Embed Size (px)
Citation preview

S. Afr. J. Bot. 1999. 65(5 & 6): 361 - 369 361
Micropropagation of members of the Hyacinthaceae with medicinal and ornamental potential- A review
S.A. McCartan and J. van Staden* Research Centre for Plant Growth and Development, School of Botany and Zoology, Un1vers1ty of Natal P1etermaritzburg,
Private Bag X01 , Scottsville, 3209 Republic of South Africa
Recetved 5 May I')')!); revtsed I September 19!.19
The Hyac1nthaceae comprises many genera, which are characterized by bulbs, thick roots, basal leaves and simple or more rarely branched racemes. These genera are widely exploited for their medicinal, pharmaceutical and ornamental potential. In South Africa, these plants are harvested without permits from wild populations, processed and then sold as traditional medicine. This is having a negative impact on wild populations. Although the active ingredients of many
of the bulbs have not been identified, several bufadienolides such as proscillaren A have been isolated from members of this family. These bufadienolides have pharmaceutical potential as cardiotonics. Thus, many members of the Hyacmthaceae, have potential as alternative horticultural/agricultural bulb and/or flower crops for medicinal, pharmaceutical and ornamental purposes. Conventional propagation of these plants, however, is usually fairly slow. M1cropropagalion provides a rapid means to propagate selected chemotypes or cultivars, servmg both conservation and commercial interests. Many members of Hyacinthaceae have been micropropagated This review summarizes th1s information, highlighting the potent1al and problems surrounding this family of flowering plants.
Keywords: adventitious shoots, axillary shoots, bufadienolides, conservation, in vitro propagation, somatic embryos, tissue culture.
Abbreviations: 2,4-D = 2,4-dichlorophenoxyacetic acid; AC = activated charcoal; 85 = Gamborg medium: BA = benzyladenine, BDS = modified 85 medium: CH = casein hydrosyla te: CM = coconut m1lk; IM = indole-3-acetiC ac1d:
IBA = indole-3-butyric acid; iP = 6-[y,y-dimethylallylamino]purine: kinetin = N6-furfuryladenine; MS = Murashige & Skoog medium: NM = •x-napthyaleneacetic acid; PAC = paclobutrazol; TDZ = thidiazuron: YE = yeast extract; (3G]BA, [7G]BA, (9G]BA = 3,7,9 glucosides of benzyladenine.
• To whom correspondence should be addressed.
Introduction The Asparngales forms a large and fairly homogenous complex of fami lies. It is believed to be a monophy letic group, which evolved in parallel with the Li liales and Dioscoralcs. The Asparagales consists of 31 fami l ies including several new families. These fami lies. which include the All iaceae, Asparagaceae, Asphodelacene, Dracennaceae, Eriospermaceae and Hyacinthaceae. compri se genera previously placed wi thin the Liliaceae Wll.\'11 fato (Dahlgren et a/. 1985; Perry 1985). The Hyacinthaccae comprises several genera including Bo111iea. F.u~·omis, Lachenalia, Ledebouria, Scilla and l/rg inea. These genera arc charncterised by bulbs with thick and sometimes contractile roots. basal leaves and simple or more rarely branched racemes. The inflorescences comprise actinomorphic, bisexual tlowers, which range from white, blue, violet, and more rarely yellow, red, brown to nearly black. T he Hyac inthaceae are w idely distributed, occurring in southern Africa and in a region from tht: Mediterranean to SW As ia (Dahlgren et af. 1985; Duncan & du Pless is 1989).
Medicinal and pharmaceutical potential
In South A fr ica, approximately 20 000 tons of plant material , worth about R270 million, is harvested, processed and sold as traditional medicine annually (Gosling 1998}. Approximately 14% of this plant material comprises bulbs (Mander 1997}, which are destruct ive ly harvested, processed and sold for the treatment of var ious ai lments (Table I ). These bulbs, which are sold whole, sliced or chopped, are usually administered as decoctions, emetics and enemas (Watt & Breyer-Brandwij k 1962; Jacot Gui llamod 1971; Hutchings 1989; Mander et af. 1995; van
Wyk eta/ 1997). $'cilia natalensis is one of the top-ten most popular medicinal
plants in South Africa (Cunningham 1988; Williams 1996; Mander 1997). Despite its specially protected conservation status. approximately 95 tons of these bulbs are sold illegally in Durban annually. The price of these bulbs is relatively low ranging from R 1.89 to R6.80 per kilogram. This is mainly due to the occurrence of large populations of these bulbs locally. T he price of another popular medicinal plant, Bowiea volubilis is substantially higher ranging from R 11.74 to R27.80 per kilogram (Mander 1997). The price of these bulbs has increased steadily wi th the decline in their avail abi lity and size (Cunningham 1988).
A lthough the active ingredients of many of these bulbs have not yet been identified. several bufadienol ides have been iso lated from members of the Hyacinthaceae including Bml'iea I'Oiubilis (Jha 1988; Finnie et a/. 1994), Drimia rohu.1·ta (van Wyk eta/ 1997; Luyt et a/. 1999}, Urginea a/tissima (Hutchings et a/. 1996), U. indica (Jha & Sen 198 1, 1983; Jha 1988) and (} maritima (Shyr & Staba 1976; Verbiscar eta/. 1986; Gentry el a/ 1987; Jha 1988). Bufadienol ides such as proscillaridin A are valuable since they can be administered to digital in sensitive/intolerant patients (Jha 1988). These bufadienol idcs. however. are usually not used as cardiotonics due to their low therapeutic indices (Jager & van Staden 1995). However, some bufadienolides identified in certain organs of Bowiea l'oluhi/is are thirty to sixty ti mes more active than those fi·om Digitalis (Hutchings et a/ 1996). Proscillaridin A, which is derived by enzyme hydrolys is from sci llaren A , is produced under several trade names including Caradin®, Cardion®, Proscillan®, Sandoscill®, Scillacrist®, Talusin® and Urgilan® (Budavari 1996). T hese cardiac

362 S. Afr. J. Bot. 1999, 65(5 & 6)
Table 1 Medicinal uses of members of the Hyacinthaceae in South Africa
Plant namc1
Alhuw mnadensis (L) Leighton syn. A. JllllfOI"
Medicinal uses
As an anthelmintic, thirst quencher anc.l to treat venereal c.liseases
References
Watt & Brcyer-Brandwijk I 962; Hutchings 1989
Albuca coopen Bak. As an anthelmintic, lotion for washing wounds and Watt & Breyer-Brandwijk 1962; llutchings to treat venereal diseases 1989
To treat illnesses caused by poisons Hutchings eta/ 1996 .·1/huca fusugaw ( L.f.) Dryanc.l.
Alhuca selosa Jacq. As an anthelmintic, lotion for washing wounds in Jacot Guillamnd 1971; Hutchings 19R9 animals and to tn:at venereal diseases
A/hum slwH"ii Bak. syn . ..1. trichophylla
Bml"iea m/uhilis Harv.
/Jtpcatli hreviofo/ium (Thunb.) Fnurc.
As an anthelmintic and to treat constipation and gonorrhoea
To treat c.lropsy. barrenness in women and headaches
To treat heart pains and breathlessness
fJtpcatlt gmcillimum Bak. syn. /J. po~vphyf- To treat gonorrhoea and pimples fum
Oipcaclt \'ll"tde (L.) Moench
!Jrimia ctliaris Jacq.
f)rimia data Jacq.
/)nmiallem!formi.l· Bak.
f)rimia mhusta Oak. syn. D. alw
Ez""OIII/.1"1111111111/Utlis (Mill.) Chill.
f:"ummts hicolor Bak.
To treat gonorrhoea and flatulence
As an emetic. expectorant and diuretic
To treat high blond pressure and stabbing pains
To treat extemal tumours that have been lanced
As an expectorant, emetic and enema to treat feverish colds
To treat co lic. llatulence, syphilis and hangovers
For colic and as a purgative
f:'ucomts mmo.w (Houtl.) var. comosa syn E. Tn treat rheumatism and teething infants JIIIIIC/(1/(1
t:ucomts JIO/eevansii N .E. Br.
f:"ucoll/1.1" regia (L.) L 'Heril
f.l!clehrwria cooperi (I look. I:) Jessop syn. Scilla cooperi. S. immdensis. S. satum
f.edehmll·w ovatifolia (Oak.) Jessop syn. Scilla m·wifolia
l.edehouno rewJ!wa (I.. f.) Jessop syn. Scilla
faiiCI!III!(Ofill
.\/a.uonut echinata L.f. syn M . bowker1
Ormtlwgalum longl/Jractealllm Jacq.
For people sut1'ering from mental disease
To treat venereal disease, lumbago, diarrhoea, respiratory conditions especially cough nnd biliousness and to prevent premature childbirth
Used in the initiation ceremony of boys; to treat gastro-intestinal, gynaecological and psychological ailments
To treat gastro-intestinal and gynaecological ailments, backache and inllucnza
For treating lumbago and gall sickness in animals; to bathe skin eruptions anti as an ointment for wounds and sores
ror ophthalmic applications. sterility and toothache
To treat swellings or growths
Ormtlwgalum duhiwn Houtt. syn 0. mima- As an anthelmintic ll/111
Ormtlwgalum lr!lllljolium De Iaroche subsp. As a rat poison tenui(ofiunz syn. 0. eckfonii & 0. viren.1·
Ormthoj<alum thyrsoides .J acq. To treat diabetes mellitus
Schiznbasis intricala To treat chest complaints
Walt & Breycr-Brandwijk 1962; Jacot Guillamnd 1971; Hutchings 1989
Walt & 13rcycr-l3rantlwijk 1962; Batten & Bokclmann 1966; Hutchings 1989; Hutchings eta/ 1996
11utchings 1989; Hutchings et al 1996
Watt & 13rcycr-13randwijk 1962
Watt & Breycr-Brandwijk 1962; l3attcn & Bokelmann 1966: Hutchings 1989; llutchings et a/ 1 996
Walt & Breyer-Brandwijk 1962
Hutchings el af 1996
Watt & Breyer-Brandwijk 1962; Jacot Guillamod 1971
Hutchings 1989; Hutchings et af 1996
Hutchings et af 1996
Watt & Breyer-Brandwijk 1962; Batten & 13okclmann 1966; Hutchings et af 1996
Walt & Brcyer-Brandwijk 1962; Batten & l3okclmann 1 966; llutchings 1989: Hutchings eta/ 1996
Wall & Brcycr-Brandwijk 1962
Watt & l3rcycr-Brandwijk 1962
Watt & Brcyer-Brandwijk 1962; Hallen & !3okclmann I 966: Hutchings 1989: Hutchings et a/ 1996
llutchings 1989; Hutchings et of 1996
Watt & Brcyer-Brandwijk 1962: Jacot Guillamod 197 1
Walt & Breyer-Brandwijk 1962; Jacot Guilh:mod 1971
Hutch ings 1989: Hutchings et a/ 1996
BaHcn & Bokelmann 1966
Watt & Brcyer-Brandwijk 1962: Jacot Guillamod 197 1
Watt & l3reyer-13randwijk 1962
Hutchings et of 1996
Scillallatafellsis Planch. syn. S. kraussii & S. As a purgative anti to treat sprains, fractures. boils. Hutchings cl a/ 1996 dracomo11tana vcldsorcs and lung sickness in cattle
Set! fa nermsa (Burch. ) Jessop syn. S. rigidi- To treat fevers, gastro-intestinal ailments and folia insanity
Bryant 1966; Hutchings I 989; Hutchings et a/ 1996

S. Ati". J. Bot. 1999, 65(5 & 6) 363
Table 1 Continued
/ .'rgineaaltJssima (I.. f.) Bak. To treat rheumatic swellings, gastro-intestinal and Watt & 13rcycr-Brandwijk 1962; Hutchings 1989; Hutchings eta/ 1996 respiratory ailments
f irginea e/11gea R.A. Dyer As a treatment f()r colds anti headaches Wall & Rrcycr-Brantlwijk 1962
l :rginea mciC-roce/ltra Bak. As an anthelmintic Hutchings eta! 1996
I "rgmea phy.mdes (Jacq.) Bak. To treat gynaecological ailments and to facilitate delivery during birth
Hutchings I 989: llutchings eta/ 1996
l ."rgmea rubella Bak. For the treatment of colic Watt & Brc)cr-Brandwijk 1962: Jacot Cluillamotll971
l r~;mea .wmgumea Schinz. syn. li. burkei As an abortifacient and lo treat paralysis, circulatory diseases and rheumatism pains
Watt & 13rcycr-Brantlwijk 1962
' Plant name and synonyms (Amok! & De Wet 1993).
glycosides increase the force of the systolic contraction, inhibit lhe atrioventricular conduction, and enhance the 'automacy' of the cardiac tissue and. therefore, are used to treat heart fai lure and certain arrhythmias (Walton eta/. 1994).
13ufadienolides such as sci ll iroside and scillaren A also are effective rodenticides with the French Pharmacopoeia Codex describing a raticidal paste composed of powdered bulb of Red Squill ( LlrKinea maritima), fl our and sugar (Balbaa eta/. 1979). The erticicncy of these strongly emetic rodenticides is largely due to the inability of rodents to regurgitate. This renders these products extremely safe and specific since humans and domestic animals including chickens, cats and dogs readily regurgitate accidentally-i ngested baits (Crabtree 1947). In the 1940's, several tons of powdered bulb of Red Squill ( Urginea maritima) were imported into the USA. This ceased with the introduction of warfarin and other anticoagulant-type rodenticides. The development of warfarin-resistance in rodents, however, has led to renewed interest in Red Squill-formulated products as co-rodenticides ( V crbiscar eta/. 1986). This also has led to the identification and quantification of bufadienolides in various clones or cytotypes of Indian Squill (U indica) (Jha & Sen 198 1, 1983) and Red Squill ( li. maritima) (Gentry et a/ 1987) to select high-yielding strains suitable for commercial exploitation.
Ornamental potential Approximately 16 000 ha are allotted to ornamental bulb production in the Netherlands, representing 55% of the world 's total production area, with significantly smaller areas allotted to ornamental bulb production in the USA (4 449 ha), the UK (4 300 ha). Japan ( I 622 ha), France ( 1 285 ha) and South Africa (425 hnl (De Hertogh & Le Nard 1993a). Despite the ornamental potential of the Hyacinthaceae. the area allotted to the production of /~un1111is (2 ha), Galtonia (I ha), 1/yacint/ws (955 ha), Ornitlwga/um (50 ha), Scilla (20 ha) and Urginea (3.2 ha) is small (De He11ogh & LeNard 1993 a, b; Le Nard & De Hertogh 1993 ), representing approximately 3% of the world 's total ornamental bulb production area. Many of these genera including Hyacinthus. Lachenalia and Scilla are worthwhile pot plants, while others including Eucomis. Ornitlwgalum and Urginea are useful garden plants (De Hertogh & Le Nard 1993b; Le Nard & De Hertogh 1993 ). A breeding programme for Lachenalia comprising five phases including the establishment of a genebank, and the development and subsequent evaluation of hybrids for commercialization has been established over the past thirty years (Nicderwieser eta/. 1998 ). The successful introduction of these hybrids onto the international flower market could initiate other breeding programmes, increasing the popularity of these and other plants.
Conventional propagation Several members of the Hyacinthaceae, therefore, have potential as alternative horticultural/agricultural crops, providing crops of both bulbs and inflorescences. Seed, however, is seldom used for commercial propagation, except where the species or cultivars produce large quantities of reliable seed with short juvenile periods (Rees 1992). Some members of the Hyacinthaceae including Scilla siberica, S. siberica cv • Alba', and S. bifolia, however, are only propagated by seed, while others including S. siberica cv 'Spring Beauty' and S. tubergeniana [= S mischtschenkoana (Bryan 1989)] are propagated vegetatively (Le Nard & De Hertagh 1993). Although offsets are used to propagate many members of the Hyacinthaceae, this is generally too slow for commercial propagation. Several artificial techniques, which include scaling, basal cuttage (scooping and scoring) and bulb cuttings, are used to increase the rate of natural multiplication. These, however, are usual ly restricted by the size of the mother bulb.
Micropropagation Several members of the Hyacinthaceae have been micropropagated using various techniques. These include the proliferation of axillary shoots, the ini tiation of adventitious shoots and the induction of somatic embryos, which are discussed in greater detail (Table 2). Some details, however, such as the frequency of shoot initiation, the average number of shoots initiated, the frequency of fasciated, deformed or mutated shoots, the frequency of plantlets successfully acclimatized, and the propagation potential within a specified time-frame are seldom reported. These details, which should be included routinely in micropropagation reports, are essential for commercial fac ilities.
Axillary shoots In Eucomis. axi llary shoots were proliferated from twin-scale explants, which were cultured on medium containing combinations of BA and NAA. These shoots, which were then sub-cultured onto the same medium, proliferated more shoots resulting in a continuous culture system. After 8- 10 weeks, the shoots were then rooted on medium with or without NAA (Ault 1995b).
Although axillary shoots are genetically-stable, the limited availability of these axillary shoots restricts the potential of this technique (Hussey 1980). Furthermore, 'mixed' cultures comprised of axillary and adventitious shoots do occur occasionally (Hussey 1976a). The origin of these axillary shoots is seldom confirmed, and thus, the frequency of 'mixed cultures' is rarely reported.
Direct adventitious shoots In several members of the Hyacinthaceae, adventitious shoots were initiated directly on, or along the periphery of, various
364 S. Afr. J. Bot. !999. 65(5 & 6)
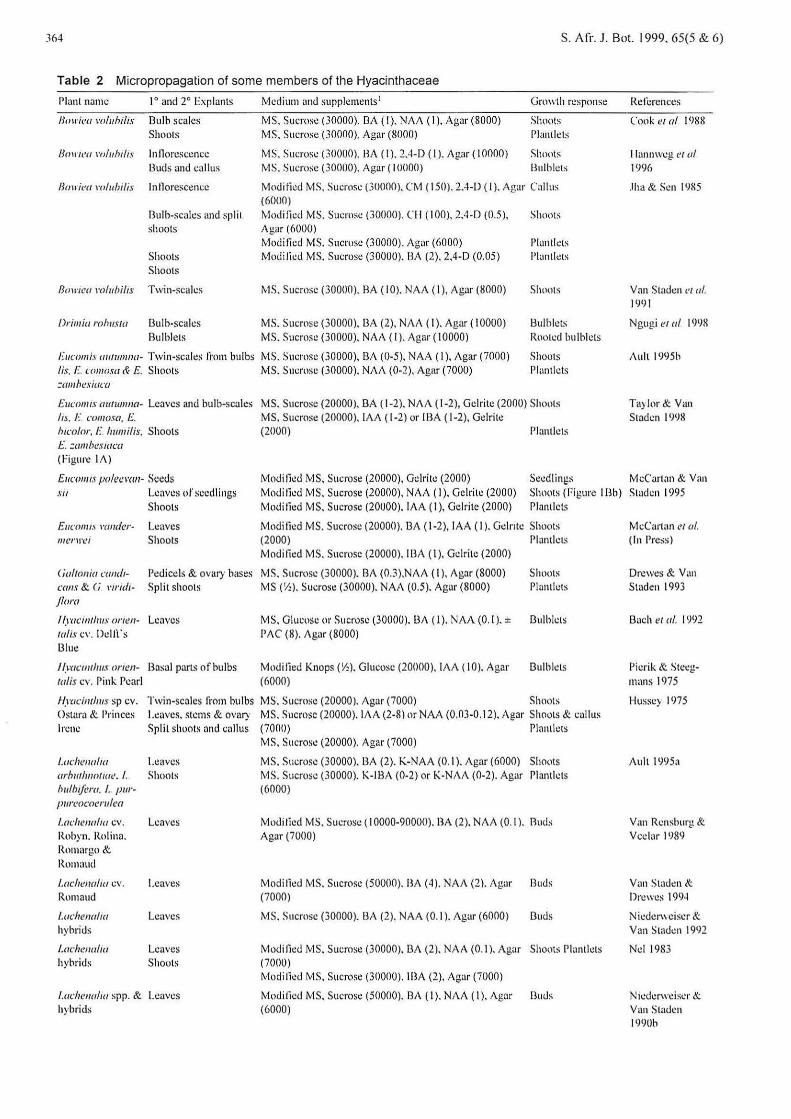
Table 2 Micropropagation of some members of the Hyacinthaceae
Plant name 1° and 2° Explants
/Jowiea \'oluhilis Bulb sc<Jlcs Shoots
/Jourea m/uhrlts In llon:sccnce Buds and callus
/Jou iea mluhilis lntlorescence
Bulb-scales ~nd split shoots
Shoots Shoots
Bowrca l'olubilts Twin-scales
Orimia rollltsta 13ulb-scalcs Rulblets
Medium an<.! supplcments 1
MS. Sucrose (30000), 13A ( I). NAA (I), Agar (8000} MS. Sucrose (30000}. Agar (8000}
MS. Sucrose (30000}. HA (I). 2.4-D ( I). Agar (I 0000) MS. Sm:rose (30000). Agar ( 10000)
Growth response
Shoots Plantlels
Shouts Bul b lets
Mo<.litic<.l MS. Sucrose (30000}. CM ( 150). 2.4-D (I ). Ag<~r Callus (6000) Modi lied MS. Sucrose (30000). Cll ( 100). 2.4 -D (0.5), Shoots Agar (6000) Modi tied MS. Sucrose (30000), Agar (6000) l'lantlets Modi fied MS. Sucrose (30000). BA (2), 2.4-D (0.05) l'lantlets
MS. Sucrnsc (30000). BA ( I 0). NAA ( I), Agar (8000)
MS. Sucrose (30000}, 13A (2), NAA ( 1 ). Ag<~r ( 1 0000) MS. Sucrose (30000). NAA ( 1 ). Agar ( 1 0000)
Shoots
Bulblets Root..:d bulblcts
J:ucomi.~ att1wn1w- Twin-scales H·om bulbs MS. Sucrose (30000), BA (0-5). NAA ( 1 ), Agar (7000) Shoots li.~. [. comosa & £. Shoots MS. Sucrose (30000). NAA (0-2), Agt1r (7000) Plantlets =umhe.1·iaca
Eucoll/1.\' aurw1ma- Leaves and bulb-scales MS. Sucrose (20000}, BA ( 1-2). NAA ( 1-2), Gel rite (2000) Shoots h I:· mnw.1·a. E. MS. Sucrose (20000). lA A ( 1-2) or I BA ( 1-2), Gelrite lun>lor, E lmmilis. Shoots (2000) l'hmtlets E. =ambeswca (rigure lA)
Refi:rences
Cook t'/ a/ I 988
llannweg et a/ 1<)96
.lim & Sen !9!15
Van Staden e1 a/. (991
Ngugi era/ I <Jl)8
Aull 1995b
Ta)lor& Van Stndcn 1998
Eun mrt.1· polccmn- Seeds Modified MS, Sucrose (20000), Gdritc (2000) Secdl in~:s McCartan & Van stt Leaves of seedli ngs
Shoots Mndi lied MS. Sucrose (20000), NAA ( 1), Gelrite (2000) Modified MS. Sucrose (20000). lA A ( 1 ), Gel rite (2000)
Shoots (Figure 1 Bb) Staden I 995 Planllcts
Eucomt.l \'(}}lder-1111!1' \1'1..'1
( ialtonio <'llllllt
mlls & (i l'trldrj/om
Leaves Shoots
l'e<.liccls & ovary bas~s Split shoots
lf.racttllhus orren- Leaves ta/1.1· cv. Delft's 131u~
I f.mcmtlm.t cmen- Basal parts nf bulbs talis cv. Pink Pearl
Modi fied MS. Sucrose (20000). 13A (1-2), lAA (I). Geln te Shoots (2000) Pluntlcts Modi tied MS. Sucrose (20000). 113A (I), Gdrite (2000)
MS. Sucrose (30000). BA (0.3),NAA ( 1 ), Agar (8000) MS (Y: ). Sucrose (30000). NAA (0.5). Agar (8000)
Shoots Plantlcts
MS. Glucose or Sucrose (30000). 13A ( l ). NAA (0. 1 ). ± 13ulblcts PAC (8), Ag<~r (8000)
Modi lied Knops (V.,). Glucose (20000). lAA ( 10). Agar 13ulblcts (6000)
flyad mhu.1· sp cv. Twin-scales ti·om bulbs MS. Sucmsc (20000). Agar (7000) Shoots Ostara & Princes Leaves. stems & ovary MS. Sucrose (20000). lA A (2-8) or NAA (0.03-0.12). Agar Shoots & callus lrcn..: Split shoots and callus (7000) Plantk ts
l.uchenolw Leaves arhurllllollll<'. I. Shoots l>ulhifem . I. purfJIIrt'c>c·oemlca
l.adrenu!ta cv. Rohyn. Rolina. Romargo & Romaud
l.acherwlw cv. Rnmaud
l.adlellalw hybrids
Lachellulw hybrids
Leaves
Leaves
Leaves
Leaves Shoots
l.achenalw spp. & Le<Jves hybrids
MS. Sucrose (20000). Agar (7000)
MS. Sucrose (30000). 13A (2). K-NAA (0.1 ). Ag<~r (6000) Shoots MS. Sucrose (30000). K-IBA (0-2) or K-NAA (0-2). Agar Plantlcts (6000)
Modi tied MS. Sucrose (I 0000-90000). BA (2). NAA (0. 1 ). 11u<.ls Agar (7000)
Modified MS. Sucrose (50000). BA (4). NAA (2). Agar Buds (7000)
MS. Sucrose (30000). BA (2). NAA (0.1 ). Agar (6000) 13uds
Modi lied MS. Sucrose (30000). 13A (2). NAA (0.1 ). Agar Shm1ts Phmth::ls (7000) Modi tied MS. Sucrose (300fl0). lBA (2), Agar (7000)
Mo<.litic<.l MS. Sucrost:: (50000). RA ( 1 ). NAA ( 1 ). Agar Buds (6000)
McCartan et of. (In Press)
Drewc:s & V;m Staden 1993
Bach et a/. I <)')2
Pi erik & Stcc:gmans 1975
Hussc} 1975
Auh 19<J5a
Van Rcnshurg &. Vcelar l 98~
Van Sta<.len & Drewes 199<1
Nic<.lcm ciser & Van Stadcn 1992
Nd 1983
N J<.:derweiser & Van Stac..lt:n 1990h
S. Afr. J. Bot. 1999, 65(5 & 6)
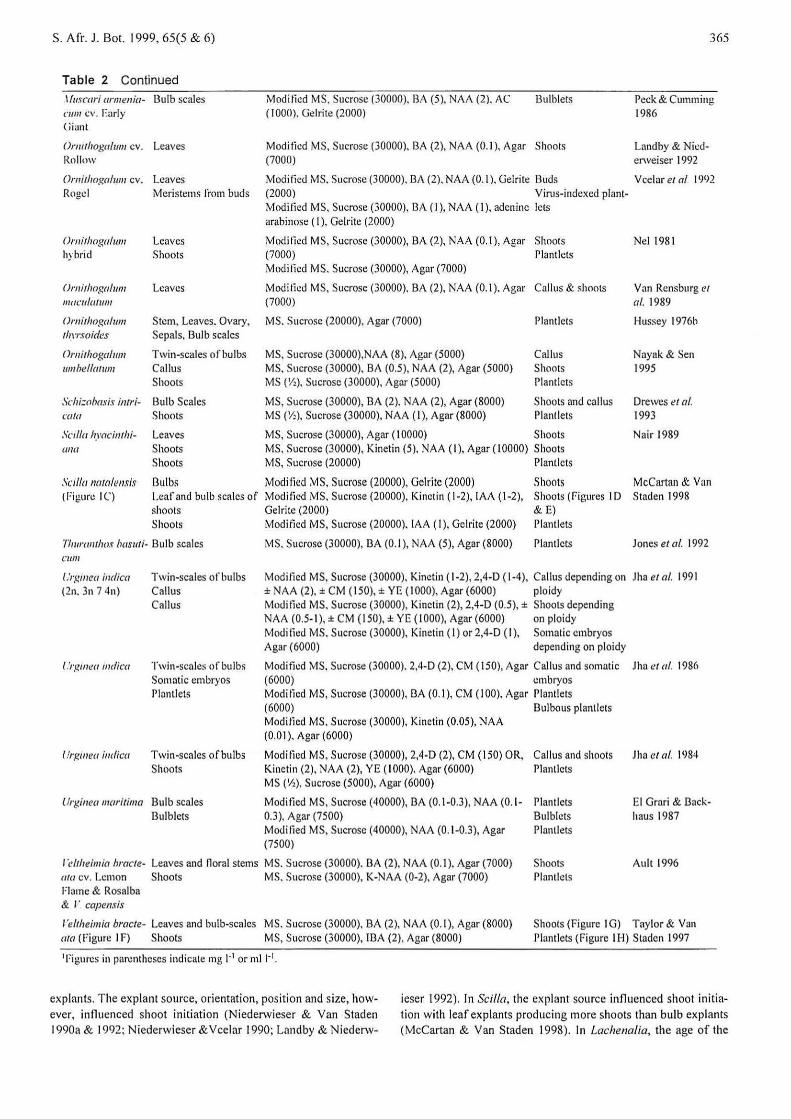
Table 2 Continued
lfmcari armenia- 13ulb scales cw11 cv. Early (jiant
Ormthogalum cv. Leaves Rollmv
Ornithor.alum cv. Leaves Rogel Meristems from buds
()mithogalum hybrid
On1ithoga/unl IIIOCII/lllllll/
( Jmithogalwn tlm·.wides
Omithogalum umhellatum
Sclli:ohmis ino·icctla
Leaves Shoots
Leaves
Stem, Leaves. Ovary, Sepals, Bulb scales
Twin-scales of bulbs Callus Shoots
Bulb Scales Shoots
Sed/a hyacinthi- Leaves ww Shoots
Shoots
Scilla nawlensi.~ l3ulbs ( Figun: I C) Leaf and bulb scales of
shoots Shoots
/lwmntho.f basuti- Bulb scales Cl/111
/ .'rgmea indica (2n. 3n 7 4n}
/ .'rgtnea mdica
Twin-scales of bulbs Callus Callus
Twin-scales of bulbs Somatic embryos Plantlets
Modi lied MS, Sucrose (30000). BA (5}, NAA (2}. AC ( I 000). Gel rite (2000}
Bulblets
Modified MS, Sucrose (30000}, BA (2}, NAA (0.1 }, Agar Shoots (7000}
Modi tied MS. Sucrose (30000}, BA (2}, NAA (0.1 ). Gel rite Buds (2000) Virus-indexed plant-Modilied MS, Sucrose (30000), BA (I}, NAA (I}, adenine lets arabinose ( I), Gelrite (2000}
Modi lied MS, Sucrose (30000), BA (2), NAA (0. 1 }, Agar Shoots (7000} Plantlcts Modified MS. Sucrose (30000), Agar (7000}
Modified MS, Sucrose (30000). BA (2), NAA (0.1). Agar Callus & shoots (7000}
MS. Sucrose (20000), Agar (7000)
MS, Sucrose (30000),NAA (8}, Ag;~r (5000) MS. Sucrose (30000), BA (0.5), NAA (2), Agar (5000} MS ('/>}, Sucrose (30000), Agar (5000)
MS, Sucrose (30000), BA (2). NAA (2), Agar (8000) MS (1/:), Sucrose (30000), NAA ( I), Agar (8000)
Plantlets
Callus Shoots Plantlcts
Shoots and callus Plantlets
365
Peck & Cumming 1986
Landby & Nicderweiser 1992
V celar et a/ I 992
Nel 198 1
Van Rcnsburg et a!. 1989
Hussey 1976h
Nayak & Sen 1995
Drewes eta/. 1993
MS, Sucrose (30000), Agar (I 0000) Shoots Nair 1989 MS. Sucrose (30000), Kinetin (5), NAA ( I), Agar ( 10000) Shoots MS, Sucrose (20000) Plantlcts
Modi tied MS. Sucrose (20000), Gelrite (2000) Shoots McCartan & Van Modi tied MS. Sucrose (20000), Kinetin ( 1-2), IAA ( 1-2), Shoots (figures I D Staden 1998 Gelrile (2000) & E) Moditicd MS, Sucrose (20000). IAA (I), Gel rite (2000) Plantlets
MS. Sucrose (30000), BA (0.1 ), NAA (5), Agar (8000) Ptantlcts Jones eta/. 1992
Modi tied MS, Sucrose (30000), Kinetin ( 1-2), 2,4-D ( 1-4), Callus depending on Jha et a/. t 991 ± NAA (2), ± CM ( 150), ± YE ( I 000), Agar (6000} ploidy Mndi fit:d MS. Sucrose (30000), Kinetin (2}, 2,4-D (0.5), ± Shoots depending NAA (0.5-1 ), ± CM ( 150), ± YE ( 1000), Agar (6000) on ploidy Modi tied MS. Sucrose (30000), Kinetin ( I) or 2,4-D ( I), Somatic embryos Agar (6000) depending on ploidy
Moditkd MS. Sucrose (30000). 2,4-D (2), CM ( 150), Agar Callus and somotic Jha ct a/. 1986 (6000) embryos Modi tied MS, Sucrose (30000), BA (0.1 ), CM ( I 00}. Agar Plantlets (6000) Bulbous plantlets Moditlcd MS. Sucrose (30000), Kinetin (0.05), NAA (0.01 ). Agar (6000}
llrgmea indica Twin-scales of bulbs Modi tied MS, Sucrose (30000), 2,4-D (2), CM (150) OR, Callus and shoots .lha eta/. !984 Shoots Kinetin (2), NAA (2), YE ( 1000), Agar (6000) Plantlets
MS ('h), Sucrose (5000), Agar (6000)
Urginea maritima Bulb scales Bulblets
Modified MS, Sucrose (40000), BA (0.1-0.3), NAA (0. 1- Plantlets 0.3 ), Agar (7500) Bulblets Modified MS, Sucrose (40000), NAA (0.1-0.3), Agar Ptantlets (7500}
El Grnri & Backhaus 1987
l"cltheimia hracte- Leaves and nora! stems MS. Sucrose (30000). BA (2), NAA (0.1 ), Agar (7000) Shoots Plnntlels
Ault 1996 Ilia cv. Lemon Shoots MS. Sucrose (30000), K-NAA (0-2), Agar (7000) Flame & Rosalba & / " capensis
/ "e/theimia bracte- Leaves and bulb-scales MS. Sucrose (30000}, BA (2), NAA (0.1 ), Agar (8000) Shoots (Figure I G) Taylor & Van Plantlets (Figure I H) Staden 1997 ala (Figure t F) Shoots MS, Sucrose (30000), IBA (2), Agar (8000)
1 Figures in parentheses indicate mg 1·1 or m11·1•
explants. T he explant source, orientation, position and s ize, however, influenced shoot initiation (Niederwieser & Van Staden t 990a & 1992: Niederwieser &Vcelar 1990; Landby & Niederw-
ieser 1992). In Scilla, the explant source influenced shoot in itia
tion with leaf ex plants producing more shoots than bulb explants (McCartan & Van Staden 1998). In Lachenalia, the age of the

:>li6 S. Afr. J. Bot. 1999, 65(5 & 6)
Figure I :\ : lntlorescences of Eucomis hybrids and species; B: Adventitious shoot initiated along the periphery of a leaf explant or E. poleevcmsii: C: lntlorescence of Scilla natalcnsis: D: Adventitious buds initiated on the adaxia l surface of a leaf explant of S. natalcnsis; E: Adventitious shoots initiated on a leaf explant of S. natalcnsis; F: Inflorescence of Veltheimia bracteata cv. Lemon rtame; G : Plantlcts of V. hrat·leata cv. Lemon Flame H: Acclimatized plants of V bracteata cv. Lemon Flame.

S. Afr. J. Bot. 1999, 65(5 & 6)
cxplants intluenced shoot initiation, with young ex plants producing more shoots than old explants. This may have been linked to endogenous cytokini n levels particularly in the intermediate and old explants (Niedcrwieser & Van Staden 1990a). In Ornithogalum, the orientat ion of the explants also influenced shoot initiation with <~polar explants producing more shoots than polar ex plants ( Landby & Niederwieser 1992), whi le in Lachenalia. callus and deformed shoots were formed when explants were cultured with the adaxial surface down (Niederwieser & Vcelar 1990). In l.ucllenalia, the explant size influenced shoot initiation wi th small explants producing more shoots than large explants. The optimum size explant was 3.3 x 15 nun {Niederwieser & Vcelar 1990). This may have been related to the wound surface, which is proportionally larger in smaller explants. In Omithogalum. however. additional wounding on the surface of the explant inhibited shoot initiation (Landby & Niederwieser 1992). In I ~mcimlws. the explant size also influenced shoot initiation, which decreased linearly with a decrease in explant size (Pierik & Post 1975).
These adventitious shoots originated from single cells or from groups of ce lls. In Lachenalia. the shoots were initiated mainly from single epidermal cells primarily derivatives of the stomatal mother cells. although a few shoots were ini tiated from groups of cells ( Niedcrwieser & Van Staden 1990b ). The frequency of mutations in shoots initiated from single cells is usually fairly high. resulti ng in ·solid mutations' (Hussey 1980). The adventitious shoots also have been restricted to certain surfaces of cxplants. In IJ111riea. the shoots were initiated on the adaxial surfaces of bulb-scales. This may be associated with anatomical differences between these surfaces. which included thicker cuticles, more chloroplasts and stomata on the abaxial surfaces, as well as the presence of unidenti fied globular bodies on the adaxial surfaces (Van Stadcn et a/ 199 1 ).
Some adventitious shoots have been initiated on medium containing no plant growth regulators, although most adventitious shoots have been initiated on medium contain ing various cytokinins and auxins. In Ladtr:nalia. the add ition of BA had no influence on shoot initiation in hybrids with high cytokinin-like activity. but increased shoot initiation in hybrids with low cytoI..inin-like activity (Niederwieser & Van Staden 1992). Furthermore, the substitution of cytokinin glucosides ([3G)BA, (7G)BA & (9G] I3A l for BA inhibited shoot initiation (Van Staden & Drewes 1994 ). In f/yacinthus, the addition of cytokinins had no influence on shoot initiation. although the addition of auxins, particularl:-- IAA and IBA. increased shoot in itiation (Pierik & Steegmans 1975: Bach & Cecot 1988, 1989). In Scilla, the addition ofNA .~ inhibited shoot initiation (McCartan & Van Staden 1998). while in Hyacintlms. the addition ofNAA promoted callus formation (Pierik & Steegmans 1975). In Hyacinlhus, the carbohydrate source also influenced shoot initiation with glucose and sucrose producing more shoots than fructose (Bach et a/. 1992). In .lluscari, the add ition of charcoal promoted bulblet formation and inhibited callus formation (Peck & Cumming 1986), while in/~\'(ld/1/hus. the addition ofpaclobutrazol promoted bulblet formation (Bach el a/ 1992). Some adventitious shoots produced roots and/or bulbs spontaneously. Other adventitious shoots have been rooted on medium containing no plant growth regulators (Cook et a/ 1988). or various auxins, usually IBA (Nel 198 1. 1983: Taylor & Van Staden 1997) or NAA (Drewes eta/ 1993: Drewes & Van Staden 1993). Although adventitious shoots arc genetically less stable than axillary shoots, particularly when they originate from single cells, (Hussey 1980) this technique is frequently used to clone medicinal and ornamental plants rapidly and economically.
367
Callus and indirect adventitious shoots In Boll'iea, callus was initiated on medium containing 2.4-D and coconut milk, and then transferred to medium containing 2,4-D and casein hydrolysate. This callus was comprised of green nodules, which produced adventitious shoots, when transferred to medium containing low concentrations of 2,4-D (Jha & Sen 1985). In Omithogalum. callus was initiated on medium containing NAA alone (Hussey 1976b; Nayak & Sen 1995) or in combination with BA (Van Rensburg et a!. 1989). The frequency and size of the callus decreased with increasing sucrose concentration (Van Rensburg eta/. 1989). Hussey (1 976b) found that the callus was comprised of a mixture of diploid and tetraploid cells. He also found that the frequency of tetraploid cells increased with callus age (Hussey 1976b). In contrast, Nayak and Sen ( 1995) found that the callus was genetically-stable for several years. The callus produced adventitious shoots when transferred to medium containing no plant growth regulators (Hussey 1976b) or medium containing combinations of BA and NAA (Nayak & Sen 1995). In Urginea, callus was initiated on medium containing 2,4-D, NAA and kinetin. The frequency of callus initiation was increased by the addition of yeast extract. This callus produced small adventitious shoots and roots when transferred to medium containing low concentrations of auxins and vitamins (Jha & Sen 1984).
Although vast quantities of adventitious shoots can be produced from callus, the frequency of genetically-aberrant shoots may be relatively high. Thus, propagation via indirect adventitious shoots is usually avoided. In certain breeding programmes, however, genetically-aberrant plantlets could be a useful source of somaclonal variation .
Direct and indirect somatic embryos In Scilla, embryogenic callus was initiated on medium containing NAA and coconut milk. The embryogenic callus, which was comprised of small thin-walled cells with large nuclei, formed shoots and roots when transferred to medium containing no plant growth regulators (Chakravarty & Sen 1989). In Urginea. embryogenic and non-embryogenic callus was initiated on medium containing 2,4-D and coconut milk. The embryogenic callus, which was comprised of large, vacuolated parenchyma cells, formed greenish zones of small cells with large nuclei when exposed to 2,4- D for prolonged periods. These greenish zones produced globular embryos, which elongated to form banana-shaped bipolar embryos when transferred to medium containing low concentrations of BA with or without coconut milk. The bipolar embryos produced bulbous plantlets when transferred to medium containing kinetin and NAA. In Urginea, the age of the callus, the frequency of sub-culturing and the cytological state ofthe callus influenced the initiation and subsequent development of the callus (Jha & Sen 1986). The ploidy of the callus (diploid & tetraploid) also influenced medium requirements (Jha eta/. 1991 ). Despi te the potential of this technique, somatic embryos have not been induced in many members of the Hyacinthaceae.
Conclusions The Hyacinthaceae comprises several genera, which are widely exploited for their medicinal, pharmaceutical and ornamental potential. In South Africa, several members of the Hyacinthaceae are harvested without permits from wi ld populations, processed and then sold as traditional medicine. This is reducing the density, distribution and genetic divers ity of wild populations. Furthermore, the enforcement of existing legislation has proved ineffective with vast quantities of plants being traded locally and internationally. Consequently, it has been suggested that ex situ

t.:onse::rvation through cultivation may alleviate pressures on natural resources, whilst meeting the demand for these plants. Conventional vegetative propagation, however, is usually fairly s low. Micropropagation, therefore, provides a rap id and economical means to propagate these endangered plants, whilst supplying an alternative source of superior quality plants for the consumer market.
Many bufadienolides have been isolated from members of the Hyacinthace::ae. These bufadienolides are used as cardiotonics and, therefore. have pharmaceutical potential. This has led to the identification and quantification of bufadienolides in various clones and cytotypes in vitro and in situ to select high yielding s trains suitable for commercial exploitation. In Bowiea, the same bufadienolides were accumulated at similar concentrations for both in 1·irro and in situ plants (Finnie et a/. 1994 ). In Urginea, however. the accumulation of bufadienolides was linked to the format ion of specific organs in vitro, but was independent of the genotype. explant source and regeneration system (Jha et a/. I 99 I). Micropropagation, therefore, not only provides a rapid and economical means to propagate selected chemotypes, but a lso a means to produce these bufadienolides in 1•itro. This is important economically s ince it obviates the need to cultivate these plants ex virrum.
Several members of the Hyacinthaceae are cultivated as ornarne::ntals. although the total area allotted to the production of these bulbs is relatively small. Breeding programmes, which have been established for certain genera, may increase the popularity of these plants. Micropropagation, therefore, not only provides a rapid and economical means to propagate newl:r-devcloped cultivars, but also a means to eliminate viruses sut.:h as Hyacinthus Mosaic Virus (B lom-Barnhoorn 1986) and Ornithogalum Mosaic Virus (Vcelar eta/. 1992), thus reducing crop losses and faci litating the export of these plants. Micropropagation also provides a means to conserve valuable germplasm (Louw 1995), therefore retaining useful character traits for future breeding programmes.
Acknowledgements The:: National Research Foundation and the University of Natal Re::search Fund are thanked for financial assistance. Ms J.L.S. Taylor kindly provided Figures I A, F, G and H.
References ARNOLD. T.H. & DE WET. B.C. 1993. Plants of Southem Africa:
Names and Distribution, pp. 135, 146-155. NBI. Pretoria. A Ul.T . .I .R 1995a. In vitro rooting and grccnhous~ ncclimntization of
l.adllmalia shoots. HortScience 30: 1304- 1305. i\ lll.T .. l.lt 1995b. In vitro propagation of Eucomis aiiii/IIIIW!is. E
t'omosa. and £. =ambesiocn by twin-scaling. HortSctence 30: 144 1-1442.
i\ ULT .. l.lt 1996. In vitro rooting nnd greenhouse acclimatization of I 'elrheunia hracteata and V. capensis shoots. HortScience 31: 1229-1230.
11ACI I. A. & CECOT. A. 1988. Micropropogation of Hyacinth us orienwlts cv. Camegie in a double-phase culture medium. I. Effect of cytokmins on growth and development. Acln Hort . 226: 607- 610.
BACH. A. & CECOT, A. 1989. Application of double-phase culture media in micropropagation of Hyacimhus orien!alts cv. Camegie. II . tfl'e.:t of auxins on growth and development. Acta Hort. 251: 215-221.
lli\Cll. /\. .. CECOT. A. & GAWEDA. M. 1992. Utilization of soluble ~:arbohydrates in shoot and bulb regeneration of Hyacimhus oriemnlis L. in vilro. Acta flort . 325: 487-492.
BALI3AA. S. l.. KHAfAGY, S.M .. KHAYYAL. S.E. & GIRGIS. A.N. 1979. TLC-spectrophotometric assay of the main glycosides of Red Squill. a speci~ic rodenticide. J. /1'01. Prod. 42: 522- 524.
131\ TIEN. A. & BOKELMANN. H. 1966. Wildflowers of the Enstcm
S. A li·. J. Bot. 1999, 65(5 & 6)
Cape Province. pp. 8- 15. Books of Africa. Cape Town. 13 LOM-BARNHOORN. G.J., VAN AARTRI.IK . .1 . & VAN DER
LINDE. P.C.G. 1986. Effect ofviraznle on thc production ofh)acinth plants free li-mn hyacinth mosaic virus (I·IMV) by meristcm cultun;. Acta Hort. 177:571-574.
BRYAN, J.E. 1989. Bulbs. Vol. 2. pp. 324. Timber Press inc .. !long Kong.
BUD/\ V ARI. S. (Ed) 1996. The Merck. Index - an Encyclopedia of Chemicals, Drugs and Biochemicals. 12th Ed. pp. 1352. Whitehou~c Station. NJ.
CHAKRAVARTY. B & SEN. S. 1989. Regeneration through somatic embryogenesis !rom anther explants of Scilla indica (Roxb.) 13akcr. Plant Cell Tiss. Org. Cult. 19: 71-75.
COOK. E.L.. CUNNINGHAM, A. & VAN STADEN . .1 . 1988. The ussue culture of an exploited Zulu medicinal plant, Bmviea mlubi!ts S Afr. J. Rot. 54: 509-5 10.
CRABTREE. D.G. 1947. Red Squill. thc most spcc ilic of the raticiucs. Eco. Bot. I: 394-40 I.
CUNNINGHAM. A.B. 1988. An invcstigiltion of the herbal mcdicinc trade in Natal/Kwazulu. INR report 29. l' ictermantzhurg.
DAHLGREN, R.M.T., CLIFFORD. H.T. & YEO. P.F. 1985. Thc limulies of the Monocotyledons- Structure, Evolution and Taxonomy. pp. 129- 133, 188- 193. Springcr-Vcrlag. Berl in.
DE HERTOGH, A.A. & I.E NARD. M. 199h World production ami horticnltural utilization of flower bulbs. In: Thc Physiology of f- lower llulbs, eds. A.A. De I lcrtogh and M. Lc Nard. pp. 21- 28. Elsevh:r. The Netherlands.
DE HER TOG H. A.A. & LENARD. M. 1993h. Gcnc:ral chapter on summer !lowering bulbs, In: The Physiology of Flowe::r Bulbs, cds. A.A. De Hertogh. and M. LeNard. pp. 74 1-774. Elsevier. The Ncthcrbnds.
DREWES. F.E., BAYLEY. A.D. & VAN STADEN. J. 1993. Tissue culture of Schi:obnsis 111tricaw. a medicinal plant. S. Afr. J /Jot 59: I 05-106.
DREWES, F.E. & VAN STADEN . .1. 1993. In l'itro propagation of Ualtonia species. S. Afr J. Bot. 59: 457-458.
DU PLESSIS. N. & DUNCAN, G. 1989. Bulbous plants of South..:m Africa. A Guide to their Cult ivation and Propagation. pp 71- 84 Tatl!lbcrg. Cape Town.
EL GRARI. R .. & 13ACKHAUS. R.A. 1987. In riu·o propagation of red squill. Urgineonwntima Baker. Plant Cell Tis.1·. Org. Cult. I 0: 65-71.
FINNIE . .J .f.. DREWES. F. F.. & VAN STADEN . .J . 1994. 80\rnmmluhi/is Harv. ex Hook.!: (Sea Onion): ht vitro cultun: uno prudu~:tion of cardiac glycosides. In: Biotechnology in Agriculture :md foorestr). Vol 28. Medicinal and Aromatic Plants VI I . cd . Y. l'.S . 13aja,i. pp 84-97. Springer-Verlag. 13erlin .
GENTRY. 1-l .S .. VERBISCAR. A . .l. & 13ANIGAN. T.F. 19!17. Red Squill ( Urginea maritima, Liliaccue). Em. Bot. 41: 267-282.
GOSLING. M. 1998. Muti mcccn taking root. Cape Times April 17. pp. 18.
IIANNWEG. K .. WATT. M.P. & 13ERJAK. P. 1996. A simple method for the micropropngation of IJ01riea \'Ofubifts li-\1111 intloreSCI!nCe explants. Bot. Bull. A end Sin. 3 7: 213-2 18.
HUSSEY. G. 1975. Propngation nf hyilcinths by tissue culture. S". Hart. 3: 21-28.
HUSSEY. G. 1976a. ln1•ifro rele<tse ofn.xi llary shot1ts from apical !.luminance in Monocotyledonous plant lets. Ann. Bot. 40: 1 32~- 1 325.
HUSSEY. G. 1976b. Plantlct regeneration from callus and pnrell! tissue in Ornithogolumtltyrsoides. J. Exp. Bot. 27: 375-382.
HUSSEY. G. 1980. Propagation of some members of the Li l iacea~. lndnceae and Amaryllidaccac by tissue culture. In: l'etaloid Mouowtyleuons (Horticultural and Botanical Research). eds. C.D. Rrickell. D.F. Cutler & M. Gregor). pp. 33-45. Academic Pn:ss. NcwYork .
HUTCHINGS. A. 1989. A survey nnd analysis of traditional medicinal plants as used by the Zulu. Xhosa and Sotho. B01ltn!in 19: 111 - 123.
HUTCHINGS. A .. SCOTI'. A. H., LEWIS. G. & CUNNINGllAM. A.B. 1996. Zulu Medicinal Plants- U\1 ln\'entury. pp. 38-44. l.lni\crsi t~ or Natal Prcss. Pietennnritzburg.
JACOT GUILLAMOD. A. 197 1. Flora of Lt:sotho (Basutoland ). pp. -108-458. Vcrlag von .1. Cramer. Gennany.

S. Afr. J. Bot. 1999, 65(5 & 6)
JAGER. A.K. & VAN STADEN. J. 1995. Screening lor cardiac glyco· sides in Schb1husis il1lricata. S. Afr. J. Bot. 61: I 01- 103.
.lilA. S. 1988. 13ufadiennlidcs.ln: Cdl Culture and Somatic Cell Genet· ics of Plants. Vnl 5. Phytocht:micals in plant cell cutures. cd. l.K. Vasil. pp. 179-191. Academic Press Inc .. Flu.
.IliA. S .. MITRA. G.C.. & SEN. S. 1984./n vuro regeneration from bulb explunts of Indian squi ll. Urgmea indica Kunth. Plant Cell Tiss. Org. Cult. 3:9 1- 100.
JfiA. S .. SAII U. N.P .. & MAHATO. S.B. 1991. Callus induction, orga· nngencsis and somatic embryogenesis in three chromosomal races of llrginea nullca and production of hui~Idiennlides. Plam Cell Tiss. Org. Cult 25: 85-90.
JHA. S. & SEN. S. 1981. Bufadienolides in different chromosomal races of Indian Squill. Phytochemistrv 20: 524-526.
.IHA. S. & SEN, S. 1983. Quantitation of principal bufadicnolides in dit~ lercnt cytntypes of Urginea indica. Planta Med1ca 47: 43-45.
.IliA. S & SEN. S. 1985. Regeneration and rapid multiplication of Boll'· 1ea voluhdis Harv. in tissue culture. Plant Cell Rep. 4: 12-14.
.lilA. S. & SEN. S. 1986. Development of lndian Squill (Urginea mdica Kunth.) through somatic embi)"ogencsis from long tcm1 culture. J. P/a/11. Physio/. 124: 43 1-439.
JONES. N.IJ .. MITCHELL . .1., BAYLEY. A.D. & VAN STADEN. J. I 992. ln1·itm propagation of l'llllmntlws baswicum. S. Afr. J. Bot. 58: 403-404.
l.ANDDY. P.A. & N!EDERWIESER. J.G. 1992./nvitro propagation of Omitlwgalum 'Rollow'. J. S Afr. Soc. Hort. SCJ. 2: 50-54.
I.E NARD. M. & DE HERTOGH. A.A. 1993. General Chapter on Spring-flowering Bulbs. In: The Physiology nf Flower Bulbs. cds. A.A. De llt!rtogh and M. Lc Nard. pp 705-739. Elsevit:r, The Nether· hinds.
l.OUW. E. 1995. Long tcm1 storage of Lachena/ia shoot tips. J. S. Afr Soc. Horl. Sci. 5: 77-80.
l.UYT. R.P .. JAG ER. A.K. & VAN STADEN. J. 1999 The rational usage of /Jrimia robusta Dak. in traditional medicine. S. Afr. J. Rot. (In Press).
MANDER. M .. MANDER . .1 .. CROUCH, N .. McKEAN. S. & NICHOI.S. G. 1995. Catchment action: Growing and knowing muthi plants. pp. I 1.14 .18.
MANDER. M. 1997. The marketing of indigenous medicinal plants in South All·ica: a case study in Kwaz.ulu-Natal. INR report 164. PicW· maritzhurg.
1v11:CARTAN. SA & VAN STADEN, J. 1995. In v1tro propagation of the medicinal plant. Eucomis poleevansli N. E. Brown. J. S. Afr. Soc. 1/ort. Sci. 5: 73-75.
McCARTAN. SA & VAN STADEN . .1. 1998. Micropropagation of the medicinal plant. Scilla natalensis Planch. Plant Growth Reg. 25: 177-180.
McCARTAN. SA. CROUCH, N.R. & KRYNAUW. S. 1999. Micm· propagatitm of a naturally rare pineapple lily, Eucomi~· vandermerwei I. Verd . .1. S. Afi'. Soc. Hort. Sci. {In Press)
NAIR. A.S. I 989. Micropropagation of Scifla hyacinthiana (Roth) Mac· bride. Proc. Indian. Natn. Sci. Amd. 2: 121- 124.
NA YAK. S. & SEN, S. 1995. Rapid and stable propagation of Ornithog· alum umbeflatum L. in long tenn culture. Plant Cell Rep. 15: 150-153.
NEL. D.O. I 98 1. Rapid propagation of Omithogalum hybrid in vitro. Awoplantae 13: 83- 84.
NE!.. D.D. 1983. Rapid propagation of Lachenalia hybrids in vitro. S Afi'. J. Bot. 2: 245- 246.
NGUGI. G.W .. JAGER. A.K. & VAN STADEN, J. 1998. In vitro prop· agation of Drimia robusta 13ak. S. Afi'. J. Bot. 64: 266- 268.
NIEDER WIESER. J .G., ANANDAJA YASEKERAM, P .. COETZEE. M. MARTELLA. D., PIETERSE, !3.1. & MARASAS, C.N. 1998. Resc;Irch impact assessment as a management tool: Lachenalia research at ARC·Roodeplaat as a case study . .1. S. Afr. Soc. Horr. Sci. 8: 80-84.
NIEDER WIESER, J .G. & VAN STADEN, .1 . 1990a. The relationship between genotype, tissue age and cndogncous cytokinin levels on adventitious bud fonnation on leaves of l.achenulia. Plant Cell Tiss.
369
& Org. Cult. 22: 223-228. NIEDER WIESER. J.G. & VAN STADEN . .1 . l990h. Origin nfmhcnti·
tious buds on cultured l.achenalm leaves. Be11r Bwl. 1'/lcm:en 65:
443-453 . NIEDER WIESER . .I.Ci . & VAN STADEN .. 1. 1992.1nteraction hetwccn
bcnzyladenine. naphthalcneacctic acid and tissue age on ad\·entil ious
bud formation on leaf sections of l.achenalia hybrids. S Aji· J /Jot
58: 13-16. NIEDERWIESER. J.G. & VCTI.AR. B.l\1. 1990. Regeneration of
Lachenalia species I rom leaf ex plants. llorJ51m:nce 25: 684-6!\7. PECK. D.E. & GUMMING. 13.Ci. I 986. Bcnetkial effects of act inned
charcoal on bulblet production in tissue cul tures of Muscan armenia·
cum. Plant Cell ?iss. Org. Cult. 6: \J- 14 . PERRY. P. 1985. The restmcturing of the t~unily l.iliaccac. f"e/d &
Flora 7 1: 66-68 . !'!ERIK, R.L.M .. & POST. A.J.M . 1975. Rapid vegetative propagation
of f~vacmthus orienta/is L. in varo. Sci. firm. 3: 293-2'.17 . !'I ERIK, R.L.M. & STEEGtv1ANS. H.H.M. 1975. Etlecl ofau:-.ins. cyto
kinins, gibbcn:llins. ubscisic acid and ethephon on regeneration and
growth of bulb lets on excised bulb scale segments of Hyacinth. /'hys
iol. Plant. 34: 14-17. REES. A.R. 1992. Onmncntul bulbs. corms and tubers. pp. 93- 11 I.
CAB lnternutionul, UK. Sl!YR. S.E. & STAB A, E.J. 1976. Examination of Squi ll tissue cult ures
ll.1r bufadicnolidcs and anthocyan ins. /'Iaiita Mec!Jca 29: 86- 90. TAYLOR. J.L.S. & VAN STADEN . .1. 1997. In wlro propagation of
Ve/Jheimia hracteata and V bracteara ' l.t:mon Flame'. S . . ·!fr . .I fim
63: 159-161. TAYLOR. J.L.S. & VAN STADEN. J. 1998. Micropwpagation of
Eummis species: a plant with horticultural and medicinal pot.:nti al. IX
111/emationa/ Congress on Plalll Tissue & Cell C'uflllre . .l~ rusalcm.
Israel. VAN HORN. D.l.. & DOMINGO. W.E. 1950. Comparison of seed <lnd
vegetative propagation methods f(1r Red Squi ll. /;'co. Bot. 4: 350- 353. VAN RENSBURG . .I .G .. VCELAR. D.M. & LANDDY. P.A. 1989.
Micropropagation of Omithogolum mawlatum. S. Afi'. J. Hot 55:
137- 139. VAN RENSBURG. J.G. & VCELAR. l3.M. 19g9_ The d'ti:ct ofsucrnsc
concentratitm on the initiatitm and growth of adventi tious bmls !hun
leuf tissue of Lachenalia. S. Afr . .!. Bot. 55: 117- 121. VAN STADEN, J. & DREWES. F.E. 1994. The efti:ct of bt:nz) ladcnim:
and its glucosides on udventitious bud formation on Laclumalia lear
s~ct i ons. S. Afr. J. Bot. 60: 191 - 192. VAN STADEN, J.. ELMER-ENGLISH. C.W. & FINNIE. J.r:. 199 1.
Phys iological and anatomical aspects related to m VI tro shoot initia
tion in Bowiea volubilis. S. rlfr. J. Bot. 57: 352-355. VANWYK, B.-E .. VAN OUDTSHOORN. B. & GERICKE. N. 1997.
Medicinal Plants of South Ali·ica. pp. 16- 18. 11 2. Brizu Publications.
Pretoria. VCELAR, D.M., FERREIRA. 0 .1. & NIEDER\VIESER. Hi. 1992.
Elimination of omithogalum mosaic virus in Omithogalum cv. Rogd through meristem tip cu ltur~ and chemotherapy. /'/ant Cell T1ss Org.
Cult. 29: 51-55. VERI3ISCAR, A.J .. PATEL. J.. f3ANIGAN. T.F. & SCHATZ. R.A.
1986. Scillirosidc and other sci lla compounds in Red Squil l. J. Agric.
Food. Chenr. 34: 973- 979. WALTON, J., BARONDESS. J.A. & LOCK. S. 1994. The Ox ford Mcd·
ical Companion, pp. 106. Oxtbrd University Press. 0:\tord. WATT. J.M. & I3REYER·I3RANDW I.I K. M.G. 1962. Medicinal and
poisonous plants of Southern and Eastern Africa. 2nd Ed. , pp. 669-
725 . E & S. Livingstone Ltd., Edinburgh. WILLIAMS, V.L. 1996. The Witwatersrand Muti Trade. J'e/d & Flora
82: 12- 14.