Embed Size (px)
Citation preview

CENTELLA ASIATICA ON HEPATOCYTES 605
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
Copyright © 2005 John Wiley & Sons, Ltd.
PHYTOTHERAPY RESEARCHPhytother. Res. 19, 605–611 (2005)Published online in Wiley InterScience (www.interscience.wiley.com). DOI: 10.1002/ptr.1684
Modification of Gamma Ray induced Changesin the Mouse Hepatocytes by Centella asiaticaextract: In Vivo Studies
Radha Sharma and Jaimala SharmaDepartment of Zoology, University of Rajasthan, Jaipur 302 004, India
Ionizing radiation, besides causing direct damage also generates reactive oxygen species that are capable ofinducing damage to various organs. Pretreatment with Centella asiatica 1 h prior to irradiation at a dose rateof 100 mg/kg body weight was found to be effective against radiation induced damage in the liver. The numberof normal hepatocytes was higher in the Centella asiatica pretreated group in comparison with the irradiatedonly group. The number of binucleated cells and abnormal hepatocytes was less in comparison with theanimals irradiated without Centella asiatica pretreatment. Copyright © 2005 John Wiley & Sons, Ltd.
Keywords: gamma radiation; liver; hepatocytes; Centella asiatica.
Received 8 April 2003Accepted 27 January 2005
* Correspondence to: Dr R. Sharma, Department of Zoology, Universityof Rajasthan, Jaipur 302 004, India.E-mail: [email protected]
The purpose of the present study was to evaluate theprotective activity of Centella asiatica plant extract forradiation induced hepatic injury, the sensitivity ofnormal liver to radiation induced interphase death andthe rate at which injured or dead cells are removedfrom this organ following irradiation were quantitativelyevaluated.
Centella asiatica (Linn.), Syn: Hydrocotyle asiatica(L), Apiaceae, commonly known as Brahmi or IndianPennywort , is used in a number of herbal preparationsand is prescribed for the treatment of various diseasesin the Indian system of medicine, Ayurved.
MATERIALS AND METHODS
Source of animals. The experiments were conductedon Swiss albino mice obtained from an inbred colonymaintained in the laboratory on standard mouse food.Six to eight week old healthy adult male mice, weigh-ing 25 g (±2 g) were selected. All animals were giventetracycline containing water whenever required, toprotect them from infection. The animals were groupedrandomly.
Source of irradiation. Irradiation of animals wascarried out at S.M.S. Hospital, Jaipur with the help ofthe Cobalt-60 ATC-C9 teletherapy unit in the cancertreatment centre. The dose rate of the source at adistance of 80 cm was 1.08 and 1.09 Gy/min for 6 and8 Gy, respectively. A dose of 6 Gy is sublethal and 8 Gyis a lethal dose of radiation for Swiss albino mice, hencethese doses were selected for the experiment.
Plant extract. The dried powdered extract of Centellaasiatica, equivalent to 16.66% w/w of the dried powderof the plant, was supplied by Amsar Private Limited,Indore, as a gift. The extract was dissolved in distilledwater and the animals were fed by gastric intubation at
INTRODUCTION
Normal and cancerous tissues have similar responses toradiation exposure. Radiation induced injury to normaltissues may present either during or after the comple-tion of radiotherapy treatment for killing malignant cells.
Ionizing radiation generates reactive oxygen speciesthat are capable of inducing damage to various cellularmolecules, and cause peroxidation. The lipid peroxideinduced damage changes the membrane permeabilityand injures both the cell and the organ (Bandyopadhyayet al., 1999). An increase in lipid peroxidation in mouseliver is related to the dose of radiation dose and thetime elapsed.
Acute and chronic hepatic injury can be a dose limit-ing, life threatening complication in patients under-going radiation for metastatic and primary liver cancers,and other neoplasms in the upper abdomen or lowerthorax when the whole or most of the liver is in theradiation field (Lewin and Millis, 1973). The principalgross manifestations are hepatomegaly, hyperaemia,jaundice and ascites. Histology of the liver shows extra-vasation, parenchymal cell loss, formation of canaliculi,fibrosis and necrosis (Mehta et al., 1975).
Some authors have proposed that histological changesin the liver are an indirect result of irradiation (Scottet al., 1962). The reported vascular changes were mostprominent in the smaller hepatic vein branches, andwere described as a loss of endothelium, splitting andthickening of subintimal argyrophilic fibers to form acellular loose fibrillar tissue sometimes filling the veinlumen. These changes were regarded as the cause ofthe accompanying hepatic cell injury. It was suggestedthat the vascular lesions may have been a direct resultof irradiation.

606 R. SHARMA AND J. SHARMA
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
a dose rate of 100 mg/kg body weight. This dose wasselected on the basis of the survival test (Sharma andSharma, 2002).
Design of experiments. Healthy adult Swiss albinomice (all males) were selected from an inbred colonymaintained in the laboratory under ideal conditionswith standard mouse feed and water ad libitum. Theweight of the animals was 25 ± 2 g, and their agewas between 6 and 8 weeks hence there was a littlevariation in the various groups. They were randomlygrouped as follows.
Group I Animals of this group were sham-irradiated.This group served as normal group.
Group II Animals of this group received plant extract1 h before irradiation at a dose rate of100 mg/kg body weight orally. Animals wereexposed to 6 Gy 60Co gamma radiation.
Group III Animals of this group were irradiated with6 Gy of 60Co gamma rays and given an equalamount of double distilled water as givenwith the plant extract.
Group IV Animals of this group received plant extract1 h before irradiation at a dose rate of100 mg/kg body weight orally. Animals wereexposed to 8 Gy of 60Co gamma radiation.
Group V Animals of this group were irradiated with8 Gy of 60Co gamma rays and given an equalamount of double distilled water as givenwith the plant extract.Groups III and V served as control groupswhile groups II and IV served as experi-mental groups.
Group VI Animals of this group received extract ofCentella asiatica at a dose rate of 100 mg/kgbody weight orally.
Autopsy. The animals of all the groups were killed bycervical dislocation at 1/4, 1, 2, 4, 7, 10, 14 and 28 dayspost irradiation. At least six animals of each group werekilled at each interval. The liver was removed, weighedand fixed in Bouin’s fluid for 24 h for histopathologicalstudies. The tissue was properly dehydrated in alco-holic series and embedded in paraffin wax followingthe routine procedure. Sections were cut at 5 µm andstained with Harris hematoxylin and eosin.
Normal hepatocytes and abnormal hepatocytesbesides binucleate cells were counted with the help ofplanimeter.
Statistical analysis. All the values related to countingare expressed as standard error of mean (±SEM). Thedata were subjected to Student’s t-test for comparisonbetween the groups.
RESULTS
Normal hepatocytes
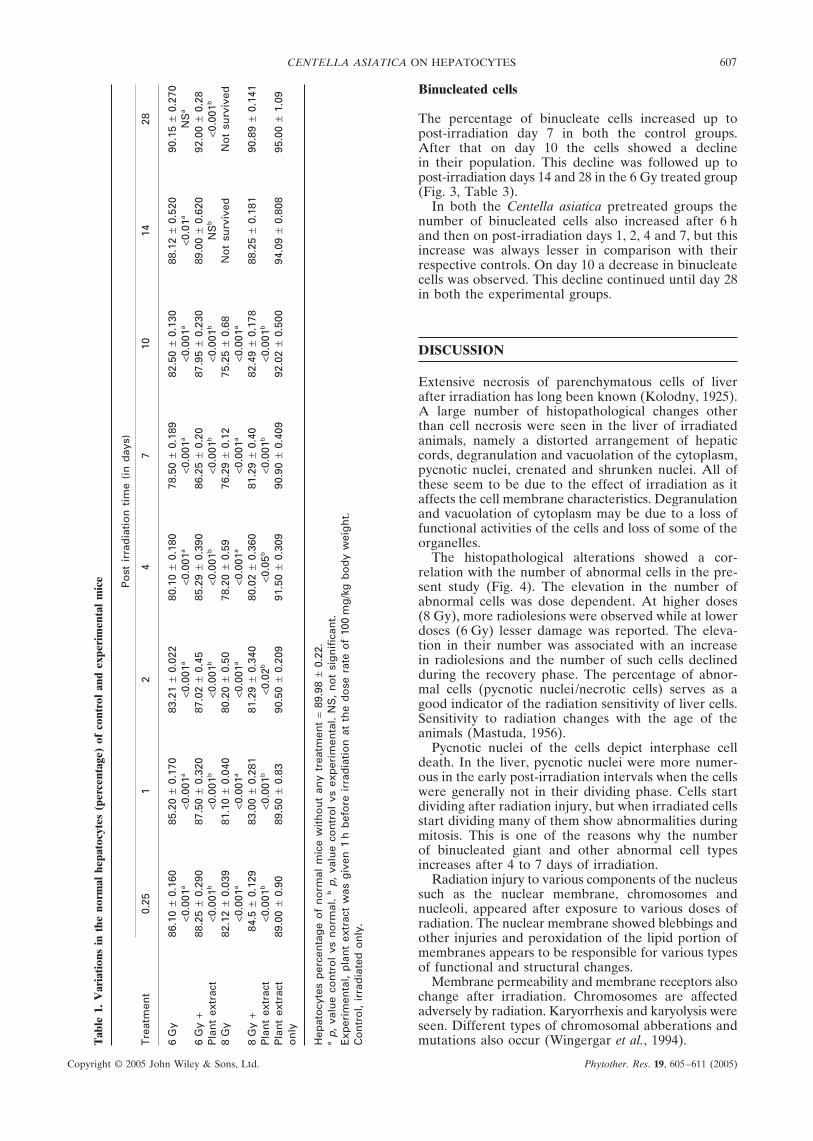
The population of normal hepatocytes decreasedafter irradiation in both the control groups. Thisdecline was higher in the 8 Gy treated group in com-parison with the 6 Gy control. The number ofcells decreased after 6 h, and continued up to post-irradiation days 7 and 10 in the 6 and 8 Gy irradiatedanimals, respectively. After that it started to increaseup to post-irradiation day 28 in the 6 Gy treated group(Fig. 1, Table 1).
The population of normal hepatocytes was alsodecreased after irradiation in the experimental groups.This decline was always lesser in comparison with theirrespective controls. Early recovery was observed in boththe experimental groups.
Abnormal hepatocytes
In the control groups the number of abnormal hepato-cytes increased up to post-irradiation day 7. The numberof abnormal cells was higher in the 8 Gy treated groupin comparison with 6 Gy, at each autopsy interval(Fig. 2, Table 2).
On day 10 the percentage of abnormal cells declined.This decline was continued up to post-irradiation day14 and 28 in the 6 Gy treated group.
In the experimental groups, the percentage of ab-normal cells was always less in comparison withtheir respective controls. The number of abnormalhepatocytes increased up to post-irradiation day 7.On days 10, 14 and 28 the number of abnormal cellsdeclined in both the experimental groups.
Figure 1. Variation in normal hepatocytes of irradiated mouse with and without Centella asiatica pretreatment.
CENTELLA ASIATICA ON HEPATOCYTES 607
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
Binucleated cells
The percentage of binucleate cells increased up topost-irradiation day 7 in both the control groups.After that on day 10 the cells showed a declinein their population. This decline was followed up topost-irradiation days 14 and 28 in the 6 Gy treated group(Fig. 3, Table 3).
In both the Centella asiatica pretreated groups thenumber of binucleated cells also increased after 6 hand then on post-irradiation days 1, 2, 4 and 7, but thisincrease was always lesser in comparison with theirrespective controls. On day 10 a decrease in binucleatecells was observed. This decline continued until day 28in both the experimental groups.
DISCUSSION
Extensive necrosis of parenchymatous cells of liverafter irradiation has long been known (Kolodny, 1925).A large number of histopathological changes otherthan cell necrosis were seen in the liver of irradiatedanimals, namely a distorted arrangement of hepaticcords, degranulation and vacuolation of the cytoplasm,pycnotic nuclei, crenated and shrunken nuclei. All ofthese seem to be due to the effect of irradiation as itaffects the cell membrane characteristics. Degranulationand vacuolation of cytoplasm may be due to a loss offunctional activities of the cells and loss of some of theorganelles.
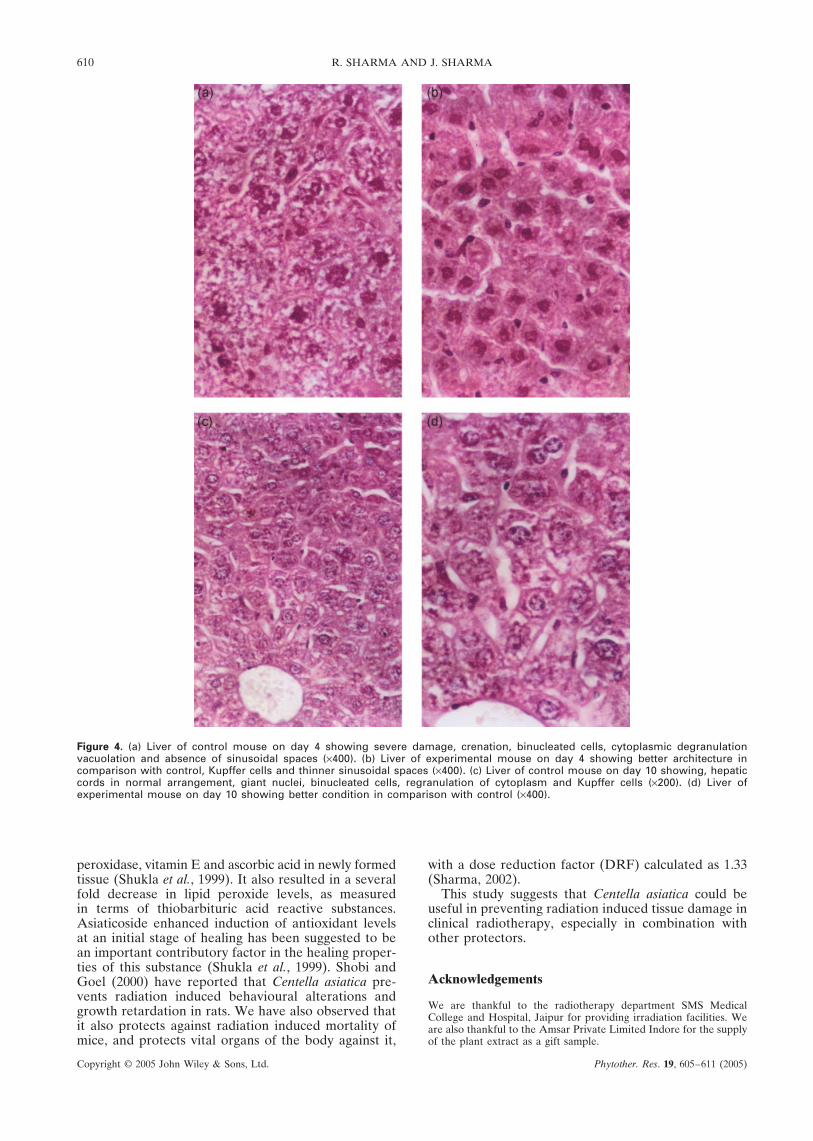
The histopathological alterations showed a cor-relation with the number of abnormal cells in the pre-sent study (Fig. 4). The elevation in the number ofabnormal cells was dose dependent. At higher doses(8 Gy), more radiolesions were observed while at lowerdoses (6 Gy) lesser damage was reported. The eleva-tion in their number was associated with an increasein radiolesions and the number of such cells declinedduring the recovery phase. The percentage of abnor-mal cells (pycnotic nuclei/necrotic cells) serves as agood indicator of the radiation sensitivity of liver cells.Sensitivity to radiation changes with the age of theanimals (Mastuda, 1956).
Pycnotic nuclei of the cells depict interphase celldeath. In the liver, pycnotic nuclei were more numer-ous in the early post-irradiation intervals when the cellswere generally not in their dividing phase. Cells startdividing after radiation injury, but when irradiated cellsstart dividing many of them show abnormalities duringmitosis. This is one of the reasons why the numberof binucleated giant and other abnormal cell typesincreases after 4 to 7 days of irradiation.
Radiation injury to various components of the nucleussuch as the nuclear membrane, chromosomes andnucleoli, appeared after exposure to various doses ofradiation. The nuclear membrane showed blebbings andother injuries and peroxidation of the lipid portion ofmembranes appears to be responsible for various typesof functional and structural changes.
Membrane permeability and membrane receptors alsochange after irradiation. Chromosomes are affectedadversely by radiation. Karyorrhexis and karyolysis wereseen. Different types of chromosomal abberations andmutations also occur (Wingergar et al., 1994).T
able
1.
Var
iati
ons
in t
he n
orm
al h
epat
ocyt
es (
perc
enta
ge)
of c
ontr
ol a
nd e
xper
imen
tal
mic
e Po
st i
rrad
iati
on
tim
e (i
n d
ays)
Tre
atm
ent
0.25
12
47
1014
28
6G
y86
.10
± 0.
160
85.2
0 ±
0.17
083
.21
± 0.
022
80.1
0 ±
0.18
078
.50
± 0.
189
82.5
0 ±
0.13
088
.12
± 0.
520
90.1
5 ±
0.27
0<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.01a
NS
a
6G
y +
88.2
5 ±
0.29
087
.50
± 0.
320
87.0
2 ±
0.45
85.2
9 ±
0.39
086
.25
± 0.
2087
.95
± 0.
230
89.0
0 ±
0.62
092
.00
± 0.
28P
lan
t ex
trac
t<0
.001
b<0
.001
b<0
.001
b<0
.001
b<0
.001
b<0
.001
bN
Sb
<0.0
01b
8G
y82
.12
± 0.
039
81.1
0 ±
0.04
080
.20
± 0.
5078
.20
± 0.
5976
.29
± 0.
1275
.25
± 0.
68N
ot
surv
ived
No
t su
rviv
ed<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a
8G
y +
84.5
± 0
.129
83.0
0 ±
0.28
181
.29
± 0.
340
80.0
2 ±
0.36
081
.29
± 0.
4082
.49
± 0.
178
88.2
5 ±
0.18
190
.89
± 0.
141
Pla
nt
extr
act
<0.0
01b
<0.0
01b
<0.0
2b<0
.05b
<0.0
01b
<0.0
01b
Pla
nt
extr
act
89.0
0 ±
0.90
89.5
0 ±
0.83
90.5
0 ±
0.20
991
.50
± 0.
309
90.9
0 ±
0.40
992
.02
± 0.
500
94.0
9 ±
0.80
895
.00
± 1.
09o
nly
Hep
ato
cyte
s p
erce
nta
ge
of
no
rmal
mic
e w
ith
ou
t an
y tr
eatm
ent
= 89
.98
± 0.
22.
a p,
valu
e co
ntr
ol
vs n
orm
al.
b p
, va
lue
con
tro
l vs
exp
erim
enta
l. N
S,
no
t si
gn
ifica
nt.
Exp
erim
enta
l, p
lan
t ex
trac
t w
as g
iven
1h
bef
ore
irr
adia
tio
n a
t th
e d
ose
rat
e o
f 10
0m
g/k
g b
od
y w
eig
ht.
Co
ntr
ol,
irra
dia
ted
on
ly.
608 R. SHARMA AND J. SHARMA
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
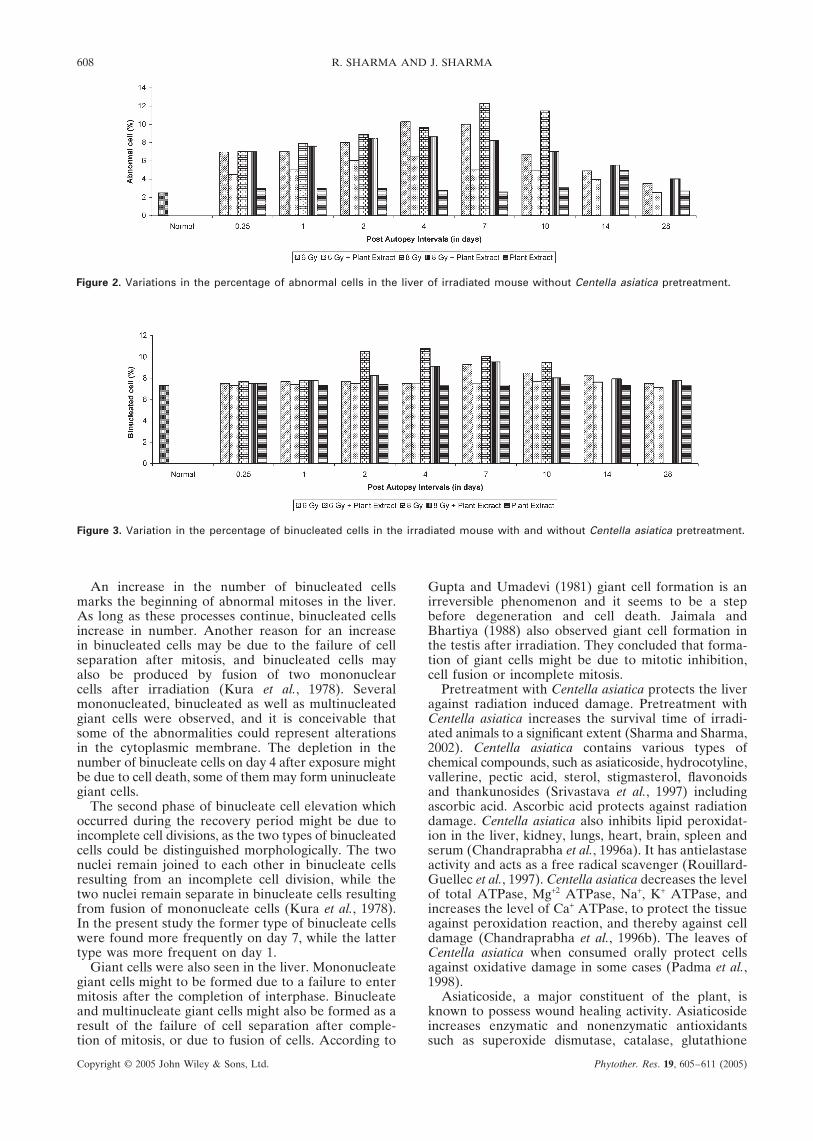
Figure 2. Variations in the percentage of abnormal cells in the liver of irradiated mouse without Centella asiatica pretreatment.
Figure 3. Variation in the percentage of binucleated cells in the irradiated mouse with and without Centella asiatica pretreatment.
Gupta and Umadevi (1981) giant cell formation is anirreversible phenomenon and it seems to be a stepbefore degeneration and cell death. Jaimala andBhartiya (1988) also observed giant cell formation inthe testis after irradiation. They concluded that forma-tion of giant cells might be due to mitotic inhibition,cell fusion or incomplete mitosis.
Pretreatment with Centella asiatica protects the liveragainst radiation induced damage. Pretreatment withCentella asiatica increases the survival time of irradi-ated animals to a significant extent (Sharma and Sharma,2002). Centella asiatica contains various types ofchemical compounds, such as asiaticoside, hydrocotyline,vallerine, pectic acid, sterol, stigmasterol, flavonoidsand thankunosides (Srivastava et al., 1997) includingascorbic acid. Ascorbic acid protects against radiationdamage. Centella asiatica also inhibits lipid peroxidat-ion in the liver, kidney, lungs, heart, brain, spleen andserum (Chandraprabha et al., 1996a). It has antielastaseactivity and acts as a free radical scavenger (Rouillard-Guellec et al., 1997). Centella asiatica decreases the levelof total ATPase, Mg+2 ATPase, Na+, K+ ATPase, andincreases the level of Ca+ ATPase, to protect the tissueagainst peroxidation reaction, and thereby against celldamage (Chandraprabha et al., 1996b). The leaves ofCentella asiatica when consumed orally protect cellsagainst oxidative damage in some cases (Padma et al.,1998).
Asiaticoside, a major constituent of the plant, isknown to possess wound healing activity. Asiaticosideincreases enzymatic and nonenzymatic antioxidantssuch as superoxide dismutase, catalase, glutathione
An increase in the number of binucleated cellsmarks the beginning of abnormal mitoses in the liver.As long as these processes continue, binucleated cellsincrease in number. Another reason for an increasein binucleated cells may be due to the failure of cellseparation after mitosis, and binucleated cells mayalso be produced by fusion of two mononuclearcells after irradiation (Kura et al., 1978). Severalmononucleated, binucleated as well as multinucleatedgiant cells were observed, and it is conceivable thatsome of the abnormalities could represent alterationsin the cytoplasmic membrane. The depletion in thenumber of binucleate cells on day 4 after exposure mightbe due to cell death, some of them may form uninucleategiant cells.
The second phase of binucleate cell elevation whichoccurred during the recovery period might be due toincomplete cell divisions, as the two types of binucleatedcells could be distinguished morphologically. The twonuclei remain joined to each other in binucleate cellsresulting from an incomplete cell division, while thetwo nuclei remain separate in binucleate cells resultingfrom fusion of mononucleate cells (Kura et al., 1978).In the present study the former type of binucleate cellswere found more frequently on day 7, while the lattertype was more frequent on day 1.
Giant cells were also seen in the liver. Mononucleategiant cells might to be formed due to a failure to entermitosis after the completion of interphase. Binucleateand multinucleate giant cells might also be formed as aresult of the failure of cell separation after comple-tion of mitosis, or due to fusion of cells. According to
CENTELLA ASIATICA ON HEPATOCYTES 609
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
Tab
le 2
. V
aria
tion
s in
the
abn
orm
al c
ell
(per
cent
age)
in
the
liver
of
cont
rol
and
expe
rim
enta
l m
ice
Po
st i
rrad
iati
on
tim
e (i
n d
ays)
Tre
atm
ent
0.25
12
47
1014
28
6G
y6.
98 ±
0.2
207.
00 ±
0.2
678.
02 ±
0.2
8410
.28
± 0.
172
10.0
1 ±
0.14
86.
72 ±
0.2
454.
90 ±
0.1
413.
50 ±
0.1
74<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.01a
6G
y +
4.52
± 0
.313
5.02
± 0
.266
6.02
± 0
.267
6.52
± 0
.011
5.02
± 0
.233
4.92
± 0
.254
3.98
± 0
.161
2.54
± 0
.082
Pla
nt
extr
act
<0.0
01b
<0.0
01b
<0.0
01b
<0.1
0b<0
.001
b<0
.001
b<0
.01b
<0.0
01b
8G
y7.
02 ±
0.4
37.
90 ±
0.6
18.
90 ±
0.0
389.
65 ±
0.0
1512
.30
± 0.
052
11.4
8 ±
0.40
No
t su
rviv
edN
ot
surv
ived
<0.0
01a
<0.0
01a
<0.0
01a
<0.0
01a
<0.0
01a
<0.0
01a
8G
y +
7.00
± 0
.006
7.62
± 0
.093
8.50
± 0
.024
8.65
± 0
.38
8.23
± 0
.40
7.03
± 0
.061
5.54
± 0
.040
4.02
± 0
.042
Pla
nt
extr
act
NS
b<0
.01b
<0.0
01b
<0.0
5b<0
.001
b<0
.001
b
Pla
nt
extr
act
3.00
± 0
.83
3.01
± 0
.900
2.98
± 0
.118
2.80
± 0
.450
2.59
± 0
.73.
05 ±
0.8
94.
95 ±
0.9
02.
70 ±
0.9
0o
nly
Ab
no
rmal
cel
l o
f n
orm
al m
ice
wit
ho
ut
any
trea
tmen
t =
2.50
± 0
.132
.a p
, va
lue
con
tro
l vs
no
rmal
. b p
, va
lue
con
tro
l vs
exp
erim
enta
l. N
S,
no
t si
gn
ifica
nt.
Exp
erim
enta
l, p
lan
t ex
trac
t w
as g
iven
1h
bef
ore
irr
adia
tio
n a
t th
e d
ose
rat
e o
f 10
0m
g/k
g b
od
y w
eig
ht.
Co
ntr
ol,
irra
dia
ted
on
ly.
Tab
le 3
. V
aria
tion
s in
the
bin
ucle
ate
cell
coun
t (p
erce
ntag
e) i
n th
e liv
er o
f co
ntro
l an
d ex
peri
men
tal
mic
e
Po
st i
rrad
iati
on
tim
e (i
n d
ays)
Tre
atm
ent
0.25
12
47
1014
28
6G
y7.
495
± 0.
043
7.70
± 0
.023
7.72
± 0
.047
7.49
5 ±
0.04
39.
315
± 0.
085
8.50
± 0
.075
8.29
± 0
.21
7.5
± 0.
020
<0.1
0a<0
.001
a<0
.001
a<0
.10a
<0.0
01a
<0.0
01a
<0.0
1a<0
.05a
6G
y +
7.34
5 ±
0.06
57.
435
± 0.
056
7.50
± 0
.083
6.54
5 ±
0.07
57.
55 ±
0.0
497.
74 ±
0.0
067.
65 ±
0.0
297.
12 ±
0.0
38P
lan
t ex
trac
t<0
.10b
<0.0
1b<0
.05b
NS
b<0
.001
b<0
.001
b<0
.05b
<0.0
01b
8G
y7.
71 ±
0.0
337.
81 ±
0.0
4310
.575
± 0
.038
10.0
0 ±
0.38
10.0
65 ±
0.5
39.
51 ±
0.4
7N
ot
surv
ived
No
t su
rviv
ed<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a<0
.001
a
8G
y +
7.51
± 0
.049
7.79
± 0
.052
8.25
± 0
.012
8.12
± 0
.042
9.54
± 0
.041
8.03
± 0
.008
7.95
± 0
.027
7.82
± 0
.055
Pla
nt
extr
act
<0.0
1bN
Sb
<0.0
01b
<0.0
1bN
Sb
<0.0
2b
Pla
nt
extr
act
7.51
± 0
.31
7.32
± 0
.28
7.42
± 0
.33
7.30
± 0
.21
7.35
± 0
.28
7.38
± 0
.389
7.32
± 0
.35
7.30
± 0
.39
on
ly
Bin
ucl
eate
d c
ell
per
cen
tag
e o
f n
orm
al m
ice
wit
ho
ut
any
trea
tmen
t =
7.31
5 ±
0.07
1a p
val
ue
con
tro
l vs
no
rmal
. b p
, va
lue
con
tro
l vs
exp
erim
enta
l. N
S,
no
t si
gn
ifica
nt.
Exp
erim
enta
l, p
lan
t ex
trac
t w
as g
iven
1h
bef
ore
irr
adia
tio
n a
t th
e d
ose
rat
e o
f 10
0m
g/k
g b
od
y w
eig
ht.
Co
ntr
ol,
irra
dia
ted
on
ly.
610 R. SHARMA AND J. SHARMA
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
Figure 4. (a) Liver of control mouse on day 4 showing severe damage, crenation, binucleated cells, cytoplasmic degranulationvacuolation and absence of sinusoidal spaces (×400). (b) Liver of experimental mouse on day 4 showing better architecture incomparison with control, Kupffer cells and thinner sinusoidal spaces (×400). (c) Liver of control mouse on day 10 showing, hepaticcords in normal arrangement, giant nuclei, binucleated cells, regranulation of cytoplasm and Kupffer cells (×200). (d) Liver ofexperimental mouse on day 10 showing better condition in comparison with control (×400).
with a dose reduction factor (DRF) calculated as 1.33(Sharma, 2002).
This study suggests that Centella asiatica could beuseful in preventing radiation induced tissue damage inclinical radiotherapy, especially in combination withother protectors.
Acknowledgements
We are thankful to the radiotherapy department SMS MedicalCollege and Hospital, Jaipur for providing irradiation facilities. Weare also thankful to the Amsar Private Limited Indore for the supplyof the plant extract as a gift sample.
peroxidase, vitamin E and ascorbic acid in newly formedtissue (Shukla et al., 1999). It also resulted in a severalfold decrease in lipid peroxide levels, as measuredin terms of thiobarbituric acid reactive substances.Asiaticoside enhanced induction of antioxidant levelsat an initial stage of healing has been suggested to bean important contributory factor in the healing proper-ties of this substance (Shukla et al., 1999). Shobi andGoel (2000) have reported that Centella asiatica pre-vents radiation induced behavioural alterations andgrowth retardation in rats. We have also observed thatit also protects against radiation induced mortality ofmice, and protects vital organs of the body against it,

CENTELLA ASIATICA ON HEPATOCYTES 611
Copyright © 2005 John Wiley & Sons, Ltd. Phytother. Res. 19, 605–611 (2005)
REFERENCES
Bandyopadhyay U, Das D, Banerjee RK. 1999. Reactive oxygenspecies: oxidative damage and pathogenesis. Curr Sci. 77:658–666.
Chandraprabha D, Annapurani S, Murthy NK. 1996a. A studyon the membrane structural alterations in the mice treatedwith the plant extract. Indian J Nutr Diet 33: 158–162.
Chandraprabha D, Annapurani S, Murthy NK. 1996b. Testingthe mutagenicity/comutageneticity/antimutagenicity ofselected medicinal plants by the oxidative mutant strainSalmonella typhimurium TA 102. Indian J Nutr Diet 33: 74–79.
Gupta ML, Umadevi P. 1981. Formation of giant hepatocytesin response to radiation. Curr Sci 50: 637–638.
Jaimala, Bharitya HC. 1988. Mechanisms of unusual cellenlargement in the testis of irradiated mouse followingtreatment with a radioprotective thiophosphate. RadiobiolRadiother 29: 429–433.
Kolodny A. 1925. Tissue changes after experimental deepRoentgen irradiation. Am J Pathol 1: 285–293.
Kura S, Sasaki H, Aramaki R, Yoshinaga H. 1978. Binucleate cellformation by x-irradiation. Radiat Res 76: 83.
Lewin K, Millis RR. 1973. Human radiation hepatitis. A morpho-logic study with emphasis on the late changes. Arch Pathol96: 21–23.
Mastuda H. 1956. Histochemical studies of irradiated liver. MedJ Osaka Univ 6: 853.
Mehta S, Mehta L, Mongia SP. 1975. Effect of whole bodyx-irradiation on liver of albino rats. Indian J Exp Biol 13:73–75.
Padma PR, Bhuvaneswari V, Silambuchelvi K. 1998. The act-ivities of the enzymatic antioxidants in selected green leaves.Indian J Nutr Diet 35: 1–3.
Rouillard-Guellec F, Robin JR, Rakotarat Simamangu A,Rastsimamangs, Rasaoanavo P. 1997. Comparative studyof C. asiatica of Madagaskar and Indian origin. Acta BotGallica 144: 489–493.
Scott RB, Budinger JM, Prendergast RAM, Ny Dick I. 1962.Hepatic veno-occlusive syndrome in an American adult.Gastroenterology 12: 631–636.
Sharma J, Sharma R. 2002. Radioprotection of Swiss albinomouse by Centella asiatica extract. Phytother Res 16: 785–786.
Sharma R. 2002. Modulation of Radiation Response by PlantExtract. PhD Thesis. University of Rajasthan, Jaipur,India.
Shobi V, Goel HC. 2000. Centella asiatica prevents gammaradiation induced conditioned taste averstion in rats.Radiobiology International Conference on Radiation Bio-logy, 17–19 February, Trivandrum, India.
Shukla A, Rasik AM, Dhawan BN. 1999. Asiaticoside inducedelevation of antioxidant levels in healing wounds. PhytotherRes 13: 50–54.
Srivastava R, Shukla YN, Kumar S. 1997. Chemistry andpharmacology of Centella asiatica. A review. Indian J MedAromatic Plant Sci 19: 1049–1056.
Wingergar RA, Lutze LH, Hamer JD. 1994. Radiation inducedpoint mutations, deletions and micronuclei in transgenicmice. Mutat Res 307: 479–487.