Embed Size (px)
Citation preview

YEASTBOOK
CELL STRUCTURE & TRAFFICKING
Mitochondrial Protein Synthesis, Import,and AssemblyThomas D. Fox1
Department of Molecular Biology and Genetics, Cornell University, Ithaca, New York 14853
ABSTRACT The mitochondrion is arguably the most complex organelle in the budding yeast cell cytoplasm. It is essential for viability aswell as respiratory growth. Its innermost aqueous compartment, the matrix, is bounded by the highly structured inner membrane,which in turn is bounded by the intermembrane space and the outer membrane. Approximately 1000 proteins are present in theseorganelles, of which eight major constituents are coded and synthesized in the matrix. The import of mitochondrial proteinssynthesized in the cytoplasm, and their direction to the correct soluble compartments, correct membranes, and correct membranesurfaces/topologies, involves multiple pathways and macromolecular machines. The targeting of some, but not all, cytoplasmicallysynthesized mitochondrial proteins begins with translation of messenger RNAs localized to the organelle. Most proteins then passthrough the translocase of the outer membrane to the intermembrane space, where divergent pathways sort them to the outermembrane, inner membrane, and matrix or trap them in the intermembrane space. Roughly 25% of mitochondrial proteins participatein maintenance or expression of the organellar genome at the inner surface of the inner membrane, providing 7 membrane proteinswhose synthesis nucleates the assembly of three respiratory complexes.
TABLE OF CONTENTS
Abstract 1203
Introduction 1204
Cytoplasmic Synthesis of Mitochondrial Proteins 1205Localization of some cytoplasmic messenger RNAs to mitochondria promotes import of the proteins thatthey encode 1205
Complex mechanisms for mRNA localization 1207
Tethering of mRNAs to mitochondria via nascent polypeptide chains 1208
Translocation and Membrane Insertion of Cytoplasmically Synthesized Mitochondrial Proteins 1209Insertion of proteins into the outer membrane 1210
Import and insertion of b-barrel proteins: 1210Insertion of other integral proteins into the outer membrane: 1210
Import of proteins into the IMS 1211Covalent attachment of heme: 1211Oxidation of paired cysteine residues to form disulfide bonds: 1212
Import of proteins into the inner membrane 1213Continued
Copyright © 2012 by the Genetics Society of Americadoi: 10.1534/genetics.112.141267Manuscript received April 16, 2012; accepted for publication June 11, 20121Address for correspondence: Department of Molecular Biology and Genetics, Biotechnology Bldg., Cornell University, Ithaca, NY 14853-2703. E-mail: [email protected]
Genetics, Vol. 192, 1203–1234 December 2012 1203

CONTENTS, continued
Insertion of metabolite carriers and other multispanning inner membrane proteins by the TIM22 insertase/translocasecomplex: 1213
Insertion of inner membrane spanning proteins with cleavable presequences by the TIM23 insertase/translocasecomplex: 1214
Insertion of multispanning inner membrane proteins with presequences: 1215
Import of presequence-containing proteins to the matrix 1216
Spatial distributions and regulation of import complexes 1217
Assembly of Complexes Containing Mitochondrially Synthesized Proteins 1218Mitochondrial protein synthesis is membrane bound 1218
Channeling of mRNAs to the inner membrane 1219
Localization of protein synthesis by mRNA-specific translational activators 1219
Assembly of cytochrome c oxidase 1220
Assembly of the bc1 complex 1221
Assembly of the ATP synthase 1222
Perspective 1223
TO think about how mitochondrial proteins are synthe-sized, imported, and assembled, it is useful to have a clear
picture of the organellar structures that they, along withmembrane lipids, compose and the functions that they carryout. As almost every schoolchild learns, mitochondria carryout oxidative phosphorylation, the controlled burning of nu-trients coupled to ATP synthesis. Since Saccharomyces cere-visiae prefers to ferment sugars, respiration is a dispensablefunction and nonrespiring mutants are viable [although theycannot undergo meiosis (Jambhekar and Amon 2008)].However, mitochondria themselves are not dispensable. Asubstantial fraction of intermediary metabolism occurs inmitochondria (Strathern et al. 1982), and at least one ofthese pathways, iron–sulfur cluster assembly, is essentialfor growth (Kispal et al. 2005). Thus, any mutation thatprevents the biogenesis of mitochondria by, for example,preventing the import of protein constituents from the cyto-plasm, is lethal (Baker and Schatz 1991).
The mitochondria of S. cerevisiae are tubular structures atthe cell cortex. While the number of distinct compartmentscan range from 1 to�50 depending upon conditions (Stevens1981; Pon and Schatz 1991), continual fusion and fissionevents among them effectively form a single dynamic net-work (Nunnari et al. 1997). The outer membrane surroundsthe tubules. The inner membrane has a boundary domainclosely juxtaposed beneath the outer membrane and cristaedomains that project internally from the boundary into thematrix (Figure 1A). The matrix is the aqueous compartmentsurrounded by the inner membrane. The aqueous intermem-brane space lies between the membranes and is continuouswith the space within cristae.
Inner membrane cristae are often depicted as bafflesemanating from the boundary domain. However, electrontomography of mitochondria from several species, includingyeast, shows that cristae actually emanate from the bound-
ary membrane as narrow tubular structures at sites termed“crista junctions” and expand as they project into the matrix(Frey and Mannella 2000; Mannella et al. 2001) (Figure1B). It seems clear that the boundary and cristae domainsof the inner membrane have distinct compositions with re-spect to the respiratory complexes that are embedded pref-erentially in the cristae membrane domains, as well as othercomponents (Vogel et al. 2006; Wurm and Jakobs 2006;Rabl et al. 2009; Suppanz et al. 2009; Zick et al. 2009;Davies et al. 2011).
The outer and inner boundary membranes are connectedat multiple contact sites, at least some of which are involvedin protein translocation and may be transient (Pon andSchatz 1991). In addition, there appear to be firm contactsites, not directly involved with protein translocation, pref-erentially colocalized with crista junctions (Harner et al.2011a).
Overall, there appear to be �1000 distinct proteins inyeast mitochondria (Premsler et al. 2009). One series ofproteomic studies on highly purified organelles identified851 proteins thought to represent 85% of the total numberof species (Sickmann et al. 2003; Reinders et al. 2006;Zahedi et al. 2006). Another study identified an additional209 candidates (Prokisch et al. 2004). A computationallydriven search for candidates involved in yeast mitochondrialfunction, coupled with experiments to assay respiratoryfunction and maintenance of mitochondrial DNA (mtDNA),identified 109 novel candidates, although many of thesemay not be mitochondrial per se (Hess et al. 2009). Takingthe boundary and cristae domains together, the inner mem-brane is the most protein-rich mitochondrial compartment,followed by the matrix (Daum et al. 1982).
Only eight of the yeast mitochondrial proteins detected inproteomic studies are encoded by mtDNA and synthesizedwithin the organelle. They are hydrophobic subunits of
1204 T. D. Fox

respiratory complexes III (bc1 complex or ubiquinol-cytochromec reductase), IV (cytochrome c oxidase), and V (ATP synthase),as well as a hydrophilic mitochondrial small subunit ribosomalprotein. The remaining �99% of yeast mitochondrial proteinsare encoded by nuclear genes, synthesized in cytoplasmic ribo-somes, and imported into the organelle.
An overview of known nuclearly encoded mitochondrialprotein functions (Figure 2) reveals that �25% of them areinvolved directly in genome maintenance and expression ofthe eight major mitochondrial genes (Schmidt et al. 2010).The functions of �20% of the proteins are not known. Fif-teen percent are involved in the well-known processes ofenergy metabolism. Protein translocation, folding, and turn-over functions occupy �10% of mitochondrial proteins.
The following discussion reviews our understanding ofthe biogenesis of mitochondria starting on the outside, thecytoplasm, and working inward through the mitochondrialcompartments.
Cytoplasmic Synthesis of Mitochondrial Proteins
Localization of some cytoplasmic messenger RNAsto mitochondria promotes import of the proteinsthat they encode
Expression of nuclear genes coding mitochondrial proteinsbegins with the transcription of messenger RNAs (mRNAs).
(Mechanisms controlling the synthesis of these mRNAs arebeyond the scope of this review.) It has been known forsome time that the synthesis of proteins destined to reside inmitochondria can occur on polysomes bound to mitochon-dria or on other polysomes, usually referred to as “free poly-somes” (Kellems et al. 1975; Ades and Butow 1980; Suissaand Schatz 1982). More recently, surveys of the intracellularlocations of specific mRNAs encoding the bulk of the mito-chondrial proteome have indicated a range, with approxi-mately half of them selectively translated at the surface ofthe outer membrane, while translation of others occurs se-lectively on free polysomes or is not biased between mito-chondrial and cytoplasmic locations (Corral-Debrinski et al.2000; Marc et al. 2002; Garcia et al. 2007a; Saint-Georgeset al. 2008; Gadir et al. 2011) (Figure 3). It is easy to imag-ine that the biological rationale for localized synthesis oforganellar proteins is to promote their efficient import andassembly. The rationale for synthesizing roughly half of mi-tochondrial proteins on free cytoplasmic polysomes remainsto be discerned.
What directs and tethers so many mitochondrially boundmRNAs to the outer surface of the organelles? Currentevidence indicates the involvement of nucleotide signals inmRNA 39-untranslated regions (39-UTRs) that function priorto translation. In addition, the familiar (Pon and Schatz1991) mitochondrial targeting signals in the amino acidsequences of the precursor proteins that the mRNAs encodealso appear to contribute mRNA localization by mechanismsthat are at least partially redundant—and poorly understood(Lithgow 2000).
The ATM1 mRNA, which encodes an essential innermembrane transporter protein, is among those that arehighly enriched on mitochondrial-bound polysomes (Corral-Debrinski et al. 2000). By examining the localization of chi-meric mRNAs, it was shown that the ATM1 39-UTR wassufficient to direct mitochondrial localization of a reportermRNA lacking any other mitochondria-related signals, al-though this did not lead to import of the GFP reporter pro-tein itself into mitochondria. The 39-UTR of the PGK1mRNA, which encodes a soluble cytoplasmic protein, didnot direct localization of the mRNA to mitochondria (Corral-Debrinski et al. 2000).
The physiological significance of mRNA localization sig-nals in 39-UTRs was demonstrated by a study in which the39-UTR of the ATP2 mRNA, which encodes the b-subunit ofthe F1 ATP synthase, was replaced by 39-UTR of the ADH1mRNA by alteration of the chromosomal ATP2 locus (Margeotet al. 2002). This alteration prevented normal growth on non-fermentable carbon sources, presumably due to decreasedATP synthase activity. The swap of 39-UTRs did not affect theoverall steady-state level of ATP2 mRNA. However, it didcause a large reduction in the fraction of ATP2 mRNA asso-ciated with mitochondria and a large increase of this mRNAin the free polysomal fraction.
The behavior of the Atp2 precursor protein translatedfrom the altered mRNA was particularly interesting: while
Figure 1 Overview of mitochondrial structure in yeast. (A) Schematic ofcompartments comprising mitochondrial tubules. The outer membranesurrounds the organelle. The inner membrane surrounds the matrix andconsists of two domains, the inner boundary membrane and the cristaemembranes, which are joined at cristae junctions. The intermembranespace lies between the outer membrane and inner membrane. (B) Elec-tron tomograph image of a highly contracted yeast mitochondrion ob-served en face (a) with the outer membrane (red) and (b) without theouter membrane. Reprinted by permission from John Wiley & Sons fromMannella et al. (2001).
Mitochondrial Proteins 1205

normal levels of protein were associated with mitochondria,it was overwhelmingly in the larger precursor form, retain-ing the 34-amino-acid N-terminal targeting signal (Margeotet al. 2002). This contrasted with Atp2 translated from thewild-type mRNA, which was overwhelmingly in the matureprocessed form. Furthermore, the pre-Atp2 protein trans-lated from the altered mRNA appeared to be on the outsideof the outer membrane. Taken together, these results sug-gest that localization of wild-type ATP2 mRNA to mitochon-dria via signals in its 39-UTR promotes synthesis of animport-competent pre-Atp2 polypeptide. Synthesis of pre-Atp2 from an mRNA lacking this 39-UTR on free polysomesyields a protein that binds the outer surface of mitochondriabut fails to be imported efficiently, presumably due to al-tered structure or interactions with other proteins. Thus,mRNA localization directed by a signal in the ATP2 mRNA39-UTR promotes efficient translocation of the protein intomitochondria.
Based on this observation, one should use caution whenconducting genetic experiments involving alteration of mito-chondrial proteins by methods that also alter the 39-UTRs oftheir mRNAs.
A large-scale survey of mRNA abundance in mitochond-rially bound polysomes vs. free polysomes was carried out byhybridization to microarrays. After corrections for the amountof mRNA in each fraction, and for cross-contamination offractions, the degree to which mRNAs encoding the yeastproteome are selectively localized to mitochondria was de-termined (Garcia et al. 2007b). The ATP2 mRNA, for exam-ple, ranks among the most highly localized to mitochondria,with 50% of the total bound to the organelles. At the otherextreme of the distribution, none of the COX4 mRNA en-coding cytochrome c oxidase subunit IV was found amongmitochondria-bound polysomes (Figure 3). The degree ofmitochondrial association for each mRNA was then scoredby the rank of its ratio among all mRNAs (Garcia et al.2007b). About half of the 423 mRNAs known to encodemitochondrial proteins at the time of this study were found
to be preferentially associated with mitochondria, while rel-atively few mRNAs encoding known non-mitochondrial pro-teins were in this group (Marc et al. 2002). The subcellularmRNA distributions observed using genomic microarrayswere confirmed by a focused study on 112 mRNAs encodingprotein components of known mitochondrial complexes, us-ing quantitative PCR to assay the fractions (Garcia et al.2007a).
The notion that the strong association of specific mRNAswith mitochondria is correlated with the behavior of theproteins that they encode received striking support froma careful proteomic study of highly purified vesicles derivedfrom the mitochondrial outer membrane (Zahedi et al. 2006).Forty-nine proteins, with a wide range of abundance, werefound to be exposed on the outer surface of mitochondria.Surprisingly, 36 of these surface proteins were already wellknown to actually reside in internal mitochondrial compart-ments. Two such anomalous surface proteins, Atp2 andCyb2, were experimentally shown to be unprocessed precur-sors of the mature internal proteins, and it is likely that atleast several others are as well (Zahedi et al. 2006). Thus,a significant group of precursors, possibly awaiting import tointernal destinations and processing, are bound to the sur-face of mitochondria. Significantly, 16 of these proteins areencoded by the 25 mRNAs most selectively localized tomitochondrial-bound polysomes as determined by Marcet al. (2002). Zahedi et al. (2006) performed comparisonsof the entire yeast proteome, the mitochondrial proteome,and the internal mitochondrial proteins detected on the sur-face of the outer membrane, with the degree of mitochon-drial localization of all yeast mRNAs as scored by rank (Marcet al. 2002). These distributions demonstrated a strong biasfor localized mRNAs to encode internal proteins that werealso detected at the surface.
Taken together, these findings appear to be paradoxical.If the purpose of localized translation at the surface ofmitochondria is to promote efficient cotranslational importof proteins, then why should the locally synthesized proteins
Figure 2 Classification of identified mitochondrial pro-teins according to function. Reprinted by permission fromNature Publishing Group from Schmidt et al. (2010).
1206 T. D. Fox

be preferentially found among the full-length unimportedand unprocessed species detected on the organellar surface?Such molecules were clearly not cotranslationally imported.Perhaps these proteins, as a group, tend to rapidly adoptfolded conformations that inhibit translocation. In this case,localized synthesis could be an adaptation that alleviatesthis problem, albeit incompletely, by facilitating cotransla-tional import of a significant fraction of molecules. It iscurrently unknown whether the fully synthesized precursorproteins bound to the mitochondrial outer surface aredestined to be imported or degraded (Zahedi et al. 2006).The observation that unprocessed pre-Atp2 accumulates toan abnormally high level outside of mitochondria in cellstranslating a chimeric ATP2 mRNA lacking the localizationsignal in its 39-UTR (Margeot et al. 2002) is consistent withthe possibility that post-translational import of pre-ATP2 isinefficient in vivo, despite the fact that it occurs in vitro(Maccecchini et al. 1979). Perhaps the pre-ATP2 moleculesdetected on the surface of mitochondria were actually trans-lated from those ATP2 mRNA molecules, �50% of the total,that were not localized to mitochondria-bound polysomes.In any event, it is clear that localized synthesis of pre-ATP2somehow facilitates its import.
Another fascinating but imperfect correlation emergedfrom the ranking of mRNAs by their propensity to bemitochondrially localized. Those mRNAs found most selec-tively in mitochondria-bound polysomes tend to encodeproteins whose evolutionary origins can be clearly traced toBacteria and/or Archaea. Conversely, those mRNAs foundmost selectively in free polysomes tend to encode proteinslacking clear homologs in those phylogenetic domains andare therefore likely to be more recently evolved inventions
of Eukarya (Marc et al. 2002; Garcia et al. 2007a). This corre-lation runs in parallel with the observation that the more locallysynthesized proteins tend to be either ancient, conserved com-ponents of the mitochondrial genetic system or respiratory com-plexes or conserved proteins with roles in assembly of thosecore components (Margeot et al. 2005). In the case of cyto-chrome c oxidase, for example, proteins with bacterial orthologsthat assemble mitochondrially coded core subunits in the innermembrane, insert metal cofactors, and synthesize the specificheme A cofactor are all selectively translated at the mitochon-drial surface (although they do not all have clear bacterial orarchaeal ancestors). In contrast, the eukaryotic-specific subunitsof cytochrome c oxidase that surround the catalytic core of theenzyme are all selectively translated on free polysomes.
There are no obvious structural or chemical similaritiesamong the set of proteins most selectively synthesized at themitochondrial surface (Marc et al. 2002). So, what selectiveconstraints maintain the localized translation of more an-ciently evolved proteins? It has been argued that synthesislocalized to mitochondria may promote efficient assembly ofcore components of complexes by directing import of pro-teins to specific regions (Margeot et al. 2005; Garcia et al.2007a). While this is an attractive hypothesis, there is nostrong evidence that localized synthesis of any mitochon-drial protein is spatially organized on the organellar surface.Nor is it obvious why nonlocalized translation of other es-sential but peripheral subunits of complexes would be ad-vantageous. Interestingly, the mitochondria-bound mRNAstend to be synthesized early during the yeast metabolic cycle(Tu et al. 2005; Lelandais et al. 2009).
Complex mechanisms for mRNA localization
Regardless of why some mRNAs are localized to mitochon-dria while others are not, the example of ATP2 demonstratesthe importance of mRNA targeting for mitochondrial bio-genesis. How are localized mRNAs brought and tetheredto the organelles? mRNA 39-UTRs contain information forlocalization in at least eight cases that have been experimen-tally examined (Corral-Debrinski et al. 2000; Marc et al.2002; Margeot et al. 2002).
One factor with apparent roles in localization of manymRNAs encoding mitochondrial proteins is Puf3, a member ofthe Pumilio-homology domain family (PUF) of RNA-bindingproteins. PUF family proteins are found in a wide variety ofeukaryotes and carry out a wide variety of functions throughtheir ability to mediate interactions between target RNAsand other proteins (Quenault et al. 2011). An initial surveyof yeast PUF protein functions (Olivas and Parker 2000)revealed that a puf3Δ mutation strongly affected theCOX17 mRNA, which encodes a (Eukarya-specific) mito-chondrial copper-binding protein required for cytochromec oxidase assembly (Glerum et al. 1996). The presence ofPuf3 was shown to stimulate deadenylation and degradationof COX17 mRNA, but did not affect its translation as mea-sured by the degree of polysome association or the level ofaccumulated Cox17 protein.
Figure 3 Cytoplasmic synthesis of some mitochondrial proteins is local-ized to the organelles, while the synthesis of others is not. The figuredepicts three examples: (1) The ATP2 mRNA is highly localized tomitochondria-bound polysomes (Garcia et al. 2007b), although factorsrequired for this localization are unknown. (2) The BCS1 mRNA is alsoselectively found in mitochondria-bound polysomes, and its localization ispartially dependent upon the mitochondrially localized RNA-binding pro-tein Puf3 and the Puf3-binding sites in its 39-UTR (Saint-Georges et al.2008). (3) The COX4 mRNA is exclusively found on free polysomes, un-associated with mitochondria (Garcia et al. 2007b). The Atp2, Bcs1, andCox4 proteins all traverse the outer membrane via the TOM complexpore.
Mitochondrial Proteins 1207

A genomic investigation of RNAs bound to Puf3 (as wellas other members of the yeast Puf protein family) revealeda striking specificity: among 154 Puf3-binding mRNAsof known function, 135 encoded mitochondrial proteins(Gerber et al. 2004). Furthermore, mitochondrial proteinscoded by 80 of the Puf3-binding mRNAs have roles in organ-ellar translation (e.g., mitochondrial ribosomal proteins)while most of the rest participate in post-translational as-sembly functions. A Puf3-binding sequence was identified inthe 39-UTRs of these mRNAs (Gerber et al. 2004). This siteoccurs twice in the COX17 mRNA, and those sites are nec-essary for Puf3-dependent mRNA destabilization (Jacksonet al. 2004).
The Puf3 protein itself was found to be located on theouter surface of mitochondria and visualized in puncta largelyassociated with mitochondrial tubules (Garcia-Rodriguezet al. 2007). Puf3 was also associated with Mdm12, a proteincomponent of the tether that connects distinct sites on mito-chondria with the endoplasmic reticulum (Garcia-Rodriguezet al. 2007; Kornmann et al. 2009). Consistent with a role inpromoting degradation of mRNAs required for respiratorymetabolism, and thereby affecting the production of respi-ratory complexes, overproduction of Puf3 caused a modestreduction in the growth of cells on the nonfermentable car-bon source glycerol (Garcia-Rodriguez et al. 2007) and ox-ygen consumption (Chatenay-Lapointe and Shadel 2011).Log-phase cells lacking Puf3 contained elevated levels ofrespiratory complex subunits and exhibited increased ratesof oxygen consumption (Chatenay-Lapointe and Shadel2011). However, Puf3 appears to be more than simplya post-transcriptional repressor of mitochondrial functionssince a puf3Δ also produces a very modest defect in growthon glycerol (Gerber et al. 2004).
Taken together, these findings suggest the possibility thatPuf3 could have a direct role in localizing a number ofmRNAs to the mitochondrial surface. Consistent with thispossibility, a majority of those Puf3-binding mRNAs thatencode known mitochondrial proteins (Gerber et al. 2004),and were examined for subcellular distribution (Marc et al.2002), were among those selectively localized to mitochon-dria. Overall, it appears that about half of the mRNAs selec-tively localized to mitochondria contain Puf3-binding sites intheir 39-UTRs, and the localization of about half of those issignificantly decreased in the absence of Puf3 (Saint-Georgeset al. 2008). In addition, mutation of the Puf3-binding site inone such mRNA (BCS1) reduced its selective association withmitochondria by a factor of two as assayed both by quantita-tive PCR of the two polysome fractions and by quantitation ofRNA granule location cytologically in FISH images (Saint-Georges et al. 2008) (Figure 3).
Similar results were obtained for a set of 24 mRNA-encoding mitochondrial proteins that were tagged withbinding sites for an RNA-binding GFP fusion protein andvisualized in granules (Gadir et al. 2011). These imagessuggest the possibility that mRNAs bound to the surface ofmitochondria may not be evenly distributed on the organ-
ellar surface. Experiments to test whether these RNA gran-ules colocalize with Puf3 puncta on mitochondria have notbeen reported. However, if they do, the fact that Puf3 asso-ciates with the mitochondrial-ER tether protein Mdm12(Garcia-Rodriguez et al. 2007; Kornmann et al. 2009) wouldbe consistent with reported partial colocalization of mito-chondrial mRNA granules with ER (Gadir et al. 2011). Suchlocalization of protein synthesis and presumably importcould facilitate assembly of mitochondrial complexes, forexample, mitochondrial ribosomes (Saint-Georges et al. 2008).
It is important to bear in mind that Puf3 promotes deg-radation of at least some mitochondrially localized mRNAs(Olivas and Parker 2000; Jackson et al. 2004; Foat et al.2005). Thus, even if Puf3 were not directly involved in lo-calization, an mRNA stabilized in the absence of Puf3, or bymutation of its Puf3-binding site, could appear to be lessselectively bound to mitochondria as measured by the ratioof its presence in bound vs. free polysomes. This would occurif an RNA’s abundance increased sufficiently to saturateother limiting localization factors on the organelle surface.The extent to which altered RNA stability may contribute toPuf3 dependence of localization has not been systematicallyexplored. Nevertheless, it seems likely that Puf3 bindingcontributes directly to localization of those mRNAs bearingits binding site in addition to influencing their rates of deg-radation. How the interplay between these two activitiesinfluences protein import and assembly of mitochondrialcomplexes remains an open question (Quenault et al. 2011).
The existence of distinct mechanisms for RNA sequence-based mRNA recognition is indicated by the fact that the ATM1and ATP2 mRNAs, whose 39-UTRs clearly cause mitochondriallocalization, lack Puf3-binding sites (Corral-Debrinski et al.2000; Margeot et al. 2002; Saint-Georges et al. 2008). Se-lection of variant sequences derived from the ATP2 39-UTRthat functionally localize the mRNA suggest that both nu-cleotide sequence and secondary structural features playa role in its recognition (Liu and Liu 2007). However, noprotein or other species that interact with this RNA elementhave been identified (Figure 3). Interestingly, the absence ofPuf3 may reduce mitochondrial localization of the ATP2mRNA, presumably by an indirect mechanism (Gadir et al.2011), although this observation is inconsistent with an ear-lier report (Saint-Georges et al. 2008).
Tethering of mRNAs to mitochondria via nascentpolypeptide chains
Actively translated mRNAs can be tethered to membranesvia nascent polypeptide chains undergoing cotranslationalmembrane translocation. This appears to occur in the case ofat least some mRNAs localized to mitochondria. A chimericmRNA encoding the Atm1 N-terminal mitochondrial targetingsignal fused to GFP, but with the ATM1 39-UTR replaced bythe PGK1 mRNA 39-UTR, localized to mitochondria (Corral-Debrinski et al. 2000). Thus, the wild-type ATM1 mRNA onmitochondria appears to be localized both by an untranslatedsignal in its 39-UTR and by the interaction of the polypeptide
1208 T. D. Fox

targeting signal with receptors on the outer mitochondrialsurface and the protein import machinery.
In the case of an ATP2 mRNA lacking its normal 39-UTR,residual localization to mitochondria required translation ofboth the N-terminal targeting sequence and sequences withinthe mature protein itself (Garcia et al. 2010). Normal associ-ation of the wild-type ATP2 mRNA also required one of thethree Translocase of the Outer Membrane (TOM) complexouter membrane import receptor proteins, Tom70 (Table 1),and was reduced by mutation of the ATP2 translation initi-ation codon (Gadir et al. 2011).
Deletion of another outer membrane import receptorprotein, Tom20, was found to lower but not eliminate selec-tive localization of most mRNAs associated with mitochon-dria (Eliyahu et al. 2010). While tom20Δ mutants are viablewith a modest respiratory defect, and puf3Δ mutants areviable and almost wild type with respect to respiratorygrowth, a tom20Δ, puf3Δ double mutant was viable witha very tight respiratory defect. This synthetic respiratoryphenotype is consistent with the picture of synergy in target-ing of mRNAs to mitochondria by factors recognizing mRNA39-UTRs and the protein import machinery acting on nascentchains to promote efficient assembly of functional mitochon-drial complexes. At the same time, the viability of thetom20Δ, puf3Δ double mutant demonstrates that proteinimport to mitochondria remains active by the action of par-tially redundant pathways for mRNA localization and pre-cursor recognition.
Translocation and Membrane Insertion ofCytoplasmically Synthesized Mitochondrial Proteins
The distant ancestors of mitochondria were bacteria fromthe a-proteobacterial lineage (Gray et al. 2001). While theorigins of all known extant eukaryotes trace back to organismsthat contained both mitochondria and nucleo-cytoplasmic ge-netic systems related to Archaea, the events leading to en-dosymbiosis and the subsequent evolution of mitochondriaas integrated cellular organelles have not been clearly dis-cerned (Embley and Martin 2006). However, since bacteriaare not known to import large polypeptides, their evolu-tion into mitochondria apparently required the evolutionof new mechanisms for the transport of cytoplasmically
synthesized proteins across one or both of the mitochon-drial (formerly bacterial) membranes. Some componentsof the present-day protein import machinery are clearlyof bacterial origin. However, most appear to have distantbacterial homologs that do not participate in protein trans-location or to have evolved de novo as endosymbionts becameorganelles (Dolezal et al. 2006; Kutik et al. 2009; Hewitt et al.2011).
Transport of cytoplasmically synthesized mitochondrialproteins or their precursors across or into the outer mem-brane is carried out by the TOM complex, which includesboth receptor proteins facing the cytoplasm and a pore inthe membrane (Table 1). It is widely believed that the pre-cursor proteins arrive at the organelle bound by chaperonesand, in that state, are recognized by receptors of the TOMcomplex, although this has been demonstrated in only a fewcases (Gautschi et al. 2001; Young et al. 2003).
Depending upon the nature of their targeting signals,proteins may be inserted into the outer membrane, trans-located into the intermembrane space (IMS), or delivered toone of the two Translocase of the Inner Membrane (TIM)complexes for insertion into the inner membrane or trans-location into the matrix (Pon and Schatz 1991; Neupert1997; Voos et al. 1999). (While the nature of targeting sig-nals for different compartments has been investigated inten-sively, it is important to note that they cannot be predictedsolely from sequence information with a high degree of cer-tainty.) A wide variety of different translocation and sortingevents must be completed prior to, or concomitant with, theassembly of imported proteins into functional multimericenzymes and higher-order complexes.
The literature on import of proteins into yeast mitochon-dria is extensive and has been extensively reviewed. Recentreviews present detailed descriptions of the componentsof import complexes and their functions (Young et al.2003; Neupert and Herrmann 2007; Chacinska et al. 2009;Koehler and Tienson 2009; Mokranjac and Neupert 2009;Walther and Rapaport 2009; Endo and Yamano 2010;Schmidt et al. 2010; Dukanovic and Rapaport 2011; Gebertet al. 2011; Hewitt et al. 2011; Marom et al. 2011a; Riemeret al. 2011; Yogev and Pines 2011). Molecular structures ofhydrophilic domains of proteins composing the importmachinery are emerging, but as yet no full structures of
Table 1 Components of the TOM complex: transport of proteins through the outer membrane
Protein ORF Known function Null phenotype
Mim1 YOL026C Insertion of transmembrane helix proteins into the outer membrane InviableMim2 YLR099W-A Insertion of transmembrane helix proteins into the outer membrane InviableTom5 YPR133W-A Viable, various defectsTom6 YOR045W Viable, various defectsTom7 YNL070W Viable, various defectsTom20 YGR082W Receptor for substrates with presequences Viable, various defectsTom22 YNL131W Central receptor facing cytoplasm and IMS; interaction with TIM23 complex Viable, various defectsa
Tom40 YMR203W Translocation channel–b-barrel structure InviableTom70 YNL121C Receptor for substrates lacking presequences Viable, various defectsTom71 YHR117W Receptor Viable, various defectsa Null was inviable in large-scale studies, but is viable if obtained by loss of TOM22 plasmid during mitotic growth (van Wilpe et al. 1999).
Mitochondrial Proteins 1209

mitochondrial translocation complexes are available (Endoet al. 2011), precluding, for the most part, precise biochem-ical descriptions of mechanisms. Outlined below are theroutes taken by cytoplasmically synthesized proteins des-tined for the outer membrane, the intermembrane space,the inner membrane, and the matrix. The known pathwaysto these compartments overlap for most proteins as theytraverse the outer membrane, but then become distinct.
Insertion of proteins into the outer membrane
All proteins entering mitochondria first encounter pre-existingouter membrane proteins and lipids. Thus, outer membraneproteins are crucial for all import, including the biogenesisof the outer membrane itself. All outer membrane proteinsare synthesized in the cytoplasm, and none are known to beproteolytically cleaved during import or assembly (Schmidtet al. 2010). The signals that target these proteins are poorlyunderstood but appear to reside in transmembrane domains(Mokranjac and Neupert 2009; Walther and Rapaport2009).
Import and insertion of b-barrel proteins: The over-whelming majority of cytoplasmically synthesized proteinsthat become incorporated into mitochondrial structures firstassociate with the organelles by interaction with the TOMcomplex in the outer membrane (Endo and Yamano 2010).Among them are the integral proteins of the mitochondrialouter membrane with b-barrel structures, including the mostabundant, Por1 (porin) (Riezman et al. 1983), a voltage-gatedanion channel. Another key b-barrel protein is Tom40 (Bakeret al. 1990), which forms the TOM complex pores in theouter membrane through which most imported proteinspass (Hill et al. 1998; Künkele et al. 1998).
Both of these b-barrel proteins are translated on freecytoplasmic polysomes (Saint-Georges et al. 2008) and di-rected to mitochondria by unknown signals and mechanisms(Mokranjac and Neupert 2009). It is also unknown whetherthey arrive at the TOM complex associated with cytoplasmicchaperones. In any event, these apparently unfolded b-barrelproteins pass through the Tom40 pore of the TOM complexafter interaction with the TOM receptor subunits Tom20 andTom22 (Krimmer et al. 2001; Model et al. 2001) (Figure 4).
At this point, the b-barrel proteins are present in the IMS,which is homologous to the periplasm of Gram-negativebacteria. In this soluble milieu, they are bound by hetero-hexameric chaperone complexes composed of the smallproteins (�100 amino acids each) Tim9-Tim10 and ofTim8-Tim13 (Hoppins and Nargang 2004; Wiedemannet al. 2004) (Table 2). The crystal structure of the Tim9-Tim10 hexamer reveals a propeller arrangement with 12mobile a-helical tentacles descending from a core stabilizedby intramolecular disulfide bonds in each subunit (Bakeret al. 2009). The N-terminal tentacle of Tim9 is especiallyimportant for substrate binding in vivo. These chaperonesaccompany the b-barrel proteins back to the inner surfaceof the outer membrane where they are delivered to the
Sorting and Assembly Machinery (SAM) complex, embeddedin the outer membrane (also known as TOB for Topogenesisof b-Barrel proteins) (Paschen et al. 2003; Wiedemann et al.2003a) (Table 3). The Sam35 component of the SAM com-plex recognizest b-barrel proteins by virtue of an amino acidsequence near their C termini (Wiedemann et al. 2003a;Kutik et al. 2008). The b-barrel proteins associate with theSAM complex from the IMS side and are then inserted lat-erally into the outer membrane lipid bilayer by an as-yet-unknown mechanism, where they assume b-barrel structure(Stroud et al. 2011) (Figure 4).
The essential core component of the SAM complex,Sam50, is itself a b-barrel protein. Thus, the assembledand functional b-barrel proteins Tom40 and Sam50 are nec-essary for import and assembly of newly synthesized Tom40and Sam50, as well as that of other outer membraneb-barrel proteins. Sam50 is homologous to the bacterial out-er membrane protein Omp85, which has a similar functionin the insertion of b-barrel proteins in the outer membraneof Gram-negative bacteria (Paschen et al. 2003; Gentle et al.2004).
Insertion of other integral proteins into the outer mem-brane: In addition to b-barrel proteins, the outer membranecontains integral proteins anchored in the lipid bilayer byone or more individual transmembrane domains. There ap-pear to be multiple pathways for such proteins, and they arenot well understood at present. At least some integral outermembrane proteins are exceptional in that they do nottraverse the membrane via the Tom40 pore of the TOMcomplex. In the case of proteins with multiple membrane-spanning helices, the newly synthesized polypeptides firstcontact the outer membrane via the Tom70 receptor, butare then inserted into the bilayer from the outside, indepen-dently of Tom40, through their interaction with the multimericcomplexes of Mim1 (Becker et al. 2011; Papic et al. 2011).Mim1 is a short (113 amino acids) single-spanning outer
Figure 4 Insertion of proteins into the outer membrane. b-Barrel proteinsare imported through the pores of the TOM complex in the outer mem-brane and then bound by IMS chaperone complexes comprising Tim9 andTim10. The b-barrel-Tim9-Tim10 complexes bind to the inner surfaces ofSAM complexes in the outer membrane, leading to insertion of b-barrelproteins into the outer membrane lipid bilayer. Some integral outer mem-brane proteins with multiple transmembrane domains (TMD) contact theTom70 receptor and are then inserted into the bilayer from the outsidethrough their interaction with multimeric complexes of Mim1.
1210 T. D. Fox

membrane protein (Ishikawa et al. 2004; Waizenegger et al.2005) that forms dimers that organize into higher-ordercomplexes (Popov-�Celeketić et al. 2008b) that were re-cently also shown to contain a second protein, Mim2(YLR099W-A) (Dimmer et al. 2012). These Mim1-Mim2complexes appear to have a membrane insertase function(Figure 4).
Mim1 is also required for insertion of at least some pro-teins with a single transmembrane domain near the theirN termini, often termed signal anchored proteins. These in-clude the outer membrane receptor proteins Tom20 andTom70 (Becker et al. 2008; Hulett et al. 2008; Popov-�Celeketić et al. 2008b). Interestingly, insertion of thesereceptors does not depend upon their own receptor function(Ahting et al. 2005).
Mim1 itself has a conserved, centrally located transmem-brane domain that is partially functional even in the absenceof both flanking hydrophilic domains (Popov-�Celeketić et al.2008b). The Mim1 C-terminal domain is exposed to thecytoplasm (Lueder and Lithgow 2009; Walther and Rapaport2009). The pathway that Mim1 takes into the outer mem-brane has not yet been studied.
There is apparently at least one additional pathway intothe outer membrane employed by proteins anchored in themembrane by a single transmembrane domain at their Ctermini, the so-called “tail-anchored proteins.” In the case ofthe tail-anchored protein Fis1, required for normal mito-chondrial fission, outer membrane insertion is independentof all known components of the TOM and SAM complexes(Kemper et al. 2008). Furthermore, the insertion of Fis1 intolipid vesicle membranes with a low ergosterol content re-sembling the mitochondrial outer membrane indicates thatlipid content may play a role in specificity in vivo. A possiblerole for Mim1 in Fis1 insertion was not tested. Another tail-anchored protein, the essential receptor Tom22, enters themembrane through the direct or indirect action of the SAMcomplex, after being recognized on the surface by TOM re-ceptors (Stojanovski et al. 2007). Thus, the SAM complex
may not be specific for the insertion of b-barrel proteins andmay recognize substrates on either side of the membrane.The insertion of Tom22 is not dependent upon Mim1 (Beckeret al. 2008).
Recently, evidence indicating the possibility of lateraldiffusion of transmembrane domains out of the TOM com-plex has been reported (Harner et al. 2011b). Chimeric fu-sion proteins were trapped across the outer membrane bythe folded structure of GFP on the outside and the multi-spanning inner membrane protein Tim23 inserted in the in-ner membrane. Fusion proteins with transmembrane domainsaccessible to the outer membrane were released from theTOM complex by an unknown mechanism. It remains to bedetermined whether any endogenous mitochondrial proteinsemploy this route into the outer membrane.
Import of proteins into the IMS
There are at least three mechanisms by which proteins arelocalized to the intermembrane space. Two involve covalentmodifications of precursors after transit across the outermembrane by enzymes located in the intermembrane spacethere. The modifications stabilize folded structures thatprevent retrograde transport out of the organelle. Trappingby noncovalent bonds may also occur in some cases. Finally,as discussed below in conjunction with transport to the innermembrane, some proteins are first targeted to the innermembrane and then released into the IMS by proteolyticcleavage.
Covalent attachment of heme: Cytochrome c (Cyc1 andCyc7), which is located in the IMS, is perhaps the most in-tensively genetically analyzed S. cerevisiae protein (Sherman2005). Surprisingly, the import of cytochrome c to the IMS isstill relatively poorly understood. Cyc1 is largely synthesizedon mitochondria-bound polysomes (Saint-Georges et al.2008) and requires the TOM complex to traverse the outermembrane in a reaction that does not require ATP or aninner membrane potential (Diekert et al. 2001; Wiedemann
Table 2 IMS import chaperones: delivery of hydrophobic proteins to the SAM complex (outer membrane) or TIM22complex (inner membrane)
Protein ORF Known function Null phenotype
Tim8 YJR135W-A Complexed with Tim13 Viable, various defectsTim9 YEL020W-A Complexed with Tim10 InviableTim10 YHR005C-A Complexed with Tim9 InviableTim12 YBR091C Associated with Tim22 complex InviableTim13 YGR181W Complexed with Tim8 Viable, various defects
Table 3 Components of the SAM complex: insertion of b-barrel proteins into the outer membrane
Protein ORF Known function Null phenotype
Mdm10 YAL010C Viable, various defectsSam35 YHR083W Receptor InviableSam37 YMR060C Viable, various defectsSam50 YNL026W b-Barrel protein of SAM complex Inviable
Mitochondrial Proteins 1211

et al. 2003b). However, the mechanism by which the TOMcomplex translocates apo-cytochrome c remains enigmatic.Blockage of the Tom40 pores used by other TOM substratesdoes not prevent import of apo-cytochrome c into membranevesicles containing purified TOM complexes. Furthermore,removal of cytosolic domains of the TOM receptor subunitsdid not affect apo-cytochrome c import into mitochondria(Wiedemann et al. 2003b; Yamano et al. 2008). Neverthe-less, complete removal of the Tom22 receptor did preventimport (Wiedemann et al. 2003b). Thus, Tom22 domainswithin the outer membrane or exposed on its inner surfacemay play a role in this as-yet-enigmatic translocation pro-cess. This behavior of apo-cytochrome c is unique amongstudied proteins.
Once in the IMS, apo-cytochrome c binds with the cyto-chrome c heme lyase, Cyc3, which is itself bound peripher-ally to the outer surface of the inner membrane (Dumontet al. 1991; Steiner et al. 1995; Bernard et al. 2005). Apo-cytochrome c is then irreversibly trapped by the covalentattachment of heme, which forms mature cytochrome c(Dumont et al. 1991) (Figure 5). The first 27 amino acidsof cytochrome c contain the residues required for heme at-tachment, and they appear to be required for import (Wanget al. 1996). It is not clear whether this region is required forinteraction with the TOM complex in addition to the hemelyase. In any event, apo-cytochrome c does not selectivelypartition to mitochondria in the absence of Cyc3, or if cyc1mutations block the heme lyase reaction, although smallamounts are associated with the organelles (Dumont et al.1991). Interestingly, even when cyc1 mutations block hemeattachment, overexpression of heme lyase increases parti-tioning of the mutant apo-cytochrome c to mitochondria,suggesting that protein–protein interactions alone initiallysequester it (Dumont et al. 1991).
The cytochrome c heme lyase Cyc3 is itself imported intothe intermembrane space via the TOM complex (Steineret al. 1995). Based on studies with Neurospora crassa, a con-served targeting signal for heme lyases has been identifiedwithin its amino acid sequence, and this 60-residue regioncan target passenger proteins to the IMS (Diekert et al.1999). This signal is believed to interact with cis and transsites of the TOM complex and may also direct binding tounknown components of the inner membrane that couldanchor it in the IMS.
Oxidation of paired cysteine residues to form disulfidebonds: A second form of covalent modification that seques-ters some IMS proteins is the generation of internal disulfidebonds between paired cysteine residues (CX3C or CX9Cmotifs) after import. There are at least 24 such proteins(Koehler and Tienson 2009), including the small chaperoneproteins Tim8, Tim9, Tim10, Tim12, and Tim13 whosefolded structures are known to be stabilized by intramoleculardisulfide bonds (Baker et al. 2009). Like the Gram-negativebacterial periplasmic space (Messens and Collet 2006), theIMS is a more oxidizing environment than the cytoplasm
(Hu et al. 2008) and contains enzymatic machinery for thecontrolled generation of intramolecular disulfide bonds(Koehler and Tienson 2009; Herrmann and Riemer 2012).
Some IMS proteins with paired Cys residues are synthe-sized on mitochondria-bound polysomes (e.g., Pet191, Cox23,Cox17), while others (e.g., Tim9, Tim13) are not (Saint-Georges et al. 2008). It is not clear what directs these proteinsto mitochondria (Riemer et al. 2011). Import of Tim13 doesnot depend upon surface receptors of the TOM complex, butit apparently does enter mitochondria through the Tom40pore of the TOM complex (Lutz et al. 2003). The Tom5 sub-unit of the TOM complex is also required (Kurz et al. 1999).Chemical modification or mutation of the Cys residues pre-vented accumulation of Tim13 in the IMS (Lutz et al. 2003).Uptake of Tim10 was blocked if Cys residues were oxidizedprior to import (Lu et al. 2004).
Mia40 is an essential protein bound to the outer surfaceof the inner membrane that is required for import of theessential Tim9-Tim10 chaperones and other twin-Cys pro-teins to the IMS (Chacinska et al. 2004; Naoe et al. 2004).These imported proteins associate with Mia40 via disulfidebonds. As one would expect, import of cytochrome c doesnot require Mia40 (Chacinska et al. 2004). A short peptidesequence containing a single Cys residue has been identifiedin several twin-Cys proteins that directs them to Mia40 in theIMS and binds covalently to it via a disulfide bond (Milenkovicet al. 2009; Sideris et al. 2009). This signal may also promotepassage from the cytoplasm to the IMS, but it is not clear whatouter membrane surface component could be involved in thisrecognition. The interaction of newly imported reducedtwin-Cys substrates with oxidized Mia40 promotes foldingof the substrate proteins and the formation of disulfidebonds, trapping the folded proteins in the intermembranespace (Banci et al. 2010) (Figure 5).
Figure 5 Trapping of proteins in the IMS by covalent modification. Apo-cytochrome c (Cyc1) traverses the outer membrane via the TOM complexby an unusual and poorly understood mechanism (see text). Covalentattachment of heme by the lyase (Cyc3), bound to the outer surface ofthe inner membrane, generates holo-cytochrome c. Holo-cytochrome ccannot translocate through the TOM complex and remains in the IMS. Inan analogous mechanism, IMS proteins with twin-Cys residue pairs inreduced form are imported through the TOM complex and then oxidizedby the Mia40-Erv1 disulfide relay system bound to the inner membrane.The internal disulfide bonds formed in the twin-Cys proteins prevent re-verse translocation.
1212 T. D. Fox

Reduced Mia40 is oxidized in turn by the essential in-termembrane space protein Erv1, a conserved flavin-linkedsulfhydryl oxidase (Mesecke et al. 2005). Electrons from theresulting reduced Erv1 can be accepted by cytochrome c andenter the respiratory chain or be accepted by molecular ox-ygen to form hydrogen peroxide that is metabolized by cy-tochrome c peroxidase (Bihlmaier et al. 2007; Dabir et al.2007). This disulfide relay system has been reconstitutedin vitro (Tienson et al. 2009).
Although Erv1 does not contain the CX3C or CX9C motifspresent in the other substrates of this system, its import doesdepend upon Mia40 action following TOM-dependent pas-sage through the outer membrane (Terziyska et al. 2007).Interestingly, the import of another protein located partiallyin the IMS and lacking the closely paired Cys residue, Ccs1,depends upon Mia40 to form a disulfide bond, but that bondis not necessary for enzymatic activity (Gross et al. 2011;Klöppel et al. 2011). The import pathway followed byMia40, and other proteins anchored in the inner membranewith hydrophilic domains in the IMS, will be describedbelow.
Import of proteins into the inner membrane
The mitochondrial inner membrane contains a very widevariety of integral proteins. All studied cytoplasmically syn-thesized inner membrane proteins are recognized by recep-tors of the TOM complex and imported through its pores.They are then inserted into the inner membrane by one ofthree mechanisms, or combinations of them, depending uponthe signals that they contain and their ultimate topology.
Insertion of metabolite carriers and other multispanninginner membrane proteins by the TIM22 insertase/translocasecomplex: A major class of inner membrane proteins areimported and assembled into multispanning topologieswithout being proteolytically processed. At least 34 ofthese proteins are members of the metabolite carrier family(Palmieri et al. 2006), which includes the ATP/ADP car-riers. Two other such proteins are Tim22 and Tim23, theessential pore-forming subunits of the TIM complexes de-scribed below.
These multispanning membrane proteins contain multi-ple internal mitochondrial targeting signals that generallyflank transmembrane domains (Neupert and Herrmann2007; Chacinska et al. 2009). Newly synthesized yeast carrierproteins have been shown to associate with the cytoplasmicchaperone Hsp70, which participates in their recognitionby the Tom70 receptor subunit of the TOM complex (Younget al. 2003; Endo and Yamano 2010). ATP-dependent re-lease from the Hsp70 chaperones allows the carrier proteinsto enter the TOM complex pore (Figure 6). The N and Ctermini initially remain on the outside while internal regionscontaining recognition signals traverse the TOM complex aslooped polypeptide chains through the Tom40 pore in the out-er membrane (Wiedemann et al. 2001; Neupert and Herrmann2007; Chacinska et al. 2009). The Tom40 pore appears to have
specific interactions with different imported proteins and thusmay also play an active role in substrate recognition (Gabrielet al. 2003; Sherman et al. 2006).
On the inside of the outer membrane, incoming carrierproteins are removed from the TOM complex by binding tothe soluble essential Tim9-Tim10 chaperone complexes(Koehler et al. 1998; Sirrenberg et al. 1998; Curran et al.2002; Vasiljev et al. 2004). Incoming Tim23 is preferentiallybound by the homologous but dispensable Tim8-Tim13 chap-erone complex (Davis et al. 2000; Paschen et al. 2000). Thehydrophobic client proteins are thus transported throughthe aqueous intermembrane space (Figure 6). Their destina-tion in the inner membrane is the TIM22 insertase/translo-case complex (Sirrenberg et al. 1996; Kerscher et al. 1997)(Table 4). This complex has at its core a voltage-gated poreformed by the essential Tim22 protein (Kovermann et al.2002), which is required for its own membrane insertion(Sirrenberg et al. 1998).
Carrier proteins, or other substrates, associated with theIMS chaperones Tim9-Tim10 or Tim8-Tim12 bind to theTIM22 complex on the outer surface of the inner membrane.The substrates are dissociated from the chaperones, a pro-cess that may involve redox reactions (Curran et al. 2004).Uptake of the substrates into the TIM22 complex, transloca-tion of substrate domains through the membrane, and in-sertion of transmembrane domains into the membranerequire the Dc-membrane potential but not ATP (Rehlinget al. 2003; Peixoto et al. 2007). The mechanistic detailsof these reactions are not well understood.
Functional carrier protein dimers appear to self-assemblein the membrane rapidly following insertion (Dyall et al.2003). Tim22 is assembled into the TIM22 complex alongwith subunits that are inserted into the inner membrane bythe pathway described below for proteins with cleavablepresequences (Wagner et al. 2008).
Figure 6 Insertion of multi-spanning carrier proteins into the inner mem-brane. Newly synthesized multi-spanning carrier proteins, complexed withcytoplasmic Hsp70, are recognized by the Tom70 receptor subunit of theTOM complex. ATP-dependent release from cytoplasmic Hsp70 leads totranslocation through the TOM complex in a looped configuration andbinding to the Tim9-Tim10 IMS chaperone complex. The multi-spanningproteins are delivered to the TIM22 insertase complex in the inner mem-brane, released from Tim9-Tim10, and inserted into the inner membrane byreactions that depend upon the Dc potential across the inner membrane.
Mitochondrial Proteins 1213

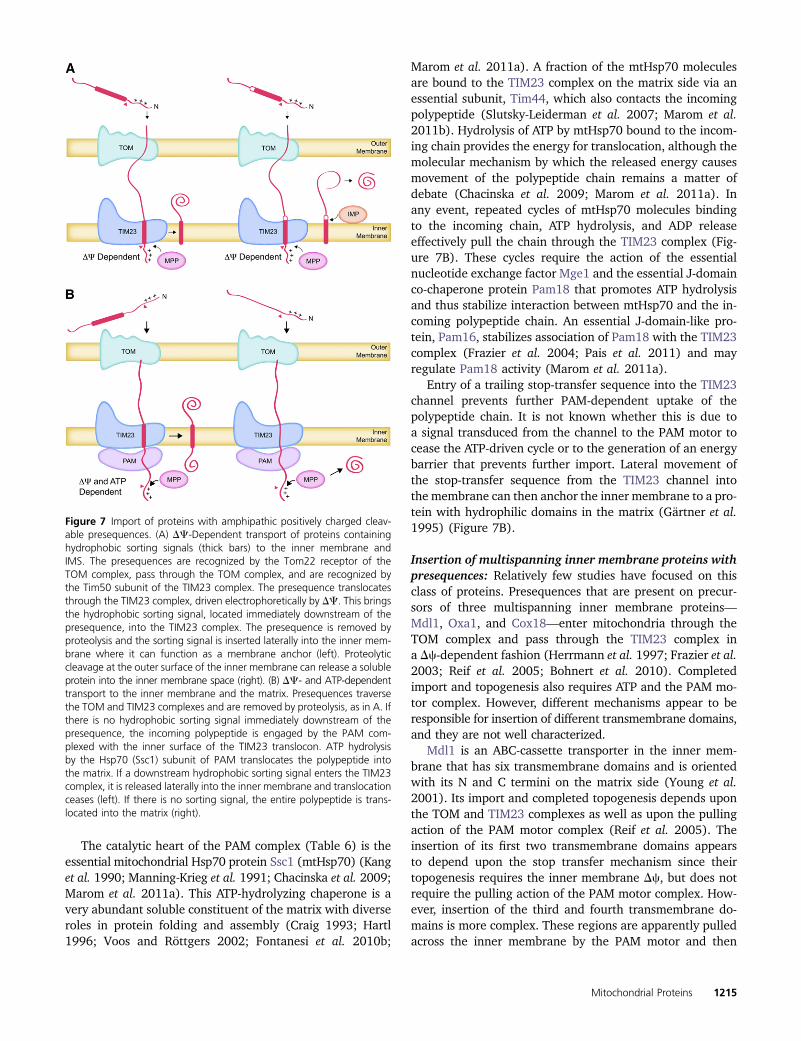
Insertion of inner membrane spanning proteins withcleavable presequences by the TIM23 insertase/translocasecomplex: Sixty percent or more of all yeast mitochondrialproteins are synthesized as precursors whose N termini arecleaved during import (Vögtle et al. 2009). The N-terminalpresequences typically contain targeting signals comprisingamphipathic a-helices with positively charged and hydro-phobic surfaces, although there is no consensus sequence.These “classical” targeting signals, which were reviewed inthe previous edition of YeastBook (Pon and Schatz 1991)and elsewhere (Neupert 1997; Voos et al. 1999), are suffi-cient on their own to target proteins to the matrix, as dis-cussed in the section below. However, these amphipathica-helices can also be combined with nearby downstreamhydrophobic sorting signals to form bipartite signals thatdirect proteins to the inner membrane (Figure 7A). Mia40is such a protein, anchored in the inner membrane by theN-terminal hydrophobic sorting signal with its hydrophilicdomains facing the IMS (Naoe et al. 2004; Neupert andHerrmann 2007). In addition, at least two well-studied IMSproteins, cytochrome b2 (Cyb2) and cytochrome c1 (Cyt1),adopt the same topology before being released from theirN-terminal membrane anchors by the inner membrane pro-tease (IMP: Imp1, Imp2, Som1) (Glick et al. 1992; Nunnariet al. 1993; Jan et al. 2000). In the case of Cyt1, a secondinternal sorting sequence near the C terminus inserts into theinner membrane by an unknown mechanism anchoring thehydrophilic N-terminal domain on the intermembrane spaceside (Arnold et al. 1998; Lange and Hunte 2002).
Different presequence-containing proteins are synthe-sized on free or mitochondria-bound polysomes (the pre-cursors of Mia40, Cyb2, and Cyt1 are all synthesized onmitochondria-bound polysomes) (Saint-Georges et al. 2008).There is relatively little information on the binding of cyto-plasmic chaperones to presequence-containing precursors.However, in at least some cases, cytoplasmic Hsp70 (Ssa1–4) is required for import. This is thought to reflect the abilityof Hsp70 to maintain precursors in partially unfolded states(Deshaies et al. 1988; Gautschi et al. 2001; Sass et al. 2003;Endo and Yamano 2010). While the pathways taken by theseprecursors to the outer surface of mitochondria are poorlyunderstood, their pathways into the organelle have beenthe subject of intensive research.
A domain of the TOM receptor subunit Tom20 on thecytoplasmic side of the outer membrane recognizes the hy-drophobic surfaces of presequence amphipathic a-helices(Abe et al. 2000; Yamamoto et al. 2011). The presequences
are in turn bound by the Tom22 receptor subunit via elec-trostatic interactions and directed into the pore formed byTom40 (Schmidt et al. 2010; Shiota et al. 2011). The Tom70subunit is not thought to play a major role in recognition ofpresequence-containing precursors. However, while yeastcells survive without either Tom70 or Tom20, deletion ofboth is lethal, indicating that they can carry out redundantfunctions (Ramage et al. 1993).
The IMS side of the TOM complex interacts transientlywith the major TIM complex, whose essential pore-formingsubunit is Tim23 (Chacinska et al. 2009; Mokranjac andNeupert 2009; Marom et al. 2011a). This TIM23 complex(Table 5) has an essential receptor, Tim50, that seals theTim23 pore in the absence of a substrate protein, preservingthe inner membrane potential (Meinecke et al. 2006).Tim50 recognizes presequences emerging from the TOMcomplex and facilitates their transit to the pore (Yamamotoet al. 2002; Mokranjac et al. 2009; Tamura et al. 2009;Marom et al. 2011b; Schulz et al. 2011) in a reaction thatmust occur at translocation contact sites between outer andinner membranes (Pon et al. 1989).
Passage of the presequence into and through the TIM23complex pore is electrophoretically driven by the innermembrane electrical potential, Dc, which is negative inside(Pon and Schatz 1991; Chacinska et al. 2009; Mokranjacand Neupert 2009; Marom et al. 2011a). It is independentof ATP hydrolysis (Glick et al. 1992). This transit of thepositively charged presequence through the TIM23 complexcan bring the downstream hydrophobic sorting signal intothe TIM23 pore if the distance between them is short (Fig-ure 7A). The presence of the hydrophobic sorting signal inthe TIM23 channel prevents further translocation of thepolypeptide chain. The presequence, now located in the ma-trix, is removed by sequence-specific activity of the solublemitochondrial processing protease (MPP) (Pon and Schatz1991; Taylor et al. 2001; Vögtle et al. 2009).
The “stop-transfer” activity of the sorting signal also trig-gers a lateral release of the polypeptide from the TIM23complex, resulting in its insertion into the lipid bilayer of theinner membrane (Neupert and Herrmann 2007; Chacinskaet al. 2009; Marom et al. 2011a) (Figure 7A). This lateralinsertion reaction can be reconstituted in vitro with purifiedTIM23 complex components in lipid vesicles containing themitochondria-specific lipid cardiolipin (van der Laan et al.2007). Once embedded in the membrane, the sorting signalfunctions as a membrane anchor that eventually sequestersthe rest of the polypeptide in the IMS after its passagethrough the TOM complex.
In the case of inner membrane proteins whose stop-transfer hydrophobic sorting signal is far downstream of thepositively charged presequence, Dc-dependent translocationof the presequence alone will not bring the sorting signalinto the TIM23 translocase. For such proteins, the interven-ing residues must be pulled into the matrix by the ATP-drivenpresequence translocase-associated motor (PAM) until thestop-transfer sequence enters the TIM23 channel (Figure 7B).
Table 4 Components of the TIM22 complex: insertion ofmultispanning carrier proteins into the inner membrane
Protein ORF Known function Null phenotype
Tim18 YOR297C Viable, various defectsTim22 YDL217C Core insertase of
the complexInviable
Tim54 YJL054W Viable, various defects
1214 T. D. Fox
The catalytic heart of the PAM complex (Table 6) is theessential mitochondrial Hsp70 protein Ssc1 (mtHsp70) (Kanget al. 1990; Manning-Krieg et al. 1991; Chacinska et al. 2009;Marom et al. 2011a). This ATP-hydrolyzing chaperone is avery abundant soluble constituent of the matrix with diverseroles in protein folding and assembly (Craig 1993; Hartl1996; Voos and Röttgers 2002; Fontanesi et al. 2010b;
Marom et al. 2011a). A fraction of the mtHsp70 moleculesare bound to the TIM23 complex on the matrix side via anessential subunit, Tim44, which also contacts the incomingpolypeptide (Slutsky-Leiderman et al. 2007; Marom et al.2011b). Hydrolysis of ATP by mtHsp70 bound to the incom-ing chain provides the energy for translocation, although themolecular mechanism by which the released energy causesmovement of the polypeptide chain remains a matter ofdebate (Chacinska et al. 2009; Marom et al. 2011a). Inany event, repeated cycles of mtHsp70 molecules bindingto the incoming chain, ATP hydrolysis, and ADP releaseeffectively pull the chain through the TIM23 complex (Fig-ure 7B). These cycles require the action of the essentialnucleotide exchange factor Mge1 and the essential J-domainco-chaperone protein Pam18 that promotes ATP hydrolysisand thus stabilize interaction between mtHsp70 and the in-coming polypeptide chain. An essential J-domain-like pro-tein, Pam16, stabilizes association of Pam18 with the TIM23complex (Frazier et al. 2004; Pais et al. 2011) and mayregulate Pam18 activity (Marom et al. 2011a).
Entry of a trailing stop-transfer sequence into the TIM23channel prevents further PAM-dependent uptake of thepolypeptide chain. It is not known whether this is due toa signal transduced from the channel to the PAM motor tocease the ATP-driven cycle or to the generation of an energybarrier that prevents further import. Lateral movement ofthe stop-transfer sequence from the TIM23 channel intothe membrane can then anchor the inner membrane to a pro-tein with hydrophilic domains in the matrix (Gärtner et al.1995) (Figure 7B).
Insertion of multispanning inner membrane proteins withpresequences: Relatively few studies have focused on thisclass of proteins. Presequences that are present on precur-sors of three multispanning inner membrane proteins—Mdl1, Oxa1, and Cox18—enter mitochondria through theTOM complex and pass through the TIM23 complex ina Dc-dependent fashion (Herrmann et al. 1997; Frazier et al.2003; Reif et al. 2005; Bohnert et al. 2010). Completedimport and topogenesis also requires ATP and the PAM mo-tor complex. However, different mechanisms appear to beresponsible for insertion of different transmembrane domains,and they are not well characterized.
Mdl1 is an ABC-cassette transporter in the inner mem-brane that has six transmembrane domains and is orientedwith its N and C termini on the matrix side (Young et al.2001). Its import and completed topogenesis depends uponthe TOM and TIM23 complexes as well as upon the pullingaction of the PAM motor complex (Reif et al. 2005). Theinsertion of its first two transmembrane domains appearsto depend upon the stop transfer mechanism since theirtopogenesis requires the inner membrane Dc, but does notrequire the pulling action of the PAM motor complex. How-ever, insertion of the third and fourth transmembrane do-mains is more complex. These regions are apparently pulledacross the inner membrane by the PAM motor and then
Figure 7 Import of proteins with amphipathic positively charged cleav-able presequences. (A) DC-Dependent transport of proteins containinghydrophobic sorting signals (thick bars) to the inner membrane andIMS. The presequences are recognized by the Tom22 receptor of theTOM complex, pass through the TOM complex, and are recognized bythe Tim50 subunit of the TIM23 complex. The presequence translocatesthrough the TIM23 complex, driven electrophoretically by DC. This bringsthe hydrophobic sorting signal, located immediately downstream of thepresequence, into the TIM23 complex. The presequence is removed byproteolysis and the sorting signal is inserted laterally into the inner mem-brane where it can function as a membrane anchor (left). Proteolyticcleavage at the outer surface of the inner membrane can release a solubleprotein into the inner membrane space (right). (B) DC- and ATP-dependenttransport to the inner membrane and the matrix. Presequences traversethe TOM and TIM23 complexes and are removed by proteolysis, as in A. Ifthere is no hydrophobic sorting signal immediately downstream of thepresequence, the incoming polypeptide is engaged by the PAM com-plexed with the inner surface of the TIM23 translocon. ATP hydrolysisby the Hsp70 (Ssc1) subunit of PAM translocates the polypeptide intothe matrix. If a downstream hydrophobic sorting signal enters the TIM23complex, it is released laterally into the inner membrane and translocationceases (left). If there is no sorting signal, the entire polypeptide is trans-located into the matrix (right).
Mitochondrial Proteins 1215

inserted into the membrane from the inside by the action ofOxa1 (Reif et al. 2005). Oxa1 is an inner membrane trans-locase/insertase known to export mitochondrially synthe-sized protein domains from the matrix (Bonnefoy et al.2009). Since Oxa1 is homologous and functionally similarto bacterial YidC proteins, the insertion of imported domainsback into the inner membrane from the inside is often re-ferred to as “conservative sorting.”
Oxa1 has five transmembrane domains and is orientedin the inner membrane with its N terminus in the IMS andits C terminus in the matrix (Bonnefoy et al. 2009). Duringits import, the first Oxa1 transmembrane domain appears tocross the inner membrane, following the presequence. Ina second step, the N-terminal domain is re-exported, concom-itant with insertion of the first transmembrane domain by thetranslocase activity of pre-existing Oxa1 itself (Herrmannet al. 1997). It is not clearly established whether the othertransmembrane domains are imported and then insertedfrom inside, transferred laterally into the membrane fromthe TIM23 complex, or perhaps inserted via some otherpathway (Herrmann et al. 1997). The bacterial homolog ofOxa1, YidC, can promote lateral insertion of transmembranedomains from the Sec translocase into the bilayer (Dalbeyand Kuhn 2004). This suggests the possibility that Oxa1could carry out an analogous function with some substrates,in conjunction with the TIM23 complex (Reif et al. 2005). Itis clear that Oxa1 cannot be absolutely required for its owntopogenesis since nonrespiring oxa1Δ mutants can be re-stored to normal phenotype by reintroduction of a wild-typeOXA1 gene (Bonnefoy et al. 1994).
Import of presequence-containing proteins to the matrix
A large fraction of presequence-containing precursors aretargeted to the innermost mitochondrial compartment, the
matrix. They contain amphipathic a-helices in their prese-quences but no stop-transfer sorting signals (Pon and Schatz1991). Following synthesis on either bound or free poly-somes, they traverse the TOM and TIM23 complexes asdescribed above. After Dc-dependent uptake of the prese-quence, ATP-dependent action of the PAM complex pulls theentire polypeptide into the matrix.
The pulling of entire proteins into the matrix by PAMdepends upon at least partial unfolding of the C-terminaldomains that are often still on the cytoplasmic side of theTOM complex when PAM engages the N-terminal end. Thishas been clearly demonstrated in in vitro reactions (Pon andSchatz 1991). The importance of this in vivo is demonstratedby the import of Fum1, the precursor of fumarase (Sass et al.2003; Karniely et al. 2006). Wild-type Fum1 has the abilityto fold rapidly into a stable conformation while the prese-quence is imported into the matrix and processed. Moleculesthat achieve this state fail to import and are released backinto the cytoplasm in mature form by retrograde movementof their N termini. On the other hand, molecules whose C-terminal domains do not fold rapidly are pulled into thematrix. This is one of several mechanisms by which proteinscan be localized both in mitochondria and in the cytoplasm(Yogev and Pines 2011).
Presequences of imported precursors are typically removedby the soluble MPP (Pon and Schatz 1991; Taylor et al. 2001).Many matrix proteins are further processed at their N terminiby removal of a single residue or eight residues by the pro-teases Icp55 and Oct1, respectively (Vögtle et al. 2009, 2011).These alterations apparently serve to generate mature productswith increased stability, following the bacterial N-end rules.
The folding of imported matrix proteins must be largelycoupled to their interaction with, and release from, mtHsp70associated with the PAM motor. Mitochondria also contain
Table 5 Components of the TIM23 complex: transport of polypeptides through the inner membrane and lateralinsertion of membrane anchors into the inner membrane
Protein ORF Known function Null phenotype
Tim17 YJL143W InviableTim21 YGR033C Viable various defectsTim23 YNR017W Pore formation InviableTim44 YIL022W Tethers PAM to TIM23 complex InviableTim50 YPL063W Substrate receptor and pore gating Inviable
Table 6 Components of the PAM complex: ATP-dependent pulling of proteins through the TIM23 complex intothe matrix
Protein ORF Known function Null phenotype
Mdj2 YNL328C Viable, various defectsMge1 YOR232W Nucleotide release factor for Ssc1 InviablePam16 YJL104W InviablePam17 YKR065C Interaction between TIM23 and PAM complexes ViablePam18 YLR008C J protein co-chaperone for Ssc1 InviableSsc1 YJR045C Hsp70; ATP hydrolysis drives import of substrates into matrix Inviable
1216 T. D. Fox

the essential chaperonin complex Hsp60-Hsp10 (Pon andSchatz 1991; Voos and Röttgers 2002), which is characteristicof bacteria and energy-transducing organelles. These chaper-ones, together with the dispensable Hsp78 and Pim1, homo-logs of bacterial ClpB and lon ATP-dependent proteases, assistin folding and maintenance of imported matrix proteins(Leonhardt et al. 1993; Suzuki et al. 1997; Bender et al.2011).
Spatial distributions and regulation of import complexes
The import of presequence-containing proteins to the innermembrane and matrix requires at least transient interactionof some TOM complexes and TIM23 complexes at translo-cation contact sites between the membranes. By incubatingisolated mitochondria with saturating amounts of an artifi-cial presequence-containing protein that become trappedboth outside the outer membrane and in the inner mem-brane, TOM complexes bound to TIM23 complexes can bedetected. Using such a trap, it could be shown that all of theTIM23 complexes were associated with �25% of the TOMcomplexes (Dekker et al. 1997). Thus, at least under theseconditions, the TIM23 complex must be located almost ex-clusively in the boundary domain of the inner membraneand form translocation contact sites with the outer mem-brane TOM complexes. However, in respiring wild-typecells, Tim23 appears to be only moderately enriched in theinner membrane boundary domain relative to cristae (Vogelet al. 2006).
The TIM23 complex must respond to signals in the sub-strate precursors that direct either their lateral insertion intothe inner membrane or their complete translocation into thematrix. There is currently some dispute about whether thelateral insertion and matrix translocation functions are car-ried out by a single multi-functional TIM23-PAM complexwith different conformations (Popov-�Celeketić et al. 2008a,2011; Mokranjac and Neupert 2009) or by two forms of theTIM23 complex in dynamic equilibrium with each other(Chacinska et al. 2009, 2010; Schmidt et al. 2010). Twoforms of static TIM23 complex were detected in solubilizedextracts of mitochondria, depending upon whether theywere trapped importing a precursor targeted for sorting tothe outer surface of the inner membrane, which does notrequire the ATP-driven PAM motor, or for import into thematrix, which does require the motor (Chacinska et al. 2005,2010). The isolated TIM23-sorting complexes contained theprotein Tim21, but not PAM complex subunits. In contrast,the isolated TIM23 motor complexes contained PAM subu-nits but only low levels of Tim21. It has been suggested thatthe PAM motor could associate with the TIM23 channel ifand when Tim21 is ejected preceding import into the matrix(Wiedemann et al. 2007).
Surprisingly, the TIM23 complex has been found to asso-ciate with proton-pumping supercomplexes of the bc1 com-plex and cytochrome c oxidase. These interactions are facilitatedby the Tim21 subunit (van der Laan et al. 2006), which alsohas a role in connecting the TOM complex to the TIM23
complex (Chacinska et al. 2005; Mokranjac et al. 2005),and by Pam16 and Pam18 (Wiedemann et al. 2007). Thephysiological relevance of this association is suggested bythe observation that Tim21 accelerates Dc-dependent pre-cursor translocation only when the respiratory complexesare active (van der Laan et al. 2006). Thus, it appears thatlocal increases in Dc may affect the rate of presequencetranslocation through TIM23. Active TIM23 complexes mustbe located in the boundary domain of the inner membrane,while the respiratory complexes are selectively, but presum-ably not completely, located in the cristae domains (Vogelet al. 2006; Wurm and Jakobs 2006; Rabl et al. 2009; Zicket al. 2009; Davies et al. 2011).
A complex of inner membrane proteins present at thecrista junctions, and required to form them, has been iden-tified in several labs and named MINOS, MitOS, and MICOS(Harner et al. 2011a; Hoppins et al. 2011; von der Malsburget al. 2011; Alkhaja et al. 2012). A core component of thiscomplex, Fcj1, was previously shown to be required forcrista junction formation (Rabl et al. 2009). The crista junc-tions are located at firm contact sites between the outer andinner membranes (Harner et al. 2011a). These firm contactsites apparently do not correspond to translocation contactsites between TOM and TIM23 complexes since TIM23-dependent precursor translocation is not directly affectedby the absence of Fcj1 (von der Malsburg et al. 2011). How-ever, Fcj1 interacts with Mia40 and with TOM complexes notassociated with TIM23, facilitating Mia40-dependent importof soluble twin-Cys proteins to the IMS (von der Malsburget al. 2011). This suggests that import of substrates depen-dent upon the Mia40-Erv1 disulfide relay system is selectivelylocated near crista junctions. Cells lacking Fcj1 or other com-ponents of MINOS/MitOS/MICOS have respiratory defects,but are viable. Thus, this complex does not appear to be ab-solutely required for any essential import pathway.
Is the mitochondrial protein import machinery simplya conduit for any targeted protein that arrives at the outermembrane, or is its activity per se subject to modulation?Recent evidence indicates that TOM complex activity isregulated by phosphorylation of the Tom70 receptor bycAMP-dependent protein kinase A (PKA) (Schmidt et al.2011). PKA activity is controlled such that its activity ishigh in the presence of glucose but low in nonfermentablecarbon sources (Zaman et al. 2008). In the presence ofglucose, PKA phosphorylates Tom40 residue Ser174(among a wide variety of targets). This modificationdecreases the ability of Tom40 to interact with cytoplasmicHsp70, the chaperone that delivers metabolite carrier pro-teins to the TOM complex (Young et al. 2003), decreasingthe efficiency with which carrier proteins are imported tothe inner membrane (Schmidt et al. 2011). Constitutiveactivation of PKA results in decreased levels of carrier pro-teins relative to mitochondrial proteins whose import isnot affected by decreased Tom70 activity. Thus, this regu-latory pathway has significant physiological effects onmitochondria.
Mitochondrial Proteins 1217

Interestingly, another kinase, casein kinase 2, quantita-tively phosphorylates two Ser residues of the central TOMreceptor Tom22 and two residues of Mim1 (Schmidt et al.2011). These modifications are required for normal assem-bly and activity of both proteins, and thus for the activityof the TOM complex. It is not clear whether these phospho-rylations are modulated in response to environmentalconditions.
Assembly of Complexes Containing MitochondriallySynthesized Proteins
Mitochondrial gene expression in S. cerevisiae is required foractive respiratory complexes located in the inner membrane,but is not required for any other physiological processes(Pon and Schatz 1991; Chacinska and Boguta 2000; Lipinskiet al. 2010). The organellar genetic system produces onlyeight major proteins in S. cerevisiae. Seven are hydrophobicintegral inner membrane proteins: Cob (Apo-cytochrome b),a subunit of the proton-pumping bc1 complex (respiratorycomplex III); Cox1, Cox2, and Cox3, subunits of proton-pumping cytochrome c oxidase (respiratory complex IV);and Atp6, Atp8, and Atp9, subunits of the F0 componentof ATP synthase (respiratory complex V). These mitochond-rially synthesized subunits are assembled with importedsubunits to form the active enzymes. (S. cerevisiae doesnot have respiratory complex I, the proton-pumping NADHdehydrogenase found in most other eukaryotes. Succinatedehydrogenase, respiratory complex II, does not contain mi-tochondrial gene products.)
The eighth major mitochondrial gene product, Var1, isa small mitochondrial ribosomal subunit protein. Aside fromVar1, all proteins known to be required for yeast mitochon-drial gene expression are imported from the cytoplasm [withthe exception of some minor proteins encoded by mitochon-drial introns that facilitate intron splicing (Pon and Schatz1991; Saldanha et al. 1993)]. Var1 is assembled inside theorganelle with mitochondrially coded 15S rRNA and at least33 imported ribosomal proteins coded by nuclear genes(Groot et al. 1979; Terpstra et al. 1979; Gan et al. 2002;Williams et al. 2005). The large mitochondrial ribosomalsubunit is assembled from the mitochondrially coded 21SrRNA and at least 44 imported proteins (Gan et al. 2002).These organellar ribosomes do not share any known compo-nents with cytoplasmic ribosomes.
The assembly of yeast cytochrome c oxidase has beenstudied and reviewed extensively (Barrientos et al. 2002a,2009; Carr and Winge 2003; Herrmann and Funes 2005;Khalimonchuk and Rodel 2005; Cobine et al. 2006; Fontanesiet al. 2006, 2008; Horn and Barrientos 2008; Mick et al.2011; Soto et al. 2011). A striking fact to emerge from thesestudies is the specific involvement of .30 nuclear geneproducts that are not constituents of cytochrome c oxidase.They are required to produce the active enzyme by promot-ing specific reactions in the expression of genes; insertion ofproteins into the inner membrane; insertion of heme, cop-
per, zinc, and magnesium; and chaperoning assembly inter-mediates (Soto et al. 2011). The assembly of the cytochromebc1 complex (Zara et al. 2009; Conte et al. 2011; Wageneret al. 2011; Smith et al. 2012) and the ATP synthase (Ackermanand Tzagoloff 2005; Rak et al. 2009) also requires similarassistance. This appears to contrast with the assembly of theTOM complex, for example. Its constituents are inserted intothe outer membrane by general mechanisms and are be-lieved to self-assemble once in the membrane (Beckeret al. 2008, 2010). Furthermore, as discussed below, themitochondrial synthesis of several respiratory complex sub-units is coupled to assembly of those complexes.
At a higher level of assembly, the cytochrome bc1 complexand cytochrome c oxidase are organized together into super-complexes that appear to facilitate efficient electron trans-port between them via cytochrome c (Heinemeyer et al.2007; Stuart 2008), although this rationale has been ques-tioned (Trouillard et al. 2011). Assembly of these supercom-plexes has recently been shown to depend upon twosupercomplex subunits, Rcf1 and Rcf2 (Chen et al. 2012;Strogolova et al. 2012; Vukotic et al. 2012).
Mitochondrial protein synthesis is membrane bound
Components of the mitochondrial genetic system have beenidentified chiefly through the isolation of mutants and thestudy of their phenotypes and of the proteins affected bytheir mutations (Tzagoloff and Dieckmann 1990). Whilemitochondrial protein synthesis can be studied in isolatedintact organelles, no true in vitro translation system hasbeen developed from yeast mitochondria. It appears thatthe mitochondrial genetic system is largely organized onthe two-dimensional inner surface of the inner membrane,and an intact membrane [although not the membrane DC
(Clarkson and Poyton 1989; He and Fox 1997)] may berequired for protein synthesis. Indeed, the lipid compositionof the inner membrane appears to affect mitochondrial pro-tein synthesis (Marzuki and Hibbs 1986; Ostrander et al.2001).
Mitochondrial ribosomes are bound to the inner mem-brane, preferentially to the cristae domains, apparently tofacilitate cotranslational protein insertion (Vogel et al. 2006;Ott and Herrmann 2010). To date, three inner membraneproteins are known to interact with large mitochondrial ri-bosomal subunits and are thought to tether them to themembrane independently of nascent chains: they are Oxa1(Jia et al. 2003, 2009; Szyrach et al. 2003), Mba1 (Ott et al.2006; Gruschke et al. 2010), and Mdm38 (Frazier et al.2006; Lupo et al. 2011). Oxa1 functions as a translocase/insertase-promoting export of mitochondrially coded hydro-philic domains, particularly those of Cox2, to the intermem-brane space and insertion of transmembrane domains (Heand Fox 1997; Hell et al. 1997, 2001). MbaI, a peripheralmembrane protein, appears to be partially redundant infunction with Oxa1 (Preuss et al. 2001; Ott et al. 2006).Similarly, Mdm38 promotes export of Cob and Atp6 domains(Frazier et al. 2006). Interestingly, Mdm38 is a bifunctional
1218 T. D. Fox

protein that also participates in K+/H+ exchange across theinner membrane (Nowikovsky et al. 2004; Froschauer et al.2005; Zotova et al. 2010; Lupo et al. 2011).
The synthesis of mitochondrially coded proteins at theinner membrane surface appears to limit their contact withmatrix chaperones. Membrane proteins emerging from mito-chondrial ribosomes do not interact with mtHsp70 (Ssc1)(Ott et al. 2006). Furthermore, soluble reporter proteinssynthesized inside mitochondria appear to fold more slowlythan identical polypeptides synthesized in the cytoplasmand imported into the matrix via the mtHsp70-dependentTIM23/PAM machinery (Demlow and Fox 2003).
Channeling of mRNAs to the inner membrane
Mitochondrial DNA molecules are present in nucleoproteinaggregates, termed “nucleoids” (Meeusen et al. 1999; Kaufmanet al. 2000; Kucej et al. 2008). A fraction of the �40 nucle-oids per cell are tethered to the inner and outer membranesby a membrane-spanning protein complex (Aiken Hobbset al. 2001; Boldogh et al. 2003; Meeusen and Nunnari2003; Chen and Butow 2005). While the distribution ofthe mitochondrial RNA polymerase, Rpo41, among thenucleoids has not been reported, transcription likely occursadjacent to the inner membrane. Furthermore, strong evi-dence suggests that mRNAs are channeled from RNA poly-merase to the membrane-bound translation apparatus.Rpo41 interacts through its amino terminal domain withthe soluble protein Mtf2 (Nam1), which in turn appears tochaperone processing of primary transcripts and delivery ofmRNAs to the inner membrane surface, together with themembrane-associated proteins Sls1 and Rmd9 (Dieckmannand Staples 1994; Wallis et al. 1994; Rouillard et al. 1996;Rodeheffer et al. 2001; Bryan et al. 2002; Rodeheffer andShadel 2003; Nouet et al. 2007;Williams et al. 2007) (Figure 8).Mtf2 (Nam1), Sls1, and Rmd9 are all required for normalmitochondrial translation.
The physiological importance of mitochondrial mRNAchanneling from synthesis to translation is most clearlydemonstrated by the fact that mutations in the mitochon-drial RNA polymerase amino terminal domain that preventinteraction with Mtf2 (Nam1) severely reduce translationbut do not prevent mRNA synthesis and accumulation(Rodeheffer and Shadel 2003). Thus, new primary tran-scripts appear to be directly transferred to Mtf2 (Nam1)from RNA polymerase for efficient gene expression, in con-junction with the interacting membrane protein Sls1 andperhaps Rmd9 among other factors. mRNAs do not, appar-ently, diffuse freely through the matrix.
Localization of protein synthesis by mRNA-specifictranslational activators
A peculiar feature of mitochondrial translation in S. cerevi-siae and other ascomycetes is its dependence on transla-tional activators for individual mRNAs (Fox 1996a; Coffinet al. 1997; Costanzo et al. 2000; Towpik 2005; Kühl et al.2011) (Table 7). The roles of mRNA-specific translational
activators have been most thoroughly explored for theexpression of the S. cerevisiae mitochondrial genes encodingthe three core cytochrome c oxidase subunits and apo-cytochrome b. The best-studied translational activators aremembrane-bound proteins that recognize targets in the mi-tochondrial mRNA 59-UTRs and interact with mitochondrialribosomes, apparently promoting translation initiation (Fox1996a; Rödel 1997; Towpik 2005). The translational activa-tors for the COX1, COX2, and COX3 mRNAs are presentat low levels that are rate limiting for gene expression(Fox 1996b; Steele et al. 1996; Green-Willms et al. 2001;Naithani et al. 2003; Perez-Martinez et al. 2009). Upsettingthe balance of translational activators can interfere withcytochrome c oxidase biogenesis (Fiori et al. 2005).
Mtf2 (Nam1) interacts physically with the activator Pet309,a pentatricopeptide repeat (PPR) protein that binds both theCOX1 mRNA 59-UTR and mitochondrial ribosomes (Mantheyand Mcewen 1995; Naithani et al. 2003; Tavares-Carreon et al.2008; F. Tavares-Carreon, A. Zamudio-Ochoa, Y. Camacho-Villasana, and X. Perez-Martinez; personal communica-tion). PPR motifs have been identified in many organellarRNA-binding proteins with sequence-specific functions(Delannoy et al. 2007) and have been identified in eightmRNA-specific translational activators as well as Rmd9and the Rpo41 amino terminal domain (Lipinski et al.2011). Thus, it is likely that the COX1 mRNA is transferredfrom the RNA polymerase Rpo41 to Mtf2 (Nam1)/Sls1and Rmd9 to Pet309 for translation by the ribosome. In-deed, Pet309 and at least two ribosomal proteins copurifywith Rpo41 (Markov et al. 2009). Mtf2 (Nam1) also inter-acts with the PPR-containing COX2 mRNA-specific activa-tor Pet111, and with Pet494, which activates COX3 mRNAtranslation, suggesting a similar pathway for those mRNAs(Naithani et al. 2003; Lipinski et al. 2011) (Figure 8).
The translational activators promoting synthesis of cyto-chrome c oxidase subunits have been shown to interact with eachother (Brown et al. 1994; Naithani et al. 2003). Furthermore,a large protein complex can be isolated from mitochondria
Figure 8 Channeling of mitochondrially coded mRNAs from RNA poly-merase (Rpo41) to membrane-bound ribosomes by Sls1, Mtf2, Rmd9,and mRNA-specific translational activators (TA). (The figure is not in-tended to suggest that mRNAs are translated while still emerging fromRNA polymerase.)
Mitochondrial Proteins 1219

that contains Pet309 and the COB mRNA-specific activatorCbp1, as well as several unidentified proteins (Krause et al.2004). These findings suggest that clusters of translationalactivators promote colocalized synthesis of mitochondriallycoded respiratory complex subunits at distinct locations.Based on the levels of translational activator proteins, thereappear to be at most a few hundred such sites per cell(Marykwas and Fox 1989; Fox 1996b; Ghaemmaghamiet al. 2003; Naithani et al. 2003). It seems likely that thesesites are adjacent to nucleoids.
The physiological significance of distinct sites or topol-ogies for mitochondrial translation is supported by experi-ments in which Cox2 was translated in vivo froma chimeric mRNA with the 59- and 39-UTRs of the VAR1mRNA, which normally flank the ORF encoding a ribo-somal protein. While the rate of Cox2 synthesis was nor-mal, Cox2 accumulation and cytochrome c oxidase activitywere severely reduced (Sanchirico et al. 1998). The puta-tive translational activator for the VAR1 mRNA is Sov1,a membrane-bound PPR protein (Sanchirico 1998; Lipin-ski et al. 2011; M. E. Sanchirico, A. R. Wiffen, T. D. Fox,and T. L. Mason; unpublished results), and translation ofthe chimeric mRNA appears to be membrane bound (Fioriet al. 2003). However, the VAR1 mRNA 59-UTR apparentlymislocalizes or misorients Cox2 synthesis on the innermembrane such that Cox2 cannot be efficiently assembledwith other cytochrome c oxidase subunits and is largelydegraded.
Interestingly, bacteria have also recently been found toemploy signals embedded in mRNA sequences to localizetranslation of membrane proteins (Nevo-Dinur et al. 2011).
It has been suggested that localization of mitochondria-bound cytoplasmic protein synthesis and import could becoordinated with localization of mitochondrial protein syn-thesis to promote assembly of complexes containing bothkinds of proteins (Garcia et al. 2007a). There is at present noevidence directly bearing on this interesting hypothesis.
Assembly of cytochrome c oxidase
The three mitochondrially coded subunits of yeast cyto-chrome c oxidase form its catalytic core (Tsukihara et al.1996). They are surrounded by eight subunits importedfrom the cytoplasm (Taanman and Capaldi 1992; Sotoet al. 2011). The order in which these subunits are assem-
bled is not known, despite efforts to analyze subassembliesdetected in mutants (Church et al. 2005; Horan et al. 2005).However, assembly is believed to be nucleated by mitochondrialsynthesis of the core subunits, in particular Cox1 (Herrmannand Funes 2005; Fontanesi et al. 2008; Mick et al. 2011; Sotoet al. 2011)
Cox1 is highly hydrophobic, with 12 transmembranedomains, and both termini are on the inside of the innermembrane. In the assembled enzyme Cox1 contains twoheme A moieties and a copper atom that participate in elec-tron transport (Tsukihara et al. 1995). Cox1 is believed to becotranslationally inserted into the inner membrane by Oxa1(Hell et al. 2001).
The synthesis of Cox1 is coupled to the formation of earlyassembly intermediates and is downregulated if assembly isartificially blocked by a variety of mutations (Barrientos et al.2004; Shingu-Vazquez et al. 2010). This assembly-feedbackregulation may prevent overproduction of Cox1 to protectyeast cells from damage due to pro-oxidant activity ofunassembled Cox1 (Khalimonchuk et al. 2007). Mss51, a mi-tochondrial inner membrane protein with at least two func-tions, is a key factor in this regulatory circuit. Mss51 activatesCOX1 mRNA translation via its 59-UTR, in conjunction withPet309, and also binds to newly synthesized Cox1 (Decosteret al. 1990; Perez-Martinez et al. 2003, 2009; Zambranoet al. 2007) (Figure 9).
Newly synthesized Cox1 nucleates the formation of anearly pre-assembly complex with Mss51 and two short mem-brane proteins, Cox14 and Coa3 (Cox25) (Perez-Martinezet al. 2009; Fontanesi et al. 2010a; Mick et al. 2010). Inaddition, whether complexed with Cox1 or not, Mss51 isstoichiometrically associated with mtHsp70 (Ssc1) (Fontanesiet al. 2010b) (Figure 9). Both cox14 and coa3 deletions pre-vent association of Mss51 with Cox1 and allow uncontrolledCox1 synthesis despite the fact that they prevent cytochromec oxidase assembly (Perez-Martinez et al. 2009; Fontanesiet al. 2010a; Mick et al. 2010). Thus, sequestration of Mss51in assembly intermediates containing Cox1 appears to pre-vent it from activating COX1 mRNA translation. Truncationof the C-terminal 11 residues of Cox1, normally exposedin the matrix, destabilizes this pre-assembly complex anddisrupts assembly-feedback regulation, but does not pre-vent cytochrome c oxidase assembly (Shingu-Vazquez et al.2010).
Table 7 Proteins that activate translation of specific mitochondrially coded mRNAs by various mechanisms
Mitochondrial mRNA Activator proteins Activator protein ORFs
ATP8-ATP6 Aep3, Assembled F1 ATPase YPL005W,YBL099W,YJR121W,YBR039W,YPL271WATP6 Atp22 YDR350CATP9 Aep1, Aep2, Atp25 YMR064W, YMR282C, YMR098CCOB Cbp1, Cbp3, Cbp6, Cbs1, Cbs2 YJL209W, YPL215W, YBR120C, YDL069C, YDR197WCOX1 Pet54, Pet309, Mss51 YGR222W, YLR067C, YLR203CCOX2 Pet111 YMR257CCOX3 Pet54, Pet122, Pet494 YGR222W, YER153C, YNR045WVAR1 Sov1 YMR066W
1220 T. D. Fox

Assembly of Cox1 proceeds by the addition of the assemblyfactor Coa1 to the pre-assembly complex nucleated by Cox1,which in turn allows the addition of Shy1 (Mick et al. 2007,2010; Pierrel et al. 2007; Khalimonchuk et al. 2010). NeitherCoa1 nor Shy1 are completely essential for production of ac-tive cytochrome c oxidase [both are virtually dispensable in theD273-10B strain background (L. S. Burwell, Z. W. Via, and T.D. Fox, unpublished results)]. Mss51 is thought to be releasedfrom assembly intermediates in a step after Shy1 association,which may involve the addition of the imported cytochrome coxidase subunits Cox5 (encoded by COX5A and COX5B) andCox6 (Barrientos et al. 2002b; Mick et al. 2007; Shingu-Vaz-quez et al. 2010), although some evidence indicates that thisoccurs farther upstream (Khalimonchuk et al. 2010). The re-leased Mss51 becomes free to activate another round of Cox1synthesis, completing the regulatory circuit (Figure 9).
Cox1 assembly proceeds by the addition of redox cofac-tors. Shy1, together with Coa2, appears to promote the in-sertion of heme A into Cox1 (Pierrel et al. 2008; Bestwicket al. 2010). The copper-binding membrane protein Cox11receives copper from the soluble shuttle protein Cox17 andinserts it into assembling Cox1 from the IMS side (Beerset al. 1997; Banting and Glerum 2006; Cobine et al. 2006;Khalimonchuk et al. 2010). The subunits Cox5 and Cox6,which directly contact Cox1, are believed to be associated atthis stage with Cox1, Cox14, Shy1, Coa1, and Coa3 in a sub-assembly that awaits the addition of mitochondrially synthe-sized Cox2 and Cox3 (Horan et al. 2005; Fontanesi et al.2010a; Khalimonchuk et al. 2010; Mick et al. 2010).
Cox2 has two transmembrane domains and acidic N- andC-terminal domains that are exported to the IMS side of theinner membrane prior to assembly (Tsukihara et al. 1996).The C-terminal domain binds two copper atoms. In S. cer-evisiae, Cox2 is synthesized as a precursor with a 15-residueleader peptide (Pratje et al. 1983) whose length and se-quence can be altered dramatically without destroying func-tion (Bonnefoy et al. 2001). However, the mRNA sequenceencoding the leader peptide contains a positive element thatis required to antagonize negative elements downstream inthe mRNA that otherwise prevent completed translation(Bonnefoy et al. 2001; Williams and Fox 2003). Taken to-gether with the functional interactions of these elementswith mitochondrial ribosomes (Williams et al. 2004, 2005;Prestele et al. 2009), these findings suggest the possibilitythat regulation of translation elongation could be coupled tosteps in membrane topogenesis of pre-Cox2.
The pre-Cox2 N-terminal domain is cotranslationally ex-ported by Oxa1 (He and Fox 1997; Hell et al. 1998; Bonnefoyet al. 2009). The leader peptide is rapidly processed in theIMS by the inner membrane protease in a reaction chaper-oned by Cox20, an inner membrane protein whose topologyresembles that of Cox2 (Nunnari et al. 1993; Hell et al.2000; Jan et al. 2000).
Export of the acidic Cox2 C-terminal domain also requiresOxa1, but appears to be post-translational (He and Fox1997; Fiumera et al. 2007). Its export also depends specif-ically upon another highly conserved inner membrane trans-locase, Cox18, which is paralogously related to Oxa1, andon two inner membrane proteins, Mss2 and Pnt1, whichinteract with Cox18 (He and Fox 1999; Broadley et al.2001; Saracco and Fox 2002; Funes et al. 2004). TheCox20 chaperone also functions in C-tail export, probablyby removing the exported protein from Cox18 (Fiumeraet al. 2009; Elliott et al. 2012). Mature but unassembledCox2 remains associated with Cox20 (Hell et al. 2000).Metallation of the Cox2 C-terminal domain is catalyzed bymembrane-bound Sco1, which receives copper from the sol-uble Cox17 shuttle and then directly interacts with Cox2(Lode et al. 2000; Cobine et al. 2006; Rigby et al. 2008).Metallation is believed to precede Cox2 addition to the Cox1subassembly complex (Soto et al. 2011).
Insertion of the seven transmembrane domains of Cox3,whose amino terminal residues remain on the matrix side, isassisted by Oxa1 (Tsukihara et al. 1996; Hell et al. 2001).There are no known specific chaperones involved in furtherassembly of Cox3 into the catalytic core of cytochrome coxidase. Nor are there data on the addition of importedsubunits to form the active enzyme.
Assembly of the bc1 complex
The catalytic core of the bc1 complex comprises three sub-units. Cob (cytochrome b) is mitochondrially synthesizedand embedded in the membrane with its two noncovalentlybound heme B moieties. The imported subunits Cyt1 (cyto-chrome c1) and Rip1 (Rieske 2Fe-2S protein) are anchoredto the membrane with their hydrophilic domains bearingheme C and 2Fe-2S cluster redox cofactors, respectively, inthe IMS (Solmaz and Hunte 2008; Zara et al. 2009; Smithet al. 2012). Seven additional imported proteins that do notdirectly participate in electron transfer complete the bc1 com-plex structure, which forms a dimer. Analysis of subassembliesdetected in mutants indicates that Cob may initially associate
Figure 9 Assembly feedback control of Cox1 synthesis byMss51 activities as a translational activator and assembly fac-tor. Mitochondria translation of the COX1 mRNA is activatedby the mRNA-specific activators Pet309 and Mss51 [Mss51 isassociated with Hsp70 (Ssc1) throughout]. Synthesis of a newCox1 polypeptide nucleates an early assembly intermediatecontaining Mss51 and the assembly factors Cox14 and Coa3.As additional assembly factors associate with newly synthe-sized Cox1, Mss51 is released from the assembly intermedi-ates and is then available to initiate additional Cox1 synthesis.
Mitochondrial Proteins 1221

with two imported subunits, Qcr7 and Qcr8, while Cyt1 asso-ciates with Cor1 and Cor2 (Crivellone et al. 1988; Zara et al.2004, 2007). Association of these subcomplexes, facilitated bythe assembly factor Bca1 (Mathieu et al. 2011), forms a nearlycomplete species that subsequently accepts Rip1.
Cob has eight transmembrane domains, and both terminiare on the inside of the inner membrane. Its two heme Bmoieties are inserted by an unknown mechanism. Oxa1 par-ticipates in its membrane insertion (Hell et al. 2001). Trans-lation of the COB mRNA is specifically activated through its59-UTR by Cbs1, Cbs2, and Cbp1 (Rödel 1997; Islas-Osunaet al. 2002). In addition, synthesis of Cob is promoted bytwo proteins, Cbp3 and Cbp6, that bind to mitochondrialribosomes, associate with newly synthesized Cob after re-lease from the ribosome, and together with Cob form a pre-assembly complex with the assembly factor Cbp4 (Gruschkeet al. 2011). While this mechanism couples translation toassembly, Cob synthesis is not reduced by a feedback mech-anism when assembly of the bc1 complex is disrupted.
The complex import pathway of Cyt1, resulting in a hy-drophilic N-terminal domain in the IMS with a C-tail anchor,was described above. Covalent attachment of heme to thishydrophilic domain is catalyzed by the lyase Cyt2 eitherduring or after maturation of Cyt1 (Bernard et al. 2003).
The maturation of Rip1 is particularly interesting. Thisprotein is initially fully imported into the matrix (Hartlet al. 1986). Since Fe-S cluster biogenesis occurs in the ma-trix, it is believed that assembly of the Rip1 polypeptide withits 2Fe-2S cluster occurs after this initial import reaction(Kispal et al. 1999; Lill 2009; Wagener et al. 2011). In anyevent, the C-terminal domain of Rip1 is subsequently ex-ported through the inner membrane to the IMS. This exportreaction specifically requires Bcs1, a member of the AAAfamily of ATPases that is tethered to the inner surface of theinner membrane (Nobrega et al. 1992; Wagener et al. 2011).Most mitochondrial AAA proteins are hexameric complexeswith a central compartment capable of ATP-dependent un-folding, proteolysis, and chaperone-like activity (Fiumeraet al. 2009; Tatsuta and Langer 2009; Truscott et al.2010). In the case of Bcs1, it appears that an enlarged cen-tral compartment translocates the folded C-terminal domainof Rip1 bearing its 2Fe-2S cluster across the membrane andreleases the Rip1 N-terminal transmembrane domain later-ally into the membrane (Wagener et al. 2011). The matrixprotein Mzm1 facilitates this export of Rip1 from the matrix(Atkinson et al. 2010, 2011), which must occur prior to itsassembly into the bc1 complex.
Assembly of the ATP synthase
The raison d’être of the respiratory chain is the synthesis ofATP. This reaction is carried out by the well-known F1F0 ATPsynthase. The F0 sector in the inner membrane capturesenergy of the proton gradient as rotary motion and turnsthe central stalk of the F1 sector, which projects into thematrix. Rotation of the central stalk relative to the catalyticsites in the Atp13-Atp23 (a3b3) hexamer of F1, which is
fastened to nonrotating proteins in F0 by the peripheral stalkor stator, drives the synthesis of ATP from ADP and inorganicphosphate (Stock et al. 1999; von Ballmoos et al. 2009). InS. cerevisiae, all subunits of F1 and of the stator are imported(Velours and Arselin 2000; Ackerman and Tzagoloff 2005).Three hydrophobic core proteins of F0,—Atp6, Atp8, andAtp9—are mitochondrial gene products, while two othersare imported. Current evidence indicates that the completeATP synthase is assembled from modular subassemblies,such that the proton-conducting pore of F0, formed by theinterface between Atp6 and Atp9 (Fillingame and Dmitriev2002), is formed at the last step (Rak et al. 2009, 2011)(Figure 10). This hypothesis predicts that the proton poreis immediately coupled to the catalytic machinery at themoment that it is formed, avoiding the deleterious effectsof unregulated proton leakage across the inner membrane.
One pre-assembly module is F1 itself, which assembles inthe absence of F0 or the stator (Schatz 1968; Tzagoloff1969; Velours and Arselin 2000). The newly imported sub-unit Atp1 is bound stoichiometrically in a dimeric assemblyintermediate with the specific chaperone Atp12, while thenewly imported Atp2 subunit is similarly bound by its chap-erone Atp11 (Ackerman and Tzagoloff 1990; Ludlam et al.2009). Release of these F1 subunits from their chaperones toallow the formation of Atp1-Atp2 dimers and hexamersrequires the presence of the subunit Atp3 (g), a componentof the central stalk. In the case of Atp1, structural evidencestrongly suggests that Atp3 binding to Atp1 directly displa-ces the chaperone Atp12 (Ludlam et al. 2009). An additionalprotein, Fmc1, is required to generate Atp1-Atp2 dimers athigh temperature (Lefebvre-Legendre et al. 2001), and Hsp90chaperones have also been implicated in this process (Francisand Thorsness 2011). Addition of the central stalk proteinsAtp16 (d) and Atp15 (e) has not been examined.
A second pre-assembly module is the rotating componentof F0, comprising a decameric ring of the highly hydrophobicmitochondrially coded Atp9 (subunit c) embedded in themembrane (Stock et al. 1999; Rak et al. 2011). [The assem-bled ring structure is not dissociated by treatment with SDS(Herrmann et al. 1994; Rak et al. 2011).] Translation and/or stability of the ATP9mRNA requires the specific activatorsAep1, Aep2, and Atp25 (Ackerman et al. 1991; Finneganet al. 1991; Payne et al. 1993; Ziaja et al. 1993; Ellis et al.1999; Zeng et al. 2008; Rak et al. 2009). Atp25 is particu-larly interesting as the 60-kDa protein is cleaved roughly inhalf to yield a C-terminal fragment that is required for ATP9mRNA stability and/or translation and an N-terminal frag-ment that is not required for Atp9 synthesis but appears topromote assembly of the Atp9 ring (Zeng et al. 2008). In-sertion of Atp9 into the inner membrane and assembly of thering structure does not require Oxa1 (Lemaire et al. 2000;Jia et al. 2007; Mathieu et al. 2010) or the other mitochond-rially coded F0 subunits Atp6 and Atp8 (Rak et al. 2011).However, Oxa1 does have a role in promoting association ofthe Atp9 ring with a single Atp6 molecule late in the assem-bly of ATP synthase (Jia et al. 2007)
1222 T. D. Fox
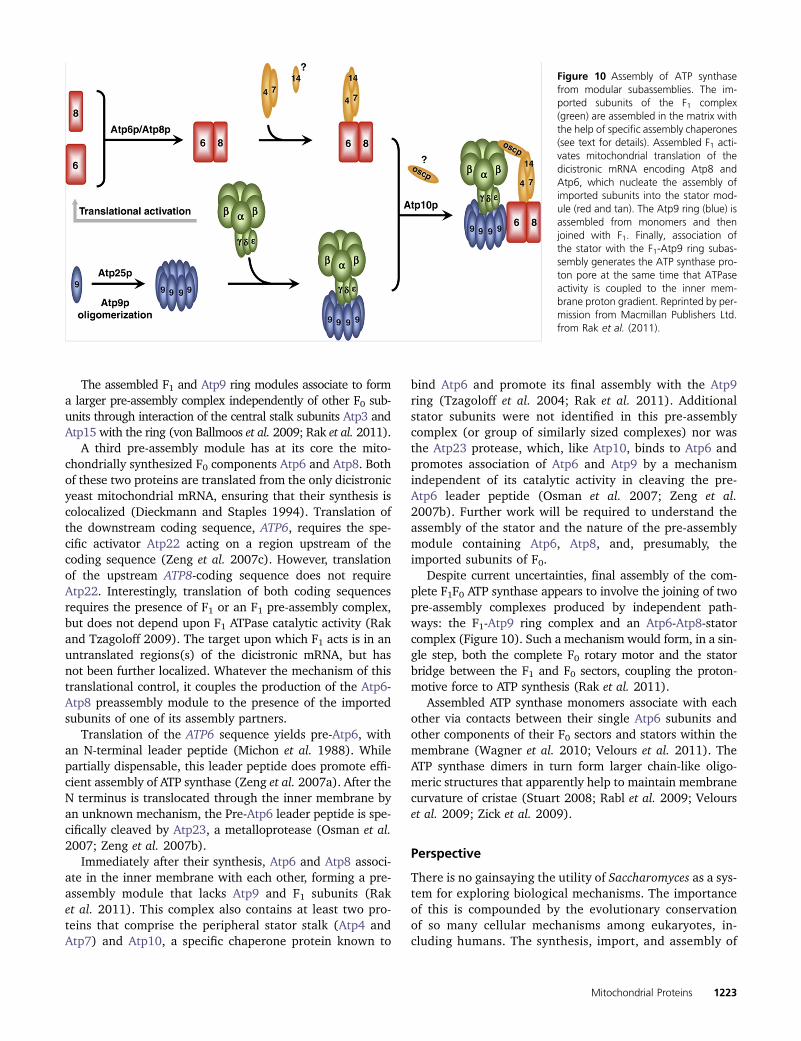
The assembled F1 and Atp9 ring modules associate to forma larger pre-assembly complex independently of other F0 sub-units through interaction of the central stalk subunits Atp3 andAtp15 with the ring (von Ballmoos et al. 2009; Rak et al. 2011).
A third pre-assembly module has at its core the mito-chondrially synthesized F0 components Atp6 and Atp8. Bothof these two proteins are translated from the only dicistronicyeast mitochondrial mRNA, ensuring that their synthesis iscolocalized (Dieckmann and Staples 1994). Translation ofthe downstream coding sequence, ATP6, requires the spe-cific activator Atp22 acting on a region upstream of thecoding sequence (Zeng et al. 2007c). However, translationof the upstream ATP8-coding sequence does not requireAtp22. Interestingly, translation of both coding sequencesrequires the presence of F1 or an F1 pre-assembly complex,but does not depend upon F1 ATPase catalytic activity (Rakand Tzagoloff 2009). The target upon which F1 acts is in anuntranslated regions(s) of the dicistronic mRNA, but hasnot been further localized. Whatever the mechanism of thistranslational control, it couples the production of the Atp6-Atp8 preassembly module to the presence of the importedsubunits of one of its assembly partners.
Translation of the ATP6 sequence yields pre-Atp6, withan N-terminal leader peptide (Michon et al. 1988). Whilepartially dispensable, this leader peptide does promote effi-cient assembly of ATP synthase (Zeng et al. 2007a). After theN terminus is translocated through the inner membrane byan unknown mechanism, the Pre-Atp6 leader peptide is spe-cifically cleaved by Atp23, a metalloprotease (Osman et al.2007; Zeng et al. 2007b).
Immediately after their synthesis, Atp6 and Atp8 associ-ate in the inner membrane with each other, forming a pre-assembly module that lacks Atp9 and F1 subunits (Raket al. 2011). This complex also contains at least two pro-teins that comprise the peripheral stator stalk (Atp4 andAtp7) and Atp10, a specific chaperone protein known to
bind Atp6 and promote its final assembly with the Atp9ring (Tzagoloff et al. 2004; Rak et al. 2011). Additionalstator subunits were not identified in this pre-assemblycomplex (or group of similarly sized complexes) nor wasthe Atp23 protease, which, like Atp10, binds to Atp6 andpromotes association of Atp6 and Atp9 by a mechanismindependent of its catalytic activity in cleaving the pre-Atp6 leader peptide (Osman et al. 2007; Zeng et al.2007b). Further work will be required to understand theassembly of the stator and the nature of the pre-assemblymodule containing Atp6, Atp8, and, presumably, theimported subunits of F0.
Despite current uncertainties, final assembly of the com-plete F1F0 ATP synthase appears to involve the joining of twopre-assembly complexes produced by independent path-ways: the F1-Atp9 ring complex and an Atp6-Atp8-statorcomplex (Figure 10). Such a mechanism would form, in a sin-gle step, both the complete F0 rotary motor and the statorbridge between the F1 and F0 sectors, coupling the proton-motive force to ATP synthesis (Rak et al. 2011).
Assembled ATP synthase monomers associate with eachother via contacts between their single Atp6 subunits andother components of their F0 sectors and stators within themembrane (Wagner et al. 2010; Velours et al. 2011). TheATP synthase dimers in turn form larger chain-like oligo-meric structures that apparently help to maintain membranecurvature of cristae (Stuart 2008; Rabl et al. 2009; Velourset al. 2009; Zick et al. 2009).
Perspective
There is no gainsaying the utility of Saccharomyces as a sys-tem for exploring biological mechanisms. The importanceof this is compounded by the evolutionary conservationof so many cellular mechanisms among eukaryotes, in-cluding humans. The synthesis, import, and assembly of
Figure 10 Assembly of ATP synthasefrom modular subassemblies. The im-ported subunits of the F1 complex(green) are assembled in the matrix withthe help of specific assembly chaperones(see text for details). Assembled F1 acti-vates mitochondrial translation of thedicistronic mRNA encoding Atp8 andAtp6, which nucleate the assembly ofimported subunits into the stator mod-ule (red and tan). The Atp9 ring (blue) isassembled from monomers and thenjoined with F1. Finally, association ofthe stator with the F1-Atp9 ring subas-sembly generates the ATP synthase pro-ton pore at the same time that ATPaseactivity is coupled to the inner mem-brane proton gradient. Reprinted by per-mission from Macmillan Publishers Ltd.from Rak et al. (2011).
Mitochondrial Proteins 1223

mitochondrial proteins can be largely included with othersuch conserved processes.
The TOM, SAM, TIM22, and TIM23 complexes appearto be nearly universally present in eukaryotes (Paschenet al. 2003; Dolezal et al. 2006; Kutik et al. 2009; Hewittet al. 2011; Rada et al. 2011) [the TIM22 complex appearsto be absent in some, such as Trypanosomes (Schneideret al. 2008)]. The core components of these complexes havemaintained significant sequence conservation. Furthermore,expression of a mitochondrially targeted protein from onespecies in cells of another typically results in correct proteinlocalization by the host import machinery.
Similarly, many of the accessory proteins required to as-semble respiratory complexes have apparent or confirmedorthologs in humans (Ackerman and Tzagoloff 2005; Sotoet al. 2011; Wagener et al. 2011). Particularly striking is thefact that the human orthologs of Oxa1 (Bonnefoy et al.1994) and Cox18 (Gaisne and Bonnefoy 2006) can supportassembly of cytochrome c oxidase and respiratory growth inthe corresponding yeast mutants. Furthermore, the ability ofthe human OXA1 mRNA to direct synthesis of the functionalmitochondrial protein depends upon interaction of the hu-man mRNA 39-UTR with yeast mitochondria, suggesting con-servation in the mechanisms of mRNA targeting (Sylvestreet al. 2003).
In contrast to the clear conservation of protein import andassembly mechanisms, mitochondrial genome structures andtheir expression systems are highly divergent among eu-karyotic phyla (Burger et al. 2003). Nevertheless, the strat-egy of linking transcription and translation together at theinner membrane, identified in yeast, appears to be employedby mammals for the synthesis and assembly of highly hydro-phobic mitochondrial gene products (Weraarpachai et al. 2009;Sasarman et al. 2010; Brown et al. 2011; He et al. 2012).
Given the increasingly evident role of a broad array ofmitochondrial functions in the maintenance of healthy hu-man cells, tissues, and bodies (Nunnari and Suomalainen2012), detailed studies of mechanisms underlying mito-chondrial activities in yeast should remain a high priority.
Acknowledgments
Research in the author’s laboratory is supported by NationalInstitutes of Health grant R01-GM29362.
Literature Cited
Abe, Y., T. Shodai, T. Muto, K. Mihara, H. Torii et al., 2000 Structuralbasis of presequence recognition by the mitochondrial proteinimport receptor Tom20. Cell 100: 551–560.
Ackerman, S. H., and A. Tzagoloff, 1990 Identification of twonuclear genes (ATP11, ATP12) required for assembly of theyeast F1-ATPase. Proc. Natl. Acad. Sci. USA 87: 4986–4990.
Ackerman, S. H., and A. Tzagoloff, 2005 Function, structure, andbiogenesis of mitochondrial ATP synthase. Prog. Nucleic AcidRes. Mol. Biol. 80: 95–133.
Ackerman, S. H., D. L. Gatti, P. Gellefors, M. G. Douglas, and A.Tzagoloff, 1991 ATP13, a nuclear gene of Saccharomyces cer-
evisiae essential for the expression of subunit 9 of the mitochon-drial ATPase. FEBS Lett. 278: 234–238.
Ades, I. Z., and R. A. Butow, 1980 The products of mitochondria-bound cytoplasmic polysomes in yeast. J. Biol. Chem. 255:9918–9924.
Ahting, U., T. Waizenegger, W. Neupert, and D. Rapaport,2005 Signal-anchored proteins follow a unique insertion path-way into the outer membrane of mitochondria. J. Biol. Chem.280: 48–53.
Aiken Hobbs, A. E., M. Srinivasan, J. M. McCaffery, and R. E. Jensen,2001 Mmm1p, a mitochondrial outer membrane protein, is con-nected to mitochondrial DNA (mtDNA) nucleoids and requiredfor mtDNA stability. J. Cell Biol. 152: 401–410.
Alkhaja, A. K., D. C. Jans, M. Nikolov, M. Vukotic, O. Lytovchenkoet al., 2012 MINOS1 is a conserved component of Mitofilincomplexes and required for mitochondrial function and cristaeorganization. Mol. Biol. Cell 23: 247–257.
Arnold, I., H. Folsch, W. Neupert, and R. A. Stuart, 1998 Twodistinct and independent mitochondrial targeting signals func-tion in the sorting of an inner membrane protein, cytochromec1. J. Biol. Chem. 273: 1469–1476.
Atkinson, A., O. Khalimonchuk, P. Smith, H. Sabic, D. Eide et al.,2010 Mzm1 influences a labile pool of mitochondrial zinc im-portant for respiratory function. J. Biol. Chem. 285: 19450–19459.
Atkinson, A., P. Smith, J. L. Fox, T. Z. Cui, O. Khalimonchuk et al.,2011 The LYR protein Mzm1 functions in the insertion of theRieske Fe/S protein in yeast mitochondria. Mol. Cell. Biol. 31:3988–3996.
Baker, K. P., and G. Schatz, 1991 Mitochondrial proteins essentialfor viability mediate protein import into yeast mitochondria.Nature 349: 205–208.
Baker, K. P., A. Schaniel, D. Vestweber, and G. Schatz, 1990 Ayeast mitochondrial outer membrane protein essential for pro-tein import and cell viability. Nature 348: 605–609.
Baker, M. J., C. T. Webb, D. A. Stroud, C. S. Palmer, A. E. Frazieret al., 2009 Structural and functional requirements for activityof the Tim9-Tim10 complex in mitochondrial protein import.Mol. Biol. Cell 20: 769–779.
Banci, L., I. Bertini, C. Cefaro, L. Cenacchi, S. Ciofi-Baffoni et al.,2010 Molecular chaperone function of Mia40 triggers consec-utive induced folding steps of the substrate in mitochondrialprotein import. Proc. Natl. Acad. Sci. USA 107: 20190–20195.
Banting, G. S., and D. M. Glerum, 2006 Mutational analysis of theSaccharomyces cerevisiae cytochrome c oxidase assembly proteinCox11p. Eukaryot. Cell 5: 568–578.
Barrientos, A., M. H. Barros, I. Valnot, A. Rotig, P. Rustin et al.,2002a Cytochrome oxidase in health and disease. Gene 286:53–63.
Barrientos, A., D. Korr, and A. Tzagoloff, 2002b Shy1p is neces-sary for full expression of mitochondrial COX1 in the yeastmodel of Leigh’s syndrome. EMBO J. 21: 43–52.
Barrientos, A., A. Zambrano, and A. Tzagoloff, 2004 Mss51p andCox14p jointly regulate mitochondrial Cox1p expression in Sac-charomyces cerevisiae. EMBO J. 23: 3472–3482.
Barrientos, A., K. Gouget, D. Horn, I. C. Soto, and F. Fontanesi,2009 Suppression mechanisms of COX assembly defects inyeast and human: insights into the COX assembly process. Bio-chim. Biophys. Acta 1793: 97–107.
Becker, T., S. Pfannschmidt, B. Guiard, D. Stojanovski, D. Milenkovicet al., 2008 Biogenesis of the mitochondrial TOM complex:Mim1 promotes insertion and assembly of signal-anchored re-ceptors. J. Biol. Chem. 283: 120–127.
Becker, T., B. Guiard, N. Thornton, N. Zufall, D. A. Stroud et al.,2010 Assembly of the mitochondrial protein import channel:role of Tom5 in two-stage interaction of Tom40 with the SAMcomplex. Mol. Biol. Cell 21: 3106–3113.
1224 T. D. Fox

Becker, T., L. S. Wenz, V. Kruger, W. Lehmann, J. M. Muller et al.,2011 The mitochondrial import protein Mim1 promotes bio-genesis of multispanning outer membrane proteins. J. Cell Biol.194: 387–395.
Beers, J., D. M. Glerum, and A. Tzagoloff, 1997 Purification, char-acterization, and localization of yeast Cox17p, a mitochondrialcopper shuttle. J. Biol. Chem. 272: 33191–33196.
Bender, T., I. Lewrenz, S. Franken, C. Baitzel,, and W. Voos,2011 Mitochondrial enzymes are protected from stress-induced aggregation by mitochondrial chaperones and thePim1/LON protease. Mol. Biol. Cell 22: 541–554.
Bernard, D. G., S. T. Gabilly, G. Dujardin, S. Merchant, and P. P.Hamel, 2003 Overlapping specificities of the mitochondrialcytochrome c and c1 heme lyases. J. Biol. Chem. 278: 49732–49742.
Bernard, D. G., S. Quevillon-Cheruel, S. Merchant, B. Guiard, andP. P. Hamel, 2005 Cyc2p, a membrane-bound flavoprotein in-volved in the maturation of mitochondrial c-type cytochromes.J. Biol. Chem. 280: 39852–39859.
Bestwick, M., O. Khalimonchuk, F. Pierrel, and D. R. Winge,2010 The role of Coa2 in hemylation of yeast Cox1 revealedby its genetic interaction with Cox10. Mol. Cell. Biol. 30: 172–185.
Bihlmaier, K., N. Mesecke, N. Terziyska, M. Bien, K. Hell et al.,2007 The disulfide relay system of mitochondria is connectedto the respiratory chain. J. Cell Biol. 179: 389–395.
Bohnert, M., P. Rehling, B. Guiard, J. M. Herrmann, N. Pfanneret al., 2010 Cooperation of stop-transfer and conservative sort-ing mechanisms in mitochondrial protein transport. Curr. Biol.20: 1227–1232.
Boldogh, I. R., D. W. Nowakowski, H. C. Yang, H. Chung, S. Karmonet al., 2003 A protein complex containing Mdm10p, Mdm12pand Mmm1p links mitochondrial membranes and DNA to thecytoskeleton-based segregation machinery. Mol. Biol. Cell 14:4618–4627.
Bonnefoy, N., M. Kermorgant, O. Groudinsky, M. Minet, P. P.Slonimski et al., 1994 Cloning of a human gene involvedin cytochrome oxidase assembly by functional complementa-tion of an oxa12 mutation in Saccharomyces cerevisiae. Proc.Natl. Acad. Sci. USA 91: 11978–11982.
Bonnefoy, N., N. Bsat, and T. D. Fox, 2001 Mitochondrial trans-lation of Saccharomyces cerevisiae COX2 mRNA is controlled bythe nucleotide sequence specifying the pre-Cox2p leader pep-tide. Mol. Cell. Biol. 21: 2359–2372.
Bonnefoy, N., H. L. Fiumera, G. Dujardin, and T. D. Fox, 2009 Rolesof Oxa1-related inner-membrane translocases in assembly ofrespiratory chain complexes. Biochim. Biophys. Acta 1793:60–70.
Broadley, S. A., C. M. Demlow, and T. D. Fox, 2001 Peripheralmitochondrial inner membrane protein, Mss2p, required for ex-port of the mitochondrially coded Cox2p C-tail in Saccharomycescerevisiae. Mol. Cell. Biol. 21: 7663–7672.
Brown, N. G., M. C. Costanzo, and T. D. Fox, 1994 Interactionsamong three proteins that specifically activate translation of themitochondrial COX3 mRNA in Saccharomyces cerevisiae. Mol.Cell. Biol. 14: 1045–1053.
Brown, T. A., A. N. Tkachuk, G. Shtengel, B. G. Kopek, D. F.Bogenhagen et al., 2011 Superresolution fluorescence imag-ing of mitochondrial nucleoids reveals their spatial range,limits, and membrane interaction. Mol. Cell. Biol. 31: 4994–5010.
Bryan, A. C., M. S. Rodeheffer, C. M. Wearn, and G. S. Shadel,2002 Sls1p is a membrane-bound regulator of transcription-coupled processes involved in Saccharomyces cerevisiae mito-chondrial gene expression. Genetics 160: 75–82.
Burger, G., M. W. Gray, and B. F. Lang, 2003 Mitochondrial ge-nomes: anything goes. Trends Genet. 19: 709–716.
Carr, H. S., and D. R. Winge, 2003 Assembly of cytochrome coxidase within the mitochondrion. Acc. Chem. Res. 36: 309–316.
Chacinska, A., and M. Boguta, 2000 Coupling of mitochondrialtranslation with the formation of respiratory complexes in yeastmitochondria. Acta Biochim. Pol. 47: 973–991.
Chacinska, A., S. Pfannschmidt, N. Wiedemann, V. Kozjak, L. K.Sanjuan Szklarz et al., 2004 Essential role of Mia40 in importand assembly of mitochondrial intermembrane space proteins.EMBO J. 23: 3735–3746.
Chacinska, A., M. Lind, A. E. Frazier, J. Dudek, C. Meisinger et al.,2005 Mitochondrial presequence translocase: switching be-tween TOM tethering and motor recruitment involves Tim21and Tim17. Cell 120: 817–829.
Chacinska, A., C. M. Koehler, D. Milenkovic, T. Lithgow, and N.Pfanner, 2009 Importing mitochondrial proteins: machineriesand mechanisms. Cell 138: 628–644.
Chacinska, A., M. van der Laan, C. S. Mehnert, B. Guiard, D. U.Mick et al., 2010 Distinct forms of mitochondrial TOM-TIMsupercomplexes define signal-dependent states of preproteinsorting. Mol. Cell. Biol. 30: 307–318.
Chatenay-Lapointe, M., and G. S. Shadel, 2011 Repression ofmitochondrial translation, respiration and a metabolic cycle-regulated gene, SLF1, by the yeast Pumilio-family protein Puf3p.PLoS ONE 6: e20441.
Chen, X. J., and R. A. Butow, 2005 The organization and inheri-tance of the mitochondrial genome. Nat. Rev. Genet. 6: 815–825.
Chen, Y. C., E. B. Taylor, N. Dephoure, J. M. Heo, A. Tonhato et al.,2012 Identification of a protein mediating respiratory super-complex stability. Cell Metab. 15: 348–360.
Church, C., B. Goehring, D. Forsha, P. Wazny, and R. O. Poyton,2005 A role for Pet100p in the assembly of yeast cytochrome coxidase: interaction with a sub-assembly that accumulates ina pet100 mutant. J. Biol. Chem. 280: 1854–1863.
Clarkson, G. H. D., and R. O. Poyton, 1989 A role for membranepotential in the biogenesis of cytochrome c oxidase subunit II,a mitochondrial gene product. J. Biol. Chem. 264: 10114–10118.
Cobine, P. A., F. Pierrel, and D. R. Winge, 2006 Copper traffickingto the mitochondrion and assembly of copper metalloenzymes.Biochim. Biophys. Acta 1763: 759–772.
Coffin, J. W., R. Dhillon, R. G. Ritzel, and F. E. Nargang, 1997 TheNeurospora crassa cya-5 nuclear gene encodes a protein witha region of homology to the Saccharomyces cerevisiae PET309protein and is required in a post-transcriptional step for theexpression of the mitochondrially encoded COXI protein. Curr.Genet. 32: 273–280.
Conte, L., B. L. Trumpower, and V. Zara, 2011 Bcs1p can rescuea large and productive cytochrome bc(1) complex assembly in-termediate in the inner membrane of yeast mitochondria. Bio-chim. Biophys. Acta 1813: 91–101.
Corral-Debrinski, M., C. Blugeon, and C. Jacq, 2000 In yeast, the39 untranslated region or the presequence of ATM1 is requiredfor the exclusive localization of its mRNA to the vicinity of mi-tochondria. Mol. Cell. Biol. 20: 7881–7892.
Costanzo, M. C., N. Bonnefoy, E. H. Williams, G. D. Clark-Walker,and T. D. Fox, 2000 Highly diverged homologs of Saccharomy-ces cerevisiaemitochondrial mRNA-specific translational activatorshave orthologous functions in other budding yeasts. Genetics154: 999–1012.
Craig, E. A., 1993 Chaperones: helpers along the pathways toprotein folding. Science 260: 1902–1903.
Crivellone, M. D., M. Wu, and A. Tzagoloff, 1988 Assembly of themitochondrial membrane system: analysis of structural mutantsof the yeast coenzyme QH2-cytochrome c reductase complex. J.Biol. Chem. 263: 14323–14333.
Mitochondrial Proteins 1225

Curran, S. P., D. Leuenberger, W. Oppliger, and C. M. Koehler,2002 The Tim9p-Tim10p complex binds to the transmem-brane domains of the ADP/ATP carrier. EMBO J. 21: 942–953.
Curran, S. P., D. Leuenberger, E. P. Leverich, D. K. Hwang, K. N.Beverly et al., 2004 The role of Hot13p and redox chemistry inthe mitochondrial TIM22 import pathway. J. Biol. Chem. 279:43744–43751.
Dabir, D. V., E. P. Leverich, S. K. Kim, F. D. Tsai, M. Hirasawa et al.,2007 A role for cytochrome c and cytochrome c peroxidase inelectron shuttling from Erv1. EMBO J. 26: 4801–4811.
Dalbey, R. E., and A. Kuhn, 2004 YidC family members are in-volved in the membrane insertion, lateral integration, folding,and assembly of membrane proteins. J. Cell Biol. 166: 769–774.
Daum, G., P. Böhni, and G. Schatz, 1982 Import of proteins intomitochondria: cytochrome b2 and cytochrome c peroxidase arelocated in the intermembrane space of yeast mitochondria. J.Biol. Chem. 257: 13028–13033.
Davies, K. M., M. Strauss, B. Daum, J. H. Kief, H. D. Osiewacz et al.,2011 Macromolecular organization of ATP synthase and com-plex I in whole mitochondria. Proc. Natl. Acad. Sci. USA 108:14121–14126.
Davis, A. J., N. B. Sepuri, J. Holder, A. E. Johnson, and R. E. Jensen,2000 Two intermembrane space TIM complexes interact withdifferent domains of Tim23p during its import into mitochon-dria. J. Cell Biol. 150: 1271–1282.
Decoster, E., M. Simon, D. Hatat, and G. Faye, 1990 The MSS51gene product is required for the translation of the COX1 mRNAin yeast mitochondria. Mol. Gen. Genet. 224: 111–118.
Dekker, P. J. T., F. Martin, A. C. Maarse, U. Bomer, H. Muller et al.,1997 The tim core complex defines the number of mitochon-drial translocation contact sites and can hold arrested prepro-teins in the absence of matrix Hsp70-Tim44. EMBO J. 16: 5408–5419.
Delannoy, E., W. A. Stanley, C. S. Bond, and I. D. Small,2007 Pentatricopeptide repeat (PPR) proteins as sequence-specificity factors in post-transcriptional processes in organelles.Biochem. Soc. Trans. 35: 1643–1647.
Demlow, C. M., and T. D. Fox, 2003 Activity of mitochondriallysynthesized reporter proteins is lower than imported proteins,and is increased by lowering cAMP in glucose-grown Saccharo-myces cerevisiae cells. Genetics 165: 961–974.
Deshaies, R. J., B. D. Koch, M. Werner-Washburne, E. A. Craig, andR. Schekman, 1988 A subfamily of stress proteins facilitatestranslocation of secretory and mitochondrial precursor polypep-tides. Nature 332: 800–805.
Dieckmann, C. L., and R. R. Staples, 1994 Regulation of mito-chondrial gene expression in Saccharomyces cerevisiae. Int. Rev.Cytol. 152: 145–181.
Diekert, K., G. Kispal, B. Guiard, and R. Lill, 1999 An internaltargeting signal directing proteins into the mitochondrial in-termembrane space. Proc. Natl. Acad. Sci. USA 96: 11752–11757.
Diekert, K., A. I. de Kroon, U. Ahting, B. Niggemeyer, W. Neupertet al., 2001 Apocytochrome c requires the TOM complex fortranslocation across the mitochondrial outer membrane. EMBOJ. 20: 5626–5635.
Dimmer, K. S., D. Papic, B. Schumann, D. Sperl, K. Krumpe et al.,2012 A crucial role of Mim2 in the biogenesis of mitochondrialouter membrane proteins. J. Cell Sci. 125: 3464–3473.
Dolezal, P., V. Likic, J. Tachezy, and T. Lithgow, 2006 Evolution ofthe molecular machines for protein import into mitochondria.Science 313: 314–318.
Dukanovic, J., and D. Rapaport, 2011 Multiple pathways in theintegration of proteins into the mitochondrial outer membrane.Biochim. Biophys. Acta 1808: 971–980.
Dumont, M. E., T. S. Cardillo, M. K. Hayes, and F. Sherman,1991 Role of cytochrome c heme lyase in mitochondrial import
and accumulation of cytochrome c in Saccharomyces cerevisiae.Mol. Cell. Biol. 11: 5487–5496.
Dyall, S. D., S. C. Agius, C. De Marcos Lousa, V. Trezeguet, and K.Tokatlidis, 2003 The dynamic dimerization of the yeast ADP/ATP carrier in the inner mitochondrial membrane is affectedby conserved cysteine residues. J. Biol. Chem. 278: 26757–26764.
Eliyahu, E., L. Pnueli, D. Melamed, T. Scherrer, A. P. Gerber et al.,2010 Tom20 mediates localization of mRNAs to mitochondriain a translation-dependent manner. Mol. Cell. Biol. 30: 284–294.
Elliott, L. E., S. A. Sarraco,, and T. D. Fox, 2012 Multiple roles ofthe Cox20 chaperone in assembly of Saccharomyces cereviaecytochrome c oxidase. Genetics 190: 559–567.
Ellis, T. P., H. B. Lukins, P. Nagley, and B. E. Corner, 1999 Suppressionof a nuclear aep2 mutation in Saccharomyces cerevisiae by a basesubstitution in the 59-untranslated region of the mitochondrial oli1gene encoding subunit 9 of ATP synthase. Genetics 151: 1353–1363.
Embley, T. M., and W. Martin, 2006 Eukaryotic evolution,changes and challenges. Nature 440: 623–630.
Endo, T., and K. Yamano, 2010 Transport of proteins across orinto the mitochondrial outer membrane. Biochim. Biophys. Acta1803: 706–714.
Endo, T., K. Yamano, and S. Kawano, 2011 Structural insight intothe mitochondrial protein import system. Biochim. Biophys.Acta 1808: 955–970.
Fillingame, R. H., and O. Y. Dmitriev, 2002 Structural model ofthe transmembrane Fo rotary sector of H+-transporting ATPsynthase derived by solution NMR and intersubunit cross-linkingin situ. Biochim. Biophys. Acta 1565: 232–245.
Finnegan, P. M., M. J. Payne, E. Kermidaris, and H. B. Lukins,1991 Characterization of a yeast nuclear gene, AEP2, requiredfor accumulation of mitochondrial mRNA encoding subunit-9 ofthe ATP synthase. Curr. Genet. 20: 53–61.
Fiori, A., T. L. Mason, and T. D. Fox, 2003 Evidence that synthesisof the Saccharomyces cerevisiae mitochondrially-encoded ribo-somal protein Var1p may be membrane localized. Eukaryot. Cell2: 651–653.
Fiori, A., X. Perez-Martinez, and T. D. Fox, 2005 Overexpressionof the COX2 translational activator, Pet111p, prevents transla-tion of COX1 mRNA and cytochrome c oxidase assembly in mi-tochondria of Saccharomyces cerevisiae. Mol. Microbiol. 56:1689–1704.
Fiumera, H. L., S. A. Broadley, and T. D. Fox, 2007 Translocationof mitochondrially synthesized Cox2p domains from the matrixto the intermembrane space. Mol. Cell. Biol. 27: 4664–4673.
Fiumera, H. L., M. J. Dunham, S. A. Saracco, C. A. Butler, J. A. Kellyet al., 2009 Translocation and assembly of mitochondriallycoded Saccharomyces cerevisiae cytochrome c oxidase subunitCox2 by Oxa1 and Yme1 in the absence of Cox18. Genetics182: 519–528.
Foat, B. C., S. S. Houshmandi, W. M. Olivas, and H. J. Bussemaker,2005 Profiling condition-specific, genome-wide regulation ofmRNA stability in yeast. Proc. Natl. Acad. Sci. USA 102:17675–17680.
Fontanesi, F., I. C. Soto, D. Horn, and A. Barrientos, 2006 Assemblyof mitochondrial cytochrome c oxidase, a complicated and highlyregulated cellular process. Am. J. Physiol. Cell Physiol. 291:c1129–c1147.
Fontanesi, F., I. C. Soto, and A. Barrientos, 2008 Cytochrome coxidase biogenesis: new levels of regulation. IUBMB Life 60:557–568.
Fontanesi, F., P. Clemente, and A. Barrientos, 2010a Cox25 teamsup with Mss51, Ssc1 and Cox14 to regulate mitochondrial cyto-chrome C oxidase subunit 1 expression and assembly in Saccha-romyces cerevisiae. J. Biol. Chem. 286: 555–566.
1226 T. D. Fox

Fontanesi, F., I. C. Soto, D. Horn, and A. Barrientos, 2010b Mss51and Ssc1 facilitate translational regulation of cytochrome c ox-idase biogenesis. Mol. Cell. Biol. 30: 245–259.
Fox, T. D., 1996a Genetics of mitochondrial translation, pp. 733–758 in Translational Control, edited by J. W. B. Hershey, M. B.Matthews, and N. Sonenberg. Cold Spring Harbor Press, ColdSpring Harbor, NY.
Fox, T. D., 1996b Translational control of endogenous and re-coded nuclear genes in yeast mitochondria: regulation andmembrane targeting. Experientia 52: 1130–1135.
Francis, B. R., and P. E. Thorsness, 2011 Hsp90 and mitochon-drial proteases Yme1 and Yta10/12 participate in ATP synthaseassembly in Saccharomyces cerevisiae. Mitochondrion 11: 587–600.
Frazier, A. E., A. Chacinska, K. N. Truscott, B. Guiard, N. Pfanneret al., 2003 Mitochondria use different mechanisms for trans-port of multispanning membrane proteins through the inter-membrane space. Mol. Cell. Biol. 23: 7818–7828.
Frazier, A. E., J. Dudek, B. Guiard, W. Voos, Y. Li et al.,2004 Pam16 has an essential role in the mitochondrial proteinimport motor. Nat. Struct. Mol. Biol. 11: 226–233.
Frazier, A. E., R. D. Taylor, D. U. Mick, B. Warscheid, N. Stoepelet al., 2006 Mdm38 interacts with ribosomes and is a compo-nent of the mitochondrial protein export machinery. J. Cell Biol.172: 553–564.
Frey, T. G., and C. A. Mannella, 2000 The internal structure ofmitochondria. Trends Biochem. Sci. 25: 319–324.
Froschauer, E., K. Nowikovsky, and R. J. Schweyen,2005 Electroneutral K+/H+ exchange in mitochondrial mem-brane vesicles involves Yol027/Letm1 proteins. Biochim. Bio-phys. Acta 1711: 41–48.
Funes, S., F. E. Nargang, W. Neupert, and J. M. Herrmann,2004 The Oxa2 protein of Neurospora crassa plays a criticalrole in the biogenesis of cytochrome oxidase and defines a ubiq-uitous subbranch of the Oxa1/YidC/Alb3 protein family. Mol.Biol. Cell 15: 1853–1861.
Gabriel, K., B. Egan, and T. Lithgow, 2003 Tom40, the importchannel of the mitochondrial outer membrane, plays an activerole in sorting imported proteins. EMBO J. 22: 2380–2386.
Gadir, N., L. Haim-Vilmovsky, J. Kraut-Cohen, and J. E. Gerst,2011 Localization of mRNAs coding for mitochondrial proteinsin the yeast Saccharomyces cerevisiae. RNA 17: 1551–1565.
Gaisne, M., and N. Bonnefoy, 2006 The COX18 gene, involved inmitochondrial biogenesis, is functionally conserved and tightlyregulated in humans and fission yeast. FEMS Yeast Res. 6: 869–882.
Gan, X., M. Kitakawa, K. Yoshino, N. Oshiro, K. Yonezawa et al.,2002 Tag-mediated isolation of yeast mitochondrial ribosomeand mass spectrometric identification of its new components.Eur. J. Biochem. 269: 5203–5214.
Garcia, M., X. Darzacq, T. Delaveau, L. Jourdren, R. H. Singer et al.,2007a Mitochondria-associated yeast mRNAs and the biogen-esis of molecular complexes. Mol. Biol. Cell 18: 362–368.
Garcia, M., X. Darzacq, F. Devaux, R. H. Singer, and C. Jacq,2007b Yeast mitochondrial transcriptomics. Methods Mol. Biol.372: 505–528.
Garcia, M., T. Delaveau, S. Goussard, and C. Jacq, 2010 Mito-chondrial presequence and open reading frame mediate asym-metric localization of messenger RNA. EMBO Rep. 11: 285–291.
Garcia-Rodriguez, L. J., A. C. Gay, and L. A. Pon, 2007 Puf3p,a Pumilio family RNA binding protein, localizes to mitochondriaand regulates mitochondrial biogenesis and motility in buddingyeast. J. Cell Biol. 176: 197–207.
Gärtner, F., W. Voos, A. Querol, B. R. Miller, E. A. Craig et al.,1995 Mitochondrial import of subunit Va of cytochrome c ox-idase characterized with yeast mutants: independence from re-
ceptors but requirement for matrix hsp70 translocase function.J. Biol. Chem. 270: 3788–3795.
Gautschi, M., H. Lilie, U. Funfschilling, A. Mun, S. Ross et al.,2001 RAC, a stable ribosome-associated complex in yeastformed by the DnaK-DnaJ homologs Ssz1p and zuotin. Proc.Natl. Acad. Sci. USA 98: 3762–3767.
Gebert, N., M. T. Ryan, N. Pfanner, N. Wiedemann, and D. Stojanovski,2011 Mitochondrial protein import machineries and lipids:a functional connection. Biochim. Biophys. Acta 1808: 1002–1011.
Gentle, I., K. Gabriel, P. Beech, R. Waller, and T. Lithgow,2004 The Omp85 family of proteins is essential for outermembrane biogenesis in mitochondria and bacteria. J. Cell Biol.164: 19–24.
Gerber, A. P., D. Herschlag, and P. O. Brown, 2004 Extensiveassociation of functionally and cytotopically related mRNAswith Puf family RNA-binding proteins in yeast. PLoS Biol. 2:E79.
Ghaemmaghami, S., W. K. Huh, K. Bower, R. W. Howson, A. Belleet al., 2003 Global analysis of protein expression in yeast. Na-ture 425: 737–741.
Glerum, D. M., A. Shtanko, and A. Tzagoloff, 1996 Char-acterization of COX17, a yeast gene involved in copper metab-olism and assembly of cytochrome oxidase. J. Biol. Chem. 271:14504–14509.
Glick, G. S., A. Brandt, K. Cunningham, S. Müller, R. L. Hallberget al., 1992 Cytochromes c1 and b2 are sorted to the intermem-brane space of yeast mitochondria by a stop-transfer mecha-nism. Cell 69: 809–822.
Gray, M. W., G. Burger, and B. F. Lang, 2001 The origin and earlyevolution of mitochondria. Genome Biol. 2: REVIEWS1018.
Green-Willms, N. S., C. A. Butler, H. M. Dunstan, and T. D. Fox,2001 Pet111p, an inner membrane-bound translational activa-tor that limits expression of the Saccharomyces cerevisiae mito-chondrial gene COX2. J. Biol. Chem. 276: 6392–6397.
Groot, G. S. P., T. L. Mason, and N. Van Harten-Loosbrock,1979 Var1 is associated with the small ribosomal subunit ofmitochondrial ribosomes in yeast. Mol. Gen. Genet. 174: 339–342.
Gross, D. P., C. A. Burgard, S. Reddehase, J. M. Leitch, V. C. Culottaet al., 2011 Mitochondrial Ccs1 contains a structural disulfidebond crucial for the import of this unconventional substrate bythe disulfide relay system. Mol. Biol. Cell 22: 3758–3767.
Gruschke, S., K. Groene, M. Heublein, S. Holz, L. Israel et al.,2010 Proteins at the polypeptide tunnel exit of the yeast mi-tochondrial ribosome. J. Biol. Chem. 285: 19022–19028.
Gruschke, S., K. Kehrein, K. Rompler, K. Grone, L. Israel et al.,2011 Cbp3-Cbp6 interacts with the yeast mitochondrial ribo-somal tunnel exit and promotes cytochrome b synthesis andassembly. J. Cell Biol. 193: 1101–1114.
Harner, M., C. Korner, D. Walther, D. Mokranjac, J. Kaesmacheret al., 2011a The mitochondrial contact site complex, a deter-minant of mitochondrial architecture. EMBO J. 30: 4356–4370.
Harner, M., W. Neupert, and M. Deponte, 2011b Lateral release ofproteins from the TOM complex into the outer membrane ofmitochondria. EMBO J. 30: 3232–3241.
Hartl, F. U., 1996 Molecular chaperones in cellular protein fold-ing. Nature 381: 571–579.
Hartl, F. U., B. Schmidt, E. Wachter, H. Weiss, and W. Neupert,1986 Transport into mitochondria and intramitochondrialsorting of the Fe/S protein of ubiquinol-cytochrome c reductase.Cell 47: 939–951.
He, J., H. M. Cooper, A. Reyes, M. Di Re, H. Sembongi et al.,2012 Mitochondrial nucleoid interacting proteins support mi-tochondrial protein synthesis. Nucleic Acids Res. 40: 6109–6121.
Mitochondrial Proteins 1227

He, S., and T. D. Fox, 1997 Membrane translocation of mito-chondrially coded Cox2p: distinct requirements for export ofamino- and carboxy-termini, and dependence on the conservedprotein Oxa1p. Mol. Biol. Cell 8: 1449–1460.
He, S., and T. D. Fox, 1999 Mutations affecting a yeast mitochon-drial inner membrane protein, Pnt1p, block export of a mito-chondrially synthesized fusion protein from the matrix. Mol.Cell. Biol. 19: 6598–6607.
Heinemeyer, J., H. P. Braun, E. J. Boekema, and R. Kouril, 2007 Astructural model of the cytochrome C reductase/oxidase super-complex from yeast mitochondria. J. Biol. Chem. 282: 12240–12248.
Hell, K., J. Herrmann, E. Pratje, W. Neupert, and R. A. Stuart,1997 Oxa1p mediates the export of the N- and C-termini ofpCoxII from the mitochondrial matrix to the intermembranespace. FEBS Lett. 418: 367–370.
Hell, K., J. M. Herrmann, E. Pratje, W. Neupert, and R. A. Stuart,1998 Oxa1p, an essential component of the N-tail protein ex-port machinery in mitochondria. Proc. Natl. Acad. Sci. USA 95:2250–2255.
Hell, K., A. Tzagoloff, W. Neupert, and R. A. Stuart, 2000 Identificationof Cox20p, a novel protein involved in the maturation andassembly of cytochrome oxidase subunit 2. J. Biol. Chem. 275:4571–4578.
Hell, K., W. Neupert, and R. A. Stuart, 2001 Oxa1p acts as a gen-eral membrane insertion machinery for proteins encoded bymitochondrial DNA. EMBO J. 20: 1281–1288.
Herrmann, J. M., and S. Funes, 2005 Biogenesis of cytochromeoxidase-sophisticated assembly lines in the mitochondrial innermembrane. Gene 354: 43–52.
Herrmann, J. M., and J. Riemer, 2012 Mitochondrial disulfiderelay: redox-regulated protein import into the intermembranespace. J. Biol. Chem. 287: 4426–4433.
Herrmann, J. M., R. A. Stuart, E. A. Craig, and W. Neupert,1994 Mitochondrial heat shock protein 70, a molecular chap-erone for proteins encoded by mitochondrial DNA. J. Cell Biol.127: 893–902.
Herrmann, J. M., W. Neupert, and R. A. Stuart, 1997 Insertioninto the mitochondrial inner membrane of a polytopic protein,the nuclear-encoded Oxa1p. EMBO J. 16: 2217–2226.
Hess, D. C., C. L. Myers, C. Huttenhower, M. A. Hibbs, A. P. Hayeset al., 2009 Computationally driven, quantitative experimentsdiscover genes required for mitochondrial biogenesis. PLoSGenet. 5: e1000407.
Hewitt, V., F. Alcock, and T. Lithgow, 2011 Minor modificationsand major adaptations: the evolution of molecular machinesdriving mitochondrial protein import. Biochim. Biophys. Acta1808: 947–954.
Hill, K., K. Model, M. T. Ryan, K. Dietmeier, F. Martin et al.,1998 Tom40 forms the hydrophilic channel of the mitochon-drial import pore for preproteins. Nature 395: 516–521.
Hoppins, S. C., and F. E. Nargang, 2004 The Tim8-Tim13 complexof Neurospora crassa functions in the assembly of proteins intoboth mitochondrial membranes. J. Biol. Chem. 279: 12396–12405.
Hoppins, S., S. R. Collins, A. Cassidy-Stone, E. Hummel, R. M.Devay et al., 2011 A mitochondrial-focused genetic interactionmap reveals a scaffold-like complex required for inner mem-brane organization in mitochondria. J. Cell Biol. 195: 323–340.
Horan, S., I. Bourges, J. W. Taanman, and B. Meunier, 2005 Analysisof COX2 mutants reveals cytochrome oxidase subassemblies inyeast. Biochem. J. 390: 703–708.
Horn, D., and A. Barrientos, 2008 Mitochondrial copper metabo-lism and delivery to cytochrome c oxidase. IUBMB Life 60: 421–429.
Hu, J., L. Dong, and C. E. Outten, 2008 The redox environment inthe mitochondrial intermembrane space is maintained sepa-
rately from the cytosol and matrix. J. Biol. Chem. 283:29126–29134.
Hulett, J. M., F. Lueder, N. C. Chan, A. J. Perry, P. Wolynec et al.,2008 The transmembrane segment of Tom20 is recognized byMim1 for docking to the mitochondrial TOM complex. J. Mol.Biol. 376: 694–704.
Ishikawa, D., H. Yamamoto, Y. Tamura, K. Moritoh, and T. Endo,2004 Two novel proteins in the mitochondrial outer mem-brane mediate beta-barrel protein assembly. J. Cell Biol. 166:621–627.
Islas-Osuna, M. A., T. P. Ellis, L. L. Marnell, T. M. Mittelmeier, andC. L. Dieckmann, 2002 Cbp1 is required for translation of themitochondrial cytochrome b mRNA of Saccharomyces cerevisiae.J. Biol. Chem. 277: 37987–37990.
Jackson, J. S. Jr., S. S. Houshmandi, F. Lopez Leban, and W. M.Olivas, 2004 Recruitment of the Puf3 protein to its mRNAtarget for regulation of mRNA decay in yeast. RNA 10: 1625–1636.
Jambhekar, A., and A. Amon, 2008 Control of meiosis by respira-tion. Curr. Biol. 18: 969–975.
Jan, P.-S., K. Esser, E. Pratje, and G. Michaelis, 2000 Som1, a thirdcomponent of the yeast mitochondrial inner membrane pepti-dase complex that contains Imp1 and Imp2. Mol. Gen. Genet.263: 483–491.
Jia, L., M. Dienhart, M. Schramp, M. McCauley, K. Hell et al.,2003 Yeast Oxa1 interacts with mitochondrial ribosomes: theimportance of the C-terminal region of Oxa1. EMBO J. 22:6438–6447.
Jia, L., M. K. Dienhart, and R. A. Stuart, 2007 Oxa1 directly in-teracts with Atp9 and mediates its assembly into the mitochon-drial F1Fo-ATP synthase complex. Mol. Biol. Cell 18: 1897–1908.
Jia, L., J. Kaur, and R. A. Stuart, 2009 Mapping the yeast Oxa1-mitochondrial ribosome interface and identification of MrpL40,a ribosomal protein in close proximity to Oxa1 and critical foroxidative phosphorylation complex assembly. Eukaryot. Cell 8:1792–1802.
Kang, P.-J., J. Ostermann, J. Shilling, W. Neupert, E. A. Craig et al.,1990 Requirement for hsp70 in the mitochondrial matrix fortranslocation and folding of precursor proteins. Nature 348:137–142.
Karniely, S., N. Regev-Rudzki, and O. Pines, 2006 The prese-quence of fumarase is exposed to the cytosol during import intomitochondria. J. Mol. Biol. 358: 396–405.
Kaufman, B. A., S. M. Newman, R. L. Hallberg, C. A. Slaughter, P. S.Perlman et al., 2000 In organello formaldehyde crosslinking ofproteins to mtDNA: identification of bifunctional proteins. Proc.Natl. Acad. Sci. USA 97: 7772–7777.
Kellems, R. E., V. F. Allison, and R. A. Butow, 1975 Cytoplasmictype 80S ribosomes associated with yeast mitochondria IV: at-tachment of ribosomes to the outer membrane of isolated mito-chondria. J. Cell Biol. 65: 1–14.
Kemper, C., S. J. Habib, G. Engl, P. Heckmeyer, K. S. Dimmer et al.,2008 Integration of tail-anchored proteins into the mitochon-drial outer membrane does not require any known import com-ponents. J. Cell Sci. 121: 1990–1998.
Kerscher, O., J. Holder, M. Srinivasan, R. S. Leung, and R. E. Jensen,1997 The Tim54p-Tim22p complex mediates insertion of pro-teins into the mitochondrial inner membrane. J. Cell Biol. 139:1663–1675.
Khalimonchuk, O., and G. Rodel, 2005 Biogenesis of cytochromec oxidase. Mitochondrion 5: 363–388.
Khalimonchuk, O., A. Bird, and D. R. Winge, 2007 Evidence fora Pro-oxidant Intermediate in the Assembly of Cytochrome Ox-idase. J. Biol. Chem. 282: 17442–17449.
Khalimonchuk, O., M. Bestwick, B. Meunier, T. C. Watts, and D. R.Winge, 2010 Formation of the redox cofactor centers during
1228 T. D. Fox

Cox1 maturation in yeast cytochrome oxidase. Mol. Cell. Biol.30: 1004–1017.
Kispal, G., P. Csere, C. Prohl, and R. Lill, 1999 The mitochondrialproteins Atm1p and Nfs1p are essential for biogenesis of cyto-solic Fe/S proteins. EMBO J. 18: 3981–3989.
Kispal, G., K. Sipos, H. Lange, Z. Fekete, T. Bedekovics et al.,2005 Biogenesis of cytosolic ribosomes requires the essentialiron-sulphur protein Rli1p and mitochondria. EMBO J. 24: 589–598.
Klöppel, C., Y. Suzuki, K. Kojer, C. Petrungaro, S. Longen et al.,2011 Mia40-dependent oxidation of cysteines in domain I ofCcs1 controls its distribution between mitochondria and thecytosol. Mol. Biol. Cell 22: 3749–3757.
Koehler, C. M., and H. L. Tienson, 2009 Redox regulation of pro-tein folding in the mitochondrial intermembrane space. Bio-chim. Biophys. Acta 1793: 139–145.
Koehler, C. M., E. Jarosch, K. Tokatlidis, K. Schmid, R. J. Schweyenet al., 1998 Import of mitochondrial carriers mediated by es-sential proteins of the intermembrane space. Science 279: 369–373.
Kornmann, B., E. Currie, S. R. Collins, M. Schuldiner, J. Nunnariet al., 2009 An ER-mitochondria tethering complex revealedby a synthetic biology screen. Science 325: 477–481.
Kovermann, P., K. N. Truscott, B. Guiard, P. Rehling, N. B. Sepuriet al., 2002 Tim22, the essential core of the mitochondrialprotein insertion complex, forms a voltage-activated and sig-nal-gated channel. Mol. Cell 9: 363–373.
Krause, K., R. Lopes de Souza, D. G. Roberts, and C. L. Dieckmann,2004 The mitochondrial message-specific mRNA protectorsCbp1 and Pet309 are associated in a high-molecular weightcomplex. Mol. Biol. Cell 15: 2674–2683.
Krimmer, T., D. Rapaport, M. T. Ryan, C. Meisinger, C. K. Kassenbrocket al., 2001 Biogenesis of porin of the outer mitochondrialmembrane involves an import pathway via receptors and thegeneral import pore of the TOM complex. J. Cell Biol. 152:289–300.
Kucej, M., B. Kucejova, R. Subramanian, X. J. Chen, and R. A.Butow, 2008 Mitochondrial nucleoids undergo remodeling inresponse to metabolic cues. J. Cell Sci. 121: 1861–1868.
Kühl, I., L. Dujeancourt, M. Gaisne, C. J. Herbert, and N. Bonnefoy,2011 A genome wide study in fission yeast reveals nine PPRproteins that regulate mitochondrial gene expression. NucleicAcids Res. 39: 8029–8041.
Künkele, K. P., P. Juin, C. Pompa, F. E. Nargang, J. P. Henry et al.,1998 The isolated complex of the translocase of the outermembrane of mitochondria. Characterization of the cation-selective and voltage-gated preprotein-conducting pore. J. Biol.Chem. 273: 31032–31039.
Kurz, M., H. Martin, J. Rassow, N. Pfanner, and M. T. Ryan,1999 Biogenesis of Tim proteins of the mitochondrial carrierimport pathway: differential targeting mechanisms and crossingover with the main import pathway. Mol. Biol. Cell 10: 2461–2474.
Kutik, S., D. Stojanovski, L. Becker, T. Becker, M. Meinecke et al.,2008 Dissecting membrane insertion of mitochondrial beta-barrel proteins. Cell 132: 1011–1024.
Kutik, S., D. A. Stroud, N. Wiedemann, and N. Pfanner,2009 Evolution of mitochondrial protein biogenesis. Biochim.Biophys. Acta 1790: 409–415.
Lange, C., and C. Hunte, 2002 Crystal structure of the yeast cy-tochrome bc1 complex with its bound substrate cytochrome c.Proc. Natl. Acad. Sci. USA 99: 2800–2805.
Lefebvre-Legendre, L., J. Vaillier, H. Benabdelhak, J. Velours, P. P.Slonimski et al., 2001 Identification of a nuclear gene (FMC1)required for the assembly/stability of yeast mitochondrial F(1)-ATPase in heat stress conditions. J. Biol. Chem. 276: 6789–6796.
Lelandais, G., Y. Saint-Georges, C. Geneix, L. Al-Shikhley, G. Dujardinet al., 2009 Spatio-temporal dynamics of yeast mitochondrialbiogenesis: transcriptional and post-transcriptional mRNA oscilla-tory modules. PLOS Comput. Biol. 5: e1000409.
Lemaire, C., P. Hamel, J. Velours, and G. Dujardin, 2000 Absenceof the mitochondrial AAA protease Yme1p restores ATPase F0subunit accumulation in an oxa1 deletion mutant of Saccharo-myces cerevisiae. J. Biol. Chem. 275: 23471–23475.
Leonhardt, S. A., K. Fearon, P. N. Danese, and T. L. Mason,1993 HSP78 encodes a yeast mitochondrial heat shock proteinin the clp family of ATP-dependent proteases. Mol. Cell. Biol. 13:6304–6313.
Lill, R., 2009 Function and biogenesis of iron-sulphur proteins.Nature 460: 831–838.
Lipinski, K. A., A. Kaniak-Golik, and P. Golik, 2010 Maintenanceand expression of the S. cerevisiae mitochondrial genome: fromgenetics to evolution and systems biology. Biochim. Biophys.Acta 1797: 1086–1098.
Lipinski, K. A., O. Puchta, V. Surandranath, M. Kudla, and P.Golik, 2011 Revisiting the yeast PPR proteins: applicationof an iterative hidden Markov model algorithm reveals newmembers of the rapidly evolving family. Mol. Biol. Evol. 28:2935–2948.
Lithgow, T., 2000 Targeting of proteins to mitochondria. FEBSLett. 476: 22–26.
Liu, J. M., and D. R. Liu, 2007 Discovery of a mRNA mitochon-drial localization element in Saccharomyces cerevisiae by nonho-mologous random recombination and in vivo selection. NucleicAcids Res. 35: 6750–6761.
Lode, A., M. Kuschel, C. Paret, and G. Rödel, 2000 Mitochondrialcopper metabolism in yeast: interaction between Sco1p andCox2p. FEBS Lett. 485: 19–24.
Lu, H., S. Allen, L. Wardleworth, P. Savory, and K. Tokatlidis,2004 Functional TIM10 chaperone assembly is redox-regulatedin vivo. J. Biol. Chem. 279: 18952–18958.
Ludlam, A., J. Brunzelle, T. Pribyl, X. Xu, D. L. Gatti et al.,2009 Chaperones of F1-ATPase. J. Biol. Chem. 284: 17138–17146.
Lueder, F., and T. Lithgow, 2009 The three domains of the mito-chondrial outer membrane protein Mim1 have discrete functionsin assembly of the TOM complex. FEBS Lett. 583: 1475–1480.
Lupo, D., C. Vollmer, M. Deckers, D. U. Mick, I. Tews et al.,2011 Mdm38 is a 14–3-3-like receptor and associates withthe protein synthesis machinery at the inner mitochondrialmembrane. Traffic 12: 1457–1456.
Lutz, T., W. Neupert, and J. M. Herrmann, 2003 Import of smallTim proteins into the mitochondrial intermembrane space.EMBO J. 22: 4400–4408.
Maccecchini, M. L., Y. Rudin, G. Blobel, and G. Schatz, 1979 Im-port of proteins into mitochondria: precursor forms of the ex-tramitochondrially made F1-ATPase subunits in yeast. Proc.Natl. Acad. Sci. USA 76: 343–347.
Mannella, C. A., D. R. Pfeiffer, P. C. Bradshaw, I. I. Moraru, B.Slepchenko et al., 2001 Topology of the mitochondrial innermembrane: dynamics and bioenergetic implications. IUBMB Life52: 93–100.
Manning-Krieg, U. C., P. E. Scherer, and G. Schatz, 1991 Se-quential action of mitochondrial chaperones in protein importinto the matrix. EMBO J. 10: 3273–3280.
Manthey, G. M., and J. E. McEwen, 1995 The product of thenuclear gene PET309 is required for translation of maturemRNA and stability or production of intron-containing RNAsderived from the mitochondrial COX1 locus of Saccharomycescerevisiae. EMBO J. 14: 4031–4043.
Marc, P., A. Margeot, F. Devaux, C. Blugeon, M. Corral-Debrinskiet al., 2002 Genome-wide analysis of mRNAs targeted to yeastmitochondria. EMBO Rep. 3: 159–164.
Mitochondrial Proteins 1229

Margeot, A., C. Blugeon, J. Sylvestre, S. Vialette, C. Jacq et al.,2002 In Saccharomyces cerevisiae, ATP2 mRNA sorting to thevicinity of mitochondria is essential for respiratory function.EMBO J. 21: 6893–6904.
Margeot, A., M. Garcia, W. Wang, E. Tetaud, J. P. di Rago et al.,2005 Why are many mRNAs translated to the vicinity ofmitochondria: a role in protein complex assembly? Gene 354:64–71.
Markov, D. A., M. Savkina, M. Anikin, M. Del Campo, K. Ecker et al.,2009 Identification of proteins associated with the yeast mito-chondrial RNA polymerase by tandem affinity purification. Yeast26: 423–440.
Marom, M., A. Azem, and D. Mokranjac, 2011a Understandingthe molecular mechanism of protein translocation across themitochondrial inner membrane: still a long way to go. Biochim.Biophys. Acta 1808: 990–1001.
Marom, M., D. Dayan, K. Demishtein-Zohary, D. Mokranjac, W.Neupert et al., 2011b Direct interaction of mitochondrial tar-geting presequences with purified components of the TIM23complex. J. Biol. Chem. 286: 43809–43815.
Marykwas, D. L., and T. D. Fox, 1989 Control of the Saccharomy-ces cerevisiae regulatory gene PET494: transcriptional repressionby glucose and translational induction by oxygen. Mol. Cell.Biol. 9: 484–491.
Marzuki, S., and A. R. Hibbs, 1986 Are all mitochondrial trans-lation products synthesized on membrane-bound ribosomes?Biochim. Biophys. Acta 866: 120–124.
Mathieu, L., M. Bourens, S. Marsy, O. Hlavacek, C. Panozzo et al.,2010 A mutational analysis reveals new functional interac-tions between domains of the Oxa1 protein in Saccharomycescerevisiae. Mol. Microbiol. 75: 474–488.
Mathieu, L., S. Marsy, Y. Saint-Georges, C. Jacq, and G. Dujardin,2011 A transcriptome screen in yeast identifies a novel assem-bly factor for the mitochondrial complex III. Mitochondrion 11:391–396.
Meeusen, S., and J. Nunnari, 2003 Evidence for a two membrane-spanning autonomous mitochondrial DNA replisome. J. CellBiol. 163: 503–510.
Meeusen, S., Q. Tieu, E. Wong, E. Weiss, D. Schieltz et al.,1999 Mgm101p is a novel component of the mitochondrialnucleoid that binds DNA and is required for the repair of oxida-tively damaged mitochondrial DNA. J. Cell Biol. 145: 291–304.
Meinecke, M., R. Wagner, P. Kovermann, B. Guiard, D. U. Micket al., 2006 Tim50 maintains the permeability barrier of themitochondrial inner membrane. Science 312: 1523–1526.
Mesecke, N., N. Terziyska, C. Kozany, F. Baumann, W. Neupertet al., 2005 A disulfide relay system in the intermembranespace of mitochondria that mediates protein import. Cell 121:1059–1069.
Messens, J., and J. F. Collet, 2006 Pathways of disulfide bondformation in Escherichia coli. Int. J. Biochem. Cell Biol. 38:1050–1062.
Michon, T., M. Galante, and J. Velours, 1988 NH2-terminal se-quence of the isolated yeast ATP synthase subunit 6 revealspost-translational cleavage. Eur. J. Biochem. 172: 621–625.
Mick, D. U., K. Wagner, M. van der Laan, A. E. Frazier, I. Perschilet al., 2007 Shy1 couples Cox1 translational regulation tocytochrome c oxidase assembly. EMBO J. 26: 4347–4358.
Mick, D. U., M. Vukotic, H. Piechura, H. E. Meyer, B. Warscheidet al., 2010 Coa3 and Cox14 are essential for negative feed-back regulation of COX1 translation in mitochondria. J. CellBiol. 191: 141–154.
Mick, D. U., T. D. Fox, and P. Rehling, 2011 Inventory control:cytochrome c oxidase assembly regulates mitochondrial trans-lation. Nat. Rev. Mol. Cell Biol. 12: 14–20.
Milenkovic, D., T. Ramming, J. M. Muller, L. S. Wenz, N. Gebertet al., 2009 Identification of the signal directing Tim9 and
Tim10 into the intermembrane space of mitochondria. Mol. Biol.Cell 20: 2530–2539.
Model, K., C. Meisinger, T. Prinz, N. Wiedemann, K. N. Truscottet al., 2001 Multistep assembly of the protein import channelof the mitochondrial outer membrane. Nat. Struct. Biol. 8: 361–370.
Mokranjac, D., and W. Neupert, 2009 Thirty years of proteintranslocation into mitochondria: unexpectedly complex and stillpuzzling. Biochim. Biophys. Acta 1793: 33–41.
Mokranjac, D., D. Popov-Celeketić, K. Hell, and W. Neupert,2005 Role of Tim21 in mitochondrial translocation contactsites. J. Biol. Chem. 280: 23437–23440.
Mokranjac, D., M. Sichting, D. Popov-Celeketić, K. Mapa, L.Gevorkyan-Airapetov et al., 2009 Role of Tim50 in the transferof precursor proteins from the outer to the inner membrane ofmitochondria. Mol. Biol. Cell 20: 1400–1407.
Naithani, S., S. A. Saracco, C. A. Butler, and T. D. Fox, 2003 In-teractions among COX1, COX2 and COX3 mRNA-specific trans-lational activator proteins on the inner surface of the mitochondrialinner membrane of Saccharomyces cerevisiae. Mol. Biol. Cell 14:324–333.
Naoe, M., Y. Ohwa, D. Ishikawa, C. Ohshima, S. Nishikawa et al.,2004 Identification of Tim40 that mediates protein sorting tothe mitochondrial intermembrane space. J. Biol. Chem. 279:47815–47821.
Neupert, W., 1997 Protein import into mitochondria. Annu. Rev.Biochem. 66: 863–917.
Neupert, W., and J. M. Herrmann, 2007 Translocation of proteinsinto mitochondria. Annu. Rev. Biochem. 76: 723–749.
Nevo-Dinur, K., A. Nussbaum-Shochat, S. Ben-Yehuda, and O. Amster-Choder, 2011 Translation-independent localization of mRNA inE. coli. Science 331: 1081–1084.
Nobrega, F. G., M. P. Nobrega, and A. Tzagoloff, 1992 BCS1,a novel gene required for the expression of functional Rieskeiron-sulfur protein in Saccharomyces cerevisiae. EMBO J. 11:3821–3829.
Nouet, C., M. Bourens, O. Hlavacek, S. Marsy, C. Lemaire et al.,2007 Rmd9p controls the processing/stability of mitochon-drial mRNAs and its overexpression compensates for a partialdeficiency of Oxa1p in Saccharomyces cerevisiae. Genetics 175:1105–1115.
Nowikovsky, K., E. M. Froschauer, G. Zsurka, J. Samaj, S. Reipertet al., 2004 The LETM1/YOL027 gene family encodes a factorof the mitochondrial K+ homeostasis with a potential role inthe Wolf-Hirschhorn syndrome. J. Biol. Chem. 279: 30307–30315.
Nunnari, J., and A. Suomalainen, 2012 Mitochondria: in sicknessand in health. Cell 148: 1145–1159.
Nunnari, J., T. D. Fox, and P. Walter, 1993 A mitochondrial pro-tease with two catalytic subunits of nonoverlapping specificities.Science 262: 1997–2004.
Nunnari, J., W. F. Marshall, A. Straight, A. Murray, J. W. Sedatet al., 1997 Mitochondrial transmission during mating in Sac-charomyces cerevisiae is determined by mitochondrial fusion andfission and the intramitochondrial segregation of mitochondrialDNA. Mol. Biol. Cell 8: 1233–1242.
Olivas, W., and R. Parker, 2000 The Puf3 protein is a transcript-specific regulator of mRNA degradation in yeast. EMBO J. 19:6602–6611.
Osman, C., C. Wilmes, T. Tatsuta, and T. Langer, 2007 Prohibitinsinteract genetically with Atp23, a novel processing peptidaseand chaperone for the F1Fo-ATP synthase. Mol. Biol. Cell 18:627–635.
Ostrander, D. B., M. Zhang, E. Mileykovskaya, M. Rho, and W.Dowhan, 2001 Lack of mitochondrial anionic phospholipidscauses an inhibition of translation of protein components ofthe electron transport chain. A yeast genetic model system for
1230 T. D. Fox

the study of anionic phospholipid function in mitochondria. J.Biol. Chem. 276: 25262–25272.
Ott, M., and J. M. Herrmann, 2010 Co-translational membraneinsertion of mitochondrially encoded proteins. Biochim. Bio-phys. Acta 1803: 767–775.
Ott, M., M. Prestele, H. Bauerschmitt, S. Funes, N. Bonnefoy et al.,2006 Mba1, a membrane-associated ribosome receptor in mi-tochondria. EMBO J. 25: 1603–1610.
Pais, J. E., B. Schilke, and E. A. Craig, 2011 Reevaluation of therole of the Pam18:Pam16 interaction in translocation of proteinsby the mitochondrial Hsp70-based import motor. Mol. Biol. Cell22: 4740–4749.
Palmieri, F., G. Agrimi, E. Blanco, A. Castegna, M. A. Di Noia et al.,2006 Identification of mitochondrial carriers in Saccharomy-ces cerevisiae by transport assay of reconstituted recombinantproteins. Biochim. Biophys. Acta 1757: 1249–1262.
Papic, D., K. Krumpe, J. Dukanovic, K. S. Dimmer, and D. Rapaport,2011 Multispan mitochondrial outer membrane protein Ugo1follows a unique Mim1-dependent import pathway. J. Cell Biol.194: 397–405.
Paschen, S. A., U. Rothbauer, K. Kaldi, M. F. Bauer, W. Neupertet al., 2000 The role of the TIM8–13 complex in the importof Tim23 into mitochondria. EMBO J. 19: 6392–6400.
Paschen, S. A., T. Waizenegger, T. Stan, M. Preuss, M. Cyrklaffet al., 2003 Evolutionary conservation of biogenesis of beta-barrel membrane proteins. Nature 426: 862–866.
Payne, M. J., P. M. Finnegan, P. M. Smooker, and H. B. Lukins,1993 Characterization of a second nuclear gene, AEP1, re-quired for expression of the mitochondrial OLI1 gene in Saccha-romyces cerevisiae. Curr. Genet. 24: 126–135.
Peixoto, P. M., F. Grana, T. J. Roy, C. D. Dunn, M. Flores et al.,2007 Awaking TIM22, a dynamic ligand-gated channel forprotein insertion in the mitochondrial inner membrane. J. Biol.Chem. 282: 18694–18701.
Perez-Martinez, X., S. A. Broadley, and T. D. Fox, 2003 Mss51ppromotes mitochondrial Cox1p synthesis and interacts withnewly synthesized Cox1p. EMBO J. 22: 5951–5961.
Perez-Martinez, X., C. A. Butler, M. Shingu-Vazquez, and T. D. Fox,2009 Dual functions of Mss51 couple synthesis of Cox1 toassembly of cytochrome c oxidase in Saccharomyces cerevisiaemitochondria. Mol. Biol. Cell 20: 4371–4380.
Pierrel, F., M. L. Bestwick, P. A. Cobine, O. Khalimonchuk, J. A.Cricco et al., 2007 Coa1 links the Mss51 post-translationalfunction to Cox1 cofactor insertion in cytochrome c oxidaseassembly. EMBO J. 26: 4335–4346.
Pierrel, F., O. Khalimonchuk, P. A. Cobine, M. Bestwick, and D. R.Winge, 2008 Coa2 is an assembly factor for yeast cytochromec oxidase biogenesis facilitating the maturation of Cox1. Mol.Cell. Biol. 28: 4927–4939.
Pon, L., and G. Schatz, 1991 Biogenesis of yeast mitochondria, pp.333–406 in The Molecular and Cellular Biology of the Yeast Sac-charomyces: Genome Dynamics, Protein Synthesis and Energetics,edited by J. R. Broach, J. R. Pringle, and E. W. Jones. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY.
Pon, L., T. Moll, D. Vestweber, B. Marshallsay, and G. Schatz,1989 Protein import into mitochondria: ATP-dependent pro-tein translocation activity in a submitochondrial fraction en-riched in membrane contact sites and specific proteins. J. CellBiol. 109: 2603–2616.
Popov-�Celeketić, D., K. Mapa, W. Neupert, and D. Mokranjac,2008a Active remodelling of the TIM23 complex during trans-location of preproteins into mitochondria. EMBO J. 27: 1469–1480.
Popov-�Celeketić, J., T. Waizenegger, and D. Rapaport, 2008b Mim1functions in an oligomeric form to facilitate the integration ofTom20 into the mitochondrial outer membrane. J. Mol. Biol.376: 671–680.
Popov-�Celeketić, D., K. Waegemann, K. Mapa, W. Neupert, and D.Mokranjac, 2011 Role of the import motor in insertion oftransmembrane segments by the mitochondrial TIM23 complex.EMBO Rep. 12: 542–548.
Pratje, E., G. Mannhaupt, G. Michaelis, and K. Beyreuther, 1983 Anuclear mutation prevents processing of a mitochondrially en-coded membrane protein in Saccharomyces cerevisiae. EMBO J.2: 1049–1054.
Premsler, T., R. P. Zahedi, U. Lewandrowski, and A. Sickmann,2009 Recent advances in yeast organelle and membrane pro-teomics. Proteomics 9: 4731–4743.
Prestele, M., F. Vogel, A. S. Reichert, J. M. Herrmann, and M. Ott,2009 Mrpl36 is important for generation of assembly compe-tent proteins during mitochondrial translation. Mol. Biol. Cell20: 2615–2625.
Preuss, M., K. Leonhard, K. Hell, R. A. Stuart, W. Neupert et al.,2001 Mba1, a novel component of the mitochondrial proteinexport machinery of the yeast Saccharomyces cerevisiae. J. CellBiol. 153: 1085–1096.
Prokisch, H., C. Scharfe, D. G. Camp II. W. Xiao, L. David et al.,2004 Integrative analysis of the mitochondrial proteome inyeast. PLoS Biol. 2: 795–804.
Quenault, T., T. Lithgow, and A. Traven, 2011 PUF proteins: re-pression, activation and mRNA localization. Trends Cell Biol.21: 104–112.
Rabl, R., V. Soubannier, R. Scholz, F. Vogel, N. Mendl et al.,2009 Formation of cristae and crista junctions in mitochondriadepends on antagonism between Fcj1 and Su e/g. J. Cell Biol.185: 1047–1063.
Rada, P., P. Dolezal, P. L. Jedelsky, D. Bursac, A. J. Perry et al.,2011 The core components of organelle biogenesis and mem-brane transport in the hydrogenosomes of Trichomonas vagina-lis. PLoS ONE 6: e24428.
Rak, M., and A. Tzagoloff, 2009 F1-dependent translation of mi-tochondrially encoded Atp6p and Atp8p subunits of yeast ATPsynthase. Proc. Natl. Acad. Sci. USA 106: 18509–18514.
Rak, M., X. Zeng, J. J. Briere, and A. Tzagoloff, 2009 Assembly ofF0 in Saccharomyces cerevisiae. Biochim. Biophys. Acta 1793:108–116.
Rak, M., S. Gokova, and A. Tzagoloff, 2011 Modular assembly ofyeast mitochondrial ATP synthase. EMBO J. 30: 920–930.
Ramage, L., T. Junne, K. Hahne, T. Lithgow, and G. Schatz,1993 Functional cooperation of mitochondrial protein importreceptors in yeast. EMBO J. 12: 4115–4123.
Rehling, P., K. Model, K. Brandner, P. Kovermann, A. Sickmannet al., 2003 Protein insertion into the mitochondrial innermembrane by a twin-pore translocase. Science 299: 1747–1751.
Reif, S., O. Randelj, G. Domanska, E. A. Dian, T. Krimmer et al.,2005 Conserved mechanism of Oxa1 insertion into the mito-chondrial inner membrane. J. Mol. Biol. 354: 520–528.
Reinders, J., R. P. Zahedi, N. Pfanner, C. Meisinger, and A. Sickmann,2006 Toward the complete yeast mitochondrial proteome: mul-tidimensional separation techniques for mitochondrial proteo-mics. J. Proteome Res. 5: 1543–1554.
Riemer, J., M. Fischer, and J. M. Herrmann, 2011 Oxidation-driven protein import into mitochondria: insights and blindspots. Biochim. Biophys. Acta 1808: 981–989.
Riezman, H., R. Hay, S. Gasser, G. Daum, G. Schneider et al.,1983 The outer membrane of yeast mitochondria: isolationof outside-out sealed vesicles. EMBO J. 2: 1105–1111.
Rigby, K., P. A. Cobine, O. Khalimonchuk, and D. R. Winge,2008 Mapping the functional interaction of Sco1 and Cox2in cytochrome oxidase biogenesis. J. Biol. Chem. 283: 15015–15022.
Rodeheffer, M. S., and G. S. Shadel, 2003 Multiple interactionsinvolving the amino-terminal domain of yeast mtRNA polymer-
Mitochondrial Proteins 1231

ase determine the efficiency of mitochondrial protein synthesis.J. Biol. Chem. 278: 18695–18701.
Rodeheffer, M. S., B. E. Boone, A. C. Bryan, and G. S. Shadel,2001 Nam1p, a protein involved in RNA processing and trans-lation, is coupled to transcription through an interaction withyeast mitochondrial RNA polymerase. J. Biol. Chem. 276: 8616–8622.
Rödel, G., 1997 Translational activator proteins required for cy-tochrome b synthesis in Saccharomyces cerevisiae. Curr. Genet.31: 375–379.
Rouillard, J. M., M. E. Dufour, B. Theunissen, E. Mandart, G.Dujardin et al., 1996 SLS1, a new Saccharomyces cerevisiaegene involved in mitochondrial metabolism, isolated as a syn-thetic lethal in association with an SSM4 deletion. Mol. Gen.Genet. 252: 700–708.
Saint-Georges, Y., M. Garcia, T. Delaveau, L. Jourdren, S. Le Cromet al., 2008 Yeast mitochondrial biogenesis: a role for the PUFRNA-binding protein Puf3p in mRNA localization. PLoS ONE 3:e2293.
Saldanha, R., G. Mohr, M. Belfort, and A. M. Lambowitz,1993 Group I and group II introns. FASEB J. 7: 15–24.
Sanchirico, M. E., 1998 Understanding Mitochondrial BiogenesisThrough Gene Relocation. Ph.D. Thesis, University of Massachu-setts, Amherst, MA.
Sanchirico, M. E., T. D. Fox, and T. L. Mason, 1998 Accumulationof mitochondrially synthesized Saccharomyces cerevisiae Cox2pand Cox3p depends on targeting information in untranslatedportions of their mRNAs. EMBO J. 17: 5796–5804.
Saracco, S. A., and T. D. Fox, 2002 Cox18p is required for exportof the mitochondrially encoded Saccharomyces cerevisiae Cox2pC-tail, and interacts with Pnt1p and Mss2p in the inner mem-brane. Mol. Biol. Cell 13: 1122–1131.
Sasarman, F., C. Brunel-Guitton, H. Antonicka, T. Wai, E. A.Shoubridge et al., 2010 LRPPRC and SLIRP interact in a ribo-nucleoprotein complex that regulates posttranscriptional geneexpression in mitochondria. Mol. Biol. Cell 21: 1315–1323.
Sass, E., S. Karniely, and O. Pines, 2003 Folding of fumarase dur-ing mitochondrial import determines its dual targeting in yeast.J. Biol. Chem. 278: 45109–45116.
Schatz, G., 1968 Impaired binding of mitochondrial adenosinetriphosphatase in the cytoplasmic “petite” mutant of Saccharo-myces cerevisiae. J. Biol. Chem. 243: 2192–2199.
Schmidt, O., N. Pfanner, and C. Meisinger, 2010 Mitochondrialprotein import: from proteomics to functional mechanisms.Nat. Rev. Mol. Cell Biol. 11: 655–667.
Schmidt, O., A. B. Harbauer, S. Rao, B. Eyrich, R. P. Zahedi et al.,2011 Regulation of mitochondrial protein import by cytosolickinases. Cell 144: 227–239.
Schneider, A., D. Bursac, and T. Lithgow, 2008 The direct route:a simplified pathway for protein import into the mitochondrionof trypanosomes. Trends Cell Biol. 18: 12–18.
Schulz, C., O. Lytovchenko, J. Melin, A. Chacinska, B. Guiard et al.,2011 Tim50’s presequence receptor domain is essential forsignal driven transport across the TIM23 complex. J. Cell Biol.195: 643–656.
Sherman, E. L., R. D. Taylor, N. E. Go, and F. E. Nargang,2006 Effect of mutations in Tom40 on stability of the translo-case of the outer mitochondrial membrane (TOM) complex,assembly of Tom40, and import of mitochondrial preproteins.J. Biol. Chem. 281: 22554–22565.
Sherman, F., 2005 The importance of mutation, then and now:studies with yeast cytochrome c. Mutat. Res. 589: 1–16.
Shingu-Vazquez, M., Y. Camacho-Villasana, L. Sandoval-Romero, C.A. Butler, T. D. Fox et al., 2010 The carboxyl-terminal end ofCox1 is required for feedback-assembly regulation of Cox1 syn-thesis in Saccharomyces cerevisiae mitochondria. J. Biol. Chem.285: 34382–34389.
Shiota, T., H. Mabuchi, S. Tanaka-Yamano, K. Yamano, and T.Endo, 2011 In vivo protein-interaction mapping of a mitochon-drial translocator protein Tom22 at work. Proc. Natl. Acad. Sci.USA 108: 15179–15183.
Sickmann, A., J. Reinders, Y. Wagner, C. Joppich, R. Zahedi et al.,2003 The proteome of Saccharomyces cerevisiae mitochondria.Proc. Natl. Acad. Sci. USA 100: 13207–13212.
Sideris, D. P., N. Petrakis, N. Katrakili, D. Mikropoulou, A. Galloet al., 2009 A novel intermembrane space-targeting signaldocks cysteines onto Mia40 during mitochondrial oxidative fold-ing. J. Cell Biol. 187: 1007–1022.
Sirrenberg, C., M. F. Bauer, B. Guiard, W. Neupert, and M. Brunner,1996 Import of carrier proteins into the mitochondrial innermembrane mediated by Tim22. Nature 384: 582–585.
Sirrenberg, C., M. Endres, H. Folsch, R. A. Stuart, W. Neupert et al.,1998 Carrier protein import into mitochondria mediated bythe intermembrane proteins Tim10/Mrs11 and Tim12/Mrs5.Nature 391: 912–915.
Slutsky-Leiderman, O., M. Marom, O. Iosefson, R. Levy, S. Maozet al., 2007 The interplay between components of the mito-chondrial protein translocation motor studied using purifiedcomponents. J. Biol. Chem. 282: 33935–33942.
Smith, P. M., J. L. Fox, and D. R. Winge, 2012 Biogenesis of thecytochrome bc1 complex and role of assembly factors. Biochim.Biophys. Acta 1817: 276–286.
Solmaz, S. R., and C. Hunte, 2008 Structure of complex III withbound cytochrome c in reduced state and definition ofa minimal core interface for electron transfer. J. Biol. Chem.283: 17542–17549.
Soto, I. C., F. Fontanesi, J. Liu, and A. Barrientos, 2011 Biogenesisand assembly of eukaryotic cytochrome c oxidase catalytic core.Biochim. Biophys. Acta 1817: 883–897.
Steele, D. F., C. A. Butler, and T. D. Fox, 1996 Expression ofa recoded nuclear gene inserted into yeast mitochondrial DNAis limited by mRNA-specific translational activation. Proc. Natl.Acad. Sci. USA 93: 5253–5257.
Steiner, H., A. Zollner, A. Haid,W. Neupert, and R. Lill, 1995 Biogenesisof mitochondrial heme lyases in yeast. Import and foldingin the intermembrane space. J. Biol. Chem. 270: 22842–22849.
Stevens, B., 1981 Mitochondrial structure, pp. 471–504 in TheMolecular Biology of the Yeast Saccharomyces, Life Cycle and In-heritance, edited by J. N. Strathern, E. W. Jones, and J. R.Broach. Cold Spring Harbor Laboratory Press, Cold SpringHarbor, NY.
Stock, D., A. G. Leslie, and J. E. Walker, 1999 Molecular architectureof the rotary motor in ATP synthase. Science 286: 1700–1705.
Stojanovski, D., B. Guiard, V. Kozjak-Pavlovic, N. Pfanner, and C.Meisinger, 2007 Alternative function for the mitochondrialSAM complex in biogenesis of alpha-helical TOM proteins. J.Cell Biol. 179: 881–893.
Strathern, J. N., E. W. Jones, and J. R. Broach (Editors), 1982 TheMolecular Biology of the Yeast Saccharomyces: Metabolism andGene Expression. Cold Spring Harbor Laboratory Press, ColdSpring Harbor, NY.
Strogolova, V., A. Furness, M. Robb-McGrath, J. Garlich, and R. A.Stuart, 2012 Rcf1 and Rcf2, members of the hypoxia-inducedgene 1 protein family, are critical components of the mitochon-drial cytochrome bc1-cytochrome c oxidase supercomplex. Mol.Cell. Biol. 32: 1363–1373.
Stroud, D. A., T. Becker, J. Qiu, D. Stojanovski, S. Pfannschmidtet al., 2011 Biogenesis of mitochondrial beta-barrel proteins:the POTRA domain is involved in precursor release from theSAM complex. Mol. Biol. Cell 22: 2823–2833.
Stuart, R. A., 2008 Supercomplex organization of the oxidativephosphorylation enzymes in yeast mitochondria. J. Bioenerg.Biomembr. 40: 411–417.
1232 T. D. Fox

Suissa, M., and G. Schatz, 1982 Import of proteins into mitochon-dria. Translatable mRNAs for imported mitochondrial proteinsare present in free as well as mitochondria-bound cytoplasmicpolysomes. J. Biol. Chem. 257: 13048–13055.
Suppanz, I. E., C. A. Wurm, D. Wenzel, and S. Jakobs, 2009 Them-AAA protease processes cytochrome c peroxidase preferen-tially at the inner boundary membrane of mitochondria. Mol.Biol. Cell 20: 572–580.
Suzuki, C. K., M. Rep, J. M. van Dijl, K. Suda, L. A. Grivell et al.,1997 ATP-dependent proteases that also chaperone proteinbiogenesis. Trends Biochem. Sci. 22: 118–123.
Sylvestre, J., A. Margeot, C. Jacq, G. Dujardin, and M. Corral-Debrinski, 2003 The role of the 39 untranslated region inmRNA sorting to the vicinity of mitochondria is conserved fromyeast to human cells. Mol. Biol. Cell 14: 3848–3856.
Szyrach, G., M. Ott, N. Bonnefoy, W. Neupert, and J. M. Herrmann,2003 Ribosome binding to the Oxa1 complex facilitates co-translational protein insertion in mitochondria. EMBO J. 22:6448–6457.
Taanman, J. W., and R. A. Capaldi, 1992 Purification of yeastcytochrome c oxidase with a subunit composition resemblingthe mammalian enzyme. J. Biol. Chem. 267: 22481–22485.
Tamura, Y., Y. Harada, T. Shiota, K. Yamano, K. Watanabe et al.,2009 Tim23-Tim50 pair coordinates functions of translocatorsand motor proteins in mitochondrial protein import. J. Cell Biol.184: 129–141.
Tatsuta, T., and T. Langer, 2009 AAA proteases in mitochondria:diverse functions of membrane-bound proteolytic machines.Res. Microbiol. 160: 711–717.
Tavares-Carreon, F., Y. Camacho-Villasana, A. Zamudio-Ochoa, M.Shingu-Vazquez, A. Torres-Larios et al., 2008 The pentatrico-peptide repeats present in Pet309 are necessary for translationbut not for stability of the mitochondrial COX1 mRNA in yeast.J. Biol. Chem. 283: 1472–1479.
Taylor, A. B., B. S. Smith, S. Kitada, K. Kojima, H. Miyaura et al.,2001 Crystal structures of mitochondrial processing peptidasereveal the mode for specific cleavage of import signal sequences.Structure 9: 615–625.
Terpstra, P., E. Zanders, and R. A. Butow, 1979 The association ofvar1 with the 38S mitochondrial ribosomal subunit in yeast. J.Biol. Chem. 254: 12653–12661.
Terziyska, N., B. Grumbt, M. Bien, W. Neupert, J. M. Herrmannet al., 2007 The sulfhydryl oxidase Erv1 is a substrate of theMia40-dependent protein translocation pathway. FEBS Lett.581: 1098–1102.
Tienson, H. L., D. V. Dabir, S. E. Neal, R. Loo, S. A. Hasson et al.,2009 Reconstitution of the mia40-erv1 oxidative folding path-way for the small tim proteins. Mol. Biol. Cell 20: 3481–3490.
Towpik, J., 2005 Regulation of mitochondrial translation in yeast.Cell. Mol. Biol. Lett. 10: 571–594.
Trouillard, M., B. Meunier, and F. Rappaport, 2011 Questioningthe functional relevance of mitochondrial supercomplexes bytime-resolved analysis of the respiratory chain. Proc. Natl. Acad.Sci. USA 108: E1027–E1034.
Truscott, K. N., B. R. Lowth, P. R. Strack, and D. A. Dougan,2010 Diverse functions of mitochondrial AAA+ proteins: pro-tein activation, disaggregation, and degradation. Biochem. CellBiol. 88: 97–108.
Tsukihara, T., H. Aoyama, E. Yamashita, T. Tomizaki, H. Yamaguchiet al., 1995 Structures of metal sites of oxidized bovine heartcytochrome c oxidase at 2.8 Å. Science 269: 1069–1074.
Tsukihara, T., H. Aoyama, E. Yamashita, T. Tomizaki, H. Yamaguchiet al., 1996 The whole structure of the 13-subunit oxidizedcytochrome c oxidase at 2.8 Å. Science 272: 1136–1144.
Tu, B. P., A. Kudlicki, M. Rowicka, and S. L. McKnight, 2005 Logicof the yeast metabolic cycle: temporal compartmentalization ofcellular processes. Science 310: 1152–1158.
Tzagoloff, A., 1969 Assembly of the mitochondrial membrane sys-tem. II. Synthesis of the mitochondrial adenosine triphospha-tase. F1. J. Biol. Chem. 244: 5027–5033.
Tzagoloff, A., and C. L. Dieckmann, 1990 PET genes of Saccharo-myces cerevisiae. Microbiol. Rev. 54: 211–225.
Tzagoloff, A., A. Barrientos, W. Neupert, and J. M. Herrmann,2004 Atp10p assists assembly of Atp6p into the F0 unit ofthe yeast mitochondrial ATPase. J. Biol. Chem. 279: 19775–19780.
van der Laan, M., N. Wiedemann, D. U. Mick, B. Guiard, P. Rehlinget al., 2006 A role for Tim21 in membrane-potential-dependentpreprotein sorting in mitochondria. Curr. Biol. 16: 2271–2276.
van der Laan, M., M. Meinecke, J. Dudek, D. P. Hutu, M. Lind et al.,2007 Motor-free mitochondrial presequence translocase drivesmembrane integration of preproteins. Nat. Cell Biol. 9: 1152–1159.
van Wilpe, S., M. T. Ryan, K. Hill, A. C. Maarse, C. Meisinger et al.,1999 Tom22 is a multifunctional organizer of the mitochon-drial preprotein translocase. Nature 401: 485–489.
Vasiljev, A., U. Ahting, F. E. Nargang, N. E. Go, S. J. Habib et al.,2004 Reconstituted TOM core complex and Tim9/Tim10complex of mitochondria are sufficient for translocation ofthe ADP/ATP carrier across membranes. Mol. Biol. Cell 15:1445–1458.
Velours, J., and G. Arselin, 2000 The Saccharomyces cerevisiaeATP synthase. J. Bioenerg. Biomembr. 32: 383–390.
Velours, J., A. Dautant, B. Salin, I. Sagot, and D. Brethes,2009 Mitochondrial F1F0-ATP synthase and organellar inter-nal architecture. Int. J. Biochem. Cell Biol. 41: 1783–1789.
Velours, J., C. Stines-Chaumeil, J. Habersetzer, S. Chaignepain, A.Dautant et al., 2011 Evidence of the proximity of ATP synthasesubunits 6 (a) in the inner mitochondrial membrane and in thesupramolecular forms of Saccharomyces cerevisiae ATP syn-thase. J. Biol. Chem. 286: 35477–35484.
Vogel, F., C. Bornhovd, W. Neupert, and A. S. Reichert, 2006 Dy-namic subcompartmentalization of the mitochondrial innermembrane. J. Cell Biol. 175: 237–247.
Vögtle, F. N., S. Wortelkamp, R. P. Zahedi, D. Becker, C. Leidholdet al., 2009 Global analysis of the mitochondrial N-proteomeidentifies a processing peptidase critical for protein stability. Cell139: 428–439.
Vögtle, F. N., C. Prinz, J. Kellermann, F. Lottspeich, N. Pfanneret al., 2011 Mitochondrial protein turnover: role of the pre-cursor intermediate peptidase Oct1 in protein stabilization. Mol.Biol. Cell 22: 2135–2143.
von Ballmoos, C., A. Wiedenmann, and P. Dimroth, 2009 Es-sentials for ATP synthesis by F1F0 ATP synthases. Annu. Rev.Biochem. 78: 649–672.
von der Malsburg, K., J. M. Muller, M. Bohnert, S. Oeljeklaus, P.Kwiatkowska et al., 2011 Dual role of mitofilin in mitochon-drial membrane organization and protein biogenesis. Dev. Cell21: 694–707.
Voos, W., and K. Röttgers, 2002 Molecular chaperones as essen-tial mediators of mitochondrial biogenesis. Biochim. Biophys.Acta 1592: 51–62.
Voos, W., H. Martin, T. Krimmer, and N. Pfanner, 1999 Mechanismsof protein translocation into mitochondria. Biochim. Biophys.Acta 1422: 235–254.
Vukotic, M., S. Oeljeklaus, S. Wiese, F. N. Vogtle, C. Meisingeret al., 2012 Rcf1 mediates cytochrome oxidase assembly andrespirasome formation, revealing heterogeneity of the enzymecomplex. Cell Metab. 15: 336–347.
Wagener, N., M. Ackermann, S. Funes, and W. Neupert, 2011 Apathway of protein translocation in mitochondria mediated bythe AAA-ATPase Bcs1. Mol. Cell 44: 191–202.
Wagner, K., N. Gebert, B. Guiard, K. Brandner, K. N. Truscott et al.,2008 The assembly pathway of the mitochondrial carrier
Mitochondrial Proteins 1233

translocase involves four preprotein translocases. Mol. Cell.Biol. 28: 4251–4260.
Wagner, K., I. Perschil, C. D. Fichter, and M. van der Laan,2010 Stepwise assembly of dimeric F(1)F(o)-ATP synthase inmitochondria involves the small F(o)-subunits k and i. Mol. Biol.Cell 21: 1494–1504.
Waizenegger, T., S. Schmitt, J. Zivkovic, W. Neupert, and D. Rapaport,2005 Mim1, a protein required for the assembly of the TOMcomplex of mitochondria. EMBO Rep. 6: 57–62.
Wallis, M. G., O. Groudinsky, P. P. Slonimski, and G. Dujardin,1994 The NAM1 protein (NAM1p), which is selectively re-quired for cox1, cytb and atp6 transcript processing/stabilisa-tion, is located in the yeast mitochondrial matrix. Eur. J.Biochem. 222: 27–32.
Walther, D. M., and D. Rapaport, 2009 Biogenesis of mitochon-drial outer membrane proteins. Biochim. Biophys. Acta 1793:42–51.
Wang, X., M. E. Dumont, and F. Sherman, 1996 Sequence re-quirements for mitochondrial import of yeast cytochrome c. J.Biol. Chem. 271: 6594–6604.
Weraarpachai, W., H. Antonicka, F. Sasarman, J. Seeger, B. Schranket al., 2009 Mutation in TACO1, encoding a translational ac-tivator of COX I, results in cytochrome c oxidase deficiency andlate-onset Leigh syndrome. Nat. Genet. 41: 833–837.
Wiedemann, N., N. Pfanner, and M. T. Ryan, 2001 The threemodules of ADP/ATP carrier cooperate in receptor recruit-ment and translocation into mitochondria. EMBO J. 20: 951–960.
Wiedemann, N., V. Kozjak, A. Chacinska, B. Schonfisch, S. Rospertet al., 2003a Machinery for protein sorting and assembly in themitochondrial outer membrane. Nature 424: 565–571.
Wiedemann, N., V. Kozjak, T. Prinz, M. T. Ryan, C. Meisinger et al.,2003b Biogenesis of yeast mitochondrial cytochrome c: aunique relationship to the TOM machinery. J. Mol. Biol. 327:465–474.
Wiedemann, N., K. N. Truscott, S. Pfannschmidt, B. Guiard, C.Meisinger et al., 2004 Biogenesis of the protein import channelTom40 of the mitochondrial outer membrane: intermembranespace components are involved in an early stage of the assemblypathway. J. Biol. Chem. 279: 18188–18194.
Wiedemann, N., M. van der Laan, D. P. Hutu, P. Rehling, and N.Pfanner, 2007 Sorting switch of mitochondrial presequencetranslocase involves coupling of motor module to respiratorychain. J. Cell Biol. 179: 1115–1122.
Williams, E. H., and T. D. Fox, 2003 Antagonistic signals withinthe COX2 mRNA coding sequence control its translation in Sac-charomyces cerevisiae mitochondria. RNA 9: 419–431.
Williams, E. H., X. Perez-Martinez, and T. D. Fox, 2004 MrpL36p,a highly diverged L31 ribosomal protein homolog with addi-tional functional domains in Saccharomyces cerevisiae mitochon-dria. Genetics 167: 65–75.
Williams, E. H., N. Bsat, N. Bonnefoy, C. A. Butler, and T. D. Fox,2005 Alteration of a novel dispensable mitochondrial ribo-somal small subunit protein, Rsm28p, allows translation of de-fective COX2 mRNAs. Eukaryot. Cell 4: 337–354.
Williams, E. H., C. A. Butler, N. Bonnefoy, and T. D. Fox,2007 Translation initiation in Saccharomyces cerevisiae mito-chondria: functional interactions among mitochondrial ribosomalprotein Rsm28p, initiation factor 2, methionyl-tRNA-formyltrans-ferase, and novel protein Rmd9p. Genetics 175: 1117–1126.
Wurm, C. A., and S. Jakobs, 2006 Differential protein distribu-tions define two sub-compartments of the mitochondrial innermembrane in yeast. FEBS Lett. 580: 5628–5634.
Yamamoto, H., M. Esaki, T. Kanamori, Y. Tamura, S. Nishikawaet al., 2002 Tim50 is a subunit of the TIM23 complex thatlinks protein translocation across the outer and inner mitochon-drial membranes. Cell 111: 519–528.
Yamamoto, H., N. Itoh, S. Kawano, Y. Yatsukawa, T. Momose et al.,2011 Dual role of the receptor Tom20 in specificity and effi-ciency of protein import into mitochondria. Proc. Natl. Acad.Sci. USA 108: 91–96.
Yamano, K., Y. Yatsukawa, M. Esaki, A. E. Hobbs, R. E. Jensen et al.,2008 Tom20 and Tom22 share the common signal recognitionpathway in mitochondrial protein import. J. Biol. Chem. 283:3799–3807.
Yogev, O., and O. Pines, 2011 Dual targeting of mitochondrialproteins: mechanism, regulation and function. Biochim. Bio-phys. Acta 1808: 1012–1020.
Young, J. C., N. J. Hoogenraad, and F. U. Hartl, 2003 Molecularchaperones Hsp90 and Hsp70 deliver preproteins to the mito-chondrial import receptor Tom70. Cell 112: 41–50.
Young, L., K. Leonhard, T. Tatsuta, J. Trowsdale, and T. Langer,2001 Role of the ABC transporter Mdl1 in peptide export frommitochondria. Science 291: 2135–2138.
Zahedi, R. P., A. Sickmann, A. M. Boehm, C. Winkler, N. Zufallet al., 2006 Proteomic analysis of the yeast mitochondrial out-er membrane reveals accumulation of a subclass of preproteins.Mol. Biol. Cell 17: 1436–1450.
Zaman, S., S. I. Lippman, X. Zhao, and J. R. Broach, 2008 HowSaccharomyces responds to nutrients. Annu. Rev. Genet. 42:27–81.
Zambrano, A., F. Fontanesi, A. Solans, R. L. de Oliveira, T. D. Foxet al., 2007 Aberrant translation of cytochrome c oxidase sub-unit 1 mRNA species in the absence of Mss51p in the yeastSaccharomyces cerevisiae. Mol. Biol. Cell 18: 523–535.
Zara, V., I. Palmisano, L. Conte, and B. L. Trumpower, 2004 Fur-ther insights into the assembly of the yeast cytochrome bc1complex based on analysis of single and double deletion mu-tants lacking supernumerary subunits and cytochrome b. Eur. J.Biochem. 271: 1209–1218.
Zara, V., L. Conte, and B. L. Trumpower, 2007 Identification andcharacterization of cytochrome bc(1) subcomplexes in mito-chondria from yeast with single and double deletions of genesencoding cytochrome bc(1) subunits. FEBS J. 274: 4526–4539.
Zara, V., L. Conte, and B. L. Trumpower, 2009 Biogenesis of theyeast cytochrome bc1 complex. Biochim. Biophys. Acta 1793:89–96.
Zeng, X., R. Kucharczyk, J. P. di Rago, and A. Tzagoloff, 2007a Theleader peptide of yeast Atp6p is required for efficient interactionwith the Atp9p ring of the mitochondrial ATPase. J. Biol. Chem.282: 36167–36176.
Zeng, X., W. Neupert, and A. Tzagoloff, 2007b The metallopro-tease encoded by ATP23 has a dual function in processing andassembly of subunit 6 of mitochondrial ATPase. Mol. Biol. Cell18: 617–626.
Zeng, X., A. Hourset, and A. Tzagoloff, 2007c The Saccharomycescerevisiae ATP22 gene codes for the mitochondrial ATPase sub-unit 6-specific translation factor. Genetics 175: 55–63.
Zeng, X., M. H. Barros, T. Shulman, and A. Tzagoloff, 2008 ATP25,a new nuclear gene of Saccharomyces cerevisiae required forexpression and assembly of the Atp9p subunit of mitochondrialATPase. Mol. Biol. Cell 19: 1366–1377.
Ziaja, K., G. Michaelis, and T. Lisowksy, 1993 Nuclear controlof the messenger RNA expression for mitochondrial ATPasesubunit 9 in a new yeast mutant. J. Mol. Biol. 229: 909–916.
Zick, M., R. Rabl, and A. S. Reichert, 2009 Cristae formation-linkingultrastructure and function of mitochondria. Biochim. Biophys.Acta 1793: 5–19.
Zotova, L., M. Aleschko, G. Sponder, R. Baumgartner, S. Reipertet al., 2010 Novel components of an active mitochondrialK(+)/H(+) exchange. J. Biol. Chem. 285: 14399–14414.
Communicating editor: A. Hinnebusch
1234 T. D. Fox

















![Cronicon OPEN ACCESS EC PULMONOLOGY AND RESPIRATORY ... · Mia40, which interacts with a mitochondrial protein Mrp10 that renders import intermediates accessible to Mia40 [9]. Components](https://img.pdfslide.us/doc/110x75/5e2f7670f5e5772d22391027/cronicon-open-access-ec-pulmonology-and-respiratory-mia40-which-interacts-with.jpg)











