Embed Size (px)
Citation preview

Micropuncture Study of Nephron
Function in the Rhesus Monkey
CLEAVES M. BENNETr, BARRY M. BRENNER, andROBERTW. BERLINER with the technical assistance of JuLHA L. TROY
From the Laboratory of Kidney and Electrolyte Metabolism, NationalHeart Institute, National Institutes of Health, Bethesda, Maryland
A B S T R A C T The function of the proximal anddistal tubule was studied in the rhesus monkeyduring antidiuresis and during the diuresis afterfurosemide administration (during which extracel-lular fluid volume was maintained).
In the proximal tubule, fluid to plasma ratios forsodium, potassium, and osmolality approximatedunity. During antidiuresis, about 30% of the fil-tered water remained at the end of the accessibleportion of this segment (92% of length). Fluidwas hypotonic to plasma throughout the distaltubule. 25%o of the filtered water was present inthe early distal tubule. Small but significant waterreabsorption (about 8% of filtered) occurred in theremainder of this segment. Tubule fluid to plasmapotassium concentration ratios tended to increasealong the distal tubule, and the amount of potas-sium, relative to the amount filtered, increased from13% in the early portion of this segment to 26%r,in the late portion.
After furosemide was administered animals ex-creted about one-third of the filtered sodiumand water. Despite this diuresis, electrolyte andwater reabsorption along the proximal tubuledid not differ from values obtained in control
An abstract of this work appears in J. Clin. Invest.1967. 46: 1035. Complete tables of data from these experi-ments are available from the American Documentation In-stitute, 2000 P Street, N. W., Washington, D. C. 20036.
Address requests for reprints to Dr. C. M. Bennett,Laboratory of Kidney and Electrolyte Metabolism, Na-tional Heart Institute, Bethesda, Md. 20014.
Dr. Brenner is a Postdoctoral Research Fellow, Na-tional Heart Institute.
Received for publication 18 July 1967 and in revisedform 15 September 1967,
animals. Osmolality and sodium concentrationof fluid from the distal tubule approached those ofplasma. 22%o of the filtered sodium (twice the con-trol values) reached the distal tubule, whereas thefraction of filtered water remaining was onlyslightly increased. These findings indicate that,after the administration of this drug, inhibition ofsodium reabsorption occurred in the water-im-permeable segment of the nephron, rather than inthe proximal tubule. After furosemide administra-tion, all tubule fluid to plasma potassium concen-tration ratios in the distal tubule were equal to orgreater than one, suggesting inhibition of activepotassium reabsorption at or prior to this site.
Fluid to plasma bicarbonate concentration ratiosfrom the midportion of the proximal tubule wereconsistently less than one in normal monkeys.After acetazolamide was administered, the bicar-bonate concentration of samples of tubule fluidrecollected from these same sites was the same as,or higher than in plasma. This fact demonstratesthe inhibition of bicarbonate reabsorption in thisportion of the tubule.
INTRODUCTION
It is likely that many of the basic transport proc-esses operative in renal tubule cells are similarin rat, dog, and man. Indeed, micropuncture stud-ies in the dog and rat have revealed that individualnephron function in these species is quite similar(1, 2). Those differences which have been foundare primarily of a quantitative nature and are notnecessarily fundamental. It has been found, how-ever, that the extent of sodium, water, and bi-
The Journal of Clinical Investigation Volume 47 1968 220.3
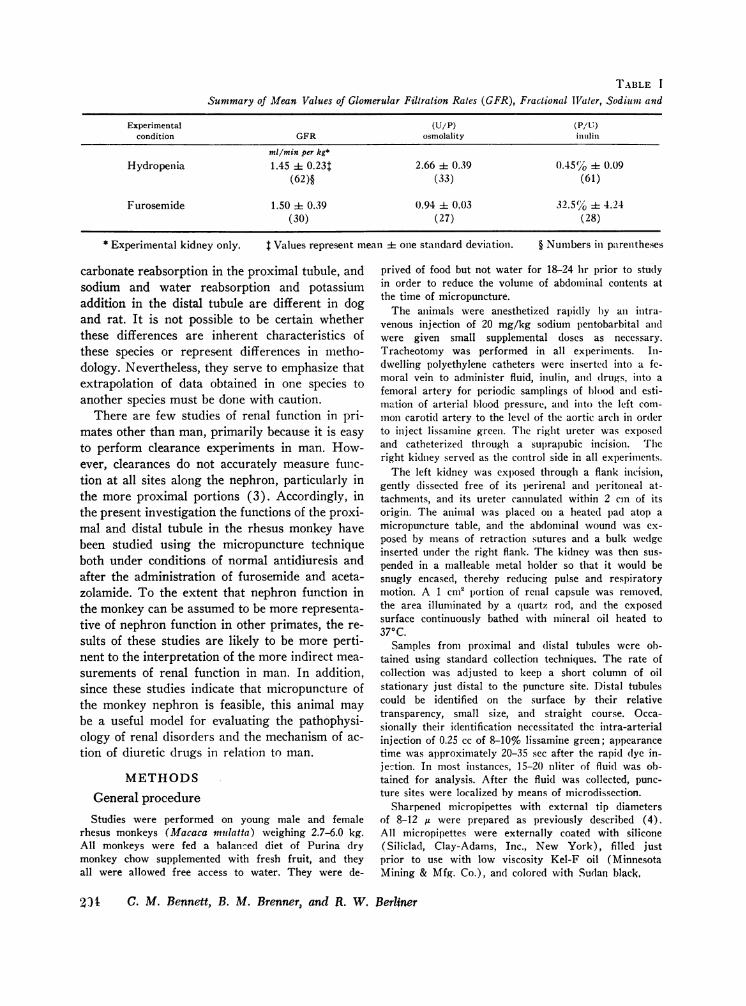
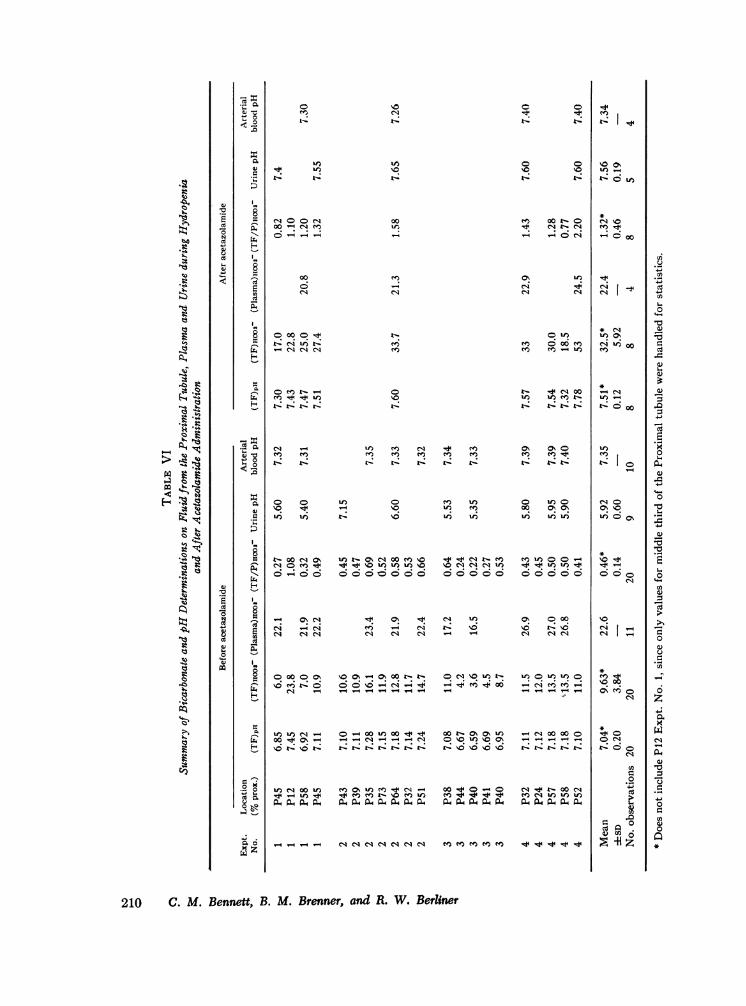
TABLE I
Summary of Mean Values of Glomerular Filtration Rates (GFR), Fractional Tigater, SodiumII and
Experimental (U/P) (Pa/n)condition GFR osmolality intilin
ml/min per kg*
Hydropenia 1.45 ± 0.23t 2.66 + 0.39 0.45(%O 4 0.09(62)§ (33) (61)
Furosemide 1.50 + 0.39 0.94 i 0.03 32.5% + 4.24(30) (27) (28)
* Experimental kidney only. $ Values represent mean + one standard deviation.
carbonate reabsorption in the proximal tubule, andsodium and water reabsorption and potassiumaddition in the distal tubule are different in dogand rat. It is not possible to be certain whetherthese differences are inherent characteristics ofthese species or represent differences in metho-dology. Nevertheless, they serve to emphasize thatextrapolation of data obtained in one species toanother species must be done with caution.
There are few studies of renal function in pri-mates other than man, primarily because it is easyto perform clearance experiments in man. How-ever, clearances do not accurately measure func-tion at all sites along the nephron, particularly inthe more proximal portions (3). Accordingly, inthe present investigation the functions of the proxi-mal and distal tubule in the rhesus monkey havebeen studied using the micropuncture techniqueboth under conditions of normal antidiuresis andafter the administration of furosemide and aceta-zolamide. To the extent that nephron function inthe monkey can be assumed to be more representa-tive of nephron function in other primates, the re-sults of these studies are likely to be more perti-nent to the interpretation of the more indirect mea-surements of renal function in man. In addition,since these studies indicate that micropuncture ofthe monkey nephron is feasible, this animal maybe a useful model for evaluating the pathophysi-ology of renal disorders and the mechanism of ac-tion of diuretic drugs in relation to man.
METHODSGeneral procedure
Studies were performed on young male and femalerhesus monkeys (Macaca niulatta) weighing 2.7-6.0 kg.All monkeys were fed a balanced diet of Purina drymonkey chow supplemented with fresh fruit, and theyall were allowed free access to water. They were de-
§ Numbers in parentheses
prived of food but not water for 18-24 hr prior to studyin order to reduce the volume of abdominal contents atthe time of micropuncture.
The animals were anesthetized rapidly by an intra-venous inj ection of 20 mg/kg sodium pentobarbital andwere given small supplemental doses as necessary.Tracheotomy was performed in all experiments. In-dwelling polyethylene catheters were inserte(l into a fe-moral vein to administer fluid, inulin, and (Irugs, into afemoral artery for periodic samplings of blood an(l esti-mation of arterial blood pressure, and into the left com-mon carotid artery to the level of the aortic arch in orderto inject lissamine green. The right ureter was exposedand catheterized through a suprapubic incision. Theright kidney served as the control side in all experiments.
The left kidney was exposed through a flank incision,gently dissected free of its perirenal andl peritoneal at-tachments, and its ureter cannulated within 2 cm of itsorigin. The animal was placed on a heated pad atop amicropuncture table, and the abdominal wound was ex-posed by means of retraction sutures and a bulk wedgeinserted under the right flank. The kidney was then sus-pended in a malleable metal holder so that it would besnugly encased, thereby reducing pulse and respiratorymotion. A 1 cm2 portion of renal capsule was removed,the area illuminated by a quartz rod, and the exposedsurface continuously bathed with mineral oil heated to370C.
Samples from proximal and distal tubules were oh-tained using standard collection techniques. The rate ofcollection was adjusted to keep a short column of oilstationary just distal to the puncture site. Distal tubulescould be identified on the surface by their relativetransparency, small size, and straight course. Occa-sionally their identification necessitated the intra-arterialinjection of 0.25 cc of 8-10% lissamine green; appearancetime was approximately 20-35 sec after the rapid (lye in-jection. In most instances, 15-20 nliter of fluid was ob-tained for analysis. After the fluid was collected, punc-ture sites were localized by means of microdissection.
Sharpened micropipettes with external tip diametersof 8-12 Ae were prepared as previously described (4).All micropipettes were externally coated with silicone(Siliclad, Clay-Adams, Inc., New York), filled justprior to use with low viscosity Kel-F oil (MinnesotaMining & Mfg. Co.), and colored with Sudan black,
931 C. M. Bennett, B. M. Brenner, and R. W. Berliner
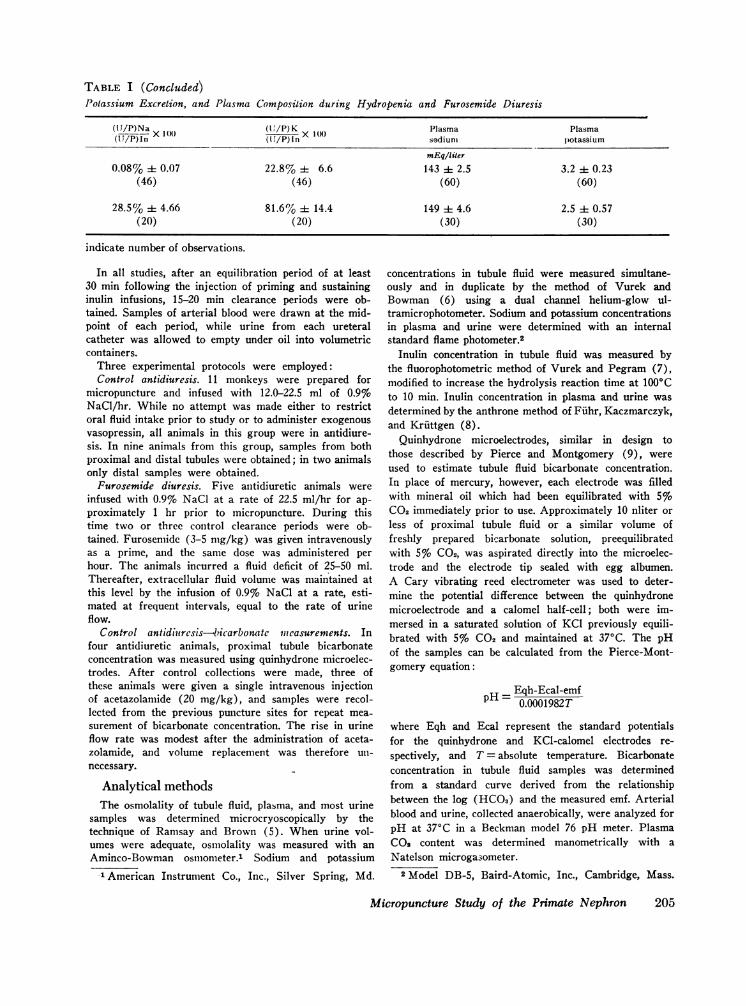
TABLE I (Concluded)Potassium Excretion, and Plasma Composition during Hydropenia and Furosemide Diuresis
(lJ/P)Na X(11PIn X1(
(U'/P) Kn Plasmasadium
Plasmapotassium
mEq/liter0.08% ± 0.07 22.8% ± 6.6 143 ± 2.5 3.2 ± 0.23
(46) (46) (60) (60)
28.5% i 4.66 81.6 % i 14.4 149 ± 4.6 2.5 :1= 0.57(20) (20) (30) (30)
indicate number of observations.
In all studies, after an equilibration period of at least30 min following the injection of priming and sustaininginulin infusions, 15-20 min clearance periods were ob-tained. Samples of arterial blood were drawn at the mid-point of each period, while urine from each ureteralcatheter was allowed to empty under oil into volumetriccontainers.
Three experimental protocols were employed:Control antidiuresis. 11 monkeys were prepared for
micropuncture and infused with 12.0-22.5 ml of 0.9%oNaCl/hr. While no attempt was made either to restrictoral fluid intake prior to study or to administer exogenousvasopressin, all animals in this group were in antidiure-sis. In nine animals from this group, samples from bothproximal and distal tubules were obtained; in two animalsonly distal samples were obtained.
Furosemide diuresis. Five antidiuretic animals wereinfused with 0.9% NaCl at a rate of 22.5 ml/hr for ap-proximately 1 hr prior to micropuncture. During thistime two or three control clearance periods were ob-tained. Furosemide (3-5 nmg/kg) was given intravenouslyas a prime, and the same dose was administered perhour. The animals incurred a fluid deficit of 25-50 ml.Thereafter, extracellular fluid volume was maintained atthis level by the infusion of 0.9% NaCl at a rate, esti-mated at frequent intervals, equal to the rate of urineflow.
Control antidiurcsis-.bicarbonate measurements. Infour antidiuretic animals, proximal tubule bicarbonateconcentration was measured using quinhydrone microelec-trodes. After control collections were made, three ofthese animals were given a single intravenous injectionof acetazolamide (20 mg/kg), and samples were recol-lected from the previous puncture sites for repeat mea-surement of bicarbonate concentration. The rise in urineflow rate was modest after the administration of aceta-zolamide, and volume replacement was therefore un-necessary.
Analytical methodsThe osmolality of tubule fluid, plasma, and most urine
samples was determined microcryoscopically by thetechnique of Ramsay and Brown (5). When urine vol-umes were adequate, osmolality was measured with anAminco-Bowman osmnometer.1 Sodium and potassium
1 American Instrument Co., Inc., Silver Spring, Md.
concentrations in tubule fluid were measured simultane-ously and in duplicate by the method of Vurek andBowman (6) using a dual channel helium-glow ul-tramicrophotometer. Sodium and potassium concentrationsin plasma and urine were determined with an internalstandard flame photometer.2
Inulin concentration in tubule fluid was measured bythe fluorophotometric method of Vurek and Pegram (7),modified to increase the hydrolysis reaction time at 1000Cto 10 min. Inulin concentration in plasma and urine wasdetermined by the anthrone method of Fiuhr, Kaczmarczyk,and Kruttgen (8).
Quinhydrone microelectrodes, similar in design tothose described by Pierce and Montgomery (9), wereused to estimate tubule fluid bicarbonate concentration.In place of mercury, however, each electrode was filledwith mineral oil which had been equilibrated with 5%C02 immediately prior to use. Approximately 10 nliter orless of proximal tubule fluid or a similar volume offreshly prepared bicarbonate solution, preequilibratedwith 5%o C02, was aspirated directly into the microelec-trode and the electrode tip sealed with egg albumen.A Cary vibrating reed electrometer was used to deter-mine the potential difference between the quinhydronemicroelectrode and a calomel half-cell; both were im-mersed in a saturated solution of KC1 previously equili-brated with 5% C02 and maintained at 370C. The pHof the samples can be calculated from the Pierce-Mont-gomery equation:
pH = Eqh-Ecal-emf0.0001982T
where Eqh and Ecal represent the standard potentialsfor the quinhydrone and KCl-calomel electrodes re-spectively, and T = absolute temperature. Bicarbonateconcentration in tubule fluid samples was determinedfrom a standard curve derived from the relationshipbetween the log (HCO3) and the measured emf. Arterialblood and urine, collected anaerobically, were analyzed forpH at 37°C in a Beckman model 76 pH meter. PlasmaC02 content was determined manometrically with aNatelson microgasometer.
2 Model DB-5, Baird-Atomic, Inc., Cambridge, Mass.
Micropuncture Study of the Primate Nephron 205
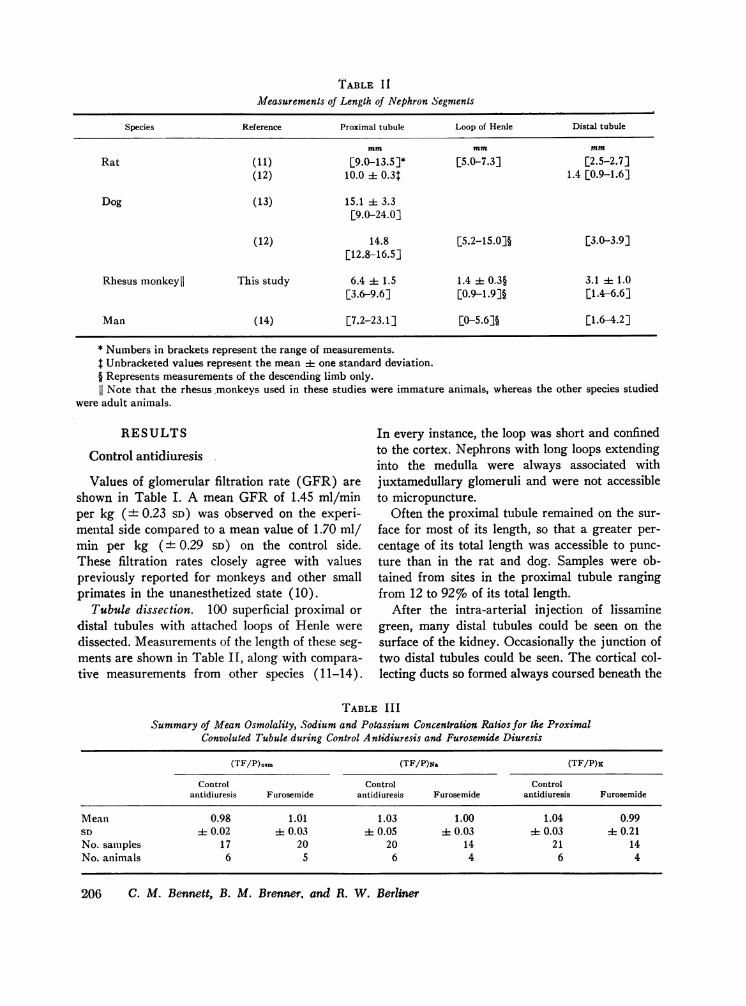
TABLE II
Measurements of Length of Nephron Segments
Species Reference Proximal tubule Loop of Henle Distal tubule
mm mm mm
Rat (I11) [9.0-13.5]* [5.0-7.3] [2.5-2.7](12) 10.0 i: 0.31 1.4 [0.9-1.6]
Dog (13) 15.1 1: 3.3[9.0-24.0]
(12) 14.8 [5.2-15.0]§ [3.0-3.9][12.8-16.5]
Rhesus monkeyll This study 6.4 :1: 1.5 1.4 4 0.3§ 3.1 ± 1.0[3.6-9.6] [0.9-1.9]§ [1.4-6.6]
Man (14) [7.2-23.1] [0-5.6]§ [1.6-4.2]
* Numbers in brackets represent the range of measurements.t Unbracketed values represent the mean 1 one standard deviation.§ Represents measurements of the descending limb only.11 Note that the rhesus monkeys used in these studies were immature animals, whereas the other species studied
were adult animals.
RESULTS
Control antidiuresis
Values of glomerular filtration rate (GFR) areshown in Table I. A mean GFRof 1.45 ml/minper kg (± 0.23 SD) was observed on the experi-mental side compared to a mean value of 1.70 ml/min per kg (± 0.29 SD) on the control side.These filtration rates closely agree with valuespreviously reported for monkeys and other smallprimates in the unanesthetized state (10).
Tubule dissection. 100 superficial proximal ordistal tubules with attached loops of Henle weredissected. Measurements of the length of these seg-ments are shown in Table II, along with compara-tive measurements from other species (11-14).
In every instance, the loop was short and confinedto the cortex. Nephrons with long loops extendinginto the medulla were always associated withjuxtamedullary glomeruli and were not accessibleto micropuncture.
Often the proximal tubule remained on the sur-face for most of its length, so that a greater per-centage of its total length was accessible to punc-ture than in the rat and dog. Samples were ob-tained from sites in the proximal tubule rangingfrom 12 to 92%o of its total length.
After the intra-arterial injection of lissaminegreen, many distal tubules could be seen on thesurface of the kidney. Occasionally the junction oftwo distal tubules could be seen. The cortical col-lecting ducts so formed always coursed beneath the
TABLE IIISummary of Mean Osmolality, Sodium and Potassium Concentration Ratios for the Proximal
Convoluted Tubule during Control Antidiuresis and Furosemide Diuresis
(TF/P)osm (TF/P)N, (TF/P)K
Control Control Controlantidiuresis Furosemide antidiuresis Furosemide antidiuresis Furosemide
Mean 0.98 1.01 1.03 1.00 1.04 0.99SD 4 0.02 4 0.03 ± 0.05 i 0.03 i 0.03 i 0.21No. samples 17 20 20 14 21 14No. animals 6 5 6 4 6 4
206 C. M. Bennett, B. M. Brenner, and R. W. Berliner
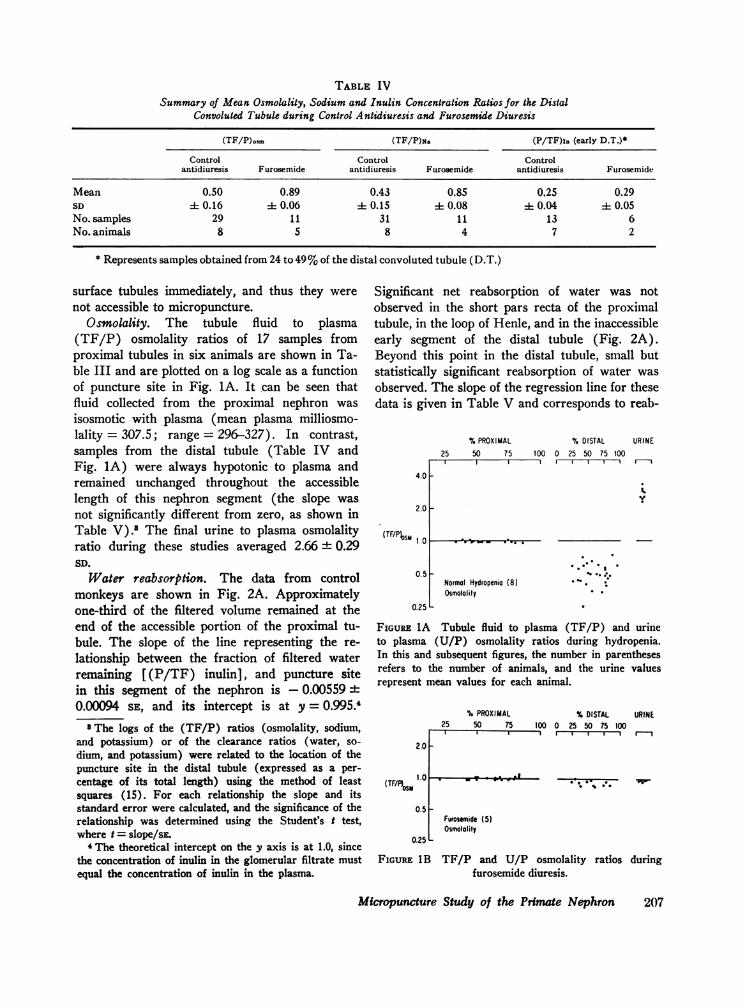
TABLE IVSummary of Mean Osmolality, Sodium and Inulin Concentration Ratios for the Distal
Convoluted Tubule during Control Antidiuresis and Furosemide Diuresis
(TF/P)osm (TF/P)N& (P/TF)ln (early D.T.)*
Control Control Controlantidiuresis Furosemide antidiuresis Furosemide antidiuresis Furosemide
Mean 0.50 0.89 0.43 0.85 0.25 0.29SD 1 0.16 0.06 4 0.15 A 0.08 4 0.04 4 0.05No. samples 29 11 31 11 13 6No. animals 8 5 8 4 7 2
* Represents samples obtained from 24 to 49 %of the distal convoluted tubule (D.T.)
surface tubules immediately, and thus they werenot accessible to micropuncture.
Osmolality. The tubule fluid to plasma(TF/P) osmolality ratios of 17 samples fromproximal tubules in six animals are shown in Ta-ble III and are plotted on a log scale as a functionof puncture site in Fig. 1A. It can be seen thatfluid collected from the proximal nephron wasisosmotic with plasma (mean plasma milliosmo-lality = 307.5; range = 296-327). In contrast,samples from the distal tubule (Table IV andFig. 1A) were always hypotonic to plasma andremained unchanged throughout the accessiblelength of this nephron segment (the slope wasnot significantly different from zero, as shown inTable V).' The final urine to plasma osmolalityratio during these studies averaged 2.66 + 0.29SD.
Water reabsorption. The data from controlmonkeys are shown in Fig. 2A. Approximatelyone-third of the filtered volume remained at theend of the accessible portion of the proximal tu-bule. The slope of the line representing the re-lationship between the fraction of filtered waterremaining [(P/TF) inulin], and puncture sitein this segment of the nephron is - 0.00559 ±0.00094 SE, and its intercept is at y = 0.995.4
S The logs of the (TF/P) ratios (osmolality, sodium,and potassium) or of the clearance ratios (water, so-dium, and potassium) were related to the location of thepuncture site in the distal tubule (expressed as a per-centage of its total length) using the method of leastsquares (15). For each relationship the slope and itsstandard error were calculated, and the significance of therelationship was determined using the Student's t test,where t = slope/sE.
4The theoretical intercept on the y axis is at 1.0, sincethe concentration of inulin in the glomerular filtrate mustequal the concentration of inulin in the plasma.
Significant net reabsorption of water was notobserved in the short pars recta of the proximaltubule, in the loop of Henle, and in the inaccessibleearly segment of the distal tubule (Fig. 2A).Beyond this point in the distal tubule, small butstatistically significant reabsorption of water wasobserved. The slope of the regression line for thesedata is given in Table V and corresponds to reab-
4.0
2.0
(TF/Psm 1 .0
0.5
%PROXIMAL25 50 75 100
"0VWW .... .~~~~~~~~~~~~
% DISTAL URINE0 25 50 75 100
mi , mI I r--l~~S
Normal Hydropenia (81Osmololily
FIGURE 1A Tubule fluid to plasma (TF/P) and urineto plasma (U/P) osmolality ratios during hydropenia.In this and subsequent figures, the number in parenthesesrefers to the number of animals, and the urine valuesrepresent mean values for each animal.
2.0
(TF/FosI
0.5 F
0.25 L
% PROXIMAL % DISTAL URINE25 50 75 100 0 25 50 75 100I T---.m-II --,r--l m--
| - | 1I 1 - , |
Furosemide (5)Osmololity
FIGURE IB TF/P and U/P osmolality ratios duringfurosemide diuresis.
Micropuncture Study of the Primate Nephron
0.25
207

0.50
0.25
0.10
(P/ TFOIn 0.05
0.025
%PROXIMAL25 50 75
* .8
0.0101_
Normal Hydropenio (11Inulin
0.0oo L
FIGURE 2A Plasma to tubuleto urine (P/U) inulin concerdropenia. The lines represent tition of filtered water remainithe puncture site.
% PROXIMAL25 50 75
(PITF)1 0.5 *0.25-
Furosemide (5)0.10 L Inulin
FIGURE 2B P/TF and P/Uduring furosemide diuresis. INdistal tubule because data froitions of this segment wereanimals.
sorption of approximatelyfiltrate, or about one-thirdto the earliest accessible pThe 95%o confidence limitline indicate that betweentrate was reabsorbed. It cthat nearly all of the remaiivolume that enters the csorbed.
Sodium reabsorption.ment of (TF/P) sodium20 proximal samples andeight animals are shownand in the upper portionseen that the TF/P ratiosapproximated unity. In ccbule, fluid to plasma concowell below unity. The slopfor the relationship betwethe puncture site is not sig
%DISTAL URINE zero, indicating no change in this variable along100 0 25 50 75 100 the distal tubule (Table V). Assuming that chlo-
ride is the predominant accompanying anion, so-dium chloride accounted for about 75 o of theosmolality.
The lower portion of Fig. 3A, which illustratesfractional 5 sodium reabsorption, shows that ap-proximately 35%o of the filtered sodium is presentat the end of the proximal tubule. Approximately
I) 10io of filtered sodium reached the earliest acces-| sible portion of the distal tubule. This value is less
than half of the fraction of filtered water remainingfluid (P/TF) and plasma at the same site, which indicates that sodium was
ntration ratios during hy- reabsorbed in hypertonic proportions by the rela-he slopes relating the frac- tively water-impermeable ascending limb. Signifi-ng [ (P/TF) Inulin], and cant net reabsorption of sodium along the re-
mainder of the distal tubule could not be detected(0.1 < P < 0.2), as shown in Table V. Nearly
% DISTAL URINE all of the remaining sodium was reabsorbed in the100 0 25 50 75 100collecting duct system since fractional sodium ex-
cretion was extremely low in these studies (less.. . than 0.1 %).
Potassium reabsorption. Data obtained from21 proximal and 30 distal tubules during anti-
inulin concentration ratios diuresis in eight animals are shown in Fig. 4A andlo slope is given for the Table III. Inspection of the (TF/P)K ratios inm the early and late por- the upper portion of Fig. 4A reveals values ap-
obtained from different proximating unity in the proximal tubule. Con-
siderable scatter is evident in the potassium con-
centration ratios in the distal tubule (range = 0.26-of thevolumedelivered 3.43), but the slope of the line indicates a sig-of the volume gme nificant rise along the tubule (Table V). When
fortion slopeof thissegm these ratios are factored by simultaneously deter-s for the slope of this mined TF/P inulin concentration ratios," net addi-
1 and 13%o. of t~he fil- tion of potassium (0.13 rising to 0.26) is demon-:an be seen i Fig. 2A strated (Fig. 4A, lower portion, and Table V).ning 20%o of the filtered Although the (U/P)K ratio (Fig. 4A) rises to)llecting ducts is reab- quite high levels, the ratio [(U/P)K/(U/P)nl]The results of measure- 5The TF/P or U/P ratio for sodium, factored by theconcentration ratios in corresponding TF/P inulin ratio, gives the fraction of31 distal samples from filtered sodium remaining at the puncture site (or final
in Tables III and IV urinS6Since potassium is not only reabsorbed, but also addedof Fig. 3A. It can be to the tubule fluid, the TF/P (or U/P) ratio for this ion,
I in the proximal tubule factored by the simultaneously determined TF/P inulincntrast, in the distal tu- ratio, is defined as the amount of potassium relative to themntration ratios were all amount filtered remaining at the puncture site. This re-
-. . lationship is intended merely to provide a convenientMe of the regression line means of evaluating the fate of potassium in the tubule
!en the (TF/P)NNa and by taking into account the simultaneous reabsorption ofnificantly different from water.
208 C. M. Bennett, B. M. Brenner, and R. W. Berliner

TABLE VSlopes of the Regression Lines Calculated from Data Obtained from the Distal Tubule of Hydropenic Monkeys*
Fraction of Fraction ofIranstubular Fraction of rranstubtflar filtered sodium Transtubular filtered potas-
osmolality filtered watcr Na conce. remaining K concen. sium remainingratio remaining ratio (TF/P)N. ratio (TF/P)K
y = (TF/P).om y = (P/TF)in y = (TF/P)N. (TF/P)in y = (TF/P)K (TF/P)In
Slope 0.00070 -0.00249 0.00048 -0.00332 0.0066 0.00485SE ±0.00102 40.00095 ±0.0017 ±0.00225 ±0.0025 ±0.00219No. observations 28 33 30 28 30 28P value 0.5 <0.025 >0.5 0.1 < P < 0.2 <0.025 <0.05
* These slopes are calculated from the relationship log y = a + b x where y represents the value for each headingdefined above, and x represents the puncture site expressed as per cent of the total 'length of the distal convolutedtubule. A P value of <0.05, by convention, indicates that the relationship is a significant one (i.e., there is greater thana 95% chance that the slope is different from zero).
%PROXIMAL25 50 75
1.00 de % . . goe ,
0.50 _
0.25 -
0.10 _0.05 _
1.00 1
0.50
0.25
(TF/P)14e 0.10
(TF/P)10 0.05
0.01 _
0.0 025 _
% DISTAL URINE100 0 25 50 75 100
3 .
** I.
(.011)
. I
0
**. *0
Normal Hydropenia (8)Sodium
FIGURE 3A Tubule fluid to plasma and urineto plasma sodium concentration ratios (up-per) and sodium clearance ratios (lower)
* .0025 during hydropenia.
(Table I) does not increase above the mean valueof this ratio in late distal tubule. To the extent thatthese surface convolutions of the distal tubule can
be assumed to be representative of all distal tubulesin the kidney, these data indicate that net potas-sium addition in the collecting ducts did not occur.
Furosemide
After the administration of this diuretic, urineflow increased promptly and in all instancesreached maximal levels within 15 min. Glomerularfiltration rates (Table I) were similar to thoseobtained during control studies in hydropenicanimals. Mean values for fractional water, sodium,
(TF/P),b
%PROXIMAL %DISTAL URINE25 50 75 100 0 25 50 75 100
1.00 c- . .r
0.50
0.25
0.10
1.001 l
(TF/P) 0.50
(TF/P)I. 0.25
0.10
I.
Furosemide (5)Sodium
A.
FIGURE 3B TF/P and U/P sodium concentration ra-
tios (upper) and sodium clearance ratios (lower) duringfurosemide diuresis.
Micropuncture Study of the Primate Nephron 209
(TF/P)o*.
twc t-: 0
0 U) %) O 0 lf),
oo m 4 t 4 m 1
.t 8 H~~
U) 8: o. X . o..W
0~~~~~~~~~~~~~~~~~~~~~~~~~~0
00 Itt~~~~~~~~~~~~~~
z~~ ~~~U vu e°°° 00U)oU)~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~j
) *-4 0 .-' 0 t-- d4 C1400 C-4-N~~~ '0 U) uo t . u) _ _
N t .Y A CsU) n N + eCh Ut
X C'~ '-4 If) UoI n 00-4~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~-
0 -
0 r. .
'0
¢~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~C1404=
U)-~~~~~~~~~~~~~~~~~~~~~~~~~~~~~~08 C)u00 '0~H~~~~~~~~~~r 0 C1o %o - CK o1w m oC14 I-e ca~e 6B~6 6 66 0 0 oooXoo b o 0
coIdI0%
W) 0 0 '0
U) @ -sOooOoOcOoOOOOOOd o 000 0
0
80o000 '00%0%00Z- 04'0
06~~~~~~~~~~~~~~~~~~~~
Ul)V) C1-4 000U)00'1 d4 00 t-%0Olt '-4 -00 00 ..4 0(z4 oo ~0%' C4 'UC 0\0 v) \00 'h -4v-'- -. 0' C1
C14
w~~~~~ Ct ox C14 0 to m C\ in m C,4 Cq I-
S~~~~~~~~~~~~~~~~~~~~~~~~~~~l Id Id 0 to tn to4-
0 4U
Aauavo) oo'0o sou)c oo
.00d
0C6 0 o o 4 C14 C14 _14 c14 C14 oq m )eo en m _ 444 o.04 cs44 -.14 Z
$ bbbb bboo bbbZ
210 C. M. Bennett, B. M. Brenner, and R. W. Berliner
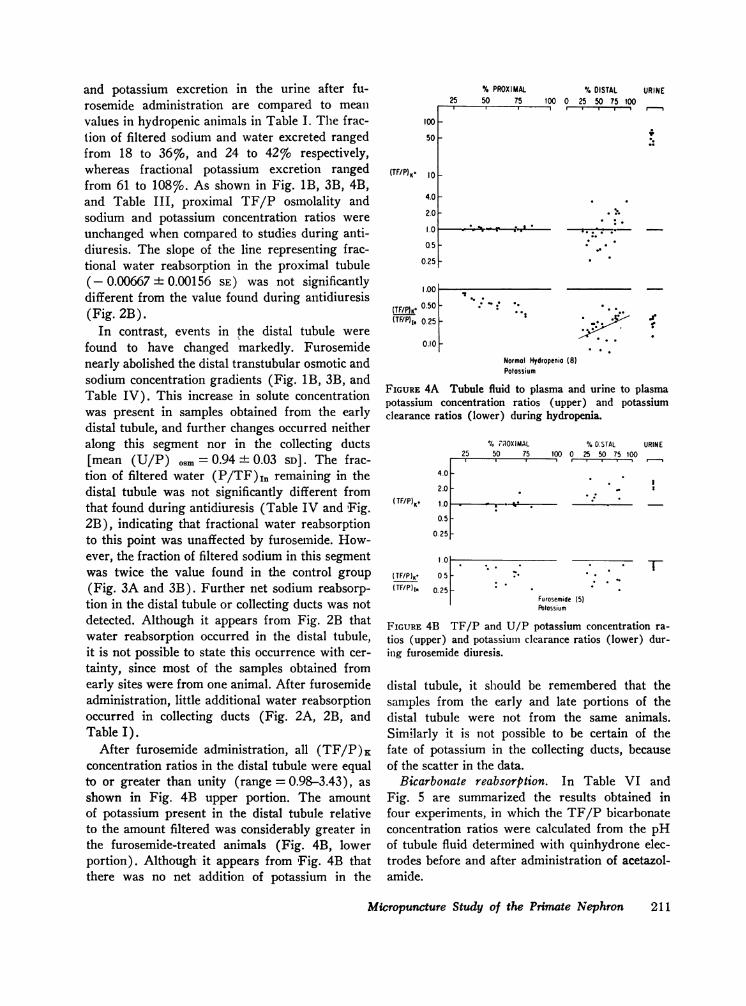
and potassium excretion in the urine after fu-rosemide administration are compared to meanvalues in hydropenic animals in Table I. The frac-tion of filtered sodium and water excreted rangedfrom 18 to 36%o, and 24 to 42%o respectively,whereas fractional potassium excretion rangedfrom 61 to 108%o. As shown in Fig. 1B, 3B, 4B,and Table III, proximal TF/P osmolality andsodium and potassium concentration ratios wereunchanged when compared to studies during anti-diuresis. The slope of the line representing frac-tional water reabsorption in the proximal tubule(- 0.00667 ± 0.00156 SE) was not significantlydifferent from the value found during antidiuresis(Fig. 2B).
In contrast, events in the distal tubule werefound to have changed markedly. Furosemidenearly abolished the distal transtubular osmotic andsodium concentration gradients (Fig. 1B, 3B, andTable IV). This increase in solute concentrationwas present in samples obtained from the earlydistal tubule, and further changes occurred neitheralong this segment nor in the collecting ducts[mean (U/P) osm = 0.94 + 0.03 SD]. The frac-
tion of filtered water (P/TF),1, remaining in thedistal tubule was not significantly different fromthat found during antidiuresis (Table IV and Fig.2B), indicating that fractional water reabsorptionto this point was unaffected by furosemide. How-ever, the fraction of filtered sodium in this segmentwas twice the value found in the control group(Fig. 3A and 3B). Further net sodium reabsorp-tion in the distal tubule or collecting ducts was notdetected. Although it appears from Fig. 2B thatwater reabsorption occurred in the distal tubule,it is not possible to state this occurrence with cer-tainty, since most of the samples obtained fromearly sites were from one animal. After furosemideadministration, little additional water reabsorptionoccurred in collecting ducts (Fig. 2A, 2B, andTable I).
After furosemide administration, all (TF/P)Kconcentration ratios in the distal tubule were equalto or greater than unity (range = 0.98-3.43), asshown in Fig. 4B upper portion. The amountof potassium present in the distal tubule relativeto the amount filtered was considerably greater inthe furosemide-treated animals (Fig. 4B, lowerportion). Although it appears from 'Fig. 4B thatthere was no net addition of potassium in the
(TF/P) re
% PROXIMAL25 50 75
100 _
50 _
10 _
4.0
2.0
1.0 , ,^ r : @.
05
0.25
I.OCl
(TF/P)5* 0.50(TF/P)1, 0.25
0.10 KIs
% DISTAL URINE100 0 25 50 75 100
*
.e,
Normal Hydropenio (8)Potassium
FIGURE 4A Tubule fluid to plasma and urine to plasmapotassium concentration ratios (upper) and potassiumclearance ratios (lower) during hydropenia.
4.0
% PROXIMAL25 50 75
%DOSTAL URINE100 0 25 50 75 100
I r
*
2.0 F
( TF/P) K
TF/P)K,(TF/P)1i
1.0 , ", * i'% .
0.5-
0.25-
1.°1025
0.25
1-
Furosemide (5)POtassium
FIGURE 4B TF/P and U/P potassium concentration ra-tios (upper) and potassium clearance ratios (lower) dur-ing furosemide diuresis.
distal tubule, it should be remembered that thesamples from the early and late portions of thedistal tubule were not from the same animals.Similarly it is not possible to be certain of thefate of potassium in the collecting ducts, becauseof the scatter in the data.
Bicarbonate reabsorption. In Table VI andFig. 5 are summarized the results obtained infour experiments, in which the TF/P bicarbonateconcentration ratios were calculated from the pHof tubule fluid determined with quinhydrone elec-trodes before and after administration of acetazol-amide.
Micropuncture Study of the Primate Nephron
I. . . .-
0
: .ft
211

(TF/P)HCo-
%PROXIMAL TUBULE0 25 50 75 100
4.0 -
20-0
i- 1, 21.0I~~0
0.51-
0.25 F
0 o
* Before Diomoxo After Diomox
Bicorbonote (4)0 125 L
FIGURE 5 Tubule fluid to plasma bicarbonate concen-tration ratios. After diamox, samples of tubule fluid wereobtained by the recollection technique.
Plasma HCO3concentrations and pH were rela-tively stable throughout each experiment andchanged only slightly after acetazolamide adminis-tration. Note that in Experiment 3 the plasmaHCO, concentrations were low, but the (TF/P)]HCO3 ratios were not obviously different fromvalues obtained in-the other experiments. The bi-carbonate concentrations and pH of all samples offluid obtained from the middle third of the prox-imal tubule prior to acetazolamide administrationwere significantly lower than those in plasma.From these data we were unable to detect a pro-gressive decline in (TF/P)Hco3 along the courseof the proximal tubule. However, it can be seenthat the only early proximal sample (P 12%o) hada bicarbonate concentration almost identical withthe concentration in plasma. After an intravenousinjection of acetazolamide, nine recollection sam-ples from three animals revealed a uniform in-crease in (TF/P)ico3. In all but two instances,the bicarbonate concentration in tubule fluid ex-ceeded the concentration in plasma.
DISCUSSION
The study of nephron function in the rhesus mon-key by use of the micropuncture technique offersseveral advantages over similar studies in rodentsand dogs. One important advantage is that in ex-tending these data to an understanding of renalfunction in man, phylogenetic differences are lessformidable. Moreover, the short loop of Henle andthe arrangement of the proximal and distal con-volutions on the surface of the kidney make almost
the entire nephron, up to the collecting duct, ac-cessible to micropuncture. Finally, the monkey ismore suitable than other animals for study of themechanism anid site of action of diuretic drugs,particularly with respect to changes iln function ofthe distal tubule. The rat is much less responsiveto diuretics than man and has been unsatisfactoryto study for this reason.
Fractional water reabsorption in the proximaltubule of the antidiuretic monkey is similar tovalues reported for the dog and rat from this andother laboratories (1, 16, 17). In earlier studiesin rat (18-20) and dog (21, 22), variable butgreater fractional water reabsorption- in the proxi-mal tubule was found than in the monkey. Thisfact may be attributable to differences in thephysiological state of the animals, particularly inthe extracellular fluid volume. The fraction offiltered water arriving in the early distal tubuleof the monkey is similar to the fraction found inthe dog (1). The values reported for the rat (20.23) are somewhat lower than in the dog andmonkey, but again may reflect the physiologicalstate of the animals studied. In the rat, fluid fromithe early distal tubule is hypotonic and duringantidiuresis becomes isotonic to plasma in themidportion (24). During this osmotic equilibra-tion, nearly three-fourths of the volume is reab-sorbed (19, 20). The remaining fraction of fil-tered water (8-10%o) enters the collecting ductswhere it is almost completely reabsorbed (2). Inthe antidiuretic monkey, as in the dog (25), fluidremains hypotonic to plasma throughout the distaltubule. Despite this considerable driving force forpassive water reabsorption, only about one-thirdof the volume is reabsorbed. Thus, as in the dog, alarger fraction of the glomerular filtrate enters thecollecting ducts of the monkey than enters thecollecting ducts of the rat (1).
The reabsorption of sodium in hypertonic pro-portionis in the loop of Henle is evidence that theprimate nephron has the potential to generate the"single effect," the process fundamental to thecountercurrent mechanism for urine concentration(26). However, it should be remembered thatcortical nephrons, such as those studied here, donot contribute to the concentrating mechanism.The fraction of filtered sodium reaching the distaltubule of the monkey is similar to values reportedfor the dog (1). In neither species could further
212 C. M. Bennett, B. M. Brenner, and R. W. Berliner

net sodium reabsorption in the distal tubule bedetected, presumably because of the low permeabil-ity to water and the presence of a limiting concen-tration gradient for sodium (1). As in the rat, thesite of the steepest transtubular concentrationgradient for sodium is in the collecting duct, notthe distal tubule (2).
On the basis of clearance data (27) it has beenshown that in the dog, furosemide administrationin doses of 0.5-5.0 mg/kg impairs free water ex-cretion and reduces free water reabsorption underappropriate experimental conditions. Since theascending limb of Henle's loop is the only, sitein the nephron where sodium reabsorption par-ticipates in the formation of both dilute and con-centrated urine, it has been concluded that theprincipal site of action of this drug is in this seg-ment. In the same study the authors infer from themagnitude of the diuresis (up to 38%o of filteredsodium) that reabsorption in the proximal tubulewas also inhibited. In a micropuncture study inrats, Deetjen (28) found an inhibitory effect offurosemide on reabsorption of fluid in the prox-imal tubule when GFRwas reduced by about 50%,but he could not detect an inhibitory effect whenGFR remained in the normal range. In anothermicropuncture study, Dirks, Cirksena and Berliner(29), using the recollection micropuncture tech-nique, were unable to detect an action of this drugin the proximal tubules in dogs. Rector and hisassociates (16), using both stopped-flow and free-flow micropuncture techniques in the rat, foundthat although furosemide inhibited intrinsic reab-sorptive capacity by about 40%o, the drug had noeffect on fractional reabsorption in the proximaltubule. The failure to detect a decrease in frac-tional reabsorption was thought to be due to a dis-proportionate rise in the volume of the tubule rela-tive to the filtration rate, as evidenced by the pro-longed transit time observed after administrationof the drug. In the present study, furosemide ad-ministration in doses comparable to those used inprevious experiments resulted in the prompt excre-tion of up to 36%o of the filtered sodium and up to42%o of the filtered water. Despite this massivediuresis, fractional reabsorption in the proximaltubule was not depressed. Since the filtration ratewas also unchanged, the absolute rate of sodiumreabsorption in the proximal tubule was similar tocontrol values. The fraction of filtered water reach-
ing the accessible portion of the distal tubule didnot differ from the control value. However, frac-tional delivery of sodium to the distal tubule afterfurosemide administration was twice that found incontrols, indicating that the inhibition of sodiumreabsorption produced by this drug occurred at asite or sites beyond the water permeable portionof the nephron, i.e., beyond the descending limb ofHenle's loop.
The interpretation of the TF/P ratio for potas-sium depends on a consideration of the electricalpotential. A recent study of the electrical potentialin the rat nephron by Frdmter and Hegel led themto the conclusion that there is no electrical poten-tial difference between the proximal tubule lumenand the surrounding interstitial fluid (30). Theirprimary observations were essentially identicalwith those obtained by other workers who havemeasured the electrical potentials in the proximalconvoluted tubules of rats (2, 31, 32) and dogs(33). When the tubule is first punctured a nega-tive deflection is encountered; in a large numberof studies, these negative deflections have alwaysvaried rather widely but have generally averagedclose to 20 mv, tip of the exploring electrode nega-tive. This potential may be maintained for a pe-riod varying from a few seconds to several minutes,generally shorter in the dog than in the rat, andthen drops to zero. Most investigators have as-sumed that the 20 mv value represents the truetranstubular potential, and that the drop to zerois the result of short-circuiting through the punc-ture hole. However, Fr6mter and Hegel found thatthe potential difference remained at zero evenwhen the tip of the exploring electrode was thrustdown the lumen far enough so that any electricalleak at the puncture site should have had littleeffect on the observed transtubular potential. Theyconcluded that the zero value was the true value,and that the substantial negative value initially ob-served is derived from some intracellular location,presumably the brush border.
Since all previous observers have found both the20 mv and the zero potentials, it has seemed to usunprofitable to explore the problem with additionalsimilar measurements. The problem is one ofchoosing which of the two populations representsthe true transtubular electrical potential. In arecent extensive series of measurements in the dog(1) and in the present study in the monkey, the
Micropuncture Study of the Primate Nephron 213

potassium concentration in the lumen of the prox-imal tubule has been found to be very nearly idein-tical with the concentration in plasma, with onlysmall variations in the mean value. This fact leadsus to extend the conclusion of Fromter and Hegelto the dog and monkey, to infer that there is prob-ably no significant transtubular electrical potentialdifference, and further to infer that the reabsorp-tion of potassium in the proximal convoluted tubuleis probably passive, with the potassium ion closeto diffusion equilibrium across the tubule wall.These conclusions derive from the fact that if therewere an electrical potential gradient and if potas-sium were passively distributed, the concentrationratio would not be one. On the other hand, if therewere active transport of potassium, with or with-out an electrical potential difference, the concen-tration ratio of one would not be more likely thanany other value. We therefore believe that theobservations are most easily explained if it is as-sumed that there is neither an electrical gradientnor active transport of potassium in the proximalconvoluted tubule.
The conclusion that potassium reabsorption inthe proximal convoluted tubule is passive is atvariance with the conclusions of several previousstudies in which TF/P potassium concentrationratios significantly above and below one werefound (and in which a significant transtubularelectrical potential was assumed) (31-35). Weareunable to explain the difference between the con-centration ratios found in this study and previousstudies. However, we are confident that the analyt-ical procedure used in the present study is con-siderably more reliable than that used in theearlier study in the dog (34) and probably thatused in most of the others as well.
A (TF/P)K less than unity in the proximaltubule has been found in the presence of a highconcentration of a poorly reabsorbable solute, suchas during the administration of mannitol (2, 35),or during microperfusion of the tubule with iso-
7 This argument does not apply to the concentration ofsodium. The concentration of sodium in the lumen must re-main close to the concentration in plasma so long as so-dium salts constitute all but a minor part of the solutein the lumen and the fluid in the lumen has the sameosmolality as the plasma. Potassium contributes so littleto the osmolality of proximal tubule fluid that its con-centration can vary independently of the osmolality.
tonic raffinose solution (34, 36). However, thosedata do not add to the evidence for an activeprocess for reabsorption of potassium, since thesame investigators found similarly low (TF/P)Kratios in the proximal tubule in the absence of apoorly reabsorbable solute, (i.e., in free-flow col-lections during hydropenia).
About one-eighth of the amount of potassiumfiltered appeared at the earliest accessible portionof the distal tubule of the monkey. It cannot beresolved whether this potassium represents a por-tion of the original amount filtered or potassiumnewly added by the inaccessible loop and earlydistal tubule. Net addition of an amount of potas-sium equal to that initially present in the distaltubule was found in this study; no further additionwas found to occur in the collecting ducts. Similarobservations have been reported in the rat (2). Ifthe electrical potential in the distal tubule of themonkey is negative with respect to peritubularfluids, as it is in the rat, then the observation thatthe (TF/P)K in the early distal tubule is less thanone indicates, by the usual criteria, active reabsorp-tion of this ion at or just before this site. Furose-mide appeared to inhibit this reabsorption. Similarresults have recently been reported for the dog(1).
The reabsorption of bicarbonate in the proximaltubule of the rhesus monkey is nearly complete, afinding similar to that in the dog and rat (21, 37).The pH of tubule fluid in quinhydrone microelec-trodes was measured at a time when the bicar-bonate was in equilibrium with carbon dioxideat a tension approximately equal to that found inextracellular fluids. If a similar equilibrium existsin the lumen of the proximal tubule, as has beensuggested by Rector and his associates (38), thenthe pH recorded in these experiments in vitro(7.04) will be approximately equal to the true pHin vivo. This degree of acidification of proximalfluid is similar in the rat (37), and considerablymore than has been found recently in the dogunder similar experimental conditions (39). Afteradministration of acetazolamide, the bicarbonateconcentration in the tubule fluid rose, often to avalue exceeding the concentration in plasma. Thisindicates that reabsorption of this ion was inhibitedto a greater degree than the reabsorption of sodiumand water.
214 C. M. Bennett, B. M. Brenner, and R. W. Berliner

REFERENCES1. Bennett, C. M., J. R. Clapp, and R. W. Berliner.
1967. Micropuncture study of the proximal and distaltubule in the dog. Am. J. Physiol. 213: 1254.
2. Malnic, G., R. M. Klose, and G. Giebisch. 1966.Micropuncture study of distal tubular potassium andsodium transport in rat nephron. Ant. J. Physiol.211: 529.
3. Orloff, J. 1966. Pitfalls in the use of stop-flow forthe localization of diuretic action, with special refer-ence to Na reabsorption. Ann. N. Y. Acad. Sci. 139:344.
4. Vurek, G. G., C. M. Bennett, R. L. Jamison, and J.L. Troy. 1967. An air-driven micropipette sharpener.J. Appl. Physiol. 22: 191.
5. Ramsay, J. A., and R. H. J. Brown. 1955. Simplifiedapparatus and procedure for freezing point determina-tions upon small volumes of fluid. J. Sci. Instr. 32:372.
6. Vurek, G. G. and R. L. Bowman. 1965. Helium-glowphotometer for picomole analysis of alkali metals.Science. 149: 448.
7. Vurek, G. G., and S. E. Pegram. 1966. Fluorometricmethod for the determination of nanogram quantitiesof inulin. Anal. Biochem. 16: 409.
8. Fuihr, J., J. Kaczmarczyk, and C. D. Krfittgen. 1955.Eine einfache colorimetrische Methode zur Inulinbestimmung fur Nierenclearanceuntersuchungen beiStoffwechselgesunden und Diabetikern. Klin. Woch-schr. 33: 729.
9. Pierce, J. A., and H. Montgomery. 1935. A micro-quinhydrone electrode: its application to the deter-mination of pH of glomerular urine of Necturus.J. Biol. Chem. 110: 763.
10. Sweet, A. Y., M. F. Levitt, and H. L. Hodes. 1961.Kidney function, body fluid compartments, and waterand electrolyte metabolism in the monkey. Am. J.Physiol. 201: 975.
11. Walker, A. M., and J. Oliver. 1941. Methods for thecollection of fluid from single glomeruli and tubulesof the mammalian kidney. AmJ. Physiol. 134: 562.
12. Sperber, I. 1944. Studies on the mammalian kidney.Zool. Bidr. Upps. 22: 249.
13. Watson, J. F. 1966. Potassium reabsorption in theproximal tubule of the dog nephron. J. Clin. Invest.45: 1341.
14. Pai, H. C. 1935. Dissections of nephrons from thehuman kidney. J. Anat. Soc. India. 69: 344.
15. Snedecor, G. W. 1956. Statistical methods applied toexperiments in agriculture and biology. State CollegePress, Ames. 5th edition. 122.
16. Rector, F. C., Jr., J. C. Sellman, M. Martinez-Mal-donado, and D. W. Seldin. 1967. The mechanism ofsuppression of proximal tubular reabsorption bysaline infusions. J. Clin. Invest. 46: 47.
17. Landwehr, D., J. Schnermann, R. M. Klose, and G.Giebisch. 1967. The effect of acute reduction in glo-merular filtration rate on renal tubular sodium andwater reabsorption. Federation Proc. 26: 547. (Abstr.)
18. Glabman, S., H. S. Aynedjian, and N. Bank. 1965.Micropuncture study of the effect of acute reductionsin glomerular filtration rate on sodium and water re-absorption by the proximal tubules of the rat. J. Clin.Invest. 44: 1410.
19. Lassiter, W. E., C. W. Gottschalk, and M. Mylle.1961. Micropuncture study of net tranistubular move-ment of water and urea in nondiuretic mammaliankidney. Am. J. Physiol. 200: 1139.
20. Giebisch, G., R. M. Klose, and E. E. Windhager.1964. Micropuncture study of hypertonic sodiumchloride loading in the rat. Am. J. Physiol. 206: 687.
21. Clapp, J. R., J. F. Watson, and R. W. Berliner. 1963.Osmolality, bicarbonate concentration, and water reab-sorption in proximal tubule of the dog nephron. Am.J. Physiol. 205: 273.
22. Watson, J. F. 1966. Effect of saline loading on sodiumreabsorption in the dog proximal tubule. Am. J.Physiol. 210: 781.
23. Cortney, M. A., M. Mylle, W. E. Lassiter, and C. W.Gottschalk. 1965. Renal tubular transport of water,solute, and PAH in rats loaded with isotonic saline.Am. J. Physiol. 209: 1199.
24. Gottschalk, C. W., and M. Mylle. 1959. Micropuncturestudy of the mammalian urinary concentrating mech-anism: evidence for the countercurrent hypothesis.Ant. J. Physiol. 196: 927.
25. Clapp, J. R., and R. R. Robinson. 1966. Osmolality ofdistal tubular fluid in the dog. J. Clin. Invest. 45: 1847.
26. Gottschalk, C. W. 1964. Osmotic concentration anddilution of the urine. Am. J. Med. 36: 670.
27. Suki, W., F. C. Rector, Jr., and D. W. Seldin. 1965.The site of action of furosemide and other sulfon-amide diuretics in the dog. J. Clin. Invest. 44: 1458.
28. Deetjen, P. 1965. Mikropunktionsuntersuchungen zurWirkung von Furosemid. Arch. Ges. Physiol. 284:184.
29. Dirks, J. H., W. J. Cirksena, and R. W. Berliner.1966. Micropuncture study of the effect of variousdiuretics on sodium reabsorption by the proximaltubules of the dog. J. Clin. Invest. 45: 1875.
30. Fromter, E., and U. Hegel. 1966. Transtubular Poten-tialdifferenzen an proximalen und distalen Tubuli derRattenniere. Arch. Ges. Physiol. 291: 107.
31. Bloomer, H. A., F. C. Rector, Jr., and D. W. Seldin.1963. The mechanism of potassium reabsorption in theproximal tubule of the rat. J. Clin. Invest. 42: 277.
32. Malnic, G., R. M. Klose, and G. Giebisch. 1964. Micro-puncture study of renal potassium excretion in therat. Am. J. Physiol. 206: 674.
33. Watson, J. F., J. R. Clapp, and R. W. Berliner. 1964.Micropuncture study of potassium concentration inproximal tubule of dog, rat and Necturus. J. Clin. In-vest. 43: 595.
34. Marsh, D. J., K. J. Ullrich, and G. Rumrich. 1963.Micropuncture analysis of the behavior of potassiumions in rat renal cortical tubules. Arch. Ges. Physiol.277: 107.
Micropuncture Study of the Primate Nephron 215

35. Rector, F. C., Jr., H. A. Bloomer, and D. W. Seldin.1964. Proximal tubular reabsorption of potassium dur-ing mannitol diuresis in rats. J. Lab. Clin. Med. 63:100.
36. Malnic, G., R. M. Klose, and G. Giebisch. 1966. Micro-perfusion study of distal tubular potassium and sodiumtransfer in rat kidney. Am. J. Physiol. 211: 548.
37. Rector, F. C., Jr., H. A. Bloomer, and D. W. Seldin.1964. Effect of potassium deficiency on the reabsorp-
tion of bicarbonate in the proximal tubule of the ratkidney. J. Clin. Invest. 43: 1976.
38. Rector, F. C., Jr., N. W. Carter, and D. W. Seldin.1965. The mechanism of bicarbonate reabsorption inthe proximal and distal tubules of the kidney. J.CiGn. Invest. 44: 278.
39. Bernstein, B. A., and J. R. Clapp. Micropuncture studyof bicarbonate reabsorption by the dog nephron. Am.J. Physiol. In press.
216 C. M. Bennett, B. M. Brenner, and R. W. Berliner





























