Embed Size (px)
Citation preview

Modulation of imprinted gene network in placentaresults in normal development of in vitromanipulated mouse embryos
Patricia Fauque1,2,�, Marie-Anne Ripoche2, Jorg Tost3, Laurent Journot4, Anne Gabory2,
Florence Busato3, Anne Le Digarcher4, Francoise Mondon2, Ivo Gut3, Pierre Jouannet1,
Daniel Vaiman2, Luisa Dandolo2 and Helene Jammes2,5
1Biology of Reproduction, Hospital Cochin and 2Department of Genetics and Development, Institut Cochin, INSERM
U567, CNRS UMR 8104, University Paris Descartes, Paris, France, 3Centre National de Genotypage, Laboratory
for Epigenetics, CEA-Institut de Genomique, Evry, France, 4Institut of Functional Genomics, Montpellier, France
and 5Biology of Development and Reproduction, INRA—ENVA, UMR 1198, Jouy en Josas, France
Received December 15, 2009; Revised and Accepted February 9, 2010
Genomic imprinting regulates the expression of a group of genes monoallelically expressed in a parent-of-origin specific manner. Allele-specific DNA methylation occurs at differentially methylated regions (DMRs)of these genes. We have previously shown that in vitro fertilization and embryo culture result in methylationdefects at the imprinted H19– Igf2 locus at the blastocyst stage. The current study was designed to evaluatethe consequences of these manipulations on genomic imprinting after implantation in the mouse.Blastocysts were produced following three experimental conditions: (i) embryos maintained in culturemedium after in vivo fertilization or (ii) in vitro fertilization and (iii) a control group with embryos obtainedafter in vivo fertilization and timed mating. Blastocysts were all transplanted into pseudopregnant females.Embryos and placentas were collected on day 10.5 of development. DNA methylation patterns of the H19,Igf2, Igf2r and Dlk1–Dio3 DMRs were analyzed by quantitative pyrosequencing. In contrast to blastocyststage, methylation profiles were normal both in embryonic and placental tissues after in vitro fertilizationand culture. Expression of a selected set of imprinting genes from the recently described imprinted gene net-work (IGN) (including Igf2 and H19) was analyzed in placental tissues by quantitative RT–PCR. Placentasobtained after in vitro fertilization and embryo culture displayed significantly disturbed levels of H19 andIgf2 mRNA, as well as of most other genes from the IGN. As embryos were phenotypically normal, we hypoth-esize that the modulation of a coordinated network of imprinted genes results in a compensatory processcapable of correcting potential dysfunction of placenta.
INTRODUCTION
Children born from assisted reproductive technologies (ART)account for �2.2% of all births in the USA and 1.6% in theUK (1,2). Although the great majority of these children arein good health, a small but worrying percentage suffers froma variety of morbid congenital problems and has twice therate of major birth defects compared with naturally conceivedoffspring (3). This risk persists even after adjusting for
increased maternal age and increased incidence of multiplegestations after ART. Birth defects include chromosomalabnormalities, musculoskeletal and cardiovascular defects,and low birth weight (4). Furthermore, several reportssuggested that in vitro fertilization (IVF) increases the riskof diseases caused by aberrant genomic imprinting such asBeckwith–Wiedemann syndrome (5–9), Angelman syndrome(8,10,11) and Silver–Russell syndrome (12,13). Imprintedgenes are expressed monoallelically in a parent-of-origin
�To whom correspondence should be addressed at: Biology of Reproduction, University Paris Descartes, Hospital Cochin, 24 rue du FaubourgSt Jacques, Paris 75014, France. Tel: þ33 1 58 41 22 76; Fax: þ33 1 58 41 15 54; Email: [email protected]
# The Author 2010. Published by Oxford University Press. All rights reserved.For Permissions, please email: [email protected]
Human Molecular Genetics, 2010, Vol. 19, No. 9 1779–1790doi:10.1093/hmg/ddq059Advance Access published on February 11, 2010
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

specific manner. These genes are generally located in clusters,epigenetically marked by DNA methylation on key regulatorysequences (differentially methylated regions, DMRs), byhistone modifications (acetylation/deacetylation and methyl-ation) and sometimes associated with antisense RNAs(14–16). The allele-specific methylation of primary DMRsor imprinting control centers (ICR) such as the H19 DMRoccurs in germ cells and provides a heritable ‘memory’ thatis maintained throughout fertilization and embryo develop-ment. This differential methylation is preserved during preim-plantation development, despite genome-wide changes inglobal DNA methylation occurring at these early stages (17).In somatic cells, the other DMRs considered as secondaryDMRs are de novo methylated and the allele-specific methyl-ation status is maintained throughout cell division.
It has been previously demonstrated that IVF disturbs DNAmethylation patterns at specific locations of the H19–Igf2locus in individual mouse blastocysts and that epigeneticchanges were influenced by culture medium (18). These dataas well as data from others (19,20) suggest that the H19–Igf2 locus could be considered as a relevant biomarker of epi-genetic alterations induced by environmental conditions.
However, the temporal evolution of these defective DNAmethylation/gene expression patterns during the developmen-tal process is so far unclear. Could the embryos exposed tovarious environmental conditions restore correct DNA methyl-ation and expression patterns of H19 and Igf2 genes at thepost-implantation stage? The current study was designed toexamine the effects of two different embryo culture mediaassociated or not with in vitro fertilization on subsequentnormal post-implantation development (Fig. 1). We studiedmethylation profiles of the H19 and Igf2 DMRs, as well asof the Igf2r and Dlk1–Dio3 DMRs in embryos and placentascollected at 10.5 days of development. The expression levelsof 19 genes belonging to the imprinted gene network (IGN)
involved in embryonic growth, were also analyzed in placen-tas (21,22). We were able to show that the collected embryosdisplay a phenotypically normal development after in vitrofertilization and different culture procedures. They alsopresent the expected methylation profile for all DMRs bothin placenta and embryos without significant alteration.Unexpectedly, expression of H19 and Igf2 was up-regulatedin the placenta from manipulated embryos when comparedwith the control group. A correlated regulation of severalother imprinted genes was also observed. We hypothesizethat this modulation of several genes from the IGN in a coor-dinated fashion could constitute a major compensatory mech-anism enabling the developing fetus to cope with changing orabnormal environment.
RESULTS
Mouse blastocyts were produced following three differentexperimental conditions: group A after in vivo fertilizationand development (as control group), group B after in vivo fer-tilization and in vitro development and group C after bothin vitro fertilization and development. Group C is the groupin which the different steps of human ART are most closelymimicked. All embryos were transferred to the uteri of pseu-dopregnant females. Embryos and placentas were then col-lected from recipient females at day 10.5 of development(Fig. 1). Upon close observation, all embryos were at thecorrect developmental stage according to the Theiler criteria(23) and were collected. However, some resorption siteswere observed. The number of resorption sites was signifi-cantly higher in the manipulated groups [with embryoculture (47.8%) or in vitro fertilization and embryo culture(64.1%)] when compared with the control group (18.5%;P , 0.05) suggesting that embryos died at an early stage ofdevelopment (24).
Figure 1. Experimental design. In vivo fertilization and preimplantation development (group A); in vivo fertilization and in vitro preimplantation embryo devel-opment (group B); in vitro fertilization and preimplantation embryo development (group C). Embryo culture was performed in two different culture media: M16and G1/G2. The timeline in hours (h) is relative to the hCG injection (0 h). For all groups, the blastocysts were transferred in pseudopregnant d3.5 F1 females.
1780 Human Molecular Genetics, 2010, Vol. 19, No. 9
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

Methylation analysis of four DMRs of the H19–Igf2 locusin embryos and placentas from control group
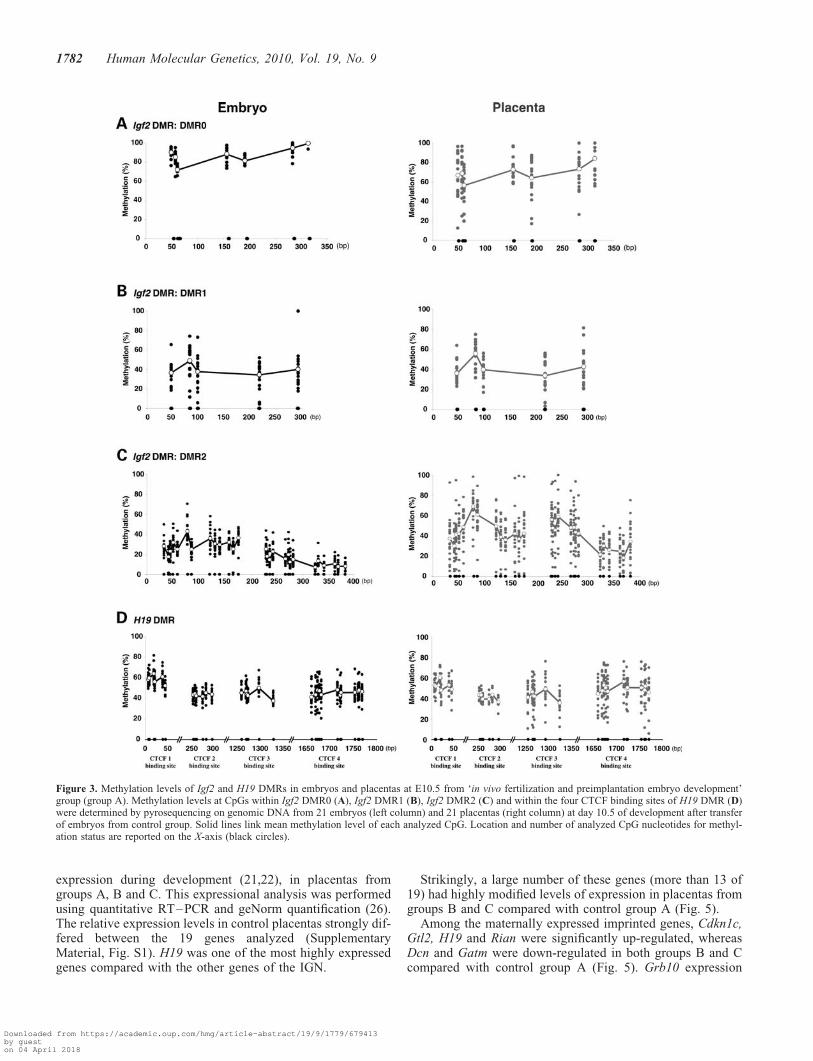
We analyzed the methylation levels of 39 CpGs of the Igf2DMR0, DMR1 and DMR2 and of the 30 CpGs located inthe four CCCTC-binding factor (CTCF) binding sites of theH19 DMR (Fig. 2). This was first done on genomic DNAfrom control embryos and placentas (group A) collected atday 10.5 of development.
For the Igf2 DMR0, the mean methylation level was 87.2+0.8% in embryo and 69.4+ 1.6% in placental DNA (Fig. 3A).This level was significantly decreased in the placenta com-pared with embryos (P , 0.001). No significant differenceof the methylation status was found between embryonic andextra-embryonic tissues for the Igf2 DMR1 (39.0+ 1.6%versus 41.4+ 1.5%, respectively) (Fig. 3B). Finally, themean methylation was significantly reduced in embryos com-pared with placentas for the Igf2 DMR2 (Fig. 3C). Theaverage methylation was 29.6+ 0.6 and 44.4+ 1.2% for thefirst 13 CpGs and 13.3+ 0.4 and 36.9+ 1.1% for the nextCpGs in embryos and placentas, respectively, demonstratinga significant hypomethylation of the DMR2 (P , 0.001) inembryos at day 10.5 of development.
For the H19 DMR, a similar mean methylation level wasobserved for the four CTCF binding sites in embryos and pla-centas (47.6+ 0.4 versus 47.1+ 0.6%, Fig. 3D). However,three placentas of 21 without morphological abnormalitiespresented a significant hypomethylation at CTCF bindingsites 3 and 4, whereas CTCF biding sites 1 and 2 displayeda normal methylation level. The three counterpart embryosdisplayed a normal methylation pattern of all CpGs.
Methylation analysis of the H19–Igf2 locus in embryosand placentas from experimental groups
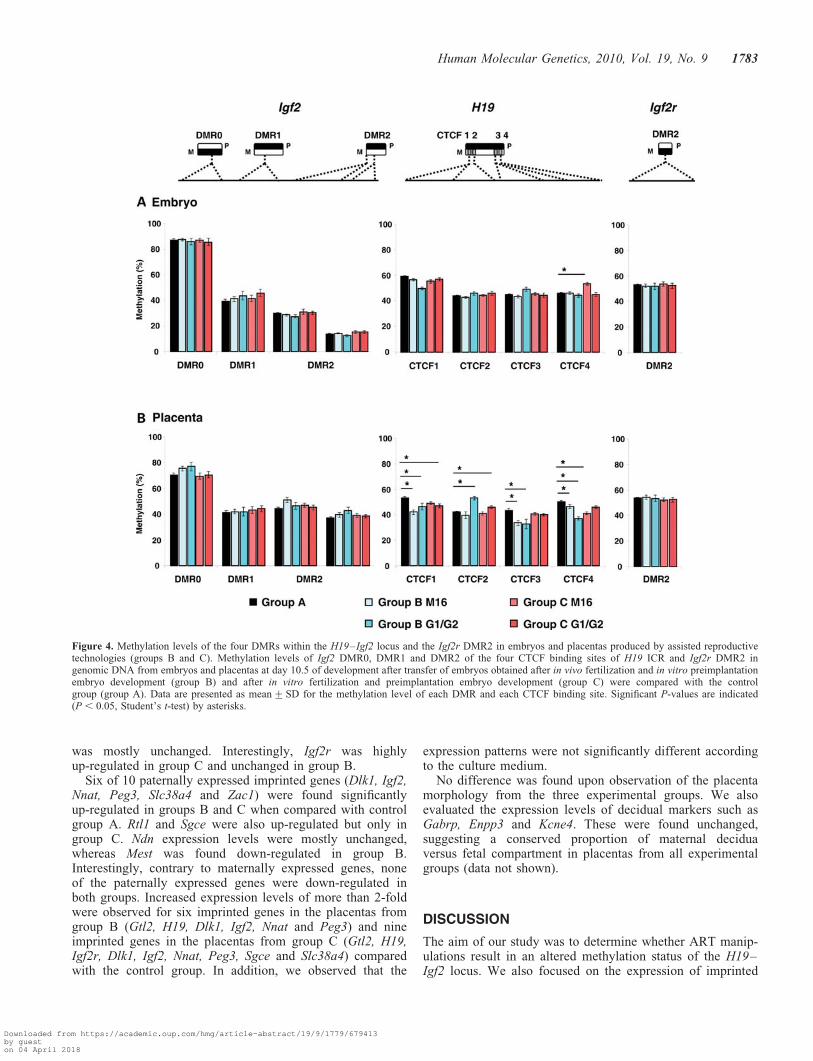
The methylation levels of the four DMRs in DNA from exper-imental groups B and C were compared with the control group(Fig. 4).
For the Igf2 DMR0, embryos from group B and C presented amean methylation level of 87.0+1.2 and 86.3+1.5% compared
with 75.8+1.7 and 69.9+1.7% in placental DNA in bothculture media (Fig. 4). These results did not differ from thecontrol group A in which, as mentioned above, the DMR0methylation level was significantly lower in placenta comparedwith embryo. No significant difference was observed neither forembryos from groups A, B and C in the two other DMRs(DMR1: 39.0+1.7%, 41.6+1.8%, 42.9+1.8% and DMR2:21.9+0.5%, 20.9+0.6%, 22.4+0.8%, respectively, t-test,P . 0.05) nor for the placentas (DMR1: 41.9+1.7%, 43.5+1.8% and 41.4+1.5%, DMR2: 45.3+1.3%, 42.4+0.8% and40.7+0.8%, respectively, t-test, P . 0.05). This suggests thatthe culture media has no influence on the methylation profile.
Comparable mean methylation levels of H19 DMR werealso observed in embryos of groups A, B and C (58.7+0.6%, 54.6+ 0.9%, 55.5+ 1.0%, respectively) (Fig. 4).However, a decrease of �10% was observed for CTCFbinding site 1, 3 and 4 in placentas from group B and C com-pared with placentas from control group. When using pyrose-quencing analysis, only differences in methylation levelsabove 5% are considered as significant (25). However, it isnot clear whether this placental hypomethylation representsan important alteration, since a high individual variability ofmethylation level was observed in all groups (Fig. 3D).
In addition, we analyzed the methylation levels of two otherDMRs, the paternally methylated Dlk1–Dio3 imprintingcontrol region (13 CpGs) and the maternally methylatedIgf2r DMR (12 CpGs). For the Igf2r DMR, comparablemethylation levels were found in embryos and placentas incontrol group A (52.7+ 0.9 and 53.6+ 1.1%, respectively,t-test, P . 0.05; Fig. 4) and no difference was observedbetween A, B and C experimental groups. Additional analysison Dlk1–Dio3 imprinting control region confirmed theabsence of methylation perturbation in groups B and C com-pared with the control group (data not shown).
Expression of the IGN
To extend our analysis, we compared the level of expressionof H19 and Igf2 as well as of 17 other imprinted genes belong-ing to the previously described IGN which displays correlated
Figure 2. Schematic representation of the mouse H19–Igf2 imprinted locus. Schematic map of the mouse H19–Igf2 imprinted locus showing the relative pos-ition of the H19 and Igf2 genes. The number and position of upstream exons (C1 and C2 for Igf2) and exons are shown as grey (Igf2 gene) and white (H19 gene)rectangles with arrows for transcription start sites. Enhancers (E) are indicated as grey circles. The locations of four DMRs within the H19–Igf2 imprinted locusrepresented by boxes are shaded to indicate preferential methylation of the maternal (M) or paternal (P) allele in each region. Grey boxes within H19 DMRrepresent the four CCCTC-binding-factor binding sites. Location and number of CpG nucleotides analyzed for methylation status are given (black circles).
Human Molecular Genetics, 2010, Vol. 19, No. 9 1781
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018
expression during development (21,22), in placentas fromgroups A, B and C. This expressional analysis was performedusing quantitative RT–PCR and geNorm quantification (26).The relative expression levels in control placentas strongly dif-fered between the 19 genes analyzed (SupplementaryMaterial, Fig. S1). H19 was one of the most highly expressedgenes compared with the other genes of the IGN.
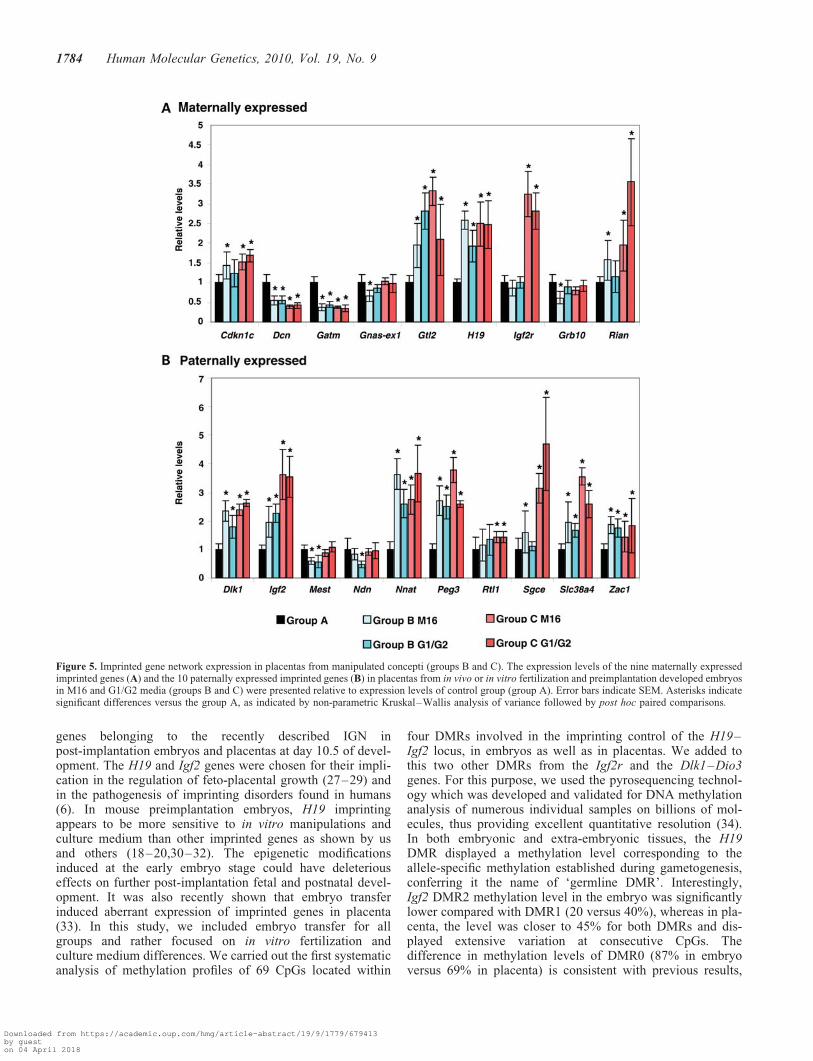
Strikingly, a large number of these genes (more than 13 of19) had highly modified levels of expression in placentas fromgroups B and C compared with control group A (Fig. 5).
Among the maternally expressed imprinted genes, Cdkn1c,Gtl2, H19 and Rian were significantly up-regulated, whereasDcn and Gatm were down-regulated in both groups B and Ccompared with control group A (Fig. 5). Grb10 expression
Figure 3. Methylation levels of Igf2 and H19 DMRs in embryos and placentas at E10.5 from ‘in vivo fertilization and preimplantation embryo development’group (group A). Methylation levels at CpGs within Igf2 DMR0 (A), Igf2 DMR1 (B), Igf2 DMR2 (C) and within the four CTCF binding sites of H19 DMR (D)were determined by pyrosequencing on genomic DNA from 21 embryos (left column) and 21 placentas (right column) at day 10.5 of development after transferof embryos from control group. Solid lines link mean methylation level of each analyzed CpG. Location and number of analyzed CpG nucleotides for methyl-ation status are reported on the X-axis (black circles).
1782 Human Molecular Genetics, 2010, Vol. 19, No. 9
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018
was mostly unchanged. Interestingly, Igf2r was highlyup-regulated in group C and unchanged in group B.
Six of 10 paternally expressed imprinted genes (Dlk1, Igf2,Nnat, Peg3, Slc38a4 and Zac1) were found significantlyup-regulated in groups B and C when compared with controlgroup A. Rtl1 and Sgce were also up-regulated but only ingroup C. Ndn expression levels were mostly unchanged,whereas Mest was found down-regulated in group B.Interestingly, contrary to maternally expressed genes, noneof the paternally expressed genes were down-regulated inboth groups. Increased expression levels of more than 2-foldwere observed for six imprinted genes in the placentas fromgroup B (Gtl2, H19, Dlk1, Igf2, Nnat and Peg3) and nineimprinted genes in the placentas from group C (Gtl2, H19,Igf2r, Dlk1, Igf2, Nnat, Peg3, Sgce and Slc38a4) comparedwith the control group. In addition, we observed that the
expression patterns were not significantly different accordingto the culture medium.
No difference was found upon observation of the placentamorphology from the three experimental groups. We alsoevaluated the expression levels of decidual markers such asGabrp, Enpp3 and Kcne4. These were found unchanged,suggesting a conserved proportion of maternal deciduaversus fetal compartment in placentas from all experimentalgroups (data not shown).
DISCUSSION
The aim of our study was to determine whether ART manip-ulations result in an altered methylation status of the H19–Igf2 locus. We also focused on the expression of imprinted
Figure 4. Methylation levels of the four DMRs within the H19–Igf2 locus and the Igf2r DMR2 in embryos and placentas produced by assisted reproductivetechnologies (groups B and C). Methylation levels of Igf2 DMR0, DMR1 and DMR2 of the four CTCF binding sites of H19 ICR and Igf2r DMR2 ingenomic DNA from embryos and placentas at day 10.5 of development after transfer of embryos obtained after in vivo fertilization and in vitro preimplantationembryo development (group B) and after in vitro fertilization and preimplantation embryo development (group C) were compared with the controlgroup (group A). Data are presented as mean+SD for the methylation level of each DMR and each CTCF binding site. Significant P-values are indicated(P , 0.05, Student’s t-test) by asterisks.
Human Molecular Genetics, 2010, Vol. 19, No. 9 1783
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018
genes belonging to the recently described IGN inpost-implantation embryos and placentas at day 10.5 of devel-opment. The H19 and Igf2 genes were chosen for their impli-cation in the regulation of feto-placental growth (27–29) andin the pathogenesis of imprinting disorders found in humans(6). In mouse preimplantation embryos, H19 imprintingappears to be more sensitive to in vitro manipulations andculture medium than other imprinted genes as shown by usand others (18–20,30–32). The epigenetic modificationsinduced at the early embryo stage could have deleteriouseffects on further post-implantation fetal and postnatal devel-opment. It was also recently shown that embryo transferinduced aberrant expression of imprinted genes in placenta(33). In this study, we included embryo transfer for allgroups and rather focused on in vitro fertilization andculture medium differences. We carried out the first systematicanalysis of methylation profiles of 69 CpGs located within
four DMRs involved in the imprinting control of the H19–Igf2 locus, in embryos as well as in placentas. We added tothis two other DMRs from the Igf2r and the Dlk1–Dio3genes. For this purpose, we used the pyrosequencing technol-ogy which was developed and validated for DNA methylationanalysis of numerous individual samples on billions of mol-ecules, thus providing excellent quantitative resolution (34).In both embryonic and extra-embryonic tissues, the H19DMR displayed a methylation level corresponding to theallele-specific methylation established during gametogenesis,conferring it the name of ‘germline DMR’. Interestingly,Igf2 DMR2 methylation level in the embryo was significantlylower compared with DMR1 (20 versus 40%), whereas in pla-centa, the level was closer to 45% for both DMRs and dis-played extensive variation at consecutive CpGs. Thedifference in methylation levels of DMR0 (87% in embryoversus 69% in placenta) is consistent with previous results,
Figure 5. Imprinted gene network expression in placentas from manipulated concepti (groups B and C). The expression levels of the nine maternally expressedimprinted genes (A) and the 10 paternally expressed imprinted genes (B) in placentas from in vivo or in vitro fertilization and preimplantation developed embryosin M16 and G1/G2 media (groups B and C) were presented relative to expression levels of control group (group A). Error bars indicate SEM. Asterisks indicatesignificant differences versus the group A, as indicated by non-parametric Kruskal–Wallis analysis of variance followed by post hoc paired comparisons.
1784 Human Molecular Genetics, 2010, Vol. 19, No. 9
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

since DMR0 is methylated on both alleles in the fetus and onlyon the maternal allele in placenta (35). The function of DMR0is still unknown although it could be involved in a complexphenomenon of IGF2 imprinting loss occurring in humandisease (36). The data obtained in embryonic tissues are inclose accordance with the model in which the DNA methyl-ation patterns of the three DMRs (H19 DMR and Igf2DMRs 1 and 2) are coordinately regulated by a hierarchy ofthese DMRs which in turn arises from long-range chromatininteractions (37).
Samples collected from the three experimental groups(A, B and C) displayed a similar pattern of methylation forall DMRs analyzed. In addition, no difference of the methyl-ation status was found between a basic culture medium(M16) and a more complex sequential medium (G1/G2) con-taining methionine known to be involved in the methylationprocess. Nevertheless, we observed a higher individual varia-bility in placenta compared with embryo in all groups,highlighting a specific epigenetic flexibility of placenta.As previously reported for the ICR2 locus on the distalchromosome 7, other epigenetic marks such as histone modi-fications could act as modulators of imprinting in placentaltissue (38).
In embryos obtained by in vitro development using Musmusculus castaneus females and C57BL/6J males, embryonictissues preserved correct genomic imprints, whereas aberrantH19 imprinting was observed in some placentas (39).However, in the reciprocal cross, both placentas andembryos showed appropriate imprint of the H19 gene underidentical conditions. This genotype effect suggests that strain-specific modifiers might affect maintenance of the H19 imprintspecifically in placental tissue. In our study, the methylationvariability observed between concepti, all with a normaldevelopment according to their gestational stage could beexplained by the formation of epigenetic mosaic embryos,probably at early stages of development.
Although our previous data had shown methylation differ-ences for H19 on individual blastocysts, our present resultsshow no major difference in the embryo methylationprofile between control and experimental groups whateverthe culture system used. This could suggest that theembryos with major epigenetic alterations do not implantor do not develop with a normal phenotype until 10.5 dpc.In contrast, embryos with minor epigenetic alterationswould be able to restore an accurate methylation status atthe H19–Igf2 and other imprinted loci and present anormal post-implantation development. We demonstratehere that for implanted and normally developed embryos,H19 and Igf2 methylation was not affected although signifi-cant demethylations were observed for several CTCF bindingsites of H19 DMR in the placenta. Our additional resultsconcerning the Dlk1–Dio3 and Igf2r DMRs confirm theappropriate epigenetic mark apposition in embryonic and pla-cental tissues at E10.5. Our data support the hypothesis thatthe feto-placental development requires a completely restoredepigenetic pattern in the embryo at least or that only embryoswith epigenetic marks associated with functional imprintinghave a satisfactory later embryonic development. Thishypothesis is reinforced by the lower implantation rate ofblastocysts and the higher number of resorption sites in the
manipulated groups when compared with control group,reported by us and others (33).
Placenta plays a crucial role in normal fetal development,and is a major organ for filtering effects coming from theoutside environment to the fetus (40). Therefore, we extendedour investigation to a significant part of the IGN which hasbeen recently described to be involved in embryonic and post-natal growth control (21,22). We demonstrated that in vitrofertilization and embryo culture modified expression of morethan 15 of 19 genes in placentas of normally developedE10.5 embryos. Both maternally and paternally expressedimprinted genes are modified and mostly up-regulated. Onlytwo maternally expressed genes were found down-regulatedin placentas from all experimental groups and only one pater-nally expressed gene was down-regulated in placentasobtained after embryo culture.
A first group of genes including H19, Igf2, Igf2r, Zac1 andSlc38a4 was up-regulated in manipulated placentas and couldbe included in a regulatory pathway. Interestingly, H19 wasstrongly affected with a 2-fold increase in expression.Although H19 is highly expressed throughout embryogenesis(41,42), its role in placenta remains unknown. A recentstudy using transgenic mice with gain or loss of H19expression suggests that H19 acts as a trans-regulator of theIGN in the embryo (22). Inactivation of Igf2 induces intrauter-ine growth restriction of embryo closely associated to thedecrease of placental functions (43,44). The lack of Igf2rresults in embryonic overgrowth (43,45–47). In our manipu-lated placentas obtained after in vitro fertilization, theseimprinted genes with reciprocal functions in fetal and placen-tal growth were all up-regulated when compared with thecontrol group. Whereas recent studies have shown thatembryo transfer reduces and superovulation increases Igf2mRNA levels (33,48), we found that embryo culture augmen-ted these levels and that in vitro fertilization further exacer-bated this effect. It is however difficult to compare theseexperiments and to conclude exactly which step of ART isinvolved in the modulation of Igf2 expression, because theaim of each study was different. Additional experimentsincluding all variables (superovulation, transfer, IVF andculture) will be required to clarify the situation.
Zac1 directly regulates the H19–Igf2 locus through bindingto a shared enhancer (21). As in vitro manipulations result inZac1 overexpression, this gene may act as an activator of H19and Igf2 expression observed in manipulated groups. An inter-esting result is the up regulation of Igf2r gene limited to placen-tas from group C. This raises the possibility of a regulation ofthe local IGF2 concentration with a para/autocrine activity inplacental tissue. Indeed, IGF2R plays a major role in bindingand regulating the tissue levels of IGF2. Finally, the paternallyexpressed Slc38a4 gene (located on chromosome 15) isimprinted in the placenta only and encodes a system A aminoacid transporter (49). It was recently suggested that the placentacould respond to fetal demand through crosstalk between theIgf2 gene and placental supply transporter genes such as theSlc38a4 gene (49). Other imprinted genes (Peg3, Cdkn1c andDlk1) involved in growth were also significantly up-regulatedby in vitro manipulations. Peg3 on mouse chromosome 7encodes a large Kruppel-type finger protein which is paternallyexpressed and associated with growth abnormalities (50).
Human Molecular Genetics, 2010, Vol. 19, No. 9 1785
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

Cdkn1c, also on mouse chromosome 7, resides in theBeckwith–Wiedmann cluster and is involved in placentaldevelopment (51). Dlk1, paternally expressed on chromosome12 encodes a member of Notch–Delta family of signalingmolecules. It was recently demonstrated that the Gtl2–Dlk1locus contributes to mouse placentation (52). In this locus, thepaternally expressed Rtl1 is a retrotransposon-like gene. Itappears essential for maintenance of the fetal capillaries, andboth its loss and its overproduction cause late-fetal and/or neo-natal lethality in mice (53). Sgce located on chromosome 6 andpaternally expressed, produces a member of the sarcoglycanfamily and a component of the transmembrane mediating cytos-keleton–extracellular matrix, highly expressed in the placenta(54,55). Gatm and Dcn, two maternally expressed genes notexclusively expressed in maternal decidua (56,57), were down-regulated in manipulated groups. Gatm, on chromosome 2,encodes an L-arginine:glycine amidinotransferase which cata-lyzes the first step in creatine biosynthesis. Gatm levels couldbe critical for normal energy metabolism in the developingembryo (57). A decreased expression of GATM and otherimprinted genes has been recently described in human intrauter-ine growth restriction cases (58). Decorin (Dcn) gene on mousechromosome 10 is maternally expressed in placenta only (59). Itencodes a member of the proteoglycan family that can bind toTGF-b and to IGF1 receptor and may be involved in processessuch as matrix assembly, cell adhesion, migration andproliferation (60).
Our study has revealed the disrupted expression of numer-ous imprinted genes in the placenta after embryo manipu-lation. However, the methylation status of the DMRs fromthe H19, Igf2, Igf2r and Dlk1 genes is either similar to controlsor very slightly modified. This would suggest that primaryimprints are correctly established and maintained on thesesequences and that it is the reading of these epigeneticmarks which is altered, possibly at a post-implantation step.These alterations could include defective binding of transcrip-tion factors, modification of histone marks or of post-transcriptional regulations.
In summary, we observed an important and complex regu-lation of a large set of imprinted genes in manipulatedgroups. This illustrates how the placenta can respond to fetaldemand signals through gene regulation involved in growthcontrol and specific placental transport systems. As allembryos analyzed in this study were phenotypically and mor-phologically normal, this suggests that the regulation of IGNin placenta may adapt to environmental changes occurringduring the first steps of development. An attractive hypothesisis that the biological function associated with the placentalIGN is to participate in the control of normal fetal growthduring post-implantation development through a fine-tunedcompensatory regulation.
Previous data had suggested that affecting one gene (such asdeletion of Zac1 or H19) resulted also in the attenuation of aphenotype through an adaptative process capable of modulat-ing the expression of the genes from the IGN (21,22). Fromthe experiments performed here, we can postulate that perturb-ing effects due to in vitro manipulation result in a counterba-lance of the members of the IGN in the placenta. This couldserve as a ‘buffering system’ and result in normal developmentof the embryo. Whether this general effect is due to one master
gene which then affects a cascade of genes or whether severalgenes drive this developmental modulation is not clear.
Finally, our data on the consequences of ART manipula-tions in the mouse model at post-implantation stage moderatethe risk of epigenetic abnormalities and appear more reassur-ing for the health of children born after ART compared withstudies investigating preimplantation embryos. Imprintedgenes appear to play a crucial role in adjusting the functionof the placenta and obtaining a fairly normal embryonicdevelopment.
MATERIALS AND METHODS
Animals
Five- to six-week-old F1 (C57BL/6/CBA) females chosen fortheir good responsiveness to hormonal stimulation and eight-to nine-week-old F1 males (Charles River Laboratories,L’Arbresle, France) were maintained in an animal facility atnormal temperature (21–238C) and 14 h light/10 h dark photo-periods with free access to water and food. Procedures forhandling and experimentation followed ethical guidelinesestablished by the Federation of European LaboratoryAnimal Science Associations.
Experimental design
Superovulated mice were divided into three different exper-imental groups (A, B and C). All females were superovulated.In group A, blastocysts were collected after in vivo fertiliza-tion and in vivo development. In group B, blastocysts wereobtained after in vivo fertilization and in vitro developmentfrom one cell to blastocyst stage. In group C, blastocystswere obtained after in vitro fertilization and development.
The embryos from groups B and C were cultured either inM16 medium (Sigma-Aldrich, Lyon, France) (B M16) or insequential G1/G2 medium (JCD Laboratories, Lyon, France)(C G1/G2).
Superovulation. Females were superovulated by intraperito-neal (i.p.) injection of 8 IU (0.1 ml) Pregnant Mare SerumGonadotropin (PMSG, Chronogest; Intervet, Beaucouze,France), followed 47 h later by an i.p. injection of 5 IU(0.1 ml) of human Chorionic Gonadotropin (hCG, Chorulon;Intervet).
In vitro fertilization and embryo culture. Thirteen hourspost-hCG, cumulus–oocyte complexes were recovered fromoviducts in M2 medium (Sigma-Aldrich) supplemented with7 mg/ml of BSA (Sigma-Aldrich). After rinsing in M16medium, cumulus–oocyte complexes were kept in the incuba-tor (378C, 5.5% CO2 in air) in 100 ml drops of M16 mediumcovered with paraffin oil (Vitrolife). Spermatozoa were col-lected from the cauda epididymis and capacitated for 90 minin M16 medium supplemented with 7 mg/ml of BSA at378C and 5.5% CO2. Oocytes were inseminated 15 hpost-hCG with 106 spermatozoa. Fertilized eggs (23 hpost-hCG) as determined by the presence of two pronucleiwere transferred to 30 ml drops of fresh medium coveredwith paraffin oil. To analyze the embryo culture medium
1786 Human Molecular Genetics, 2010, Vol. 19, No. 9
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018
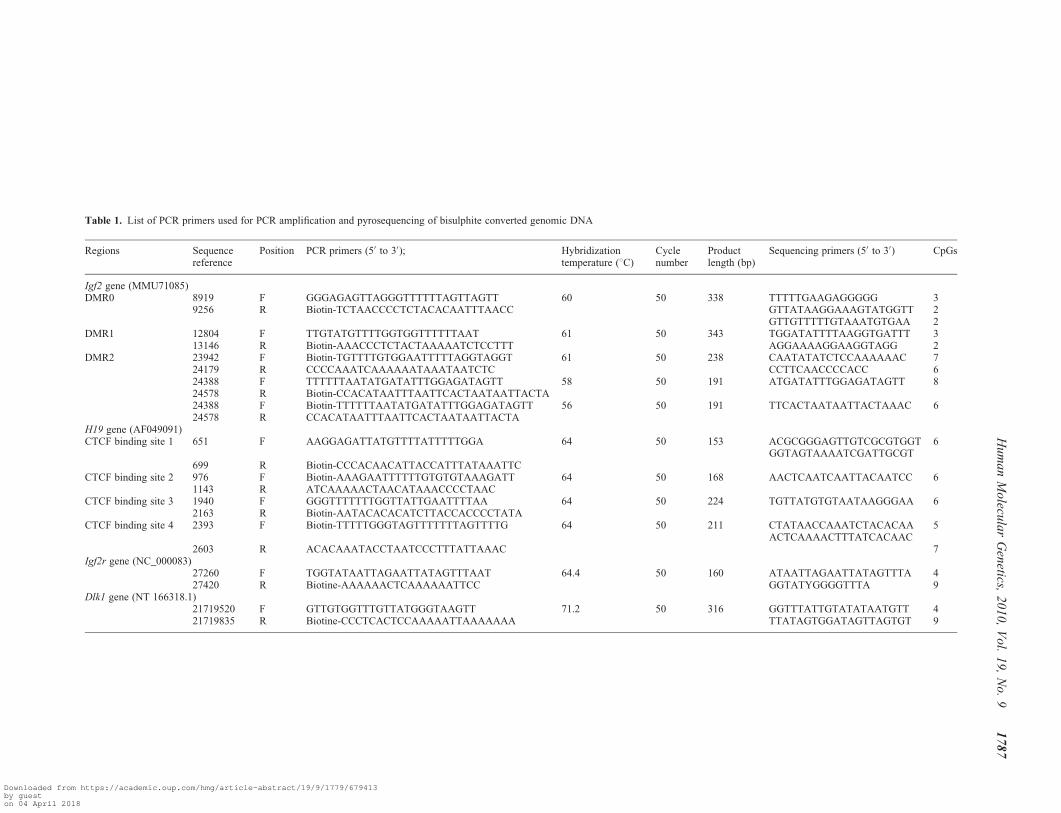
Table 1. List of PCR primers used for PCR amplification and pyrosequencing of bisulphite converted genomic DNA
Regions Sequencereference
Position PCR primers (50 to 30); Hybridizationtemperature (8C)
Cyclenumber
Productlength (bp)
Sequencing primers (50 to 30) CpGs
Igf2 gene (MMU71085)DMR0 8919 F GGGAGAGTTAGGGTTTTTTAGTTAGTT 60 50 338 TTTTTGAAGAGGGGG 3
9256 R Biotin-TCTAACCCCTCTACACAATTTAACC GTTATAAGGAAAGTATGGTT 2GTTGTTTTTGTAAATGTGAA 2
DMR1 12804 F TTGTATGTTTTGGTGGTTTTTTAAT 61 50 343 TGGATATTTTAAGGTGATTT 313146 R Biotin-AAACCCTCTACTAAAAATCTCCTTT AGGAAAAGGAAGGTAGG 2
DMR2 23942 F Biotin-TGTTTTGTGGAATTTTTAGGTAGGT 61 50 238 CAATATATCTCCAAAAAAC 724179 R CCCCAAATCAAAAAATAAATAATCTC CCTTCAACCCCACC 624388 F TTTTTTAATATGATATTTGGAGATAGTT 58 50 191 ATGATATTTGGAGATAGTT 824578 R Biotin-CCACATAATTTAATTCACTAATAATTACTA24388 F Biotin-TTTTTTAATATGATATTTGGAGATAGTT 56 50 191 TTCACTAATAATTACTAAAC 624578 R CCACATAATTTAATTCACTAATAATTACTA
H19 gene (AF049091)CTCF binding site 1 651 F AAGGAGATTATGTTTTATTTTTGGA 64 50 153 ACGCGGGAGTTGTCGCGTGGT
GGTAGTAAAATCGATTGCGT6
699 R Biotin-CCCACAACATTACCATTTATAAATTCCTCF binding site 2 976 F Biotin-AAAGAATTTTTTGTGTGTAAAGATT 64 50 168 AACTCAATCAATTACAATCC 6
1143 R ATCAAAAACTAACATAAACCCCTAACCTCF binding site 3 1940 F GGGTTTTTTTGGTTATTGAATTTTAA 64 50 224 TGTTATGTGTAATAAGGGAA 6
2163 R Biotin-AATACACACATCTTACCACCCCTATACTCF binding site 4 2393 F Biotin-TTTTTGGGTAGTTTTTTTAGTTTTG 64 50 211 CTATAACCAAATCTACACAA
ACTCAAAACTTTATCACAAC5
2603 R ACACAAATACCTAATCCCTTTATTAAAC 7Igf2r gene (NC_000083)
27260 F TGGTATAATTAGAATTATAGTTTAAT 64.4 50 160 ATAATTAGAATTATAGTTTA 427420 R Biotine-AAAAAACTCAAAAAATTCC GGTATYGGGGTTTA 9
Dlk1 gene (NT 166318.1)21719520 F GTTGTGGTTTGTTATGGGTAAGTT 71.2 50 316 GGTTTATTGTATATAATGTT 421719835 R Biotine-CCCTCACTCCAAAAATTAAAAAAA TTATAGTGGATAGTTAGTGT 9
Hu
ma
nM
olecu
lar
Gen
etics,2
01
0,
Vo
l.1
9,
No
.9
17
87
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

effects, two different media were tested: the M16 medium andthe sequential G1/G2 medium (61) containing amino acids.The embryo culture was conducted to blastocyst stage at378C and 5.5% CO2. The culture medium was changedevery day.
Collection of in vivo fertilized eggs. F1 females were matedindividually with F1 males after hCG injection. The followingmorning, females were checked for vaginal copulation plug.One cell stage zygotes were retrieved by flushing the oviductswith M2 medium supplemented with BSA (7 mg/ml) 21–23 hpost-hCG. After mechanical decoronization and washes,zygotes selected by the presence of two pronuclei were cul-tured in the two different culture media under similar con-ditions as described for in vitro fertilization.
Collection of blastocysts. Embryos at the blastocyst stage wereobtained from F1 females, mated individually with F1 malesas described above, by flushing the uterus 3.5 days p.c. withM2 medium supplemented with BSA (7 mg/ml).
Embryo transfer recipients. F1 (C57BL/6/CBA) females of atleast 6 weeks of age were mated to vasectomized F1 males1 day prior to embryo transfer. The morning after mating,females were checked for the presence of a vaginal plug andthis was considered as day 0.5 of pseudopregnancy. A totalof 40, 36 and 51 embryos (at early, non-expanded and fullyexpanded blastocyst stages, classification described in our pre-vious study) (18) were transferred to the uteri of pseudopreg-nant females of groups A, B and C, respectively, onpseudopregnant day 3.5 according to standard procedures(33). On day 10.5, females were sacrificed. Concepti wereimmediately collected and the developmental stage was esti-mated following the recommendations of Theiler et al. (23).The placenta, yolk sac and embryos were mechanically separ-ated. The placenta was cut in half, whereas the embryo wasnot divided. Both tissues (embryonic tissues and one half ofplacental tissue) were used for DNA methylation analysis.The other half of each placenta, reserved for expression analy-sis, was dropped in 250 ml of Trizol reagent (Invitrogen LifeTechnology, Cergy, France), snap-frozen and stored at2808C until further use. The 21, 12 and 14 concepti from 4,4 and 5 females were used for the experiments in groups A,B and C, respectively.
Methylation assays
DNA extraction. Nucleic acid isolation: genomic DNA wasisolated from placental and fetal tissues by incubation withproteinase K (0.2 mg/ml) in lysis saline buffer following stan-dard methods.
The bisulphite conversion of genomic DNA (1 mg) andpyrosequencing analysis were performed as previouslydescribed (34,62). Table 1 lists the accession numbers andnucleotide positions of the analyzed regions, the PCRprimers and annealing temperatures used, the size of thePCR products and the number of CpGs analyzed. Primersfor PCR amplification and pyrosequencing were purchasedfrom Biotez (Buch, Germany). Briefly, 50 ng of bisulphiteconverted DNA was added as template into PCR buffer with
1 mM MgCl2, 200 mM dNTP, 0.4 mM of each forward andreverse biotinylated primer and 2.0 U Platinum Taq polymer-ase (Invitrogen Life Technology). The PCR program consistedof a denaturing step of 4 min at 958C followed by 50 cycles of30 s at 958C, 30 s at hybridization temperature (Table 1) and20 s at 728C, with a final extension of 5 min at 728C. Purifi-cation of the PCR product with streptavidin Sepharosew HPbeads (GE Healthcare, Uppsala, Sweden) and hybridizationof the biotinylated PCR products and the sequencing primerwere conducted following the PSQ96 sample preparationguide using a vacuum filtration sample transfer device (Pyro-sequencing AB, Uppsala, Sweden). Sequencing was per-formed on a PSQ 96MA system with the PyroGold SQAreagent kit according to the manufacturer’s instructions (Pyr-osequencing AB). For one DMR, all the samples were ana-lyzed inside the same experiment. The methylation analysiswas performed in duplicate. The mean and SD were calculatedusing the individual methylation values (with n ¼ 21, n ¼ 12and n ¼ 14 placentas and embryos, from groups A, B and C,respectively). Variables in the study groups were comparedby using two-tailed t-tests, with P , 0.05 as the limit of sig-nificance.
Expression analysis of imprinted genes
Total RNA was extracted from a pool of five placental samplescoming from at least 2 L with Trizol (Invitrogen Life Technol-ogy) according to the manufacturer’s instructions. RT–PCRanalysis of total DNase-treated RNA was carried out withrandom hexamer oligonucleotides in three independentreverse transcription reactions. The expression analyses wereperformed as previously described (21). Sequences of theprimers used for the determination of imprinted gene expressionlevels were those described by Varrault et al. (21). The selectionof appropriate housekeeping genes was performed with geNorm(26). The level of expression of each imprinted gene (X) wasnormalized to the geometric mean of expression levelsof three housekeeping genes (Gapdh, Mrpl32 andTbp2), according to the formula: X/geometric mean (R1, R2,R3) ¼ 2(Ct[X]-arithmetic mean [Ct(R1, Ct(R2), CT(R3)]), where Ct isthe threshold cycle, and R1, R2, R3 are the three housekeepinggenes. Data were analyzed using a non-parametric Kruskal–Wallis one-way analysis of variance with Benjamini–Hochbergcorrection (FDR 5%) for multiple testing, as implemented in theMeV 4.2 package (www.tm4.org), followed by post hoc pairedcomparisons.
Statistics
The x2 test was used for the comparison of binary variablesand continuous variables were compared using the indepen-dent Student’s t-test, when appropriate. The significancelevel was set at 5% (P , 0.05).
SUPPLEMENTARY MATERIAL
Supplementary Material is available at HMG online.
Conflict of Interest statement. None declared.
1788 Human Molecular Genetics, 2010, Vol. 19, No. 9
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

FUNDING
This research was supported by grants from the ‘Fond d’Aidea la Recherche d’Organon’ (ALA-570/06) and from the‘Agence de BioMedecine’ (06.21.DPM OMA 09), France.
REFERENCES
1. Andersen, A.N., Goossens, V., Ferraretti, A.P., Bhattacharya, S.,Felberbaum, R., de Mouzon, J. and Nygren, K.G. (2008) Assistedreproductive technology in Europe, 2004: results generated fromEuropean registers by ESHRE. Hum. Reprod, 23, 756–771.
2. Wright, V.C., Chang, J., Jeng, G. and Macaluso, M. (2008) Assistedreproductive technology surveillance—United States, 2005. MMWRSurveill. Summ., 57, 1–23.
3. Hansen, M., Kurinczuk, J.J., Bower, C. and Webb, S. (2002) The risk ofmajor birth defects after intracytoplasmic sperm injection and in vitrofertilization. N. Engl. J. Med., 346, 725–730.
4. Schieve, L.A., Meikle, S.F., Ferre, C., Peterson, H.B., Jeng, G. andWilcox, L.S. (2002) Low and very low birth weight in infants conceivedwith use of assisted reproductive technology. N. Engl. J. Med., 346, 731–737.
5. DeBaun, M.R., Niemitz, E.L. and Feinberg, A.P. (2003) Association ofin vitro fertilization with Beckwith–Wiedemann syndrome and epigeneticalterations of LIT1 and H19. Am. J. Hum. Genet., 72, 156–160.
6. Gicquel, C., Gaston, V., Mandelbaum, J., Siffroi, J.P., Flahault, A. and LeBouc, Y. (2003) In vitro fertilization may increase the risk of Beckwith–Wiedemann syndrome related to the abnormal imprinting of the KCN1OTgene. Am. J. Hum. Genet., 72, 1338–1341.
7. Maher, E.R., Brueton, L.A., Bowdin, S.C., Luharia, A., Cooper, W., Cole,T.R., Macdonald, F., Sampson, J.R., Barratt, C.L., Reik, W. et al. (2003)Beckwith–Wiedemann syndrome and assisted reproduction technology(ART). J. Med. Genet., 40, 62–64.
8. Ludwig, H. (2005) Archives of gynecology and obstetrics: 135 years.Arch. Gynecol. Obstet., 271, 1–5.
9. Halliday, J., Oke, K., Breheny, S., Algar, E. and J. Amor, D. (2004)Beckwith–Wiedemann syndrome and IVF: a case–control study.Am. J. Hum. Genet., 75, 526–528.
10. Orstavik, K.H., Eiklid, K., van der Hagen, C.B., Spetalen, S., Kierulf, K.,Skjeldal, O. and Buiting, K. (2003) Another case of imprinting defect in agirl with Angelman syndrome who was conceived by intracytoplasmicsemen injection. Am. J. Hum. Genet., 72, 218–219.
11. Cox, G.F., Burger, J., Lip, V., Mau, U.A., Sperling, K., Wu, B.L. andHorsthemke, B. (2002) Intracytoplasmic sperm injection may increase therisk of imprinting defects. Am. J. Hum. Genet., 71, 162–164.
12. Bliek, J., Terhal, P., van den Bogaard, M.J., Maas, S., Hamel, B.,Salieb-Beugelaar, G., Simon, M., Letteboer, T., van der Smagt, J., Kroes,H. et al. (2006) Hypomethylation of the H19 gene causes not only Silver–Russell syndrome (SRS) but also isolated asymmetry or an SRS-likephenotype. Am. J. Hum. Genet., 78, 604–614.
13. Kagami, M., Nagai, T., Fukami, M., Yamazawa, K. and Ogata, T. (2007)Silver–Russell syndrome in a girl born after in vitro fertilization: partialhypermethylation at the differentially methylated region of PEG1/MEST.J. Assist. Reprod. Genet., 24, 131–136.
14. Holmes, R. and Soloway, P.D. (2006) Regulation of imprinted DNAmethylation. Cytogenet. Genome Res., 113, 122–129.
15. Lewis, A. and Reik, W. (2006) How imprinting centres work. Cytogenet.Genome Res., 113, 81–89.
16. Beechey, C.V., Blake, A. and Peters, J. (2008) Mouse imprinting data andreferences. http://www.har.mrc.ac.uk/research/genomic_imprinting/.
17. Reik, W. and Walter, J. (2001) Evolution of imprinting mechanisms: thebattle of the sexes begins in the zygote. Nat. Genet., 27, 255–256.
18. Fauque, P., Jouannet, P., Lesaffre, C., Ripoche, M.A., Dandolo, L.,Vaiman, D. and Jammes, H. (2007) Assisted Reproductive Technologyaffects developmental kinetics, H19 Imprinting Control Regionmethylation and H19 gene expression in individual mouse embryos. BMCDev. Biol., 7, 116.
19. Doherty, A.S., Mann, M.R., Tremblay, K.D., Bartolomei, M.S. andSchultz, R.M. (2000) Differential effects of culture on imprinted H19expression in the preimplantation mouse embryo. Biol. Reprod., 62,1526–1535.
20. Sasaki, H., Ferguson-Smith, A.C., Shum, A.S., Barton, S.C. and Surani,M.A. (1995) Temporal and spatial regulation of H19 imprinting in normaland uniparental mouse embryos. Development, 121, 4195–4202.
21. Varrault, A., Gueydan, C., Delalbre, A., Bellmann, A., Houssami, S.,Aknin, C., Severac, D., Chotard, L., Kahli, M., Le Digarcher, A. et al.
(2006) Zac1 regulates an imprinted gene network critically involved in thecontrol of embryonic growth. Dev. Cell, 11, 711–722.
22. Gabory, A., Ripoche, M.A., Le Digarcher, A., Watrin, F., Ziyyat, A.,Forne, T., Jammes, H., Ainscough, J.F., Surani, M.A., Journot, L. et al.(2009) H19 acts as a trans regulator of the imprinted gene networkcontrolling growth in mice. Development, 136, 3413–3421.
23. Nagy, A., Vintersten, K. and Behringer, R. (2003) Manipulating the
Mouse Embryo: a Laboratory Manual., 3rd edn.24. Laissue, P., Burgio, G., l’Hote, D., Renault, G., Marchiol-Fournigault, C.,
Fradelizi, D., Fellous, M., Serres, C., Montagutelli, X., Monget, P. et al.(2009) Identification of Quantitative Trait Loci responsible for embryoniclethality in mice assessed by ultrasonography. Int. J. Dev. Biol., 53,623–629.
25. Tost, J. and Gut, I.G. (2007) DNA methylation analysis bypyrosequencing. Nat. Protoc., 2, 2265–2275.
26. Vandesompele, J., De Preter, K., Pattyn, F., Poppe, B., Van Roy, N.,De Paepe, A. and Speleman, F. (2002) Accurate normalization ofreal-time quantitative RT–PCR data by geometric averaging of multipleinternal control genes. Genome Biol., 3, RESEARCH0034.
27. Leighton, P.A., Ingram, R.S., Eggenschwiler, J., Efstratiadis, A. andTilghman, S.M. (1995) Disruption of imprinting caused by deletion of theH19 gene region in mice. Nature, 375, 34–39.
28. Eggenschwiler, J., Ludwig, T., Fisher, P., Leighton, P.A., Tilghman, S.M.and Efstratiadis, A. (1997) Mouse mutant embryos overexpressing IGF-IIexhibit phenotypic features of the Beckwith–Wiedemann and Simpson–Golabi–Behmel syndromes. Genes Dev., 11, 3128–3142.
29. Reik, W., Constancia, M., Fowden, A., Anderson, N., Dean, W.,Ferguson-Smith, A., Tycko, B. and Sibley, C. (2003) Regulation of supplyand demand for maternal nutrients in mammals by imprinted genes.J. Physiol., 547, 35–44.
30. Khosla, S., Dean, W., Brown, D., Reik, W. and Feil, R. (2001) Culture ofpreimplantation mouse embryos affects fetal development and theexpression of imprinted genes. Biol. Reprod., 64, 918–926.
31. Li, T., Vu, T.H., Ulaner, G.A., Littman, E., Ling, J.Q., Chen, H.L., Hu,J.F., Behr, B., Giudice, L. and Hoffman, A.R. (2005) IVF results in denovo DNA methylation and histone methylation at an Igf2–H19imprinting epigenetic switch. Mol. Hum. Reprod., 11, 631–640.
32. Mann, M.R., Chung, Y.G., Nolen, L.D., Verona, R.I., Latham, K.E. andBartolomei, M.S. (2003) Disruption of imprinted gene methylation andexpression in cloned preimplantation stage mouse embryos. Biol. Reprod.,69, 902–914.
33. Rivera, R.M., Stein, P., Weaver, J.R., Mager, J., Schultz, R.M. andBartolomei, M.S. (2008) Manipulations of mouse embryos prior toimplantation result in aberrant expression of imprinted genes on day 9.5 ofdevelopment. Hum. Mol. Genet., 17, 1–14.
34. Tost, J., El Abdalaoui, H. and Gut, I.G. (2006) Serial pyrosequencing forquantitative DNA methylation analysis. Biotechniques, 40, 721–722.724, 726.
35. Moore, T., Constancia, M., Zubair, M., Bailleul, B., Feil, R., Sasaki, H.and Reik, W. (1997) Multiple imprinted sense and antisense transcripts,differential methylation and tandem repeats in a putative imprintingcontrol region upstream of mouse Igf2. Proc. Natl Acad. Sci. USA, 94,12509–12514.
36. Murrell, A., Ito, Y., Verde, G., Huddleston, J., Woodfine, K., Silengo,M.C., Spreafico, F., Perotti, D., De Crescenzo, A., Sparago, A. et al.(2008) Distinct methylation changes at the IGF2–H19 locus in congenitalgrowth disorders and cancer. PLoS ONE, 3, e1849.
37. Lopes, S., Lewis, A., Hajkova, P., Dean, W., Oswald, J., Forne, T.,Murrell, A., Constancia, M., Bartolomei, M., Walter, J. et al. (2003)Epigenetic modifications in an imprinting cluster are controlled by ahierarchy of DMRs suggesting long-range chromatin interactions. Hum.Mol. Genet., 12, 295–305.
38. Umlauf, D., Goto, Y., Cao, R., Cerqueira, F., Wagschal, A., Zhang, Y. andFeil, R. (2004) Imprinting along the Kcnq1 domain on mousechromosome 7 involves repressive histone methylation and recruitment ofPolycomb group complexes. Nat. Genet., 36, 1296–1300.
39. Mann, M.R., Lee, S.S., Doherty, A.S., Verona, R.I., Nolen, L.D., Schultz,R.M. and Bartolomei, M.S. (2004) Selective loss of imprinting in the
Human Molecular Genetics, 2010, Vol. 19, No. 9 1789
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018

placenta following preimplantation development in culture. Development,131, 3727–3735.
40. Buffat, C., Boubred, F., Mondon, F., Chelbi, S.T., Feuerstein, J.M.,Lelievre-Pegorier, M., Vaiman, D. and Simeoni, U. (2007) Kidney geneexpression analysis in a rat model of intrauterine growth restrictionreveals massive alterations of coagulation genes. Endocrinology, 148,5549–5557.
41. Leighton, P.A., Saam, J.R., Ingram, R.S., Stewart, C.L. and Tilghman,S.M. (1995) An enhancer deletion affects both H19 and Igf2 expression.Genes Dev., 9, 2079–2089.
42. Poirier, F., Chan, C.T., Timmons, P.M., Robertson, E.J., Evans, M.J. andRigby, P.W. (1991) The murine H19 gene is activated during embryonicstem cell differentiation in vitro and at the time of implantation in thedeveloping embryo. Development, 113, 1105–1114.
43. DeChiara, T.M., Efstratiadis, A. and Robertson, E.J. (1990) Agrowth-deficiency phenotype in heterozygous mice carrying aninsulin-like growth factor II gene disrupted by targeting. Nature, 345,78–80.
44. Coan, P.M., Fowden, A.L., Constancia, M., Ferguson-Smith, A.C.,Burton, G.J. and Sibley, C.P. (2008) Disproportional effects of Igf2knockout on placental morphology and diffusional exchangecharacteristics in the mouse. J. Physiol., 586, 5023–5032.
45. Ludwig, T., Eggenschwiler, J., Fisher, P., D’Ercole, A.J., Davenport, M.L.and Efstratiadis, A. (1996) Mouse mutants lacking the type 2 IGF receptor(IGF2R) are rescued from perinatal lethality in Igf2 and Igf1r nullbackgrounds. Dev. Biol., 177, 517–535.
46. Lau, M.M., Stewart, C.E., Liu, Z., Bhatt, H., Rotwein, P. and Stewart,C.L. (1994) Loss of the imprinted IGF2/cation-independent mannose6-phosphate receptor results in fetal overgrowth and perinatal lethality.Genes Dev., 8, 2953–2963.
47. Wang, Z.Q., Fung, M.R., Barlow, D.P. and Wagner, E.F. (1994)Regulation of embryonic growth and lysosomal targeting by the imprintedIgf2/Mpr gene. Nature, 372, 464–467.
48. Fortier, A.L., Lopes, F.L., Darricarrere, N., Martel, J. and Trasler, J.M.(2008) Superovulation alters the expression of imprinted genesin the midgestation mouse placenta. Hum. Mol. Genet., 17,1653–1665.
49. Constancia, M., Angiolini, E., Sandovici, I., Smith, P., Smith, R., Kelsey,G., Dean, W., Ferguson-Smith, A., Sibley, C.P., Reik, W. et al. (2005)Adaptation of nutrient supply to fetal demand in the mouse involvesinteraction between the Igf2 gene and placental transporter systems. Proc.
Natl Acad. Sci. USA, 102, 19219–19224.
50. Li, L., Keverne, E.B., Aparicio, S.A., Ishino, F., Barton, S.C. and Surani,M.A. (1999) Regulation of maternal behavior and offspring growth bypaternally expressed Peg3. Science, 284, 330–333.
51. Takahashi, K., Kobayashi, T. and Kanayama, N. (2000) p57(Kip2)regulates the proper development of labyrinthine and spongiotrophoblasts.Mol. Hum. Reprod., 6, 1019–1025.
52. Kawahara, M., Wu, Q., Yaguchi, Y., Ferguson-Smith, A.C. and Kono, T.(2006) Complementary roles of genes regulated by two paternallymethylated imprinted regions on chromosomes 7 and 12 in mouseplacentation. Hum. Mol. Genet., 15, 2869–2879.
53. Sekita, Y., Wagatsuma, H., Nakamura, K., Ono, R., Kagami, M.,Wakisaka, N., Hino, T., Suzuki-Migishima, R., Kohda, T., Ogura, A. et al.(2008) Role of retrotransposon-derived imprinted gene, Rtl1,in the feto-maternal interface of mouse placenta. Nat. Genet., 40,243–248.
54. Smallwood, A., Papageorghiou, A., Nicolaides, K., Alley, M.K., Jim, A.,Nargund, G., Ojha, K., Campbell, S. and Banerjee, S. (2003) Temporalregulation of the expression of syncytin (HERV-W), maternallyimprinted PEG10, and SGCE in human placenta. Biol. Reprod., 69,286–293.
55. Ono, R., Shiura, H., Aburatani, H., Kohda, T., Kaneko-Ishino, T. andIshino, F. (2003) Identification of a large novel imprinted gene cluster onmouse proximal chromosome 6. Genome Res., 13, 1696–1705.
56. Iacob, D., Cai, J., Tsonis, M., Babwah, A., Chakraborty, C., Bhattacharjee,R.N. and Lala, P.K. (2008) Decorin-mediated inhibition of proliferationand migration of the human trophoblast via different tyrosine kinasereceptors. Endocrinology, 149, 6187–6197.
57. Sandell, L.L., Guan, X.J., Ingram, R. and Tilghman, S.M. (2003) Gatm, acreatine synthesis enzyme, is imprinted in mouse placenta. Proc. NatlAcad. Sci. USA, 100, 4622–4627.
58. McMinn, J., Wei, M., Schupf, N., Cusmai, J., Johnson, E.B., Smith, A.C.,Weksberg, R., Thaker, H.M. and Tycko, B. (2006) Unbalanced placentalexpression of imprinted genes in human intrauterine growth restriction.Placenta, 27, 540–549.
59. Mizuno, Y., Sotomaru, Y., Katsuzawa, Y., Kono, T., Meguro, M.,Oshimura, M., Kawai, J., Tomaru, Y., Kiyosawa, H., Nikaido, I. et al.(2002) Asb4, Ata3, and Dcn are novel imprinted genes identified byhigh-throughput screening using RIKEN cDNA microarray. Biochem.Biophys. Res. Commun., 290, 1499–1505.
60. Xu, G., Guimond, M.J., Chakraborty, C. and Lala, P.K. (2002) Control ofproliferation, migration, and invasiveness of human extravilloustrophoblast by decorin, a decidual product. Biol. Reprod., 67, 681–689.
61. Guerif, F., Le Gouge, A., Giraudeau, B., Poindron, J., Bidault, R., Gasnier,O. and Royere, D. (2007) Limited value of morphological assessment atdays 1 and 2 to predict blastocyst development potential: a prospectivestudy based on 4042 embryos. Hum. Reprod., 22, 1973–1981.
62. Dupont, J.M., Tost, J., Jammes, H. and Gut, I.G. (2004) De novoquantitative bisulfite sequencing using the pyrosequencing technology.Anal. Biochem., 333, 119–127.
1790 Human Molecular Genetics, 2010, Vol. 19, No. 9
Downloaded from https://academic.oup.com/hmg/article-abstract/19/9/1779/679413by gueston 04 April 2018