Embed Size (px)
Citation preview

143
Modular Protein Domains. Edited by G. Cesareni, M. Gimona, M. Sudol, M. YaffeCopyright © 2005 WILEY-VCH Verlag GmbH & Co. KGaA, WeinheimISBN: 3-527-30813-X
7The FHA Domain
Daniel Durocher
7.1Introduction
One of the major and universal roles of reversible protein phosphorylation is theregulation of protein–protein interactions [1]. Protein phosphorylation can influenceprotein–protein interactions in multiple ways. For example, phosphorylation of aserine, threonine, or tyrosine residue can result in the masking of an epitopenecessary for the interaction of a protein with another. Conversely, phosphorylationcan induce a conformational change that allows two proteins to interact. Finally,the phosphorylated residue can be an essential and integral part of an epitope usedin a protein–protein interaction. This latter mode of influencing protein functionvia protein phosphorylation is a recurring theme in many cellular processes and inalmost all signal transduction pathways of eukaryotes [2, 3]. Phospho-dependentprotein–protein interactions are so versatile and powerful that, during the courseof evolution, protein domains specialized in mediating such interactions have arisenmultiple times [2, 3]. The main advantage of using protein modules resides intheir ‘transferability’, since they are independently folding units for which the codingDNA can be shuffled around the genome [4]. This shuffling results in the generationof multidomain proteins with a diverse array of functions. The class of phospho-recognition modules is composed of the SH2, PTB, BRCT, and WW domains,Polo-box, 14-3-3 proteins [3, 5–8], and the subject of this chapter, the forkhead-associated (FHA) domain (reviewed in [9–12]).
The FHA domain was discovered in 1995 by Hofmann and Bucher, whorecognized a protein sequence motif in a subset of forkhead-type transcriptionfactors [13]. A set of approximately 20 proteins containing this domain signaturewas identified by profile searches, and a first consensus alignment of the FHAdomain was derived [13]. Today, using other methods of database searching suchas hidden Markov models (HMMs) and iterative BLAST searches, the FHA domaincan be found in more than 400 different proteins (see SMART and PFAM databases)from eukaryotic, eubacterial, and some archeal species. For humans, the ENSEMBLdatabase lists 23 unique FHA-containing proteins encoded by the human genome.
1246vch07.pmd 10.09.2004, 14:41143
144 7 The FHA Domain
However, this number is likely to be higher, given that some additional FHA-containing proteins are clearly being missed by current HMM searches (Table 7.1).For example, the mammalian DNA repair enzyme polynucleotide kinase (PNK)was recently proposed by Caldecott and colleagues to have a divergent FHA domainthat shares homology with the histidine triad-containing protein, aprataxin (APTX)[14]. The gene encoding APTX is mutated in the neurological disease ataxia withoccular apraxia (AOA) [15, 16] and, given the high degree of similarity betweenAPTX and PNK, is proposed to be involved in the repair of single-strand breaks asis PNK. Recent work from our laboratory suggests that the divergent FHA domainof PNK does function as a phosphopeptide-binding domain (A. Koch, R. Agyei,and D. Durocher, unpublished data). Interestingly, iterative PSI-BLAST searchesreadily identify a third member of this subgroup (MGC47799, gi:27734905) thus,in total, adding three new FHA-containing proteins to the 23 reported in theENSEMBL database. Since the three noted FHA domains are not detected by manyof the bioinformatics tools used at present, it is tempting to wonder how manyadditional FHA domains exist unnoticed in protein databases.
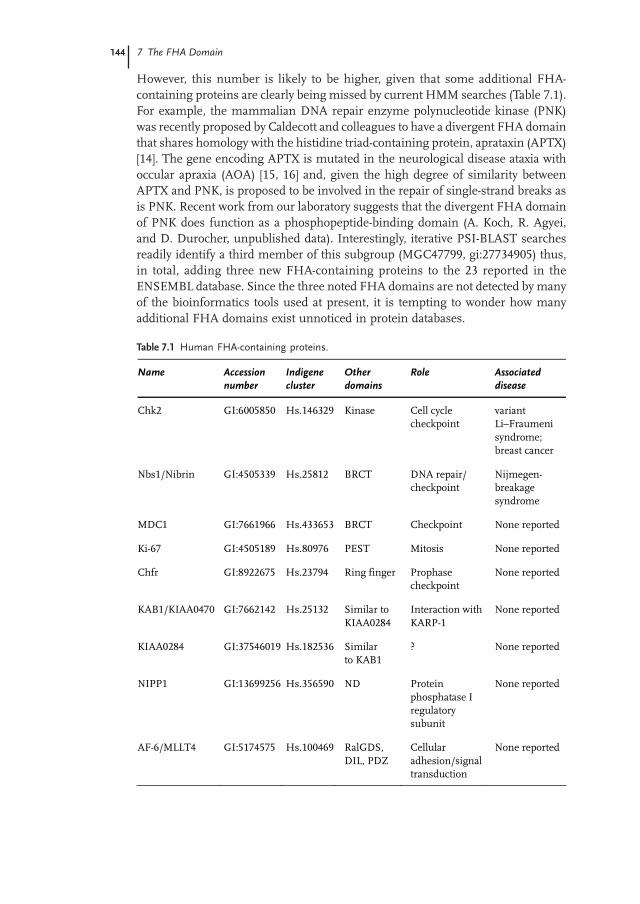
Table 7.1 Human FHA-containing proteins.
Name Accession number
Indigene cluster
Other domains
Role Associated disease
Chk2 GI:6005850 Hs.146329 Kinase Cell cycle checkpoint
variant Li–Fraumeni syndrome; breast cancer
Nbs1/Nibrin GI:4505339 Hs.25812 BRCT DNA repair/ checkpoint
Nijmegen-breakage syndrome
MDC1 GI:7661966 Hs.433653 BRCT Checkpoint None reported
Ki-67 GI:4505189 Hs.80976 PEST Mitosis None reported
Chfr GI:8922675 Hs.23794 Ring finger Prophase checkpoint
None reported
KAB1/KIAA0470 GI:7662142 Hs.25132 Similar to KIAA0284
Interaction with KARP-1
None reported
KIAA0284 GI:37546019 Hs.182536 Similar to KAB1
? None reported
NIPP1 GI:13699256 Hs.356590 ND Protein phosphatase I regulatory subunit
None reported
AF-6/MLLT4 GI:5174575 Hs.100469 RalGDS, DIL, PDZ
Cellular adhesion/signal transduction
None reported
1246vch07.pmd 10.09.2004, 14:41144
1457.1 Introduction
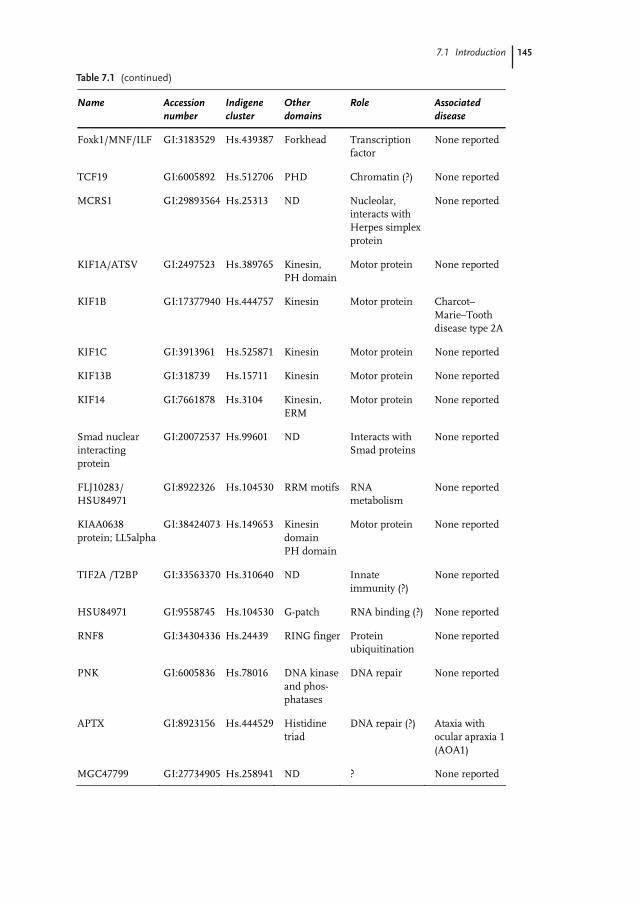
Table 7.1 (continued)
Name Accession number
Indigene cluster
Other domains
Role Associated disease
Foxk1/MNF/ILF GI:3183529 Hs.439387 Forkhead Transcription factor
None reported
TCF19 GI:6005892 Hs.512706 PHD Chromatin (?) None reported
MCRS1 GI:29893564 Hs.25313 ND Nucleolar, interacts with Herpes simplex protein
None reported
KIF1A/ATSV GI:2497523 Hs.389765 Kinesin, PH domain
Motor protein None reported
KIF1B GI:17377940 Hs.444757 Kinesin Motor protein Charcot–Marie–Tooth disease type 2A
KIF1C GI:3913961 Hs.525871 Kinesin Motor protein None reported
KIF13B GI:318739 Hs.15711 Kinesin Motor protein None reported
KIF14 GI:7661878 Hs.3104 Kinesin, ERM
Motor protein None reported
Smad nuclear interacting protein
GI:20072537 Hs.99601
ND Interacts with Smad proteins
None reported
FLJ10283/ HSU84971
GI:8922326 Hs.104530 RRM motifs RNA metabolism
None reported
KIAA0638 protein; LL5alpha
GI:38424073 Hs.149653
Kinesin domain PH domain
Motor protein None reported
TIF2A /T2BP GI:33563370 Hs.310640 ND Innate immunity (?)
None reported
HSU84971 GI:9558745 Hs.104530 G-patch RNA binding (?) None reported
RNF8 GI:34304336 Hs.24439
RING finger Protein ubiquitination
None reported
PNK GI:6005836 Hs.78016
DNA kinase and phos-phatases
DNA repair None reported
APTX GI:8923156
Hs.444529
Histidine triad
DNA repair (?) Ataxia with ocular apraxia 1 (AOA1)
MGC47799 GI:27734905 Hs.258941 ND ? None reported
1246vch07.pmd 10.09.2004, 14:41145

146 7 The FHA Domain
Prior to the bioinformatics work that led to the recognition of the FHA domainsignature, work from the Walker laboratory [17] gave the first clues as to what theFHA domain might do. In a search for proteins that could interact with the plantreceptor-like protein kinase RLK5, they identified a PP2C-type phosphatase thatthey named KAPP, for kinase-associated protein phosphatase [17]. The region ofKAPP required for the interaction was termed the kinase-interaction (KI) domain,and it was shown that only the phosphorylated form of the kinase can be bound byKAPP [17]. This observation suggested a mechanism analogous to that of othereukaryotic signaling systems, in which tyrosine-phosphorylated receptor tyrosinekinases are bound by effectors and regulators via phospho-dependent interactions.
Following the demonstration by Hoffmann and Bucher [13] that the minimal KIregion of KAPP has homology to the FHA domain, Walker and colleagues [18]sought to investigate the relevance of the FHA homology for KI activity. Theyemployed site-directed mutagenesis to disrupt amino acid residues conservedamong FHA domains. Mutation of these conserved residues abolished KI activity,suggesting that the FHA domain of KAPP mediates phospho-dependent protein–protein interactions [18]. However, this result did not resolve the important issueof whether these interactions directly involve the recognition of a phosphorylatedresidue or whether they are initiated by a conformational change triggered byphosphorylation.
In a parallel set of experiments, studies on the budding yeast DNA damagecheckpoint brought in the last elements to complete the picture of FHA domainfunction. In eukaryotes, a conserved protein-phosphorylation cascade is initiatedin response to DNA damage detection [19]. This cascade is governed by ATM-likekinases, and the DNA damage signal is propagated downstream to members ofthe Chk1 and Chk2 kinase families (reviewed in [19]). Chk2 and its homologs arerecognizable by the presence of one or two FHA domains flanking a kinase domainof the Ca2+/calmodulin kinase family [20]. In budding yeast, two Rad53 paralogsare also observed: Dun1 and Mek1/Mre4 are both protein kinases that play roles inthe mitotic and meiotic checkpoint responses, and for which the FHA domainplays an essential role [21, 22]. Early on, the presence of FHA domains on Rad53suggested a role in protein–protein interactions and, consistent with this, Rad53was found to interact with Rad9 following DNA damage [23–26]. The interactionbetween Rad9 and Rad53 is dependent on Rad9 phosphorylation and was shownto require the C-terminal FHA domain (FHA2) of Rad53, although the N-terminalFHA domain (FHA1) has now been shown to play an important role in binding toRad9 ([27] and D. Durocher and F. Sweeney, unpublished data).
To determine whether the FHA-dependent interaction involved direct recognitionof a phospho epitope on Rad9, Durocher et al. [25] tested a series of phosphopeptidesfor their ability to compete with the Rad9–Rad53FHA1 interaction. A phospho-threonine-containing peptide derived from the N terminus of p53, but not itsunphosphorylated counterpart, was found to efficiently compete with the Rad9–Rad53FHA1 interaction [25]. In addition, this p53-derived phosphopeptide was shownto directly bind Rad53FHA1, in experiments making use of a variety of assays suchas pulldowns, surface plasmon resonance, and isothermal titration calorimetry.
1246vch07.pmd 10.09.2004, 14:41146

147
Mutation of the conserved Arg70 and His88 residues, but not of the variable Asp117residue, in Rad53FHA1 abolished its interaction with both synthetic phosphopeptidesand phosphorylated Rad9 [25]. Furthermore, the Rad53FHA1 domain was shown toprotect the phosphorylated residue from phosphatase treatment, suggesting thatthe phosphate moiety is in intimate contact with the FHA domain [25]. Intriguingly,substitution of the phosphothreonine with a phosphoserine residue abolished theinteraction, suggesting that the FHA1 domain of Rad53 is specific for phospho-threonine-containing epitopes [25].
These latter results indicated that the Rad53FHA1 domain recognizes phospho-threonine-containing epitopes. In an effort to test whether most if not all FHAdomains possess the same binding capacity, Durocher et al. [25] tested the bindingof various FHA domains from various species (including plants and bacteria) todegenerate peptide libraries using surface plasmon resonance. This assay unequi-vocally indicated that all the FHA domains tested could recognize the phospho-threonine peptide library but not the unphosphorylated counterpart [25]. Collectively,these results strongly suggested that the FHA domain directly recognizes threonine-phosphorylated epitopes on proteins.
7.2FHA Domain Structure
7.2.1Topology
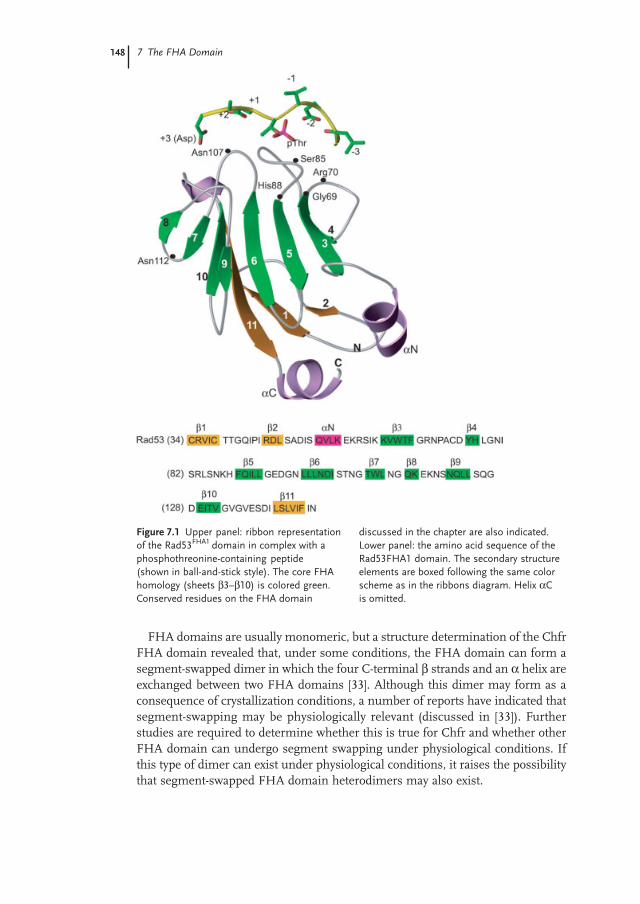
In their bioinformatics work describing the FHA domain, Hoffmann and Bucher[13] predicted that the FHA domain would consist mainly of β strands. Theirprediction was confirmed with the structure determinations of the Rad53 FHA1and FHA2 domains by X-ray crystallography and NMR, respectively [28–30]. Thesestructures revealed that the FHA domain is composed of an 11-stranded β sandwichconsisting of two twisted β sheets (Figure 7.1) [28–30]. To date, four other FHAdomain structures have been reported (Chfr, Ki67, Chk2, KAPP), and the abovetopology is shared by all four of these FHA domains [31–34]. Small variationsbetween structures include the length and the spatial arrangements of the loopsconnecting the β strands and the presence of small helical insertions. Furthermore,the Chfr FHA domain lacks the eleventh β strand, since the domain forms asegment-swapped dimer under crystallographic conditions (discussed in more detailbelow [33]).
The FHA domain, as a folded unit, is approximately 100–120 amino acids long,much larger than the FHA homology region used for profile or HMM searches(which is around 75 amino acids) [28, 29, 35]. The core homology motif encompassessix of the 11 β strands (β3 to β8) and loops β3/4 to β7/8. As explained below, thehomology motif contains the three loops that directly contact the phosphopeptide,as well as the structural framework needed to organize these loops in a spatialarrangement conducive to phosphopeptide binding [28, 31].
7.2 FHA Domain Structure
1246vch07.pmd 10.09.2004, 14:41147
148 7 The FHA Domain
FHA domains are usually monomeric, but a structure determination of the ChfrFHA domain revealed that, under some conditions, the FHA domain can form asegment-swapped dimer in which the four C-terminal β strands and an α helix areexchanged between two FHA domains [33]. Although this dimer may form as aconsequence of crystallization conditions, a number of reports have indicated thatsegment-swapping may be physiologically relevant (discussed in [33]). Furtherstudies are required to determine whether this is true for Chfr and whether otherFHA domain can undergo segment swapping under physiological conditions. Ifthis type of dimer can exist under physiological conditions, it raises the possibilitythat segment-swapped FHA domain heterodimers may also exist.
Figure 7.1 Upper panel: ribbon representationof the Rad53FHA1 domain in complex with aphosphothreonine-containing peptide(shown in ball-and-stick style). The core FHAhomology (sheets β3–β10) is colored green.Conserved residues on the FHA domain
discussed in the chapter are also indicated.Lower panel: the amino acid sequence of theRad53FHA1 domain. The secondary structureelements are boxed following the same colorscheme as in the ribbons diagram. Helix αCis omitted.
1246vch07.pmd 10.09.2004, 14:41148

149
7.2.2FHA–Phosphopeptide Interaction
The FHA–phosphopeptide interaction buries between 600 Å2 (Rad53FHA1) and1000 Å2 (Chk2) of the FHA domain’s solvent-accessible surface [28, 31]. Thephosphopeptide, when bound to the FHA domain, adopts an extended conformationand interacts via the loops connecting the β strands (loops β3/4, β4/5 and β6/7;Figure 7.1 [28, 31]). Interestingly, this mode of binding is reminiscent of the bindingof the complementary determining region (CDR) of antibodies (also β sandwiches)to antigen epitopes [36]. In the latter interaction, the antigen epitope is also boundin an extended conformation and interacts with the CDR via loops connecting theβ strands [36]. As mentioned above, the determinants of phosphopeptide recognitionby the FHA domain are all embedded in the FHA domain homology. Most of theconserved residues within the FHA domain are involved in binding the phospho-peptide. In the Rad53FHA1 domain, six amino acid residues of the phosphopeptide(–2, –1, phosphothreonine, +1, +2, +3) are contacted via either main-chain inter-actions (for residues –2, +1, and +3) or side-chain interactions (for the phospho-threonine and +3 residue). The conserved residues involved in this network ofinteraction includes, on Rad53FHA1: Arg70, Ser82, and Asn107 located on loopsβ3/4, β4/5, and β6/7, respectively (Figure 7.1).
Unlike many phospho-dependent interactions, such as those mediated by SH2domains or 14-3-3 domains, the phosphothreonine residue does not make directsalt interactions with arginine residues but is rather bound through an array ofhydrogen bonds involving Arg70 and Ser85 in Rad53FHA1 [28]. Arg70 in Rad53FHA1
(Arg117 in Chk2) is the only conserved arginine residue involved in a direct contactwith the phosphoamino acid residue, and it contacts it solely via a charged hydrogenbond with the γ oxygen of phosphothreonine. The second conserved interactionbetween the FHA domain and the phosphate moiety is the hydrogen bond betweenthe hydroxyl group of Ser85 and the ε oxygen of the phosphate [28, 31].
The two other functionally important and strongly conserved residues in theFHA domain, Gly69 and His88 in Rad53FHA1, do not play a role in directly contactingthe phosphopeptide, despite being essential for phosphopeptide binding activity[18, 25]. The function of His88 appears to be to stabilize the architecture of thephosphopeptide binding site [28]. In particular, the imidazole ring of His88 tethersthe β4/5 and β6/7 loops through hydrogen bonds with the main-chain atoms ofSer85 [28].
A unique feature of the FHA domain is its ability to discriminate phospho-threonine from phosphoserine residues [25]. Threonine differs from serine onlyby the presence of a γ methyl group on the threonine side-chain. The crystal structureof the Chk2 FHA and Rad53FHA1 domains indicate that phosphothreonine is selectedas a consequence of the docking of the γ methyl group to a small pocket consistingof a conserved Asn residue (Asn166 in Chk2, Asn107 in Rad53) and surroundingamino acid residues, via van der Waals interactions [28, 31]. These interactionsfacilitate the orientation of the phosphothreonine residue in a position suitable forestablishing a network of hydrogen bonds necessary to contact the FHA domain.
7.2 FHA Domain Structure
1246vch07.pmd 10.09.2004, 14:41149

150 7 The FHA Domain
Several FHA domains seem to be selective for particular amino acid residues atthe +3 position relative to the phosphothreonine. For example, Rad53FHA1 selectsfor an Asp residue at this position, whereas the Chk2 FHA domain selects forIle/Leu, and the mycobacterial protein Rv1827 strongly selects for Tyr [28, 30, 37].Some light on the structural basis of this +3 selectivity was shed by comparing thestructures of the Rad53FHA1 and Chk2 FHA domains bound to high-affinity-bindingphosphopeptides [28, 31]. Sequence selectivity in the Rad53FHA1 domain is impartedby the formation of a salt bridge between the guanidinium moiety of Arg83 (β4/5loop) and the carbonyl group of the +3 Asp side-chain [28]. However, in Chk2, thehydrophobic selectivity at +3 can be explained by a complementary shallowhydrophobic pocket formed by the side-chain atoms of Thr138, equivalent to theposition occupied by Arg83 in Rad53, along with the side-chain atoms of Asn166(β6/7 loop) and Ser192/Leu193 (β10/11 loop) [31]. Since the β6/7–β10/11 loopsare poorly conserved in sequence and size among various FHA domains, thesefindings suggest that each FHA domain may display its own specificity.
7.2.3A Second Binding Interface?
Recent work suggests that the FHA domain may be able to bind proteins outsidethe phosphopeptide binding surface or that an ancillary binding surface cooperateswith the phosphopeptide binding activity. A strong argument for an ancillary bindinginterface first emerged from studying the impact of the Ile157–Thr mutation ofChk2 found in families displaying a rare variant of the Li–Fraumeni syndrome[38], a highly penetrant familial cancer disease. Ile157 is located far from thephosphopeptide binding site, at the C-terminal end of the β5/6 loop, some 25 Åaway from the phosphothreonine-binding area [31]. Accordingly, mutation of Ile157to Thr does not affect binding to phosphopeptides in vitro nor does it affect proteinstability in vitro and in vivo [31]. Surprisingly however, this mutation abrogates thebinding of the Chk2 FHA domain to a number of proteins, including Brca1 andCdc25C [31, 39]. The interaction with Brca1 also requires Arg117, indicating thatthe interaction is phospho-dependent [31]. Thus, the Arg117- and Ile157-dependentsurfaces act cooperatively to bind certain proteins. Li et al. [34] proposed that Ile157lies at the heart of a hydrophobic surface that stabilizes an interaction initiated bythe phospho-dependent engagement of Arg117 [31]. Interestingly, evolutionary trace(ET) analysis performed on a large number of FHA domains suggests that thesurface surrounding Ile157 in Chk2 may be functionally conserved in a large numberof FHA domains [34].
The recent characterization Ki-67 and its interaction with hNIFK provides a secondargument supporting the possibility that some FHA domains require additionalbinding surfaces for optimal binding [32]. The human NIFK protein was discoveredas a binding partner of the mitotic/chromosome condensation regulator Ki-67 in atwo-hybrid screen using the FHA domain of Ki-67 [40]. The interaction region ofhNIFK was mapped to a peptide of 44 amino acid residues containing two threonineresidues that are required for binding [32]. Tsai’s group [32] detected high-affinity
1246vch07.pmd 10.09.2004, 14:41150

151
phospho-dependent binding between synthetic peptides corresponding to theminimal interaction region of hNIFK and the Ki-67 FHA domain, as expected [32].Furthermore, they deduced that the phosphothreonine at a position equivalent toThr234 in hNIFK was required for the interaction. However, two unexpected bindingproperties were noted: first, using HSQC NMR experiments, a weak interactionwas detected between the unphosphorylated 44-mer and the Ki-67 FHA domain(with a KD of approximately 100 µM), indicating that a binding determinant outsidethe canonical phosphothreonine-recognition determinants may exist [32]. Further-more, the interaction requires a phosphopeptide longer than the 6 amino acidresidue phosphopeptide normally sufficient for binding to the Rad53, Chk2, Rv1827,and other FHA domains [32]. This requirement for a longer phosphopeptideindicates either that Ki-67 binds structured epitopes or that a second set of bindingdeterminants is present on the hNIFK-derived peptide. In support of the latterpossibility, NMR analysis of chemical shift changes on the Ki-67 FHA domainindicated that residues outside the phosphopeptide-binding loops are involved inbinding the hNIFK 44-mer [32]. This potential binding surface includes the β1/2loop and residues located on β10, a strand neighboring β1 and β2. If this surface isexperimentally confirmed to be required for binding, it would be a distinct surfacefrom the ancillary binding surface of Chk2 which, as mentioned above, is locatedat the C terminus of the β5/6 loop.
7.2.4The FHA Domain is Part of a Domain Superfamily
Structural homology searches place the FHA domain in a superfamily of domainswith similar topology. The topology of the FHA domain is similar to that of theSmad MH2 domain and the recently described IRF-3 activation domain (IAD) [9,10, 41–43]. All three share an 11-stranded β sandwich core of nearly identicaltopology. The IRF IAD and SMAD MH2 domains are more closely related to eachother, having additional structural features that surround the β sandwich not presentin the FHA domain [41].
Interestingly, all three domains can act as transcriptional activators (see below)and two of them (the MH2 and FHA domains) have the capacity to engage inphospho-dependent protein–protein interactions [28, 44]. IRF-3 IAD also engagesin phospho-dependent dimerization, although it is yet unclear if this occurs viaphospho-epitope recognition [41]. As noted previously, this structural and functionalhomology suggests that the FHA/MH2/IAD superfamily shares a common ancestor[9, 44], and it is tempting to speculate that this ancestor may have been involved inthe regulation of transcription. As structural genomics programs progress, novelmembers of this superfamily may emerge. If so, it will be interesting to determineif they too are involved in phospho-dependent interactions and/or in the regulationof transcription.
7.2 FHA Domain Structure
1246vch07.pmd 10.09.2004, 14:41151

152 7 The FHA Domain
7.3Molecular and Signaling Function
To date, the only molecular function of the FHA domain is its phosphopeptidebinding activity. However, this function is used in a number of different ways toregulate the activity of the proteins containing it. Below, I briefly review some ofthe ways FHA-mediated protein–protein interactions are used to accomplish thistask.
7.3.1FHA Domain Can Regulate Protein Localization
Protein interaction modules such as the PDZ and PH domains often regulate theactivity of proteins by localizing them to distinct subcellular compartments [2].The FHA domain is no exception and has also been shown to act in this manner.For example, the FHA-containing protein NIPP1 is a regulatory subunit of proteinphosphatase 1. NIPP1 localizes to nuclear speckles, which are sites of pre-mRNAprocessing where it recruits PP1 [45, 46]. The Bollen group [45] has shown that theFHA domain NIPP1 is critical for targeting the NIPP1 to nuclear speckles and fornuclear retention. The spliceosomal protein that binds the FHA domain of NIPP1remains unknown, but both CDC5L and SAP155 are excellent candidates, becauseboth can interact with NIPP1 in an FHA- and phosphorylation-dependent manner[47, 48]. Interestingly, the interaction between NIPP1 and SAP155 is controlled bymitotic phosphorylation of SAP155 [48]. It will be interesting to determine whetherthe proline residue, characteristic of mitotic phosphorylation sites, is an integralpart of the motif recognized by the FHA domain of NIPP1.
Another striking example of FHA-dependent protein localization is the localizationof Dma1 to spindle pole bodies (SPBs). Dma1 is an FHA-containing ring fingerprotein from Schizosaccharomyces pombe analogous to human Chfr, a protein oftenmutated in tumors [49, 50]. Dma1 is a component of the septation-initiation network(S/N), a signaling pathway that links the end of mitosis with cytokinesis (calledseptation in fission yeast) [49, 51]. Elegant work by Guertin and colleagues [52]recently showed that Dma1 localizes to SPBs in an FHA-dependent manner via aninteraction with the spindle pole component Sid4 [52]. The localization of Dma1 toSPBs is essential for its function in regulating SIN by modulating the activity ofPlo1, the fission yeast polo kinase [52]. Interestingly, this function of Dma1 is alsoconserved in humans, since Chfr regulates the destruction and activity of humanPolo kinase, Plk1 [53].
7.3.2FHA Domain Binding to Enzyme Substrates
A number of FHA-containing proteins also display catalytic activity. These includekinases, phosphatases, ubiquitin ligases, protein motors, prolyl isomerases, andoxidases. It is therefore not surprising to find that the FHA domain can serve to
1246vch07.pmd 10.09.2004, 14:41152

153
recruit substrates. The first and most characterized example concerns the plantPP2C-type phosphatase KAPP [17]. As mentioned above, Walker and colleagues[17] identified KAPP in 1994 by virtue of its ability to interact with the phosphorylatedRLK5 receptor, suggesting that KAPP may modulate its activity [17]. More recentgenetic studies in Arabidopsis indicate that KAPP is a negative regulator of CLAVATA1 (CLV1)-dependent signaling. As with the KAPP–RLK5 interaction, KAPP wasfound to directly bind to activated CLV1, a plant receptor-like kinase [54]. Thesegenetic and physical interactions suggest that recruitment of KAPP by activatedreceptors during plant signal transduction acts in an analogous manner to therecruitment of SH2-containing tyrosine phosphatases to activated receptor tyrosinekinases, and that phosphorylation-dependent attenuation of receptor-mediatedsignaling is a common feature of both plant and animal cells [10].
The cis–trans prolyl isomerase PinA in the slime mould Dictyostelium discoideummay represent a particularly interesting example of FHA-dependent recruitmentof substrates. PinA is the slime-mould homolog of Pin1, a cis–trans prolyl isomerasewith specificity for proline residues preceded by phosphoserine or phospho-threonine. In most metazoans, the phospho dependence of Pin1 substrate recogni-tion requires a specialized WW domain [55]. However, in PinA, a WW domain isnot apparent, but the presence of an FHA domain in its place indicates that it mayserve an equivalent function. The FHA domain of PinA could therefore help torecruit phosphorylated proteins for proline isomerization. If this is true, PinA mayrepresent an instance of convergent evolution or domain shuffling.
7.3.3FHA Domain Binding to Regulators
The function of FHA domain-containing proteins can also be modulated uponFHA-dependent phosphoprotein binding. In fact, it is likely that a large proportionof FHA-dependent interactions will be found to regulate the function of the FHA-containing protein. For example, the FHA-dependent Rad53–Rad9 interaction,which led to the discovery of the FHA domain as a phosphothreonine bindingdomain, is essential for the activation of Rad53 in response to DNA damage [56].However, the exact mechanism by which Rad9 regulates Rad53 activity remainspoorly understood and is still under investigation.
7.3.4Reversible Protein–Protein Interactions
Intuitively, the presence of a phosphopeptide binding domain is suggestive of anability to form protein–protein interactions in response to signals. However, insome instances, the interaction between the FHA domain and the phosphoproteinappears constitutive under normal conditions but is rapidly inhibited underperturbing conditions. In these examples, the phospho-dependent interaction mayfunction as a switch to enforce rapid reversibility. This variation on the theme ofdynamic interactions appears to underlie the interaction between Rad53 and Asf1
7.3 Molecular and Signaling Function
1246vch07.pmd 10.09.2004, 14:41153

154 7 The FHA Domain
[27, 57]. In normally cycling budding yeast cells, Rad53 binds constitutively to Asf1in an FHA-dependent manner [27]. However, after DNA damage, Rad53 is releasedfrom Asf1 by an as-yet-uncharacterized mechanism. Thus, Rad53 appears to be‘released’ from Asf1 upon detection of DNA damage, allowing it to accomplish itsfunctions at the DNA replication fork or to signal DNA damage to the transcriptionand cell cycle machineries. Hence, in this particular instance, the main advantageof an FHA-dependent interaction may reside in the ability to break the interactionin response to a stimulus.
7.3.5FHA Domain as a Transcriptional Activator Domain
The forkhead-type transcription factor Fkh2 of budding yeast is a key regulator ofthe transcription program governing mitosis [58–60]. Fkh2 binds alongside theMADS-box Mcm1 protein to the promoter region of mitosis-specific genes. Fkh2binds to regulatory sequences known as SFF elements, which act as positiveelements during mitosis and repressor elements during interphase [58–60]. Fkh2binds the SFF element throughout the cell cycle, indicating that an additional levelof regulation must exist to allow SFF-dependent transcriptional activation in Mphase and SFF-dependent repression of M-phase genes in interphase. Thisregulation is imparted by the binding of the coactivator Ndd1 during mitosis(Figure 7.2) [58, 61, 62]. The Ndd1–Fkh2 interaction is controlled by Cdk phosphory-lation and was recently demonstrated to be mediated by a phospho-dependentinteraction between the FHA domain of Fkh2 and phThr239 of Ndd1 [61, 62].These results demonstrate that the FHA domain of Fkh2 can act as a transcriptionalactivator. Indeed, mutation of the critical Arg residue in the Fkh2 FHA domainmimics an NDD1 deletion [61, 62]. In the absence of Ndd1 binding, the FHA domainof Fkh2 appears to act as repressor. In support of this idea, NDD1 deletion is lethal,whereas the double mutation ndd1∆ fkh2∆ is viable [58]. This genetic interaction isconsistent with the model whereby Thr239-phosphorylated Ndd1 acts to convertFkh2 from a repressor to an activator of the G2/M transcription program [61, 62].
Fkh2 is part of a eukaryotic subfamily of forkhead transcription factors. In buddingyeast, the Fkh2 paralogs Fkh1 and Fhl1 have a similar FHA domain–forkhead DNA-binding domain structure. Fkh1 plays a role in the regulation of G2/M transcriptionbut is also critical for regulation of the recombination enhancer of the mating-typelocus, whereas Fhl1 is a transcriptional regulator of ribosome biogenesis [63].Interestingly, Fhl1 binds to a small regulator called Ifh1 and the IFH1 gene isessential only when FHL1 is present, suggesting a mode of transcriptional regulationessentially identical to the Fkh2–Ndd1 regulation of G2/M transcription (Figure 7.2)[64]. It will be interesting to determine if the interaction of Fhl1 to Ifh1 is phospho-and FHA-dependent and if it behaves similarly in other respects to the Fkh2–Ndd1interaction.
The parallel between Fkh2–Ndd1 and Fhl1–IFh1 is striking (Figure 7.2). Giventhat at least one Fkh2/Fhl1 homologue is present in most eukaryotes sequenced todate, it will be interesting to determine whether this mode of phospho-dependent
1246vch07.pmd 10.09.2004, 14:41154

155
transcriptional activation is also conserved. In mammals, MNF/ILF/Foxk1 is thesole Fkh2-like protein present and was identified as a regulator of muscle geneexpression and of the IL-2 promoter [65–67]. Interestingly, the N terminus of MNF/ILF/Foxk1 binds to the transcriptional corepressor mSIN3A [68]. The mSin3A-binding domain encompasses the FHA domain, perhaps indicating that phospho-dependent engagement of this forkhead transcription factor may convert it from arepressor to an activator. Further studies will be required to test this possibility.Nevertheless, these results indicate that Foxk1 may act in a manner similar to Fkh2/Fhl1-dependent regulation of transcription (Figure 7.2).
7.4Emerging Research Direction
7.4.1Bacterial FHA Domains
Prokaryotic FHA domains contain all the amino acid residues required forphosphothreonine binding and, indeed, two mycobacterial FHA domains (Rv1827and Rv0020c) have been shown to bind to phosphothreonine libraries in vitro.
Figure 7.2 The FHA–forkhead paradigm.In budding yeast, FHA-containing forkhead-type transcription factors Fkh2 and Fhl1(upper and middle panels) appear to act ina similar way. In conditions under which theFHA domain is not engaged in a phospho-
dependent interaction, they act as trans-criptional repressors. Upon FHA-dependentbinding to their coactivator proteins, they areconverted into transcriptional activators.See text for further details.
7.4 Emerging Research Direction
1246vch07.pmd 10.09.2004, 14:41155

156 7 The FHA Domain
This result raised the exciting possibility that modular phosphopeptide recognitionis widespread in bacteria [69]. Recent analysis of bacterial genomes has lent furthersupport to this possibility. Pallen et al. [70] demonstrated a near-perfect correlationbetween the presence of genes encoding for FHA domain-containing proteins andthe presence of genes coding for eukaryotic-type kinases and phosphatases [70],extending the earlier observations of Ponting et al [71]. This striking associationsuggests that bacterial FHA-containing proteins are engaging in processes in whichphosphorylation-dependent protein–protein interactions play an important role.Such a possibility has very recently gained further support after the demonstrationthat EmbR, a putative transcriptional regulator, contacts the eukaryote-type proteinkinase pknH in a phospho-dependent manner via its FHA domain [72]. Whetherthis interaction occurs in vivo remains to be demonstrated, but this recentobservation supports the notion that ‘eukaryotic-type’ signaling systems operate inbacteria.
7.4.2A Potential Role for FHA Domains During Innate Immunity?
The presence of bacterial FHA-containing proteins in type III secretion systemsindicates that FHA-containing proteins may play a role during pathogenic bacterialinfections [70]. The recent identifycation of T2BP/TIFA as a TRAF2/6-interactingFHA-containing protein suggests that at least one FHA-containing protein mayplay a role in innate immunity by influencing IL-1 and/or TNF signaling [73, 74].T2BP/TIFA appears to play a positive role in NFκB and MAP kinase signaling,because its overexpression potentiates both signaling processes [73, 74]. Thepresence of FHA-containing proteins in these important signaling processes, aswell as their presence in pathogenic bacteria, may suggest a role for bacterial FHAdomains in pathogen–host interactions. Although this possibility remains specula-tive, some pathogenic bacteria may use their FHA domains to remodel TRAF2/6complexes during bacterial infection. This hypothesis should be relatively straight-forward to assess.
7.4.3FHA Domain and Phosphothreonine-Proline Motifs
The characterization of the Rad53 and Chk2 FHA domains has led to a modelwhereby the main determinants for the FHA domain–phosphopeptide interactionare the phosphorylated residue along with the +3 residue [28]. Recent work on anumber of FHA domains is changing this view of FHA-dependent phosphopeptiderecognition. Indeed, the FHA domains of NIPP1, Ki-67, and possibly Fkh2/Fkh1appear to specifically require a proline residue immediately C-terminal to thephosphothreonine residue [32, 48, 62]. As mentioned above, recent work from theTsai laboratory indicates that, in the FHA domain of Ki67, the +1 proline residuemay play an essential and direct role in phosphopeptide recognition. This strikingcharacteristic places the FHA domain in a growing class of domains that recognize
1246vch07.pmd 10.09.2004, 14:41156
157
pSer/pThr-Pro motifs and which include the WD40 domain, the WW domain, andthe polo box domain [8, 55, 75, 76]. It will be interesting to determine whether allthe above FHA domains employ a common mechanism of proline recognition atthe +1 position. If so, such determinants could prove valuable for predicting whethera given FHA-containing protein participates in processes regulated by proline-directed kinases.
7.4.4FHA Domain Chimeras as Phosphorylation Biosensors
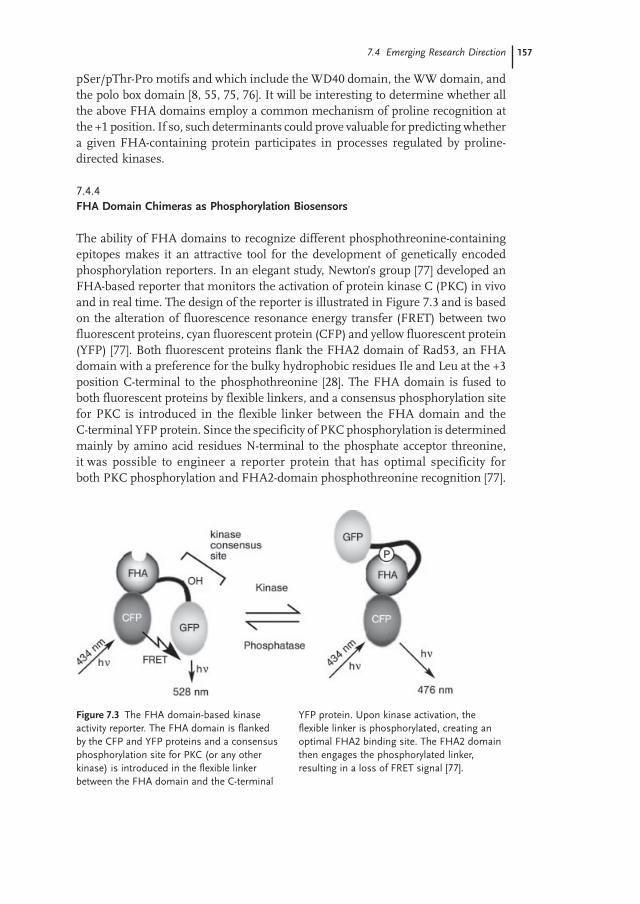
The ability of FHA domains to recognize different phosphothreonine-containingepitopes makes it an attractive tool for the development of genetically encodedphosphorylation reporters. In an elegant study, Newton’s group [77] developed anFHA-based reporter that monitors the activation of protein kinase C (PKC) in vivoand in real time. The design of the reporter is illustrated in Figure 7.3 and is basedon the alteration of fluorescence resonance energy transfer (FRET) between twofluorescent proteins, cyan fluorescent protein (CFP) and yellow fluorescent protein(YFP) [77]. Both fluorescent proteins flank the FHA2 domain of Rad53, an FHAdomain with a preference for the bulky hydrophobic residues Ile and Leu at the +3position C-terminal to the phosphothreonine [28]. The FHA domain is fused toboth fluorescent proteins by flexible linkers, and a consensus phosphorylation sitefor PKC is introduced in the flexible linker between the FHA domain and theC-terminal YFP protein. Since the specificity of PKC phosphorylation is determinedmainly by amino acid residues N-terminal to the phosphate acceptor threonine,it was possible to engineer a reporter protein that has optimal specificity forboth PKC phosphorylation and FHA2-domain phosphothreonine recognition [77].
Figure 7.3 The FHA domain-based kinaseactivity reporter. The FHA domain is flankedby the CFP and YFP proteins and a consensusphosphorylation site for PKC (or any otherkinase) is introduced in the flexible linkerbetween the FHA domain and the C-terminal
YFP protein. Upon kinase activation, theflexible linker is phosphorylated, creating anoptimal FHA2 binding site. The FHA2 domainthen engages the phosphorylated linker,resulting in a loss of FRET signal [77].
7.4 Emerging Research Direction
1246vch07.pmd 10.09.2004, 14:41157

158 7 The FHA Domain
Upon PKC activation, the flexible linker is phosphorylated by PKC, creating anoptimal FHA2 binding site [77]. The FHA2 domain then engages the phosphorylatedlinker in an intramolecular interaction, thereby moving one fluorescent proteinaway from the other, leading to a loss of FRET signal [77]. Interestingly, the relativelylow affinity of the FHA2 domain for its optimal binding site appears to be a majorfactor in enabling the monitoring of dynamic phosphorylation events in real time.Indeed, the lower binding affinity enables the access of phosphatase to thephosphothreonine residue, with concomitant restoration of the FRET signal [77].
In addition to the PKC system, it is easy to imagine how this design could beapplicable to the study of a wide variety of phosphorylation events. For example,FHA domains with proline-directed phospho recognition could be used to monitorCdk and MAP kinase activation. Furthermore, since FHA2 domain phosphopeptiderecognition is mainly dependent on the presence of an Ile residue three residuesC-terminal to the phosphorylated residue, the reporter described by Newton et al.can be easily adapted to monitor the regulation of a number of kinases, includingATM-like kinases and basic-directed kinases.
7.5Concluding Remarks
The discovery of the FHA domain as a phosphothreonine binding module was asignificant step toward gaining a better understanding of Ser/Thr-based signalingsystems in prokaryotes and eukaryotes. The understanding of FHA domain functionis also helping us to decipher the molecular basis of some diseases, such as thevariant Li–Fraumeni and Nijmegen-breakage syndromes. In the years to come,I expect that the study of FHA-containing proteins will continue to further of ourunderstanding of various cellular processes and that the discovery of novel phospho-dependent protein interaction modules will have a similar impact as that of theFHA domain.
Acknowledgements
I thank the members of my laboratory for their help and support. I would also liketo thank Frank Sicheri for his critical reading of this manuscript. I am a recipientof the Hitchings–Elion fellowship of the Burroughs–Wellcome Fund and I hold aCanada Research Chair (tier II) in Bioinformatics, Proteomics and FunctionalGenomics. Work in my laboratory is supported by grants from the CanadianInstitutes for Health Research and the National Cancer Institute of Canada.
1246vch07.pmd 10.09.2004, 14:41158

159
1 Pawson, T., Nash, P., Protein–proteininteractions define specificity in signaltransduction. Genes. Dev. 2000, 14,1027–1047.
2 Pawson, T., Nash, P., Assembly of cellregulatory systems through proteininteraction domains. Science 2003, 300,445–452.
3 Yaffe, M. B., Elia, A. E., Phosphoserine/threonine-binding domains. Curr. Opin.Cell Biol. 2001, 13, 131–138.
4 Ponting, C. P., Russell, R. R., The naturalhistory of protein domains. Annu. Rev.Biophys. Biomol. Struct. 2002, 31, 45–71.
5 Manke, I. A., Lowery, D. M., Nguyen, A.,Yaffe, M. B., BRCT repeats as phospho-peptide-binding modules involved inprotein targeting. Science 2003, 302,636–639.
6 Rodriguez, M., Yu, X., Chen, J.,Songyang, Z., Phosphopeptide bindingspecificities of BRCA1 COOH-terminal(BRCT) domains. J. Biol. Chem. 2003,278, 52914–52918.
7 Yu, X., Chini, C. C., He, M., Mer, G.,Chen, J., The BRCT domain is aphospho-protein binding domain.Science 2003, 302, 639–642.
8 Elia, A. E., Cantley, L. C., Yaffe, M. B.,Proteomic screen finds pSer/pThr-binding domain localizing Plk1 to mitoticsubstrates. Science 2003, 299, 1228–1231.
9 Durocher, D., Jackson, S. P., The FHAdomain. FEBS Lett. 2002, 513, 58–66.
10 Li, J., Lee, G. I., Van Doren, S. R.,Walker, J. C., The FHA domainmediates phosphoprotein interactions.J. Cell Sci. 2000, 113 Pt 23, 4143–4149.
11 Tsai, M. D., FHA: a signal transductiondomain with diverse specificity andfunction. Structure (Camb) 2002, 10,887–888.
12 Hammet, A., Pike, B. L., McNees, C. J.,Conlan, L. A., Tenis, N., Heierhorst, J.,FHA domains as phospho-threoninebinding modules in cell signaling.IUBMB Life 2003, 55, 23–27.
13 Hofmann, K., Bucher, P., The FHAdomain: a putative nuclear signallingdomain found in protein kinases andtranscription factors. Trends Biochem. Sci.1995, 20, 347–349.
14 Caldecott, K. W., DNA single-strandbreak repair and spinocerebellar ataxia.Cell 2003, 112, 7–10.
15 Date, H., et al., Early-onset ataxia withocular motor apraxia and hypoalbumi-nemia is caused by mutations in a newHIT superfamily gene. Nat. Genet. 2001,29, 184–188.
16 Moreira, M. C., et al., The gene mutatedin ataxia-ocular apraxia 1 encodes thenew HIT/Zn-finger protein aprataxin.Nat. Genet. 2001, 29, 189–193.
17 Stone, J. M., Collinge, M. A., Smith,R. D., Horn, M. A., Walker, J. C.,Interaction of a protein phosphatase withan Arabidopsis serine-threonine receptorkinase. Science 1994, 266, 793–795.
18 Li, J., Smith, G. P., Walker, J. C., Kinaseinteraction domain of kinase-associatedprotein phosphatase, a phosphoprotein-binding domain. Proc. Natl. Acad. Sci.USA 1999, 96, 7821–7826.
19 Rouse, J., Jackson, S. P., Interfacesbetween the detection, signaling, andrepair of DNA damage. Science 2002, 297,547–551.
20 Bartek, J., Lukas, J., Chk1 and Chk2kinases in checkpoint control and cancer.Cancer Cell 2003, 3, 421–429.
21 Bashkirov, V. I., Bashkirova, E. V.,Haghnazari, E., Heyer, W. D., Directkinase-to-kinase signaling mediated bythe FHA phosphoprotein recognitiondomain of the Dun1 DNA damagecheckpoint kinase. Mol. Cell Biol. 2003,23, 1441–1452.
22 Wan, L., De Los Santos, T., Zhang, C.,Shokat, K., Hollingsworth, N. M.,Mek1 kinase activity functions down-stream of RED1 in the regulation ofmeiotic double strand break repair inbudding yeast. Mol. Biol. Cell 2004, 15,11–23.
23 Sun, Z., Hsiao, J., Fay, D. S., Stern,D. F., Rad53 FHA domain associatedwith phosphorylated Rad9 in the DNAdamage checkpoint. Science 1998, 281,272–274.
24 Vialard, J. E., Gilbert, C. S., Green,C. M., Lowndes, N. F., The budding yeastRad9 checkpoint protein is subjected toMec1/Tel1-dependent hyperphosphory-
References
References
1246vch07.pmd 10.09.2004, 14:41159

160 7 The FHA Domain
lation and interacts with Rad53 afterDNA damage. EMBO J. 1998, 17,5679–5688.
25 Durocher, D., Henckel, J., Fersht,A. R., Jackson, S. P., The FHA domainis a modular phosphopeptide recognitionmotif. Mol. Cell 1999, 4, 387–394.
26 Emili, A., MEC1-dependent phosphory-lation of Rad9p in response to DNAdamage. Mol. Cell 1998, 2, 183–189.
27 Schwartz, M. F., Lee, S. J., Duong, J. K.,Eminaga, S., Stern, D. F., FHA domain-mediated DNA checkpoint regulation ofRad53. Cell Cycle 2003, 2, 384–396.
28 Durocher, D., Taylor, I. A., Sarbassova,D., Haire, L. F., Westcott, S. L.,Jackson, S. P., Smerdon, S. J., Yaffe,M. B., The molecular basis of FHAdomain: phosphopeptide bindingspecificity and implications for phospho-dependent signaling mechanisms.Mol. Cell 2000, 6, 1169–1182.
29 Liao, H., Byeon, I. J., Tsai, M. D.,Structure and function of a new phospho-peptide-binding domain containing theFHA2 of Rad53. J. Mol. Biol. 1999, 294,1041–1049.
30 Liao, H., et al., Structure of the FHA1domain of yeast Rad53 and identificationof binding sites for both FHA1 and itstarget protein Rad9. J. Mol. Biol. 2000,304, 941–951.
31 Li, J., et al., Structural and functionalversatility of the FHA domain inDNA-damage signaling by the tumorsuppressor kinase Chk2. Mol. Cell 2002,9, 1045–1054.
32 Li, H., Byeon, I. J., Ju, Y., Tsai, M. D.,Structure of human Ki67 FHA domainand its binding to a phosphoproteinfragment from hNIFK reveal uniquerecognition sites and new views to thestructural basis of FHA domain func-tions. J. Mol. Biol. 2004, 335, 371–381.
33 Stavridi, E. S., Huyen, Y., Loreto, I. R.,Scolnick, D. M., Halazonetis, T. D.,Pavletich, N. P., Jeffrey, P. D., Crystalstructure of the FHA domain of theChfr mitotic checkpoint protein and itscomplex with tungstate. Structure (Camb)2002, 10, 891–899.
34 Lee, G. I., Ding, Z., Walker, J. C.,Van Doren, S. R., NMR structure of theforkhead-associated domain from the
Arabidopsis receptor kinase-associatedprotein phosphatase. Proc. Natl. Acad. Sci.USA 2003, 100, 11261–11266.
35 Hammet, A., Pike, B. L., Mitchelhill,K. I., Teh, T., Kobe, B., House, C. M.,Kemp, B. E., Heierhorst, J., FHAdomain boundaries of the Dun1p andRad53p cell cycle checkpoint kinases.FEBS Lett. 2000, 471, 141–146.
36 Davies, D. R., Cohen, G. H., Inter-actions of protein antigens with anti-bodies. Proc. Natl. Acad. Sci. USA 1996,93, 7–12.
37 Qin, D., Lee, H., Yuan, C., Ju, Y., Tsai,M. D., Identification of potential bindingsites for the FHA domain of humanChk2 by in vitro binding studies.Biochem. Biophys. Res. Commun. 2003,311, 803–808.
38 Bell, D. W., et al., Heterozygous germline hCHK2 mutations in Li–Fraumenisyndrome. Science 1999, 286, 2528–2531.
39 Falck, J., Mailand, N., Syljuasen, R. G.,Bartek, J., Lukas, J., The ATM–Chk2–Cdc25A checkpoint pathway guardsagainst radioresistant DNA synthesis.Nature 2001, 410, 842–847.
40 Takagi, M., Sueishi, M., Saiwaki, T.,Kametaka, A., Yoneda, Y., A novelnucleolar protein, NIFK, interacts withthe forkhead associated domain of Ki-67antigen in mitosis. J. Biol. Chem. 2001,276, 25386–25391.
41 Moustakas, A., Heldin, C. H., The nutsand bolts of IRF structure. Nat. Struct.Biol. 2003, 10, 874–876.
42 Qin, B. Y., Liu, C., Lam, S. S., Srinath,H., Delston, R., Correia, J. J., Derynck,R., Lin, K., Crystal structure of IRF-3reveals mechanism of autoinhibition andvirus-induced phosphoactivation. Nat.Struct. Biol. 2003, 10, 913–921.
43 Takahasi, K., et al., X-ray crystalstructure of IRF-3 and its functionalimplications. Nat. Struct. Biol. 2003, 10,922–927.
44 Wu, J. W., et al., Crystal structure of aphosphorylated Smad2: recognition ofphosphoserine by the MH2 domain andinsights on Smad function in TGF-betasignaling. Mol. Cell 2001, 8, 1277–1289.
45 Jagiello, I., Van Eynde, A., Vulsteke, V.,Beullens, M., Boudrez, A., Keppens, S.,Stalmans, W., Bollen, M., Nuclear and
1246vch07.pmd 10.09.2004, 14:41160

161
subnuclear targeting sequences of theprotein phosphatase-1 regulator NIPP1.J. Cell Sci. 2000, 113 Pt 21, 3761–3768.
46 Trinkle-Mulcahy, L., Ajuh, P.,Prescott, A., Claverie-Martin, F.,Cohen, S., Lamond, A. I., Cohen, P.,Nuclear organisation of NIPP1, a regula-tory subunit of protein phosphatase 1that associates with pre-mRNA splicingfactors. J. Cell Sci. 1999, 112, 157–168.
47 Boudrez, A., et al., NIPP1-mediatedinteraction of protein phosphatase-1with CDC5L, a regulator of pre-mRNAsplicing and mitotic entry. J. Biol. Chem.2000, 275, 25411–25417.
48 Boudrez, A., Beullens, M., Waelkens,E., Stalmans, W., Bollen, M.,Phosphorylation-dependent interactionbetween the splicing factors SAP155and NIPP1. J. Biol. Chem. 2002, 277,31834–31841.
49 Murone, M., Simanis, V., The fissionyeast dma1 gene is a component of thespindle assembly checkpoint, requiredto prevent septum formation andpremature exit from mitosis if spindlefunction is compromised. EMBO J. 1996,15, 6605–6616.
50 Scolnick, D. M., Halazonetis, T. D.,Chfr defines a mitotic stress checkpointthat delays entry into metaphase. Nature2000, 406, 430–435.
51 Bardin, A. J., Amon, A., Men and sin:what’s the difference? Nat. Rev. Mol. CellBiol. 2001, 2, 815–826.
52 Guertin, D. A., Venkatram, S., Gould,K. L., McCollum, D., Dma1 preventsmitotic exit and cytokinesis by inhibitingthe septation initiation network (SIN).Dev Cell 2002, 3, 779–790.
53 Kang, D., Chen, J., Wong, J., Fang, G.,The checkpoint protein Chfr is a ligasethat ubiquitinates Plk1 and inhibits Cdc2at the G2 to M transition. J. Cell Biol.2002, 156, 249–259.
54 Stone, J. M., Trotochaud, A. E.,Walker, J. C., Clark, S. E., Control ofmeristem development by CLAVATA1receptor kinase and kinase-associatedprotein phosphatase interactions. PlantPhysiol 1998, 117, 1217–1225.
55 Lu, P. J., Zhou, X. Z., Shen, M.,Lu, K. P., Function of WW domains asphosphoserine- or phosphothreonine-
binding modules. Science 1999, 283,1325–1328.
56 Durocher, D., Smerdon, S. J., Yaffe,M. B., Jackson, S. P., The FHA domainin DNA repair and checkpoint signaling.Cold Spring Harbor Symposia on Quantita-tive Biology, Vol. LXV. 2000, 423–431.
57 Emili, A., Schieltz, D. M., Yates, J. R.3rd, Hartwell, L. H., Dynamic inter-action of DNA damage checkpointprotein Rad53 with chromatin assemblyfactor Asf1. Mol. Cell 2001, 7, 13–20.
58 Koranda, M., Schleiffer, A., Endler,L., Ammerer, G., Forkhead-like trans-cription factors recruit Ndd1 to thechromatin of G2/M-specific promoters.Nature 2000, 406, 94–98.
59 Kumar, R., Reynolds, D. M.,Shevchenko, A., Goldstone, S. D.,Dalton, S., Forkhead transcriptionfactors, Fkh1p and Fkh2p, collaboratewith Mcm1p to control transcriptionrequired for M-phase. Curr. Biol. 2000,10, 896–906.
60 Pic, A., et al., The forkhead protein Fkh2is a component of the yeast cell cycletranscription factor SFF. EMBO J. 2000,19, 3750–3761.
61 Darieva, Z., et al., Cell cycle-regulatedtranscription through the FHA domainof Fkh2p and the coactivator Ndd1p.Curr. Biol. 2003, 13, 1740–1745.
62 Reynolds, D., Shi, B. J., McLean, C.,Katsis, F., Kemp, B., Dalton, S.,Recruitment of Thr 319-phosphorylatedNdd1p to the FHA domain of Fkh2prequires Clb kinase activity: a mechanismfor CLB cluster gene activation. Genes.Dev. 2003, 17, 1789–1802.
63 Hermann-Le Denmat, S., Werner, M.,Sentenac, A., Thuriaux, P., Suppressionof yeast RNA polymerase III mutations byFHL1, a gene coding for a fork head proteininvolved in rRNA processing. Mol. Cell Biol.1994, 14, 2905–2913.
64 Cherel, I., Thuriaux, P., The IFH1gene product interacts with a fork headprotein in Saccharomyces cerevisiae. Yeast1995, 11, 261–270-
65 Nirula, A., Moore, D. J., Gaynor, R. B.,Constitutive binding of the transcriptionfactor interleukin-2 (IL-2) enhancerbinding factor to the IL-2 promoter.J. Biol. Chem. 1997, 272, 7736–7745.
References
1246vch07.pmd 10.09.2004, 14:41161

162 7 The FHA Domain
66 Li, C., Lai, C. F., Sigman, D. S., Gaynor,R. B., Cloning of a cellular factor, inter-leukin binding factor, that binds to NFAT-like motifs in the human immunodefi-ciency virus long terminal repeat. Proc.Natl. Acad. Sci. USA 1991, 88, 7739–7743.
67 Bassel-Duby, R., Hernandez, M. D.,Yang, Q., Rochelle, J. M., Seldin,M. F., Williams, R. S., Myocyte nuclearfactor, a novel winged-helix transcriptionfactor under both developmental andneural regulation in striated myocytes.Mol. Cell Biol. 1994, 14, 4596–4605.
68 Yang, Q., Kong, Y., Rothermel, B.,Garry, D. J., Bassel-Duby, R., Williams,R. S., The winged-helix/forkhead proteinmyocyte nuclear factor beta (MNF-beta)forms a co-repressor complex withmammalian sin3B. Biochem. J. 2000, 345Pt 2, 335–343.
69 Durocher, D., Bacterial signal trans-duction: a FHAscinating glimpse at theorigins of phospho-dependent signaltransduction. Trends Microbiol 2003, 11,67–68.
70 Pallen, M., Chaudhuri, R., Khan, A.,Bacterial FHA domains: neglected playersin the phospho-threonine signaling game?Trends Microbiol 2002, 10, 556–563.
71 Ponting, C. P., Aravind, L., Schultz,J., Bork, P., Koonin, E. V., Eukaryoticsignalling domain homologues inarchaea and bacteria: ancient ancestryand horizontal gene transfer. J. Mol. Biol.1999, 289, 729–745.
72 Molle, V., Kremer, L., Girard-Blanc,C., Besra, G. S., Cozzone, A. J., Prost,J. F., An FHA phosphoprotein recogni-tion domain mediates protein EmbRphosphorylation by PknH, a Ser/Thrprotein kinase from Mycobacteriumtuberculosis. Biochemistry 2003, 42,15300–15309.
73 Takatsuna, H., et al., Identification ofTIFA as an adapter protein that linkstumor necrosis factor receptor-associatedfactor 6 (TRAF6) to interleukin-1 (IL-1)receptor-associated kinase-1 (IRAK-1) inIL-1 receptor signaling. J. Biol. Chem.2003, 278, 12144–12150.
74 Kanamori, M., Suzuki, H., Saito, R.,Muramatsu, M., Hayashizaki, Y.,T2BP, a novel TRAF2 binding protein,can activate NF-kappaB and AP-1without TNF stimulation. Biochem.Biophys. Res. Commun. 2002, 290,1108–1113.
75 Nash, P., et al., Multisite phosphorylationof a CDK inhibitor sets a threshold forthe onset of DNA replication. Nature2001, 414, 514–521.
76 Elia, A. E., et al., The molecular basis forphosphodependent substrate targetingand regulation of Plks by the Polo-boxdomain. Cell 2003, 115, 83–95.
77 Violin, J. D., Zhang, J., Tsien, R. Y.,Newton, A. C., A genetically encodedfluorescent reporter reveals oscillatoryphosphorylation by protein kinase C.J. Cell Biol. 2003, 161, 899–909.
Websites related to FHA domains
SMART database: http://smart.embl-heidelberg.de/
Pfam database: http://www.sanger.ac.uk/Software/Pfam/
Ensembl genome browser: http://www.ensembl.org/
FHA domain entry, Pawson laboratory: http://www.mshri.on.ca/pawson/fha.html
1246vch07.pmd 10.09.2004, 14:41162





























