Embed Size (px)
DESCRIPTION
Stomatal conductance in fruit species depend upon conditions such as solar irradiance, leaf temperature, leaf-to-air vapour pressure deficit (VPD), soil and plant water status and mineral nutrition. Among apple cultivars, stomatal regulation has been related to tree vigour. Selected standardized apple saplings of cv. Gala Galaxy on M9 rootstocks treated with Progerbalin (GA 4 + 7 with 6-Benzyladenine) which is known to encourage shape improvement and advancing leaf apparatus emission and development, were subjected to a series of plant stress based assessment compared to untreated controlled plants. Using a model based on a complex parameterized P-M equation re-written to already include the gsw as the function of R and vapour pressure deficit (VPD), we have a produced the dynamics of stomata conductance of these saplings, 6 treated and 2 untreated controls. Transpiration data were calculated from sap flow measurements. Stomata response to both important radiation and VPD was analysed showing differences in their stomata behaviour. Stomatal sensitivity to VPD varies both within and between species. This ecophysiological model of canopy transpiration and stomata conductance could be a valuable tool for studying complex processes, i.e. interrelated physiological functions, particularly for comparing cultivars or various treatments.
Citation preview
IntroductionStomata have a major regulatory role in gas exchange in leaves and the efficiency of this regulation can impact yields of agricultural crops. Beside various environmental varia-bles like solar irradiance, leaf temperature, leaf-to-air vapour pressure deficit (VPD), soil and plant water status, stomatal behaviour is influenced by hormones and other natu-rally occuring compounds. Abscisic acid (ABA) is intimately involved in the regulation of stomatal behaviour; other groups of hormones, such as the cytokinins, may also be involved in the control of stomata. Among apple cultivars, stomatal regulation has been related to tree vigour. Gibber-elins/cytokinins are known to encourage shape improvement and advancing leaf appa-ratus emission and development. Moreover, the effects of certain environmental factors on stomatal behaviour may be mediated by hormones. However, most of reports on the effects of gibberelins and cytokinins on stomatal behaviour either refer to grasses or use excised leaves and therefore effects are not clear and not supported by most of investigators.
Material and MethodsPlant MaterialTwelve standardized apple saplings bench grafted of cv. Gala Galaxy on M9 rootstocks. Plants were selected to have the same biometric features as the treated plants, namely rootstock diameter 10 cm below the grafting point and scion diameter 10 cm above the grafing point, branch number, length, crotch angle and branch diameter 5 cm from the base, number of lateral shoots, leaf area, etc. Root systems were compared as well for uniformity and size. These trees were not producing fruits. From these 12 plants planted in spring, 6 were then treated with Progerbalin LG (GA4-7 1,8% + BA 1,8%) in order to stimulate branching. The soil in which saplings were planted was of good quality, up to 60 cm deep and in average contained: humus 2.36 %, (moderate) N 0.13 % (moderate), P2O5 10.69 mg/100g soil (low), K2O 43 mg (high), Ca 101.73 mg (moderate), Mg 47.14 mg (moderate); pH value in water was 6.8 whereas in KCl 5.8 (slightly acid). Ploughing was made at 40 cm depth, organic and mineral fertilizer were distributed in advance: organic 5 kg/m2 and mineral NPK 15 : 15 : 15 100g/m2. The plot was tilled 5 times, plants were drip irrigated and have received 3 treatments with fungicides and insecticides.
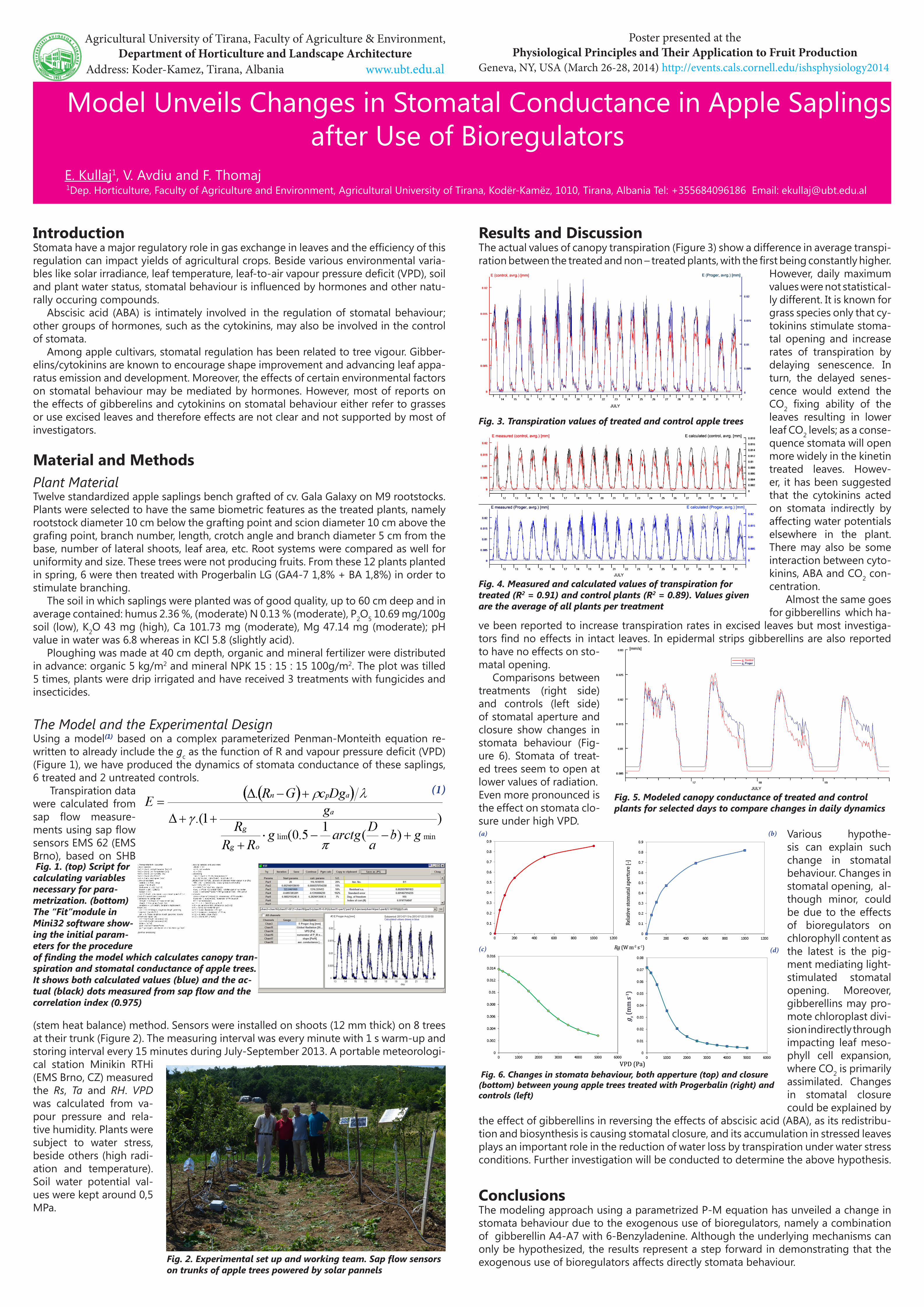
The Model and the Experimental DesignUsing a model(1) based on a complex parameterized Penman-Monteith equation re-written to already include the gc as the function of R and vapour pressure deficit (VPD) (Figure 1), we have produced the dynamics of stomata conductance of these saplings, 6 treated and 2 untreated controls. Transpiration data were calculated from sap flow measure-ments using sap flow sensors EMS 62 (EMS Brno), based on SHB
(stem heat balance) method. Sensors were installed on shoots (12 mm thick) on 8 trees at their trunk (Figure 2). The measuring interval was every minute with 1 s warm-up and storing interval every 15 minutes during July-September 2013. A portable meteorologi-cal station Minikin RTHi (EMS Brno, CZ) measured the Rs, Ta and RH. VPD was calculated from va-pour pressure and rela-tive humidity. Plants were subject to water stress, beside others (high radi-ation and temperature). Soil water potential val-ues were kept around 0,5 MPa.
Agricultural University of Tirana, Faculty of Agriculture & Environment, Department of Horticulture and Landscape Architecture
Address: Koder-Kamez, Tirana, Albania www.ubt.edu.al
Model Unveils Changes in Stomatal Conductance in Apple Saplings after Use of Bioregulators
E. Kullaj1, V. Avdiu and F. Thomaj 1Dep. Horticulture, Faculty of Agriculture and Environment, Agricultural University of Tirana, Kodër-Kamëz, 1010, Tirana, Albania Tel: +355684096186 Email: [email protected]
Results and DiscussionThe actual values of canopy transpiration (Figure 3) show a difference in average transpi-ration between the treated and non – treated plants, with the first being constantly higher.
However, daily maximum values were not statistical-ly different. It is known for grass species only that cy-tokinins stimulate stoma-tal opening and increase rates of transpiration by delaying senescence. In turn, the delayed senes-cence would extend the CO2 fixing ability of the leaves resulting in lower leaf CO2 levels; as a conse-quence stomata will open more widely in the kinetin treated leaves. Howev-er, it has been suggested that the cytokinins acted on stomata indirectly by affecting water potentials elsewhere in the plant. There may also be some interaction between cyto-kinins, ABA and CO2 con-centration. Almost the same goes for gibberellins which ha-
ve been reported to increase transpiration rates in excised leaves but most investiga-tors find no effects in intact leaves. In epidermal strips gibberellins are also reported to have no effects on sto-matal opening. Comparisons between treatments (right side) and controls (left side) of stomatal aperture and closure show changes in stomata behaviour (Fig-ure 6). Stomata of treat-ed trees seem to open at lower values of radiation. Even more pronounced is the effect on stomata clo-sure under high VPD.
Various hypothe-sis can explain such change in stomatal behaviour. Changes in stomatal opening, al-though minor, could be due to the effects of bioregulators on chlorophyll content as the latest is the pig-ment mediating light-stimulated stomatal opening. Moreover, gibberellins may pro-mote chloroplast divi-sion indirectly through impacting leaf meso-phyll cell expansion, where CO2 is primarily assimilated. Changes in stomatal closure could be explained by
the effect of gibberellins in reversing the effects of abscisic acid (ABA), as its redistribu-tion and biosynthesis is causing stomatal closure, and its accumulation in stressed leaves plays an important role in the reduction of water loss by transpiration under water stress conditions. Further investigation will be conducted to determine the above hypothesis.
ConclusionsThe modeling approach using a parametrized P-M equation has unveiled a change in stomata behaviour due to the exogenous use of bioregulators, namely a combination of gibberellin A4-A7 with 6-Benzyladenine. Although the underlying mechanisms can only be hypothesized, the results represent a step forward in demonstrating that the exogenous use of bioregulators affects directly stomata behaviour.
Poster presented at the Physiological Principles and Their Application to Fruit Production
Geneva, NY, USA (March 26-28, 2014) http://events.cals.cornell.edu/ishsphysiology2014
Fig. 1. (top) Script for calculating variables necessary for para-metrization. (bottom) The “Fit”module in Mini32 software show-ing the initial param-eters for the procedure of finding the model which calculates canopy tran-spiration and stomatal conductance of apple trees. It shows both calculated values (blue) and the ac-tual (black) dots measured from sap flow and the correlation index (0.975)
Fig. 6. Changes in stomata behaviour, both apperture (top) and closure (bottom) between young apple trees treated with Progerbalin (right) and controls (left)
Fig. 2. Experimental set up and working team. Sap flow sensors on trunks of apple trees powered by solar pannels
(1)
Fig. 3. Transpiration values of treated and control apple trees
Fig. 4. Measured and calculated values of transpiration for treated (R2 = 0.91) and control plants (R2 = 0.89). Values given are the average of all plants per treatment
Fig. 5. Modeled canopy conductance of treated and control plants for selected days to compare changes in daily dynamics
(a) (b)
(c) (d)

![Stomatal Biology of CAM Plants1[CC-BY] - Plant … on Stomatal Biology Stomatal Biology of CAM Plants1[CC-BY] Jamie Males* and Howard Griffiths Department of Plant Sciences, University](https://img.pdfslide.us/doc/110x75/5ae808d37f8b9a08778f24b1/stomatal-biology-of-cam-plants1cc-by-plant-on-stomatal-biology-stomatal.jpg)




![Stomatal Defense a Decade Later1[OPEN] - Plant Physiology · Update on Stomatal Defense Stomatal Defense a Decade Later1[OPEN] Maeli Melotto*, Li Zhang, Paula R. Oblessuc, and Sheng](https://img.pdfslide.us/doc/110x75/5eddc0a3ad6a402d6668efaa/stomatal-defense-a-decade-later1open-plant-update-on-stomatal-defense-stomatal.jpg)

![Evolution of the Stomatal Regulation of Plant Water ...Update on Stomatal Evolution Evolution of the Stomatal Regulation of Plant Water Content[OPEN] Timothy J. Brodribb* and Scott](https://img.pdfslide.us/doc/110x75/5e87e202c27a1d71d24f112b/evolution-of-the-stomatal-regulation-of-plant-water-update-on-stomatal-evolution.jpg)




















