Embed Size (px)
Citation preview

Chapter 2
Microsporogenesis and Microspore
Microsporangium
The anther is known as microsporangium. It is the future pollen sac is a cylindrical sac which appears circular in transverse section. It consists of two parts, outer wall and central homogeneous sporogenous tissue. Microsporangial wall has four types of layers:
a. Epidermis (common anther covering)b. Endotheciumc. Middle layer (1-3 middle layers)d. Ttapetum.
The outer three perform the function of protection in the young anther and mechanism of dehiscence in the ripe anther. Both endothecium and tapetum consist of larger cells. In a typical anther the endothecial cells develop fibrous thickenings of a-cellulose on the inner and radial walls and become dead.
Microsporogenesis: The sequential steps of microspore or pollen grain formation
During the development of the microsporangium, the anther is seen at first as a homogeneous mass of meristematic cells, oblong in cross-section and surrounded by an epidermis. It then becomes more or less four-lobed and four longitudinal rows of archesporial cells are differentiated. The archesporial cells are marked off from the surrounding cells by their more deeply staining cytoplasm and conspicuous nuclei.
There may be only one such archesporial cell in each of the four lobes (fig. 404B) as in Boerhaavia, etc., or there may be more of them forming a plate (Ophiopogon, etc.).
Longitudinally, also, there may be one to many of them. Each archesporial cell now cuts off a primary parietal cell towards the epidermis and a primary sporogenous cell on the inner side (Fig. 404C).
The parietal cell now divides by periclinal and anticlinal walls giving rise to several layers of cells forming the wall of the anther while the sporogenous cell usually divides a few times giving rise to a number of microspore or pollen mother cells (Fig. 404D).
The innermost layer of wall cells directly abutting on the sporogenous tissue forms the tapetum which is a nutritive tissue nourishing the developing microspores (Fig. 404E). The wall cells just below the

epidermis form the endothecium which later loses the cell contents, usually becomes fibrous, and forms the dry coat of the mature anther in which the epidermis becomes rather inconspicuous.
Between the tapetum and the endothecium there are one to three middle layers of cells. The middle layers and the tapetum are usually crushed by the time actual meiosis occurs in the sporogenous cells.
ADVERTISEMENTS:
During microsporogenesis (i.e., development of microspores or pollens) the nucleus of each microspore mother cell undergoes meiosis or reduction division ultimately giving rise to four haploid (i.e., possessing ‘n’ number of chromosomes) nuclei (Fig. 404—F & G).
These four nuclei are arranged tetrahedrally (Fig. 404—H & I) and are soon invested with cell walls. Many variations are known of this typical pattern of meiosis, e.g., in maize a wall is formed across the dyad (2-nucleated condition).
They are now the microspores or pollens which soon dry up and become powdery while the tapetum becomes absorbed.
The anther now becomes a dry structure, the partition walls between the sporangia (i.e., loculi) are usually destroyed (Fig. 404J) and the microspores (pollens) are soon liberated by dehiscence of the anther.
The tapetal cells often become multinucleate and play a great part in the nutrition of the pollens. Sometimes they develop a Plasmodium after disintegration and play a part in the development of the exine of the pollen. Even a part of the sporogenous tissue may break down and serve for nutrition instead of developing spores.
While the pollens are dry and powdery in most flowers, peculiar conditions are often let with. In Annona, Elodea, Typha, etc., the four spores in a tetrad never separate but form compound pollen grains.
In the Mimoseae 8 to 64 pollens often aggregate together while in the gynostegium of Calotropis and the gynostemium of orchids all the pollens of each anther lobe from a characteristic mass called pollinium (Fig. 405). Each pollinium is provided with a stalk called caudicle and a sticky base called disc or corpusculum.
The pollen or the microspore is a very minute structure (0.025 to 0.125 mm in diameter). It is unicellular and usually round although it may be oval, pyramidal, polyhedral, etc. It is provided with two coats—an inner, delicate cellulose layer called intine and an outer tough, cutinised layer called exine or extine. The exine is often sculptured or provided with spines, warts, etc. Occasionally, it is smooth.

ADVERTISEMENTS:
The exine may have a waxy coating to render the pollen more or less waterproof. Very often, there are some definitely thinner, circular spots or slits in the exine called germ pores or slits.
These weak spots are utilised during the germination of the pollen. The pores are sometimes provided with lids (Fig. 409 B) which open out like valves during germination. Very often, before the pollen is discharged from the anther it becomes binucleate (Fig. 406), the original single nucleus dividing-into a tube nucleus and a generative nucleus.
The latter, with some cytoplasm surrounding it, becomes the generative cell (described later). Sometimes the pollen may even become trinucleate (Fig. 410), as in most cereal crops, by complete development of the male gametophyte even before it is shed. Fig. 407 shows some different forms of mature pollens. Palynology is the science involving the study of pollens.
Development of The Male Gametophyte:The nucleus of the microspore begins to divide very soon after it is formed. Its division is usually quicker in the warmer climate than in the cooler regions. Such division may begin even before the microspores dissociate from the tetrad condition.
When the pollen is lodged on the stigma, usually its nucleus has already divided. The microspore cell divides into two very equal cells with the smaller cell attached to the wall (Fig. 408A).
The latter soon loses contact with the wall (Fig. 408 B) and becomes the lens-shaped generative cell floating freely in the cytoplasm of the large vegetative or tube cell (Fig. 409A).
It is then in the binucleate (or two-celled) stage. As the pollen germinates and the pollen tube comes out through a germ pore (Fig. 409B) the vegetative nucleus precedes the generative cell (Fig. 409C).
The generative cell soon divides mitotically to form two male gametes (Fig. 409 D) and this act is known as spermatogenesis as the male gametes of Angiosperms are equivalent to the ciliated sperms of the lower groups of plants. The gamete nuclei are enveloped in cytoplasmic sheaths, this forming gamete cells. At a later stage the two male gamete cells are seen to follow the vegetative ‘nucleus (Fig. 409E).
Soon the vegetative nucleus, which seems to be of no importance being a mere vestigial organ, disappears. As the pollen is the microspore, the

contents within the pollen and the pollen tube formed by the germination of the microspore is the male gametophyte of Angiosperms.
While the male gametophyte, thus, usually develops after the pollen has germinated, there are many instances where it develops within the pollen before the pollen tube is formed and, in some cases, even before pollination. Trinucleate pollens with tube nucleus and two gamete cells are observed in such cases (Fig. 410). Such trinucleate pollen formation is the rule in many plants (e.g., Portulaca), specially the cereal crops like wheat, rice, maize of Gramineae and sugarcane of the same family.
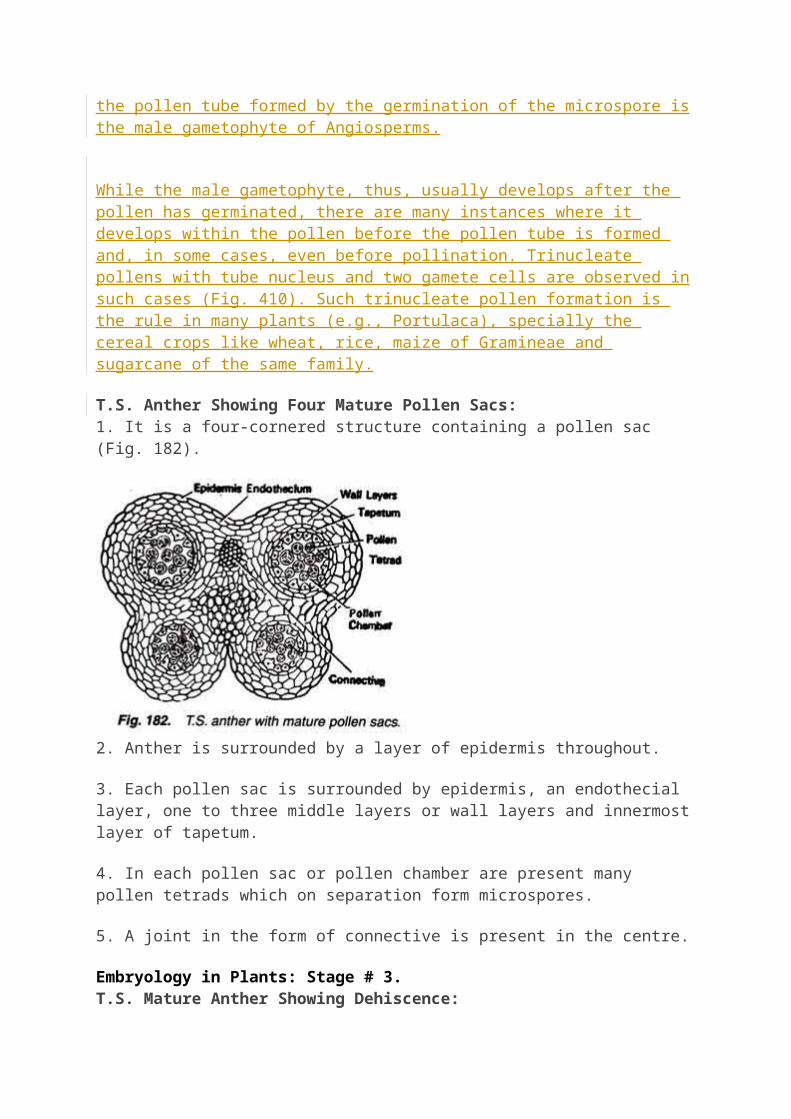
T.S. Anther Showing Four Mature Pollen Sacs:1. It is a four-cornered structure containing a pollen sac (Fig. 182).
2. Anther is surrounded by a layer of epidermis throughout.
3. Each pollen sac is surrounded by epidermis, an endothecial layer, one to three middle layers or wall layers and innermost layer of tapetum.
4. In each pollen sac or pollen chamber are present many pollen tetrads which on separation form microspores.
5. A joint in the form of connective is present in the centre.
Embryology in Plants: Stage # 3.T.S. Mature Anther Showing Dehiscence: It is a four-cornered, four-chambered, multicellular body surrounded by a layer of epidermis.
2. Partition wall between the two pollen sacs is dissolved (Fig. 183).
3. Many pollen grains or microspores are present in the pollen sacs in the form of fine, powdery or granular mass.
4. Endothecium, middle layers and tapetal layers are present below the epidermis.
5. Along the line of dehiscence of each lobe, thin- walled cells of endothecium form the stomium.
6. A connective is very clear.
Embryology in Plants: Stage # 4.
Types of microspore
Types of microspore :-
i) Tetrad antherii) Isobilateral antheriii) Linear tetrad antheriv ) T-shaped antherv) Decussate anthervi) Compound anther :-
Found In asdepiadacae Family , Microspore of an ather lobe remain united with each other & form sac like structure pollirium

@ Microsporangium Structure :-
They are haploid , Minute , Uninucleated, Brown Structure found in many number inside microsporangium formation after meiosis of spore mother Cell- Outer most layer Called Sporoderm Consist Exine & Intine
- Exine :-
it is outermost layer composed by Sporopollenin ( Organic Chemical polymer which is resistant from oxidation leaching ) Ubish Body
- Intine :-
These Intine consist Pectocellulose- In Angisperm Exine form From tapetum during Sporogenesis- In angiosperms Intine form from development of male gametophyte

@ Exine Part :-
i) Sexinei.i) Endoexinei.ii) Tactumii) Nexineii.i) Endexineii.ii) Foot layer
@ Palynology :-
Shape, Size Of microspore & Pollen grain. Exine mIcrosculpturing & Number position Character Of aperture Are great taxonomic importance , These are study under separate Branch Called Polynology- Exine Surface Of Pollen Grain Having Various Pattern of Microsculpturing & Ornamentation Or Discontinues One or more Placed Exine Is Thin Called aperture or Intactate Area through . which pollen tube emerge out at time of germination on stigma
Embryology in Plants: Stage # 5.
Pollen Grain
1. It is a unicellular, uninucleate structure . But pollen grains are always 2- or-3 nucleate, when shed.

2. It is surrounded by a double-layered wall, i. e., outer exine and inner intine.
3. Exine is thick, cutinized, pigmented, sculptured and perforated by germ pores.
4. Intine is thin, colourless, smooth and consists of cellulose.
5. In the cytoplasm are present water, protein, fats, carbohydrates, etc.
Dehiscence of Anther
The mature anther dries up. The sterile strip present between the two pollen sacs of each anther lobe disintegrates to form a single cavity. Therefore, the mature anther has only two cavities or thecae, with one theca in each anther lobe. With the loss of water the differentially thickened dead cells of endothecium contract from their outer thin walls.
The latter become concave. It brings their outer radial walls nearer. As a result the endothecium shortens and ruptures the anther lobe wall in the region of stomium. The line of dehiscence is longitudinal in such cases, e.g., Mustard. Line of dehiscence as well as pollen grains can be observed by placing a finger over the ripe anther. A line of yellowish powdery mass of pollen grains will appear over the finger.
The same can be placed over a drop of water taken on a slide and observed under the microscope. The exposed spores are picked by various agencies for pollination. Local degradation of wall occurs in other types of dehiscence like valves in Barberry, pores in Solanum and irregular rupturing in Najas.
(c) Structure of Pollen Grain:
It is commonly globular in outline, though several other shapes are also found. The diameter is 25-50 µm. There is a highly resistant wall on the outside and cellular contents inside. Its cytoplasm is rich in starch and unsaturated oils. The latter protect the chromosomes from radiation damage.
Pollen grain protoplast is uni-nucleate in the beginning but at the time of liberation it becomes 2-3 celled. Wall or covering of pollen grain is called sporoderm. It has two layers, outer exine and inner intine. Intine is pecto-cellulosic in nature. At places it contains enzymatic proteins (Knox and Heslop-Harrison, 1971). Exine is made of a highly resistant fatty substance called sporopollenin (Zelisch, 1932). Sporopollenin is not degraded by any enzyme.
It is not affected by high temperature, strong acid or strong alkali. Because of the sporopollenin, pollen grains are well preserved as microfossils. At places, exine possesses proteins for enzymatic and compatibility reactions. Exine is differentiated into outer ektexine (sexine) and inner endexine (nexine). Ektexine is further made up

of an inner continuous foot layer, a middle discontinuous baculate layer and outermost discontinuous tectum.
Tectum provides a characteristic sculpturing or designs over the surface of pollen grain, e.g., ridges, tubercles, spines, reticulations. It can help experts to identify the pollen grains and refer them to their family, genus or species. The study of external morphology of mature pollen grains is called palynology.
In insect pollinated pollen grains the exine is spiny as well as covered over by a yellowish, viscous sticky and oily layer called pollenkit. Pollenkit is made up of lipids and carotenoids. At certain places the exine is thin or absent. The areas may have thickened intine or deposition of callose. They are called germ pores (if rounded) or germinal furrows (if elongated). Pollen grains are generally tricolpate (with three germ pores) in dicots and monocolpate (with single germinal furrow) in monocots.
Pollen Viability:It is the period for which pollen grains retain the ability to germinate. Pollen viability is little in flowers which are pollinated in bud condition. It is 30 minutes in Rice and Wheat. In others the period of viability is long, even months in some members of family rosaceae, leguminosae and solanaceae. It, however, depends upon envi-ronmental conditions of temperature and humidity. It is possible to store pollen grains for years in liquid nitrogen (- 196°C) in pollen banks for later use in plant breeding programmes.
Pollen Allergy:Pollen grains are produced in large number, especially in anemophilous species. They float in air and enter respiratory tracts. Some individuals develop allergy to them, producing respiratory disorders like rhinitis, asthma and bronchitis— bronchial allergy. (Hay Fever) The major contributor to pollen allergy is Carrot Grass, Parthenium.
It entered India as contaminant with imported Wheat but has spread in all parts of the country. Chenopodium, Amaranthus, Sorghum, Ricinus, Prosopis, Cynodon are other common sources of pollen allergy. Since different plants produce pollen grains in different seasons, pollen calenders of atmosphere can be scanned and the allergic reaction pinpointed to particular plants.
Pollen Products

The pollen grains especially the ones collected by Bees are being used for a variety of purposes like nature cure, cosmetics and as food supplements. Pollen grains are believed to be rich in nutrients (Protein 7-26%, carbohydrates 24—48%, fats 0.9 – 14.5%). They are taken as tablets of syrups to improve health, enhance performance of athletes and race horses.
(d) Structure and Development of Male Gametophyte (Microgametogenesis
Pre-Pollination Development
Pollen grain or microspore is the first cell of male gametophyte and represents immature male gametophyte. Development of male gametophyte is precocious, that is, it begins inside the microsporangium or pollen sac. When first formed the microspore or young pollen grain has a centrally placed nucleus embedded in dense cytoplasm covered by plasma membrane. It grows in size with the inflow of nutrients. Vacuoles develop and bring about rapid growth of pollen grain.
The vacuoles push the pollen grain nucleus to one side near the wall. The protoplast then divides mitotically to form two unequal cells— small generative cell and large tube or vegetative cell. A layer of callose develops around the generative cell which separates the cell from the pollen grain wall. Later on, callose dissolves and the naked generative cell comes to lie freely in the cytoplasm of the tube cell.
The tube cell has a vacuolate cytoplasm which is rich in the food reserve (starch, protein, fat with mostly unsaturated fatty acids) and cell organelles. Its nucleus becomes large and irregular. The generative cell is spindle shaped to spherical in outline with thin dense cytoplasm surrounding a prominent nucleus.
In some species the generative cell divides into two nonmotile male gametes prior to the dehiscence of anther and release of the pollen grains. Therefore, at the time of pollination, the pollen grain is either 2-celled (tube cell + generative cell; found in over 60% of flowering plants, mostly dicots) or 3-celled (tube cell + two male gametes).

Post-Pollination Development:On the stigma the compatible pollen grain absorbs water and nutrients from the stigmatic secretion through its germ pores. The tube or vegetative cell enlarges. It comes out of the pollen grain through one of the germ pores or germinal furrows to form a pollen tube. The pollen tube is covered over by in-tine.
It was reported by an Italian mathematician Amici (1824) in Portulaca. It secretes pectinases and other hydrolytic enzymes to create a passage for it in the style if the latter is solid. The pollen tube absorbs nourishment from the cells of the style for its growth.
The tube nucleus descends to the tip of the pollen tube. The generative cell (or its products) also passes into it. It soon divides into two nonmotile male gametes if it is not already divided. Each male gamete is lenticular to spherical in outline. It has a large nucleus which is surrounded by a thin sheath of cytoplasm. Each male gamete is considered to be one cell. The tube nucleus may degenerate completely.
The pollen tube is glandular, secretory and absorptive. It has (i) Growth zone at tip (cap block) with dense active cytoplasm rich in vesicles, (ii) Nuclear zone containing tube nucleus and male gametes, (iii) Vacuolization zone rich in vacuolated cytoplasm between active and inactive cytoplasm with a series of callose plugs to separate older inactive cytoplasm.
Pollen tube not only carries male gametes but also secretes hormones and absorb food from style. A pollen grain with pollen tube carrying male gametes represents mature male gameto- phyte. It is 3 celled (one tube cell + 2 male gametes) and 3 nucleated structure formed by two mitotic divisions.

Microgametogenesis (Development of Male Gametophyte):Microspore i.e., the pollen grain, is the first cell of the male gametophyte, which contains only one haploid nucleus. During early stage of development, it remains within the microsporangium. The cell undergoes unequal division and forms a small generative cell and a large vegetative or tube cell (Fig. 3.5B). Initially the generative cell remains lying at one corner of the spore wall.
Within short time, it gets detached and becomes ellipsoid or fusiform in shape (Fig. 3.5C) and remains suspended in the cytoplasm of the vegetative cell (2-celled stage i.e., vegetative cell and generative cell). Later on, the generative cell divides and gives rise to two ellipsoidal or lenticular or spherical cells — the male gametes (3-celled stage i.e., vegetative cell and two male gametes,.
The second division i.e., the division of generative cell, may take place either in the pollen grain or in the pollen tube which develops through germ pore after ollination.
The nucleus of the vegetative cell is commonly known as tube nucleus. It usually shows sign of degeneration with the maturation of generative cell. Finally the tube nucleus remains within spore or may enter the pollen tube. Sooner or later it may be degenerated completely.
Significance of tube nucleus:Earlier workers thought that the tube nucleus had great significance in the direction of growth of the pollen tube, as it is usually present just behind the growing point within the pollen tube.

However, recent workers differ with the above opinion and consider it as a purely non-functional vestigial structure, based on the following facts:1. In branched pollen tube, the tube nucleus remains in one tube, but all the tubes grow normally.
2. It does not always occupy the position behind the growing point within the pollen tube, but in many cases it lies behind the male gametes.
3. In some cases, the growing pollen tube does not have any tube nucleus as it degenerates prior to the development of pollen tube.

Gynoecium is the female reproductive organ, and unit of gynoecium is called pistil or carpel. Carpel is also known as megasporophyll. The carpel is differentiated into three part: Stigma, Style and Ovary.
The gynoecium may consist of a single pistil (monocarpellary) or may have more than one pistil (multicarpellary).
When there are more than one, the pistils may be fused together (syncarpous) or may be free (apocarpous).
A typical carpel is differentiate into 3 parts stigma, style and ovary.
The free end of the carpel which serves as landing plat form pollen grains is called stigma.
A long narrow tubular structure is present in between the stigma and ovary called style.
The basal swollen part of the carpel is called ovary. Ovarian cavity or locule is present inside the ovary. Ovarian
cavity contains the ovules. Each ovule encloses an embryo sac. Number of ovules in an ovary may be one (wheat, paddy, and
mango) to many (papaya, water melon, orchids).
Placentation in ovary
A special tissue that forms the inner wall of ovary bearing one or more ovules is called placenta. The ovules are called megasporangia. Placentation refers to the arrangement of placenta or ovule with in the ovary. There are many types of placentation: marginal, axial, parietal, basal, central and superficial.
The Megasporangium (ovule)
The ovary contains one or more round or oval bodies, known as the ovule. Ovule is also knwon as integumented megasporangium and develops from the base or the inner surface of the ovary.

The ovule is a small structure attached to the placenta by means of a stalk called funicle.
The body of the ovule fuses with funicle in the region called hilum. Thus, hilum represents the junction between ovule and funicle.
Each ovule has one or two protective envelopes called integuments.
Integuments encircle the nucellus except at the tip where a small opening called the micropyle is organized.
Opposite the micropyle end there is the chalaza Chalaza, representing the basal part of the ovule.
Nucellus is mass of cells enclosed within the integuments. Cells of the nucellus have abundant reserve food materials.
Embryo sac or female gametophyte is present in between nucellus. An ovule generally has a single embryo sac formed from a megaspore.
Embryo sac contain 8 cells. 3 antipodal cells, 1 egg cell, 2 synergids, 2 polar nuclei. When embryo sac becomes developed polar nuclei fuse to form a single cell, but nucleus remain unfused. Hence matured embryo sac is 7 cell, 8 nucleated structure.

Egg cell and synergids collectively known as egg apparatus.
Megasporogenesis
The process of formation of megaspores from megaspore mother cell is called megasporogenesis. During the development of ovule, initially all cells of nucellus are undifferentiated, homogneous and meristematic. This mass of cells surrounded by single celled thick layer of epidermis. Any one of hypodermal cell of nucellus is differentiated and increase in size. It becomes different from rest of the cells due to presence of distinct nucleus. It called archesporial cell. [Archesporial cell is hypodermal in origin].
Archesporium divides periclinally to form an outer primary parietal cell and inner primary sporogenous cell.
The primary parietal cell may divide further several times both by anticlinal and periclinical divisions and form parietal tissue. Sometimes it remains undivided.
The primary sporogenous cell usually does not divide further and functions directly as the megaspore mother cell.

It divides meiotically to form, four haploid megaspore. The four haploid megaspore arranged in linear tetrad.
Generally the lower most or chalazal megaspore remains functional out of tetrad of megaspore and other three lie towards the micropyle degenerates.
This functional megaspore produce female gametophyte.
Development of Embryosac or Female gametophyte
Megaspore is first cell of female gametophyte. This megaspore grows in size and obtains nutrition from the
nucellus. The nucleus of megaspore grows in size and obtains nutrition
from the nucellus. The nucleus of megaspore divides mitotically to form a two
nuclei. Each nucleus moves towards the opposite pole and reached at their respective poles.
Both the nuclei lie at poles divide twice mitotically. Resulting 4-4 nuclei are formed at each poles [Total 8-nuclei].
Out of the four, one-one nucleus migrates from the both poles towards the centre. They are known as polar nuclei.
Remaining 3-3 nuclei at each pole surrounded by cytoplasm to form cells as a result of cytokinesis.
Three cells are formed towards the micropyle, collectively known as egg-apparatus. One cell is large and more distinct out of three, called as egg cell (n) and remaining two smaller cells are konwn as synergids (n).
Three cells are formed towards the chalaza are called antipodal cells (n).
Both polar nuclei present at centre, but just before the fertilisation they unite or fuse together in the centre to form secondary nucleus (2n).
This eight nucleated and seven celled structure is called female gametophyte or embryosac.
Fingers like processes are produced from the outer wall of the synergids are known as filifrom apparatus. With the help of these structures, synergids absorb food from the nucellus and

transfer to the embryosac. Filiform also secrete chemicals which attracts the pollen tube.










![Microsporogenesis, structure and viability of pollen in · humidity can contribute to pollen infertility [5]. Survey of literature indicated that there are no report on cytological](https://img.pdfslide.us/doc/110x75/5e19aed5bb69e26dd03fa5d0/microsporogenesis-structure-and-viability-of-pollen-in-humidity-can-contribute.jpg)


















