Embed Size (px)
Citation preview

From the SelectedWorks of David A Bainbridge
July, 2014
Mitigation site soil characteristics and the effectsof inoculation, nursery fertilizer practices andirrigation on survival and growth of mesquite(Prosopis glandulosa) in the Coachella Valley,CaliforniaDavid A BainbridgeMarcelle M. Darby, University of San Diego
Available at: https://works.bepress.com/david_a_bainbridge/42/

Restoration Notes. Volume 2:Number 5 July 2014
Mitigation site soil characteristics and the effects of inoculation, nursery fertilizer practices and irrigation on survival and growth of mesquite (Prosopis glandulosa) in the Coachella Valley, California
Marcelle M. Darby1
David A. Bainbridge2
1Assistant Director, Center for Educational Excellence, University of San Diego, CA 92110 USA2Restoration consultant, San Diego, USA
Corresponding author David A. BainbridgePhone (858) 693-1451 e-mail [email protected]
Abstract Prosopis glandulosa var. torreyana (mesquite, honey mesquite) is a small to medium sized leguminous tree that was once common in the low desert of California. Widespread destruction of mesquite woodlands in the Coachella Valley of southern California for fuel wood, agricultural and urban development, and urbanization has reduced once vast stands to isolated remnants. The rerouting and widening of Highway 86 in the Coachella Valley was a typical example of ongoing mesquite woodland removal. The California Department of Transportation supported this research to mitigate habitat loss. The abandoned agricultural land used for the test was provided by agreement with the Federal Wildlife Service and a large scale mitigation effort was anticipated. Despite the recognized need for mitigation, relatively little information on mesquite establishment in California is available. This research explored mesquite establishment by replanting on an abandoned farm field where the effects of inoculation and nutrient levels during greenhouse plant preparation for outplanting were evaluated for impacts on field survival and growth.
Introduction Prosopis glandulosa var. torreyana (L. D. Benson, M. C. Johnston) is a small to medium sized tree in the family Fabaceae (Leguminosae) (Burkart and Simpson, 1977). Mesquite was once found in extensive woodlands in the low deserts of southern California. Its distribution and occurrence have been greatly restricted by harvesting for fuel wood, agricultural and urban development, declining water tables from overdraft, salt buildup, intensive agriculture, off-road vehicle activity, tamarisk invasion (Tamarix ramosissima), and urban development (Bainbridge, 2007; Lovich and Bainbridge, 1999). Only isolated stands now remain. This is unfortunate because mesquite trees increase soil fertility and may play an important role in long-term productivity of desert plant communities (Virginia, 1990; Virginia, 1986). Indigenous people utilized this trait by transferring mesquite soils to gardens to improve fertility (Nabhan, 1982). Mesquite provides valuable food and protected habitat for many desert species (Dahl, 1982; Bainbridge and Tiszler, 2014). Mesquite was once a critical food resource for indigenous
1

populations (Bean and Saubel, 1972; Bell and Castetter, 1937), and they planted, transplanted, and managed this species (Shipek, 1989). Mesquite is a valuable multipurpose tree crop for dry land agroforestry, although it can be invasive in non-native settings (Bainbridge et al., 1990; Meyer, 1984; Zolfgahari and Harden, 1982). In the western Sonoran Desert (sometimes called the Colorado Desert), mesquite is a phreatophyte found in washes, along the edges of playas, and in other areas where ground water reserves are within 3-7 m of the surface, figure 1. Mesquite usually has a fibrous root system near the surface, exploiting moisture from infrequent rains, and a fast growing tap root that can reach great depths in its search for water (Bainbridge et al., 1990).
Mesquite forms symbiotic associations with both nitrogen fixing rhizobial bacteria and mycorrhizal fungi (Benthlenfalvay et al., 1984; Virginia, 1990). A mesquite stand near Harper's Well was found to be fixing approximately 60% of its required nitrogen (Shearer et al., 1983). Rhizobia strains collected in association with mesquite, include both fast-growing Rhizobium and slow-growing Bradyrhizobium (Jenkins et al., 1987; Waldon et al., 1989). The slow-growing rhizobia dominate the lowest depths (Jenkins et al., 1989). Nodules are commonly found in relatively moist soil in the phreatic zone, and have been found at depths of up to 8 m (Jenkins et al., 1988). Mesquite was the most effective nitrogen fixer in a comparative study in Riverside, CA (Abrams et al., 1990). Vesicular-arbuscular mycorrhizae (VAM) can enhance plant growth by improving uptake of phosphorus, water, and other nutrients (M. F. Allen, 1992). This can be very valuable for restoration project sites (E. B. Allen, 1989; M. F. Allen, 1988, 1984; Reeves et al., 1979). In soils with low water potential mycorrhizal plants show decreased resistance to water flux, i.e. mycorrhizal plants can get more water out of the soil than non-mycorrhizal plants (Allen and
2
Figure 1. Mesquite with pods

Allen, 1986). VAM fungi benefit from higher levels of available nitrogen (M. F. Allen, 1991; Azcón-Aguilar and Barea, 1992), while their formation and persistence is often inhibited by higher soil concentrations of phosphorus (Menge, 1984; Louis and Lim, 1988). The sensitivity of the VAM symbiosis to phosphorus may be species specific (Allen, 1991) and should be determined for the particular fungal and plant associations being studied. Mycorrhizal fungi benefit from higher oxygen levels and well aerated soil mixes. Reestablishing these trees in disturbed and degraded environments may require careful attention to reintroducing or facilitating microsymbiont associations through preplant preparation, field inoculation and irrigation strategies. Rhizobial associations have both nutrient requirements and limitations. Root hair infection and nodule development are inhibited by higher nitrogen concentrations (Gibson and Jordan, 1983), while higher concentrations of phosphorus can increase nodulation and nitrogen fixation in phosphorus limited soils (Louis and Lim, 1988). These symbionts both influence plant growth, root development and response to environmental stress. Among many effects the VAM are particularly helpful for phosphorus and the rhizobial bacterial in nodules can fix nitrogen. Dual inoculation with VAM fungi and rhizobia of plants that benefit from both associations would be expected to increase survival and growth more than single inoculation. There appear to be beneficial and perhaps synergistic interaction between rhizobia and VA mycorrhizal fungi in some plant species (Barea et al., 1987; Carpenter and Allen, 1988; Manjunath et al., 1984). Jenkins et al. (1988) suggested that effective nodulation of seedlings of the desert legume, smoke tree (Psorothamnus spinosus), was important for rapid root elongation and establishment in desert wash habitats occupied by this species. Daniels Hetrick et al. (1988) found that mycorrhizal inoculation increased root length as well as the number and diameter of primary and branch roots in big bluestem grass (Andropogon gerardii). It is important to understand the soil fertility, constraints such as salt and limited infiltration and the soil ecology of a restoration site. This study was also expected to provide information that would help nursery managers better prepare plants for difficult sites and assist revegetation specialists and foresters develop field support systems for both restoration and agroforestry projects in arid lands. While high nutrient levels are commonly used in nurseries this may be inappropriate for producing plants for difficult sites. Knowledge of the proper level and balance of nutrients to promote growth with a favorable allocation to roots, while supporting both symbioses is critical to improve guidelines for plant preparation and recommendations for nursery management. Materials and Methods
Site description This site is -56 m below sea level about 2 km west of Mecca on the western edge of the Sonoran Desert (33°33’56”N, 116°06’01”). This part of the Sonoran desert is commonly referred to as the Colorado Desert for its proximity to the Colorado River. The site is north of the Salton Sea in the Coachella Valley of California. This site had been used for agriculture but abandoned
3

several years before the study. Weed development was extensive and dumped trash was common around the abandoned dirt air strip.1 The two soil types in the area are Indio loam and Indio clay (Kocher and Harper, 1928). These are deep alluvial soils with good agricultural potential but at risk for salt buildup. The site was used to grow truck crops (tomatoes, lettuce, corn, etc.) until the early 1980s when farming ceased as a result of increasing water prices and perhaps salinization. Relict furrow and ditch patterns suggested the fields had been flood irrigated and drained with subsurface tile drains. The site was invaded by salt seep shrubs and trees including Atriplex lentiformis (saltbush), Suaeda moquinnii (bush seepweed), Baccharis salicifolia (seep-willow), B. emoryi, and Tamarix sp. (tamarisk) after abandonment (Hickman, 1993). A dense thicket had developed over much of the site. Saturated soil was encountered at 3 to 5 m. The annual rainfall at Indio, the closest recording station, averages 75 mm (U.S. Weather Bureau, 1955). Tropical storms moving north from the Gulf of California may drop rain equal to the yearly average in a few minutes, causing extensive sheet and stream flow and flash floods. Potential evaporation measured near the site was 3,600 mm (Hughes, 1963), more than 40 times the annual precipitation, figure 2.
Field investigation The first step was a detailed soil analysis to determine soil fertility and symbiont availability. Infiltration tests were also run, one between plants and one on the edge of the dirt runway. In September 1990, two soil cores were taken for physical and chemical analyses. The first was collected under a saltbush (Atriplex lentiformis) canopy located at the northwest end of the site and the second between saltbush canopies. The soil was sampled using a 5 cm diameter AMS bucket hand auger in 30 cm increments to a depth of 60 cm, then in 60 cm increments until the water table was reached. In May 1991, soil was sampled for microsymbiont analyses (rhizobia and VAM fungi). Two samples were taken down to the water table, one under and one between saltbush canopies. The soil was sampled in the same depth increments as the September 1990 samples. All sampling tools were flame sterilized with alcohol between samples (depth increments) to minimize microbial contamination. Each sample was sealed in a plastic bag to prevent moisture loss and refrigerated (4°C) until analyzed. Soil samples from the two cores taken to the water table at the reference site were also analyzed for physical and chemical properties.
4
1 The county sheriff warned us it was still used by narco-trafficantes and to always call before we went to the site.
Figure 2. Precipitation and evaporation from a Class A

Sub-samples for gravimetric soil moisture were taken from each sample within three days of sampling. Samples were weighed, dried for 48 hours at 105˚C and reweighed. Another portion of the sample was air-dried and ground to pass through a 2 mm sieve for physical and chemical analyses. Soil saturation pastes (Rhoades, 1982) were prepared and allowed to sit overnight. The pH of the saturated paste was then measured with a glass electrode. Saturation percentage (SP )= mls H2O added / weight dry soil x 100, was determined as a proxy for soil texture. Saturation extracts were obtained by transferring the pastes to a 120 mm Büchner funnel fitted with #5 Whatman filter paper and extracted under a 0.084 Mpa vacuum. The extract was collected in plastic centrifuge tubes and 3 drops of sodium hexametaphosphate were added to each tube as a dispersant and to prevent precipitation of CaCO3. Electrical conductivity (EC), sodium, calcium, and magnesium were measured from the soil extracts. Electrical conductivity was determined using a conductance meter (Yellow Springs Instruments (YSI) model 32. Sodium (Na+), calcium (Ca+2), and magnesium (Mg+2) concentrations of the extract were determined on a Perkin Elmer ICP/6500 Inductively Coupled Plasma Emission Spectrometer (Rhoades, 1982). The sodium adsorption ratio, SAR = Na /((Ca + Mg)/ 2)1/2, was calculated for the saturation extract from the salt concentrations. Plant available nitrogen (NH4+ and NO3-) and phosphorus (PO4-3) were extracted using potassium chloride (2N KCl) (Keeney and Nelson, 1982) and sodium bicarbonate (NaHCO3) (Olsen and Sommers, 1982) methods, respectively, and concentrations were determined by colorimetry on a Technicon Autoanalyzer II (Technicon Industrial Systems, 1977). Soil organic nitrogen was liberated using Kjeldahl digestion block techniques (Bremner and Mulvaney, 1982) and analyzed colorimetrically (Technicon Autoanalyzer II). Mesquite seedlings were grown in a sterile medium inoculated with field soil from the site to estimate the rhizobial nodulation potential of the soil and the VAM fungi inoculum density (Virginia et al., 1986). Mesquite seeds were surface sterilized and scarified with a 5-minute soak in concentrated H2SO4 and rinsed several times with sterile water. The plants were grown for four months in 4 cm x 20.5 cm Conetainers™ (supercells) in a greenhouse located at San Diego State University. Each supercell contained 150 ml of a steam sterilized sand and vermiculite mix (1:1), inoculated with eight grams (calculated dry weight) of soil collected from the site. The supercells were placed in three racks, with each rack representing a block containing three controls (uninoculated growth medium) and one replicate of each soil sample mixed with growth medium. The supercells were randomized in each rack resulting in a randomized complete block design. The plants were watered with sterilized2 water for the first two months then with a low nutrient concentration solution at every other watering. When plants were harvested, the roots and shoots were separated 2.5 cm below the cotyledon scar; the shoot lengths were measured from this point to the tip of the apical meristem. Plant samples were dried at 60°C for three days and weighed. Before the roots were dried, nodules were picked with forceps and counted. Nodules were dried and weighed. Fine roots from each dried sample were stained (Phillips and Hayman, 1970), examined for the presence of VA mycorrhizae, and assessed for percent VA mycorrhizal infection (Allen and Allen, 1980).
5
2 We had discovered that the tap water at the nursery greenhouse included rhizobial bacteria and other organisms.

Vesicular-arbuscular mycorrhizal fungi spores were extracted from 10 grams of each soil sample using the differential water/sucrose centrifugation method (Allen et al., 1979; Ianson and Allen, 1986). The spore count was used as an additional measure of inoculum density.
Greenhouse plant preparation Eight combinations of microsymbiont inoculum (none, dual, rhizobia, VAM fungi) and nutrient level (low, medium, high) were compared. Nutrient levels in the three nutrient solutions were low compared to conventional nursery practice. All started with one-fifth strength Hoagland's nutrient solution base with differing nitrogen and phosphorus concentrations. The high nutrient treatment had nitrogen (N) and phosphorus (P) concentrations at 20% of standard Hoagland's (N = 1.5 mM, P = 0.1 mM); the medium nutrient treatment had one tenth the N and P of the high nutrient solution (N = 0.15 mM, P = 0.01 mM); and the low nutrient treatment had one thirtieth the N and P concentrations (N = 0.05 mM, P = 0.003 mM) in an attempt to bracket the most appropriate nutrient level for growth, vigor and symbiont development. Mesquite seedlings from locally collected seeds were grown in a sterile medium inoculated with field soil to estimate the rhizobial nodulation potential of the soil (Virginia et al., 1986) and the VAM inoculum density. VAM spores were collected from soil under a remnant mesquite canopy. Spores were extracted from the soil using the water/sucrose differential centrifugation method (Ianson and Allen, 1986). Three genera were identified: Glomus, Aculospora, and Scutilospora. Glomus spores were the most abundant. Two different methods were used to produce the VAM inoculum, a soil-based and an aeroponic system, figure 3, with Panicum miliaceum (proso millet). Seeds were surface sterilized in a 0.5% sodium hypochlorite solution for two minutes, then rinsed with sterile water. Fluorescence was used to insure infection (Ames et al., 1982). Plants were grown in pots for two months and then soil and roots from these cultures were mixed and chopped together for use as a whole soil inoculum. The other plants were grown aeroponically (Hung and Sylvia, 1988). Samples were inspected for mycorrhizal infection using fluorescence microscopy.
6
Figure 3. Aeroponic system for inoculum production

Strains of fast and slow growing rhizobia, Rhizobium (strain 17b) and Bradyrhizobium (strain 10h) respectively, previously isolated from a mesquite woodland near Harper's Well (Waldon et al., 1989) were grown up in solution for the rhizobial inoculum. Viable rhizobia in solution were estimated with a dilution plate technique (Vincent, 1970). Mesquite seedlings were inoculated twice to ensure ample opportunity for nodulation by one or both strains. Approximately 9x108 cells of Rhizobium (in a 250 ml suspension) from the fast-growing strain were mixed directly in with the growth medium (just prior to planting). A second 250 ml suspension, with approximately 1x107 cells of Bradyrhizobium from the slow-growing strain, was added to the top of the growth medium 61 days after planting. Seedlings were grown in plant containers made from 1 m long 7.5 cm diameter PVC plastic drain pipe to encourage root proliferation and deep rooting (Bainbridge, 2007). The growth medium was well mixed flame sterilized medium grain size sand and vermiculite. Seeds were surface sterilized in concentrated H2SO4, rinsed in sterile water, and placed in 95% ethanol for 1.5 minutes before being scarified with a sterilized razor to ensure germination. Plants received 750 ml of the appropriate nutrient solution when the seeds were planted. The rhizobial inoculum was added in solution, plants without rhizobial inoculation received an equivalent amount of reverse osmosis water. Mycorrhizal inoculum was added to the containers in three bands, two of soil/root and one of roots only from the aeroponic system. A killed soil inoculum control was not used as a control because of limited supply. The volume of soil and, therefore, nutrient content in this inoculum was minimal compared to the nutrients received from the nutrient solutions.
Field Planting Ten matched plants from each of the eight treatments were installed on April 16, 1992 (152 days post planting in container) in a completely randomized design at 3 x 3 meter spacing. Ten centimeter diameter, 100 cm deep holes were drilled with a power auger and a liter of water was added to each hole. The PVC containers were split with a power saw. Roots were vigorous and taproots were several times the shoot length, figure 4. The roots were carefully removed from the medium and then the seedlings were held by the stem while the holes were gently backfilled and compacted. Another liter of water was added after soil was firmed up. Translucent twinwall Tubex™ tree shelters (approx. 7.5 cm x 20 cm) were staked down over each plant to provide protection from herbivory (Bainbridge, 1991; McAuliffe, 1986). Two liters of water were poured into the tree shelters. Netting was added to the top of the shelters to avoid trapping small animals, reptiles or birds. A
7
Figure 4. Planting mesquite seedlings

0.5 m square piece of black weed control fabric was placed around each plant to help control weeds, figure 5. The plants (almost 3 hours drive from the office) were watered with two liters of water periodically but not frequently enough to avoid significant water stress. Plant heights, from the cotyledon scar to the tip of the apical meristem, and plant survival were measured at each watering.
ResultsSoil Properties Soil infiltration rates were relatively low at 2 cm/hr at zero head and 3.4 cm hour at 5 cm head. Soils were alkaline, with mitigation site values from 7.6 to 8.8. At the mitigation site, pH values were comparable throughout the soil profile, Table 1. Only the most salt tolerant crops can grow at ECs of 4 dS m-1, and most prefer EC <2 (SICCFA, 1998).
Table 1. Soil Properties to the Water Table. September 1990. EC dSm-1 pH Sodium Ads. RatioDepth (cm) under between under between under between0-30 4.2 0.7 7.6 8.2 5 430-60 1.4 1.5 8 8 3 260-120 1 0.9 8.1 8.3 2 2120-180 0.6 0.5 8.3 8.4 2 2180-240 0.8 0.8 8.3 8.4 2 4240-300 0.9 0.6 8.3 8.3 2 4300-360 1.5 0.6 8.3 8.5 5 5360-420 1.6 3.4 8.5 8.1 10 12420-480 1.8 1.6 8.6 8.3 29 12
Sodium adsorption ratio (SAR) is an indicator of soil sodicity and the relative contribution of sodium to the soil cation exchange complex (Brady, 1990). The adverse effects on plant growth increase as soil SAR increases because of sodium toxicity, sodium induced changes in soil
8
Figure 5. Planting out with detail of Tubex treeshelter and weed mat installed
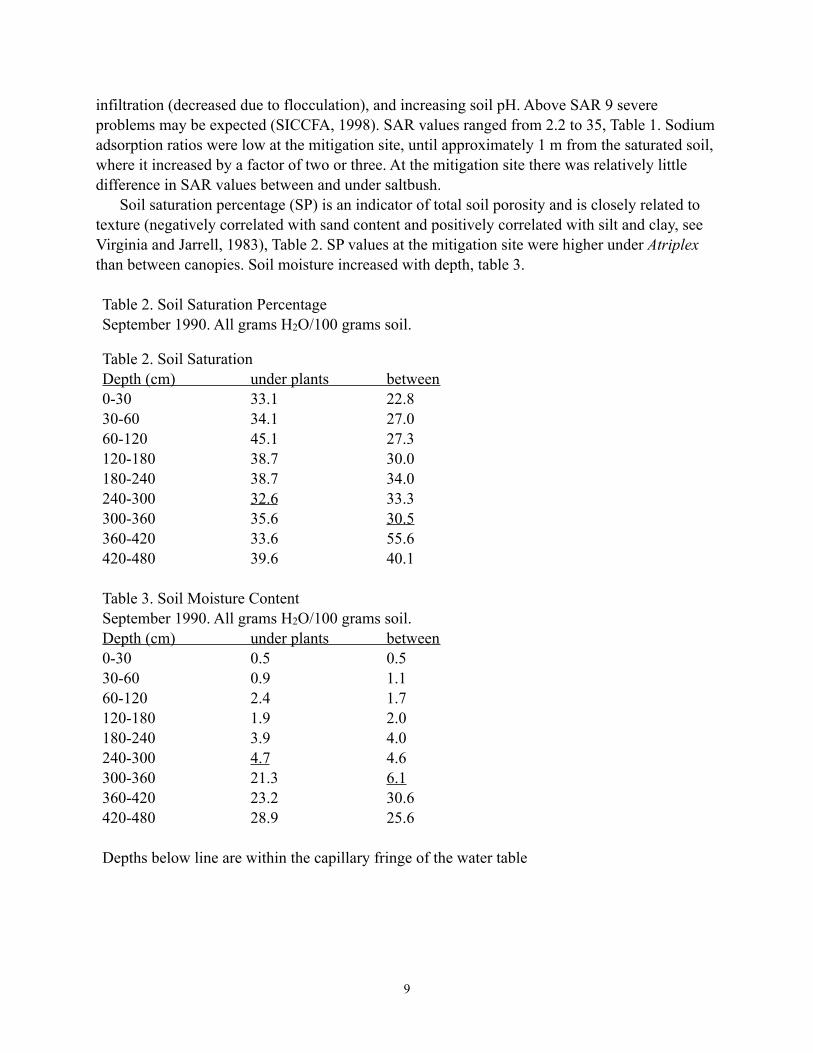
infiltration (decreased due to flocculation), and increasing soil pH. Above SAR 9 severe problems may be expected (SICCFA, 1998). SAR values ranged from 2.2 to 35, Table 1. Sodium adsorption ratios were low at the mitigation site, until approximately 1 m from the saturated soil, where it increased by a factor of two or three. At the mitigation site there was relatively little difference in SAR values between and under saltbush. Soil saturation percentage (SP) is an indicator of total soil porosity and is closely related to texture (negatively correlated with sand content and positively correlated with silt and clay, see Virginia and Jarrell, 1983), Table 2. SP values at the mitigation site were higher under Atriplex than between canopies. Soil moisture increased with depth, table 3.
Table 2. Soil Saturation PercentageSeptember 1990. All grams H2O/100 grams soil.
Table 2. Soil Saturation Depth (cm) under plants between 0-30 33.1 22.8 30-60 34.1 27.0 60-120 45.1 27.3 120-180 38.7 30.0 180-240 38.7 34.0 240-300 32.6 33.3 300-360 35.6 30.5 360-420 33.6 55.6 420-480 39.6 40.1 Table 3. Soil Moisture ContentSeptember 1990. All grams H2O/100 grams soil. Depth (cm) under plants between 0-30 0.5 0.5 30-60 0.9 1.1 60-120 2.4 1.7 120-180 1.9 2.0 180-240 3.9 4.0 240-300 4.7 4.6 300-360 21.3 6.1 360-420 23.2 30.6 420-480 28.9 25.6 Depths below line are within the capillary fringe of the water table
9

Soil fertility is shown in table 4. Organic nitrogen (TKN) and readily available forms of nitrogen (NO3--N, NH4+-N) and phosphorus (PO4-3-P) were measured to assess soil fertility. Soil nutrient concentrations generally decreased with increasing soil depth. Organic nitrogen in the soil ranged from 0.01 to 0.24 g kg-1.
Table 4. Soil fertility September 1990 TKN g/kg soil NO3---N mg/kg soilDepth (cm) under between under between0-30 0.24 0.17 44.89 4.5830-60 0.14 0.13 21.08 7.3160-120 0.17 0.10 15.5 2.96120-180 0.10 0.07 2.89 0.45180-240 0.08 0.03 2.10 0.28240-300 0.03 0.06 1.25 0.25300-360 0.04 0.04 1.47 0.13360-420 0.03 0.12 2.38 1.25420-480 0.09 0.10 1.16 3.68Means 0.10 0.09 10.30 2.32 NH4+-N mg/kg soil PO4-3-P mg/kg soilDepth (cm) under between under between0-30 3.34 3.08 4.01 2.5030-60 0.55 1.69 3.04 2.1160-120 0.63 1.13 2.39 1.45120-180 0.25 0.72 0.78 0.40180-240 0.25 <0.25 0.55 0.25240-300 <0.25 <0.25 0.25 0.25300-360 <0.25 <0.25 0.08 0.10360-420 <0.25 1.65 0.20 0.18420-480 1.38 0.75 0.17 0Means 1.07 1.50 1.27 0.80
Nitrate (NO3-) concentrations generally decreased with depth but rose again near the water table. Higher levels of NO3- were found under plants compared to unvegetated areas between plants. The NO3- concentrations ranged from 0.1 to 44 mg kg-1 at the mitigation site. Nitrate concentrations were much greater than ammonium (NH4+). This is typical of desert soils reflecting high N mineralization rates. NH4+ concentrations ranged from <0.25 to 3.3 mg kg-1, and tended to decrease with depth until about 0.5 m from the saturated soil where they increased sharply. The plant available phosphorus (PO4-3) concentrations varied widely ranging from undetectable to 4.01 mg kg-1. PO4-3 concentrations below 10 mg kg-1 are often considered inadequate for crop production. Phosphorus levels were generally higher under plants/ Both sites
10

showed decreasing concentrations with depth; the surface soil was usually over 90% higher in PO4-3 than the deep soil. The growth of seedlings inoculated with field soil reflects the combined effects of soil fertility and soil inoculation potential, and their interactive effects on plant nutrient acquisition. Total plant dry weights with soil inoculant under canopies ranged from 0.19 to 0.23 g. The plant dry weights from soil taken between canopies varied more but were similar, 0.14 and 0.23 g. Nodulation potential (the number of nodules induced on mesquite seedlings by inoculating with field soil) has been correlated with the more traditional most probable number (MPN) estimates of rhizobial population densities (Virginia et al., 1986). Each nodule usually corresponds to one rhizobial infection point. Nodulation potentials ranged from <10 to 60 nodules per plant at the mitigation site figure 6. Nodulation potential under canopies was comparable between the reference and mitigation sites. There was more variation in the nodulation potentials of soil collected between the shrub canopies at both sites.
Figure 6. Nodulation potential from samples taken a different depths.
Spores of VA mycorrhizal fungi ranged from 0 to 32 spores/10 g soil, figure 7. Spores were absent or much reduced below 60 cm, especially between saltbush canopies. Unlike the clear presence of nodules, there was no clear evidence of VA mycorrhizal infection (hyphae and vesicles present) in the stained roots of mesquite seedlings inoculated with soil from the site.
11

Plant growth and symbiont development The effects of inoculation and fertilization on growth were revealing and statistically significant, figure 8. As expected the high nutrient levels led to greater growth. At low and medium nutrient solution levels the dual inoculation provided a significant benefit. Dual inoculation led to the lowest root mass relative to plant growth. This is often the case as the symbionts improve nutrient acquisition at lower cost to the plant than growing roots. Root nodules were counted, weighed and measured. Figure 9 shows a typical root.The nodulation by depth over time was also instructive. As the roots grew deeper the symbionts did as well.
12
Figure 7. Mycorrhizal spore counts with depth

13
Figure 9. Nodules with medium fertility and dual inoculation
Figure 8. Total plant weight and root to plant ratio with differing treatments.

By the third harvest nodules were most extensive on the low fertility rhizobial inoculation and low dual, figure 10. This suggests the inoculation was occurring as expected. Nodules also developed on the low VAM treatment as rhizobial bacteria get around by wind or water even in the greenhouse.
Plant survival and growth in the field Plant survival and growth were limited by water stress. Deaths in all treatments occurred between July and September, the hottest and driest period. High-No, High-Dual, Medium-Dual, and Low-VAM treatments survived best. The Med-No and Low-No transplants suffered the greatest loss showing the value of inoculation. Transplants that were dual inoculated grew the tallest from November 1992 to May 1993. After one year the treatment ranks according to height were as follows: Low-Dual>High-Dual>Med-Dual>Low-Rhiz>Low-VAM>High-No. Med-No and Low-No treatments were not ranked because each had only one surviving transplant. The site was swept by a wildfire the following year and the remaining trees were killed.
14
Figure 10. Nodulation by treatment and depth

Discussion It proved practical to sample deep soil using only a hand auger at this site, figure 11. Soil sampling is often confined to the surface, but this misses essential details of soil properties and soil ecology. The changes in soil saturation percent were probably the result of lost fines to wind erosion during tillage and fallow periods on this exposed site. The increased saturation percent under saltbush canopies at the restoration site probably reflects the capture of wind blown clays and silts. Plants in these environments often improved the sites where they grow. Adequate and deep infiltration is especially important during plant establishment and may be essential for mesquite revegetation and deep ripping to 3 m at this site might improve survival and growth. Rapid root extension is considered to be important for successful establishment of mesquite and other deep-rooted desert trees and shrubs (Bainbridge, 2007; 1993). The ground water is well within the rooting zone of mesquite (Phillips, 1963; Freckman and Virginia, 1989); but the dry intermediate layer may slow or block mesquite establishment. A fluctuating water table may periodically wet these intermediate layers and redistribute salts preventing excessive accumulations near the water table (Jarrell and Virginia, 1990a). Or it may only occur with rare floods with standing water. The EC and SAR values decreased with depth until the capillary fringe of the water table was reached. This is different from the pattern found at long established woodland at Harper's Well, about 85 km due south of the site (Jarrell and Virginia, 1990a). At the Harper's Well site, cation concentrations increase until about two meters above the water table (active rooting zone, area of ion uptake by roots), then steadily decrease to the water table. This pattern reflects the occurrence of a stable water table and the presence of mesquite roots extracting water for a prolonged period. The increase of ion concentrations with depth at the mitigation site could be the result of irrigation leaching and drainage. Mature mesquite stands can tolerate very saline surface soils (Virginia and Jarrell, 1983). An EC of 28 dS m-1 is the estimated threshold for mesquite salt tolerance in greenhouse studies (Jarrell and Virginia 1990a), well above all the EC values at the site. However, increasing salinity decreases growth and water use efficiency of mesquite and may inhibit establishment. Excessive salinity and sodium toxicity may be causing the decline of the few remaining mesquite stands in the area, with much of the damage attributable to off-site agricultural practices including leaching salts and flood irrigation. These factors combine with removal of native vegetation to facilitate soil salinization. The increase in nitrate under saltbush canopies may, in part, be attributed to residual nitrate from agricultural fertilizer. Although it might be reasonable to hypothesize that saltbush has been
15
Figure 11. Soil sampling to depth

growing on the site long enough to accumulate residual nitrogen under its canopy through years of uptake and litter decomposition, an evaluation of mineralization, organic matter, and litter quality at the reference and mitigation sites would be required to test such an hypothesis. The low ratio of NH4+-N to NO3--N is typical for desert soils where mineralization and denitrification rates are high (Virginia et al., 1982). Ammonium is lost fairly readily in desert soils by ammonia volatilization and nitrification by microorganisms. Both processes are accelerated at high temperature and pH. Ammonia volatilization can occur at high rates with pH values greater than 8.3 (Freney et al., 1983), suggesting a high potential for NH3 losses at both sites, but especially for the reference site. Soil pH may be responsible for the pattern of higher NH4+ levels at the mitigation site. The concentration and depth distribution of inorganic P may play an important role in regulating VA mycorrhizal infection and function. Phosphorus levels in the surface layers of the mitigation site are adequate, compared to the Harper's Well system, but P concentrations at depth are low, the pattern also observed at Harper's Well. VA mycorrhizal infection of deep roots may be essential if mesquite is to have access to these limited phosphorus reserves at depth. The two to three fold higher soil phosphorus concentrations at the reference site, compared with the mitigation site, can be attributed to mesquite's ability to capture nutrients and improve soil fertility. Mesquite was able to grow in supercells inoculated with soil from the mitigation site. This suggests that there are no harmful diseases that might inhibit mesquite establishment. Plants, like mesquite, that develop symbioses with both rhizobia and mycorrhiza often require both for establishment (Carpenter and Allen, 1988; Herrera et al., 1993). Agricultural practices did not destroy the site rhizobial populations. This suggests that mesquite transplants would probably become infected in the deep soil, even if the rhizobia used to inoculate the transplant roots fail to proliferate and follow the root deep into the soil. Rhizobia are free-living in the soil and are not as dependent on a host for continued survival as VAM. Bacteria are also more easily transported by water flows. Rhizobial bacteria prefer moist but oxygenated (i.e. not saturated or anaerobic) soil. This is reflected in increasing nodulation potential with depth peaking at 2 to 3 m followed by a decline to the water table. These patterns are consistent with the distribution of mesquite-nodulating rhizobia most probable number (MPN) estimates found in a number of natural systems including dune and playa systems of the Sonoran Desert of California and the dune, playa, and arroyo systems of the Chihuahuan Desert of New Mexico (Virginia et al., 1986; Jenkins et al., 1988; Virginia et al., 1992). The VA mycorrhizal spore distribution in the soil suggests that agricultural practices at the mitigation site may have caused a decline in numbers, but did not inhibit their ability to persist or re-establish in the surface soils. Wind deposition is a common mode of dispersal of VA mycorrhizal spores in arid environments (Allen, 1988; Allen et al., 1989), and the high winds and widespread soil movement in this area could easily return fungi to the surface soils. The shrubs at the site (e.g., Atriplex sp.) were probably factors in the re-establishment (capture and maintenance) of spores and finer soil particles revealed in the higher SP values under the saltbush canopies. Vertical movement of mycorrhizal propagules through the soil is not well understood. Fungal hyphae can apparently extend deep into the soil, but only along an actively growing root (Allen, 1991). The question is whether or not roots will become infected at depths below 2.4 m at the
16

site when spores are absent. Mesquite pre-inoculated with VA mycorrhizal fungi and maintaining well-developed root systems (encouraged in deep containers) may enhance the recovery of a VA mycorrhizal fungi population at depth, when transplanted onto the site. Mesquite grown with the highest nutrient solution level (20% Hoagland's) appeared to be the best adapted. The low and very low were apparently too low. After outplanting, transplants from some of the low nutrient pre-treatments experienced shoot die-back, but the transplants from this highest level did not. Dual inoculated plants grown with moderate nutrient levels in the greenhouse responded more favorably than uninoculated plants. Low nutrient transplants did not do well even when inoculated. High-Dual inoculated plants managed the stress of symbioses well, and survived the low nutrient environment of the field well. Several limitations in the field study prevented a definitive assessment of transplanting success by treatment. Limited replication and considerable variation in plant response within treatments obscured differences between treatments. The critical long-term effects of the eight greenhouse pre-treatments on transplant establishment were not well determined because monitoring could only be done for one year after outplanting. Outplanting just prior to the hottest and driest season, coupled with infrequent watering and weeding, limited growth and survival. Although post-planting growing conditions were marginal, they reflect field conditions on many agroforestry and restoration sites. The belowground response of the transplants was not included in the field study, although it was recognized as an important aspect of plant establishment. It is not known if transplants treated with high nutrients were still restricting carbon allocation to roots, if symbiotic transplants were still infected, or if uninoculated transplants had become infected. Follow-up was limited by budget constraints and a wildfire that swept the site. Many questions remain to be answered in more detailed nursery and field studies.
Conclusions The main goal of this study was to explore the potential for successful mesquite establishment of an agriculturally disturbed and abandoned site, with an emphasis on the role of soil chemistry and biology in the full soil profile. This detailed evaluation of soil from the surface to greater than 3 m and research on symbionts and nutrient interactions in mesquite seedlings suggested that mitigation work would be relatively straightforward on this abandoned agricultural land. The physical, chemical, and biological conditions were suitable for mesquite growth. Successful establishment of microsymbionts is especially important as an important element of the belowground ecosystem (E. B. Allen 1989; M. F. Allen, 1988). Native mesquite stands form symbiotic associations with rhizobia and VA mycorrhizal fungi and pre-inoculation with symbionts may hasten the return of symbionts to the deep soil. Successful revegetation of the site would have been possible if the plants were inoculated, root shoot ratios were appropriate, and if they were deep irrigated and protected from herbivory (Bainbridge et al., 2007). Unfortunately a change in Fish and Wildlife Service personnel decided this mitigation site should be left to salt seep weeds and shrubs rather than converted to mesquite woodland. The cost of extensive testing, like that undertaken here, is not high. With simple hand augers and relatively simple tests full soil profiles were developed. Analyses did not require sophisticated or expensive tools or equipment, but did benefit from determined and capable
17

students and staff. The analysis was not expensive, representing perhaps 2-5% of implementation costs for a restoration project of this size. In this case it showed that there would be only minor challenges at the mitigation site related to soil chemistry and biology, but when this type of testing reveals serious problems or potential opportunities for improvement it can prevent very costly disasters. Ironically, interagency conflicts moved mitigation from this site to another site that was not evaluated before planting, with considerable plant losses apparently from high salinity and sodicity. Successful establishment of microsymbionts may be essential when the goal of the revegetation project is to reconstruct natural conditions (M. F. Allen, 1984). Native mesquite stands form symbiotic associations with rhizobia and VAM fungi; and pre-inoculating seedlings is a reasonable strategy for reintroducing symbionts to severely degraded sites. One or both of the root symbionts may be lacking in soils and reestablishment may be slow. While a field assessment of symbionts is not difficult, it is not routine, and dual inoculation of seedlings may be justified for severely disturbed sites. The objective should be to restore both ecosystem function and structure. Dual inoculation was best. Microsymbiont inoculation does not eliminate the need for moderate fertilization in the nursery. Plants given 20% Hoagland's N and P concentrations in the greenhouse were larger, had more favorable nutrient concentrations, and were better able to survive the rigors of outplanting than plants receiving low and very low N and P levels. High nutrient levels depress symbiont development and this can adversely affect long term survival. Managing root symbiotic associations is complex because of the interactions between soil moisture, soil biology, soil chemistry, container shape, container mix and nutrient levels (Bainbridge, 2007; M. F. Allen, 1992; Barea, 1986; Davidson, 1969). Pre-inoculating plants is more difficult than simple adding fertilizer because of the time and energy needed to collect, isolate, produce, and introduce site adapted inoculum. The management of microsymbionts in containers is still not well understood and can be challenging. Further investigation is needed to determine the optimum water regime, nutrient concentrations, growth medium, and container size and shape for optimal development of symbiotic associations. Mesquite survival would have improved for all pre-treatments if the transplants had received more water during the very hot, very dry months of the summer after outplanting. Although not used in this case deep pipe or wick irrigation would improve deep root development and would reduce the time required for mesquite tree roots to reach groundwater (Bainbridge, 2006; Bainbridge 2013). Even without deep irrigation plants that make it through the first season survive reasonable well and may survive until a flood event occurs. Successful mesquite establishment on disturbed sites is feasible. Mesquite is a desirable plant for reconstruction because it provides useful products and plays an important role in desert ecosystem functioning. Further research is still needed on deep root development and methods to speed root extension to the ground water (Bainbridge, 2013; Virginia and Jarrell, 1987).
18

ACKNOWLEDGMENTS This project was made possible by support and guidance from many people, figure 12. John Rieger and Pam Beare from the California Department of Transportation provided funding under CalTrans standard agreement 45912335. Permission to use the site was granted by the Fish and Wildlife Service. Ross Virginia provided oversight and guidance throughout the project. Robin McBride applied her attention to detail, experience and care to assist with, oversee and undertake lab analysis as well as assisting with fieldwork. Richard Darby, Betsy Bainbridge, Miguel Herrera, Debbie Waldecker, NaDene Sorenson, Matthew Fidelibus, Robert MacAller, Steve Netto, Julie Green, Mike and Edie Allen, Barbara Weinbaum and Fred Edwards also provided assistance. Students from William Carey International University helped with deep cores one day.
19
Figure 12. Complex and detailed analysis takes teamwork

REFERENCES
Abrams, M. M., W. M. Jarrell, H. A. Smith and P. R. Clark. 1990. Nitrogen accretion in soil and biomass by three Prosopis species. Agroforestry Systems 10:93-97.
Allen, E. B. 1989. The restoration of disturbed arid landscapes with special reference to mycorrhizal fungi. Journal of Arid Environments 17:279-286.
Allen, E. B. and M. F. Allen. 1986. Water relations of xeric grasses in the field: interactions of mycorrhizas and competition. New Phytologist 104:559-571.
Allen, E. B. and M. F. Allen. 1980. Natural re-establishment of vesicular-arbuscular mychorrizae following strip-mine reclamation in Wyoming. Journal of Applied Ecology 17:139-147.
Allen, E. B. and M. F. Allen. 1986. Water relations of xeric grasses in the field: interactions of mycorrhizas and competition. New Phytologist 104:559-571.
Allen, M.F. 1992. Mycorrhizal Functioning. Chapman Hall, New York, NY. Allen, M.F. 1991. The Ecology of Mycorrhizae. Cambridge University Press, New York, NY.Allen, M. F. 1989. Mycorrhizae and rehabilitation of disturbed arid soils: processes and
practices. Arid Soil Research 3:229-241.Allen, M.F. 1992. Mycorrhizal Functioning. Chapman Hall. NY NY Allen, M. F. 1988. Below ground structure: a key to reconstructing a productive arid ecosystem.
Pages 113-135 in E.B. Allen, ed. The Reconstruction of Disturbed Arid Lands: an Ecological Approach. Westview Press, Boulder, CO.
Allen, M. F. 1984. Physiology of mycorrhizae: a key to understanding successful plant establishment. Pages 69-80 in S. E. Williams and M. F. Allen, eds. VA Mycorrhizae and Reclamation of Arid and Semi-Arid Lands. University of Wyoming Experiment Station, Laramie, WY.
Allen, M.F., L.E. Hipps and G.L. Woolridge. 1989. Wind dispersal and subsequent establishment of VA mycorrhizal fungi across a successional arid landscape. Landscape Ecology 2:165-171.
Allen, M.F., T.S. Moore, Jr. and M. Christensen. 1979. Growth of vesicular-arbuscular- mycorrhizal and nonmycorrhizal Bouteloua gracilis in a defined medium. Mycologia 71:666-669.
Ames R. N., E. R. Ingham and C. P. P. Reid. 1982. Ultraviolet-induced autofluorescence of arbuscular mycorrhizal root infections: an alternative to clearing and staining methods for assessing infections. Canadian Journal of Microbiology 28:351-355.
Azcon-Aguilar, C. and J. M. Barea. 1992. Interactions between mycorrhizal fungi and other rhizosphere microorganisms. In Mycorrhizal Functioning. An Integrative Plant-Fungal Process. Ed. M F Allen. pp 163-198. New York, NY: Chapman & Hall.
Bainbridge, D. A. 2013. Wick irrigation. Overstory: The Agroforestry Journal #249.Bainbridge, D. A. 2007. A Guide for Desert and Dryland Restoration. Washington, DC: Island
Press. Bainbridge, D. A. 2006. Deep pipe irrigation. Overstory: The Agroforestry Journal #175.Bainbridge, D. A. 1993. Soil compaction. Land and Water 37(1):42-43.Bainbridge, D. A. 1991. Successful tree establishment on difficult dry sites. Proceedings of the
Third International Windbreaks and Agroforestry Symposium. Ridgetown, Canada. pp. 78-81.
20

Bainbridge, D. A. and J. Tiszler. 2014. Recreating mesquite mounds in the Colorado Desert. http://works.bepress.com/david_a_bainbridge/36/
Bainbridge, D. A. and R. A. Virginia. 1990. Restoration in the Sonoran desert of California. Restoration and Management Notes 8(1):3-14.
Bainbridge, D. A., R. A. Virginia and W. M. Jarrell. 1990. Honey Mesquite (Prosopis glandulosa). Species Note, NFTA 90-07. Nitrogen Fixing Tree Association, Hawaii 2p.
Bainbridge, D. A., R. A. Virginia and W. M. Jarrell. 1990. Honey mesquite: Multipurpose tree for arid lands. NFT Highlights 90-07. Nitrogen Fixing Tree Association, Waimanalo, HI 2 pp.
Barea, J. M. 1986. Importance of hormones and root exudates in mycorrhizal phenomena. In Physiological and Genetical Aspects of Mycorrhizae. Eds. V. Gianinazzi-Pearson and S. Gianinazzi. pp 177-187. INRA, Paris.
Barea, J. M., C. Azcon-Aguilar and R. Azcon. 1987. Vesicular arbuscular mycorrhiza improve both symbiotic N2 Fixation and N uptake from soil as assessed with a 15N technique under field conditions. New Phytologist 106, 717-725.
Bean, L. J. and K. S. Saubel. 1972. Temalpakh: Cahuilla Indian Knowledge and Usage of Plants. Banning, CA: Malki Museum Press.
Bell, W.H. and E.F. Castetter. 1937. Utilization of mesquite and screwbean by the aborigines in the American Southwest. University of New Mexico Bulletin 314, Biological Series 5(2):1-55.
Benthlenfalvay, G. J., S. Dakessian and R. S. Pacovsky. 1984. Mycorrhizae in a southern California desert: ecological implications. Canadian Journal of Botany 62:519-523.
Brady, N. C. 1990. The Nature and Properties of Soils. Macmillan Publishing Company, Bremner, J. M. and C. S. Mulvaney. 1982. Nitrogen-total. Pages 595-624 in A. L. Page, R. H.
Miller and D. R. Keeney, eds. Methods of Soil Analysis Part 2- Chemical and Microbiological Properties, 2nd edition. American Society of Agronomy, Inc. and Soil Science Society of America, Inc., Madison, WI.
Burkart, A. and B. B. Simpson. 1977. Appendix: The genus Prosopis and annotated key to the species of the world. In Mesquite, It's Biology in Two Desert Scrub Ecosystems. Ed. B. B. Simpson. pp 201-215. Stroudsburg, PA: Dowden, Hutchinson and Ross, Inc.
Carpenter, A. T. and M. F. Allen. 1988. Responses of Hedysarum boreale Nutt. to mycorrhizas and Rhizobium: plant and soil nutrient changes in a disturbed shrub-steppe. New Phytologist. 109:125-132.
Dahl, B. E. 1982. Mesquite as a rangeland plant. In Mesquite Utilization. Ed. H. W. Parker. pp A1-A20. College of Agricultural Sciences, Texas Tech University, Lubbock, TX.
Daniels Hetrick, B. A., J. F. Leslie, W. G. Thompson and K. D. Gerschefske. 1988. Physical and topological assessment of effects of a vesicular-arbuscular mycorrhizal fungus on root architecture of big bluestem. New Phytologist 110:85-96.
Darby, M. M. 1973. Distribution of Rhizobia and Mycorrhizal Fungi in Two Desert Soils and their Effects on the Growth of Prosopis glandulosa var Torreyana at Varying Nutrient Levels. MS Thesis, San Diego State University. San Diego, CA 106 p.
Davidson, R. L. 1969 Effects of soil nutrients and moisture on root/shoot ratios in Lolium perenne L. and Trifolium repens L. Annals of Botany 33:571-577.
21

Freckman, D. W. and R. A. Virginia. 1989. Plant-feeding nematodes in deep-rooting desert ecosystems. Ecology 70(6):1665-1678.
Freney, J. R., J. R. Simpson and O.T. Denmead. 1983. Volatilization of ammonia. Pages 1-32 in J. R. Freney and J. R. Simpson, eds. Gaseous Loss of Nitrogen from Plant-Soil Systems. The Hague: Martinus Nijhoff.
Gibson, A. H. and D. C. Jordan. 1983. Ecophysiology of nitrogen-fixing systems. In Physiological Plant Ecology III: Responses to the Chemical and Biological Environment. Eds. O. L. Lange, P. S. Nobel, C. B. Osmond and H. Ziegler. pp 301-390. New York, NY: Springer-Verlag.
Gupta, R. P. and V. Punj. 1990. Interaction between Rhizobium, mycorrhiza, nitrogen and phosphorus and their effect on growth and symbiotic behavior of Leucaena leucocephala. pp 165-166. In Jalali, B. L. and Chand, H. eds. Trends in mycorrhizal research. Proceedings of the National Conference on Mycorrhiza. Haryana Agricultural University, Hisar, India.
Herrera, M. A., C. P. Salamanca and J. M. Barea. 1993. Inoculation of woody legumes with selected arbuscular mycorrhizal fungi and rhizobia to recover desertified Mediterranean ecosystems. Applied Environmental Microbiology 59(1):129-133.
Hickman, J. C. ed. 1993. The Jepson Manual of Higher Plants of California. University of California Press, Los Angeles, CA.
Hughes, C. L. 1963. A study of the evaporation from Salton Sea, California. U.S. Geological Survey Open File Report, Yuma, Arizona.
Hung, L. I. and D. M. Sylvia. 1988 Production of vesicular-arbuscular mycorrhizal fungus inoculum in aeroponic culture. Applied and Environmental Microbiology 54(2):353-357.
Ianson, D. C. and M. F. Allen. 1986. The effects of soil texture on extraction of vesicular- arbuscular mycorrhizal fungal spores from arid sites. Mycologia 78(2):164-168.
Jarrell, W. M. and R. A. Virginia. 1990a. Response of mesquite to nitrate and salinity in a simulated phreatic environment: water use, dry matter and mineral nutrient accumulation. Plant and Soil 125:185-196.
Jarrell, W. M. and R. A. Virginia. 1990b. Soil cation accumulation in a mesquite woodland: sustained production and long-term estimates of water use and nitrogen fixation. Journal of Arid Environments. 18:51-58.
Jenkins, M. B., R. A. Virginia and W. M. Jarrell. 1989. Ecology of fast-growing and slow-growing mesquite-nodulating rhizobia in Chihuahuan and Sonoran Desert ecosystems. Soil Science Society of America Journal 53( ):543-549.
Jenkins, M.B., R.A. Virginia and W.M. Jarrell. 1988. Depth distribution and seasonal populations of mesquite-nodulating rhizobia in warm desert ecosystems. Soil Science Society of America Journal. 52(6):1644-1650.
Jenkins, M. B., R. A. Virginia and W. M. Jarrell. 1987. Rhizobial ecology of the woody legume mesquite (Prosopis glandulosa) in the Sonoran Desert. Applied Environmental Microbiology. 53(1):36-40.
Keeney, D. R. and D. W. Nelson. 1982. Nitrogen-inorganic forms. Pages 643-698 in A. L. Page, R. H. Miller and D. R. Keeney, eds. Methods of Soil Analysis Part 2- Chemical and Microbiological Properties. American Society of Agronomy, Inc. and Soil Science Society of America, Inc., Madison, WI.
22

Kocher, A. E. and W. G. Harper. 1928. Soil Survey: The Coachella Valley Area California. Soil Survey Report No. 16 Series 1923. U.S. Govt. Printing Office, Washington DC.
Louis, I. and G. Lim. 1988. Differential response in growth and mycorrhizal colonization of soybean to inoculation with two isolates of Glomus clarum in soils of different P availability. Plant and Soil 112:37-43.
Lovich, J. E. and D. A. Bainbridge. 1999. Anthropogenic degradation of the Southern California desert ecosystems and prospects for natural recovery and restoration. Environmental Management 24(3):309-326.
Manjunath, A., D. J. Bagyaraj and H. S. Gopalala Gowda. 1984 Dual inoculation with VA mycorrhizae and Rhizobium is beneficial to Leucaena. Plant and Soil 78:445-448.
McAuliffe, J. R. 1986. Herbivore-limited establishment of a Sonoran Desert tree, Cercidium microphyllum. Ecology 67(1):276-280.
Menge, J. A. 1984. Inoculum production. In VA Mycorrhiza. Eds. C. L. Powell and D. J. Bagyaraj. pp 187-203. Boca Raton, FL: CRC Press.
Meyer, D. 1984. Processing, utilization, and economics of mesquite pods. PhD Thesis Swiss Federal Institute of Technology, Zurich, Switzerland.
Nabhan, G. P. 1982. The Desert Smells Like Rain: A Naturalist in Papago Indian Country. San Francisco, CA: North Point Press.
Olsen, S. R. and L. E. Sommers. 1982. Phosphorus. Pages 403-430 in A. L. Page, R. H. Miller and D. R. Keeney, eds. Methods of Soil Analysis Part 2- Chemical and Microbiological Properties. American Society of Agronomy, Inc. and Soil Science Society of America, Inc., Madison, WI.
Phillips, J. M. and D. S. Hayman. 1970. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Transactions British Mycology Society 55(1):158-161.
Phillips, W. S. 1963. Depth of roots in soil. Ecology 44:424.Reeves, B. F., D. Wagner, T. Moorman and J. Kiel. 1979. The role of endomycorrhizae in
revegetation practices in the semi-arid west I: a comparison of incidence of mycorrhizae in severely disturbed vs. natural environments. American Journal of Botany 66(1):6-13.
Rhoades, J. D. 1982. Soluble salts. Pages 167-179 in A. L. Page, R. H. Miller and D. R. Keeney, eds. Methods of Soil Analysis Part 2- Chemical and Microbiological Properties, 2nd edition. American Society of Agronomy, Inc. and Soil Science Society of America, Inc., Madison, WI.
Shearer, G., D. H. Kohl, R. A. Virginia, B. A. Bryan, J. L. Skeeters, E. T. Nilsen, M. R. Sharifi and P. W. Rundel. 1983. Estimates of N2-fixation from variation in the natural abundance of 15N in Sonoran Desert ecosystems. Oecologia 56:365-373.
Soil Improvement Committee, California Fertilizer Association (SICCFA). 1998. Western Fertilizer Handbook. Interstate Publishers. Danville, IL.
Shipek F. C. 1989. An example of intensive plant husbandry: the Kumeyaay of southern California. In Foraging and Farming. Eds. D R Harris and G C Hillman. pp 159-170. Unwin Hyman, London.
Technicon Industrial Systems. 1977. Individual/simultaneous determinations of nitrogen and/or phosphorus in BD acid digests. Industrial method number 329-74 w/b. Tarrytown, NY.
U.S. Weather Bureau.
23

Vincent, J. M. 1970. A Manual for the Practical Study of the Root Nodule Bacteria. London: Blackwell.
Virginia, R. A. 1990. Desert restoration: the role of woody legumes. pp 23-30. In: J.J. Berger, ed. Environmental Restoration. Island Press, Washington, DC.
Virginia, R. A. 1986. Soil development under legume tree canopies. Forest Ecology and Management 16:69-79.
Virginia, R.A. and D.A. Bainbridge. 1987. Revegetation in the Colorado desert: lessons from the study of natural systems. pp. 52-63. In: Rieger, J.P. and Williams, B.K. eds. Proceedings of the 2nd Native Plant Revegetation Symposium. Society for Ecological Restoration and Management, Madison, Wisconsin.
Virginia, R.A. and W.M. Jarrell. 1987. Approaches for studying the function of deep root systems. Pages 107-127 in J.D. Tenhunen et al. eds. Plant Response to Stress. Berlin: Springer-Verlag.
Virginia, R. A. and W. M. Jarrell. 1983. Soil properties in a Prosopis (mesquite) dominated Sonoran Desert ecosystem. Soil Science Society of America Journal 47:138-144.
Virginia, R. A., W. M. Jarrell, W. G. Whitford and D. W. Freckman. 1992. Soil biota and soil properties in the surface rooting zone of Mesquite (Prosopis glandulosa) in historical and recently desertified Chihuahuan Desert habitats. Biology Fertility Soils. 14:90-98.
Virginia, R. A., M. B. Jenkins and W. M. Jarrell. 1986. Depth of root symbiont occurrence in soil. Biology and Fertility of Soils. 2:127-130.
Virginia R. A., W. M. Jarrell and E. Franco-Vizcaino. 1982. Direct measurement of denitrification in a Prosopis (mesquite) dominated Sonoran Desert ecosystem. Oecologia 53:120-122.
Zolfgahari, R. and M. Harden. 1982. Nutritional value of mesquite beans, Prosopis glandulosa. pp K1-16. In Parker, H. W. ed. Mesquite Utilization, College of Agricultural Sciences, Texas Tech, Lubbock, TX.
Waldon, H. B., M. B. Jenkins, R. A. Virginia and E. E. Harding. 1989. Characteristics of woodland rhizobial populations from surface- and deep-soil environments of the Sonoran Desert. Applied and Environmental Microbiology 55(12):3058-3064.
24





























