Embed Size (px)
Citation preview

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 1/8
BioSystems 36 (1995) 101-108
M odeling insect olfactory neuron signaling by a network
utilizing disinhibition
Evyatar Av-Ron*a9b, Jean-Pierre RosparsC
‘ B3E, INSERM I J263, I SARS, Facultk de Mdecine Saint-Antoine. 27 rue Chaligny. 7S571 Paris Cedex 12. Fr ance
bGroupe de Bioin formatique. URA 686, Ecole Normol e SupCri eure, 46 rue d’ lll m, 75230, Pari s Cedex 05, Fr ance
=L .uboratoire de Biomktr ie, lnstitut National de la Recherche Agronomique, 78026 Versailles Cedex, F rance
Received 24 February 1995; accepted 27 March 1995
Ah&met
A male moth locates a conspecific female by detecting her sexual-pheromone blend. This detection is carried out
in the antenna1 lobe, the first stage of olfactory information processing , wh ere local inhibitory neurons and projection
(relay) neurons interact. A ntennal-lobe neurons exhibit low-frequency (C 10 Hz) backgrou nd activity and bursting
(> 100 Hz ) activity in respons e to pherom one stimulation. We describe this behavior by a realistic biophysical neuron
mod el. The bursting behavior of the mode l is the result of both intrinsic cellular pro pertieS and n etwork interaction.A slowly activating and inactivating calcium channel provid es a depolarizing current for bursting and disinhibition
is show n to be. a feasible network mechanism for triggering this calcium chan nel. Small neural n etwork s utilizing
disinhibition are presented w ith local neurons intercalated between recept or and projection neurons. The tiring
behavio rs of projection neurons in respons e to stimulation by the pherom one blend or its compo nents are in accor-
dance with experimental results. This network architecture offers an alternative view of olfactory processing from the
classical architecture derived from vertebrate studies.
Keyw ords: Insect olfaction; Ion conductance neuron mo del; Netw ork disinhibition
1. Introduction
The first stage of insect olfaction processing
resides in the brain antenna1 lobes (ALs). Receptor
neurons (RNs ) of the antennae send afferent in-
puts into well-organized structures of the AL, call-
??Corresponding author, Tel.: +33 1 44738433;Fax: +33 I
43738462;E-mail: [email protected].
ed glomeruli, in which they make synapse with ALneurons. Two main types of AL neurons are found
in glomeruli, local neurons (LNs) with arboriza-
tions confined to the AL and projection neurons
(PNs) with axons that relay information to higher
brain centers in the protocerebrum (for reviews see
Christensen and Hildebrand 1987a, Rospars 1988,
Homberg et al. 1989, Masson and M ustaparta
1990, Boeckh et al. 199 0, Hildebrand et al. 1992 ).
As a case study we examined olfaction process-
0303-264 7/95609.50 0 1995 Elsevier Science Ireland Ltd. All rights reservedSSDI 0303-2647(95)01531-O

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 2/8
10 2 E. Av -Ron, .-P. Rospars Bi oSystems 6 f T99.5) 101-108
ing in a specific glomerular AL neuropil, the so-
called macroglomerular complex (MGC ) of male
moths (which is also present in other insects, e.g.
cockroaches and honey bees (Rospars 1988,
Masson and M ustaparta 1990, Boeckh et al. 1990,Hildebrand et al. 1992). This complex processes
sex-pheromone inputs coded by specialized RNs.
Sex pheromones released by female moths attract
their conspecific males. This system p lays a crucial
role for the survival a nd evolution of the species.
The sex pheromone blend is composed of a num-
ber of different compound s. For example, in the
moth M unducu sex& (Kaissling et al. 1989,
Tumlinson et al. 1989), twelve aldehydes have been
identified. Of these, two 16carbon compoun ds
have been found ‘major’ from quantitative and be-havioral points of view, a dienal (bombykal, Bal)
and a trienal mimicked by a 15carbon aldehyde
(Cl 5). In wind tunnel experiments, a blend of these
two components is sufficient to induce up wind
flight of males towards the odor source (Tumlin-
son et al., 1 989). This blend was also found to elicit
neuronal response similar to extract from the fe-
male sex-pheromone gland (see Fig. 12 in
(Christensen a nd Hildebrand, 1987b)). Similar
neuronal responses were found in other moth spe-
cies (Christensen et al. 1989 ).Intracellular recordings from PNs in the MG C
show various types of responses to the two major
components and sex-pheromone gland extract (see
Fig. 9 in (Christensen et al. 1989)). Some neurons
are found to be generalists, i.e. they exhibit either
excitatory response to Bal, Cl5 and blend or in-
hibitory response to all three. Other neu rons are
specialists. One type of specialist respond s with ex-
citation to Bal and blend, bu t exhibits no response
to C15, or vice-versa, respond s with excitation to
Cl5 and blend, but not to Bal. A second type ofspecialist exhibits excitation to one major compo-
nent and inhibition to the other (e.g. see Figs. la,
b). The response to blend is a mixture of the two
(Fig. lc).
The connections between neurons in the
glomeruli in general and the MGC in particular
can be summ arized as follows. The RN s send ex-
citatory input to LNs, which are intercalated be-
tween RNs and PNs. Most (or all) LNs are
GAB Aergic and probably inhibitory (Hoskins et
al. 1986 , W aldrop et al. 1987). The delay in PN ex-
citation suggests that there are no direct synapses
from RN s to PNs (Christensen et al. 1993).
Our goal is to study neuronal signaling
mechanisms in the MGC . This small network(there are 86 000 RNs and 30 PNs in the M GC of
M& UCU (Homberg et al. 1989)), with its well
defined biological function, is attractive for a theo-
retical study. In this article we examine disinhibi-
tion as a possible mec hanism for neural processing
and signal recognition. W e use a realistic biophysi-
cal model to describe the PN s of the network. In
this manner the biophysical features of the basic
neural comp onents can be related to the overall
computational properties of the network.
2. Neuron model
PNs respond to an odorant stimulation with a
burst followed by a period of inactivation (Fig.
la). We have chosen to describe four properties of
this response: duration and spiking frequency of
the burst, duration of the inactivation period and
spiking frequency of the background activity.
---
c-15. 50ng L
Fig. 1. Response of a neuron to odor stimulation. (a) Bal. (b)
Cl5 (c) Pheromone-gland wash. Early inhibition is indicated
by open arrow. Bars indicate stimuli. Scale markers: 50 mV,
625 ms. Taken w ith permission from Christensen and
Hildebrand (1987b).

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 3/8
E. Av -Ron , J.-P. Rospars/ B ioSystems 36 (1995) 101-108 103
The biophysical model (Av-Ro n 1994, see Ap
per&x) consists of two inward currents (sodium
INo and calcium Z cJ and three ou tward potassium
currents (delayed ZK, transient IA and calcium-
dependent ZK(,-~J) nd a leak current IL. An ioncurrent Zi is described by the product of three
MI NIS; the maxim al conductance gig the activation
and inactivation variables or functions, and the
driving force (V - Vi). The steady-state functions
are modeled as sigmoid curves, determined by two
parameters, the half maxim um voltage Vi,2 (in-
flection point), and the slope a of the curve at this
point. This model wa s developed in a stepwise
manne r. For a single action potential the currents
Z,, and ZK are required (Hodgkin and Huxley
1952, FitzHugh 1961, Av-Ron et al. 1991). Forbursting behavior the currents Ia and ZKccalwere
incorporated (Plant 1981, Rinzel and Lee 1987,
Av-Ron et al. 1993). Finally, to exhibit both low
0
-6 0
1 ----__ _ _ ‘_
0 50 0 1000 1500 2(
Time (msec)
Fig. 2. Low-frequency activity and bu rsting behavior of neuron
model. Depicted are membrane potential V (solid line), intra-
cellular calcium concentration C (dashed tine), calcium channel
activation X (solid line marked X) and transient potassium
channel inactivation B (solid line marked B). Burst initiated by
an input current Li, = 5 @cm2 from t = 300 to 320 ms.
Mod el parameters: C, = 1 pFlcm2, gNU= 120 mS/cm2,
&= I5 mW m2, zr. = 0.3 m Wm2, &ca, = 0.5 rnWcm2,
&. = I mS/cm2, & = 12.5 m!Ycm2, V,, = 55 mV, V,, = 12 4
mV, V,= -72 mV, V, = -50 mV, P)rn = -31 m V atrn)
0.065, Vc”), = -35 mV a@+= 0.055, K = 0.08, s = I, Wr,2 =
-70 mV, a$) = -0.1, r6 = IO ms, W)tj2 = -45 mV acx) = 2,
rx = 25 ms, v(l)ro) = -20 mV, a@ = 0.02, K, = 2, KP= 0.0002,
R = 0.006, Kd = 0.5.
and high firing frequencies the current IA was in-
troduced (Connor et al. 1977, Rose an d Hind-
marsh 1989, Av-Ron 1994).
A typical behavior of the model is shown in Fig.
2. W ith no stimulation, the model exhibits slowoscillations (6 Hz with the specific values chosen
for the parameters). For a short excitatory stimu-
lation, a burst respon se is displayed. The stimula-
tion activates (variable x) the current Zca which
remains active for the duration of the burst due to
the slow time constant rX (see curve x). During
the burst, calcium enters the cell, activating the
current ZKlcol (see (10)) and inactivating the cur-
rent I,, (see (7)); eventually bringing about the
quiescence (inactive) period. C alcium w as assum ed
to reside in a thin shell under the membrane. Forsimplicity a single process is used for calcium dif-
fusion, buffering and sequestration. As well the re-
versal potential for calcium is assum ed to be
constant. During quiescence, intracellular calcium
is removed (see curve C ). Upon returning to
resting levels, low-frequency oscillations com-
mence. This is mainly determ ined by the current
IA, which has a slow time constant rb for inactiva-
tion (see curve Z?).
In this paper, a single neuron model was used,
i.e. the values of the parameters that define the ki-netics of the different currents were kept constant.
Only two parameters were varied to achieve the
different behaviors, the maxim al potassium con-
ductance g’Aand the rate of calcium removal (R).
The ordinary differential equations were solved
numerically using the fourth-order Runge-K utta
method with a time step of 0.01 ms.
3. Results
3.1. Serial networkA basic network utilizing disinhibition consists
of four neurons in series (Fig. 3a). In the absence
of stimulation, we assum e the projection neuron
(PN) has an oscillatory behavior (ca. 20 Hz, see
right-hand side of Fig. 4a). Wh en connected to a
local neuron (LN2) the low frequency spontaneous
activity of LN2 is sufficient to inhibit the PN a nd
keep it below the bursting threshold. For the PN
to exhibit a bursting response to odor stimulation,
another local neuron (LNl) connected to LN2 is

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 4/8
10 4 E. Av-Ron. J.-P. Rospars/BioSyslems 36 (1995) 101-108
a
b
c
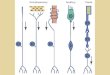
Fig. 3. Neural networks of receptor (RN), local (LN) and pro-
jection (PN) neurons with excitatory (+) and inhibitory (-)
connections. (a) Generalist network utilising disinhibition. PN
exhibits an excitatory response (see Fig. 4a) w hen R Ns a~
stimulated with odors A and/or B. (b) Specialist network
uti!ising disinhibition. PN exhibits an excitatory resp onse (see
Fig. 4a) to stimulation with odor B and an inhibitory esponse
(see Fig. 4a) to stimulation with odor A. Stimulation with both
odors A and B elicits a ‘mixed’ response (see Fig. 4c). (c) Sp e-
cialist network b ased on the ‘classical’ architecture of the
vertebrate olfactory bulb with lateral inhibition. Responses to
odor A or B is as (b). For odors A and B, a short excitatory
response is elicited.
required. W hen stimulated by the excitatory RN s,
LNl inhibits LN2, which leads to the disinhibition
of the PN, allowing it to fire a burst.
Fig. 4a shows the behavior of a PN resulting
from this serial network. During the first second
the neuron receives inhibition (Z,im = -0.5
@U cm’, mimick ing tonic inhibition by LN2) and
exhibits slow spontaneous oscillations (3 Hz for
the chosen param eters). At time 0.5 s the in-
hibitory current is suppressed (mimicking inhibi-tion of LN2 by LNl during odor stimulation) and
the neuron exhibits its bursting response. The
duration of the burst depends on the intracellular
calcium concentration which activates the current
IKfcoJ and inactivates the current Zca The burst is
followed by a period of inactivation during which
intracellular calcium is removed. At time 2.2 s in-
tracellular calcium concentration has reached its
base level and the model begins to oscillate at 20
Hz. Wh en the inhibitory current is re-established
the neuron returns to its spontaneous 3 Hzoscillations.
This simulation describes the experimentally
observed behavior shown in Fig. la, excluding the
20-Hz oscillation after time 2.2 s shown in Fig. 4a.
In the experimental recordings the duration of the
burst coincides with the duration of stimulation
(0.5 s). However, for the model, a shorter period
(120 ms) of disinhibition is sticient to obtain a
bursting response. T his can be compared with
observed experimental results where short odor
stimulations (about 50 ms) triggered b urstingresponses (see Fig. 1 in Christensen and
Hildebrand 1988).
3.2. Para l le l network
A network based on inhibition and disinhibition
is shown in Figure 3b. The PN has the sam e pro-
perties as above. For stimulation with odorant B
(e.g. Bal) it behaves as shown in Fig. 4a. For stim-
ulation with odorant A (e.g. C15) it is strongly in-
hibited by LN2 for the duration of the odorantstimulation and responds as shown in Fig. 4b.
Wh en the stimulation ceases the PN returns to its
slow background activity (3 Hz). For stimulation
with a mixture of odorants A and B the PN
responds as shown in Fig. 4c. An initial inhibition
is followed by a burst. Both LN l and LN2 are ex-
cited simultaneously by their respective RN s.
Therefore, LN2 inhibits the PN. After a synaptic
delay LN2 receives inhibition from LNl. This
input inhibits LN2 and consequently releases the

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 5/8
E. Av-Ron. J.-P. Rospars/ BioSystems 36 (1995) 101-108 105
PN which fires a burst of activity. Following the
burst the PN is inactive and eventually returns to
its low-frequency spontaneous background ac-
tivity.
. . . . . . . .
--.- /
1. . I,
- . _ _ _ _
_ _ _ _ _ _ - -
500 1000 1500 2000
60 -
-60 -
r
I”0 ,500 1000 1500 2000
- 0.5
n
7
;
4. Discussion
In this paper we have studied disinhibition, first-
ly as a biophysical mechan ism for explaining intra-
cellular recordings (individual neuron properties)
and secondly, as an information-processing mech-
anism (local network properties).
4.1. Disinhibition of neuron membrane potential
Cell culture work suggests that the channel
types used in the biophysical model are present in
receptor (Zufall et al. 1991, Stengl 1994) and A L
neurons (Hayashi and Hildebrand 1990,
Hildebrand et al. 1992, Mercer et al. 1995, Klop
penburg and Hildebrand 1995). This model ex-
hibits intrinsic low-frequency background activity(6 Hz in Fig. 2 and 20 Hz in Fig. 4a on the right,
due to different values of g” a nd R ) and shows a
burst in response to a short excitatory stimulation.
This behavior results from the existence of two
thresholds, a threshold for action potentials (acti-
vation of the sodium current) and a threshold for
burst discharge (activation of the calcium current).
Applying inhibition reduces the background ac-
tivity further (Fig. 4a, from 20 Hz on the right to
3 Hz on the left). When inhibition is stopped
(disinhibition) a bursting response occurs, calledpostinhibitory rebound (Selverston and Mo ulins
1985). The mecha nism of this response is as
follows. Inhibition hyperpolarixes the mem brane
potential which in turn increases the threshold for
tiring. A decrease of the firing rate follows which
brings about the lower intracellular calcium con-
Fig. 4. Beh avior of projection-neuron model. Depicted are
membrane potential V (solid line), intracellular calcium con -centration (dashed line) and stimulating current (dotted line).
(a) Low frequency background activity resulting from constant
inhibition (&,, = -0.5 nNcm*) until r = 0.5 s. Respo nse to
disinhibition (ISrim 0 for t>0.5 s) includes a burst followed
by a period of inactivation. Without inhibition (I,, = 0) the
model exhibits 20-Hz background activity (I > 2.2 s). (b) Cons-
tant inhibition IStim -0.5 @cm * for the entire duration (0 to
2 s), except inhibition IS,i,,, -5 fiA/cm* from r = 0.5 to I s. (c)
Constant inhibition Isrh = -0.5 pA/cm* for the entire duration
except inhibition I,,h = -5 @/cm* from 0.50 to 0.52 s and
disinhibition (Istim 0) from t = 0.52 to 1 s.Model parameters:
as Fig. 2 except fA = IO mS/crn*, R = 0.0025.

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 6/8
106 E. Av-Ron. .-P . Rospars/Bi oSysrems 6 (1995) 101-108
centration C. W hen inhibition is removed the
mem brane depolarizes rapidly. If C would also
increase rapidly the system would reach a stable
oscillation as seen on the right-hand side of Fig.
4a. However this is not the case, the slow increasein C results in a relatively weak current IK(c+
This allows the firing frequency and consequently
the depolarizing calcium current to increase, in-
stigating a positive feedback which results in the
burst discharge.
Presented here is a simple biophysical model
which can describe the qualitative behavior
observed in PNs. The next step would be to choose
parameters based on patch clamp experiments. As
well, the sensitivity of the model to parameter
changes should be studied. This may be examinedin relation to both neuronal development (Lockery
and Spitzer 1992) and neuromodulation (Mercer et
al. 1995, Kloppenburg and Hildebrand 1995).
4.2. D & i n h i b i t i o n i n l o c al - c i r cu i t p r o cessi n g
The local circuit network is derived from experi-
mental evidence.
(a) The latency in response of PNs in comparison
with LNs suggests that PNs do not receive direct
excitatory inputs from receptor cells (Christensen
and Hildebrand 1987, Christensen et al. 1993).(b) In some PNs (Fig. lc) the burst response
is preceded by a short hyperpolarization
(Christensen and Hildebrand 1988). (c) M ost (or
all) of the LNs are probably inhibitory in-
temeurons (Hildebrand et al. 1992). (d) Correla-
tion between activities in LN and PN pairs was
found both in the MG C an d regular glomeruli.
The depolarization of the LN brought about the
inactivation of the PN. Conversely, hyperpolariz-
ing the LN caused an increase in activity of the
PN. This shows that at least some PN excitation is
the result of disinhibition (Christensen et al. 1993).
The disinhibition model proposed offers an alter-
native to the ‘classical’ model (Fig. 3c) of the
vertebrate olfactory bulb in term of information
processing. However, d isinhibition may also apply
to the vertebrate olfactory bulb because serial
synapses between dendrites of presumed inhibitory
LNs within the glomeruli have been documented
(Pinching and Pow ell 1 971; W hite 1972). These ar-
rangemen ts are in addition to the direct connec-
tions from RN terminals onto LN and PN
dendrites, an d the reciprocal contacts between
these dendrites (cf. Shepherd and Greer 1990).
However, it is not evident w hether the disinhibi-
tion model is capable of coding the quantitativefeatures of the olfactory message, e .g. concentra-
tions of A and B for specialist P Ns (parallel net-
work), ratio of concentrations A:B for generalist
PNs (serial network). More generally, the differ-
ences in terms of information processing between
the ‘classical model’ (e.g. L&rsky and Rospars
1993, Rospa rs and Fort 1994, Rospars et al., 1994)
and the model of disinhibition remain to be further
investigated.
The mecha nisms of disinhibition observed in the
insect glomerulus and in the vertebrate reticularthalamic nucleus seem analogous. I&inhibition of
thalamocortical neurons reduces the inactivation
of the low-threshold calcium current bringing
about a rebound calcium plateau which causes a
burst of action potentials (Jahnsen and Llinas
1984, Steriade, et al. 199 3). This m echanism is
similar to that of Fig. 4a, where the inactivation
component is the current IKIca,.
In conclusion, disinhibition provides an altema-
tive to lateral inhibition’as a functional role for in-
hibitory LNs in a network. One possibleadvantage of disinhibition over excitation is that
the PNs spontaneously return to the resting (inac-
tive) state, whereas excitatory mecha nisms may
lead to propagating waves of activity (W ilson and
Bower 1989).
Acknowledgements
E. Av-Ron was supported by program ‘Poste
Vert’, INSERM , France.
AppediX
C ,d V l d t = I , ,- I N ,- I I C ,- Z I K - I A - I K ( C o l - I L (1 )
dW l d t = [ W , ( I ’ ) - W J/ r ,( V ) , (2)
dX l d t = [X ,( J’ ) - X J/ r ,, (3)
dB l d t = [B ,( V ) - B ] l q , (4 )
d Cl d t = K & - I & - R . C (5)

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 7/8
E. Av-Ron, J.-P. Rospars / BioSystems 36 (1995) 101-108 107
P, (v) = (1 + exp[-2dp)(V - v@)1,2)])-1for = [ W,m,X, A, B]
r,(v) = (Xexp[a(W)V -V(W),,2)]+ xkxp[-dw)( V - P)&])-l
(12)
(13)
References
Av-Ron, E., Pamas, H. and Segel, L.A. 1991 , A minimal bio-
physical model for an excitable and oscillatory neuron.
Biol. Cybem. 65,487-500.
Av-Ron, E., Pamas, H. and Segel, L.A. 1993 , A basic
biophyscial model for bursting neurons. Biol. Cybern. 6 9,
87-94.
Av-Ron, E, 1994 , The role of a transient potassium current ina bursting neuron model. J. Math. Biol. 33, 71-87 .
Boeckh, J., Distler, P., Ernst, K.D., H&l, M. and Malun, D.,
1990 , Olfactory bulb and antenna1 lobe, in: Chemosensory
information processing, D . Schild (ed.), (Springer Verlag,
Berlin) pp. 201-227.
Christensen, T.A . and Hildebrand J.G. 1987 a, Functions,
organization, and physiology of the olfactory pathw ays in
the Lepidoteran brain, in: Arthropod Brain: its evolution,
development, structure and functions, A.P. Gupta (ed.),
(John Wiley and Sons) pp. 457-484.
Christensen, T.A. and Hildebrand J.G. 1987 b, Male-specific,
sex pheromone-selective projection neurons in the antenna1
lobes of the moth Man&a sexta. J. Comp. Physiol. A 160,
553-569.
Christensen, T.A . and Hildebrand J.G. 1988 , Frequency coding
by central olfactory neurons in the sphinx m oth Manduca
sexta. Chemical Senses 13, 123-130.
Christensen, T.A., Mustap arta, H. and Hildebrand, J.G. 1989 ,
Discrimination of sex pheromone blends in the olfactory
system of the moth. C hemical Senses 14 , 463-4 77.
Christensen, T.A., Hildebrand J.G., Tumhnson J,H. and
Doolittle, R.E . 1989 , Sex pheromone blend of Manduca
sexta: Responses of central olfactory intemeurons to anten-
nal stimulation in male moths. Arch. Insect Biochem.
Physiol. IO, 281-291.
Christensen, T.A., Waldrop, B.R., Harrow, I.D. and
Hildebrand, J.G. 1 993, Local interneurons and information
processing in the olfactory glomeruli in the moth Manduca
sexta. J. Comp. Physiol. A 173, 385-399.
Connor, J.A., Walter, D. and McKnown, R. 1977, Neural re-
petitive Bring - modifications of the Hodgkin-Huxley axon
suggested by experimental results from crustacean axons.
Biophys. J. 18, 81-102.
FitxHugh, R. 1 961, Impulses and physiological states in theo-
retical models of nerve membrane. Biophys. J. I, 445-466.
Hayashi, J.H. and Hildebrand, J.G. 1990 , Insect olfactory
neurons in vitro: Morphological and physiological
characterization of cells from the developing antenna1 lobes
of Manduca sexta. J. Neurosci. IO, 848-859.
Hildebrand, J.G., Christensen, T.A., Arbas, E.A., Hayashi,
J.H., Homberg, U., Kanxaki, R. and Stengl, M. 1992,
Olfaction in Manduca sexta: Cellular mechanisms of
responses to sex pheromone, in: Neurotox 199 1 Molecular
basis of drug and pesticide action, 1.R. Duce (ed.), (ElsevierApplied Science, London and New York) pp. 323-338.
Hodgkin, A.L. and Huxley, A.F. 1952 , A quantitative descrip-
tion of membrane current and its application to conduction
and excitation in nerve. J. Physiol. 117, 500-5 44.
Homberg, U., Christensen, T .A. and Hildebrand, J.G . 198 9,
Structure and function of the deutocerebrum in insects.
Ann. Rev. Entomol. 34, 477-501.
Hoskins, S.G., Homb urg, U., Kingan, T.G., Christensen, T.A.
and Hildebrand, J .G. 198 6, Immunocytochemistry of
GABA in the antenna1 lobes of the sphinx moth Manduca
sexta. Cell Tissue Res. 244, 243-252.
Jahnsen, H . and Llinh, R. 19 84, Electrophysiological proper-
ties of guinea-pig thalamic neurones: An in vitro study. J.Physiol. 349, 205-226.
Kaissling, K.-E., Hildebrand, J.G. and Tumlinson, J.H. 1989,
Pheromone receptor cells in the male moth Manduca sexta.
Arch. Insect Biochem. Physiol. IO, 273-2 79.
Kloppenburg P. and Hildebrand, J.G. 1995 , Neurom odulation
by 5-hydroxytryptamine in the antenna1 lobe of the sphinx
moth Manduca sexta. J. Exp. Bio. 198 , 603-611.
Ldnsky, P. and R ospars, J.P. 1993 , Coding of odor intensity.
BioSystems 3 1, 15-38.
Lockery, S.R. and Spitzer, N.C. 1992 , Reconstruction of action
potential development from whole-ceil currents of differen-
tiating spinal neurons. J. Neurosci. 12, 2268 -2287 .
Masson, C . and Mustaparta, H. 1990 , Chemical information
processing in the olfactory system of insects. Physiol. Rev.
70, 199-245.
Mercer, A.R., Hayashi, J.H. and Hildebrand, J.G. 1995,
Modu latory effects of 5-hydroxytryptamine on voltage-
activated currents in cultured antenna1 lobe neurones of the
sphinx moth Manduca sexta. J. Exp. Bio. 19 8, 613-627.
Pinching, A.L. and Powell, T.P.S. 1971 , The neuropil of the
glomeruli of the olfactory bulb. J. Cell Sci. 9, 347-377.
Plant, R.E . 1981 , Bifurcation and resonance in a model for
bursting nerve cells. J. Math. Biol. 11,15-32.
Rinzel, J. and Lee, Y.S. 1987 , Dissection of a model for neuro-
nal parabolic bursting. J . Math. B iol. 25, 653-6 75.

7/30/2019 Modeling Insect Olfactory Neuron Signaling by a Network Utilizing Disinhibition
http://slidepdf.com/reader/full/modeling-insect-olfactory-neuron-signaling-by-a-network-utilizing-disinhibition 8/8
10 8 E. Av- Ron, J.-P. Rospars / Bi oSystems 36 (199s) 101-108
Rospars, J.P. 1988 , Structure and development of the insect
antennodeutocerebral system. Int. J. Insect Morphol. Em-
bryol. 17, 243-294.
Rospars, J.-P. and Hildebrand, J.G. 1992 , Anatomical identili-
cation of glomeruli in the antenna1 lobes of the male sphinx
moth Mumfncu sexra. Cell Tissue R es. 270, 205-227.
Rospars, J.P., Lrlnsky, P., Vaillant, J., Duchamp-V iret, P. and
Ducham p, A. 19 94, Spontaneous activity of first- and
second-order neurons in the frog olfactory system. Brain
Res. 662, 3 l-44.
Rospars, J.P. and Fort, J.C . 1994 , Coding of odor quality: role
of convergence and inhibition. Network 5, 121-1 45.
Rose, R.M . and Hindm arsh, J.L. 198 9, The assembly of ionic
currents in a thalamic neuron I. The threedimensional
model. Proc. R. Sot. Lond. B 237, 267-288.
Selverston, AI. and Moulins, M. 19 85, Oscillatory neural net-
works. Ann. R ev. Physiol. 47, 29-48.
Shepherd, G.M . and Greer, 1990 , Olfactory bulb, in: The
Synaptic Organization of the Brain G.M. S hepherd (rd.),(Oxford Univ. Press, Oxford) pp. 133-169.
Steriade, M., McCormick, D.A. and Sejnowski, T.J. 1993,
Stengl, M. 199 4, Inositol-triphosphatedependent calcium cur-
rents precede cation currents in insect olfactory receptor
neurons i n vi t ro . J. Comp. Physiol. 174, 187- 194.
Tumlinson, J.H., Brennan, M.M., Doolittle, R.E., Mitchell,
E.R., Brabham. A., Maxomenos, B.E., Baumhover, A.H.
and Jackson, D.M . 1989 , Identification of a pheromone
blend attractive to Mun ducu sexta (L.) males in a wind tun-
nel. Arch. Insect B&hem . Physiol. 10, 255-2 71.
Waldrop, B ., Christensen, T.A . and Hildebrand, J.G . 19 87,
GABA -mediated synaptic inhibition of projection neurons
in the antennal lobes of the sphinx moth, Mon ducu sexfu. J.
Comp. Physiol. A 161, 23-32.
Wilson, M .A. and Bower, J.M. 1989 , The simulation of large-
scale neural networks, in: Methods in neuronal modeling:
from synapses to networks, C. Koch and I. Segev (eds.)
(MI T Press, Cambridge, Massachusetts) pp. 291-3 33.
White, E.L. 1972 , Synaptic organ ization in the olfactory
glomerulus of the mouse. B rain Res. 37, 69-80.
Zufall, F., Stengl, M., Franke, C., Hildebrand, J.G. and Hatt,H. 1 991, Ionic currents of cultured olfactory receptor neu-
rons from antennae of male Munduca sextu. J. Neurosci. 11,
Thalamo cortical oscillations in the sleeping and aroused
brain. Science 262, 679-6 85.
956-965.