Embed Size (px)
Citation preview

MODELS FOR PIGMENT PATTERN FORMATION IN THESKIN OF FISHES
K.J. PAINTER
DEPARTMENT OF MATHEMATICS, UNIVERSITY OF UTAH, SALT
LAKE CITY, UT 84112,USA.
Abstract. Thecoloursandpatternsof theskin providesa fascinatingsystemusedfor the studyof patternformationin experimentalandtheoreticalresearchalike. In this article, a brief review of recentwork on the pigmentationof theskin is presented.A mathematicalmodel is shown to be able to capturemanyfeaturesassociatedwith theevolving colourpatternsonjuvenilesbelongingto thegenusof marineangelfish,Pomacanthus. Differentformsof growth leadto verydifferentpatterningphenomena.Thedevelopmentof computationaltoolswhichcanaccuratelyreflectthegeometryandgrowth of the realsystemwill allow thestudyingof the relationshipbetweengrowth and patterningin speciessuchasPomacanthusor zebrafish.
1. Introduction.Acrossthe animal kingdom, a large numberof speciesrely on the colours
andmarkingsof their skin for purposessuchasconcealmentandwarning. Thesophisticationandcontrol hasreachedastonishinglevels in someanimals. Forexample,speciesof bottomdwelling flatfishhave beenshown to adaptrapidly tobackgroundshades[53] andcanassumeacheckerboardtypepatternwhenplacedon theappropriatesurface[45].
Theability to changeskin colourandpatternis essentialfor survival in manyspecies.In speciessuchastheflatfishor thechameleon,thechangeoccursrapidly(on theorderof seconds):suchchangesaretermedphysiological colourchangesand are controlled by hormonalor nerve signals. The more slowly evolvingchangesaretermedmorphological colour changes,andtheseoccur in responseto continuousexposureto stimuli, ascanbeseenby prolongedexposureof skinto sunlight.A particularlystriking exampleoccursin membersof themarinean-gelfishgenusPomacanthus[14, 15]. Several speciesof Pomacanthusdisplayasimilar juvenilepatternconsistingof severalcurvedverticalwhite barson a darkbluebackground.As thefishgrowsin size,new whitebarsareaddedbetweentheolderstripes,first emerging faintandnarrow but wideningasthefishcontinuestogrow. This processrepeatsonceor twice beforethepatternevolvesto the adultform, which canvary drasticallybetweenspecies.The aggressive adultsattackfishwith similarcolorpatterns,andthereforejuvenilesadoptadistinctlydifferentpatternto allow themto safelyswim in anadultsterritory, [16].
The relationshipsbetweenage,growth andpatterninghave attractedinterestfrom experimentalistsandtheoreticiansalike. A simpleone-dimensionalmodel
59

60 From: MathematicalModelsfor Biological PatternFormation
basedon a Turing mechanism[61] wasproposedto accountfor basicaspectsofpatternevolution in the above angelfish[29], and further extensionshave beenincorporatedto accountfor patterningdetails[62, 50]. An experimentalstudyof a link betweengrowth andpatterningin membersof thegenusDanio (whichincludesthemuchstudiedzebrafish)hasalsobeenundertaken[35].
Pigmentcells containnaturalcell markers, namely, the pigmentitself, andthusstudiesof pigmentationhave providedexperimentalistswith a valuablesys-tem for understandingpatternformationin the developingembryo. In addition,studyingthesemechanismsmayhaveimportantconsequencesfor theclinical sci-ences.A numberof diseasesareattributedto defective pigmentation,includingthe condition vitiligo which affects approximately1% of the population. Thisdiseaseis aresultof destructionof melanocytesandcausespatchesof whiteskin.Understandingthemechanismsby whichpigmentcellsmigrateandproliferatetopatterntheskinmayleadto amoresuccessfulcourseof treatment.
In this paperwe presenta brief review of the recentexperimentalandmod-elling researchonthedevelopmentof patternsandcolours.Weproceedto presentanumberof mathematicalmodelsfor pigmentationin speciesof fish.
2. Formation of pigment patterns.A greatbody of researchexists on the physiology of pigmentcells and the
formationof patternswithin theskin. Wereferto thefollowing for greaterdetail:[10, 55,12,18, 17].
2.1. Pigments and chromatophores.Colourandpatternis theresultof pigmentcells(alsocalledchromatophores)
in the dermaland epidermalskin layers. Thesecells are large, branchedandhighly motile andcontainmembraneorganellescalledchromosomesholdingthepigmentgranules.Theorganellesarerapidlyaggregatedor dispersedin responseto nerve or hormonalsignals. A numberof chromatophoretypesexist, corre-spondingto thedifferentpigmentsandcolour. Theseinclude,� Melanophoresareacommonchromatophoretyperesidingin dermaland
epidermalskin layers. In mammalsandbirds,they residein thedermiswherethey areresponsiblefor orangesandblacksby thetransferof pig-mentfrom melanophoredendritesinto hairs,feathersandepithelialcells.In fish, melanophoresgive rise to blackpigmentation,andarefound inthedermallayerswherethey canrespondtonervalandhormonalsignals.� Iridophoresaresmaller, roundcells found in the dermallayersof fish.Thesecellscontainplateletswhich reflectlight resultingin a white/ sil-verycolour.� Xanthophores and erythophores result in bright yellows, orangesandreds.

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 61
2.2. Origin of pigment cells and migration to the skin.Theidentificationof theneuralcrest,atransitorysubpopulationof cellswhich
developsabove theneuraltubein vertebrates,asthesolesourceof pigmentcellswasfirst establishedin amphibians[56] andthenin otherspecies.In fish,it is lessclearasmany speciesdonotdevelopadistinctneuralcrest.It has,however, beenclearlydemonstratedin speciessuchasthelamprey [46].
Pigmentcell precursorsmigratefrom theneuralcrestin a wave-like mannerto uniformly seedthe skin. It is thoughtthat the variouschromatophoretypesoriginatefrom acommonneuralcrestprecursor[3] andcommitmentto aspecifictypeis not establisheduntil localizationin theskin. Themechanismscontrollingtiming andmigrationof pigmentcellsfrom theneuralcrestarelargely unknown.Cellsdo not acquiretheir characteristicpigmentuntil aftermigrationhasended,yet a numberof markershave beendevelopedwhich allow identificationprior topigmentaccumulation.A numberof candidateshave beenproposedto controltiming andmigrationof pigmentcell precursors,including(i) achangein compo-sitionof theextracellularmatrix,(ii) theappearanceof chemoattractant/repellentmolecules,(iii) achangeof cell adhesionproperties.It is alsopossiblethatneuralcrestcell precursorsmaybeforcedontoaspecificpathwaydueto theunattractivenatureof otherregions.
Studiesof mousemutationsaffectingcoatpigmentationhaveprovidedanim-portanttool for understandingfactorsinvolvedin melanocyte development.Twoessentialreceptor-ligand interactionshave beenrevealed: the receptortyrosinekinase,c-kit, with its ligandSteelfactor(SLF, alsocalledstemcell factor),e.g.[21, 64] andthe endothelinreceptorB togetherwith endothelin3 [5]. Solublesteelfactorappearsto have a role in regulatingmelanocyte precursordispersalfrom theneuralcrest,whereasmembrane-boundSteelfactoris requiredfor sur-vival of theprecursorswithin thedermis[63]. In additionto theseobservations,exogeneousSLF hasbeenshown to be importantfor proliferationanddifferen-tiation of themelanocytes[31]. Intriguingly, SLF mayhave a role in promotingchemotacticactivity in melanocytes,[20, 63, 31]. Cellsexpressingfunctionalc-kit receptorsmaybeselectively attractedontothelateralpathway by SLF, whichdiffusesfrom its siteof productionin thedermatomalepithelium.
In addition to SLF, a numberof melanocyte mitogens(chemicalsinducingcell division)havebeenidentified,includingleukotrines,endothelin-1andcertainfibroblastgrowth factors[39, 54,23,65]. Severalof thesehaveadditionallybeenshown to inducemelanocytechemotaxisandchemokineticmovement[24].
2.3. Formation of colours, patterns and colour change.Whenoneareaof the skin is dominatedby a specificcolour, this canbe at-
tributed to an accumulationof pigmentcells of the type producingthat colour.White stripes(suchasthoseon growing Pomacanthus) areoftendueto anabun-danceof iridophores,the redspotsof certaincarparelocal aggregationsof ery-thophorecellsandblackstripesin theangelfisharedueto melanphores.
Despitefew available pigments,a brief glancethroughan encyclopediaof

62 From: MathematicalModelsfor Biological PatternFormation
fishes[9] revealsa staggeringnumberof differentcolours.Themacroscopicallyperceived colour can be attributed to the microscopicorganizationof pigmentcells in theskin. In amphibiansandreptiles,chromatophoresarearrangedin or-deredlayersin the dermisto form the dermalchromatophore unit, [2]. Thesestructurescan createcolours like greenwhich cannotbe formed by the avail-ablepigmentsalone. Similar arrangementsexist in fishes. For example,bluesof the damselfishare derived by a layer of iridophoresbacked by a layer ofmelanophores,[26, 19].
Increasesand decreasesin the numberof dermalchromatophoresleadstomorphologicalcolour changes.The changesresult from proliferation of chro-matophoresand/orthedegradationof terminallydifferentiatedpigmentcells[53].By countingthe numberof chromatophoresover a long periodof adaptationtolight or dark backgroundsin Oryzias, it hasbeenshown that dark adaptationoccursvia increasednumbersof melanophoresand decreasednumbersof leu-cophores,andviceversafor thereverseprocess[58].
Theinteractionsandmechanismswhich leadto patternformationremainlar-gely unknown, however establishmentof patternsin larval salamandershasre-ceived attention. In the larval salamanderAmbyostomatigrinum tigrinum, mel-anophoresscatteruniformly over the flank of the embryo,while xanthophoresremainin aggregatesin premigratorypositions.As thexanthophoresmigrate,themelanophoresrecedeshort distancesto form alternatingbars[11, 47, 51]. Atthis time, a horizontalstripeover the lateralsurfaceof the myotomesdevelopswhich is free of the otherwiseabundantmelanophores.A seriesof experiments[52] suggestthat several factorsare involved in developmentof this stripe, in-cluding interactionsbetweenxanthophoresandmelanophoresandextracellularfactors.Themelanophorefreeregionappearsto developthroughactive retreatofmelanophoresfrom theforming lateralline. Thehorizontalstripesin othersala-mandersarethoughtto developby responseof pigmentcellsto cuesin theECM,[60, 11].
2.4. Reaction-diffusion models in pigmentation.Turing[61] demonstratedthatasimplesystemcomprisingof two reactingand
diffusingchemicalscan,underappropriateconditions,leadto stationarynonho-mogeneouspatterns.This wasproposedby Turing asa mechanismfor morpho-genesis.Theapplicationof Turingpatternsto pigmentationwasmadeby Murray[40,41,42, 43]. In essence,Murrayproposedthatareaction-diffusionmechanismprovidesamorphogenprepatternwhichdictatescell differentiation.An attractivepartof this theoryis thatthemajorityof mammalianpatternscanbegeneratedbysucha mechanism.Similar modelshave beenproposedby Bard [4] andYoung[66].
Thefirstmodellingattemptto fishpigmentationpatternsvia reaction-diffusiontheorywasproposedby KondoandAsai [29]. After observingthe relationshipbetweenfishsizeandthenumberof stripesin themarineangelfish,Pomacanthussemicirculatus, a Turing modelwasproposedwhich predictsthedoublingof the

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 63
numberof peaksof chemicalconcentrationasthedomainlengthdoubles.A com-parisonof striperearrangementin adultP. imperator andnumericalsimulationsof themodelshowedcloseagreement.Attemptsto addressadditionalaspectsofpigmentationin Pomacanthushave sincebeenproposed.Vareaet al. [62] con-sidereddomainswherebyonesideis shapedto reflectthecurvedgeometryof thefish skin. Throughconsidering“enhanced”boundaryconditionssuchthatmodelparametersareelevatedalong certainboundaries,they have demonstratedsev-eral additionalaspectsof Pomacanthuspatterning,suchasthe complicatedpat-ternsseenalongboundaryedgesin Pomacanthusimperator andthe orientationof stripeswith respectto theboundary. Painteret al. (1999)have augmentedthemodelto includecell movement.Thismodelis ableto replicatetheslow insertionof new stripesbetweenolderstripes.Additionally, featuressuchasthecurvatureof juvenile stripesand the transitionfrom stripesto spotsas P. semicirculatusmaturesto adulthavebeenshown in this model.
An activatorinhibitor modelhasalsobeenappliedto theshellpatternsof mol-lusks.Thebookby Meinhardt[38] demonstrateshow many of theshellpatternsseenin naturecanby replicatedin a reaction-diffusionmodel.
3. The cell movement model.In this sectionwe considera mathematicalmodel for pigmentpatternfor-
mationin the skin of fishes. In this model,the following aspectsaretaken intoaccount:� Whena specificareaof theskin is dominatedby onecolour, it is dueto
anaggregationof chromatophoresof thetypeproducingthatcolour.� Morphologicalcolourchangesof theskin aredueto anincreasedpopu-lationof chromatophores.Weshouldmention,however, thatsuchcolourchangescanalsobebroughtaboutby increaseddepositionof pigmentinepithelialcells.� Patterningin theskin is controlledby distributionof oneor morechemi-cal factors.A numberof candidateshavebeendiscovered,yetnodefini-tive identity existsfor a so-calledmorphogen.Kirschbaum[28] demon-stratedvia transplantationexperimentsthat differentiationof melano-phoresin the zebrafishoccurredthroughresponseto local cuesin thedermis.� Increasesin the total numberof pigmentcellsarevia a combinationofproliferationfrom chromatophoresor from undifferentiatedstemcells.Thestemcellsareundifferentiatedpigmentcell precursors.Furtherdif-ferentiationof stemcells into a specificpigmentcell type is likely toresultfrom extracellularfactorsin thedermis/extracellularmatrix.� Initial pigmentcell distributionvariesfrom speciesto species.In theze-brafish,thelarvalpigmentpatternconsistsof four laterallinesof melano-phores:dorsal,septal,abdominalandventral.Pigmentcellsin larval Po-macanthusarcuatusinitially form auniformgraypigmentationwhenthefish is between5 and7 mmin length.Thejuvenilepigmentpatterncon-

64 From: MathematicalModelsfor Biological PatternFormation
sistingof fiveverticalwhitebarsdevelopsfrom thisuniformdistribution[27].� Several lines of evidencepoint to chemotacticguidanceof migratingmelanocyte precursorsfrom the neuralcrestto the dermis[10, 6, 59,20]. Potentialchemoattractantsincludesteelfactor[31] andendothelin-1 amongstothers,[24]. In reality, cell movementmay be inducedbymany mechanisms,includingrandommotility, haptotaxisandcell adhe-sion.
Very little is currentlyknown concerningthevariousinteractionsbetweenthecell types,betweenthecellsandthechemicals,andbetweenthechemicalswitheachother, andit is not feasibleatthismomentto developahighly detailedmodelfor pigmentationpatterning.We shall restrictattentionto simplemodelsof pat-terningwhich replicatethebiologicaldata.
In this sectionwe considera mechanismfor generatingthe stripedpatternsin Pomacanthusbasedon a model for cell movementin responseto gradientsof chemicalprepatternsderived from the reaction-diffusion system. The domi-nantchromatophoretypesinvolved in juvenile stripedevelopmentare likely tobemelanophores,whichwedenote
���������, andiridophores,� ��� ����
. During thetransitionto adult stages,it is probablethat otherchromatophoretypesbecomeimportant,andtheeffectsof thesemaybeafactorin theobservedtransformation.For example,yellow horizontalstripesof P. imperator are likely to be derivedfrom xanthophores.In this model,we make the simplificationof assumingthatjust onecell type is chemotactic,andthat it is chemotacticto a singlechemicalspecies.Here we assumeit is the iridophorecells, althoughwe could equallypostulatethat it is themelanophoreswithout qualitative differencesin modelbe-haviour. Onafixeddomain,� , theequationsaregivenby� �� ������������������� � �!�"#��$%&�
(3.1)� �� �'����������()�*��( � � ��"#��$%&�� "� � �,+.-��0/ " �*� ��1���"#�$�&�� $� � �,+.23� / $ �54 ��1��"#��$%&6��� and ��( areflux termsfor themelanophoresandiridophores,respectively, andthis containscontributions from randomcell movementfrom chemotaxis. Wesimplify themodelby assumingthatonly thetwo cell typesresidein thedermis,andthat the total cell densityremainsconstant.Therefore,on a constantsizeddomain,we cantake ���7�8��(9�;: (proliferationanddegradationof pigmentcells balance)and �<� � �%���!��( �=: . Thus,melanophoresthemselvesdo notrespondto the chemicalgradients,however they experiencea chemotacticfluxtypecomponentthroughdisplacementby themoving iridophorecells. Zerofluxis assumedat theboundary.

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 65
3.1. Patterning for different growth types.During embryonicand juvenile stages,animalsundergo considerabletissue
growth anddeformation.Theeffectsof suchgrowth on patterningis beautifullyillustratedby theevolving pigmentationpatternsseenonspeciesof amphibia,rep-tiles andfish and,clearly, modelsproposedto explain featuresof embryonicde-velopmentmustconsiderthepotentialeffect of domaingrowth on thepatterningprocess.For example,by consideringthegrowth of thedevelopingalligator jaw,it hasbeendemonstratedthatasimplereaction-diffusiontypemechanismmayac-countfor thesequenceof toothprimordia,[30]. Theneedto considertheeffectsof growth onpatterninghasbeenillustratedfor thereaction-diffusionmechanismononeandtwo-dimensionaldomains,[50,49]. In onedimension,patterningsuchthatthenumberof concentrationpeaksdevelopingthroughthereaction-diffusionmechanismevolves throughthe modedoublingsequence>?�,@0�BAC�,D0� 6E6E6only occursfor certainparameters.In two dimensions,fasterratesof growth canleadto highly convolutedstripesor no spatialpatternat all, ratherthana regularpatternof stripes.
In the modelsconsideredto datedomaingrowth hasbeenincorporatedin asimple manner, anda rigorousincorporationof tissuegrowth anddeformationhasyet to beundertaken. Evenundera simpletreatmentit is possibleto demon-stratethatdifferenttypesof domaingrowth mayleadto verydifferentpatterning.This is demonstratedin the following examplefor a reaction-diffusionequation.Supposeevolution of a chemical,denotedby F �������
, evolveson theconstantdo-mainaccordingto � F� � �,+G� / FH�*� � F &6If we assumethat � changesasa function of time, thenit is straightforward toshow throughapplicationof ReynoldsTransportequation[?, 8]� F� �I�J����KLFM�,+G� / F ��� � F &� (3.2)
whereu � � �LN � �definesthefluid flow. Weconsidertwo simpletypesof growth
for agrowing one-dimensionaldomain, O : �P�����RQ.
3.1.1. Uniform growth.For uniform growth, we assumefor SUT � : WVX� : �&PY
, SUT ���� �ZSUT � : [P������N�P�� : .
Clearly, we have" ��S P \�N�P
, (wherePH\
is thederivative with respectto�), and
Equation(3.2) is givenby,� F� �I� F PH\P]� S PH\P � F� S �,+ � / F� S / ��� � F &6We canconvert theaboveequationon a one-dimensionalgrowing domainontoadomainof constantsizeby thetransformation,
� S ����M^_��`U��ab � � S N�P�����&����. It
is easyto demonstratethatthis transformationleadsto� F� �.� +P / � / F� ` / ��� � F � PH\P5F 6

66 From: MathematicalModelsfor Biological PatternFormation
Theuniform domaingrowth modelleadsto anequationfor evolution of patternin the reaction-diffusion systemwhich can be solved with a simple numericalscheme.Thismayrepresentagoodapproximationfor growth of theskin,wereweto assumethatnutrientsrequiredfor cell proliferationweresupplieduniformly totheskin from beneath.It is, however, a simplificationof domaingrowth in livingsystems.Experimentaldatacollectedon growth of zebrafishandrelatedspeciesthroughlarval andjuvenilestagesto adult [35] indicatethatdifferentregionsofthebodyaregrowing atdifferentrates.
3.1.2. Boundary growth.Underboundarygrowth, we assumethat for SUT � : cVd� : �PY
, SUT ���� �=SUT � : .
This representsgrowth at theboundaryandwe have" �!: . We follow theabove
procedureandusethesametransformationontoa domainof constantsize. Thisgivesthefollowing equationdefiningpatternevolution,� F� �.� +P / � / F� ` / � `ePH\P � F� `f�*� � F Boundarygrowth occursduringextensionof thedevelopingaxonsin thenervoussystem.Thedevelopingneuronconsistsof anervecell body(or soma),alongthinaxonand,at theaxontip, thegrowth conefrom which filopodiaextendto sensetheenvironmentfor guidancesignals.Growth of theaxonoccursat the level ofthe growth cone,[22]. Boundarygrowth may alsobe importantwith respecttoskingrowth if thenutrientsweresuppliedfrom specificbodyregions.
3.1.3. Simulations under uniform and boundary growth.We comparethe different typesof growth above by numericalsimulation.
Kineticsfor thereaction-diffusionsystemarebasedona two-speciessystempro-posedby LengyelandEpstein[32] to accountfor spatialpatternsgeneratedin theCIMA chemicalreaction,[7, 48]. Themodelis,� "� � �Z+g- � / "� S / �*h�ij� " � A "U$>)� " / (3.3)� $� �G�khmln+g2 � / $� S / ��h / hml o " � "U$>)� " /qpwhere +g- � +g2 � h�i � h / � h3l areall positive constants.It is straightforwardto deter-minetheparameterspacefor Turingpatterning.In Figure1 we compareresultsof numericalsimulationsfor the following fourcases:� Exponentialuniformandboundarygrowth
P����� � PHrtsEuwvx��y���, Figure1
(a)and(b).� Linear uniform andboundarygrowthPz���� � Pjr{� >z� y���
, Figure1 (c)and(d).
Under uniform exponentialdomaingrowth, Figure 1 (a), we seethe modedoublingsequenceof patterningwhichhasbeendescribedpreviously, [1, 29,50],
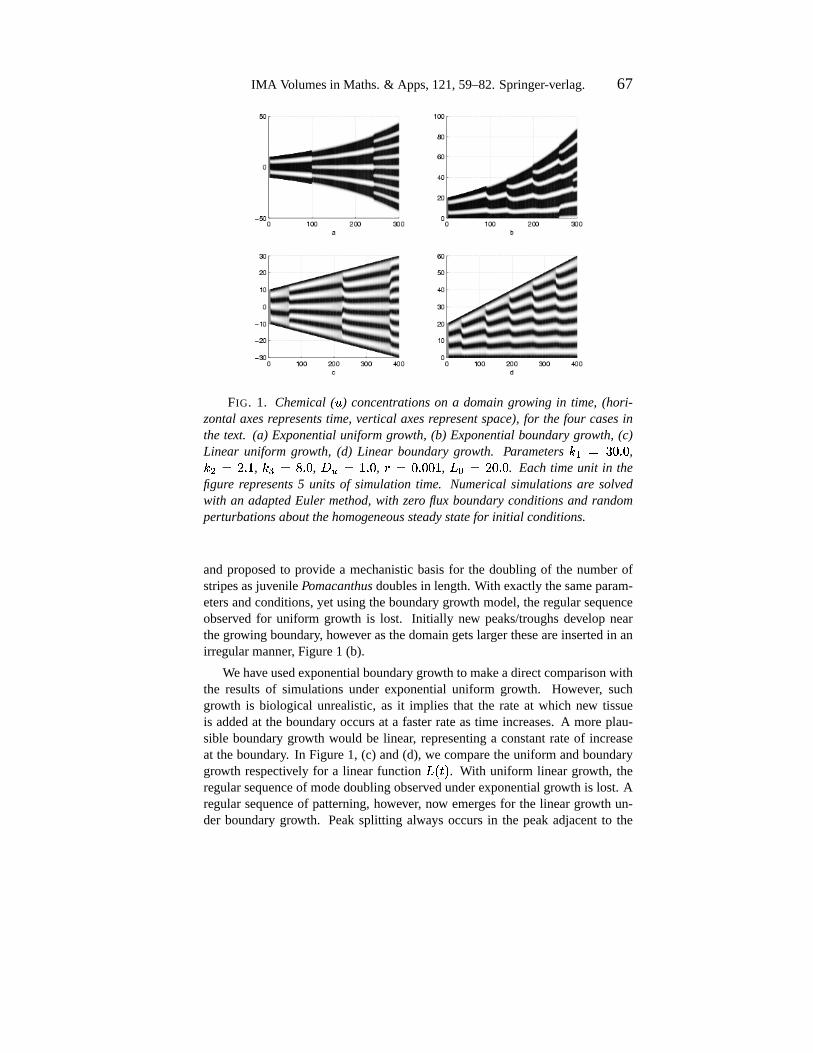
IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 67
FIG. 1. Chemical(") concentrationson a domaingrowing in time, (hori-
zontalaxesrepresentstime, vertical axesrepresentspace),for the four casesinthe text. (a) Exponentialuniform growth, (b) Exponentialboundarygrowth, (c)Linear uniform growth, (d) Linear boundarygrowth. Parameters hei|�~}3: 6 : ,h / ��@ 6 > , h3l���D 6 : , +g-|��> 6 : ,
y ��: 6 :m:%> , PHr ��@3: 6 : . Each time unit in thefigure represents5 units of simulationtime. Numericalsimulationsare solvedwith an adaptedEuler method,with zero flux boundaryconditionsand randomperturbationsaboutthehomogeneoussteadystatefor initial conditions.
andproposedto provide a mechanisticbasisfor the doublingof the numberofstripesasjuvenilePomacanthusdoublesin length.With exactly thesameparam-etersandconditions,yet usingtheboundarygrowth model,theregularsequenceobserved for uniform growth is lost. Initially new peaks/troughsdevelop nearthegrowing boundary, however asthedomaingetslargertheseareinsertedin anirregularmanner, Figure1 (b).
Wehaveusedexponentialboundarygrowth to makeadirectcomparisonwiththe resultsof simulationsunder exponentialuniform growth. However, suchgrowth is biological unrealistic,as it implies that the rate at which new tissueis addedat the boundaryoccursat a fasterrateastime increases.A moreplau-sible boundarygrowth would be linear, representinga constantrateof increaseat theboundary. In Figure1, (c) and(d), we comparetheuniform andboundarygrowth respectively for a linear function
P�����. With uniform linear growth, the
regularsequenceof modedoublingobservedunderexponentialgrowth is lost. Aregularsequenceof patterning,however, now emergesfor the lineargrowth un-der boundarygrowth. Peaksplitting alwaysoccursin the peakadjacentto the

68 From: MathematicalModelsfor Biological PatternFormation
growing boundary, and thosefurther from the growth retain their spatial loca-tion throughoutgrowth. Thus the patterningof peaksevolves in the sequence}��5A������|���|���|D�� 6E6E6
.
FIG. 2. Order of adult stripe developmentin the zebrafish. Stripes1 and2 appearalmostsimultaneouslyfrom larval lines (dashed).Subsequentstripesappearin theorder indicated.
This latter behaviour is reminiscentof the mechanismby which new stripesemerge alongthe body of the adult zebrafish,Figure2. Stripes1 and2 appearsimultaneouslyfrom thelarval pattern.Additional stripesappearin a specificse-quence:3 appearsventrallyof 1, 4 appearsdorsallyof 1, 5 appearsventrallyof 3,and6 appearsdorsallyof 4. Stripescontinueto beadded,retainingthissequence,asspacedictates. This sequencemay suggestthat growth in the zebrafishmaybe morecloselyapproximatedby boundarygrowth alongthe dorsalandventraledges.
In summary, via the two typesof growth we can force the patterningintodifferent typesof sequence.In boundarygrowth, the numberof peaksof thereaction-diffusionsequenceprogressthroughtheorder >x��@H�'}j�CAY�'�j�'�H� 6�6�6
,whereasexponentialuniformgrowth givesapeakdoublingsequence> ��@W��A��D���>n��� 6E6n6
.
3.2. Chemotactic-cell model under uniform growth.We considernumericalsimulationof the full chemotactic-cellmodelwith a
uniformly growing domainincorporated.A detailedinvestigationinto thevariousbehaviours this model can show hasbeenpresentedelsewhere,[50]: Here webriefly explain how this model replicatesthe patterningphenomenaof juvenilePomacanthusdevelopment.Thegrowth hereis classifiedas logistic, stipulatingthat initially the fish grows in a mannerapproximatingexponentialgrowth, buteventuallythegrowth rateslowsandthefish approachesa maximumsize.
This sectionconsiderstwo model formulations: (i) The zero cell feedbackmodel,and(ii) thecell feedbackmodel. In the formerwe have no effect on thechemicalsby thecells:Chemicalconcentrationpatternsevolveindependentlyandcellsmove in responseto thegradients.In thesecondmodelwe considera formof chemicalregulationby the cells by control of the rateof chemicalsynthesis.For the two-dimensionalgrowing domain, O : �P i ����RQj� O : �P / ����RQ , scaledonto a

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 69
domainof constantsize,wehave� �� �'� >P / i �� S o +0( � �� S � � r��� � "� / � � "� S p � >P //�� ` o +0( � �� `�� � r��� � "� / � � "� ` p� "� � �,+.- o >P / i � / "� S / � >P //
� / "� ` /�p � �J� � � " � A "U$>)� " / � ��y iL� y / �"#�� $� � �!hmln+g2 o >P / i � / $� S / � >P //� / $� ` /qp �*h / hml o " � "U$>)� " /%p � ��y i�� y / �$q�
(3.4)
whereP i ���� � P / ���� � Pjrts�uevx��y����N���� � s�uevx��y����
. See[50] for a derivationoftheseequations.Weconsiderzeroflux boundaryconditions,andinitial conditionsweightedsuchthatastripepatternwill initially form.
3.2.1. Case 1: No cell feedback, �=���x�?���m�%�M e¡E¢b�)¡g�¤£#¥ .Simulationsin this casehave beendescribedpreviously, [50], anda succes-
sionof framesatdifferenttimesareplottedin Figure3 for thechemoattractant,",
(a) – (e),andthecell density, (f) – (j). Chemicalconcentrationsevolve throughastripedoublingsequenceand,with nofeedbackfrom cellsto chemicals,cellswillsimplymovein responseto themorphogengradients.Thiscreatesstripesappear-ing faint andnarrow at first, but growing with time. As the domainapproachesthemaximumsize,theregularsequenceof stripesbreaksinto a patternof spots.Domaingrowth appearsto hold thestripepatternduringinitial stages,but asthethedomainapproachesits maximumsize,thepatternrelaxesto its favoredTuringwavelength.This hasbeencomparedto thesequenceof stripesforming in juve-nile Pomacanthussemicirculatusandthesubsequenttransitionto thespottedadultpattern.During thetransitionfrom juvenileto adultpattern,a “mixedpattern”isobserved wherebythe iridophoresarereorganizinginto spots,yet the stripesofthejuvenilecanstill beseen.Such“transitionpatterns”arealsoobservedduringthe transitionfrom juvenile to adult patternin semicirculatus. Thesetransitionstagesarecomparedin Figure4.
To understandtheeffectsof differing chemotacticstrengths,we vary thepa-rameter � r
and examine the resultingcell densitypatterns. One dimensionalresultsfor threeseparatesimulationsareshown in Figure5. For the “normal”chemotacticsensitivities,weobserve theslow insertionof new stripes,consistentwith thepatterningon Pomacanthus. Similar patterningis observedwhenvary-ing � r
by plusor minusanorderof magnitude.Increasesof � rby plusor minus
two ordersof magnitude,however, result in very differentpatterning.In Figure5 (b), � r
hasbeenincreasedby two ordersin magnitude.Althoughinitial peaksdevelopaspreviously, no new stripesform. Dueto thestrongchemotacticeffect,all available iridophoresarepulled into the initial stripes,leaving few to createsecondarystripes.A decreaseby two ordersof magnitude,(c), resultsin thedis-appearanceof pattern.Whenthechemotacticeffect is very weak,emerging cellaggregateshave smallamplitudes,suchthat they arelikely to beindiscernibleatthemacroscopiclevel.

70 From: MathematicalModelsfor Biological PatternFormation
FIG. 3. Numericalsimulationsfor the cell movementmodelon a growingtwo-dimensionaldomain.For convenienceof representation,thegrowingdomainhas beenscaledonto one of constantsize. (a) – (j) The zero feedback modelshownfor chemical
", (a) – (e), and cell density, (f) – (j), at
� ��@3:3: , (a) and(f),
� �¦>n�3:m: , (b) and (g),� �8}m�3:m: , (c) and (h),
� �;Aw@�:m: , (d) and (i), and� ���m@3:m: , (e) and (j). Correspondingplots for the feedback modelare shownin (k) – (t). We use h�i§�8>E: 6 : , h / �¨@ 6 @ , hml��¨D 6 : , +g-X�©: 6 :%> , +.29�©: 6 @m� ,+0(0� � r �<� 6 : � >n:�ªb« . y �<: 6 :m:%> , � �<� 6 : ,
PHr ��> 6 � ,� ��> 6 : . Numerical
simulationsusean ADI methodwhich hasbeenadaptedto includechemotacticcell movement.
3.2.2. Case 2: Cell feedback, �=���L�'�¬£Um�x®x�n¯b°²±'³��L� . We con-sidertheeffect of a cell feedbackwhich takestheform of regulationof chemical
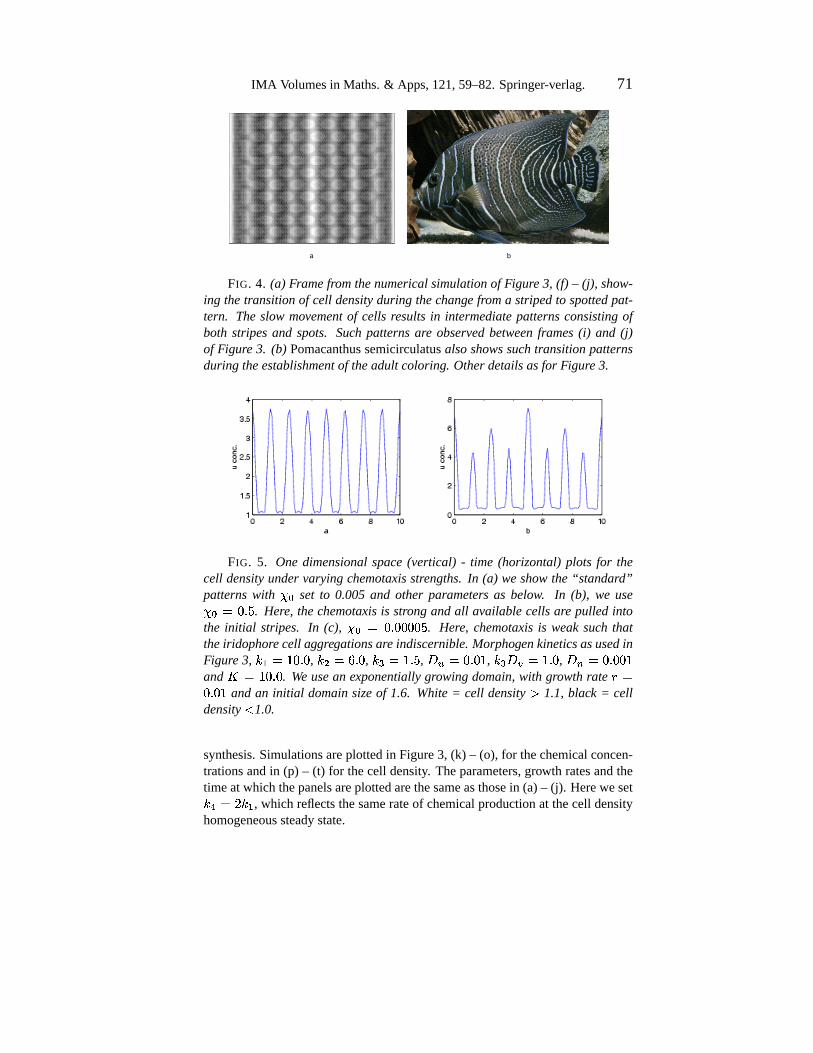
IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 71
a b
FIG. 4. (a) Framefromthenumericalsimulationof Figure3, (f) – (j), show-ing thetransitionof cell densityduring thechange froma stripedto spottedpat-tern. Theslow movementof cells resultsin intermediatepatternsconsistingofboth stripesand spots. Such patternsare observedbetweenframes(i) and (j)of Figure 3. (b) Pomacanthussemicirculatusalsoshowssuch transitionpatternsduring theestablishmentof theadult coloring. Otherdetailsasfor Figure3.
FIG. 5. One dimensionalspace(vertical) - time (horizontal) plots for thecell densityundervaryingchemotaxisstrengths.In (a) weshowthe“standard”patternswith � r
set to 0.005 and other parameters as below. In (b), we use� r �<: 6 � . Here, thechemotaxisis strongandall availablecellsare pulled intothe initial stripes. In (c), � r �8: 6 :m:3:m:{� . Here, chemotaxisis weaksuch thattheiridophorecell aggregationsare indiscernible. MorphogenkineticsasusedinFigure 3, hei��´>n: 6 : , h / �d� 6 : , h3lI��> 6 � , +g-G�k: 6 :�> , h3ln+.2g�´> 6 : , +.µ'�k: 6 :3:%>and
� ��>E: 6 : . We usean exponentiallygrowingdomain,with growthratey �: 6 :%> and an initial domainsizeof 1.6. White= cell density ¶ 1.1, black = cell
density· 1.0.
synthesis.Simulationsareplottedin Figure3, (k) – (o), for thechemicalconcen-trationsandin (p) – (t) for thecell density. Theparameters,growth ratesandthetimeatwhich thepanelsareplottedarethesameasthosein (a)– (j). Hereweseth3¸��d@mh�i , which reflectsthesamerateof chemicalproductionat thecell densityhomogeneoussteadystate.
72 From: MathematicalModelsfor Biological PatternFormation
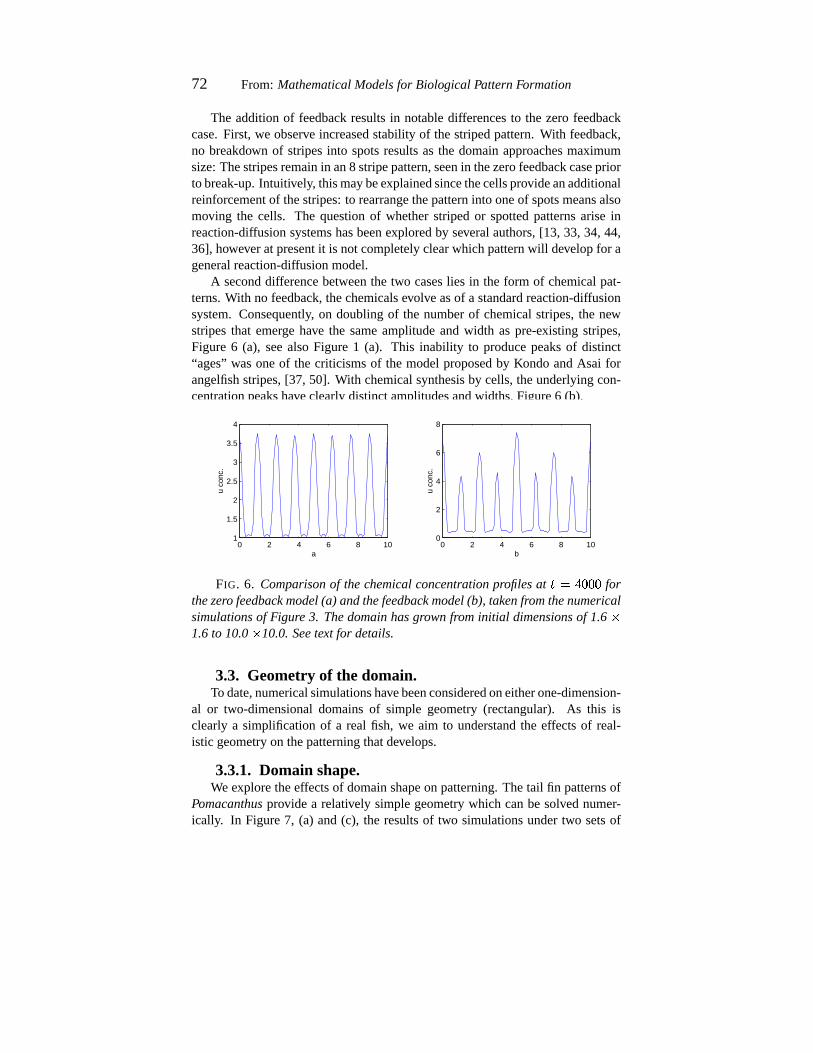
The additionof feedbackresultsin notabledifferencesto the zerofeedbackcase.First, we observe increasedstability of thestripedpattern.With feedback,no breakdown of stripesinto spotsresultsas the domainapproachesmaximumsize:Thestripesremainin an8 stripepattern,seenin thezerofeedbackcasepriortobreak-up.Intuitively, thismaybeexplainedsincethecellsprovideanadditionalreinforcementof thestripes:to rearrangethepatterninto oneof spotsmeansalsomoving the cells. The questionof whetherstripedor spottedpatternsariseinreaction-diffusionsystemshasbeenexploredby severalauthors,[13, 33,34, 44,36], howeveratpresentit is notcompletelyclearwhichpatternwill developfor ageneralreaction-diffusionmodel.
A seconddifferencebetweenthe two caseslies in the form of chemicalpat-terns.With no feedback,thechemicalsevolve asof a standardreaction-diffusionsystem.Consequently, on doublingof the numberof chemicalstripes,the newstripesthat emerge have the sameamplitudeand width as pre-existing stripes,Figure 6 (a), seealsoFigure 1 (a). This inability to producepeaksof distinct“ages” wasoneof the criticismsof the modelproposedby KondoandAsai forangelfishstripes,[37, 50]. With chemicalsynthesisby cells,theunderlyingcon-centrationpeakshaveclearlydistinctamplitudesandwidths,Figure6 (b).
0 2 4 6 8 101
1.5
2
2.5
3
3.5
4
a
u co
nc.
0 2 4 6 8 100
2
4
6
8
b
u co
nc.
FIG. 6. Comparisonof thechemicalconcentration profilesat� �´A{:m:3: for
thezero feedback model(a) andthefeedback model(b), takenfromthenumericalsimulationsof Figure 3. Thedomainhasgrownfrominitial dimensionsof 1.6
�1.6to 10.0
�10.0.Seetext for details.
3.3. Geometry of the domain.To date,numericalsimulationshavebeenconsideredoneitherone-dimension-
al or two-dimensionaldomainsof simple geometry(rectangular). As this isclearly a simplificationof a real fish, we aim to understandthe effectsof real-istic geometryon thepatterningthatdevelops.
3.3.1. Domain shape.We exploretheeffectsof domainshapeon patterning.Thetail fin patternsof
Pomacanthusprovide a relatively simplegeometrywhich canbe solved numer-ically. In Figure7, (a) and(c), the resultsof two simulationsundertwo setsof

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 73
initial conditionsareconsidered.Thosefor (a)createhighly curvedstripeswhichresultsin complexity of patternat theedgesof thetail fin. This is comparedwiththe patterningon the tail fin of P. imperator, (b). The initial conditionsin (c)giveriseto “gently” curvedstripes,resultingin asimplepatternof curvedstripes,asseenon the tail fins of juvenile P. semicirculatus, (d). For both simulations,thegeometryof thedomainaffectsthepatternthatdevelopsby arrangingstripesperpendicularto theboundaries.This forcesthestripesinto maintainingcurvedpatterns.
FIG. 7. Comparisonsof patternsof the reaction-diffusionmodelwith Po-macanthustail fins on a domaingeometryreflectingthe actual fin. Simulation,(a), and P. imperatortail fin, (b). Simulation,(c), and P. semicirculatustail fin(d). Thesimulationsin (a) and(c) havebeenobtainedusingslightlydifferentsetsof initial conditions,with thesamereaction-diffusionmodelasconsideredbefore.Initial conditionsin (a) give rise to more highly curvedstripes. See(Painter etal., 1999)for details. Parameters: h�i���}3: 6 : , h / ��> 6º¹ � , hml.��D 6 : , +g-f��> 6 : ,+g2��»> 6 � .
Themethodof incorporatingboundaryshapehasinvolvedmeasuringdimen-sionsof the fin from photographicimages,calculatingsuitablefunctionsto ap-proximatethe shape,andprogrammingboundaryconditionsaccordingly. This,of course,turns out to be a time-consumingand laboriousprocess.A numer-ical schemehasbeendeveloped(seecontribution by D. Bottino, this volume)which will solve a systemof reaction-diffusionequationsfor a Voronoi tessella-tion. In Figure8 (a), theresultof a numericalsimulationis shown, which solvesthereaction-diffusionsystemgivenby Equations(3.3)onanirregulardomainus-ing thismethod.Theboundaryitself wascreatedby tracinganimageof ajuvenileP. semicirculatusat 6 monthsold (shown in Figure8 (b)) which is convertedintoaVoronoigrid by theschemeof Shewchuk,[57].

74 From: MathematicalModelsfor Biological PatternFormation
FIG. 8. (a) Patterngeneratedbysolvingthereaction-diffusionsystemon theirr egular domaindeterminedby tracing the boundaryof the real fish shownin(b). Equationssolvedusing the numericalschemeof Bottino (seethis volume)on a Voronoi grid. Figure (b) taken from Kondoand Asai (1995). Parameters:h�i��´}3: 6 : , h / �=> 6¼¹ , hmlc�´D 6 : , +g-½�»: 6 :m:{@m� , +g2C��: 6 :%>¾@m� . Initial conditionsconsidera disturbanceof thehomogeneoussteadystatein theareaof thetail fin.Thisforcesa patternof curvedstripes.
3.3.2. Three dimensional patterning.Fishchromatophorecellsresideprimarily in thedermisand,sincebodylength
anddeptharemuchgreaterthanthe thicknessof the skin, (on the orderof 0.1-1 mm, althoughthereis wide variationbetweenspecies),it is oftenadequatetomodeltwo dimensionsonly. In somespecies,however, patterningoccursonafinescale:thefirst two stripesappearingin zebrafishareapproximately1 mm apart.In suchcasesit maybenecessaryto considerthreedimensions.
Numericalsimulationsfor Turingstructuresdevelopingin a two speciesreac-tion-diffusionmechanismon a three-dimensionaldomainareplottedin Figure9.Thetwo standardpatternsareshown onthin layers:stripesandspots.Here,depthis smallandhasnoeffect on thepatterning.
Depth hasnegligible effect on pigmentationwhen the reactionoccursuni-formly throughout,but mayplay a role whennon-uniformityof reactionis intro-duced.For example,if reactantscomefrom lower layers,agradientin thesupplymay occur. The potentialeffect canbe understoodby imposinga gradienton aparameterthroughthe depth. Jensenet al. [25] consideredsucheffects in twodimensionsfor the kineticsgiven by Equations(3.3) undervariationof the pa-rameterh / . It wasnotedthat structureof the patternmoved throughregionsofspatialhomogeneityto spottedpatternsto stripes.Variationof this sameparam-eter in threedimensionsshows similar transitions,aswe show in variousslicesof the threedimensionalpatternplotted in Figure 10. At lower h / the patternconsistsof stripes,howeverasthis is increasedweobserve transitionto spotsandhomogeneity.
In summary, whena parametervariesthroughdepthit may disrupt the pat-terning. This may be significantif the parameterregion for patterningis small,

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 75
FIG. 9. Three-dimensionalpatternsin a Turing system.Top left: On a thindomain,the patternof stripesis maintainedthroughoutthe thicknessof the do-main. Bottomleft: A similar effectoccurs whenthepatternis oneof spots.Topandbottomright: Correspondingpatternsfor thicker layers. Slicesthroughthepatternshowsa largeamountof structureand,in threedimensions,patternsshowmuch lessregularity thanthosein twodimensions.Theregularityof thehexagonalpatternsseenon thetwo dimensionallayer is unlikely to developwhenthicknessbecomessignificant.Parameters use h�i���}3: 6 : , h3l?��D 6 : , +g-'��> 6 : , +g20�<> 6 �and h / ��> 6 A for stripedpatterns,h / �k@ 6 D for spots.EquationssolvedusinganEulermethodwith zero flux boundaryconditions.
in which casesmallvariationsin theparametermayshift thereactionoutsidetheparameterspacefor patterning.

76 From: MathematicalModelsfor Biological PatternFormation
FIG. 10. Simulationsof threedimensionalTuring patternsunderlinear vari-ation of a kineticparameter. Slicesshowhowpatternvariesalong thedirectionof thisvariation. Patterningchangesfromstripes(left side)to spots(middle)andhomogeneity(right). Simulationsusethesamekineticsasin previoussimulationswith h�iC�©}3: 6 : , h3l½�¨D 6 : , +g-X��> 6 : , +g29�8> 6 � and h / variesfrom 1.4 (left)to 5.6 (right). Domaindimensionsare @�: � A{: � @3: . Solutionsshowpatternat� ��>n@m� . Simulationsusean Eulermethodwith zero fluxboundaryconditions.
4. Discussion. In this article, recentexperimentaldataon the develop-mentof skin pigmentationpatternshasbeenreviewed. Themathematicalmodelpresentedhereandin earlierwork, [50], hasbeenshown to becapableof repli-catingmany of thefeaturesobservedduringevolution of thepigmentpatternsinspeciessuchasPomacanthussemicirculatus.
Theunderstandingof patternsdevelopingvia a reaction-diffusiontypemech-anismon anirregulargrid hasreceivedcomparatively little attentionto date.Thedevelopmentof numericalschemesin whichadeformingirregulardomainis con-sideredwill allow for simple incorporationof existing biological dataon fishgrowth into the modeland the understandingof patternformation in suchsys-tems.
The modelling to datehasconsidereda continuumapproachto understandmacroscopicpatterningfeatures. Experimentaldatain zebrafishand salaman-derssuggeststhat local interactionsbetweenpigmentcell typesmay contributeto the patterningprocesses.One future direction is to employ a hybrid dis-

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 77
crete/continuumapproachwherebycellsareconsideredasdiscreteparticles.Thisapproachwould allow understandingof how differenttypesof microscopicrulesfor movementgive riseto differentmacroscopicpatterning.
A numberof areasareopento theoreticaltreatment.As mentionedabove,thequestionof whetherstripesor spotsoccurin a generalreaction-diffusion modelhasyet to beansweredsatisfactorily. Patternshavealsobeenobservedto undergobreakupfrom regularstripesinto convolutedstripesundervariousformsandratesof growth, [49, 50]. Whetherthesepatternsrepresentsa form of spatialchaosisanintriguing question.
Themodelhasbeenshown to reproducemany of thefeaturesassociatedwithpigmentationin Pomacanthussemicirculatus. A principalpredictionof this andprevious modelsis that the appearancesof the new “interstripes” occurswhenthe fish hasapproximatelydoubledits previous length. Growth of angelfishinanaquariumenvironmentis limited by thesizeof thetank(e.g. see[9]) andtheabove predictioncould thereforebetestedby limiting thegrowth rateof thefishin this manner. However, dueto theslow rateof growth (approximatelyoneanda half yearsto reachadult),largesize(15 inches)andtheterritorial natureof Po-macanthus, thesefishareunsuitableaslaboratoryanimals.A morewidely studiedspeciesis thezebrafish,Daniorerio, whichis small,easyto breedandhasatrans-parentskinallowing for relatively straightforwardobservationsof cell movementandpatternformation.We arecurrentlyapplyingthemodelto patternformationin thezebrafishto developanumberof experimentallytestablepredictions.Exci-sionandtransplantexperiments,wherebya fragmentof skin tissuefrom a donoris removed andeither replacedat a new orientationor transplantedonto a hostfish (which hasalsohada fragmentof skin removed), canbe easilyperformedwithin themodellingframework. Comparisonof themodelresultswith existingtransplantexperiments(e.g. [28]) in zebrafish,togetherwith thedevelopmentofnew predictionswill beusedto testthesuitability of themodelasa mechanismfor pigmentation.
Themechanismsunderlyingpatternformationin areassuchaspigmentationarestill poorlyunderstood.For thisreason,modellingthusfarhasconcentratedondevelopingsimplemodelswhichcapturebasicfeaturesof thepatterning.As moresophisticatedtechniquesbecomeavailable,andmany of the currentambiguitiesareresolved,themodelwill beadaptedaccordingly.
Acknowledgments I would like to thankD. Bottino for providing thenumericalcodeandassistingmewith simulationsto solve the reaction-diffusionequationson anirregulargrid.

78 From: MathematicalModelsfor Biological PatternFormation
REFERENCES
[1] P. Arcuri andJ.D.Murray. Patternsensitivity to boundaryandinitial condi-tionsin reaction-diffusionmodels.J. Math.Biol., 24:141–165,1986.
[2] J.T. Bagnaraand M.E. Hadley. Chromatophores and Color Change.Prentice-Hall,EaglewoodCliffs, New Jersey., 1973.
[3] J.T. Bagnara,J.Matsumoto,W. Ferris,S.K.Frost,W.A. Turner, T.T. Tchen,andJ.D.Taylor. Commonorigin of pigmentcells. Science, 182:1034–1035,1979.
[4] J.B.L.Bard. A modelfor generatingaspectsof zebraandothermammaliancoatpatterns.J. Theor. Biol., 93:363–385,1981.
[5] A. GreensteinBaynash,K. Hosoda,A Giaid, J.A. Richardson,N. Emoto,R.E. Hammer, and M. Yanagisawa. Interaction of endothelin-3with endothelin-breceptoris essentialfor developmentof epidermalmelanocytesandentericneurons.Cell, 79:1277–1285,1994.
[6] P. Blume-Jensen,L. Claesson-Welsh,A. Siegbahn,K.M Zsebo,B. Wester-mark, andC.I. Heldin. Activation of the humanc-kit productby theligandinduceddimerizationmediatescircularactin reorganizationandchemotaxis.EMBOJ., 10:4121–4128,1991.
[7] V. Castets,E. Dulos, J. Boissonade,andP. De Kepper. Experimentalevi-denceof a sustainedstandingturing-typenonequilibriumchemicalpat-tern. Phys.Rev. Lett., 64:2953–2956,1990.
[8] E. Crampin,E. Gaffney, andP.K. Maini. submitted.[9] N. Dakin. TheMacmillanbookof the marineaquarium. MacmillanPub-
lishingCompany, New York, 1992.[10] N. M. Le Douarin.TheNeural Crest. CUP, Cambridge,1982.[11] H.-H. Epperleinand J. Lofberg. The developmentof the larval pigment
patternsin triturus alpestrisand ambystomamexicanum. Adv. Anat.Embrol. Cell. Biol., 118:1–101,1990.
[12] C.A. Erickson. From the crestto the periphery: Control of pigmentcellmigrationandlineagesegregation.PigmentCell Res., 6:336–347,1993.
[13] B. Ermentrout.Stripesor spots?nonlineareffectsin bifurcationof reaction-diffusionequationson thesquare.Proc. Roy. Soc.Lond.A., 434:413–417,1991.
[14] A. Fraser-Brunner. A revision of the chaetodontfishesof the subfamilypomacanthinae. Proc.Zool.Soc., 36:543–596,1933.
[15] A. Fraser-Brunner. Patterndevelopmentin thechaetodontfishpomacanthusannularis (bloch), with a noteon the statusof euxiphipops. Copeia.,1:88–89,1951.
[16] H.W. Fricke. Juvenile-adultcolourpatternsandcoexistencein theterritorialcoral reef fish pomacanthusimperator. Marine Ecology, 1:133–141,1980.
[17] R. Fujii. Cytophysiologyof fish chromatophores.Int. Rev. Cytol., 143:191,1993.

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 79
[18] R.Fujii. ThePhysiologyof Fishes, chapterColorationandChromatophores,pages535–562. Marine ScienceSeries.CRC press,BocaRaton,AnnArbor, London,Tokyo, 1993.
[19] R. Fujii, H. Kasuwaka,, K. Miyaji, andN. Oshima.Mechanismof skincol-orationandits changesin theblue-greendamselfish.Zool.Sci., 6:477–486,1989.
[20] S.J.Galli, K.M. Zsebo,andE.N. Geissler. Thekit ligand,stemcell factor.AdvancesImmunol., 55:1–96,1993.
[21] E.N.Geissler, M.A. Ryan,andD.E.Housman.Thedominant-whitespotting(w) locusof themouseencodesthec-kit proto-oncogene.Cell, 55:185–192,1988.
[22] S. F. Gilbert. DevelopmentalBiology. SinauerAssociates,fifth edition,1997.
[23] R. Halaban,S. Ghosh,and S. Baird. bfgf is the putative naturalgrowthfactorfor humanmelanovytes. In Vitro, 23:47–52,1987.
[24] T. Horikawa, D.A. Norris, J.J. Yohn, T. Zekman,and J.B. Travers J.G.Morelli. Melanocyte mitogensinduceboth melanocyte chemokinesisandchemotaxis.J. Invest.Derm., 104:256–259,1995.
[25] O. Jensen,E. Mesekilded, P. Borckmans,and G. Dewel. Computer-simulationof turing structuresin the chlorite-iodide-malonicacid sys-tem. Phys.Scripta., 53:243–251,1996.
[26] H. Kasuwaka,N. Oshima,andR. Fujii. Mechanismof light reflectioninbluedamselfishmotile iridophores.Zool.Sci., 4:243–257,1987.
[27] S. Kelley. Pigmentation,squamationandthe osteologicaldevelopmentoflarval andjuvenilegrayangelfishpomacanthusarcuatus(pomacanthi-dae:Pisces).Bull. Mar. Sci., 56(3):826–848,1995.
[28] F. Kirschbaum. Untersuchungenuber dans farbmusterder zebrabarbebrachydanio rerio (cyprindae, teleostei). Roux’s. Arch. Dev. Biol.,177:129–152,1975.
[29] S.KondoandR. Asai. A reaction-diffusionwave on theskin of themarineangelfishpomacanthus. Nature, 376:675–768,1995.
[30] P.M. Kulesa,G.C. Cruywagen,S.R.Lubkin, P.K. Maini, J. Sneyd, M.W.J.Ferguson,andJ.D.Murray. On a modelmechanismfor thespatialpat-terningof teethprimordiain thealligator. J. Theor. Biol., 180:287–296,1996.
[31] T. Kunisada, H. Yoshida, H. Yamazaki, A. Miyamoto, H. Hemmi,E. Nishimura,L. D. Shultz,S. Nishikawa, andS. Hayashi. Transgeneexpressionof steelfactor in the basallayer of the epidermispromotessurvival, proliferation,differentiationandmigrationof melanocytepre-cursors.Development, 125:2915–2923,1998.
[32] I. LengyelandI. R. Epstein.Modelling of Turingstructuresin thechlorite-iodide-malonicacid-starchreactionsystem. Science, 251:650–652,1991.

80 From: MathematicalModelsfor Biological PatternFormation
[33] M.J. LyonsandL.G. Harrison. A classof reaction-diffusion mechanismswhichpreferentiallyselectstripedpatterns.Chem.Phys.Lett., 183:158–164,1991.
[34] M.J. LyonsandL.G. Harrison. Stripeselection:An intrinsic propertyofsomepattern-formingmodelswith nonlineardynamics. Dev. Dyn.,195:201–215,1992.
[35] M. McClure. Chapter3: Growth, ShapeChange, and the developmentofpigmentpatternsin fishesof the genusDanio (Teleostei: cyprindae).PhDthesis,CornellUniversity, 1998.
[36] H. Meinhardt. Models for positional signalling with applicationto thedorsoventral patterningof insectsand segregation into different celltypes.Development, supplement:169–180,1989.
[37] H. Meinhardt.Dynamicsof stripeformation.Nature, 376:722–723,1995.[38] H. Meinhardt.Thealgorithmicbeautyof seashells. Springer, Berlin, New
York, 2ndedition,1998.[39] J.G. Morelli, J.J. Yohn, B. Lyons, R.C. Murphy, and D.A. Norris.
Leukotrinesc and d aspotentmitogensfor culturedhumanneona-tal melanocytes.J. Invest.Dermatol., 93:719–722,1989.
[40] J.D. Murray. A patternformationmechanismandits applicationto mam-maliancoatmarkings.volume39 of Lecture Notesin Biomathematics,pages360–399.Springer, Berlin, Heidelberg, New York., 1979.
[41] J.D.Murray. A pre-patternformationmechanismfor animalcoatmarkings.J. Theor. Biol., 88:161–199,1981.
[42] J.D.Murray. How theleopardgot its spots.Sci.Am., 258:80–87,1988.[43] J.D. Murray. MathematicalBiology. Springer-Verlag,Berlin, Heidelberg,
New York, secondeditionedition,1993.[44] B. N. Nagorcka.Fromstripesto spots:Prepatternswhich canbeproduced
in theskin by a reaction-diffusionsystem.IMA. J. Math.Appl.Med.&Biol., 9:249–267,1992.
[45] T. Naitoh,A. Morioka,andY. Omura.Adaptationof a commonfreshwatergoby, yoshinobori,rhinogobiusbrunneustemmincket schlegel to vari-ousbackgroundsincludingthosecontainingdifferentsizesof blackandwhitecheckerboardsquares.Zool.Sci., 2:59,1985.
[46] D.R. Newth. On theneuralcrestof the lamprey embryo. J. Embryol.Exp.Morphol., 4:358–375,1956.
[47] L. OlssonandJ.Lofberg. Pigmentpatternformationin larval ambystomatidsalamanders:Ambystomatigrinum tigrinum. J. Morphol., 211:73–85,1992.
[48] Q. OuyangandH.L. Swinney. Transitionfrom auniformstateto hexagonalandstripedturingpatterns.Nature, 352:610–612,1991.
[49] K.J. Painter. Chemotaxisasa Mechanismfor Morphogenesis. PhDthesis,Universityof Oxford.,1997.
[50] K.J. Painter, H.G.Othmer, andP.K. Maini. Stripeformationin juvenilepo-

IMA Volumesin Maths.& Apps,121,59–82.Springer-verlag. 81
macanthusexplainedby a generalizedturing mechanismwith chemo-taxis. Submittedarticle,1999.
[51] D. M. Parichy. Pigmentpatternsof larval salamanders(ambystomatidae,salamandridae):Theroleof thelateralline sensorysystemandtheevo-lution of pattern-formingmechanisms.Dev. Biol., 175:265–282,1996.
[52] D. M. Parichy. When neuralcrestandplacosescollide: Interactionsbe-tweenmelanophoresandthelaterallinesthatgeneratestripesin thesala-manderambystomatigrinum tigrinum (ambystomatidae).Dev. Biol.,175:283–300,1996.
[53] G.H.Parker. AnimalColourChangesandTheirNeurohumours. CUP, Cam-bridge,1948.
[54] J.S.Rubin, A.M.L. Chan,D.P. Bottaro,W.H. Burgess,W.G. Taylor, A.CCech,D.W. Hirschfield,J.Wong,T. Miki, P.W. Finch,andS.T. Aaron-son.A broadspectrumhumanlungfibroblast-derivedmitogenis avari-antof hepatocytegrowth factor. Proc.Natl Acad.Sci.USA, 88:415–419,1991.
[55] M. Schliwa. Biology of the Integument2: Vertebrates, chapterPigmentCells, pages65–77. Springer-Verlag, Berlin Heidelberg New YorkTokyo, 1986.
[56] G. P. Du Shane.Theorigin of pigmentcellsin amphibia.Science, 80:620–621,1934.
[57] JonathanRichardShewchuk. Triangle: Engineeringa 2D Quality MeshGeneratorand DelaunayTriangulator. In Ming C. Lin and DineshManocha,editors, Applied ComputationalGeometry: Towards Geo-metric Engineering, volume 1148 of Lecture Notesin ComputerSci-ence, pages203–222.Springer-Verlag,May 1996.FromtheFirstACMWorkshoponApplied ComputationalGeometry.
[58] M. Sugimoto.Morphologicalcolourchangesin themedaka,oryziaslatipes,after prolongedbackgroundadaptation— i. changesin the popula-tion andmorphologyof the melanophores.Comp.Biochem.Physiol.,104A:513,1993.
[59] K. W. Tosney. A long distancecuefrom emerging dermisstimulatesneuralcrestmigration.Soc.Neurosci.Abs., 18:1284,1992.
[60] R. P. Tucker and C. A. Erickson. Pigmentpatternformationin tarichatorosa: The role of theextracellularmatrix in controllingpigmentcellmigrationanddifferentition.Dev. Biol., 118:268–285,1986.
[61] A.M. Turing. Thechemicalbasisfor morphogenesis.Phil. Trans.Roy. Soc.Lond.B., 237:37–72,1952.
[62] C. Varea,J.L. Aragon,andR. A. Barrio. Confinedturing patternsin grow-ing systems.Phys.Rev. E., 56:1250–1253,1997.
[63] B. Wehrle-HallerandJ.A. Weston. Solubleandcell-boundforms of steelfactoractivity play distinct rolesin melanovyte precursodispersalandsurvival on the lateral neuarlcrestmigration pathway. Development,

82 From: MathematicalModelsfor Biological PatternFormation
121:731–742,1995.[64] D.E. Williams, J.Eisenman,A. Baird, C. Ruach,K. VanNess,C.J.March,
L.S. Park, U. Martin., D.Y. Mochizuki, H.S. Boswell, G.S. Burgess,D. Cosman,and S.D. Lyman. Identificationof a ligand for the c-kitproto-oncogene.Cell, 63:167–174,1990.
[65] Y. Yada,K. Higuchi, andG. Imokawa. Effects on endothelinson signaltransductionandproliferation in humanmelanocytes. J. Biol. Chem.,266:18352–18357,1991.
[66] D.A. Young. A local activator-inhibitor modelof vertebrateskin patterns.Math.Biosci., 72:51–58,1984.