Embed Size (px)
Citation preview

MiG
KM*NIJJ
R
copcwtgspap
g
lctTfta
DCAf
rctH
Biochemical and Biophysical Research Communications 262, 610–614 (1999)
Article ID bbrc.1999.1223, available online at http://www.idealibrary.com on
0CA
igration of Exogenous Immature Hematopoietic Cellsnto Adult Mouse Brain Parenchyma underFP-Expressing Bone Marrow Chimera
enji Ono,*,† Takemasa Takii,* Kikuo Onozaki,* Masahito Ikawa,‡asaru Okabe,‡ and Makoto Sawada†,§,1
Department of Hygienic Chemistry, Faculty of Pharmaceutical Sciences, Nagoya City University, Mizuho,agoya, Aichi 467-8603, Japan; †Joint Research Division for Therapies against Intractable Diseases,
nstitute for Comprehensive Medical Science, Fujita Health University, Toyoake, Aichi 470-1192,apan; ‡Research Institute for Microbial Diseases, Osaka University, Suita, Osaka 565-0871,apan; and §PRESTO, Japan Science and Technology Corporation, Japan
eceived July 14, 1999
mmprebpiwbsnmmpd
diflctfltmempfi
Gtte
Bone marrow transplantation with GFP-expressingells from GFP-transgenic mice resulted in migrationf GFP-positive cells into peripheral tissues and brainarenchyma. Most of these cells were observed asolony-like clusters. GFP-positive clusters in the brainere stained by antibody for ER-MP12, but those in
he peripheral tissues were not. Since ER-MP12 anti-en has been reported as a marker for murine early-tage myeloid precursor, this might suggest that somearts of phagocytic cells in the brain parenchyma suchs microglia are derived from undifferentiated pluri-otent hematopoietic cells. © 1999 Academic Press
Key Words: bone marrow transplantation; ER-MP12;reen fluorescent protein; microglia.
The brain contains many identifiable cell subpopu-ations in addition to neurons, which are collectivelyalled glial cells. Microglia, macrophage-like cells inhe brain parenchyma, are multifunctional cells (1).hey play important roles in the development, dif-
erentiation and maintenance of neural cells viaheir phagocytic activity and production of cytokinesnd trophic factors (2). Microglia are thought to be of
1 To whom correspondence should be addressed at Joint Researchivision for Therapies against Intractable Diseases, Institute foromprehensive Medical Science, Fujita Health University, Toyoake,ichi 470-1192, Japan. Fax: 181-562-93-2487. E-mail: msawada@
ujita-hu.ac.jp.Abbreviations used: BBB, blood brain barrier; GFP, green fluo-
escent protein; 5-FU, 5-fluorouracil; FACS, fluorescence activatedell sorter; RT, reverse transcription; PCR, polymerase chain reac-ion; EGFP, enhanced green fluorescent protein; b-actin, beta-actin;E, hematoxylin–eosin.
610006-291X/99 $30.00opyright © 1999 by Academic Pressll rights of reproduction in any form reserved.
esodermal origin and probably belong to theonocyte/macrophage cell lineage (3). The usual re-
lacement of peripheral phagocytic cells, tissue mac-ophages, is from blood-borne monocytes (4, 5). How-ver, such turnover appears unlikely to occur in therain because the blood brain barrier (BBB) wouldrevent such cell migration under normal physiolog-cal conditions (6). Furthermore, microglia appearithin the CNS before it is vascularized (7–10) andefore monocytes are produced in hemopoietic tis-ues (11, 12). Therefore, not all microglia can origi-ate from circulating monocytes during develop-ent. Another possibility is that some or allicroglia are derived from undifferentiated hemato-
oietic cells which colonize the developing CNS in-ependently of its vascularization (12).Recently, green fluorescent protein (GFP) has been
emonstrated to be a useful tool for identifying genet-cally modified hematopoietic cells (13). Because theuorescence activity of GFP requires no substrates,ofactors, or additional gene products, cells expressinghe intracellular GFP marker can be scored directly byuorescent microscopy and/or flow cytometry withouthe requirement for antibody staining or extensive cellanipulation. Therefore, GFP is thought to be a pow-
rful in vivo marker for identification and tracking ofurine multipotential homatopoietic cells and their
rogeny following transplantation of genetically modi-ed bone marrow.In this paper, we made bone marrow chimeras withFP-expressing bone marrow cells derived from GFP-
ransgenic mice as a donor and investigated distribu-ion and characterization of GFP-positive cells in vivo,specially in the brain parenchyma.
M
rm5G(wMswmivmwbgAca
eFfi
c7mf
stctw
frT9es(sUB
oFfCrpa1amdctTy
Biaatp
ppptcmpw
Vol. 262, No. 3, 1999 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ATERIAL AND METHODS
Bone marrow transplantation. Adult male B6 mice were used asecipients. At 96 h and 48h before bone marrow transplantation,ice (weighing 18 –38 g) were intraperitoneally injected with
-fluorouracil (5-FU) at a dose of 150 and 75 mg/kg. Adult maleFP-transgenic mice (TgN(b-act-EGFP)Osb) were used as donors
14). Upon harvest, bone marrow cells from GFP-transgenic miceere collected by flushing the bone shafts of femora with RPMIedium 1640 (GIBCO BRL) supplemented with 10% fetal calf
erum and 1% penicillin/streptomycin, using a 5 ml syringe fittedith a 21-G needle. Then the cells were washed once in the aboveedium by centrifugation at 1500 rpm for 5 min at 4°C and were
njected into recipient male B6 mice treated with 5-FU via the tailein at a concentration of 1 3 107 cells/mouse. At 7 days after bonearrow transplantation, mice were anesthetized and treatedith about 100 ml isotonic saline by cardiac perfusion, then therain, liver, spleen and lung were isolated, frozen in liquid nitro-en, and embedded in O.C.T. compound (Tissue Tek, Elkhart, IN).t the same time, the femora were also isolated and bone marrowells were collected as described above for the following FACSnalysis.
FACS analysis. GFP-positive cells in bone marrow and periph-ral blood cells were analyzed by a flow cytometry using aACSCalibur cell sorter (Becton–Dickinson) equipped with a 530-nmlter at a bandwidth of 615 nm.
Western blotting. The brain, liver, spleen, lung and bone marrowells were homogenized in 3-fold TNE buffer (10 mM Tris–HCl, pH.5, 1% NP-40, 0.15 M NaCl, 1 mM EDTA, 10 mg/ml approtinin, 10g/ml leupeptin, 1 mM ortho-vanadate), centrifuged at 13,000 rpm
or 30 min at 4°C, then supernatant fractions were collected. Their
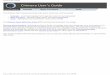
FIG. 1. Replacement of bone marrow cells by GFP-positive cells.one marrow cells derived from a control B6 mouse (A) and a recip-
ent B6 mouse at Day 7 after bone marrow transplantation (B) werenalyzed by flow cytometry. Results were summarized in C. Negativend positive indicates bone marrow cells derived from B6 and GFPransgenic mice, respectively. % indicates percentage of GFP-ositive cell detected.
611
upernatants were analyzed by 15% SDS–polyacrylamide gel elec-rophoresis and western blotting. To detect GFP, anti-GFP poly-lonal rabbit antibody (CLONTECH Laboratories, Inc., 1:1000 dilu-ion) and ECL detection system (Amersham Pharmacia Biotech)ere used.
RNA preparation and RT-PCR. Total RNA were extracted fromrozen sections using RNeasy RNA preparation kit (QIAGEN). RTeaction PCR were performed on RoboCycler (STRATAGENE).he PCR conditions were 94°C for 5 min, followed by 35 cycles of4°C for 1 min, 55°C for 2 min and 72°C for 2 min, with a finalxtension at 72°C for 4 min. The following primers were synthe-ized and used to detect the enhanced green fluorescent proteinEGFP) mRNA and mouse beta-actin (b-actin) mRNA: EGFPense and antisense, 794 – 811 and 1249 –1266 (Gene Bank,55763); b-actin sense and antisense, 25– 45 and 541–564 (Geneank, M12481).
Immunochemical staining. Sections (7 mm) were cut with a cry-stat microtome, transferred to a slide glass and immediately dried.ollowing fixation with 4% paraformaldehyde in PBS (pH 7.2) at 4°C
or 5 min, sections were labeled with monoclonal antibodies againstD34 (hematopoietic stem cell marker; PharMingen), Mac-1 (C3bieceptor marker; BMA) or ER-MP12 (murine early stage myeloidrecursor marker; BMA). In brief, sections were incubated for 30 mint room temperature in PBS containing 1% bovine serum albumin,0% normal goat serum and 0.01% sodium azide, then labeled withmonoclonal antibody against mouse Mac-1 at a dilution of 1:50, theouse ER-MP12 at a dilution of 1:100 or the mouse CD34 at a
ilution of 1:25. The reaction was visualized with R-phycoerythrin-onjugated goat F(ab9)2 anti-rat IgG (CALTAG) at a dilution of 1:100,hen photographed under a fluorescent microscope (BX-50, Olympus,okyo, Japan). One of each serial section were stained by Hematox-lin Eosin (HE).
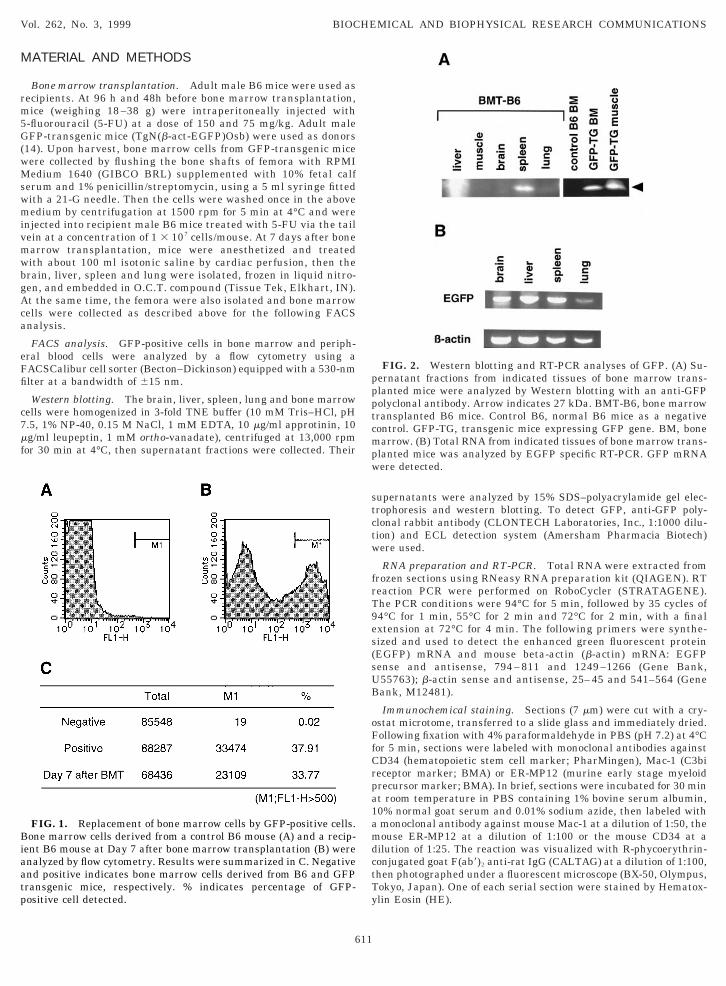
FIG. 2. Western blotting and RT-PCR analyses of GFP. (A) Su-ernatant fractions from indicated tissues of bone marrow trans-lanted mice were analyzed by Western blotting with an anti-GFPolyclonal antibody. Arrow indicates 27 kDa. BMT-B6, bone marrowransplanted B6 mice. Control B6, normal B6 mice as a negativeontrol. GFP-TG, transgenic mice expressing GFP gene. BM, bonearrow. (B) Total RNA from indicated tissues of bone marrow trans-
lanted mice was analyzed by EGFP specific RT-PCR. GFP mRNAere detected.
R
prmctmps
taa(tfPlo
oos3Tbo
oalbt
goCC(n4nngs
mtPmMwe
cc
(G
Vol. 262, No. 3, 1999 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
ESULTS AND DISCUSSIONS
At day 7 after bone marrow transplantation, GFP-ositive cells were detected in the bone marrow cells ofecipients (Fig. 1). The replacement seemed to be al-ost maximal because the percentage of GFP-positive
ells in the recipient’s bone marrow cells was similar tohat from the positive control of donor GFP-transgenicouse. This was confirmed by FACS analysis using
eripheral blood cells from recipient mice (data nothown).Tissue distribution of GFP-positive cells were inves-
igated biochemically. Western blotting revealed thepparent GFP-specific 27-kDa band in spleen and lungnd very faint but significant band in liver and brainFig. 2A). By RT-PCR analysis, GFP mRNA was de-ected in brain, lung, liver and spleen (Fig. 2B). There-ore, migration of GFP-positive cells was detected byCR. Since the number of GFP-positive colonies in
iver and brain was much less than other tissues, bandf GFP protein in these tissues was thought to be faint.Frozen tissue sections were also investigated by flu-
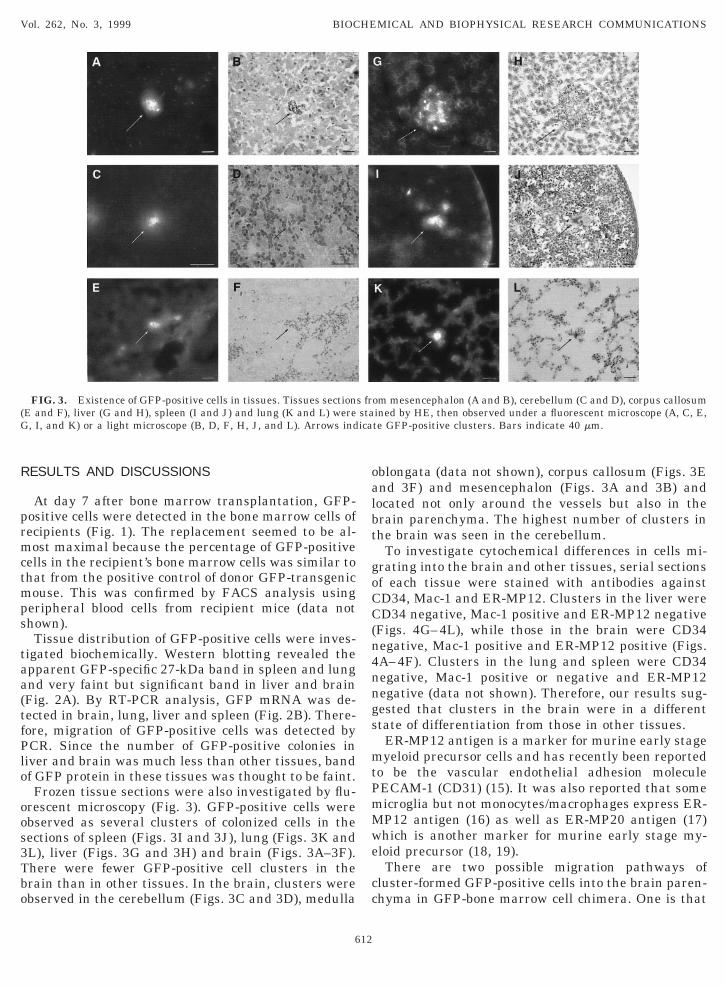
rescent microscopy (Fig. 3). GFP-positive cells werebserved as several clusters of colonized cells in theections of spleen (Figs. 3I and 3J), lung (Figs. 3K andL), liver (Figs. 3G and 3H) and brain (Figs. 3A–3F).here were fewer GFP-positive cell clusters in therain than in other tissues. In the brain, clusters werebserved in the cerebellum (Figs. 3C and 3D), medulla
FIG. 3. Existence of GFP-positive cells in tissues. Tissues sectionE and F), liver (G and H), spleen (I and J) and lung (K and L) were, I, and K) or a light microscope (B, D, F, H, J, and L). Arrows ind
612
blongata (data not shown), corpus callosum (Figs. 3End 3F) and mesencephalon (Figs. 3A and 3B) andocated not only around the vessels but also in therain parenchyma. The highest number of clusters inhe brain was seen in the cerebellum.
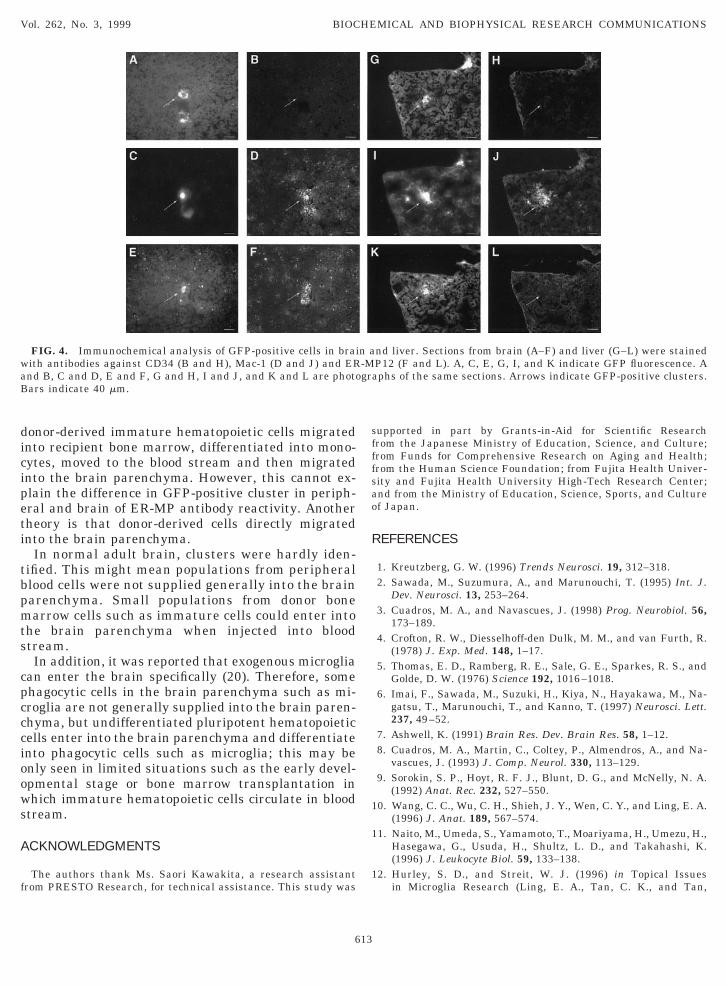
To investigate cytochemical differences in cells mi-rating into the brain and other tissues, serial sectionsf each tissue were stained with antibodies againstD34, Mac-1 and ER-MP12. Clusters in the liver wereD34 negative, Mac-1 positive and ER-MP12 negative
Figs. 4G–4L), while those in the brain were CD34egative, Mac-1 positive and ER-MP12 positive (Figs.A–4F). Clusters in the lung and spleen were CD34egative, Mac-1 positive or negative and ER-MP12egative (data not shown). Therefore, our results sug-ested that clusters in the brain were in a differenttate of differentiation from those in other tissues.ER-MP12 antigen is a marker for murine early stageyeloid precursor cells and has recently been reported
o be the vascular endothelial adhesion moleculeECAM-1 (CD31) (15). It was also reported that someicroglia but not monocytes/macrophages express ER-P12 antigen (16) as well as ER-MP20 antigen (17)hich is another marker for murine early stage my-loid precursor (18, 19).There are two possible migration pathways of
luster-formed GFP-positive cells into the brain paren-hyma in GFP-bone marrow cell chimera. One is that
om mesencephalon (A and B), cerebellum (C and D), corpus callosumined by HE, then observed under a fluorescent microscope (A, C, E,te GFP-positive clusters. Bars indicate 40 mm.
s frstaica
dicipeti
tbpmts
cpcccioows
A
f
sfffsao
R
1
1
1
waB
Vol. 262, No. 3, 1999 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
onor-derived immature hematopoietic cells migratednto recipient bone marrow, differentiated into mono-ytes, moved to the blood stream and then migratednto the brain parenchyma. However, this cannot ex-lain the difference in GFP-positive cluster in periph-ral and brain of ER-MP antibody reactivity. Anotherheory is that donor-derived cells directly migratednto the brain parenchyma.
In normal adult brain, clusters were hardly iden-ified. This might mean populations from peripherallood cells were not supplied generally into the brainarenchyma. Small populations from donor bonearrow cells such as immature cells could enter into
he brain parenchyma when injected into bloodtream.In addition, it was reported that exogenous microglia
an enter the brain specifically (20). Therefore, somehagocytic cells in the brain parenchyma such as mi-roglia are not generally supplied into the brain paren-hyma, but undifferentiated pluripotent hematopoieticells enter into the brain parenchyma and differentiatento phagocytic cells such as microglia; this may benly seen in limited situations such as the early devel-pmental stage or bone marrow transplantation inhich immature hematopoietic cells circulate in blood
tream.
CKNOWLEDGMENTS
The authors thank Ms. Saori Kawakita, a research assistantrom PRESTO Research, for technical assistance. This study was
FIG. 4. Immunochemical analysis of GFP-positive cells in braiith antibodies against CD34 (B and H), Mac-1 (D and J) and ERnd B, C and D, E and F, G and H, I and J, and K and L are photoars indicate 40 mm.
613
upported in part by Grants-in-Aid for Scientific Researchrom the Japanese Ministry of Education, Science, and Culture;rom Funds for Comprehensive Research on Aging and Health;rom the Human Science Foundation; from Fujita Health Univer-ity and Fujita Health University High-Tech Research Center;nd from the Ministry of Education, Science, Sports, and Culturef Japan.
EFERENCES
1. Kreutzberg, G. W. (1996) Trends Neurosci. 19, 312–318.2. Sawada, M., Suzumura, A., and Marunouchi, T. (1995) Int. J.
Dev. Neurosci. 13, 253–264.3. Cuadros, M. A., and Navascues, J. (1998) Prog. Neurobiol. 56,
173–189.4. Crofton, R. W., Diesselhoff-den Dulk, M. M., and van Furth, R.
(1978) J. Exp. Med. 148, 1–17.5. Thomas, E. D., Ramberg, R. E., Sale, G. E., Sparkes, R. S., and
Golde, D. W. (1976) Science 192, 1016–1018.6. Imai, F., Sawada, M., Suzuki, H., Kiya, N., Hayakawa, M., Na-
gatsu, T., Marunouchi, T., and Kanno, T. (1997) Neurosci. Lett.237, 49–52.
7. Ashwell, K. (1991) Brain Res. Dev. Brain Res. 58, 1–12.8. Cuadros, M. A., Martin, C., Coltey, P., Almendros, A., and Na-
vascues, J. (1993) J. Comp. Neurol. 330, 113–129.9. Sorokin, S. P., Hoyt, R. F. J., Blunt, D. G., and McNelly, N. A.
(1992) Anat. Rec. 232, 527–550.0. Wang, C. C., Wu, C. H., Shieh, J. Y., Wen, C. Y., and Ling, E. A.
(1996) J. Anat. 189, 567–574.1. Naito, M., Umeda, S., Yamamoto, T., Moariyama, H., Umezu, H.,
Hasegawa, G., Usuda, H., Shultz, L. D., and Takahashi, K.(1996) J. Leukocyte Biol. 59, 133–138.
2. Hurley, S. D., and Streit, W. J. (1996) in Topical Issuesin Microglia Research (Ling, E. A., Tan, C. K., and Tan,
nd liver. Sections from brain (A–F) and liver (G–L) were stainedP12 (F and L). A, C, E, G, I, and K indicate GFP fluorescence. Aphs of the same sections. Arrows indicate GFP-positive clusters.
n a-Mgra

C. B. C., Eds.), pp. 1–19, Singapore Neuroscience Assoc., Sin-
1
1
1
16. Sawada, M., Suzumura, A., Tanaka, M., Kato, K., Hosoya, H.,
1
1
1
2
Vol. 262, No. 3, 1999 BIOCHEMICAL AND BIOPHYSICAL RESEARCH COMMUNICATIONS
gapore.3. Persons, D. A., Allay, J. A., Riberdy, J. M., Wersto, R. P.,
Donahue, R. E., Sorrentino, B. P., and Nienhuis, A. W. (1998)Nat. Med. 4, 1201–1205.
4. Okabe, M., Ikawa, M., Kominami, K., Nakanishi, T., and Ni-shimune, Y. (1997) FEBS Lett. 407, 313–319.
5. Ling, V., Luxenberg, D., Wang, J., Nickbarg, E., Leenen, P. J.,Neben, S., and Kobayashi, M. (1997) Eur. J. Immunol. 27, 509–514.
614
and Marunouchi, T. (1996) Dev. Neurosci. 19, 127.7. Tanaka, M., Marunouchi, T., and Sawada, M. (1997) Neurosci.
Lett. 239, 17–20.8. Leenen, P. J., Melis, M., Slieker, W. A., and Van Ewijk, W. (1990)
Eur. J. Immunol. 20, 27–34.9. Slieker, W. A., de Rijk-de Bruijn, M. F., Leenen, P. J., and van
Ewijk, W. (1993) Int. Immunol. 5, 1093–1098.0. Sawada, M., Imai, F., Suzuki, H., Hayakawa, M., Kanno, T., and
Nagatsu, T. (1998) FEBS Lett. 433, 37–40.