., 2001; Lim et al., 2002; Chen 147
and Li, 2017). As a consequence, it is still unclear whether DHBA glycosylation 148
functions in plant defense response and, if so, what its molecular mechanism entails. 149
Here, we identified and characterized the pathogen-induced glycosyltransferase 150
UGT76D1. We found that UGT76D1 was involved in the lesion mimic HR, including 151
the formation of natural necrotic spots, ROS accumulation, SA homeostasis, and PR 152
gene expression in rosette leaves of Arabidopsis. Furthermore, our experiments 153
revealed that UGT76D1 can catalyze the formation of 2,3-DHBA and 2,5-DHBA 154
glycosides in vitro and in vivo. We found that blocking DHBA glycosylation 155
abolished the UGT76D1-induced HR. Together, our data suggest that the 156
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

6
glycosylation of DHBA plays a previously unrecognized role in the plant innate 157
immune response through modulating SA homeostasis. 158
RESULTS 159
UGT76D1 expression is induced by the plant pathogen Pst DC3000 160
Through searching the publicly available microarray data for UDP-dependent 161
glycosyltransferase (UGT) genes associated with plant secondary metabolism, we 162
found that several Arabidopsis UGT genes were induced by Pseudomonas syringae pv. 163
tomato DC3000 (Pst DC3000). UGT76D1 is one of the most highly 164
pathogen-responsive UGT genes. Since the UGT76D subfamily contains only this 165
unique member, UGT76D1 may have an important and specific role in plant immune 166
responses. To examine the reliability of microarray data, RT-qPCR was used to 167
analyze the expression of UGT76D1 at different time points after treatment with Pst 168
DC3000 or SA. The results showed that expression of UGT76D1 was strongly 169
induced by either Pst DC3000 or SA (Figs.1A and 1B). UGT76D1Pro::GUS 170
transgenic plants were constructed and GUS staining also demonstrated the high 171
induction of UGT76D1 expression by pathogen infection (Fig.1C), but not by MgCl2 172
treatment used as a negative control (Fig.1D). Thus, UGT76D1 is a 173
pathogen-responsive gene, and may be involved in plant defense responses. 174
UGT76D1 is involved in the spontaneous HR-like lesion mimic 175
response 176
To study the function of UGT76D1 in plants, we constructed UGT76D1 177
knockout mutants and transgenic lines overexpressing UGT76D1. Through the 178
CRISPR/Cas9 system, two independent mutant lines of UGT76D1 (ko-33, ko-36) 179
were isolated, each with a nucleotide deletion in the target site of the 5′ coding region 180
(Fig.S1, A–D), which causes a frameshift mutation. Nineteen independent UGT76D1 181
overexpression lines were obtained, and two lines (OE-17, OE-32) with a high 182
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

7
expression of UGT76D1 were used in this study (Fig.S1, E). Under long-day 183
conditions, mutant lines exhibited a phenotype comparable to that of WT plants. On 184
the other hand, UGT76D1 overexpression lines showed obvious spontaneous necrotic 185
lesions, a phenotype that mimics hypersensitive cell death (Fig.2A). In the T2 186
generation of OE-17 and OE-32 lines, the lesion phenotype co-segregated with the 187
antibiotic resistance marker (Table S1). The lesions became more severe under 188
short-day conditions (Fig.S2). In addition to the spontaneous necrotic lesions, leaf 189
senescence in UGT76D1 overexpression lines also occurred earlier than in WT plants 190
(Fig.S3). 191
LMMs are mutants with spontaneous HR-like cell death without pathogen 192
infection (Lorrain et al., 2003; Bruggeman et al., 2015). The phenotype of UGT76D1 193
overexpression lines was similar to that of LMM mutants, suggesting a constitutively 194
activated hypersensitive response and localized cell death in the UGT76D1 195
overexpression lines. Trypan blue staining further confirmed cell death in UGT76D1 196
overexpression lines (Fig.2B). Reactive oxygen species (ROS) overproduction can 197
lead to cell death and is a family of crucial signal molecules in the activation of PCD 198
during HR (Malik et al., 2014). DAB staining showed that UGT76D1 overexpression 199
lines accumulated considerable amounts of hydrogen peroxide, whereas UGT76D1 200
knockout mutant lines exhibited hydrogen peroxide levels comparable to those in the 201
WT plants (Fig.2C), a phenomenon which correlated with the HR-like phenotypes of 202
these lines. We next examined the expression of the 203
SA-regulated PR1 and PR2 defense genes. PR1 and PR2 were drastically up-regulated 204
in UGT76D1 overexpression lines, whereas they were downregulated in the 205
UGT76D1 knockout mutant lines (Figs.2D, 2E), suggesting the involvement of 206
UGT76D1 in the SA-associated innate immune response. 207
The difference in changes of PR1 and PR2 expression between UGT76D1 208
overexpression and knockout mutant lines, as well as the spontaneous HR-like lesion 209
mimic phenotype occurring in the UGT76D1 overexpression lines, but not in the 210
mutant lines, prompted us to investigate SA levels in these lines. Our analysis showed 211
that free and total SA levels were considerably higher in UGT76D1 overexpression 212
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

8
lines than that in WT plants, but were moderately reduced in the mutant lines 213
compared to the WT (Figs.2F, 2G). These data suggest that UGT76D1 may be 214
involved in the plant innate immune response through modulating SA homeostasis. To 215
further explore whether UGT76D1 was involved in pathogen-induced HR, we tested 216
the ability of UGT76D1 mutants to trigger the HR against Pst DC3000 carrying 217
avrRpt2. We found that UGT76D1 knockout mutants suppressed the HR induced by 218
Pst DC3000 carrying avrRpt2 and the disease symptoms occurring at 48 h after 219
inoculation (Fig.3A). Cell death in each line was visualized with trypan blue staining 220
(Fig.3B). These results suggested that UGT76D1 may be a positive activator in at 221
least the Pst DC3000 (avrRpt2)-induced HR. 222
SA is required for the UGT76D1-activated lesion mimic phenotype 223
To determine whether the UGT76D1-activated lesion mimic phenotype is SA 224
dependent, UGT76D1 overexpression lines were crossed transgenic plants expressing 225
NahG, a SA hydroxylase that degrades SA (Delaney et al., 1994). Hybrids with high 226
expression of both UGT76D1 and NahG were selected, and named OE-17 NahG and 227
OE-32 NahG. It was shown that NahG can restore a WT phenotype (Fig. 4A) and 228
prevent a spontaneous lesion mimic phenotype despite UGT76D1 overexpression in 229
hybrid lines (Fig. 4B). Corresponding with the rescued leaf phenotype, cell death 230
(indicated by Trypan blue staining), the H2O2 content (indicated by DAB staining), 231
and the expression of PR1 and PR2 were restored to the levels of WT plants following 232
NahG expression in UGT76D1 overexpression lines (Figs. 4C, 4D and 4E). Most 233
importantly, the contents of free and total SA were reduced to undetectable levels in 234
the UGT76D1 overexpression NahG hybrids (Figs. 4F and 4G). Thus, we propose that 235
SA accumulation is the main factor triggering the UGT76D1-activated lesion mimic 236
phenotype. 237
UGT76D1 possesses biochemical activity towards DHBA 238
glycosylation in vitro 239
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

9
To investigate the mechanism by which UGT76D1 was involved in maintaining 240
SA homeostasis, we next carried out in vitro screening of substrates catalyzed by this 241
glycosyltransferase. Fusion proteins of UGT76D1 with the GST-tag 242
were heterologously expressed in E.coli and then purified (Fig. 5A). When SA was 243
tested as a candidate substrate for UGT76D1, UGT76D1 exhibited no detectable 244
enzymatic activity towards SA, suggesting that SA was not the natural substrate (Fig. 245
5B). We further extended the candidate substrates to SA metabolites, phenolic 246
compounds, plant hormones, and flavonoids (Supplemental Table 2). UGT76D1 had 247
high enzymatic activities towards only the hydroxylated products of SA, namely 248
2,3-DHBA and 2,5-DHBA (Figs. 5C and 5D). No activity or only trace activity was 249
observed towards other compounds in our analyses (Table S2). It was previously 250
reported that 2,3-DHBA and 2,5-DHBA mainly form both glucosides and xylosides in 251
planta, although the physiological roles of these glycosides were unknown (Dempsey 252
et al., 2011). Thus, both UDP-glucose and UDP-xylose were used as the sugar donors 253
in our biochemical assays. We found that UGT76D1 can catalyze 2,3-DHBA and 254
2,5-DHBA to form their corresponding glucosides and xylosides (Figs. 5C and 5D). 255
LC-MS analysis confirmed the identity of the glucosides and xylosides produced from 256
2,3-DHBA and 2, 5-DHBA (Fig. S4). A previous study had found that 2,3-DHBA and 257
2,5-DHBA can be conjugated by UGT89A2 to form 2,3-DHBA or 2,5-DHBA 258
xylosides (Li et al., 2014). To determine the DHBA and sugar donor preferences of 259
UGT76D1 and UGT89A2, specific enzyme activity was tested in vitro. UGT89A2 260
exhibited a strong preference toward UDP-xylose over UDP-glucose. In contrast, 261
UGT76D1 preferred UDP-glucose to UDP-xylose (Table S3). 262
UGT76D1 functions in DHBA glycosylation in vivo 263
To further investigate the biochemical function of UGT76D1 in vivo, glycosides 264
of 2,3-DHBA and 2,5-DHBA were extracted and measured from UGT76D1 265
overexpression and knockout mutant lines. The high performance liquid 266
chromatography (HPLC) profiling of the DHBA sugar conjugates showed that 267
UGT76D1 overexpression lines accumulated much more 2,3-DHBA and 2,5-DHBA 268
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

10
glycosides than that in WT under non-challenged conditions. However, in 269
non-challenged UGT76D1 mutant lines, only 2,3-DHBA xyloside was significantly 270
affected, accumulating at levels lower than that in WT (Figure 6A and 6B). The 271
relative level of DHBA glucosides in the UGT76D1 knockout mutants was very low 272
and comparable to that in WT. 273
The very low base levels of DHBA glucosides made them difficult to measure 274
accurately. To overcome this difficulty, we applied 100 μm 2,3-DHBA and 2,5-DHBA 275
to plants and re-measured the DHBA glucoside concentrations. Although the level of 276
2,5-DHBA glucosides was comparable between UGT76D1 knockout mutants and WT, 277
2,3-DHBA glucoside level in the mutants was much lower than that in WT (Figs. S5A 278
and S5B). Meanwhile, it was observed that UGT76D1 overexpression lines 279
accumulated much higher concentrations of both 2,3-DHBA and 2,5-DHBA 280
glucosides than that in WT upon application of DHBA; a similar result occurred with 281
UGT76D1 overexpression lines without DHBA application. Because expression of 282
UGT76D1 is Pst DC3000 inducible, the concentrations of 2,3-DHBA glycoside and 283
2,5-DHBA glycoside in UGT76D1 mutants and WT were re-measured 48 h after 284
infiltration with Pst DC3000 or with MgCl2 as the negative control. Our analysis 285
revealed that the accumulation of both 2,3-DHBA and 2,5-DHBA glucosides was 286
significantly reduced in Pst DC3000-inoculated UGT76D1 mutant plants compared to 287
that in WT (Figs. 6C and 6D). The concentrations of both 2,3-DHBA and 2,5-DHBA 288
xylosides were only slightly reduced in Pst DC3000-inoculated UGT76D1 mutants. 289
These data suggested that UGT76D1 functions in DHBA glycosylation in vivo and 290
may be the main enzyme responsible for the glucosylation of 2,3-DHBA and 291
2,5-DHBA. 292
We also attempted to determine the concentrations of the free forms of 2,3-DHBA 293
and 2,5-DHBA in the UGT76D1 overexpression and knockout mutant lines. We found 294
that the 2,3-DHBA level was too low to be detectable with or without pathogen 295
inoculation, a result which is consistent with those from previous studies (Bartsch et 296
al., 2010; Zhang et al., 2013). For free 2,5-DHBA, however, an interesting and 297
unexpected change was observed. After Pst DC3000 inoculation, UGT76D1 298
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

11
overexpression lines accumulated much more of the free form of 2,5-DHBA than was 299
determined in the WT, whereas mutant lines had greatly reduced levels of the free 300
form of 2,5-DHBA compared to that in WT (Fig. S6). There are several hypotheses to 301
explain this unexpected change. Firstly, overexpression or mutation of UGT76D1 302
could alter metabolic flux towards the conjugated DHBA form, resulting in feedback 303
activation or inhibition of upstream enzymes of DHBA biosynthesis. Similar 304
situations have been found in previous studies (Chong et al., 2002; Tognetti et al., 305
2010). Secondly, one part of the DHBA pool may arise from cleavage of the 306
glucosylated form by glucosidases. Thirdly, because of the cytotoxicity of DHBAs, an 307
overcompensated degradation of DHBAs may occur in UGT76D1 mutants. 308
Immune response activated by UGT76D1 depends on the formation 309
of 2,3-DHBA and 2,5-DHBA glycosides 310
Previous research indicated that SA 3-hydroxylase (S3H) can convert SA to 311
2,3-DHBA, whereas the formation of 2,3-DHBA is blocked in the mutant s3h (Zhang 312
et al., 2013). To investigate the effects of UGT76D1 on the immune response in the 313
absence of 2,3-DHBA, the preferred substrate of UGT76D1, we produced hybrids 314
between UGT76D1 overexpression lines and s3h mutants. It was found that the s3h 315
background inhibited the spontaneous lesion mimic phenotype caused by UGT76D1 316
overexpression (Fig. 7A). DAB and Trypan blue staining also showed reduced levels 317
of H2O2 and cell death in the UGT76D1 overexpression s3h hybrid lines (Fig. 7B). 318
RT-qPCR analysis revealed that, although the expression of UGT76D1 remained at 319
high levels in the UGT76D1 overexpression s3h hybrids (Figs. 7C), the expression of 320
both PR1 and PR2 was significantly reduced (Figure 7D). We also measured the SA 321
levels in UGT76D1 overexpression s3h hybrid plants and found that the 322
concentrations of both free SA and total SA were markedly reduced (Fig. 7E). These 323
data indicated that the formation of 2,3-DHBA glycosides under the presence of 324
substrate 2,3-DHBA would be important for UGT76D1-activated immune response. 325
Recently, SA 5-hydroxylase (S5H) has also been described (Zhang et al., 2017). 326
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

12
It was reported that the concentrations of 2,3-DHBA and 2,5-DHBA were reduced in 327
the s3h s5h double mutant, which exhibited a dwarf and early-senescence phenotype 328
(Zhang et al., 2017). Here, we further examined the effect of increased UGT76D1 329
expression on the defense response in the s3h s5h double mutant background. We 330
found that UGT76D1 overexpression in s3h s5h double mutants did not cause the 331
HR-like lesion mimic phenotype seen in the WT background (Fig. 7F), further 332
demonstrating that the function of UGT76D1 in activating the immune response is 333
dependent on DHBA glycosylation. 334
DHBA glycosylation catalyzed by UGT76D1 modulates SA 335
biosynthesis during pathogen infection 336
SA is believed to be the crucial signaling molecule in SA-dependent plant 337
defense responses, where PR1 and PR2 are known to be important marker genes, the 338
expression of which is regulated by SA. It appears that the levels of SA and the 339
expression of PR1/PR2 always correlate with the lesion mimic phenotype of the 340
UGT76D1 overexpression and mutant lines, suggesting an important relationship 341
between UGT76D1-catalyzed DHBA glycosylation and SA synthesis. To further 342
investigate the physiological effects of UGT76D1 on SA synthesis in the plant 343
defense response, we analyzed the changes in SA concentration and PR expression in 344
UGT76D1 overexpression and mutant lines challenged by Pst DC3000 or the MgCl2 345
control. The expression of PR1 and PR2 was detected at different time points after 346
plants were inoculated with Pst DC3000 or the control buffer. It was found that PR1 347
and PR2 had high expression levels in the overexpression lines even at the beginning 348
of the treatment. As the time after pathogen challenge increased, the expression of 349
PR1 and PR2 increased rapidly in UGT76D1 overexpression lines. However, the 350
expression of PR1 and PR2 increased only slowly in the WT. In the UGT76D1 mutant 351
lines, the expression levels of PR1 and PR2 were even lower than that in WT at every 352
time point of pathogen treatment (Figs. 8A and 8B), suggesting that the defense 353
response was retarded by the loss of function of UGT76D1. We analyzed the free and 354
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

13
total SA concentrations in the UGT76D1 overexpression and mutant lines after 355
exposure to Pst DC3000 or MgCl2 for 48 h. Although SA concentrations in UGT76D1 356
overexpression lines were little more than that in WT 48 h after inoculation, the SA 357
concentrations in the knockout mutant lines were much lower than that in WT (Figs. 358
8C and 8D). These data suggested that the physiological role of UGT76D1 in the 359
SA-dependent defense response was a consequence of modulating SA levels. 360
The mechanism by which SA concentration is modulated by UGT76D1 was also 361
investigated. We analyzed the expression of SA synthesis-related genes in UGT76D1 362
overexpression and mutant lines with or without challenge by Pst DC3000. Without 363
Pst DC3000 treatment, the expression of several key SA-related genes, namely the 364
SA biosynthesis gene ICS1 and the SA biosynthesis regulatory genes EDS1 and PAD4, 365
were substantially increased in the UGT76D1 overexpression lines, whereas they 366
were only slightly decreased in the mutant lines (Fig. 8E). After challenge by Pst 367
DC3000 for 24 h or 48 h, the expression of the key SA biosynthetic gene ICS1 was 368
dramatically decreased in the UGT76D1 mutant lines (Fig. 8F), although the 369
expression levels of EDS1 and PAD4 were still only slightly decreased. It is likely that 370
UGT76D1 was involved in the positive feedback regulation of SA biosynthesis, 371
particularly via the regulation of the ICS1 gene. 372
To verify the role of ICS1 in the UGT76D1-mediated innate immune response, 373
we generated hybrids between UGT76D1 overexpression lines with sid2, an ICS1 374
mutant. Although expression of UGT76D1 remained at a high level in the hybrid lines, 375
the UGT76D1-activated lesion mimic phenotype was totally suppressed (Figs. 9A and 376
9B). We also measured the SA concentrations in the hybrid lines. It was found that 377
both the free SA and total SA were reduced to levels lower than that in WT plants (Fig. 378
9C). These data indicated that the pathogen-induced glycosyltransferase UGT76D1 379
functions in the defense response through modulating salicylic acid biosynthesis. The 380
glycosylation of DHBA by UGT76D1 may be involved in a positive feedback loop of 381
SA biosynthesis, accelerating SA accumulation in the process of plant defense 382
response. 383
384
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

14
DISCUSSION 385
UGT76D1 is a novel DHBA glycosyltransferase 386
Previous experiments had shown that SA is the main precursor of 2,3-DHBA and 387
2,5-DHBA (Ibrahim and Towers, 1959). The two DHBAs usually exist in a 388
glycosylated form (glucoside or xyloside), and were considered to be among the 389
major inactive forms of SA (Bartsch et al., 2010). However, the enzyme responsible 390
for the formation of those DHBA glycosides had previously been unknown. Recent 391
studies indicated that UGT89A2 is a 2,5-DHBA xylosyltransferase in Arabidopsis (Li 392
et al., 2014). However, overexpression of the UGT89A2 gene did not result in any 393
overt phenotype (Chen and Li, 2017). In this study, UGT76D1 was identified as a 394
novel DHBA glycosyltransferase in Arabidopsis. UGT76D1 can glycosylate both 395
2,3-DHBA and 2,5-DHBA, using either UDP-glucose or UDP-xylose as sugar donors. 396
Our analyses indicated that lines overexpressing UGT76D1 accumulated much higher 397
concentrations of DHBA glycosides, particularly the DHBA glucosides, than did the 398
WT. However, knockout mutant lines of UGT76D1 contained much lower 399
concentrations of DHBA glucosides following pathogen challenge. These data suggest 400
that the role of UGT76D1 in the defense response is mainly with respect to the 401
glucosylation of DHBAs. This hypothesis received support from our enzyme activity 402
analyses. Our results indicated that UGT76D1 prefers UDP-glucose to UDP-xylose as 403
the sugar donor for DHBA glycosylation. By contrast, although UGT89A2 exhibited 404
enzyme activity towards the DHBAs, it exhibited a strong preference toward 405
UDP-xylose over UDP-glucose. UGT76D1 and UGT89A2 seem to be too distant on 406
the phylogenetic tree, whereas they can act on the same substrate, DHBA. Actually, 407
this is a common phenomenon. For example, UGT73C6 and UGT78D1, two UGTs 408
belonging to different groups in the GT1 family, can act on the same flavonol 409
molecules (Jones et al., 2003). Another example involves auxin glycosylation. Both 410
UGT74E2 and UGT75D1 can catalyze the glycosylation of the auxin indole-3-butyric 411
acid (IBA) (Tognetti et al., 2010; Zhang et al., 2016), whereas UGT84B1 is reported 412
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

15
to act on the auxin indole-3-acetic acid (IAA) (Jackson et al., 2002). What is the 413
biological significance for plants to have several different UGTs acting on the same 414
substrate? It is speculated that plant evolution has involved the formation of 415
functionally redundant multiple glycosyltransferases active towards the same type of 416
substrates. A synergistic or coordinated interaction between different 417
glycosyltransferase members may be meaningful for the fine tuning of metabolic 418
homeostasis. On the other hand, the activities of these enzymes will depend on cell 419
specificity of enzyme expression, relative availability of substrates and sugar donors, 420
and relative compartmentalization of the enzyme and substrates, which may have the 421
potential to enhance the plant’s flexibility in development or in adaptation to diverse 422
environments. 423
Pathogen-induced UGT76D1 may act as a positive component to promote 424
hypersensitive response 425
In plants, the innate immune system has evolved to protect them from the 426
pathogens which attack them, but the mechanisms involved in mediating this defense 427
are poorly understood. Plant cells are usually capable of perceiving signals from 428
pathogens and inducing resistance to the challenge, which is achieved sometimes by 429
launching a cell-suicide mechanism to produce a localized PCD, known as HR, at the 430
infection site (Heath, 2000). The fates of cells at the infection site and in adjacent 431
positions are different due to the SA concentration gradient. In the infection site, high 432
levels of SA induced the degradation of NPR1 (an inhibitor of PCD in ETI), and then 433
cell death. By contrast, in cells adjacent to the infection site, the SA level is not high 434
enough to lead to cell death, so the cells survive and enhance resistance (Fu and Dong, 435
2013). UGT76D1 is a glycosyltranferase gene induced by pathogens. Transgenic 436
plants with high levels of expression of UGT76D1 displayed spontaneous local cell 437
death, a lesion mimic and HR-like phenotype. This finding suggested that UG76D1 438
may be a positive component in the regulation of the HR-related immune response. 439
When the SA hydroxylase gene NahG was transferred into UGT76D1 overexpression 440
lines, the lesion mimic phenotype, the high levels of H2O2 and the upregulated PR 441
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

16
expression disappeared. Thus, high levels of SA are required for lesion formation in 442
UGT76D1 overexpression lines. Under normal conditions, the basal expression of 443
UGT76D1 is very low. When plants were challenged by pathogens such as Pst 444
DC3000, UGT76D1 expression was induced. Based on these findings, we proposed 445
that UGT76D1 acts as a positive component, playing an important role in the 446
SA-activated hypersensitive response. 447
The glycosylation of DHBAs may be an important process responsible for the 448
UGT76D1-mediated innate immune response 449
2,5-DHBA glucosides accumulated in tomato and cucumber plants after infection 450
by different pathogens (Bellés et al., 1999; Fayos et al., 2006), while 2,3-DHBA 451
xylosides increased in Arabidopsis after induction by Pst DC3000 (avrRpm1) 452
(Bartsch et al., 2010). It is likely that DHBA glycosides have important roles in the 453
plant defense response. In this study, UGT76D1 overexpression lines can accumulate 454
much higher concentrations of 2,3- and 2,5-DHBA glycosides (glucosides and 455
xylosides) than that in WT. By contrast, the UGT76D1 mutants produced less 2,3- and 456
2,5-DHBA glucosides than that in WT after inoculation with Pst DC3000, suggesting 457
the greater role of 2,3-DHBA and 2,5-DHBA glucosylation in the 458
UGT76D1-activated immune response. S3H and S5H are enzymes responsible for the 459
conversion of SA to 2,3-DHBA and 2,5-DHBA, and the s3h s5h double mutants 460
abolished the production of 2,3-DHBA and 2,5-DHBA (Zhang et al., 2013; Zhang et 461
al., 2017). In our experiments, when UGT76D1 was overexpressed in s3h mutant or 462
s3h s5h double mutant backgrounds, we found that the HR-like lesion mimic 463
phenotype caused by UGT76D1 overexpression was totally suppressed. These results 464
suggested that the lack of glycosylation of 2,3-DHBA on its own or of both 465
2,3-DHBA and 2,5-DHBA abolished the UGT76D1-activated immune response. How 466
does the s3h single mutant suppress the lesion mimic phenotype caused by UGT76D1? 467
We proposed several possible reasons. Firstly, it was reported that S3H can convert 468
SA to both 2,3-DHBA and 2,5-DHBA in vitro (Zhang et al., 2013), therefore, s3h 469
mutation may be efficient for blocking the formation of 2,3-DHBA and 2,5DHBA. In 470
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

17
addition, s3h mutant nearly abolished the formation of 2,3-DHBA, while s5h mutant 471
still maintain the formation of 2,5-DHBA to a greater extent (Zhang et al., 2017), 472
Which may be due to the enzyme activity of S3H in s5h mutant and also suggest a 473
more crucial role for S3H in the metabolic flux from SA to DHBAs. Consistent with 474
this hypothesis, our results indicated that SA level was restored to the s3h level in the 475
UGT76D1OE s3h hybrid plants. Secondly, because of the difference in molecular 476
structures, 2,3-DHBA glucoside may have different roles from 2,5-DHBA glucoside 477
in the defense responses, thus resulting in the elimination of the lesion phenotype 478
when UGT76D1 was overexpressed in a s3h background. Thirdly, 479
UGT76D1-activated immune response is likely to depend on a threshold of 480
2,3-DHBA and 2,5-DHBA concentrations. 481
It was reported that SA accumulated in the s3h s5h double mutant or the s3h 482
single mutant. However, the s3h and s3h s5h mutants did not show the HR-like lesion 483
phenotype as was exhibited in the UGT76D1 overexpression lines, but only showed 484
dwarf or earlier leaf senescence traits when compared to the WT (Zhang et al., 2013; 485
Zhang et al., 2017). Previous studies had indicated that leaf senescence and HR are 486
both examples of PCD, triggered by different stimuli (Love et al., 2008; Rivas-San 487
Vicente and Plasencia, 2011). Here, UGT76D1 was demonstrated to be involved in 488
the HR-like immune response, which was different from the involvement of S3H or 489
S5H in senescence, suggesting that UGT76D1 may respond to the different stimuli 490
from S3H and S5H. In the past, DHBA glycosides were viewed simply as the storage 491
forms of DHBA. In this study, however, we revealed that the sugar conjugates of 492
DHBA, or the process towards their formation, are likely to be an important trigger to 493
initiate PCD in the plant HR response. 494
In this study, we also found that UGT76D1 has trace activity toward coumarins 495
and some other compounds in vitro. Previous studies indicated that scopoletin, a 496
coumarin compound, may be a reactive oxygen intermediate (ROI) scavenger, and 497
play a role in redox homeostasis during the HR of tobacco to TMV (Chong, 2002). 498
Here, because UGT76D1 has only a trace activity toward coumarins in vitro, we 499
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

18
propose that the glycosylation of coumarins may not be the major stimulus to HR 500
phenotype mediated by UGT76D1. 501
The glycosylation of DHBA may be involved in the positive feedback regulation 502
of SA synthesis 503
SA is a crucial signal for plant defense responses. Its levels and dynamic changes 504
in tissues will directly influence the SA-dependent defense responses of plants. Thus, 505
SA concentrations in plant defense responses must be precisely controlled (Shah, 506
2003; Fu and Dong, 2013). When challenged by a pathogen, plants need to rapidly 507
synthesize more SA at the challenge sites to promote cell death. An amplification loop 508
of immune response may exist. For example, SA itself can activate the expression of 509
R genes and further promote downstream signals to synthesize more SA (Yang and 510
Hua, 2004). Previous studies have usually focused their attentions on the upstream 511
regulation of SA, but with very little attention being paid to downstream regulation of 512
SA. Whether or not the downstream events of SA biosynthesis pathway have an 513
important role in modulating SA concentration is largely unknown. Recently, it was 514
reported that the concentrations of DHBA glycosides increased after plants were 515
inoculated with Pst DC3000 (Bartsch et al., 2010). It was also noticed that expression 516
of both S3H and S5H were induced by plant pathogens, affecting the SA level (Zhang 517
et al., 2013; Zhang et al., 2017). These findings suggested the importance in SA 518
signaling of downstream metabolic processes of SA. In a previous study, 519
glycosyltransferase UGT76B1 was found to be involved in the modulation of SA 520
levels and defense responses (von Saint Paul et al., 2011). Isoleucic acid 521
(2-hydroxy-3-methyl-pentanoic acid) was identified to be a substrate for UGT76B1, 522
and UGT76B1 is known to function as a novel player in SA-JA signaling crosstalk 523
(von Saint Paul et al., 2011). However, the step at which isoleucic acid influences SA 524
level or its action mechanism is currently not clear. In another report, UGT76B1 was 525
found to have SA glycosyltransferase (SAGT) activity, and some immune-priming 526
compounds can repress the activities of SAGTs including UGT74F1and UGT76B1 527
(Noutoshi et al., 2012). These examples suggest the complexity of regulation 528
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

19
mechanisms in SA levels and defense response. 529
In this study, our focus was on the glycosylation of 2,3- and 2,5-DHBA, the two 530
main forms of hydroxylated SA. UGT76D1 was found to encode DHBA 531
glycosyltransferase, which can convert both 2,3-DHBA and 2,5-DHBA to their 532
glycosides, particularly the glucosides. More importantly, we found that the levels of 533
SA can be changed after disturbing the expression of UGT76D1 in the pathogen 534
infection process. Our analyses indicated that the key SA synthase gene, ICS1, was 535
significantly upregulated in UGT76D1 overexpression lines, but downregulated in 536
UGT76D1 knockout mutants. The findings presented here suggested that the 537
glucosylation of DHBA catalyzed by UGT76D1 may be involved in the positive 538
feedback regulation of SA synthesis. Therefore, this study revealed that the 539
glucosylation of DHBA plays a previously unrecognized role in modulating SA 540
homeostasis and plant immune response. 541
In conclusion, we propose a putative working model for UGT76D1 (Figure 10). 542
When plants are challenged by pathogens, expression of UGT76D1 is induced to a 543
high level, and the formation of DHBA glycosides is accelerated. At the same time, 544
DHBA glycosylation may activate SA synthase and then increase SA synthesis 545
through an unknown positive feedback loop, possibly through increasing the 546
metabolic flux towards SA and the DHBAs. High levels of SA accumulation lead to 547
oxidative burst and PR gene expression, and ultimately lead to PCD and increased 548
plant resistance to pathogens. Considering the xylosyltransferase activity of 549
UGT89A2 towards DHBAs, we cannot exclude the role of UGT89A2 in this 550
immune-priming process. However, the detailed mechanism by which DHBA 551
glycosylation regulates SA biosynthesis requires further investigation. 552
553
554
555
556
557
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

20
MATERIALS AND METHODS 558
Plant materials and growth conditions 559
Arabidopsis ecotype Columbia-0 (Col-0) was used in this study. For selection of 560
transgenic plants, SA treatment, and GUS staining, seeds were sterilized and imbibed 561
for 3 d at 4°C in the dark and sown on plates with Murashige and Skoog medium and 562
1% (w/v) Suc, solidified with 0.8% (w/v) phytoagar. The plates were transferred to a 563
growth room with a temperature at 22°C in a long day (16 h light/8 h dark). For all 564
other analyses, plants were grown for 1 week as described above and transferred to 565
soil under a long day condition (16 h light/8 h dark, approximately 100 µmol photons 566
m−2
s−1
) for the indicated times. A short day condition (8 h light/16 h dark) was used 567
to determine whether the lesion phenotype of UGT76D1 overexpression lines is 568
influenced by light cycle. 569
GUS staining 570
Sequences about 1500 bp upstream of the UGT76D1 transcription start were 571
amplified from the Arabidopsis genome by PCR using UGT76D1 promoter specific 572
primers (Table S4). The amplified sequences were inserted into the pBI121 plasmid to 573
replace the CaMV35S promoter. Arabidopsis was transformed with the 574
UGT76D1pro:GUS vector to generate UGT76D1pro:GUS transgenic plants through 575
the floral dip method (Clough and Bent, 1998). 576
For GUS staining assays, leaves of two-week-old transgenic seedlings of 577
UGT76D1pro:GUS were soaked in 10 mM MgCl2 (control) or Pst DC3000 (OD600 = 578
0.01). Seedlings were then collected in different time points for GUS staining. At each 579
time point, GUS activity was measured in at least twelve individual plants, most of 580
them with similar results were used to take photos. 581
GUS staining was performed as described by Wang et al. (2012). Briefly, two 582
independent lines of the UGT76D1pro:GUS transgenic plants were first fixed with 583
90% acetone on ice for 20 min, and then washed with staining buffer (50 mM sodium 584
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

21
phosphate, pH 7.2, 0.2 mM K 3 Fe(CN) 6 , 0.2 mM K 4 Fe(CN) 6, and 0.2% Triton 585
X-100) without X-Gluc (5-bromo-4-chloro-3-indolyl-β-D-galactopyr-anoside) twice 586
before adding staining buffer with X-Gluc (final concentration of 2 mM) and 587
incubation overnight at 37°C. Two independent transgenic lines showing similar 588
results were used as sources for the photos. 589
Plasmid construction and creation of transgenic Arabidopsis lines 590
Full-length cDNA of UGT76D1 was obtained from total RNA of Arabidopsis 591
Col-0 by reverse transcription PCR using specific primers (Table S4). The cDNA was 592
inserted into plasmid pBluescript SK, and then transferred to the plant expression 593
vector pBI121 to replace the GUS gene. The constructed plant expression plasmids 594
were used to transform Arabidopsis, using the floral dip method (Clough and Bent, 595
1998). The transgenic plants were selected for kanamycin resistance. After selection 596
for three generations, the homozygous UGT76D1 overexpression lines were obtained. 597
Two independent overexpression lines (OE-17 and OE-32), with high expression 598
levels of UGT76D1 and similar phenotypes, were selected for further study. 599
UGT76D1 mutant lines were obtained, using the CRISPR/Cas9 system (Feng et al., 600
2013; Li et al., 2013). The 19-bp sequence (5′-GCGTCCTACCTTTCTTCCC-3′), 601
with high specificity to the UGT76D1 gene, was selected as the targeting sequence for 602
UGT76D1. Two knockout mutant lines (ko-33 and ko-36) with different deletions in 603
the 5′ coding region of UGT76D1 were selected for further study. 604
DAB and trypan blue staining 605
Approximately four-week-old plants were used in these staining experiments. 606
For detecting H2O2, DAB staining was performed as described by 607
Thordal-Christensen et al. (1997). Whole rosette leaves were immersed in a 1% 608
solution of 3,3'-diaminobenzidine (DAB) in Tris-HCl buffer (pH 6.5) for 16 h at 25°C 609
under dark conditions, and then cleared by boiling destaining solution (ethanol:acetic 610
acid:glycerol = 3:1:1) for 10 min. H2O2 accumulation was visualized by the 611
yellow-brown color. 612
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

22
For detecting cell death, trypan blue staining was carried out according to the 613
method described previously (Yang et al., 2015). Plant leaves were boiled 1 min in 614
Trypan Blue solution (10 mL of lactic acid, 10 mL of glycerol, 10 g of phenol, and 10 615
mg of Trypan Blue, dissolved in 10 mL of distilled water) diluted in 1 volume of 616
100% ethanol. Leaves were then bleached in choral hydrate (2.5 g mL−1
) overnight 617
and conserved in 50% glycerol. 618
Protein purification and enzyme assays 619
Full-length cDNA of UGT76D1 was inserted into plasmid pBluescript SK, and 620
then transferred to the prokaryotic expression vector pGEX-2T. The UGT76D1-GST 621
fusion protein was expressed in E. coli XL1-Blue and then purified using the method 622
described previously (Hou et al., 2004). The enzyme assays were performed at 30°C 623
using methods described previously (Lim et al., 2001) with modifications. About 2μg 624
of UGT76D1 fusion protein were added to 200 μL enzyme reaction mixture. The 625
reaction mixture contained 100 mM Tris-HCl (pH 7.6), 14 mM 2-mercaptoethanol, 5 626
mM UDP-glucose or UDP-xylose, 2.5 mM MgSO4, 10 mM KCl and 1mM substrate. 627
The tested substrates in this study are listed in Table S2. The reaction products were 628
analyzed by HPLC, and confirmed by LC-MS. 629
Measurements of SA, DHBA, and DHBA glycosides 630
Four-week-old plants were harvested for the analysis of SA and DHBA glycoside 631
concentrations. Free SA and total SA were extracted and measured by the methods 632
described previously (Nawrath and Metraux, 1999). The internal reference, 633
2,6-dichloroisonicotinic acid (INA), was used to estimate the loss ratesduring 634
extraction. SA was detected with a 295-nm excitation wavelength and a 405-nm 635
emission wavelength, using a fluorescence detector. DHBA glycosides were extracted 636
and measured by the protocol described previously (Zhang et al., 2012; Zhang et al., 637
2013), with modifications. Briefly, DHBA glycosides were extracted from leaves 638
homogenized in 80% (v/v) methanol overnight at 4°C. Caffeic acid and ferulic acid 639
were used as the internal references to estimate the loss rates during extraction. Target 640
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

23
compounds were analyzed by HPLC (Shimadzu, Kyoto, Japan) on a C18 columnat a 641
flow rate of 0.7 ml min-1
. The mobile phases were composed of sodium acetate (50 642
mM, pH 5.5) and MeOH. DHBA glycosides were detected using a UV detector at 310 643
nm wavelength. The identity of these compounds following HPLC profiling by 644
retention time were confirmed by liquid chromatography-mass spectrometer (LC-MS) 645
(Thermo Scientific, Waltham, MA, USA), and UV absorbance. 2,3-DHBA and 646
2,5-DHBA were extracted and measured as previously described (Zhang et al., 2017). 647
To quantify SA concentration, a SA standard curve was constructed. The peak 648
area of SA showed a close linear relationship with concentration in the standard curve. 649
Because the DHBA glycosides are not commercially available, the relative 650
concentrations of DHBA glycosides were analyzed in this study. The peak areas of 651
DHBA glycosides from the overexpression lines and knockout mutant lines were 652
determined and compared to the corresponding values from the WT. 653
RNA extraction and gene expression analysis 654
For analysis of gene expression, the rosette leaves from four-week-old plants (6 655
plants grown in different pots at the same time for each technical replicates) were 656
collected for RNA extraction. Total RNA was extracted using the Trizol reagent 657
(TaKaRa, Tokyo, Japan). Reverse transcription was performed using Prime Script RT 658
reagent kit (TaKaRa, Tokyo, Japan). RT-qPCR were performed with a Bio-Rad 659
thermal-cycling system (CFX Connect, Hercules, CA, USA), using a SYBR Green 660
PCR Master Mix kit (TaKaRa, Tokyo, Japan). ACTIN2 was used as the internal 661
control. Results were normalized to the reference gene ACTIN2 using the ΔΔCt 662
method. Primers used in RT-qPCR are listed in Supplemental Table S4. 663
Pst DC3000 cultivation and challenge assays 664
Pst DC3000 was used for most of the pathogen assays, while Pst DC3000 665
(avrRpt2) were used for HR assays. Both bacterial strains were cultured in King’s B 666
liquid medium (King et al., 1954) at 30°C, washed twice in 10 mM MgCl2, and 667
resuspended at OD600 = 0.001 (for Pst DC3000) or 0.02 [for Pst DC3000 (avrRpt2)]. 668
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

24
The resulting suspension solution was injected into the leaves of four-week-old plants; 669
MgCl2 (10 mM) was used as the negative control. Inoculated leaves were collected at 670
different time points to determine the expression of SA-related genes in response to 671
pathogen challenge. SA levels were determined at 48h after injection. 672
Statistical analysis 673
All experiments were carried out with at least three independent biological 674
replicates, with each measurement also being carried out in triplicate. Data are 675
presented as the mean ± SD. Data were statistically analyzed using Student’s t-test. 676
Asterisks indicate significant differences relative to the wild type or control (*P < 677
0.05, **P < 0.01). 678
679
ACCESSION NUMBERS 680
Sequence data from this article can be found in the GenBank/EMBL data libraries 681
under accession numbers UGT76D1 (At2g26480), PR1 (At2g14610), PR2 682
(At3g57260), EDS1 (At3g48090), PAD4 (At3g52430), ICS1 (At1g74710). 683
684
Supplemental Data 685
Supplemental Figure S1. Generation and identification of UGT76D1 mutants 686
and overexpression lines. 687
688
Supplemental Figure S2. Exaggerated necrotic lesion phenotype of 689
UGT76D1 overexpression lines under short day conditions (8 h light/16 h 690
dark). 691
Supplemental Figure S3. The leaf senescence phenotype of UGT76D1 692
overexpression and mutant lines. 693
694
Supplemental Figure S4. LC-MS analyses confirmed the glycoside formation 695
from UGT76D1 catalyzed 2,3-DHBA and 2,5-DHBA under positive ion mode. 696
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

25
697
Supplemental Figure S5. Analyses of DHBA glucoside in UGT76D1 698
overexpression and mutant lines after application of DHBA. 699
700
Supplemental Figure S6. LC-MS analyses of free forms of 2,3-DHBA and 701
2,5-DHBA in UGT76D1 overexpression and mutant lines with or without 702
inoculation of Pst DC3000. 703
704
Supplemental Table S1. Co-segregation data of LMM phenotype and 705
kanamycin resistance marker in T2 generation 706
707
Supplemental Table S2. Candidate substrates detected for UGT76D1 708
enzymatic activity 709
710
Supplemental Table S3. Glucosyltransferase activity of UGT76D1 and 711
UGT89A2 towards 2,3-DHBA and 2,5-DHBA with different sugar donors 712
713
Supplemental Table S4. Primers used in this study. 714
715
ACKNOWLEDGMENTS 716
We thank Dr. Shuhua Yang (China Agricultural University) for providing the 717
bacteria strains Pst DC3000, Pst DC3000 (avrRpt2) and the sid2 mutant seeds. We 718
also thank Dr. Kewei Zhang (Zhejiang Normal University, China) for providing the 719
s3h mutant and s3h s5h double mutant seeds, Dr. Jia Li (Lanzhou University, China) 720
for providing the NahG transgenic seeds. This research was supported by National 721
Natural Science Foundation of China (no. 91217301, 31570299, 31770313 to B.K.H). 722
723
724
725
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

26
FIGURE LEGENDS 726
Figure 1. Expression of UGT76D1 is induced by SA and Pst DC3000. A and B, 727
RT-qPCR analysis of UGT76D1 expression induction by SA and Pst DC3000 728
treatment. Mock treatment in A is DMSO solvent. Mock treatment in B is 10 729
mM MgCl2 suspension buffer. The value of WT with mock treatment for 0 h 730
was set at 1. Data are means ± SD of three biological replicates (**P < 0.01, *P 731
< 0.05, Student’s t-test). C and D, GUS staining of UGT76D1Pro:GUS 732
expression following Pst DC3000 (C) and MgCl2 (mock, D) inoculation. Bars = 733
1 mm (upper panels) and 200 μm (lower panels). 734
735
Figure 2. Analysis of UGT76D1 mutant and overexpression lines. A, 736
Phenotype of four-week-old wild-type (WT), UGT76D1 overexpression lines 737
(OE-17 and OE-32) and UGT76D1 mutant lines (ko-33 and ko-36) grown 738
under 22°C and long day conditions (16 h light/8 h dark). Right panel is a 739
magnified photograph showing the spontaneous lesion mimic phenotype in 740
rosette leaves of overexpression lines (Bar = 1cm for both panels). B, Trypan 741
blue staining showing cell death in UGT76D1 overexpression and mutant lines. 742
Four-week-old plants grown under long-day condition were used for the 743
staining assay. The bottom panels are the magnified photographs of top panels. 744
Bars = 1 cm (top panels) and 100 μm (bottom panels). C, DAB staining 745
showing the H2O2 content of UGT76D1 overexpression and mutant lines. 746
Four-week-old plants grown under long-day condition were used for the 747
staining assay. The bottom panels are magnified photographs of top panels. 748
Bars = 1 cm (top panels) and 100 μm (bottom panels). D and E, RT-qPCR 749
analysis of the expression of defense response genes PR1 and PR2 in 750
four-week-old UGT76D1 overexpression lines and mutant lines. The value of 751
WT plants was set at 1.0. Data are means ± SD of three biological replicates 752
(**P < 0.01, *P < 0.05, Student’s t-test). F and G, Quantification of free SA and 753
total SA levels in UGT76D1 overexpression and mutant lines. Leaves of 754
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

27
four-week-old plants grown under long day conditions at 22°C were collected 755
and used for the measurements. Data are means ± SD of three biological 756
replicates (**P < 0.01, *P < 0.05, Student’s t-test). 757
758
Figure 3. Hypersensitive response phenotype of UGT76D1 mutant lines 759
following avirulent pathogen induction. A and B, HR phenotypes (A) and 760
Trypan Blue staining showing cell death (B) of wild-type (WT) and UGT76D1 761
mutant (ko-33 and ko-36) leaves following inoculation with Pst DC3000 762
(avrRpt2) (OD600 = 0.02). MgCl2 treatment was used as mock control. At least 763
36 leaves were assessed for each genotype during each experiment, with a 764
similar phenotype displayed in most. Representative leaves are shown at 24 h 765
and 48 h post inoculation. Bars = 1cm. 766
767
Figure 4. SA is required for UGT76D1-activated lesion mimic phenotype. A, 768
Phenotypes of four-week-old wild-type (WT), UGT76D1 overexpression 769
(OE-17 and OE-32), NahG-expressing (NahG), and UGT76D1 770
overexpression/NahG hybrid (OE-17 NahG and OE-32 NahG) plants. Bar = 1 771
cm. B, RT-PCR analysis of the expression of UGT76D1 in UGT76D1 772
overexpression/NahG hybrid lines alongside control lines described in A. 773
ACTIN2 was used as the internal control. C, Trypan blue staining (top panel) 774
showing the cell death level and DAB staining (bottom panel) showing the 775
H2O2 contents in leaves from four-week-old plants of UGT76D1 776
overexpression/NahG hybrid lines alongside control lines described in A. Bars 777
= 1 cm. D and E, RT-qPCR analysis of PR1 and PR2 expression in UGT76D1 778
overexpression/NahG hybrid lines alongside control lines described in A. The 779
value of WT plants was set at 1. Data represent mean ± SD of three biological 780
replicates (**P < 0.01, *P < 0.05, Student’s t-test). F and G, Free and total SA 781
contents in UGT76D1 overexpression/NahG hybrid lines alongside control 782
lines described in A. Data are means ± SD of three biological replicates. 783
(*P<0.05, Student’s t-test). ND: not detectable. 784
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

28
785
Figure 5. UGT76D1 catalyzes the in vitro glycosylation of 2,3-DHBA and 786
2,5-DHBA, but not SA. A, SDS-PAGE detection of the purified recombinant 787
UGT76D1-GST fusion protein. Proteins were visualized by Coomassie Blue 788
staining. GST: glutathione-s-transferase; UGT76D1: UGT76D1 fusion protein. 789
B, HPLC analysis of SA glycosylation by UGT76D1. C and D, HPLC analysis 790
of 2,3-DHBA and 2,5DHBA glycosylation by UGT76D1 G: UDP-glucose; X: 791
UDP-xylose. GST protein was used as the negative control. 792
793
Figure 6. The relative levels of sugar conjugates of 2,3-DHBA and 2,5-DHBA 794
in UGT76D1 overexpression and mutant lines. A and B, HPLC profiling (A) and 795
the quantitation (B) of DHBA sugar conjugates in wild type (WT), UGT76D1 796
overexpression lines (OE-17 and OE-32), and UGT76D1 mutant lines (ko-33 797
and ko-36) in the absence of pathogen exposure. Data were normalized to the 798
corresponding DHBA glycoside in WT to account for the changes in UGT76D1 799
overexpression and mutant lines. C and D, HPLC profiling and the quantitation 800
of DHBA sugar conjugates in wild type and UGT76D1 mutant lines following 801
Pst DC3000 (DC) or 10 mM MgCl2 (Mock) treatment for 48 h. Data were 802
normalized to the corresponding DHBA glycoside in WT treated with Pst 803
DC3000 to account for the changes in UGT76D1 mutant lines. The DHBA 804
sugar conjugates formed by UGT76D1 catalysis in vitro were used as the 805
glycoside standards. Peak1: 2,5-DHBA glucosides; Peak2: 2,3-DHBA 806
glucosides; Peak3: 2,5-DHBA xlyoside; Peak4: 2,3-DHBA xylosides; Peak5: 807
internal reference (caffeic acid). 2,3DHBA-G: 2,3-DHBA glucosides; 808
2,3DHBA-X: 2,3DHBA xyloside; 2,5DHBA-G: 2,5-DHBA glucosides; 809
2,5DHBA-X: 2,5-DHBA xlyoside. Data are means ± SD of three biological 810
replicates (**P < 0.01, *P < 0.05, Student’s t-test). 811
812
Figure 7. UGT76D1-activated immune response depends on DHBA 813
glycosylation. A, Phenotypes of four-week-old wild-type (WT), UGT76D1 814
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

29
overexpression (OE-17 and OE-32), s3h mutant (s3h), and UGT76D1 815
overexpression/s3h hybrid (OE-17 s3h and OE-32 s3h) plants grown under 816
long day conditions at 22°C. Bar = 1 cm. B, DAB (top row panels) and Trypan 817
blue (bottom row panels) staining showing levels of H2O2 and cell death, 818
respectively, in leaves from four-week-old plants of UGT76D1 819
overexpression/s3h hybrid lines alongside control lines described in A. Bars = 820
100 μm. C, RT-PCR analysis of UGT76D1 expression in UGT76D1 821
overexpression/s3h hybrid lines alongside control lines described in A. 822
ACTIN2 was used as the internal control. D, RT-qPCR analysis of relative PR1 823
and PR2 expression in UGT76D1 overexpression/s3h hybrid lines alongside 824
control lines described in A. The value of WT plants was set at 1. Data are 825
means ± SD of three biological replicates (**P < 0.01, *P < 0.05, Student’s 826
t-test). E, The free and total SA levels in UGT76D1 overexpression/s3h hybrid 827
lines alongside control lines described in A. Data are means ± SD of three 828
biological replicates (**P < 0.01, *P < 0.05, Student’s t-test). F, Phenotypes of 829
transgenic plants overexpressing UGT76D1 in a WT or s3h s5h double mutant 830
background. The right panel shows the enlarged leaf phenotype. Leaf lesions 831
are indicated with red arrowheads. Bars = 1 cm (left panel) and 0.5 cm (right 832
panel). 833
834
Figure 8. The immune response and SA biosynthesis in UGT76D1 835
overexpression and mutant lines following treatment with Pst DC3000. A and B, 836
Relative PR1 and PR2 expression in three-week-old wild-type (WT) and 837
UGT76D1 overexpression (OE) and mutant (ko) plants following Pst DC3000 838
(DC) or MgCl2 (Mock) treatment. For either treatment, expression levels in WT 839
at 0 h were set at 1. C and D, Free and total SA levels in three-week-old 840
UGT76D1 overexpression (OE-17 and OE-32) and mutant (ko-33 and ko-36) 841
lines following treatment with Pst DC3000 or MgCl2 (Mock) for 48 h. E, 842
RT-qPCR analysis of EDS1, PAD4, and ICS1 expression in UGT76D1 843
overexpression and mutant lines without any experimental treatment. Gene 844
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

30
expression levels in WT were set at 1.0. F, RT-qPCR analysis of EDS1, PAD4, 845
and ICS1 expression in UGT76D1 mutant lines following treatment with Pst 846
DC3000 for 24 h and 48 h. Gene expression levels in WT following 24 h Pst 847
DC3000 treatment were set at 1.0. A to F, Data are means ± SD of three 848
biological replicates (**P < 0.01, *P < 0.05, Student’s t-test). 849
850
Figure 9. UGT76D1-activated immune response depends on SA biosynthesis. 851
A, Phenotypes of four-week-old wild-type (WT), UGT76D1 overexpression 852
(OE-17 and OE-32), sid2 mutant (sid2-1), and UGT76D1 overexpression/sid2 853
hybrid (OE-17 sid2-1 and OE-32 sid2-1) plants. Bar = 1 cm. B, RT-PCR 854
analysis of UGT76D1 expression in UGT76D1 overexpression/sid2 hybrid 855
lines alongside control lines described in A. ACTIN2 was used as the internal 856
control. C, Quantification of free SA and total SA levels in UGT76D1 857
overexpression/sid2 hybrid lines alongside control lines described in A. Leaves 858
of four-week-old plants grown under long day conditions at 22°C were 859
collected and used for the measurements. Data are means ± SD of three 860
biological replicates (Student’s t-test, *P<0.05, * * P <0.01). 861
862
Figure 10. Proposed scheme for the regulation of SA homeostasis and plant 863
defense responses by UGT76D1. When plants are challenged by pathogens, 864
expression of UGT76D1 is induced to a high level, and the formation of DHBA 865
glycosides is accelerated. At the same time, DHBA glycosylation may activate 866
SA synthase and then increase SA biosynthesis through an unknown positive 867
feedback loop, possibly by increasing the metabolic flux towards SA and the 868
DHBAs. High levels of SA accumulation lead to an oxidative burst and PR 869
gene expression, and ultimately lead to PCD and increased plant resistance to 870
pathogens. Considering the xylosyltransferase activity of UGT89A2 towards 871
DHBAs, we cannot exclude the role of UGT89A2 in this immune-priming 872
process. 873
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

31
LITERATURE CITED 874
Aarts N, Metz M, Holub E, Staskawicz BJ, Daniels MJ, Parker JE (1998) 875
Different requirements for EDS1 and NDR1 by disease resistance genes define 876
at least two R gene-mediated signaling pathways in Arabidopsis. Proceedings 877
of the National Academy of Sciences 95: 10306-10311 878
Alvarez ME (2000) Salicylic acid in the machinery of hypersensitive cell death and 879
disease resistance. In Programmed Cell Death in Higher Plants. Springer, pp 880
185-198 881
Bartsch M, Bednarek P, Vivancos PD, Schneider B, von Roepenack-Lahaye E, 882
Foyer CH, Kombrink E, Scheel D, Parker JE (2010) Accumulation of 883
isochorismate-derived 2, 3-dihydroxybenzoic 3-O-β-D-xyloside in 884
Arabidopsis resistance to pathogens and ageing of leaves. Journal of 885
Biological Chemistry 285: 25654-25665 886
Bellés JM, Garro R, Fayos J, Navarro P, Primo J, Conejero V (1999) Gentisic 887
acid as a pathogen-inducible signal, additional to salicylic acid for activation 888
of plant defenses in tomato. Molecular plant-microbe interactions 12: 227-235 889
Bruggeman Q, Raynaud C, Benhamed M, Delarue M (2015) To die or not to die? 890
Lessons from lesion mimic mutants. Frontiers in plant science 6 891
Chen HY, Li X (2017) Identification of a residue responsible for UDP-sugar donor 892
selectivity of a dihydroxybenzoic acid glycosyltransferase from Arabidopsis 893
natural accessions. Plant J 89: 195-203 894
Chen Y, Shen H, Wang M, Li Q, He Z (2013) Salicyloyl-aspartate synthesized by 895
the acetyl-amido synthetase GH3. 5 is a potential activator of plant immunity 896
in Arabidopsis. Acta Biochim Biophys Sin 45: 827-836 897
Chong J, Baltz R, Schmitt C, Beffa R, Fritig B, Saindrenan P (2002) 898
Downregulation of a pathogen-responsive tobacco UDP-Glc:phenylpropanoid 899
glucosyltransferase reduces scopoletin glucoside accumulation, enhances 900
oxidative stress, and weakens virus resistance. Plant Cell 14: 1093-1107 901
Chong J, Pierrel M-A, Atanassova R, Werck-Reichhart D, Fritig B, Saindrenan P 902
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

32
(2001) Free and conjugated benzoic acid in tobacco plants and cell cultures. 903
Induced accumulation upon elicitation of defense responses and role as 904
salicylic acid precursors. Plant Physiology 125: 318-328 905
Clough SJ, Bent AF (1998) Floral dip: a simplified method forAgrobacterium‐906
mediated transformation ofArabidopsis thaliana. The plant journal 16: 907
735-743 908
Coll N, Epple P, Dangl J (2011) Programmed cell death in the plant immune system. 909
Cell death and differentiation 18: 1247 910
Coll NS, Vercammen D, Smidler A, Clover C, Van Breusegem F, Dangl JL, Epple 911
P (2010) Arabidopsis type I metacaspases control cell death. Science 330: 912
1393-1397 913
Dean JV, Delaney SP (2008) Metabolism of salicylic acid in wild‐type, ugt74f1 and 914
ugt74f2 glucosyltransferase mutants of Arabidopsis thaliana. Physiologia 915
plantarum 132: 417-425 916
Delaney TP, Uknes S, Vernooij B, Friedrich L (1994) A central role of salicylic acid 917
in plant disease resistance. Science 266: 1247 918
Dempsey DMA, Vlot AC, Wildermuth MC, Klessig DF (2011) Salicylic acid 919
biosynthesis and metabolism. The Arabidopsis Book: e0156 920
Fayos J, Bellés JM, Lopez-Gresa MP, Primo J, Conejero V (2006) Induction of 921
gentisic acid 5-O-β-D-xylopyranoside in tomato and cucumber plants infected 922
by different pathogens. Phytochemistry 67: 142-148 923
Feng Z, Zhang B, Ding W, Liu X, Yang D-L, Wei P, Cao F, Zhu S, Zhang F, Mao 924
Y (2013) Efficient genome editing in plants using a CRISPR/Cas system. Cell 925
research 23: 1229 926
Feys BJ, Moisan LJ, Newman MA, Parker JE (2001) Direct interaction between 927
the Arabidopsis disease resistance signaling proteins, EDS1 and PAD4. The 928
EMBO journal 20: 5400-5411 929
Feys BJ, Wiermer M, Bhat RA, Moisan LJ, Medina-Escobar N, Neu C, Cabral A, 930
Parker JE (2005) Arabidopsis senescence-associated gene101 stabilizes and 931
signals within an enhanced disease susceptibility1 complex in plant innate 932
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

33
immunity. The Plant Cell 17: 2601-2613 933
Fu ZQ, Dong X (2013) Systemic acquired resistance: turning local infection into 934
global defense. Annual review of plant biology 64: 839-863 935
Greenberg JT, Silverman FP, Liang H (2000) Uncoupling salicylic acid-dependent 936
cell death and defense-related responses from disease resistance in the 937
Arabidopsis mutant acd5. Genetics 156: 341-350 938
Heath MC (2000) Hypersensitive response-related death. In Programmed cell death 939
in higher plants. Springer, pp 77-90 940
Hou B, Lim E-K, Higgins GS, Bowles DJ (2004) N-glucosylation of cytokinins by 941
glycosyltransferases of Arabidopsis thaliana. Journal of Biological Chemistry 942
279: 47822-47832 943
Ibrahim RK, Towers G (1959) Conversion of salicylic acid to gentisic acid and 944
o-pyrocatechuic acid, all labelled with carbon-14, in plants. Nature 184: 945
1803-1803 946
Jabs T (1999) Reactive oxygen intermediates as mediators of programmed cell death 947
in plants and animals. Biochemical pharmacology 57: 231-245 948
Jackson RG, Kowalczyk M, Li Y, Higgins G, Ross J, Sandberg G, Bowles DJ 949
(2002) Overexpression of an Arabidopsis gene encoding a glucosyltransferase 950
of indole-3-acetic acid: phenotypic characterization of transgenic lines. Plant J 951
32:573–583 952
Jones P, Messner B, Nakajima J, Schaffner AR, Saito K (2003) UGT73C6 and 953
UGT78D1, glycosyltransferases involved in glycoside biosynthesis in 954
Arabidopsis thaliana. J Biol Chem. 278(45):43910-43918 955
King EO, Ward MK, Raney DE (1954) Two simple media for the demonstration of 956
pyocyanin and fluorescin. The journal of laboratory and clinical medicine 44: 957
301-307 958
Li J-F, Norville JE, Aach J, McCormack M, Zhang D, Bush J, Church GM, 959
Sheen J (2013) Multiplex and homologous recombination-mediated genome 960
editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. 961
Nature biotechnology 31: 688-691 962
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

34
Li X, Svedin E, Mo H, Atwell S, Dilkes BP, Chapple C (2014) Exploiting natural 963
variation of secondary metabolism identifies a gene controlling the 964
glycosylation diversity of dihydroxybenzoic acids in Arabidopsis thaliana. 965
Genetics 198: 1267-1276 966
Lim EK, Doucet CJ, Li Y, Elias L, Worrall D, Spencer SP, Ross J, Bowles DJ 967
(2002) The activity of Arabidopsis glycosyltransferases toward salicylic acid, 968
4-hydroxybenzoic acid, and other benzoates. J Biol Chem 277: 586-592 969
Lim EK, Li Y, Parr A, Jackson R, Ashford DA, Bowles DJ (2001) Identification of 970
glucosyltransferase genes involved in sinapate metabolism and lignin 971
synthesis in Arabidopsis. J Biol Chem 276: 4344-4349 972
Lorrain S, Vailleau F, Balagué C, Roby D (2003) Lesion mimic mutants: keys for 973
deciphering cell death and defense pathways in plants? Trends in plant science 974
8: 263-271 975
Love AJ, Milner JJ, Sadanandom A (2008) Timing is everything: regulatory 976
overlap in plant cell death. Trends in plant science 13: 589-595 977
Malik B, Pirzadah TB, Tahir I, Rehman RU, Hakeem KR, Abdin M (2014) Plant 978
signaling: response to reactive oxygen species. In Plant signaling: 979
Understanding the molecular crosstalk. Springer, pp 1-38 980
Nawrath C, Metraux JP (1999) Salicylic acid induction-deficient mutants of 981
Arabidopsis express PR-2 and PR-5 and accumulate high levels of camalexin 982
after pathogen inoculation. Plant Cell 11: 1393-1404 983
Noutoshi Y, Okazaki M, Kida T, Nishina Y, Morishita Y, Ogawa T, Suzuki H, 984
Shibata D, Jikumaru Y, Hanada A, Kamiya Y, Shirasu K (2012) Novel 985
plant immune-priming compounds identified via high-throughput chemical 986
screening target salicylic acid glucosyltransferases in Arabidopsis. Plant Cell 987
24: 3795-3804 988
Rate DN, Cuenca JV, Bowman GR, Guttman DS, Greenberg JT (1999) The 989
gain-of-function Arabidopsis acd6 mutant reveals novel regulation and 990
function of the salicylic acid signaling pathway in controlling cell death, 991
defenses, and cell growth. The Plant Cell 11: 1695-1708 992
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

35
Rivas-San Vicente M, Plasencia J (2011) Salicylic acid beyond defence: its role in 993
plant growth and development. Journal of experimental botany 62: 3321-3338 994
Ryals J, Uknes S, Ward E (1994) Systemic acquired resistance. Plant Physiology 995
104: 1109Shah J (2003) The salicylic acid loop in plant defense. Current 996
opinion in plant biology 6: 365-371 997
Shah J (2003) The salicylic acid loop in plant defense. Current opinion in plant 998
biology 6: 365-371 999
Shah J, Kachroo P, Nandi A, Klessig DF (2001) A recessive mutation in the 1000
Arabidopsis SSI2 gene confers SA-and NPR1-independent expression of PR 1001
genes and resistance against bacterial and oomycete pathogens. The Plant 1002
Journal 25: 563-574 1003
Shirano Y, Kachroo P, Shah J, Klessig DF (2002) A gain-of-function mutation in an 1004
Arabidopsis Toll Interleukin1 Receptor–Nucleotide Binding 1005
Site–Leucine-Rich Repeat type R gene triggers defense responses and results 1006
in enhanced disease resistance. The Plant Cell 14: 3149-3162 1007
Straus MR, Rietz S, Ver Loren van Themaat E, Bartsch M, Parker JE (2010) 1008
Salicylic acid antagonism of EDS1-driven cell death is important for immune 1009
and oxidative stress responses in Arabidopsis. The Plant Journal 62: 628-640 1010
Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB (1997) Subcellular 1011
localization of H2O2 in plants. H2O2 accumulation in papillae and 1012
hypersensitive response during the barley—powdery mildew interaction. The 1013
Plant Journal 11: 1187-1194 1014
Tognetti VB, Van Aken O, Morreel K, Vandenbroucke K, van de Cotte B, De 1015
Clercq I, Chiwocha S, Fenske R, Prinsen E, Boerjan W, Genty B, Stubbs 1016
KA, Inze D, Van Breusegem F (2010) Perturbation of indole-3-butyric acid 1017
homeostasis by the UDP-glucosyltransferase UGT74E2 modulates 1018
Arabidopsis architecture and water stress tolerance. Plant Cell 22: 2660-2679 1019
Van Doorn WG (2011) Classes of programmed cell death in plants, compared to 1020
those in animals. Journal of Experimental Botany 62: 4749-4761Vlot AC, 1021
Dempsey DMA, Klessig DF (2009) Salicylic acid, a multifaceted hormone to 1022
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

36
combat disease. Annual review of phytopathology 47: 177-206 1023
Vlot AC, Dempsey DMA, Klessig DF (2009) Salicylic acid, a multifaceted hormone 1024
to combat disease. Annual review of phytopathology 47: 177-206 1025
von Saint Paul V, Zhang W, Kanawati B, Geist B, Faus-Kessler T, 1026
Schmitt-Kopplin P, Schaffner AR (2011) The Arabidopsis 1027
glucosyltransferase UGT76B1 conjugates isoleucic acid and modulates plant 1028
defense and senescence. Plant Cell 23: 4124-4145 1029
Walbot V, Hoisington DA, Neuffer M (1983) Disease lesion mimic mutations. In 1030
Genetic engineering of plants. Springer, pp 431-442 1031
Wang B, Jin SH, Hu HQ, Sun YG, Wang YW, Han P, Hou BK (2012) UGT87A2, 1032
an Arabidopsis glycosyltransferase, regulates flowering time via 1033
FLOWERING LOCUS C. New Phytologist 194: 666-675 1034
Wildermuth MC, Dewdney J, Wu G, Ausubel FM (2001) Isochorismate synthase is 1035
required to synthesize salicylic acid for plant defence. Nature 414: 562-565 1036
Yalpani N, León J, Lawton MA, Raskin I (1993) Pathway of salicylic acid 1037
biosynthesis in healthy and virus-inoculated tobacco. Plant physiology 103: 1038
315-321 1039
Yang L, Li B, Zheng X-y, Li J, Yang M, Dong X, He G, An C, Deng XW (2015) 1040
Salicylic acid biosynthesis is enhanced and contributes to increased biotrophic 1041
pathogen resistance in Arabidopsis hybrids. Nature communications 6: 7309 1042
Yang S, Hua J (2004) A haplotype-specific Resistance gene regulated by BONZAI1 1043
mediates temperature-dependent growth control in Arabidopsis. The Plant Cell 1044
16: 1060-1071 1045
Yoshioka K, Kachroo P, Tsui F, Sharma SB, Shah J, Klessig DF (2001) 1046
Environmentally sensitive, SA-dependent defense responses in the cpr22 1047
mutant of Arabidopsis. The Plant Journal 26: 447-459 1048
Zhang GZ, Jin SH, Jiang XY, Dong RR, Li P, Li YJ, Hou BK (2016) Ectopic 1049
expression of UGT75D1, a glycosyltransferase preferring indole-3-butyric 1050
acid, modulates cotyledon development and stress tolerance in seed 1051
germination of Arabidopsis thaliana. Plant Mol Biol 90: 77-93 1052
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

37
Zhang K, Bhuiya M-W, Pazo JR, Miao Y, Kim H, Ralph J, Liu C-J (2012) An 1053
engineered monolignol 4-O-methyltransferase depresses lignin biosynthesis 1054
and confers novel metabolic capability in Arabidopsis. The Plant Cell 24: 1055
3135-3152 1056
Zhang K, Halitschke R, Yin C, Liu CJ, Gan SS (2013) Salicylic acid 3-hydroxylase 1057
regulates Arabidopsis leaf longevity by mediating salicylic acid catabolism. 1058
Proc Natl Acad Sci U S A 110: 14807-14812 1059
Zhang Y, Xu S, Ding P, Wang D, Cheng YT, He J, Gao M, Xu F, Li Y, Zhu Z 1060
(2010) Control of salicylic acid synthesis and systemic acquired resistance by 1061
two members of a plant-specific family of transcription factors. Proceedings of 1062
the National Academy of Sciences 107: 18220-18225 1063
Zhang Y, Zhao L, Zhao J, Li Y, Wang J, Guo R, Gan S-S, Liu C-J, Zhang K 1064
(2017) S5H/DMR6 Encodes a Salicylic Acid 5-Hydroxylase That Fine-Tunes 1065
Salicylic Acid Homeostasis. Plant physiology 175: 1082–1093 1066
Zhou F, Menke FL, Yoshioka K, Moder W, Shirano Y, Klessig DF (2004) High 1067
humidity suppresses ssi4-mediated cell death and disease resistance upstream 1068
of MAP kinase activation, H2O2 production and defense gene expression. The 1069
Plant Journal 39: 920-932 1070
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
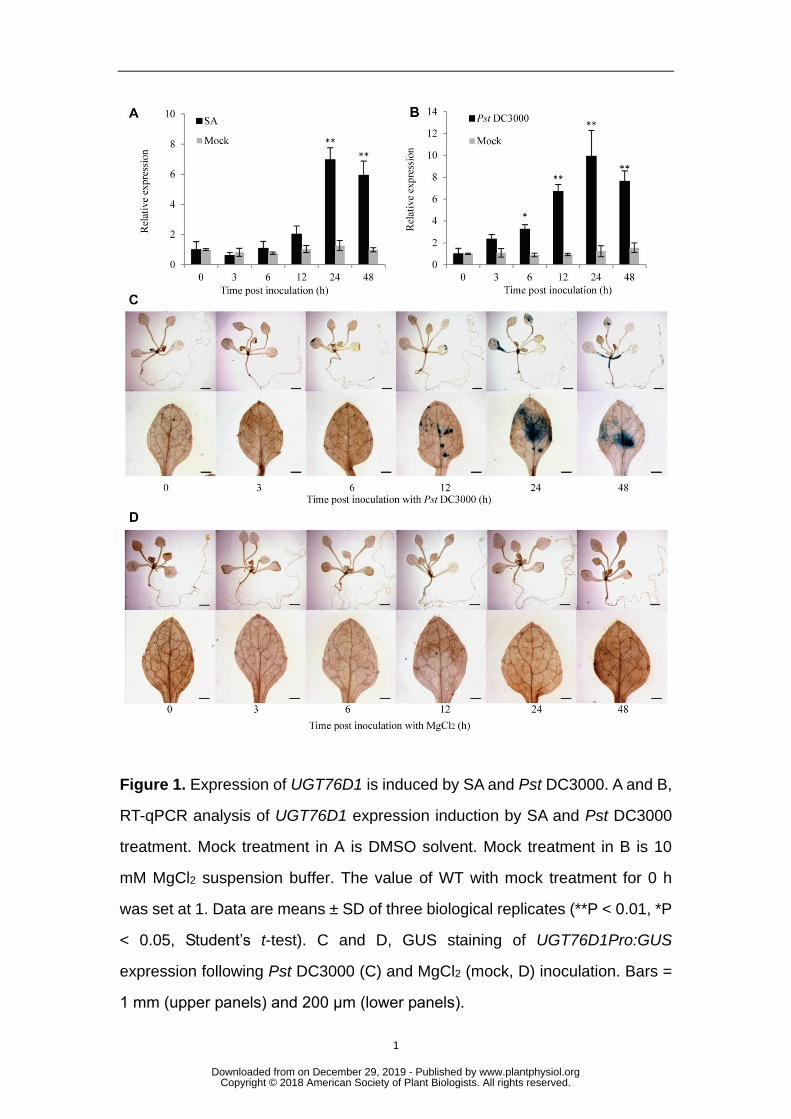
1
Figure 1. Expression of UGT76D1 is induced by SA and Pst DC3000. A and B,
RT-qPCR analysis of UGT76D1 expression induction by SA and Pst DC3000
treatment. Mock treatment in A is DMSO solvent. Mock treatment in B is 10
mM MgCl2 suspension buffer. The value of WT with mock treatment for 0 h
was set at 1. Data are means ± SD of three biological replicates (**P < 0.01, *P
< 0.05, Student’s t-test). C and D, GUS staining of UGT76D1Pro:GUS
expression following Pst DC3000 (C) and MgCl2 (mock, D) inoculation. Bars =
1 mm (upper panels) and 200 μm (lower panels).
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
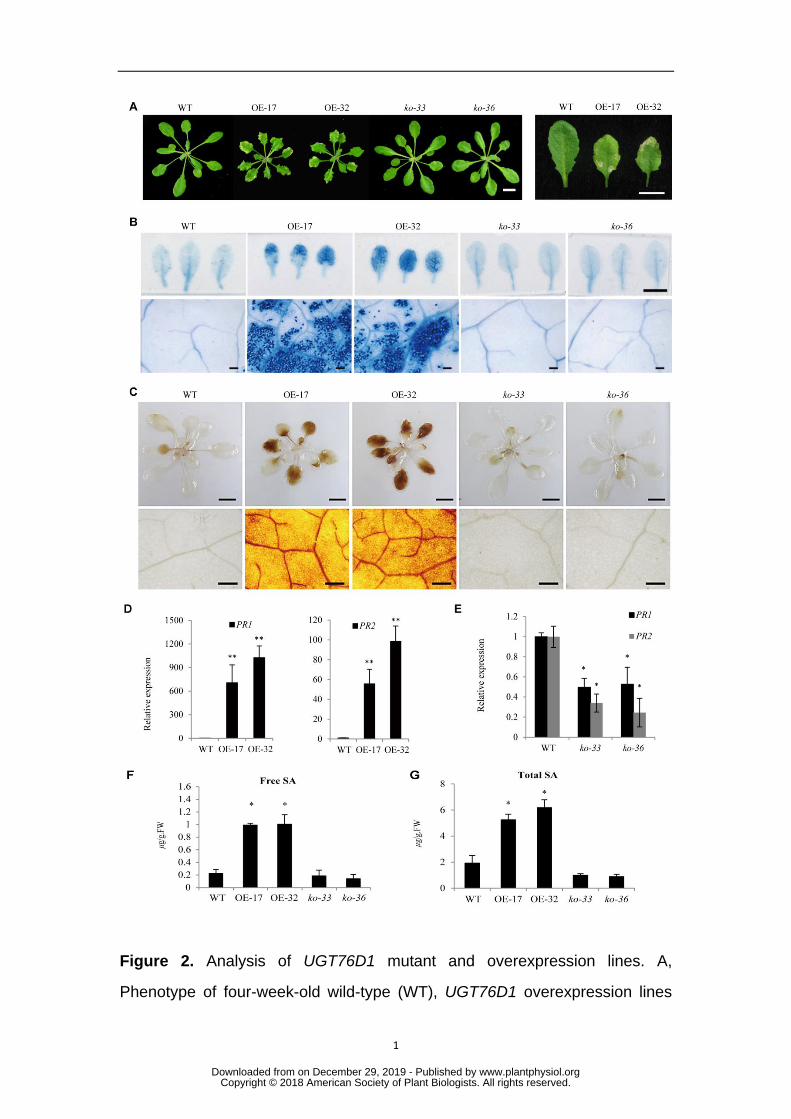
1
Figure 2. Analysis of UGT76D1 mutant and overexpression lines. A,
Phenotype of four-week-old wild-type (WT), UGT76D1 overexpression lines
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
(OE-17 and OE-32) and UGT76D1 mutant lines (ko-33 and ko-36) grown
under 22°C and long day conditions (16 h light/8 h dark). Right panel is a
magnified photograph showing the spontaneous lesion mimic phenotype in
rosette leaves of overexpression lines (Bar = 1cm for both panels). B, Trypan
blue staining showing cell death in UGT76D1 overexpression and mutant lines.
Four-week-old plants grown under long-day condition were used for the
staining assay. The bottom panels are the magnified photographs of top panels.
Bars = 1 cm (top panels) and 100 μm (bottom panels). C, DAB staining
showing the H2O2 content of UGT76D1 overexpression and mutant lines.
Four-week-old plants grown under long-day condition were used for the
staining assay. The bottom panels are magnified photographs of top panels.
Bars = 1 cm (top panels) and 100 μm (bottom panels). D and E, RT-qPCR
analysis of the expression of defense response genes PR1 and PR2 in
four-week-old UGT76D1 overexpression lines and mutant lines. The value of
WT plants was set at 1.0. Data are means ± SD of three biological replicates
(**P < 0.01, *P < 0.05, Student’s t-test). F and G, Quantification of free SA and
total SA levels in UGT76D1 overexpression and mutant lines. Leaves of
four-week-old plants grown under long day conditions at 22°C were collected
and used for the measurements. Data are means ± SD of three biological
replicates (**P < 0.01, *P < 0.05, Student’s t-test).
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
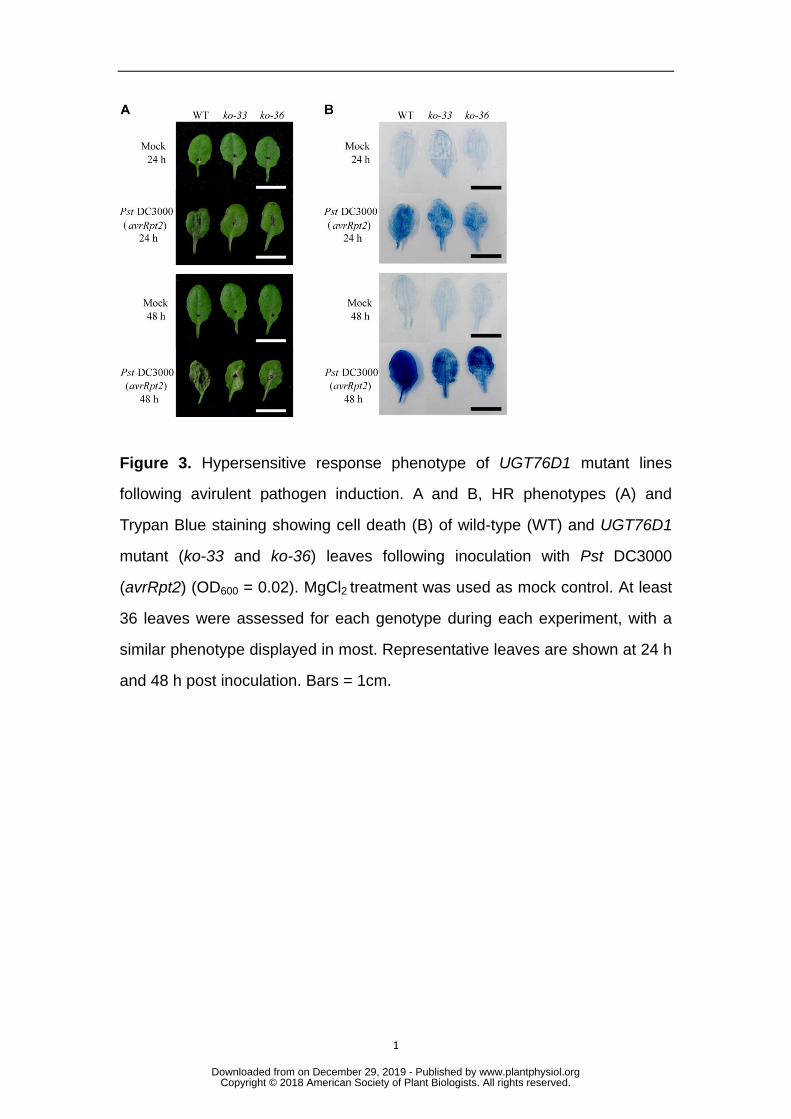
1
Figure 3. Hypersensitive response phenotype of UGT76D1 mutant lines
following avirulent pathogen induction. A and B, HR phenotypes (A) and
Trypan Blue staining showing cell death (B) of wild-type (WT) and UGT76D1
mutant (ko-33 and ko-36) leaves following inoculation with Pst DC3000
(avrRpt2) (OD600 = 0.02). MgCl2 treatment was used as mock control. At least
36 leaves were assessed for each genotype during each experiment, with a
similar phenotype displayed in most. Representative leaves are shown at 24 h
and 48 h post inoculation. Bars = 1cm.
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
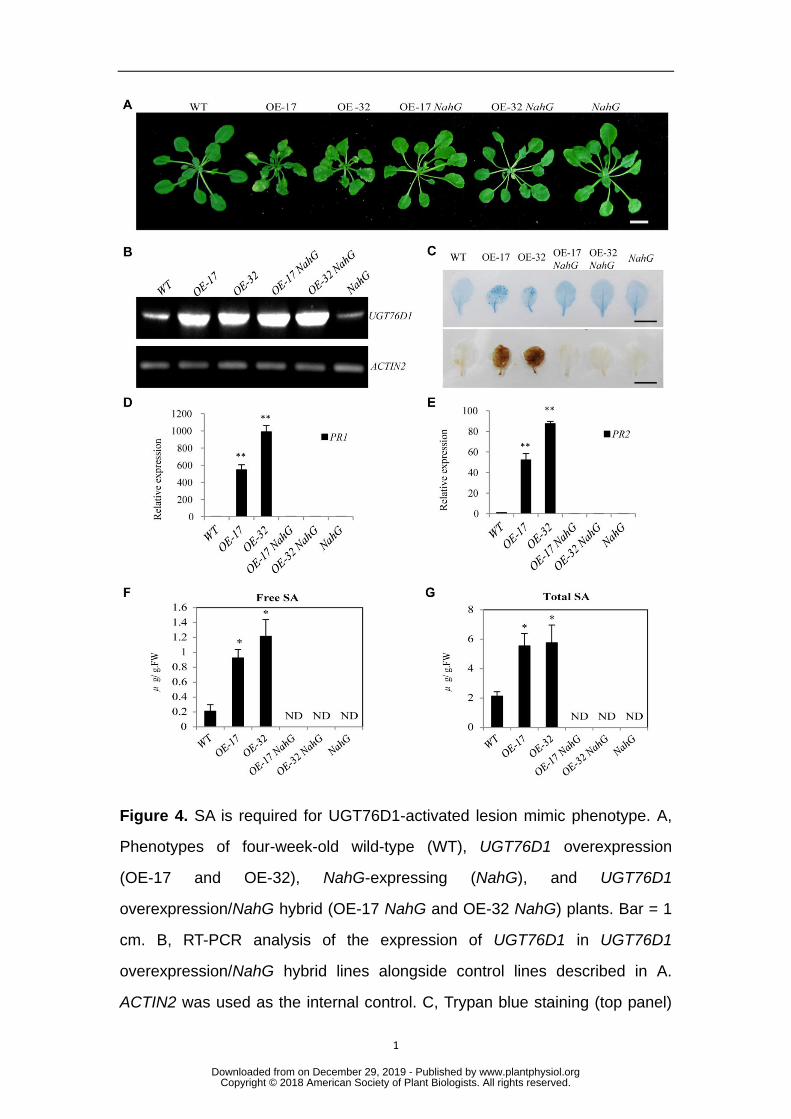
1
Figure 4. SA is required for UGT76D1-activated lesion mimic phenotype. A,
Phenotypes of four-week-old wild-type (WT), UGT76D1 overexpression
(OE-17 and OE-32), NahG-expressing (NahG), and UGT76D1
overexpression/NahG hybrid (OE-17 NahG and OE-32 NahG) plants. Bar = 1
cm. B, RT-PCR analysis of the expression of UGT76D1 in UGT76D1
overexpression/NahG hybrid lines alongside control lines described in A.
ACTIN2 was used as the internal control. C, Trypan blue staining (top panel)
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
showing the cell death level and DAB staining (bottom panel) showing the
H2O2 contents in leaves from four-week-old plants of UGT76D1
overexpression/NahG hybrid lines alongside control lines described in A. Bars
= 1 cm. D and E, RT-qPCR analysis of PR1 and PR2 expression in UGT76D1
overexpression/NahG hybrid lines alongside control lines described in A. The
value of WT plants was set at 1. Data represent mean ± SD of three biological
replicates (**P < 0.01, *P < 0.05, Student’s t-test). F and G, Free and total SA
contents in UGT76D1 overexpression/NahG hybrid lines alongside control
lines described in A. Data are means ± SD of three biological replicates.
(*P<0.05, Student’s t-test). ND: not detectable.
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
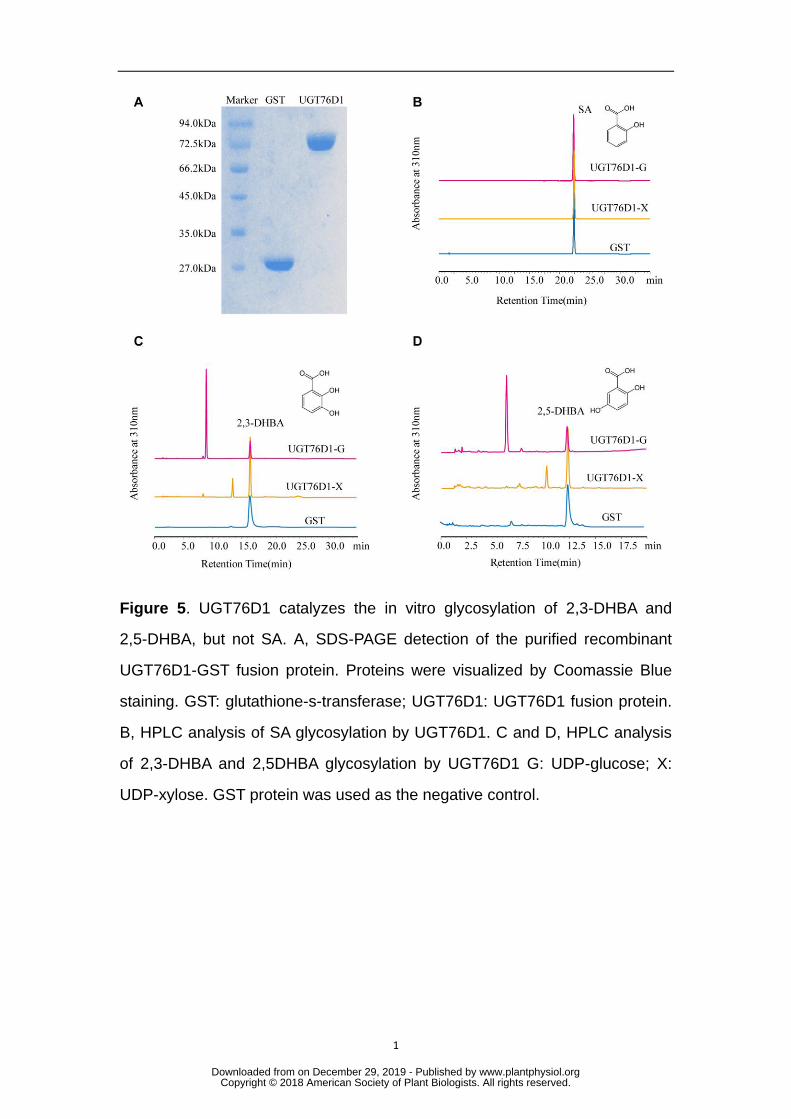
1
Figure 5. UGT76D1 catalyzes the in vitro glycosylation of 2,3-DHBA and
2,5-DHBA, but not SA. A, SDS-PAGE detection of the purified recombinant
UGT76D1-GST fusion protein. Proteins were visualized by Coomassie Blue
staining. GST: glutathione-s-transferase; UGT76D1: UGT76D1 fusion protein.
B, HPLC analysis of SA glycosylation by UGT76D1. C and D, HPLC analysis
of 2,3-DHBA and 2,5DHBA glycosylation by UGT76D1 G: UDP-glucose; X:
UDP-xylose. GST protein was used as the negative control.
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
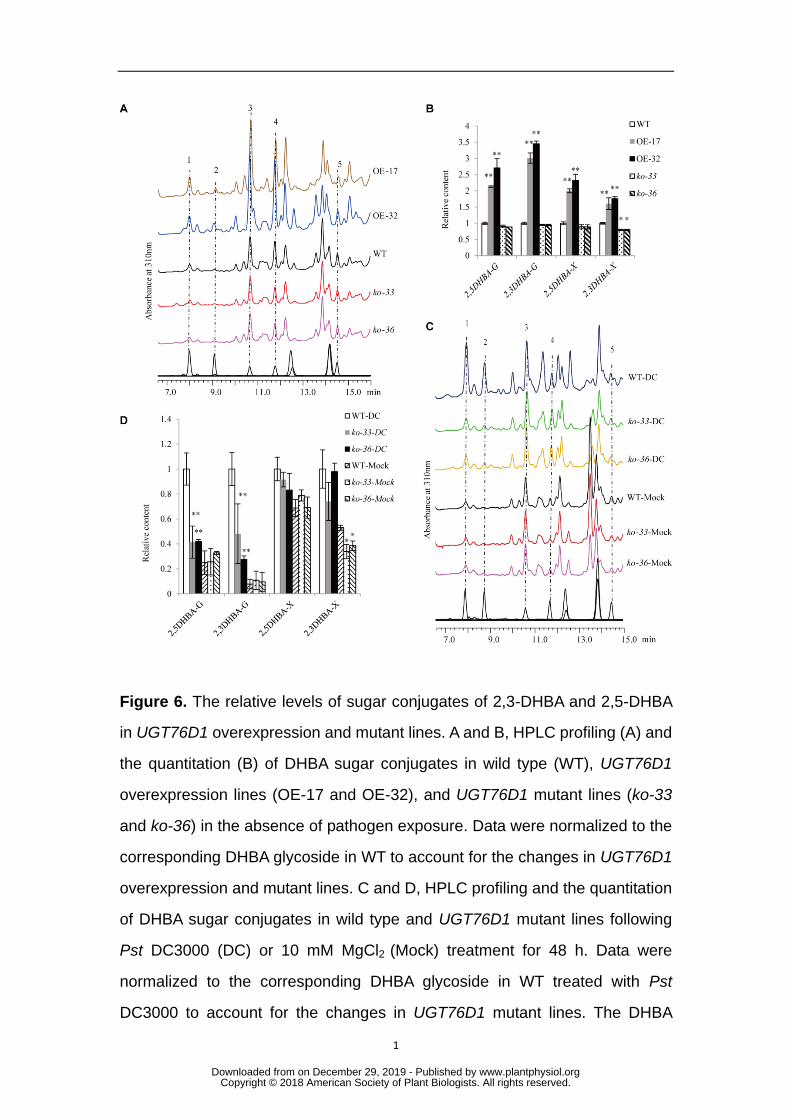
1
Figure 6. The relative levels of sugar conjugates of 2,3-DHBA and 2,5-DHBA
in UGT76D1 overexpression and mutant lines. A and B, HPLC profiling (A) and
the quantitation (B) of DHBA sugar conjugates in wild type (WT), UGT76D1
overexpression lines (OE-17 and OE-32), and UGT76D1 mutant lines (ko-33
and ko-36) in the absence of pathogen exposure. Data were normalized to the
corresponding DHBA glycoside in WT to account for the changes in UGT76D1
overexpression and mutant lines. C and D, HPLC profiling and the quantitation
of DHBA sugar conjugates in wild type and UGT76D1 mutant lines following
Pst DC3000 (DC) or 10 mM MgCl2 (Mock) treatment for 48 h. Data were
normalized to the corresponding DHBA glycoside in WT treated with Pst
DC3000 to account for the changes in UGT76D1 mutant lines. The DHBA
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
sugar conjugates formed by UGT76D1 catalysis in vitro were used as the
glycoside standards. Peak1: 2,5-DHBA glucosides; Peak2: 2,3-DHBA
glucosides; Peak3: 2,5-DHBA xlyoside; Peak4: 2,3-DHBA xylosides; Peak5:
internal reference (caffeic acid). 2,3DHBA-G: 2,3-DHBA glucosides;
2,3DHBA-X: 2,3DHBA xyloside; 2,5DHBA-G: 2,5-DHBA glucosides;
2,5DHBA-X: 2,5-DHBA xlyoside. Data are means ± SD of three biological
replicates (**P < 0.01, *P < 0.05, Student’s t-test).
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

1
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
Figure 7. UGT76D1-activated immune response depends on DHBA
glycosylation. A, Phenotypes of four-week-old wild-type (WT), UGT76D1
overexpression (OE-17 and OE-32), s3h mutant (s3h), and UGT76D1
overexpression/s3h hybrid (OE-17 s3h and OE-32 s3h) plants grown under
long day conditions at 22°C. Bar = 1 cm. B, DAB (top row panels) and Trypan
blue (bottom row panels) staining showing levels of H2O2 and cell death,
respectively, in leaves from four-week-old plants of UGT76D1
overexpression/s3h hybrid lines alongside control lines described in A. Bars =
100 μm. C, RT-PCR analysis of UGT76D1 expression in UGT76D1
overexpression/s3h hybrid lines alongside control lines described in A.
ACTIN2 was used as the internal control. D, RT-qPCR analysis of relative PR1
and PR2 expression in UGT76D1 overexpression/s3h hybrid lines alongside
control lines described in A. The value of WT plants was set at 1. Data are
means ± SD of three biological replicates (**P < 0.01, *P < 0.05, Student’s
t-test). E, The free and total SA levels in UGT76D1 overexpression/s3h hybrid
lines alongside control lines described in A. Data are means ± SD of three
biological replicates (**P < 0.01, *P < 0.05, Student’s t-test). F, Phenotypes of
transgenic plants overexpressing UGT76D1 in a WT or s3h s5h double mutant
background. The right panel shows the enlarged leaf phenotype. Leaf lesions
are indicated with red arrowheads. Bars = 1 cm (left panel) and 0.5 cm (right
panel).
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
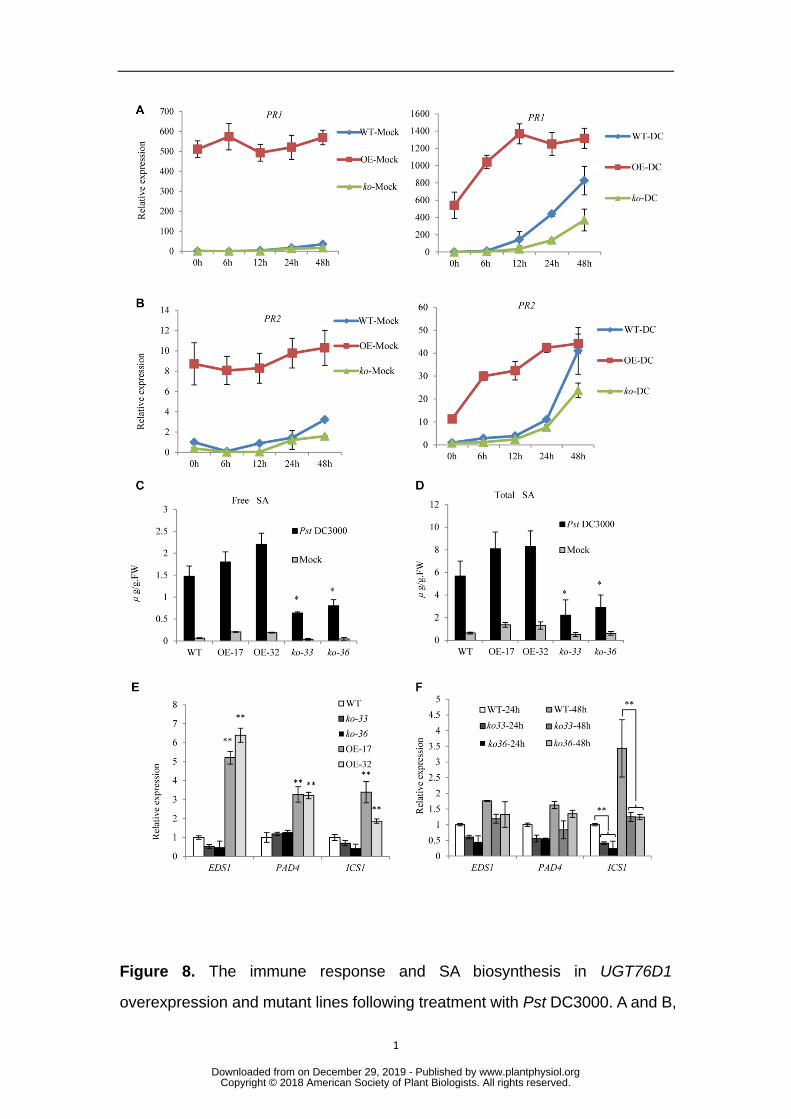
1
Figure 8. The immune response and SA biosynthesis in UGT76D1
overexpression and mutant lines following treatment with Pst DC3000. A and B,
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
Relative PR1 and PR2 expression in three-week-old wild-type (WT) and
UGT76D1 overexpression (OE) and mutant (ko) plants following Pst DC3000
(DC) or MgCl2 (Mock) treatment. For either treatment, expression levels in WT
at 0 h were set at 1. C and D, Free and total SA levels in three-week-old
UGT76D1 overexpression (OE-17 and OE-32) and mutant (ko-33 and ko-36)
lines following treatment with Pst DC3000 or MgCl2 (Mock) for 48 h. E,
RT-qPCR analysis of EDS1, PAD4, and ICS1 expression in UGT76D1
overexpression and mutant lines without any experimental treatment. Gene
expression levels in WT were set at 1.0. F, RT-qPCR analysis of EDS1, PAD4,
and ICS1 expression in UGT76D1 mutant lines following treatment with Pst
DC3000 for 24 h and 48 h. Gene expression levels in WT following 24 h Pst
DC3000 treatment were set at 1.0. A to F, Data are means ± SD of three
biological replicates (**P < 0.01, *P < 0.05, Student’s t-test).
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
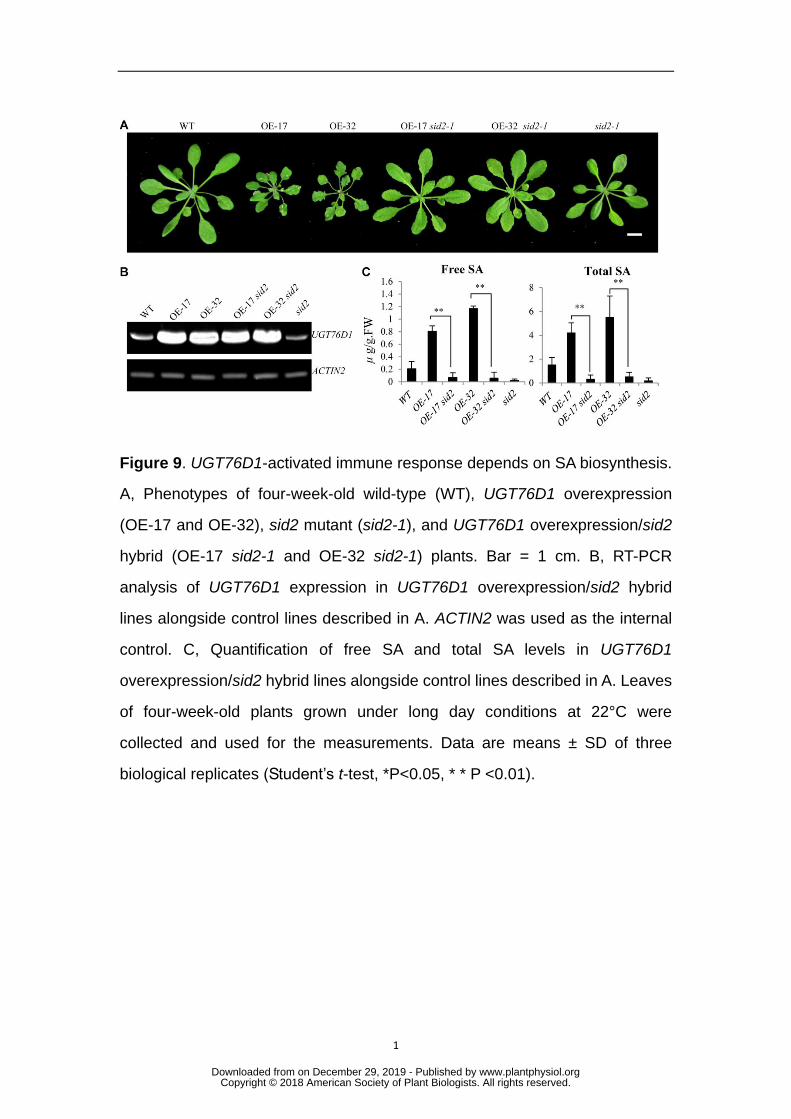
1
Figure 9. UGT76D1-activated immune response depends on SA biosynthesis.
A, Phenotypes of four-week-old wild-type (WT), UGT76D1 overexpression
(OE-17 and OE-32), sid2 mutant (sid2-1), and UGT76D1 overexpression/sid2
hybrid (OE-17 sid2-1 and OE-32 sid2-1) plants. Bar = 1 cm. B, RT-PCR
analysis of UGT76D1 expression in UGT76D1 overexpression/sid2 hybrid
lines alongside control lines described in A. ACTIN2 was used as the internal
control. C, Quantification of free SA and total SA levels in UGT76D1
overexpression/sid2 hybrid lines alongside control lines described in A. Leaves
of four-week-old plants grown under long day conditions at 22°C were
collected and used for the measurements. Data are means ± SD of three
biological replicates (Student’s t-test, *P<0.05, * * P <0.01).
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.
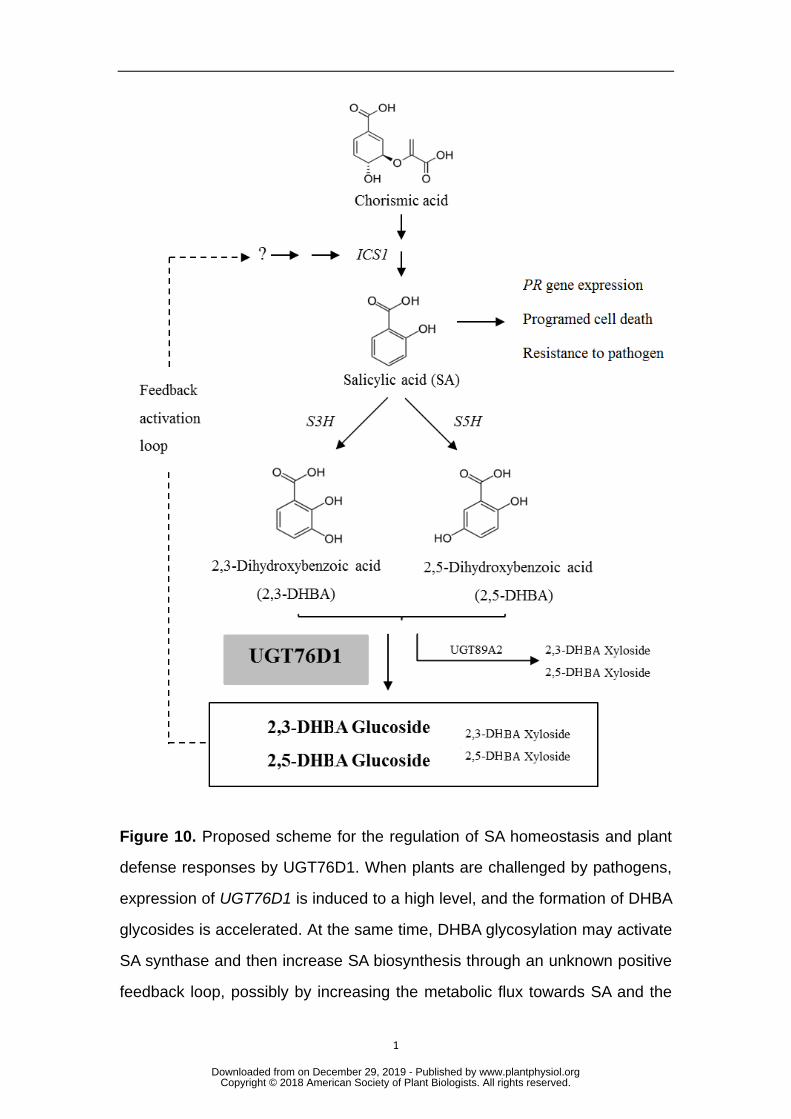
1
Figure 10. Proposed scheme for the regulation of SA homeostasis and plant
defense responses by UGT76D1. When plants are challenged by pathogens,
expression of UGT76D1 is induced to a high level, and the formation of DHBA
glycosides is accelerated. At the same time, DHBA glycosylation may activate
SA synthase and then increase SA biosynthesis through an unknown positive
feedback loop, possibly by increasing the metabolic flux towards SA and the
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

2
DHBAs. High levels of SA accumulation lead to an oxidative burst and PR
gene expression, and ultimately lead to PCD and increased plant resistance to
pathogens. Considering the xylosyltransferase activity of UGT89A2 towards
DHBAs, we cannot exclude the role of UGT89A2 in this immune-priming
process.
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Parsed CitationsAarts N, Metz M, Holub E, Staskawicz BJ, Daniels MJ, Parker JE (1998) Different requirements for EDS1 and NDR1 by diseaseresistance genes define at least two R gene-mediated signaling pathways in Arabidopsis. Proceedings of the National Academy ofSciences 95: 10306-10311
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Alvarez ME (2000) Salicylic acid in the machinery of hypersensitive cell death and disease resistance. In Programmed Cell Death inHigher Plants. Springer, pp 185-198
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bartsch M, Bednarek P, Vivancos PD, Schneider B, von Roepenack-Lahaye E, Foyer CH, Kombrink E, Scheel D, Parker JE (2010)Accumulation of isochorismate-derived 2, 3-dihydroxybenzoic 3-O-β-D-xyloside in Arabidopsis resistance to pathogens and ageing ofleaves. Journal of Biological Chemistry 285: 25654-25665
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bellés JM, Garro R, Fayos J, Navarro P, Primo J, Conejero V (1999) Gentisic acid as a pathogen-inducible signal, additional to salicylicacid for activation of plant defenses in tomato. Molecular plant-microbe interactions 12: 227-235
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Bruggeman Q, Raynaud C, Benhamed M, Delarue M (2015) To die or not to die? Lessons from lesion mimic mutants. Frontiers in plantscience 6
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen HY, Li X (2017) Identification of a residue responsible for UDP-sugar donor selectivity of a dihydroxybenzoic acidglycosyltransferase from Arabidopsis natural accessions. Plant J 89: 195-203
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chen Y, Shen H, Wang M, Li Q, He Z (2013) Salicyloyl-aspartate synthesized by the acetyl-amido synthetase GH3. 5 is a potentialactivator of plant immunity in Arabidopsis. Acta Biochim Biophys Sin 45: 827-836
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chong J, Baltz R, Schmitt C, Beffa R, Fritig B, Saindrenan P (2002) Downregulation of a pathogen-responsive tobacco UDP-Glc:phenylpropanoid glucosyltransferase reduces scopoletin glucoside accumulation, enhances oxidative stress, and weakens virusresistance. Plant Cell 14: 1093-1107
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Chong J, Pierrel M-A, Atanassova R, Werck-Reichhart D, Fritig B, Saindrenan P (2001) Free and conjugated benzoic acid in tobaccoplants and cell cultures. Induced accumulation upon elicitation of defense responses and role as salicylic acid precursors. PlantPhysiology 125: 318-328
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Clough SJ, Bent AF (1998) Floral dip: a simplified method forAgrobacterium‐mediated transformation ofArabidopsis thaliana. The plantjournal 16: 735-743
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Coll N, Epple P, Dangl J (2011) Programmed cell death in the plant immune system. Cell death and differentiation 18: 1247Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Coll NS, Vercammen D, Smidler A, Clover C, Van Breusegem F, Dangl JL, Epple P (2010) Arabidopsis type I metacaspases control celldeath. Science 330: 1393-1397
Pubmed: Author and Title www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

CrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dean JV, Delaney SP (2008) Metabolism of salicylic acid in wild‐type, ugt74f1 and ugt74f2 glucosyltransferase mutants of Arabidopsisthaliana. Physiologia plantarum 132: 417-425
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Delaney TP, Uknes S, Vernooij B, Friedrich L (1994) A central role of salicylic acid in plant disease resistance. Science 266: 1247Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Dempsey DMA, Vlot AC, Wildermuth MC, Klessig DF (2011) Salicylic acid biosynthesis and metabolism. The Arabidopsis Book: e0156Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fayos J, Bellés JM, Lopez-Gresa MP, Primo J, Conejero V (2006) Induction of gentisic acid 5-O-β-D-xylopyranoside in tomato andcucumber plants infected by different pathogens. Phytochemistry 67: 142-148
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Feng Z, Zhang B, Ding W, Liu X, Yang D-L, Wei P, Cao F, Zhu S, Zhang F, Mao Y (2013) Efficient genome editing in plants using aCRISPR/Cas system. Cell research 23: 1229
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Feys BJ, Moisan LJ, Newman MA, Parker JE (2001) Direct interaction between the Arabidopsis disease resistance signaling proteins,EDS1 and PAD4. The EMBO journal 20: 5400-5411
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Feys BJ, Wiermer M, Bhat RA, Moisan LJ, Medina-Escobar N, Neu C, Cabral A, Parker JE (2005) Arabidopsis senescence-associatedgene101 stabilizes and signals within an enhanced disease susceptibility1 complex in plant innate immunity. The Plant Cell 17: 2601-2613
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Fu ZQ, Dong X (2013) Systemic acquired resistance: turning local infection into global defense. Annual review of plant biology 64: 839-863
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Greenberg JT, Silverman FP, Liang H (2000) Uncoupling salicylic acid-dependent cell death and defense-related responses fromdisease resistance in the Arabidopsis mutant acd5. Genetics 156: 341-350
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Heath MC (2000) Hypersensitive response-related death. In Programmed cell death in higher plants. Springer, pp 77-90Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Hou B, Lim E-K, Higgins GS, Bowles DJ (2004) N-glucosylation of cytokinins by glycosyltransferases of Arabidopsis thaliana. Journal ofBiological Chemistry 279: 47822-47832
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ibrahim RK, Towers G (1959) Conversion of salicylic acid to gentisic acid and o-pyrocatechuic acid, all labelled with carbon-14, inplants. Nature 184: 1803-1803
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jabs T (1999) Reactive oxygen intermediates as mediators of programmed cell death in plants and animals. Biochemical pharmacology57: 231-245
Pubmed: Author and Title www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

CrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jackson RG, Kowalczyk M, Li Y, Higgins G, Ross J, Sandberg G, Bowles DJ (2002) Overexpression of an Arabidopsis gene encoding aglucosyltransferase of indole-3-acetic acid: phenotypic characterization of transgenic lines. Plant J 32:573–583
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Jones P, Messner B, Nakajima J, Schaffner AR, Saito K (2003) UGT73C6 and UGT78D1, glycosyltransferases involved in glycosidebiosynthesis in Arabidopsis thaliana. J Biol Chem. 278(45):43910-43918
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
King EO, Ward MK, Raney DE (1954) Two simple media for the demonstration of pyocyanin and fluorescin. The journal of laboratoryand clinical medicine 44: 301-307
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li J-F, Norville JE, Aach J, McCormack M, Zhang D, Bush J, Church GM, Sheen J (2013) Multiplex and homologous recombination-mediated genome editing in Arabidopsis and Nicotiana benthamiana using guide RNA and Cas9. Nature biotechnology 31: 688-691
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Li X, Svedin E, Mo H, Atwell S, Dilkes BP, Chapple C (2014) Exploiting natural variation of secondary metabolism identifies a genecontrolling the glycosylation diversity of dihydroxybenzoic acids in Arabidopsis thaliana. Genetics 198: 1267-1276
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lim EK, Doucet CJ, Li Y, Elias L, Worrall D, Spencer SP, Ross J, Bowles DJ (2002) The activity of Arabidopsis glycosyltransferasestoward salicylic acid, 4-hydroxybenzoic acid, and other benzoates. J Biol Chem 277: 586-592
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lim EK, Li Y, Parr A, Jackson R, Ashford DA, Bowles DJ (2001) Identification of glucosyltransferase genes involved in sinapatemetabolism and lignin synthesis in Arabidopsis. J Biol Chem 276: 4344-4349
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Lorrain S, Vailleau F, Balagué C, Roby D (2003) Lesion mimic mutants: keys for deciphering cell death and defense pathways in plants?Trends in plant science 8: 263-271
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Love AJ, Milner JJ, Sadanandom A (2008) Timing is everything: regulatory overlap in plant cell death. Trends in plant science 13: 589-595
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Malik B, Pirzadah TB, Tahir I, Rehman RU, Hakeem KR, Abdin M (2014) Plant signaling: response to reactive oxygen species. In Plantsignaling: Understanding the molecular crosstalk. Springer, pp 1-38
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Nawrath C, Metraux JP (1999) Salicylic acid induction-deficient mutants of Arabidopsis express PR-2 and PR-5 and accumulate highlevels of camalexin after pathogen inoculation. Plant Cell 11: 1393-1404
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Noutoshi Y, Okazaki M, Kida T, Nishina Y, Morishita Y, Ogawa T, Suzuki H, Shibata D, Jikumaru Y, Hanada A, Kamiya Y, Shirasu K (2012)Novel plant immune-priming compounds identified via high-throughput chemical screening target salicylic acid glucosyltransferases inArabidopsis. Plant Cell 24: 3795-3804
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Rate DN, Cuenca JV, Bowman GR, Guttman DS, Greenberg JT (1999) The gain-of-function Arabidopsis acd6 mutant reveals novelregulation and function of the salicylic acid signaling pathway in controlling cell death, defenses, and cell growth. The Plant Cell 11:1695-1708
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Rivas-San Vicente M, Plasencia J (2011) Salicylic acid beyond defence: its role in plant growth and development. Journal ofexperimental botany 62: 3321-3338
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Ryals J, Uknes S, Ward E (1994) Systemic acquired resistance. Plant Physiology 104: 1109Shah J (2003) The salicylic acid loop in plantdefense. Current opinion in plant biology 6: 365-371
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shah J (2003) The salicylic acid loop in plant defense. Current opinion in plant biology 6: 365-371Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shah J, Kachroo P, Nandi A, Klessig DF (2001) A recessive mutation in the Arabidopsis SSI2 gene confers SA-and NPR1-independentexpression of PR genes and resistance against bacterial and oomycete pathogens. The Plant Journal 25: 563-574
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Shirano Y, Kachroo P, Shah J, Klessig DF (2002) A gain-of-function mutation in an Arabidopsis Toll Interleukin1 Receptor–NucleotideBinding Site–Leucine-Rich Repeat type R gene triggers defense responses and results in enhanced disease resistance. The PlantCell 14: 3149-3162
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Straus MR, Rietz S, Ver Loren van Themaat E, Bartsch M, Parker JE (2010) Salicylic acid antagonism of EDS1-driven cell death isimportant for immune and oxidative stress responses in Arabidopsis. The Plant Journal 62: 628-640
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Thordal-Christensen H, Zhang Z, Wei Y, Collinge DB (1997) Subcellular localization of H2O2 in plants. H2O2 accumulation in papillaeand hypersensitive response during the barley—powdery mildew interaction. The Plant Journal 11: 1187-1194
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Tognetti VB, Van Aken O, Morreel K, Vandenbroucke K, van de Cotte B, De Clercq I, Chiwocha S, Fenske R, Prinsen E, Boerjan W,Genty B, Stubbs KA, Inze D, Van Breusegem F (2010) Perturbation of indole-3-butyric acid homeostasis by the UDP-glucosyltransferase UGT74E2 modulates Arabidopsis architecture and water stress tolerance. Plant Cell 22: 2660-2679
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Van Doorn WG (2011) Classes of programmed cell death in plants, compared to those in animals. Journal of Experimental Botany 62:4749-4761Vlot AC, Dempsey DMA, Klessig DF (2009) Salicylic acid, a multifaceted hormone to combat disease. Annual review ofphytopathology 47: 177-206
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Vlot AC, Dempsey DMA, Klessig DF (2009) Salicylic acid, a multifaceted hormone to combat disease. Annual review of phytopathology47: 177-206
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
von Saint Paul V, Zhang W, Kanawati B, Geist B, Faus-Kessler T, Schmitt-Kopplin P, Schaffner AR (2011) The Arabidopsisglucosyltransferase UGT76B1 conjugates isoleucic acid and modulates plant defense and senescence. Plant Cell 23: 4124-4145
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Walbot V, Hoisington DA, Neuffer M (1983) Disease lesion mimic mutations. In Genetic engineering of plants. Springer, pp 431-442Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wang B, Jin SH, Hu HQ, Sun YG, Wang YW, Han P, Hou BK (2012) UGT87A2, an Arabidopsis glycosyltransferase, regulates floweringtime via FLOWERING LOCUS C. New Phytologist 194: 666-675
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Wildermuth MC, Dewdney J, Wu G, Ausubel FM (2001) Isochorismate synthase is required to synthesize salicylic acid for plant defence.Nature 414: 562-565
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yalpani N, León J, Lawton MA, Raskin I (1993) Pathway of salicylic acid biosynthesis in healthy and virus-inoculated tobacco. Plantphysiology 103: 315-321
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang L, Li B, Zheng X-y, Li J, Yang M, Dong X, He G, An C, Deng XW (2015) Salicylic acid biosynthesis is enhanced and contributes toincreased biotrophic pathogen resistance in Arabidopsis hybrids. Nature communications 6: 7309
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yang S, Hua J (2004) A haplotype-specific Resistance gene regulated by BONZAI1 mediates temperature-dependent growth control inArabidopsis. The Plant Cell 16: 1060-1071
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Yoshioka K, Kachroo P, Tsui F, Sharma SB, Shah J, Klessig DF (2001) Environmentally sensitive, SA-dependent defense responses inthe cpr22 mutant of Arabidopsis. The Plant Journal 26: 447-459
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang GZ, Jin SH, Jiang XY, Dong RR, Li P, Li YJ, Hou BK (2016) Ectopic expression of UGT75D1, a glycosyltransferase preferringindole-3-butyric acid, modulates cotyledon development and stress tolerance in seed germination of Arabidopsis thaliana. Plant MolBiol 90: 77-93
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang K, Bhuiya M-W, Pazo JR, Miao Y, Kim H, Ralph J, Liu C-J (2012) An engineered monolignol 4-O-methyltransferase depresseslignin biosynthesis and confers novel metabolic capability in Arabidopsis. The Plant Cell 24: 3135-3152
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang K, Halitschke R, Yin C, Liu CJ, Gan SS (2013) Salicylic acid 3-hydroxylase regulates Arabidopsis leaf longevity by mediatingsalicylic acid catabolism. Proc Natl Acad Sci U S A 110: 14807-14812
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang Y, Xu S, Ding P, Wang D, Cheng YT, He J, Gao M, Xu F, Li Y, Zhu Z (2010) Control of salicylic acid synthesis and systemicacquired resistance by two members of a plant-specific family of transcription factors. Proceedings of the National Academy ofSciences 107: 18220-18225
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhang Y, Zhao L, Zhao J, Li Y, Wang J, Guo R, Gan S-S, Liu C-J, Zhang K (2017) S5H/DMR6 Encodes a Salicylic Acid 5-Hydroxylase ThatFine-Tunes Salicylic Acid Homeostasis. Plant physiology 175: 1082–1093
Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
Zhou F, Menke FL, Yoshioka K, Moder W, Shirano Y, Klessig DF (2004) High humidity suppresses ssi4-mediated cell death and diseaseresistance upstream of MAP kinase activation, H2O2 production and defense gene expression. The Plant Journal 39: 920-932
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.

Pubmed: Author and TitleCrossRef: Author and TitleGoogle Scholar: Author Only Title Only Author and Title
www.plantphysiol.orgon December 29, 2019 - Published by Downloaded from Copyright © 2018 American Society of Plant Biologists. All rights reserved.





























