Embed Size (px)
Citation preview

MicroRNA-206 Regulates Cell Movements during ZebrafishGastrulation by Targeting prickle1a and Regulating c-Jun N-TerminalKinase 2 Phosphorylation
Xiuli Liu,a Guozhu Ning,a Anming Meng,a,b and Qiang Wanga
State Key Laboratory of Biomembrane and Membrane Biotechnology, Institute of Zoology, Chinese Academy of Sciences, Beijing, China,a and Tsinghua-Peking Center forLife Sciences, School of Life Sciences, Tsinghua University, Beijing, Chinab
During vertebrate gastrulation, both concurrent inductive events and cell movements are required for axis formation. Conver-gence and extension (CE) movements contribute to narrowing and lengthening the forming embryonic axis. MicroRNAs(miRNAs) play a critical role in regulating fundamental cellular functions and developmental processes, but their functions inCE movements are not well known. Zebrafish mir206 is maternally expressed and present throughout blastulation and gastrula-tion periods. Either gain or loss of function of mir206 leads to severe defects of convergent extension movements both cell auton-omously and non-cell autonomously. Mosaic lineage tracing studies reveal that the formation of membrane protrusions andactin filaments is disturbed in mir206-overexpressing embryos or mir206 morphants. Mechanistically, mir206 targets prickle1a(pk1a) mRNA and as a result regulates c-Jun N-terminal protein kinase 2 (JNK2) phosphorylation. pk1a overexpression orknockdown can rescue convergent extension defects induced by mir206 overexpression or knockdown, respectively. Therefore,mir206 is an essential, novel regulator for normal convergent and extension movements by regulating mitogen-activated proteinkinase (MAPK) JNK signaling.
During vertebrate gastrulation, the embryonic body plan is es-tablished by coordinated movements of large populations of
cells to generate the ectodermal, mesodermal, and endodermallayers (22, 49, 66). A key driving force of gastrulation is conver-gence and extension (CE) movements. The convergence of cellsnarrows the germ layers and the embryonic body mediolaterally,while extension movement elongates the embryonic tissues fromhead to tail (23, 46). In vertebrates, the dorsal axial and paraxialmesoderms, the notochordal and somitic mesoderms, convergeand extend (24). CE movements also occur in the neuroectodermto narrow and elongate the neural floor plate, which then foldsappositionally and the neural tube is formed (10). Wnt/PCP andBmp pathways play important roles in cell movements duringgastrulation (19, 29, 45, 57, 63). c-Jun N-terminal protein kinase(JNK), the signal transducer and activator of transcription 3(Stat3), and Prickle1 have also been shown to be required fornormal CE movements (5, 47, 52, 70).
The regulation of gene expression by microRNAs (miRNAs)plays a critical role in regulating fundamental cellular functionsand developmental processes (9, 32, 37, 44, 71). Inactivation ofmiRNA biogenesis by the loss of dicer in zebrafish maternal zy-gotic dicer (MZdicer) mutants causes abnormal morphogenesisduring gastrulation and reduces axis extension at later stages (15),suggesting that the CE movements may be disrupted. However,the functional role and molecular mechanisms of miRNAs in reg-ulating cell movements during vertebrate gastrulation have notbeen uncovered.
Previous reports have shown that mir206 is one of the skeletalmuscle-specific miRNAs (8, 16, 43, 67, 71), and it regulates theproliferation and differentiation of muscle progenitor cells (7, 16,27). mir206 is also required for efficient regeneration of neuro-muscular synapse after acute nerve injury, and deficiency ofmir206 in the amyotrophic lateral sclerosis (ALS) mouse modelaccelerates disease progression (68). However, the function of
mir206 in early development of vertebrate embryos has not beenreported.
In zebrafish, mature mir206 is processed from two pre-miRNAtranscripts, mir206-1 and mir206-2. In the present study, we showthat mir206 is maternally expressed, and its transcripts existthroughout the early development of zebrafish embryos. We dem-onstrate that mir206 is essential for normal gastrulation cell move-ments by regulating JNK2 phosphorylation through inhibition ofpk1a expression.
MATERIALS AND METHODSZebrafish strains and antibodies. Wild-type embryos were obtainedfrom natural matings of the zebrafish Tuebingen strain. Embryos weremaintained in Holtfreter’s solution at 28.5°C and staged morphologicallyas described previously (28). The expression of enhanced green fluores-cent protein (EGFP), Pk1a, and phosphorylated and total JNK2 were de-tected by Western blot analysis using the following antibodies: anti-GFPantibody (M20004L; Abmart), anti-Pk1a antibody (55637; ANA SPEC),anti-p-JNK2 antibody (9251; Cell Signaling Technology), and anti-JNK2antibody (sc-571; Santa Cruz).
Constructs. Total RNAs were extracted from 75%-epiboly stage wild-type embryos using TRIzol reagent (Invitrogen) and reversely transcribedwith the ReverTra kit (TOYOBO). The expanded 3= untranslated region(3=UTR) of zebrafish pk1a (NM_183342.2) with a 50-nucleotide sequenceupstream of the stop codon was amplified by reverse transcription (RT)-PCR using the forward primer 5=-ACAGAAGAGAGGACGGAAAGG-3=
Received 27 January 2012 Returned for modification 11 March 2012Accepted 12 May 2012
Published ahead of print 21 May 2012
Address correspondence to Anming Meng, [email protected], orQiang Wang, [email protected].
Copyright © 2012, American Society for Microbiology. All Rights Reserved.
doi:10.1128/MCB.00134-12
2934 mcb.asm.org Molecular and Cellular Biology p. 2934–2942 July 2012 Volume 32 Number 14
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.

and the reverse primer 5=-AATTCCCTCTCAAAGTGGGC-3= and theninserted downstream of the open reading frame of EGFP to generate thereporter EGFP-pk1a-ed3=UTR. Another two reporters for confirmation ofthe mir206 site on pk1a mRNA, EGFP-206wtsite-CDS and EGFP-206mutsite-CDS, were made by fusing the wild-type and mutant forms ofthe mir206 binding site within the pk1a coding region in frame to theEGFP open reading frame. The primers were as follows: for EGFP-206wtsite-CDS, forward primer 5=-CCGGAATTCATGGTGAGCAAGGGCGAG-3= and reverse primer 5=-CCGATCGATTTATGAAATAATACAGTTTTTGCCTTTCTTGTACAGCTCGTC-3=; for EGFP-206mutsite-CDS,forward primer 5=-CCGGAATTCATGGTGAGCAAGGGCGAG-3= andreverse primer 5=-CCGATCGATTTAACTTTTAATACAGTTTTTGCCTTTCTTGTACAGCTCGTC-3= (mir206 binding sites are underlined, andthe mutations are italic). The full coding sequence of pk1a was amplifiedusing the forward primer 5=-ATGGAGCTGGAGAATCACGG-3= and thereverse primer 5=-TTATGAAATAATACAGTTTTTGCCTTTC-3= andthen cloned into the pCS-Flag vector. All the sequences were confirmed byDNA sequencing, and the expression of pk1a was verified by Westernblotting using anti-Flag antibody (M20008L; Abmart).
Morpholinos, microinjection, and in situ hybridization. Themir206-1 morpholino (206-MO1) (5=-ACCACACACTTCCTTACATTCCATAACTTG-3=) and mir206-2 morpholino (206-MO2) (5=-GCCACACACTTCCTTACATTCCATAGATTA-3=) were designed complemen-tary to the miRNA guide strand and the Dicer nucleolytic processing sites,respectively, according to the sequences of mir206-1 and mir206-2 pre-cursors. The following control MO, including six mismatched nucleotidesboth in the miRNA guide strand and the Dicer nucleolytic processing sites(underlined, mis-MO), was designed: 5=-ACGACACAGTTCCTTAGATTGCATAAGTTC-3=. The pk1a morpholino (pk1a-MO) (5=-GCCCACCGTGATTCTCCAGCTCCAT-3=) was designed to block the translationstart site as reported (5). All of these MOs and the morpholino targetingp53 (p53-MO; 5=-GCGCCATTGCTTTGCAAGAATTG-3=) were synthe-sized by Gene Tools and resuspended in nuclease-free water. For mir206knockdown experiments, 8 ng 206-MO1 or 206-MO2 or 4 ng each MOs(206-MOs) per embryo was injected, and 8 ng mir206 mis-MO was in-jected as a control. A total of 4 ng p53-MO was injected with 8 ng 206-MOs to exclude the possibility of the off-target effect. For pk1a-MO, 0.5ng per embryo was injected in the knockdown experiments, while 0.1 ngwas coinjected with 8 ng 206-MOs in rescue experiments. Whole-mountin situ hybridization was performed as previously described (36, 67).
Stem-loop RT-PCR. Stem-loop RT-PCR was performed as previouslydescribed (30, 64). Total RNAs were reversely transcribed using themir206 RT primer 5=-GTCGTATCCAGTGCAGGGTCCGAGGTATTCGCACTGGATACGACCCACAC-3= and the U6 RT primer 5=-AAAAATATGGAGCGCTTCACG-3=. The PCR primers were listed as follows: forU6, forward primer 5=-TTGGTCTGATCTGGCACATATAC-3= and re-verse primer 5=-AAAAATATGGAGCGCTTCACG-3=; for mir206, for-ward primer 5=-GCGTCTGGAATGTAAGGAAGTG-3= and reverseprimer 5=-GTGCAGGGTCCGAGGT-3=.
RNA synthesis. Capped RNAs for pk1a, mir206-1 precursor, mem-brane-located mCherry, or EGFP-pk1a-3=UTR were in vitro transcribedusing the mMessage mMachine kit (Ambion) according to the manufac-turer’s instructions. The mir206 duplex was synthesized by GenePharmaCompany. Digoxigenin-UTP-labeled antisense RNA probes were in vitrotranscribed using the MEGAscript kit (Ambion) according to the manu-facturer’s instructions. The digoxigenin-labeled locked nucleic acid(LNA) antisense mir206 probe was synthesized by Exiqon.
Measurement of angles between the anterior and posterior ends. Forembryos at 10 h postfertilization (hpf), the angle was defined as that be-tween the anterior prechordal plate (polster) and the middle of the tail-bud; and for embryos at 13 hpf, it was between the anterior prechordalplate and the end of the tailbud. Such analyses were repeated three times(with at least 40 embryos each).
Cell transplantation. For cell-autonomous assay, donor embryoswere first coinjected with 1 nl of 0.5% rhodamine-dextran (molecular
weight [MW] of 10,000; Molecular Probes) together with 200 pg of syn-thesized mir206 precursor RNA or 8 ng 206-MOs at the one-cell stage.Lateral or axial mesendodermal cells (30 to 50 cells) of donor embryoswere transplanted to the lateral margin or embryonic shield of wild-typehosts at the same developmental stage (shield stage, about 6 hpf). Imageswere taken at 8 hpf and 10 hpf under a Zeiss AxioPlan-2 microscope.
For non-cell-autonomous assay, wild-type donor embryos were in-jected with rhodamine-dextran, and host embryos were injected withmir206 precursor RNA or 206-MOs at the one-cell stage. Lateral or axialmesendodermal cells of donor embryos were also transplanted at theshield stage, and the locations of the labeled donor cells were monitored at8 hpf and 10 hpf.
Detection of membrane protrusions. The convergent extensionmovements of small clones within the whole embryo during gastrulationwere analyzed as previously described (14, 34). One cell of a 64-cell-stageembryo was injected with 10 pg membrane mCherry mRNA alone or incombination with either 13 pg mir206 precursor or 0.5 ng 206-MOs. Thelabeled cells were imaged at the 75%-epiboly stage by confocal micros-copy. The number of membrane protrusions in the cytoplasm wascounted (20 cells per embryo, a total of five embryos for each group).These experiments were repeated three times, and Student’s t tests wereperformed to assess the significance of differences between groups.
Staining of actin filaments in embryos. Embryos were injected with200 pg mir206 precursor RNA or 8 ng 206-MOs at the one-cell stage andharvested at the shield and 75%-epiboly stages for fixing in 4% parafor-maldehyde (PFA) in 1� phosphate-buffered saline (PBS). On the secondday, fixed embryos were stained by phalloidin-fluorescein isothiocyanate(FITC) (P5282; Sigma) for 25 min and washed 3 times with 1� PBS. Thelateral epiblast cells were photographed by confocal microscopy. The cy-toplasmic actin filaments were counted (20 cells per embryo, a total of fiveembryos for each group). These experiments were repeated three times,and Student’s t tests were performed.
RESULTSmir206 is required for cell movements during normal gastrula-tion. mir206 is evolutionally conserved in human and other ver-tebrates, and its expression is restricted to skeletal myoblasts insomites, limb buds, and head muscles (7, 27, 71). Zebrafish mir206has been found to be one of the most abundant miRNAs fromsegmentation onward during embryogenesis (8, 67). We exam-ined mir206 expression in zebrafish embryos from the one-cellstage to the tailbud stage by stem-loop RT-PCR (Fig. 1A). Wenoted that mir206 was maternally expressed since its transcriptswere present from the one-cell to the 1,000-cell stages. Theamount of mir206 was obviously decreased at the sphere stage andthen started to increase at the onset of gastrulation (shield stage),suggesting that its zygotic expression compensates for clearance ofmaternal transcripts.
To efficiently block mir206 activity during early development,two independent antisense morpholinos (206-MO1 and 206-MO2) complementary to the miRNA guide strand and the Dicernucleolytic processing sites of each pre-miRNA transcripts weredesigned to prevent correct maturation of mir206. Injection of 8ng 206-MO1 or 206-MO2 or coinjection of 4 ng each MOs (206-MOs) resulted in an effective blockade of mir206 (Fig. 1B). Mor-phological observations revealed that mir206 knockdown with206-MO1, 206-MO2, or 206-MOs led to a wider angle betweenthe anterior and posterior ends of embryos from 10 to 13 hpf(Fig. 1C and D), which was indicative of defective anterior-poste-rior extension. The defect could neither be aggravated nor com-promised by co-knockdown of p53 (Fig. 1C), which excluded theinvolvement of morpholino-mediated cell apoptosis (40). Since206-MOs appeared to be more effective, we therefore used 206-
mir206 Regulates CE Movements during Gastrulation
July 2012 Volume 32 Number 14 mcb.asm.org 2935
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.
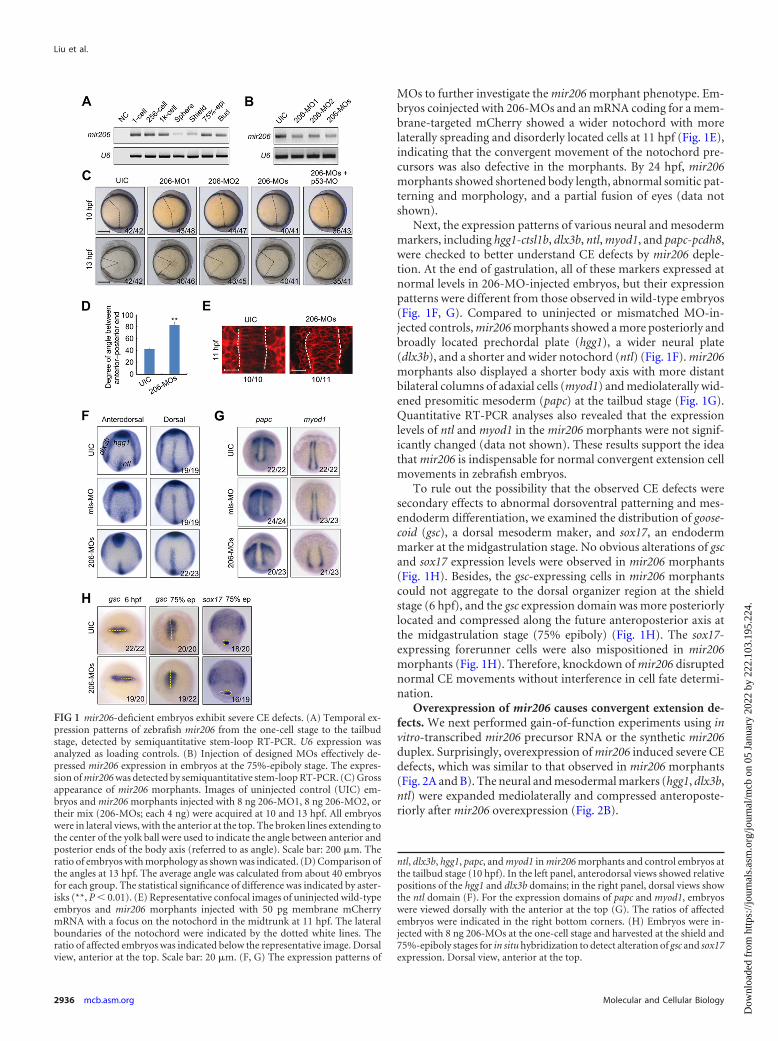
MOs to further investigate the mir206 morphant phenotype. Em-bryos coinjected with 206-MOs and an mRNA coding for a mem-brane-targeted mCherry showed a wider notochord with morelaterally spreading and disorderly located cells at 11 hpf (Fig. 1E),indicating that the convergent movement of the notochord pre-cursors was also defective in the morphants. By 24 hpf, mir206morphants showed shortened body length, abnormal somitic pat-terning and morphology, and a partial fusion of eyes (data notshown).
Next, the expression patterns of various neural and mesodermmarkers, including hgg1-ctsl1b, dlx3b, ntl, myod1, and papc-pcdh8,were checked to better understand CE defects by mir206 deple-tion. At the end of gastrulation, all of these markers expressed atnormal levels in 206-MO-injected embryos, but their expressionpatterns were different from those observed in wild-type embryos(Fig. 1F, G). Compared to uninjected or mismatched MO-in-jected controls, mir206 morphants showed a more posteriorly andbroadly located prechordal plate (hgg1), a wider neural plate(dlx3b), and a shorter and wider notochord (ntl) (Fig. 1F). mir206morphants also displayed a shorter body axis with more distantbilateral columns of adaxial cells (myod1) and mediolaterally wid-ened presomitic mesoderm (papc) at the tailbud stage (Fig. 1G).Quantitative RT-PCR analyses also revealed that the expressionlevels of ntl and myod1 in the mir206 morphants were not signif-icantly changed (data not shown). These results support the ideathat mir206 is indispensable for normal convergent extension cellmovements in zebrafish embryos.
To rule out the possibility that the observed CE defects weresecondary effects to abnormal dorsoventral patterning and mes-endoderm differentiation, we examined the distribution of goose-coid (gsc), a dorsal mesoderm maker, and sox17, an endodermmarker at the midgastrulation stage. No obvious alterations of gscand sox17 expression levels were observed in mir206 morphants(Fig. 1H). Besides, the gsc-expressing cells in mir206 morphantscould not aggregate to the dorsal organizer region at the shieldstage (6 hpf), and the gsc expression domain was more posteriorlylocated and compressed along the future anteroposterior axis atthe midgastrulation stage (75% epiboly) (Fig. 1H). The sox17-expressing forerunner cells were also mispositioned in mir206morphants (Fig. 1H). Therefore, knockdown of mir206 disruptednormal CE movements without interference in cell fate determi-nation.
Overexpression of mir206 causes convergent extension de-fects. We next performed gain-of-function experiments using invitro-transcribed mir206 precursor RNA or the synthetic mir206duplex. Surprisingly, overexpression of mir206 induced severe CEdefects, which was similar to that observed in mir206 morphants(Fig. 2A and B). The neural and mesodermal markers (hgg1, dlx3b,ntl) were expanded mediolaterally and compressed anteroposte-riorly after mir206 overexpression (Fig. 2B).
FIG 1 mir206-deficient embryos exhibit severe CE defects. (A) Temporal ex-pression patterns of zebrafish mir206 from the one-cell stage to the tailbudstage, detected by semiquantitative stem-loop RT-PCR. U6 expression wasanalyzed as loading controls. (B) Injection of designed MOs effectively de-pressed mir206 expression in embryos at the 75%-epiboly stage. The expres-sion of mir206 was detected by semiquantitative stem-loop RT-PCR. (C) Grossappearance of mir206 morphants. Images of uninjected control (UIC) em-bryos and mir206 morphants injected with 8 ng 206-MO1, 8 ng 206-MO2, ortheir mix (206-MOs; each 4 ng) were acquired at 10 and 13 hpf. All embryoswere in lateral views, with the anterior at the top. The broken lines extending tothe center of the yolk ball were used to indicate the angle between anterior andposterior ends of the body axis (referred to as angle). Scale bar: 200 �m. Theratio of embryos with morphology as shown was indicated. (D) Comparison ofthe angles at 13 hpf. The average angle was calculated from about 40 embryosfor each group. The statistical significance of difference was indicated by aster-isks (**, P � 0.01). (E) Representative confocal images of uninjected wild-typeembryos and mir206 morphants injected with 50 pg membrane mCherrymRNA with a focus on the notochord in the midtrunk at 11 hpf. The lateralboundaries of the notochord were indicated by the dotted white lines. Theratio of affected embryos was indicated below the representative image. Dorsalview, anterior at the top. Scale bar: 20 �m. (F, G) The expression patterns of
ntl, dlx3b, hgg1, papc, and myod1 in mir206 morphants and control embryos atthe tailbud stage (10 hpf). In the left panel, anterodorsal views showed relativepositions of the hgg1 and dlx3b domains; in the right panel, dorsal views showthe ntl domain (F). For the expression domains of papc and myod1, embryoswere viewed dorsally with the anterior at the top (G). The ratios of affectedembryos were indicated in the right bottom corners. (H) Embryos were in-jected with 8 ng 206-MOs at the one-cell stage and harvested at the shield and75%-epiboly stages for in situ hybridization to detect alteration of gsc and sox17expression. Dorsal view, anterior at the top.
Liu et al.
2936 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.

Recently, many off-target effects have been observed in mi-croRNA or RNA interference (RNAi) experiments (3, 60). Here,we coinjected 1 nl synthetic mir206 duplex at different concentra-tions with a fixed dose (8 ng) of 206-MOs and examined myod1and ntl expression at the tailbud stage. Results showed that themir206 duplex could gradually rescue CE defects of mir206 mor-phants in a dose-dependent manner; in particular, injection of 20�M mir206 duplex rescued mir206 morphants to almost normalmorphology; a higher concentration (40 �M) of the mir206 du-plex failed to rescue CE defects in mir206 morphants (Fig. 2C).These data suggest the specific effects of mir206 knockdown andoverexpression. Thus, mir206 plays a vital role in controlling cellmovements during gastrulation, but the correct amount isimportant.
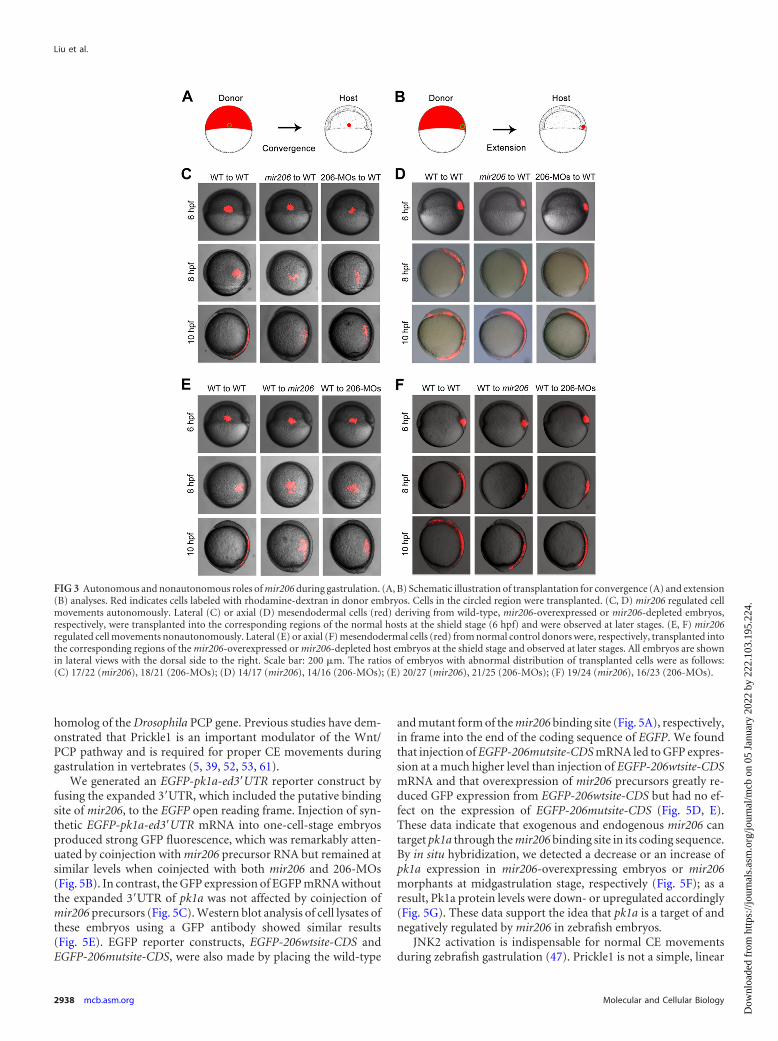
mir206 regulates cell movements in both cell-autonomousand non-cell-autonomous manners. To further clarify howmir206 affects cell movements during gastrulation, rhodamine-dextran and mir206 precursor RNA or 206-MOs were coinjectedinto one-cell-stage donor embryos. Lateral mesendodermal cells(LME) from the lateral blastoderm margin of these donor em-bryos were transplanted to the same location of host normal em-bryos at the shield stage and imaged during gastrulation (Fig. 3A).During gastrulation, the LME donor cells deriving from unin-jected embryos migrated into the dorsal sides of host embryos atthe end of gastrulation as expected (48, 70). However, the conver-
gence movements toward the dorsal side were severely impaired inmir206-overexpressed or -deficient LME donor cells (Fig. 3C). Wethen tested the extension movements by transplanting axial mes-endodermal cells (AME) from different donor embryos deeplyinto the embryonic shield of normal host embryos (Fig. 3B). Asexpected, the AME cells from the control donors migrated alongthe anterior-posterior axis and nearly reached the forebrain andtailbud at the end of gastrulation. However, the AME cells frommir206-overexpressed or -deficient donors showed severely re-duced extension ability, as these cells were distributed in a shorterregion along the anterioposterior axis (Fig. 3D). These results to-gether suggest that mir206 regulates the dorsal convergence of thelateral mesendoderm and the anterior-posterior extension of theaxial mesendoderm in a cell-autonomous manner.
We next tested whether mir206 could also achieve its functionin a non-cell-autonomous fashion. Rhodamine-dextran-labeledLME and AME cells from normal donor embryos at the shieldstage were transplanted, respectively, to normal, mir206-overex-pressed or 206-MO-injected embryos. The transplanted wild-typeLME and AME cells could not migrate into their destinations inthe mir206-overexpressed or 206-MO-injected embryos duringgastrulation compared with the control (Fig. 3E and F), indicatingthat mir206 also regulates gastrulation cell movements non-cellautonomously.
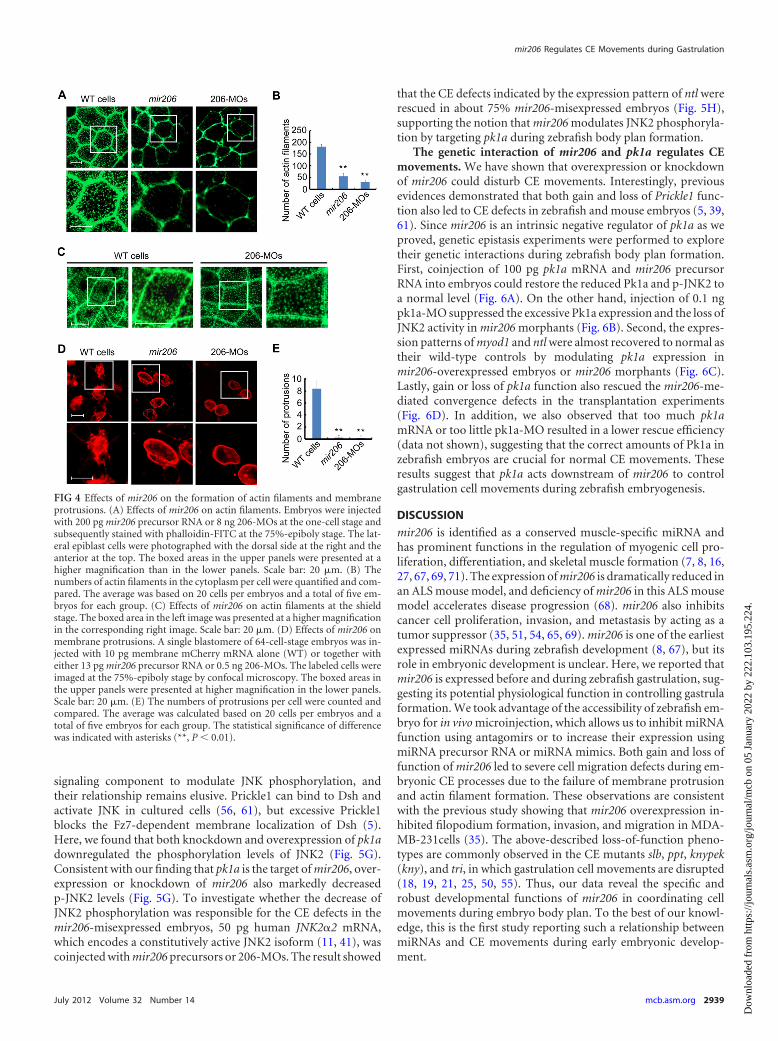
Overexpression or knockdown of mir206 impairs membraneprotrusion and actin filament formation. Migration of gastrula-tion cell movements depends on the formation of actin filamentsand cell membrane protrusions, including lamellipodia and filop-odia (1, 20, 63). We examined actin filaments, by phalloidin-FITCstaining, in embryos injected with mir206 precursor RNA or 206-MOs. Compared to the uninjected control embryos, injected em-bryos at the 75%-epiboly stage had less abundant cortical actinfilaments at cell-cell junctions and fewer F-actin bundles in thecytoplasm (Fig. 4A and B). In addition, we also examined actinfilaments in mir206-deficient embryos at the onset of gastrulation.It appeared that the actin filament formation was not obviouslyaffected in mir206 morphants at the shield stage (Fig. 4C), whichmay be explained by the fact that small amounts of mir206(Fig. 1A) due to the clearance of its maternal transcripts andshorter period of its zygotic expression would not be sufficient forinterfering with actin filament formation. The membrane protru-sions were observed by confocal microscopy following injection ofan mRNA encoding the membrane-targeted mCherry togetherwith mir206 precursor RNA or 206-MOs into one cell of 64-cell-stage embryos. The descendants of mir206-overexpressed or 206-MO-injected cells at the 75%-epiboly stage displayed roundedmorphology with few or absent protrusions, which were strikinglydifferent from wild-type control cells (Fig. 4D and E). These ob-servations suggest that the membrane protrusion- and actin fila-ment-mediated cell migration is disturbed during gastrulation.
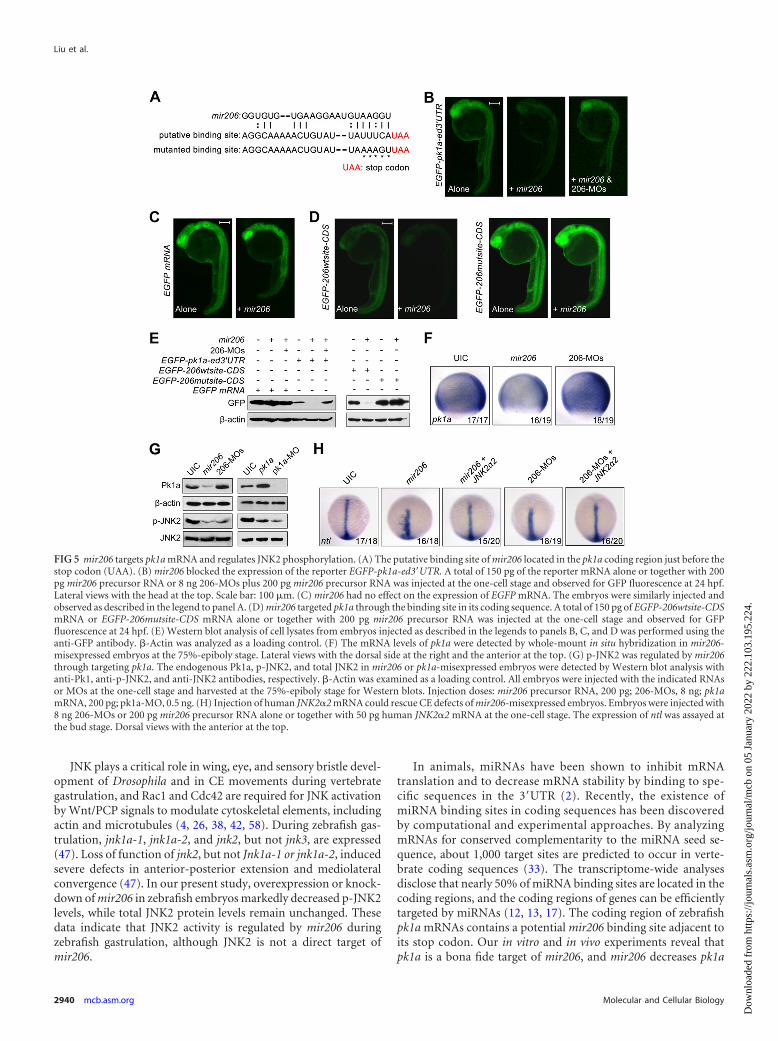
mir206 modulates mitogen-activated protein kinase(MAPK) JNK2 phosphorylation by regulating pk1a expression.In animals, recognition sites for miRNAs have been reported to belocated in the 3=UTR as well as in the coding regions of mRNAs,and miRNAs have the ability to directly target transcripts withintheir coding region to suppress gene expression (12, 13, 17, 33).Using the target prediction algorithm (RNA22 microRNA targetdetection [http://cbcsrv.watson.ibm.com/rna22.html]), we foundone putative binding site of mir206 located in the pk1a codingregion adjacent to the stop codon (Fig. 5A). pk1a is a zebrafish
FIG 2 Overexpression of mir206 causes CE defects. (A) mir206-overexpressedembryos showed increased angles between the anterior-posterior ends com-pared with the uninjected control. A total of 200 pg mir206 precursor RNA wasinjected at the one-cell stage. The stages and ratios of affected embryos areindicated. All embryos were viewed laterally with the anterior at the top. Scalebar: 200 �m. (B) Expression patterns of hgg1, dlx3b, and ntl in UIC andmir206-overexpressed embryos at the tailbud stage. Embryos were anterodor-sally viewed to show the hgg1 and dlx3 domains (top) and dorsally viewed forthe ntl domain (bottom), with the anterior at the top. (C) Injection of mir206duplex could rescue CE defects of mir206 morphants. Embryos were coin-jected with 8 ng 206-MOs and an indicated amount of mir206 duplex at theone-cell stage. The expression domains of myod1 and ntl were assayed at thebud stage. Dorsal views with the anterior at the top.
mir206 Regulates CE Movements during Gastrulation
July 2012 Volume 32 Number 14 mcb.asm.org 2937
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.
homolog of the Drosophila PCP gene. Previous studies have dem-onstrated that Prickle1 is an important modulator of the Wnt/PCP pathway and is required for proper CE movements duringgastrulation in vertebrates (5, 39, 52, 53, 61).
We generated an EGFP-pk1a-ed3=UTR reporter construct byfusing the expanded 3=UTR, which included the putative bindingsite of mir206, to the EGFP open reading frame. Injection of syn-thetic EGFP-pk1a-ed3=UTR mRNA into one-cell-stage embryosproduced strong GFP fluorescence, which was remarkably atten-uated by coinjection with mir206 precursor RNA but remained atsimilar levels when coinjected with both mir206 and 206-MOs(Fig. 5B). In contrast, the GFP expression of EGFP mRNA withoutthe expanded 3=UTR of pk1a was not affected by coinjection ofmir206 precursors (Fig. 5C). Western blot analysis of cell lysates ofthese embryos using a GFP antibody showed similar results(Fig. 5E). EGFP reporter constructs, EGFP-206wtsite-CDS andEGFP-206mutsite-CDS, were also made by placing the wild-type
and mutant form of the mir206 binding site (Fig. 5A), respectively,in frame into the end of the coding sequence of EGFP. We foundthat injection of EGFP-206mutsite-CDS mRNA led to GFP expres-sion at a much higher level than injection of EGFP-206wtsite-CDSmRNA and that overexpression of mir206 precursors greatly re-duced GFP expression from EGFP-206wtsite-CDS but had no ef-fect on the expression of EGFP-206mutsite-CDS (Fig. 5D, E).These data indicate that exogenous and endogenous mir206 cantarget pk1a through the mir206 binding site in its coding sequence.By in situ hybridization, we detected a decrease or an increase ofpk1a expression in mir206-overexpressing embryos or mir206morphants at midgastrulation stage, respectively (Fig. 5F); as aresult, Pk1a protein levels were down- or upregulated accordingly(Fig. 5G). These data support the idea that pk1a is a target of andnegatively regulated by mir206 in zebrafish embryos.
JNK2 activation is indispensable for normal CE movementsduring zebrafish gastrulation (47). Prickle1 is not a simple, linear
FIG 3 Autonomous and nonautonomous roles of mir206 during gastrulation. (A, B) Schematic illustration of transplantation for convergence (A) and extension(B) analyses. Red indicates cells labeled with rhodamine-dextran in donor embryos. Cells in the circled region were transplanted. (C, D) mir206 regulated cellmovements autonomously. Lateral (C) or axial (D) mesendodermal cells (red) deriving from wild-type, mir206-overexpressed or mir206-depleted embryos,respectively, were transplanted into the corresponding regions of the normal hosts at the shield stage (6 hpf) and were observed at later stages. (E, F) mir206regulated cell movements nonautonomously. Lateral (E) or axial (F) mesendodermal cells (red) from normal control donors were, respectively, transplanted intothe corresponding regions of the mir206-overexpressed or mir206-depleted host embryos at the shield stage and observed at later stages. All embryos are shownin lateral views with the dorsal side to the right. Scale bar: 200 �m. The ratios of embryos with abnormal distribution of transplanted cells were as follows:(C) 17/22 (mir206), 18/21 (206-MOs); (D) 14/17 (mir206), 14/16 (206-MOs); (E) 20/27 (mir206), 21/25 (206-MOs); (F) 19/24 (mir206), 16/23 (206-MOs).
Liu et al.
2938 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.
signaling component to modulate JNK phosphorylation, andtheir relationship remains elusive. Prickle1 can bind to Dsh andactivate JNK in cultured cells (56, 61), but excessive Prickle1blocks the Fz7-dependent membrane localization of Dsh (5).Here, we found that both knockdown and overexpression of pk1adownregulated the phosphorylation levels of JNK2 (Fig. 5G).Consistent with our finding that pk1a is the target of mir206, over-expression or knockdown of mir206 also markedly decreasedp-JNK2 levels (Fig. 5G). To investigate whether the decrease ofJNK2 phosphorylation was responsible for the CE defects in themir206-misexpressed embryos, 50 pg human JNK2�2 mRNA,which encodes a constitutively active JNK2 isoform (11, 41), wascoinjected with mir206 precursors or 206-MOs. The result showed
that the CE defects indicated by the expression pattern of ntl wererescued in about 75% mir206-misexpressed embryos (Fig. 5H),supporting the notion that mir206 modulates JNK2 phosphoryla-tion by targeting pk1a during zebrafish body plan formation.
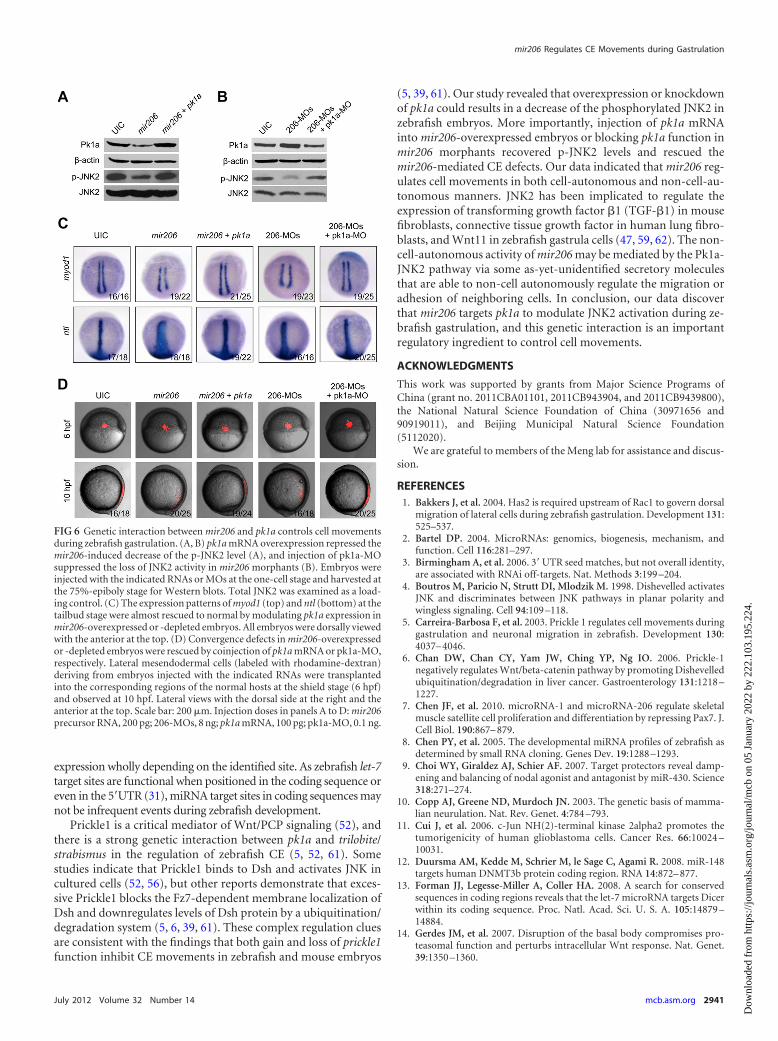
The genetic interaction of mir206 and pk1a regulates CEmovements. We have shown that overexpression or knockdownof mir206 could disturb CE movements. Interestingly, previousevidences demonstrated that both gain and loss of Prickle1 func-tion also led to CE defects in zebrafish and mouse embryos (5, 39,61). Since mir206 is an intrinsic negative regulator of pk1a as weproved, genetic epistasis experiments were performed to exploretheir genetic interactions during zebrafish body plan formation.First, coinjection of 100 pg pk1a mRNA and mir206 precursorRNA into embryos could restore the reduced Pk1a and p-JNK2 toa normal level (Fig. 6A). On the other hand, injection of 0.1 ngpk1a-MO suppressed the excessive Pk1a expression and the loss ofJNK2 activity in mir206 morphants (Fig. 6B). Second, the expres-sion patterns of myod1 and ntl were almost recovered to normal astheir wild-type controls by modulating pk1a expression inmir206-overexpressed embryos or mir206 morphants (Fig. 6C).Lastly, gain or loss of pk1a function also rescued the mir206-me-diated convergence defects in the transplantation experiments(Fig. 6D). In addition, we also observed that too much pk1amRNA or too little pk1a-MO resulted in a lower rescue efficiency(data not shown), suggesting that the correct amounts of Pk1a inzebrafish embryos are crucial for normal CE movements. Theseresults suggest that pk1a acts downstream of mir206 to controlgastrulation cell movements during zebrafish embryogenesis.
DISCUSSION
mir206 is identified as a conserved muscle-specific miRNA andhas prominent functions in the regulation of myogenic cell pro-liferation, differentiation, and skeletal muscle formation (7, 8, 16,27, 67, 69, 71). The expression of mir206 is dramatically reduced inan ALS mouse model, and deficiency of mir206 in this ALS mousemodel accelerates disease progression (68). mir206 also inhibitscancer cell proliferation, invasion, and metastasis by acting as atumor suppressor (35, 51, 54, 65, 69). mir206 is one of the earliestexpressed miRNAs during zebrafish development (8, 67), but itsrole in embryonic development is unclear. Here, we reported thatmir206 is expressed before and during zebrafish gastrulation, sug-gesting its potential physiological function in controlling gastrulaformation. We took advantage of the accessibility of zebrafish em-bryo for in vivo microinjection, which allows us to inhibit miRNAfunction using antagomirs or to increase their expression usingmiRNA precursor RNA or miRNA mimics. Both gain and loss offunction of mir206 led to severe cell migration defects during em-bryonic CE processes due to the failure of membrane protrusionand actin filament formation. These observations are consistentwith the previous study showing that mir206 overexpression in-hibited filopodium formation, invasion, and migration in MDA-MB-231cells (35). The above-described loss-of-function pheno-types are commonly observed in the CE mutants slb, ppt, knypek(kny), and tri, in which gastrulation cell movements are disrupted(18, 19, 21, 25, 50, 55). Thus, our data reveal the specific androbust developmental functions of mir206 in coordinating cellmovements during embryo body plan. To the best of our knowl-edge, this is the first study reporting such a relationship betweenmiRNAs and CE movements during early embryonic develop-ment.
FIG 4 Effects of mir206 on the formation of actin filaments and membraneprotrusions. (A) Effects of mir206 on actin filaments. Embryos were injectedwith 200 pg mir206 precursor RNA or 8 ng 206-MOs at the one-cell stage andsubsequently stained with phalloidin-FITC at the 75%-epiboly stage. The lat-eral epiblast cells were photographed with the dorsal side at the right and theanterior at the top. The boxed areas in the upper panels were presented at ahigher magnification than in the lower panels. Scale bar: 20 �m. (B) Thenumbers of actin filaments in the cytoplasm per cell were quantified and com-pared. The average was based on 20 cells per embryos and a total of five em-bryos for each group. (C) Effects of mir206 on actin filaments at the shieldstage. The boxed area in the left image was presented at a higher magnificationin the corresponding right image. Scale bar: 20 �m. (D) Effects of mir206 onmembrane protrusions. A single blastomere of 64-cell-stage embryos was in-jected with 10 pg membrane mCherry mRNA alone (WT) or together witheither 13 pg mir206 precursor RNA or 0.5 ng 206-MOs. The labeled cells wereimaged at the 75%-epiboly stage by confocal microscopy. The boxed areas inthe upper panels were presented at higher magnification in the lower panels.Scale bar: 20 �m. (E) The numbers of protrusions per cell were counted andcompared. The average was calculated based on 20 cells per embryos and atotal of five embryos for each group. The statistical significance of differencewas indicated with asterisks (**, P � 0.01).
mir206 Regulates CE Movements during Gastrulation
July 2012 Volume 32 Number 14 mcb.asm.org 2939
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.
JNK plays a critical role in wing, eye, and sensory bristle devel-opment of Drosophila and in CE movements during vertebrategastrulation, and Rac1 and Cdc42 are required for JNK activationby Wnt/PCP signals to modulate cytoskeletal elements, includingactin and microtubules (4, 26, 38, 42, 58). During zebrafish gas-trulation, jnk1a-1, jnk1a-2, and jnk2, but not jnk3, are expressed(47). Loss of function of jnk2, but not Jnk1a-1 or jnk1a-2, inducedsevere defects in anterior-posterior extension and mediolateralconvergence (47). In our present study, overexpression or knock-down of mir206 in zebrafish embryos markedly decreased p-JNK2levels, while total JNK2 protein levels remain unchanged. Thesedata indicate that JNK2 activity is regulated by mir206 duringzebrafish gastrulation, although JNK2 is not a direct target ofmir206.
In animals, miRNAs have been shown to inhibit mRNAtranslation and to decrease mRNA stability by binding to spe-cific sequences in the 3=UTR (2). Recently, the existence ofmiRNA binding sites in coding sequences has been discoveredby computational and experimental approaches. By analyzingmRNAs for conserved complementarity to the miRNA seed se-quence, about 1,000 target sites are predicted to occur in verte-brate coding sequences (33). The transcriptome-wide analysesdisclose that nearly 50% of miRNA binding sites are located in thecoding regions, and the coding regions of genes can be efficientlytargeted by miRNAs (12, 13, 17). The coding region of zebrafishpk1a mRNAs contains a potential mir206 binding site adjacent toits stop codon. Our in vitro and in vivo experiments reveal thatpk1a is a bona fide target of mir206, and mir206 decreases pk1a
FIG 5 mir206 targets pk1a mRNA and regulates JNK2 phosphorylation. (A) The putative binding site of mir206 located in the pk1a coding region just before thestop codon (UAA). (B) mir206 blocked the expression of the reporter EGFP-pk1a-ed3=UTR. A total of 150 pg of the reporter mRNA alone or together with 200pg mir206 precursor RNA or 8 ng 206-MOs plus 200 pg mir206 precursor RNA was injected at the one-cell stage and observed for GFP fluorescence at 24 hpf.Lateral views with the head at the top. Scale bar: 100 �m. (C) mir206 had no effect on the expression of EGFP mRNA. The embryos were similarly injected andobserved as described in the legend to panel A. (D) mir206 targeted pk1a through the binding site in its coding sequence. A total of 150 pg of EGFP-206wtsite-CDSmRNA or EGFP-206mutsite-CDS mRNA alone or together with 200 pg mir206 precursor RNA was injected at the one-cell stage and observed for GFPfluorescence at 24 hpf. (E) Western blot analysis of cell lysates from embryos injected as described in the legends to panels B, C, and D was performed using theanti-GFP antibody. �-Actin was analyzed as a loading control. (F) The mRNA levels of pk1a were detected by whole-mount in situ hybridization in mir206-misexpressed embryos at the 75%-epiboly stage. Lateral views with the dorsal side at the right and the anterior at the top. (G) p-JNK2 was regulated by mir206through targeting pk1a. The endogenous Pk1a, p-JNK2, and total JNK2 in mir206 or pk1a-misexpressed embryos were detected by Western blot analysis withanti-Pk1, anti-p-JNK2, and anti-JNK2 antibodies, respectively. �-Actin was examined as a loading control. All embryos were injected with the indicated RNAsor MOs at the one-cell stage and harvested at the 75%-epiboly stage for Western blots. Injection doses: mir206 precursor RNA, 200 pg; 206-MOs, 8 ng; pk1amRNA, 200 pg; pk1a-MO, 0.5 ng. (H) Injection of human JNK2�2 mRNA could rescue CE defects of mir206-misexpressed embryos. Embryos were injected with8 ng 206-MOs or 200 pg mir206 precursor RNA alone or together with 50 pg human JNK2�2 mRNA at the one-cell stage. The expression of ntl was assayed atthe bud stage. Dorsal views with the anterior at the top.
Liu et al.
2940 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.
expression wholly depending on the identified site. As zebrafish let-7target sites are functional when positioned in the coding sequence oreven in the 5=UTR (31), miRNA target sites in coding sequences maynot be infrequent events during zebrafish development.
Prickle1 is a critical mediator of Wnt/PCP signaling (52), andthere is a strong genetic interaction between pk1a and trilobite/strabismus in the regulation of zebrafish CE (5, 52, 61). Somestudies indicate that Prickle1 binds to Dsh and activates JNK incultured cells (52, 56), but other reports demonstrate that exces-sive Prickle1 blocks the Fz7-dependent membrane localization ofDsh and downregulates levels of Dsh protein by a ubiquitination/degradation system (5, 6, 39, 61). These complex regulation cluesare consistent with the findings that both gain and loss of prickle1function inhibit CE movements in zebrafish and mouse embryos
(5, 39, 61). Our study revealed that overexpression or knockdownof pk1a could results in a decrease of the phosphorylated JNK2 inzebrafish embryos. More importantly, injection of pk1a mRNAinto mir206-overexpressed embryos or blocking pk1a function inmir206 morphants recovered p-JNK2 levels and rescued themir206-mediated CE defects. Our data indicated that mir206 reg-ulates cell movements in both cell-autonomous and non-cell-au-tonomous manners. JNK2 has been implicated to regulate theexpression of transforming growth factor �1 (TGF-�1) in mousefibroblasts, connective tissue growth factor in human lung fibro-blasts, and Wnt11 in zebrafish gastrula cells (47, 59, 62). The non-cell-autonomous activity of mir206 may be mediated by the Pk1a-JNK2 pathway via some as-yet-unidentified secretory moleculesthat are able to non-cell autonomously regulate the migration oradhesion of neighboring cells. In conclusion, our data discoverthat mir206 targets pk1a to modulate JNK2 activation during ze-brafish gastrulation, and this genetic interaction is an importantregulatory ingredient to control cell movements.
ACKNOWLEDGMENTS
This work was supported by grants from Major Science Programs ofChina (grant no. 2011CBA01101, 2011CB943904, and 2011CB9439800),the National Natural Science Foundation of China (30971656 and90919011), and Beijing Municipal Natural Science Foundation(5112020).
We are grateful to members of the Meng lab for assistance and discus-sion.
REFERENCES1. Bakkers J, et al. 2004. Has2 is required upstream of Rac1 to govern dorsal
migration of lateral cells during zebrafish gastrulation. Development 131:525–537.
2. Bartel DP. 2004. MicroRNAs: genomics, biogenesis, mechanism, andfunction. Cell 116:281–297.
3. Birmingham A, et al. 2006. 3= UTR seed matches, but not overall identity,are associated with RNAi off-targets. Nat. Methods 3:199 –204.
4. Boutros M, Paricio N, Strutt DI, Mlodzik M. 1998. Dishevelled activatesJNK and discriminates between JNK pathways in planar polarity andwingless signaling. Cell 94:109 –118.
5. Carreira-Barbosa F, et al. 2003. Prickle 1 regulates cell movements duringgastrulation and neuronal migration in zebrafish. Development 130:4037– 4046.
6. Chan DW, Chan CY, Yam JW, Ching YP, Ng IO. 2006. Prickle-1negatively regulates Wnt/beta-catenin pathway by promoting Dishevelledubiquitination/degradation in liver cancer. Gastroenterology 131:1218 –1227.
7. Chen JF, et al. 2010. microRNA-1 and microRNA-206 regulate skeletalmuscle satellite cell proliferation and differentiation by repressing Pax7. J.Cell Biol. 190:867– 879.
8. Chen PY, et al. 2005. The developmental miRNA profiles of zebrafish asdetermined by small RNA cloning. Genes Dev. 19:1288 –1293.
9. Choi WY, Giraldez AJ, Schier AF. 2007. Target protectors reveal damp-ening and balancing of nodal agonist and antagonist by miR-430. Science318:271–274.
10. Copp AJ, Greene ND, Murdoch JN. 2003. The genetic basis of mamma-lian neurulation. Nat. Rev. Genet. 4:784 –793.
11. Cui J, et al. 2006. c-Jun NH(2)-terminal kinase 2alpha2 promotes thetumorigenicity of human glioblastoma cells. Cancer Res. 66:10024 –10031.
12. Duursma AM, Kedde M, Schrier M, le Sage C, Agami R. 2008. miR-148targets human DNMT3b protein coding region. RNA 14:872– 877.
13. Forman JJ, Legesse-Miller A, Coller HA. 2008. A search for conservedsequences in coding regions reveals that the let-7 microRNA targets Dicerwithin its coding sequence. Proc. Natl. Acad. Sci. U. S. A. 105:14879 –14884.
14. Gerdes JM, et al. 2007. Disruption of the basal body compromises pro-teasomal function and perturbs intracellular Wnt response. Nat. Genet.39:1350 –1360.
FIG 6 Genetic interaction between mir206 and pk1a controls cell movementsduring zebrafish gastrulation. (A, B) pk1a mRNA overexpression repressed themir206-induced decrease of the p-JNK2 level (A), and injection of pk1a-MOsuppressed the loss of JNK2 activity in mir206 morphants (B). Embryos wereinjected with the indicated RNAs or MOs at the one-cell stage and harvested atthe 75%-epiboly stage for Western blots. Total JNK2 was examined as a load-ing control. (C) The expression patterns of myod1 (top) and ntl (bottom) at thetailbud stage were almost rescued to normal by modulating pk1a expression inmir206-overexpressed or -depleted embryos. All embryos were dorsally viewedwith the anterior at the top. (D) Convergence defects in mir206-overexpressedor -depleted embryos were rescued by coinjection of pk1a mRNA or pk1a-MO,respectively. Lateral mesendodermal cells (labeled with rhodamine-dextran)deriving from embryos injected with the indicated RNAs were transplantedinto the corresponding regions of the normal hosts at the shield stage (6 hpf)and observed at 10 hpf. Lateral views with the dorsal side at the right and theanterior at the top. Scale bar: 200 �m. Injection doses in panels A to D: mir206precursor RNA, 200 pg; 206-MOs, 8 ng; pk1a mRNA, 100 pg; pk1a-MO, 0.1 ng.
mir206 Regulates CE Movements during Gastrulation
July 2012 Volume 32 Number 14 mcb.asm.org 2941
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.

15. Giraldez AJ, et al. 2005. MicroRNAs regulate brain morphogenesis inzebrafish. Science 308:833– 838.
16. Goljanek-Whysall K, et al. 2011. MicroRNA regulation of the paired-boxtranscription factor Pax3 confers robustness to developmental timing ofmyogenesis. Proc. Natl. Acad. Sci. U. S. A. 108:11936 –11941.
17. Hafner M, et al. 2010. Transcriptome-wide identification of RNA-binding protein and microRNA target sites by PAR-CLIP. Cell 141:129 –141.
18. Hammerschmidt M, et al. 1996. Mutations affecting morphogenesisduring gastrulation and tail formation in the zebrafish, Danio rerio. De-velopment 123:143–151.
19. Heisenberg CP, et al. 2000. Silberblick/Wnt11 mediates convergent ex-tension movements during zebrafish gastrulation. Nature 405:76 – 81.
20. Huang H, et al. 2007. Amotl2 is essential for cell movements in zebrafishembryo and regulates c-Src translocation. Development 134:979 –988.
21. Jessen JR, et al. 2002. Zebrafish trilobite identifies new roles for Strabis-mus in gastrulation and neuronal movements. Nat. Cell Biol. 4:610 – 615.
22. Keller R. 2002. Shaping the vertebrate body plan by polarized embryoniccell movements. Science 298:1950 –1954.
23. Keller R, Danilchik M. 1988. Regional expression, pattern and timing ofconvergence and extension during gastrulation of Xenopus laevis. Devel-opment 103:193–209.
24. Keller R, et al. 2000. Mechanisms of convergence and extension by cellintercalation. Philos. Trans. R Soc. Lond. B Biol. Sci. 355:897–922.
25. Kilian B, et al. 2003. The role of Ppt/Wnt5 in regulating cell shape andmovement during zebrafish gastrulation. Mech. Dev. 120:467– 476.
26. Kim GH, Han JK. 2005. JNK and ROKalpha function in the noncanonicalWnt/RhoA signaling pathway to regulate Xenopus convergent extensionmovements. Dev. Dyn. 232:958 –968.
27. Kim HK, Lee YS, Sivaprasad U, Malhotra A, Dutta A. 2006. Muscle-specific microRNA miR-206 promotes muscle differentiation. J. Cell Biol.174:677– 687.
28. Kimmel CB, Ballard WW, Kimmel SR, Ullmann B, Schilling TF. 1995.Stages of embryonic development of the zebrafish. Dev. Dyn. 203:253–310.
29. Klein TJ, Mlodzik M. 2005. Planar cell polarization: an emerging modelpoints in the right direction. Annu. Rev. Cell Dev. Biol. 21:155–176.
30. Kloosterman WP, et al. 2006. Cloning and expression of new microRNAsfrom zebrafish. Nucleic Acids Res. 34:2558 –2569.
31. Kloosterman WP, Wienholds E, Ketting RF, Plasterk RH. 2004. Sub-strate requirements for let-7 function in the developing zebrafish embryo.Nucleic Acids Res. 32:6284 – 6291.
32. Leucht C, et al. 2008. MicroRNA-9 directs late organizer activity of themidbrain-hindbrain boundary. Nat. Neurosci. 11:641– 648.
33. Lewis BP, Burge CB, Bartel DP. 2005. Conserved seed pairing, oftenflanked by adenosines, indicates that thousands of human genes are mi-croRNA targets. Cell 120:15–20.
34. Li S, et al. 2011. Rack1 is required for Vangl2 membrane localization andplanar cell polarity signaling while attenuating canonical Wnt activity.Proc. Natl. Acad. Sci. U. S. A. 108:2264 –2269.
35. Liu H, Cao YD, Ye WX, Sun YY. 2011. Effect of microRNA-206 oncytoskeleton remodelling by downregulating Cdc42 in MDA-MB-231cells. Tumori 96:751–755.
36. Liu Z, et al. 2011. Global identification of SMAD2 target genes reveals arole for multiple co-regulatory factors in zebrafish early gastrulas. J. Biol.Chem. 286:28520 –28532.
37. Martello G, et al. 2007. MicroRNA control of nodal signalling. Nature449:183–188.
38. Montcouquiol M, Crenshaw EB III, Kelley MW. 2006. NoncanonicalWnt signaling and neural polarity. Annu. Rev. Neurosci. 29:363–386.
39. Narimatsu M, et al. 2009. Regulation of planar cell polarity by Smurfubiquitin ligases. Cell 137:295–307.
40. Nasevicius A, Ekker SC. 2000. Effective targeted gene ‘knockdown’ inzebrafish. Nat. Genet. 26:216 –220.
41. Nitta RT, Chu AH, Wong AJ. 2008. Constitutive activity of JNK2 alpha2is dependent on a unique mechanism of MAPK activation. J. Biol. Chem.283:34935–34945.
42. Park E, Kim GH, Choi SC, Han JK. 2006. Role of PKA as a negativeregulator of PCP signaling pathway during Xenopus gastrulation move-ments. Dev. Biol. 292:344 –357.
43. Rao PK, Kumar RM, Farkhondeh M, Baskerville S, Lodish HF. 2006.Myogenic factors that regulate expression of muscle-specific microRNAs.Proc. Natl. Acad. Sci. U. S. A. 103:8721– 8726.
44. Reinhart BJ, et al. 2000. The 21-nucleotide let-7 RNA regulates develop-mental timing in Caenorhabditis elegans. Nature 403:901–906.
45. Roszko I, Sawada A, Solnica-Krezel L. 2009. Regulation of convergenceand extension movements during vertebrate gastrulation by the Wnt/PCPpathway. Semin. Cell Dev. Biol. 20:986 –997.
46. Schoenwolf GC, Smith JL. 2000. Gastrulation and early mesodermalpatterning in vertebrates. Methods Mol. Biol. 135:113–125.
47. Seo J, et al. 2010. Negative regulation of wnt11 expression by Jnk signalingduring zebrafish gastrulation. J. Cell Biochem. 110:1022–1037.
48. Sepich DS, et al. 2000. Role of the zebrafish trilobite locus in gastrulationmovements of convergence and extension. Genesis 27:159 –173.
49. Solnica-Krezel L, Stemple DL, Driever W. 1995. Transparent things: cellfates and cell movements during early embryogenesis of zebrafish. Bioes-says 17:931–939.
50. Solnica-Krezel L, et al. 1996. Mutations affecting cell fates and cellularrearrangements during gastrulation in zebrafish. Development123:67– 80.
51. Song G, Zhang Y, Wang L. 2009. MicroRNA-206 targets notch3, acti-vates apoptosis, and inhibits tumor cell migration and focus formation. J.Biol. Chem. 284:31921–31927.
52. Takeuchi M, et al. 2003. The prickle-related gene in vertebrates is essen-tial for gastrulation cell movements. Curr. Biol. 13:674 – 679.
53. Tao H, et al. 2009. Mouse prickle1, the homolog of a PCP gene, is essentialfor epiblast apical-basal polarity. Proc. Natl. Acad. Sci. U. S. A. 106:14426 –14431.
54. Taulli R, et al. 2009. The muscle-specific microRNA miR-206 blockshuman rhabdomyosarcoma growth in xenotransplanted mice by promot-ing myogenic differentiation. J. Clin. Invest. 119:2366 –2378.
55. Topczewski J, et al. 2001. The zebrafish glypican knypek controls cellpolarity during gastrulation movements of convergent extension. Dev.Cell 1:251–264.
56. Tree DR, et al. 2002. Prickle mediates feedback amplification to generateasymmetric planar cell polarity signaling. Cell 109:371–381.
57. Tucker JA, Mintzer KA, Mullins MC. 2008. The BMP signaling gradientpatterns dorsoventral tissues in a temporally progressive manner along theanteroposterior axis. Dev. Cell 14:108 –119.
58. Unterseher F, et al. 2004. Paraxial protocadherin coordinates cell polarityduring convergent extension via Rho A and JNK. EMBO J. 23:3259 –3269.
59. Utsugi M, et al. 2003. c-Jun-NH2-terminal kinase mediates expression ofconnective tissue growth factor induced by transforming growth factor-beta1 in human lung fibroblasts. Am. J. Respir. Cell Mol. Biol. 28:754 –761.
60. van Dongen S, Abreu-Goodger C, Enright AJ. 2008. Detecting mi-croRNA binding and siRNA off-target effects from expression data. Nat.Methods 5:1023–1025.
61. Veeman MT, Slusarski DC, Kaykas A, Louie SH, Moon RT. 2003.Zebrafish prickle, a modulator of noncanonical Wnt/Fz signaling, regu-lates gastrulation movements. Curr. Biol. 13:680 – 685.
62. Ventura JJ, Kennedy NJ, Flavell RA, Davis RJ. 2004. JNK regulatesautocrine expression of TGF-beta1. Mol. Cell 15:269 –278.
63. von der Hardt S, et al. 2007. The Bmp gradient of the zebrafish gastrulaguides migrating lateral cells by regulating cell-cell adhesion. Curr. Biol.17:475– 487.
64. Wang Q, et al. 2008. MicroRNA miR-24 inhibits erythropoiesis by tar-geting activin type I receptor ALK4. Blood 111:588 –595.
65. Wang X, Ling C, Bai Y, Zhao J. 2011. MicroRNA-206 is associated withinvasion and metastasis of lung cancer. Anat. Rec. (Hoboken) 294:88 –92.
66. Warga RM, Kimmel CB. 1990. Cell movements during epiboly and gas-trulation in zebrafish. Development 108:569 –580.
67. Wienholds E, et al. 2005. MicroRNA expression in zebrafish embryonicdevelopment. Science 309:310 –311.
68. Williams AH, et al. 2009. MicroRNA-206 delays ALS progression andpromotes regeneration of neuromuscular synapses in mice. Science 326:1549 –1554.
69. Winbanks CE, et al. 2011. TGF-beta regulates miR-206 and miR-29 tocontrol myogenic differentiation through regulation of HDAC4. J. Biol.Chem. 286:13805–13814.
70. Yamashita S, et al. 2002. Stat3 controls cell movements during zebrafishgastrulation. Dev. Cell 2:363–375.
71. Zhao Y, et al. 2007. Dysregulation of cardiogenesis, cardiac conduction,and cell cycle in mice lacking miRNA-1-2. Cell 129:303–317.
Liu et al.
2942 mcb.asm.org Molecular and Cellular Biology
Dow
nloa
ded
from
http
s://j
ourn
als.
asm
.org
/jour
nal/m
cb o
n 05
Jan
uary
202
2 by
222
.103
.195
.224
.





























