Embed Size (px)
Citation preview

Gene 578 (2016) 263–273
Contents lists available at ScienceDirect
Gene
j ourna l homepage: www.e lsev ie r .com/ locate /gene
Research paper
Microarray-based gene expression profiling reveals genes and pathwaysinvolved in the oncogenic function of REG3A on pancreatic cancer cells
Qianqian Xu a, Rong Fu a, Guoxiao Yin a, Xiulan Liu a, Yang Liu b, Ming Xiang a,⁎a Department of Pharmacology, School of Pharmacy, Tongji Medical College, Huazhong University of Science and Technology, Wuhan 430030, Chinab Synergy Innovation Center of Biological Peptide Antidiabetics of Hubei Province, School of Life Science, Wuchang University of Technology, Wuhan 430223, China
Abbreviations:AP, Acute pancreatitis; BACH1, BTB andfactor 2; CARD9, Caspase recruitment domain family, memoncogene homolog; c-Jun, Jun proto-oncogene; CLDN2, CDOK3, Docking protein 3; EGF, Epidermal growth factor;GATA binding protein 2 (globin transcription factor 2); G1; JAK2, Janus kinase 2; KEGG, Kyoto Encyclopedia of GenMitogen activated kinase-like protein 3;MAPK4, Mitogen14;MRAS, Muscle RAS oncogene homolog;MYC, V-myc avbox 4; PDAC, Pancreatic ductal adenocarcinoma; PDGF,bisphosphate 3-kinase, catalytic subunit alpha; PPP2CB, Prnine nucleotide exchange factor (GEF) 6; RASAL2, RAS prohomology 2 domain containing transforming protein 1; SM(Drosophila); STAT3, Signal transducer and activator of trahibitor of metalloproteinase 1; TIMP2, Tissue inhibitor of mVascular endothelial growth factor.⁎ Corresponding author.
E-mail address: [email protected] (M. Xia
http://dx.doi.org/10.1016/j.gene.2015.12.0390378-1119/© 2015 Elsevier B.V. All rights reserved.
a b s t r a c t
a r t i c l e i n f oArticle history:Received 4 July 2015Received in revised form 18 October 2015Accepted 16 December 2015Available online 21 December 2015
We previously reported that regenerating islet-derived protein 3 alpha (REG3A) exacerbates pancreatic malig-nancies. The mechanism of this effect has not been clearly elucidated. Here we first identified key differentiallyexpressed genes (DEGs) and signal pathways in the pancreatic cancer cell line SW1990, compared to two controlcell lines, bymicroarray analysis.We then identified key genes and pathways regulated by REG3A or the cytokineIL6 in SW1990 cells. Afterwards, these DEGs induced by REG3A or IL6 were subjected to KEGG pathway enrich-ment analysis and GO function analysis by the DAVID online tool. Ultimately, we constructed protein–protein in-teraction networks among the DEGs by Cytoscape. Among the three pancreatic cell lines, SW1990 exhibitedhighly deterioration with the activation of genes and pathways related to proliferation, survival, angiogenesis,and invasion. As a result, 50 DEGs enriched in 11 pathways were identified in REG3A-treated SW1990 cells,and 28 DEGs enriched in 9 pathways were detected in IL6-treated cells. Overall, results of microarray analysisfollowed by qRT-PCR and Western blotting suggest that REG3A regulates pancreatic cell growth by increasingthe expression of at least 8 genes: JAK1, STAT3, IL10, FOXM1, KRAS, MYC, CyclinD1, and c-fos; and activation of atleast 4 signal pathways: TGFβ, PDGF, angiogenesis and RAS. Similar results were obtained with IL6 treatment.Regulation network analysis confirmed the cell growth related DEGs, and further uncovered three transcriptionfactor families with immune functions regulated by REG3A.
© 2015 Elsevier B.V. All rights reserved.
Keywords:REG3ADifferentially expressed genesPancreatic cancerProliferation
1. Introduction
Pancreatic cancer, one of themost lethal and intractable humanma-lignancies, has a reported five year survival rate of less than 5% (Hidalgo,2010). It was estimated that the number of patients with pancreaticcancer in 2015 would increase to 48,960 in the United States, and of
CNC homology 1, basic leucine zipperber 9; CDK4, Cyclin-dependent kina
laudin 2; DAVID, The Database for AFOXM1, Forkhead box M1; FOXO3, FO, Gene Ontology; GRB2, Growth faces and Genomes; KRAS, Kirsten rat saactivated kinase-like protein 4;MAPKianmyelocytomatosis viral oncogenePlatelet derived growth factor; PID1otein phosphatase 2, catalytic subunittein activator like 2; REG3A, RegeneraAD4, SMAD familymember 4; SOCS3nscription 3; STAT5A, Signal transducetalloproteinase 2; TP53, Tumor pro
ng).
those, 40,560 would die (Siegel et al., 2015). Nevertheless, this lethaldisease is still difficult to treat effectively, in part because its pathogen-esis has not been thoroughly characterized.
One widely-held hypothesis is that infaust changes of certain genesare implicated in the etiology of pancreatic cancer. Specifically,alterations of Kirsten rat sarcoma viral oncogene homolog (KRAS),
transcription factor 1; BACH2, BTB and CNC homology 1, basic leucine zipper transcriptionse 4; CDKN2A, Cyclin-dependent kinase inhibitor 2A; c-fos, FBJ murine osteosarcoma viralnnotation, Visualization, and Integrated Discovery; DEGs, Differentially expressed genes;orkhead box O3; GATA1, GATA binding protein 1 (globin transcription factor 1); GATA2,tor receptor-bound protein 2; IL10, Interleukin-10; IL6, Interleukin-6; JAK1, Janus kinasercoma viral oncogene homolog;MAPK1, Mitogen activated kinase-like protein 1;MAPK3,9, Mitogen activated kinase-like protein 9;MAPK14, Mitogen activated kinase-like proteinhomolog;NF-κB, Nuclear factor kappa B; PAP, Pancreatitis-associated protein; PAX4, Paired, Phosphotyrosine interaction domain containing 1; PIK3CA, Phosphatidylinositol-4,5-, beta isozyme; RALB, V-ral simian leukemia viral oncogene homolog B; RAPGEF6, Rap gua-ting islet-derived protein 3 alpha; RT-PCR, Real-time polymerase chain reaction; SHC1, Src, Suppressor of cytokine signaling 3; SPRY1, Sprouty homolog 1, antagonist of FGF signalinger and activator of transcription 5A; TGFβ, Transforming growth factor β; TIMP1, Tissue in-tein p53; TWIST1, Twist family bHLH transcription factor 1; Tyk2, Tyrosine kinase 2; VEGF,

264 Q. Xu et al. / Gene 578 (2016) 263–273
cyclin-dependent kinase inhibitor 2A (p16/CDKN2A), tumor proteinp53 (TP53) and SMAD family member 4 (SMAD4) are all detected inmore than 50% of pancreatic cancer patients (Sheng et al., 2014). Theexcessive activation of KRAS in particular is an essential step in thedevelopment of many cancers (Yu et al., 2010). KRAS exerts its carcino-genic effects through the activation of mitogen activated kinase-likeprotein (MAPK) and PI3 kinase signaling pathway (Ji et al., 2007). In ap-proximately 80% of pancreatic ductal adenocarcinoma (PDAC) cases,there is a loss-of-function of p16/CDKN2A, a tumor suppressor gene in-volved in cell cycle regulation (Heilmann et al., 2014). Mutations inTP53 are also prevalent. TP53 codes tumor protein p53 to inhibit cancerformation through several pathways involved in cell cycle arrest and ap-optotic death (Mohamadkhani et al., 2013). SMAD4 codes proteinswhich are intracellular mediators of transforming growth factor β(TGFβ). Previous studies indicate that the absence of SMAD4 is neces-sary for the progression of pancreatic cancer (Legendre et al., 2014).
REG3A (regenerating islet-derived protein 3 alpha, also known aspancreatitis-associated protein or PAP) is a C-type lectin protein secret-ed by acini during acute pancreatitis. REG3A is a member of a family ofREG proteins. It exerts its tissue protection function at least in partthrough activating nuclear factor kappa B (NF-κB) signaling pathwaysin acute pancreatitis (AP) (Okochi et al., 2014). However, overexpres-sion of REG3 in pancreas may also promote cell proliferation of islettumor cells in mice and acinar epithelial cells in rats (Folch-Puy et al.,2006; Cui et al., 2009). In addition, REG3 stimulates cell growth by acti-vating Akt kinase and up-regulating cyclinD1/cyclin-dependent kinase4 (CDK4) signaling pathway (Cui et al., 2009). Our previous studieshave indicated that REG3 is involved in the regeneration or growth ofislet β cell (Hou et al., 2011). We reported that REG3A overexpression,in synergy with suppressor of cytokine signaling 3 (SOCS3) methyla-tion, promoted cell proliferation in pancreatic cancer as well as duringinflammation-linked pancreatic carcinogenesis (Wang et al., 2014).Further, we found that REG3A could constitute a REG3A-janus kinase2 (JAK2)/signal transducer and activator of transcription 3 (STAT3) pos-itive feedback loop, and that REG3A shared the canonical inflammation-related carcinogenic JAK2/STAT3 pathway with the inflammatory cyto-kine interleukin 6 (IL6) (Liu et al., 2015). IL6 is secreted by T cells andmacrophages to stimulate immune response. The cytokine can alsopromote the progression of pancreatic intraepithelial neoplasia anddevelopment of pancreatic cancer by activating STAT3/SOCS3 signalingpathways (Lesina et al., 2011) and by stimulating the secretion ofmultiple Th2 cytokines (Feurino et al., 2007).
mRNA expression array analysis is an efficient tool for highthroughput analysis and for the screening of differentially expressedgenes involved in carcinogenesis, including the pathogenesis of pan-creatic cancers (Xie et al., 2015). Here, we applied this technique toidentify and compare abnormalities in gene expression during
Table 1The primers of human genes used in qRT-PCR.
Gene Primer sequence (5′ → 3′)
IL10 F: ATG CCC CAA GCT GAG AAC CAA GAC CCAR: TCT CAA GGG GCT GGG TCA GCT ATC CCA
FOXM1 F: AAC CGC TAC TTG ACA TTGGR: GCA GTG GCT TCA TCT TTCC
JAK1 F: CTG GTA TGC TCC AAA TCGR: CAT CCC TAG ACA CTC GTT CT
CyclinD1 F: TGC ACA GTG TCA CGA ACA GAR: ACC TCG GAG AAG GCT AAA CA
c-fos F: TGT CAA CGC GCA GGA CTT CTR: CCT TCT CCT TCA GCA GGT TG
MYC F: CAA ACC TCC TCA CAG CCC ACTR: TGA CAC TGT CCA ACT TGA CCC
KRAS F:GAG GCC TGC TGA AAA TGA CTGR:ATT ACT ACT TGC TTC CTG TAGG
STAT3 F:GTG TAT GCG TCG GCT TCAR:GAC TCT GCG GGT CCT GTT
tumor initiation in the high malignant pancreatic cancer cell lineSW1990 (Zhou et al., 2015), with noncancerous HPDE6c7, an immor-talized but not transformed pancreatic ductal epithelial cell line, andPANC-1, a less malignant pancreatic ductal cancer cell line, as con-trols. The genetic background of the primary pancreatic cancer cellline PANC-1 is mutant KRAS, mutant TP53, wild type (wt) SMAD4,and homozygous deletion of p16. The genetic background of themetastatic pancreatic cancer cell line SW1990 is mutant KRAS, wtTP53, not determined (ND) SMAD4, and ND p16. The noncancerouscell line HPDE6c7 is near normal genotype and phenotype (Ouyanget al., 2000; Deer et al., 2010). We focused on, but did not restrict ouranalysis to, the expressions of KRAS, p16/CDKN2A, TP53, and SMAD4,genes that have been identified to be mutation in these cells (Oshimaet al., 2013). Our previous studies have revealed that among thesefive pancreatic cancer cells AsPC-1, BXPC-3, Mia Paca-2, PANC-1 andSW1990, and the normal control cell HPDE6c7, REG3A expressedhighest in SW1990, secondly in PANC-1, and nearly not expressed inHPDE6c7. Besides, after incubation with 50 ng/mL REG3A protein for24 h, SW1990 possessed higher cell viability, more colony numbersthan PANC-1. Scarcely any studies locate the receptor of REG3A, andour previous research speculated that EGFR might function as the re-ceptor of REG3A (Liu et al., 2015). In addition, IL6 receptor gp130expressed equally in pancreatic cancer cell lines as well as noncancer-ous pancreatic cell HPDE6c7 (Goumas et al., 2015). On the basis ofthose findings, we then investigated the mechanism of action of thegrowth factor REG3A in inducing inflammation-linked pancreatic can-cer development in SW1990 (Wang et al., 2014).
In this study, we approached this problem by: 1) comparing geneticprofiles of the SW1990 cancer cell line with those of two control celllines; and 2) identifying genes and pathways in SW1990 cells that areactivated or induced by treatmentwith REG3A or IL6. Based on compar-isons of results obtained bymicroarray, KEGG pathway enrichment, andGO function analyses, we confirmed the involvement of eight key genes,as well as four specific signal pathways, in the induction of tumorigen-esis by REG3A in the SW1990 cell model of pancreatic cancer.
2. Methods and materials
2.1. Cell lines and reagents
Human pancreatic cancer cell lines SW1990 and PANC-1 wereobtained from American Type Culture Collection (ATCC, USA). The non-cancerous cell lineHPDE6c7, purchased fromGuangZhou Jennio BiotechCo., Ltd. was used as a normal pancreas cell line. The cell lines were cul-tivated in RPMI 1640 (Hyclone, USA) supplemented with 10% fetal bo-vine serum (FBS) (Invitrogen, USA) and 100 units/mL penicillin/streptomycin (Invitrogen, USA) at 37 °C under a 5% CO2 environment.HPDE6c7 cells and SW1990 cells were incubated with 100 ng/mL IL6protein (SAB, USA) or 50 ng/mL REG3A protein (Sino Biological Inc.,China), respectively, for 24 h with the control group disposed with thesame volume of phosphate buffered saline (PBS) (Wang et al., 2014).Afterwards, total cellular RNA was extracted with TRIzol reagent (LifeTechnologies, USA) in accordance with the manufacturer's instructions.RNA quality was evaluated by ultraviolet spectrophotometer, Agilent2200 Bioanalyzer (Agilent, USA), and agarose gel electrophoresis(A260/A280 ≥ 1.8, A260/A230 ≥ 1, RIN value ≥ 7). Synthetic antisenseRNA was generated in the condition of Amino Allyl message Amp IIKit 9 (Life Technologies, USA), subsequently hybridized to RiboArray™Custom Array 1 × 2 K (RiboBio, China). Cells were treated identicallyfor the microarray and real-time PCR studies described below.
2.2. Microarray data and screening of differentially expression genes
Our microarray analysis targeted 444 genes which have beenimplicated in pancreatic cancer on a genome-wide scale. Data withthe coefficient of variation [CV = SD (standard deviation)/mean
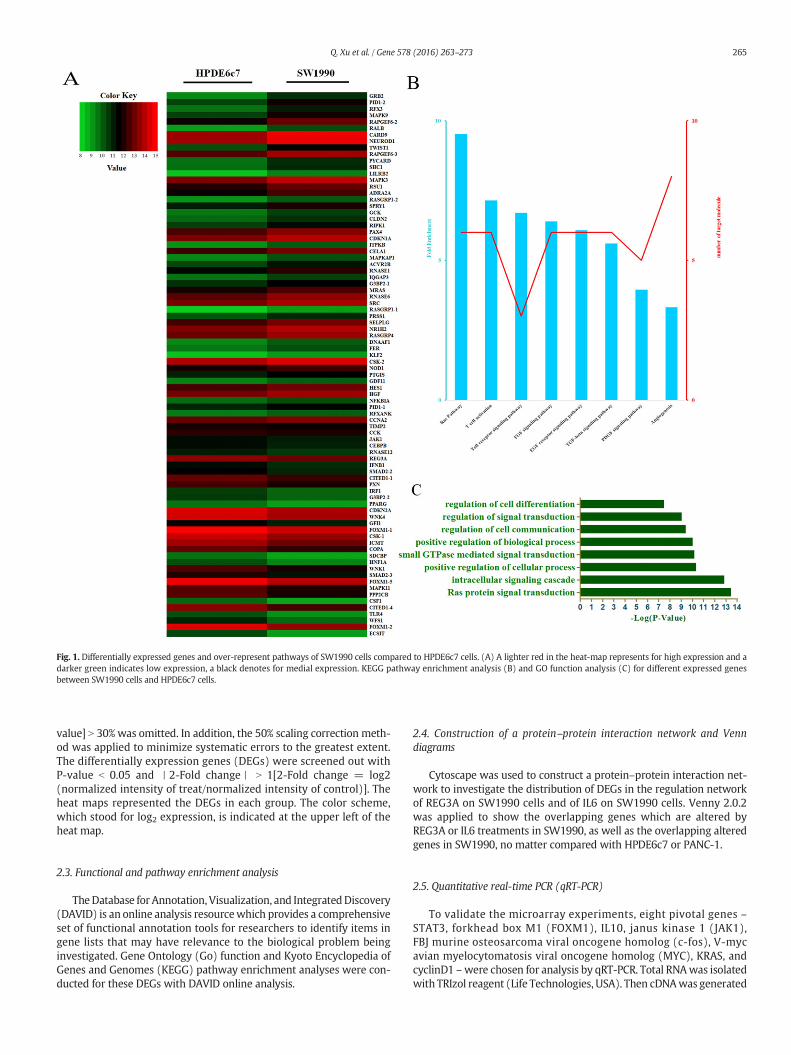
Fig. 1. Differentially expressed genes and over-represent pathways of SW1990 cells compared to HPDE6c7 cells. (A) A lighter red in the heat-map represents for high expression and adarker green indicates low expression, a black denotes for medial expression. KEGG pathway enrichment analysis (B) and GO function analysis (C) for different expressed genesbetween SW1990 cells and HPDE6c7 cells.
265Q. Xu et al. / Gene 578 (2016) 263–273
value] N 30% was omitted. In addition, the 50% scaling correction meth-od was applied to minimize systematic errors to the greatest extent.The differentially expression genes (DEGs) were screened out withP-value b 0.05 and 丨2-Fold change丨 N 1[2-Fold change = log2(normalized intensity of treat/normalized intensity of control)]. Theheat maps represented the DEGs in each group. The color scheme,which stood for log2 expression, is indicated at the upper left of theheat map.
2.3. Functional and pathway enrichment analysis
TheDatabase for Annotation, Visualization, and IntegratedDiscovery(DAVID) is an online analysis resourcewhich provides a comprehensiveset of functional annotation tools for researchers to identify items ingene lists that may have relevance to the biological problem beinginvestigated. Gene Ontology (Go) function and Kyoto Encyclopedia ofGenes and Genomes (KEGG) pathway enrichment analyses were con-ducted for these DEGs with DAVID online analysis.
2.4. Construction of a protein–protein interaction network and Venndiagrams
Cytoscape was used to construct a protein–protein interaction net-work to investigate the distribution of DEGs in the regulation networkof REG3A on SW1990 cells and of IL6 on SW1990 cells. Venny 2.0.2was applied to show the overlapping genes which are altered byREG3A or IL6 treatments in SW1990, as well as the overlapping alteredgenes in SW1990, no matter compared with HPDE6c7 or PANC-1.
2.5. Quantitative real-time PCR (qRT-PCR)
To validate the microarray experiments, eight pivotal genes –STAT3, forkhead box M1 (FOXM1), IL10, janus kinase 1 (JAK1),FBJ murine osteosarcoma viral oncogene homolog (c-fos), V-mycavian myelocytomatosis viral oncogene homolog (MYC), KRAS, andcyclinD1 –were chosen for analysis by qRT-PCR. Total RNAwas isolatedwith TRIzol reagent (Life Technologies, USA). Then cDNAwas generated
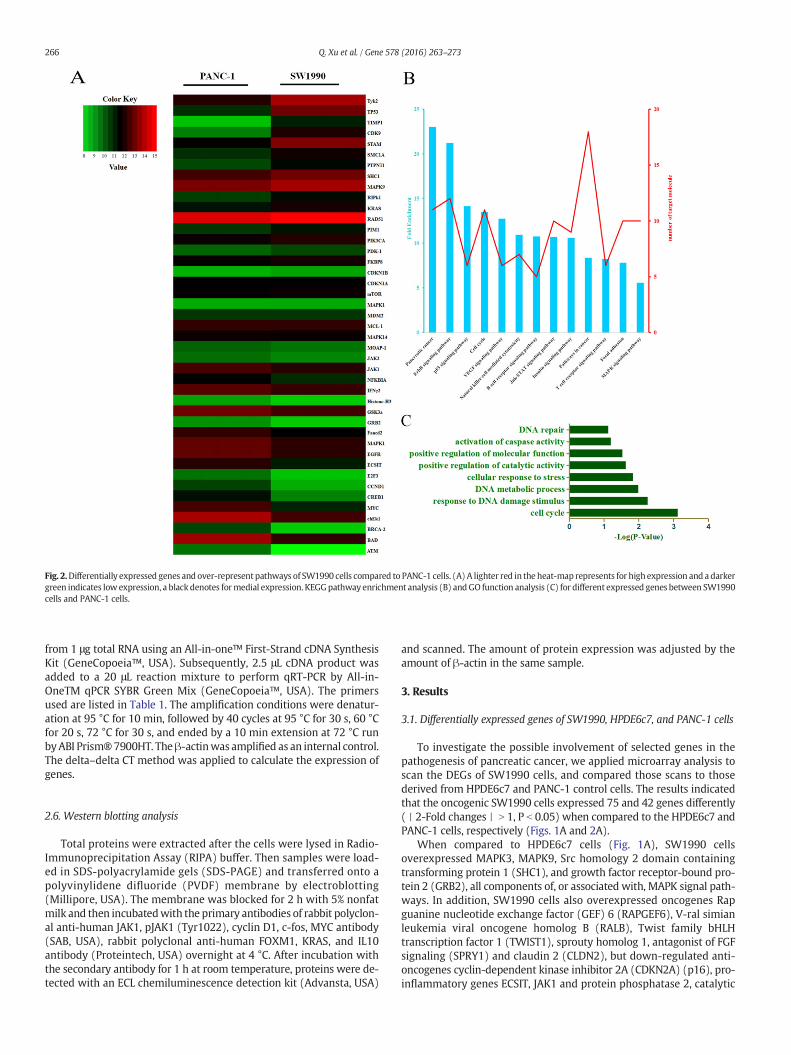
Fig. 2.Differentially expressed genes and over-represent pathways of SW1990 cells compared to PANC-1 cells. (A) A lighter red in the heat-map represents for high expression and a darkergreen indicates low expression, a black denotes formedial expression. KEGGpathway enrichment analysis (B) andGO function analysis (C) for different expressed genes between SW1990cells and PANC-1 cells.
266 Q. Xu et al. / Gene 578 (2016) 263–273
from 1 μg total RNA using an All-in-one™ First-Strand cDNA SynthesisKit (GeneCopoeia™, USA). Subsequently, 2.5 μL cDNA product wasadded to a 20 μL reaction mixture to perform qRT-PCR by All-in-OneTM qPCR SYBR Green Mix (GeneCopoeia™, USA). The primersused are listed in Table 1. The amplification conditions were denatur-ation at 95 °C for 10 min, followed by 40 cycles at 95 °C for 30 s, 60 °Cfor 20 s, 72 °C for 30 s, and ended by a 10 min extension at 72 °C runbyABI Prism®7900HT. Theβ-actinwas amplified as an internal control.The delta–delta CT method was applied to calculate the expression ofgenes.
2.6. Western blotting analysis
Total proteins were extracted after the cells were lysed in Radio-Immunoprecipitation Assay (RIPA) buffer. Then samples were load-ed in SDS-polyacrylamide gels (SDS-PAGE) and transferred onto apolyvinylidene difluoride (PVDF) membrane by electroblotting(Millipore, USA). The membrane was blocked for 2 h with 5% nonfatmilk and then incubatedwith the primary antibodies of rabbit polyclon-al anti-human JAK1, pJAK1 (Tyr1022), cyclin D1, c-fos, MYC antibody(SAB, USA), rabbit polyclonal anti-human FOXM1, KRAS, and IL10antibody (Proteintech, USA) overnight at 4 °C. After incubation withthe secondary antibody for 1 h at room temperature, proteins were de-tected with an ECL chemiluminescence detection kit (Advansta, USA)
and scanned. The amount of protein expression was adjusted by theamount of β-actin in the same sample.
3. Results
3.1. Differentially expressed genes of SW1990, HPDE6c7, and PANC-1 cells
To investigate the possible involvement of selected genes in thepathogenesis of pancreatic cancer, we applied microarray analysis toscan the DEGs of SW1990 cells, and compared those scans to thosederived from HPDE6c7 and PANC-1 control cells. The results indicatedthat the oncogenic SW1990 cells expressed 75 and 42 genes differently(丨2-Fold changes丨 N 1, P b 0.05) when compared to the HPDE6c7 andPANC-1 cells, respectively (Figs. 1A and 2A).
When compared to HPDE6c7 cells (Fig. 1A), SW1990 cellsoverexpressed MAPK3, MAPK9, Src homology 2 domain containingtransforming protein 1 (SHC1), and growth factor receptor-bound pro-tein 2 (GRB2), all components of, or associated with, MAPK signal path-ways. In addition, SW1990 cells also overexpressed oncogenes Rapguanine nucleotide exchange factor (GEF) 6 (RAPGEF6), V-ral simianleukemia viral oncogene homolog B (RALB), Twist family bHLHtranscription factor 1 (TWIST1), sprouty homolog 1, antagonist of FGFsignaling (SPRY1) and claudin 2 (CLDN2), but down-regulated anti-oncogenes cyclin-dependent kinase inhibitor 2A (CDKN2A) (p16), pro-inflammatory genes ECSIT, JAK1 and protein phosphatase 2, catalytic
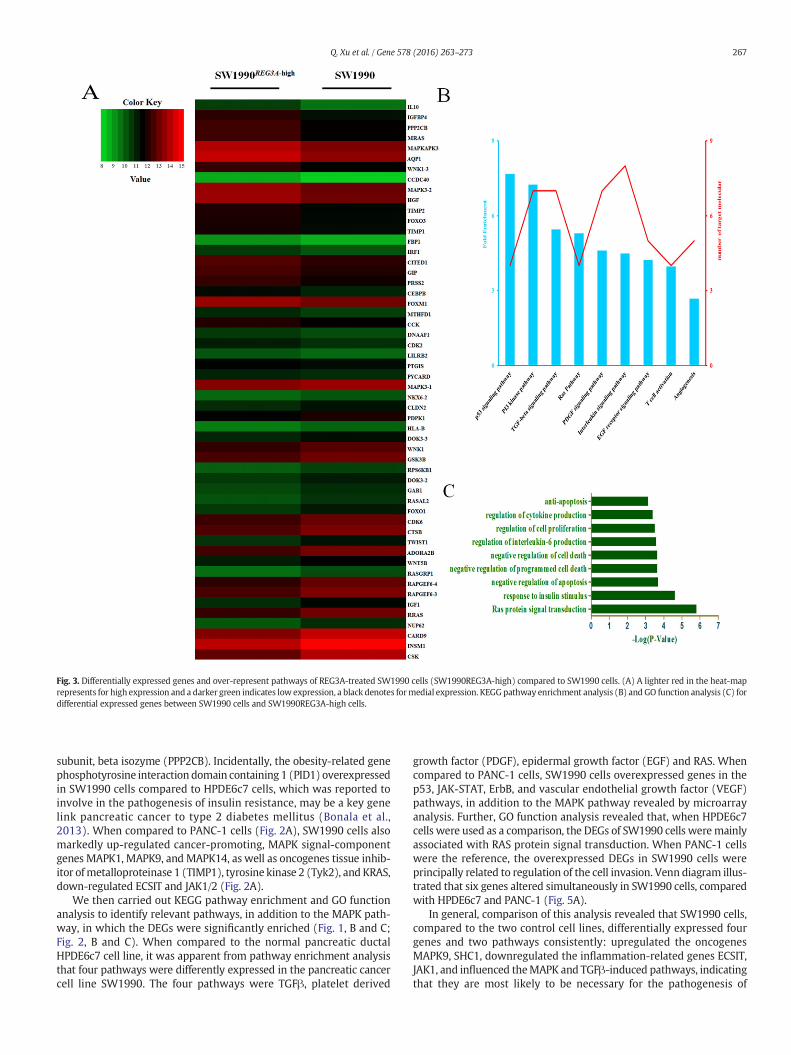
Fig. 3. Differentially expressed genes and over-represent pathways of REG3A-treated SW1990 cells (SW1990REG3A-high) compared to SW1990 cells. (A) A lighter red in the heat-maprepresents for high expression and a darker green indicates low expression, a black denotes formedial expression. KEGGpathway enrichment analysis (B) and GO function analysis (C) fordifferential expressed genes between SW1990 cells and SW1990REG3A-high cells.
267Q. Xu et al. / Gene 578 (2016) 263–273
subunit, beta isozyme (PPP2CB). Incidentally, the obesity-related genephosphotyrosine interaction domain containing 1 (PID1) overexpressedin SW1990 cells compared to HPDE6c7 cells, which was reported toinvolve in the pathogenesis of insulin resistance, may be a key genelink pancreatic cancer to type 2 diabetes mellitus (Bonala et al.,2013). When compared to PANC-1 cells (Fig. 2A), SW1990 cells alsomarkedly up-regulated cancer-promoting, MAPK signal-componentgenes MAPK1, MAPK9, and MAPK14, as well as oncogenes tissue inhib-itor ofmetalloproteinase 1 (TIMP1), tyrosine kinase 2 (Tyk2), and KRAS,down-regulated ECSIT and JAK1/2 (Fig. 2A).
We then carried out KEGG pathway enrichment and GO functionanalysis to identify relevant pathways, in addition to the MAPK path-way, in which the DEGs were significantly enriched (Fig. 1, B and C;Fig. 2, B and C). When compared to the normal pancreatic ductalHPDE6c7 cell line, it was apparent from pathway enrichment analysisthat four pathways were differently expressed in the pancreatic cancercell line SW1990. The four pathways were TGFβ, platelet derived
growth factor (PDGF), epidermal growth factor (EGF) and RAS. Whencompared to PANC-1 cells, SW1990 cells overexpressed genes in thep53, JAK-STAT, ErbB, and vascular endothelial growth factor (VEGF)pathways, in addition to the MAPK pathway revealed by microarrayanalysis. Further, GO function analysis revealed that, when HPDE6c7cells were used as a comparison, the DEGs of SW1990 cells weremainlyassociated with RAS protein signal transduction. When PANC-1 cellswere the reference, the overexpressed DEGs in SW1990 cells wereprincipally related to regulation of the cell invasion. Venn diagram illus-trated that six genes altered simultaneously in SW1990 cells, comparedwith HPDE6c7 and PANC-1 (Fig. 5A).
In general, comparison of this analysis revealed that SW1990 cells,compared to the two control cell lines, differentially expressed fourgenes and two pathways consistently: upregulated the oncogenesMAPK9, SHC1, downregulated the inflammation-related genes ECSIT,JAK1, and influenced theMAPK and TGFβ-induced pathways, indicatingthat they are most likely to be necessary for the pathogenesis of
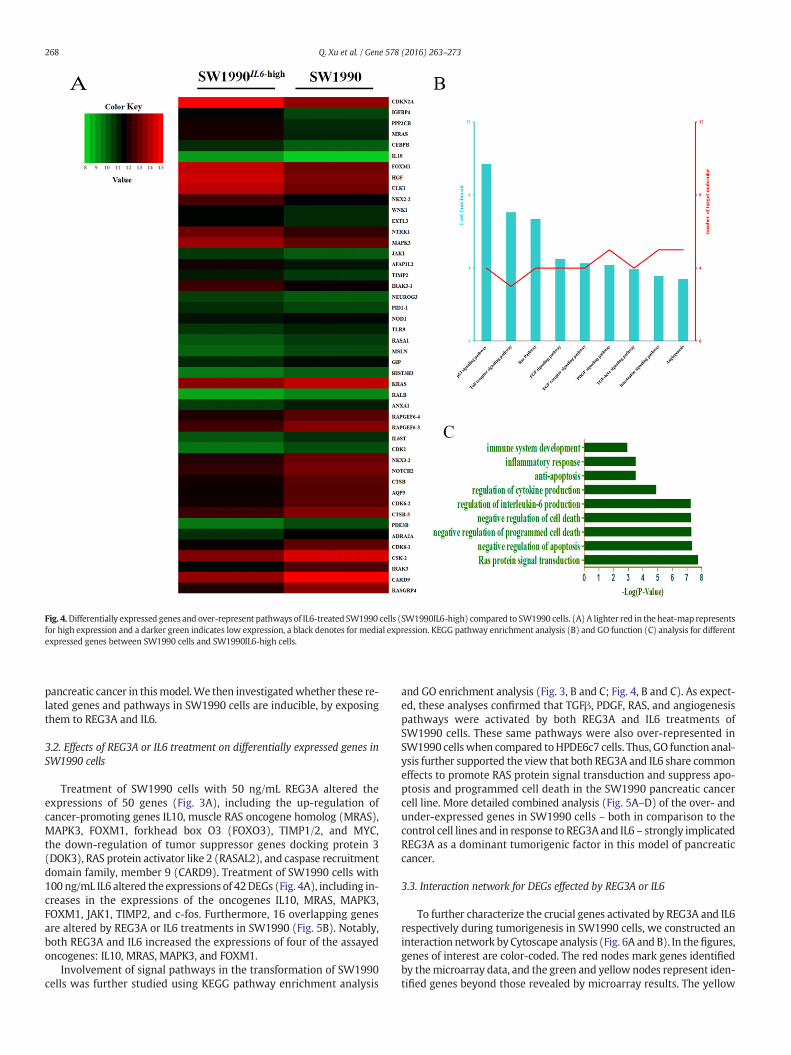
Fig. 4.Differentially expressed genes and over-represent pathways of IL6-treated SW1990 cells (SW1990IL6-high) compared to SW1990 cells. (A) A lighter red in the heat-map representsfor high expression and a darker green indicates low expression, a black denotes for medial expression. KEGG pathway enrichment analysis (B) and GO function (C) analysis for differentexpressed genes between SW1990 cells and SW1990IL6-high cells.
268 Q. Xu et al. / Gene 578 (2016) 263–273
pancreatic cancer in thismodel.We then investigatedwhether these re-lated genes and pathways in SW1990 cells are inducible, by exposingthem to REG3A and IL6.
3.2. Effects of REG3A or IL6 treatment on differentially expressed genes inSW1990 cells
Treatment of SW1990 cells with 50 ng/mL REG3A altered theexpressions of 50 genes (Fig. 3A), including the up-regulation ofcancer-promoting genes IL10, muscle RAS oncogene homolog (MRAS),MAPK3, FOXM1, forkhead box O3 (FOXO3), TIMP1/2, and MYC,the down-regulation of tumor suppressor genes docking protein 3(DOK3), RAS protein activator like 2 (RASAL2), and caspase recruitmentdomain family, member 9 (CARD9). Treatment of SW1990 cells with100 ng/mL IL6 altered the expressions of 42DEGs (Fig. 4A), including in-creases in the expressions of the oncogenes IL10, MRAS, MAPK3,FOXM1, JAK1, TIMP2, and c-fos. Furthermore, 16 overlapping genesare altered by REG3A or IL6 treatments in SW1990 (Fig. 5B). Notably,both REG3A and IL6 increased the expressions of four of the assayedoncogenes: IL10, MRAS, MAPK3, and FOXM1.
Involvement of signal pathways in the transformation of SW1990cells was further studied using KEGG pathway enrichment analysis
and GO enrichment analysis (Fig. 3, B and C; Fig. 4, B and C). As expect-ed, these analyses confirmed that TGFβ, PDGF, RAS, and angiogenesispathways were activated by both REG3A and IL6 treatments ofSW1990 cells. These same pathways were also over-represented inSW1990 cellswhen compared toHPDE6c7 cells. Thus, GO function anal-ysis further supported the view that both REG3A and IL6 share commoneffects to promote RAS protein signal transduction and suppress apo-ptosis and programmed cell death in the SW1990 pancreatic cancercell line. More detailed combined analysis (Fig. 5A–D) of the over- andunder-expressed genes in SW1990 cells – both in comparison to thecontrol cell lines and in response to REG3A and IL6 – strongly implicatedREG3A as a dominant tumorigenic factor in this model of pancreaticcancer.
3.3. Interaction network for DEGs effected by REG3A or IL6
To further characterize the crucial genes activated by REG3A and IL6respectively during tumorigenesis in SW1990 cells, we constructed aninteraction network by Cytoscape analysis (Fig. 6A and B). In thefigures,genes of interest are color-coded. The red nodes mark genes identifiedby themicroarray data, and the green and yellow nodes represent iden-tified genes beyond those revealed by microarray results. The yellow

Fig. 5. Venn diagrams illustrating the number of DEGs in the overlapping genes in SW1990 compared with HPDE6c7 or PANC-1 (A), as well as the overlapping genes which are altered byREG3A or IL6 treatments in SW1990 (B).
269Q. Xu et al. / Gene 578 (2016) 263–273
nodes also indicate commongenes activated by REG3A or IL6 treatment.The number and length of the lines signify the interaction frequency andmagnitude among these nodes. The smaller the distance between thenodes and the center, the stronger the influences those nodes have onthe networks of REG3A. The results of this pathway analysis indicatethat REG3A and IL6 regulate similar transcription factor pathways,most prominently the paired box (PAX), BTB and CNC homology 1,basic leucine zipper transcription factor (BACH), and GATAbinding pro-tein (globin transcription factor, GATA) families (Fig. 7).
3.4. Verification of vital genes by qRT-PCR
With the purpose of supporting or refuting the results of microarrayanalysis, we selected eight pivotal genes to determine their transcrip-tion levels by qRT-PCR (Fig. 8A–H). The results uniformly supportedthe microarray findings that had implicated specific involvement ofthe JAK1, STAT3, IL10, FOXM1, KRAS, MYC, CyclinD1 and c-fos genes.By qRT-PCR analysis, all eight genes were up-regulated by REG3A treat-ment of SW1990 cells, while five of them, JAK1, KRAS, MYC, CyclinD1and c-fos, were up-regulated by REG3A treatment of PANC-1 cells. Asexpected, the twomethods were also largely in agreementwith respectto identifying genes that were variably affected by REG3A treatment.
3.5. Verification of vital genes by Western blotting
Among the target genes, some undergo posttranslational modifica-tions (PTMs). It is not clear whether these genes experienced differ-ences in PTMs upon REG3A or IL6 stimulation. Hence Western blotting
was conducted to detect the protein expression of pJAK1 (Tyr1002),JAK1, KRAS, MYC, FOXM1, CyclinD1, c-fos and IL10 in SW1990 uponREG3A or IL6 treatments. We previously reported that both REG3Aand IL6 could up-regulate the expression of pSTAT3 (Ser727), pSTAT3(Tyr705), and STAT3. It met our expectation that REG3A as well as IL6can enhance the expression of those eight proteins, in accordancewith the microarray data (Fig. 8I).
4. Discussion
Previous work has shown that mutation in multiple genes andspecific signal pathways contribute to the initiation and progression ofpancreatic tumors (Yu et al., 2010; Oshima et al., 2013; Xie et al.,2015). Our results identified specific differentially expressed genesand pathways in the metastatic pancreatic cell line SW1990 that werenot expressed in the non-cancerous HPDE6c7 or primary pancreaticcell line PANC-1. Microarray, KEGG pathway, and GO function analysesimplicated the overexpression of oncogenes, including GRB2, MAPK9,SHC1, and KRAS, as well as activation of oncogenic pathways includingMAPK and TGFβ-activated pathways in SW1990 cells. These findingsare largely consistent with previous clinical studies (Leahy et al., 2004;Chang et al., 2014). Human cancers, including pancreatic cancers, havealso been shown to display upregulation of genes in the MAPK families– such as Tyk2, MAPK1, MAPK3, and MAPK9 – along with activation ofJAK/STAT and MAPK signaling pathways (Wagner and Nebreda, 2009;Nam et al., 2013). Overexpression of the KRAS oncogene elicits itsprotumor function at least in part through the activation of MAPK-dependent pathways (Wee et al., 2009). Moreover, the persistent
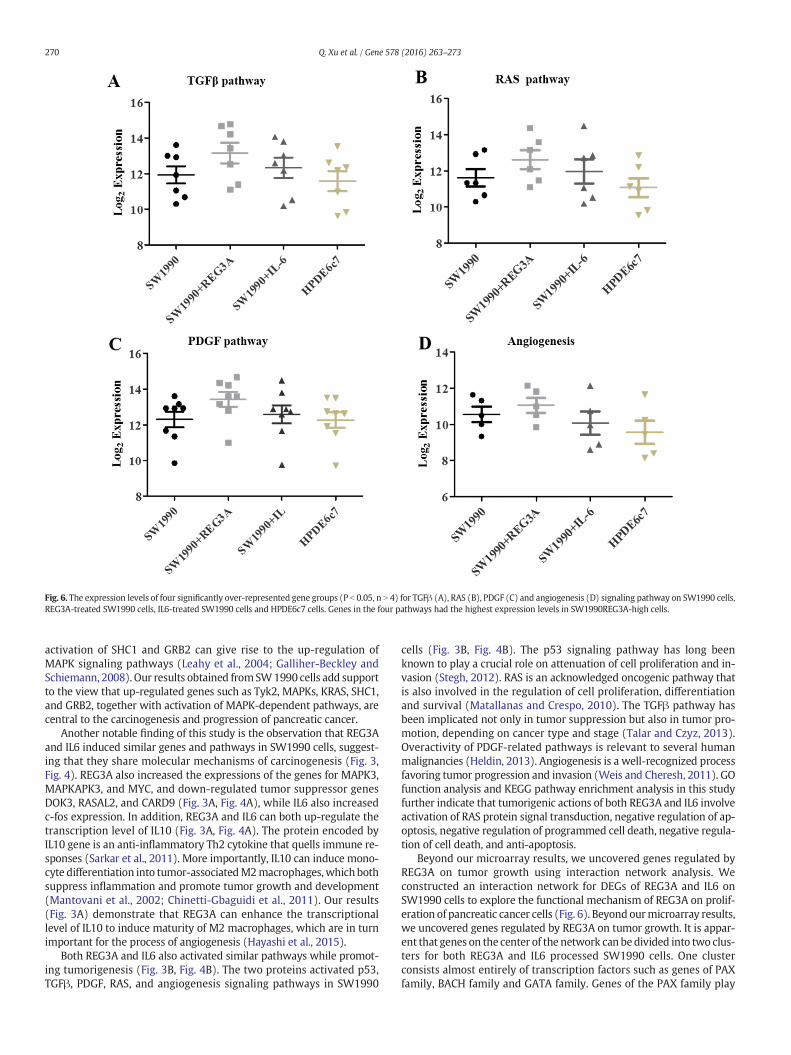
Fig. 6. The expression levels of four significantly over-represented gene groups (P b 0.05, n N 4) for TGFβ (A), RAS (B), PDGF (C) and angiogenesis (D) signaling pathway on SW1990 cells,REG3A-treated SW1990 cells, IL6-treated SW1990 cells and HPDE6c7 cells. Genes in the four pathways had the highest expression levels in SW1990REG3A-high cells.
270 Q. Xu et al. / Gene 578 (2016) 263–273
activation of SHC1 and GRB2 can give rise to the up-regulation ofMAPK signaling pathways (Leahy et al., 2004; Galliher-Beckley andSchiemann, 2008). Our results obtained fromSW1990 cells add supportto the view that up-regulated genes such as Tyk2, MAPKs, KRAS, SHC1,and GRB2, together with activation of MAPK-dependent pathways, arecentral to the carcinogenesis and progression of pancreatic cancer.
Another notable finding of this study is the observation that REG3Aand IL6 induced similar genes and pathways in SW1990 cells, suggest-ing that they share molecular mechanisms of carcinogenesis (Fig. 3,Fig. 4). REG3A also increased the expressions of the genes for MAPK3,MAPKAPK3, and MYC, and down-regulated tumor suppressor genesDOK3, RASAL2, and CARD9 (Fig. 3A, Fig. 4A), while IL6 also increasedc-fos expression. In addition, REG3A and IL6 can both up-regulate thetranscription level of IL10 (Fig. 3A, Fig. 4A). The protein encoded byIL10 gene is an anti-inflammatory Th2 cytokine that quells immune re-sponses (Sarkar et al., 2011). More importantly, IL10 can induce mono-cyte differentiation into tumor-associatedM2macrophages,which bothsuppress inflammation and promote tumor growth and development(Mantovani et al., 2002; Chinetti-Gbaguidi et al., 2011). Our results(Fig. 3A) demonstrate that REG3A can enhance the transcriptionallevel of IL10 to induce maturity of M2 macrophages, which are in turnimportant for the process of angiogenesis (Hayashi et al., 2015).
Both REG3A and IL6 also activated similar pathways while promot-ing tumorigenesis (Fig. 3B, Fig. 4B). The two proteins activated p53,TGFβ, PDGF, RAS, and angiogenesis signaling pathways in SW1990
cells (Fig. 3B, Fig. 4B). The p53 signaling pathway has long beenknown to play a crucial role on attenuation of cell proliferation and in-vasion (Stegh, 2012). RAS is an acknowledged oncogenic pathway thatis also involved in the regulation of cell proliferation, differentiationand survival (Matallanas and Crespo, 2010). The TGFβ pathway hasbeen implicated not only in tumor suppression but also in tumor pro-motion, depending on cancer type and stage (Talar and Czyz, 2013).Overactivity of PDGF-related pathways is relevant to several humanmalignancies (Heldin, 2013). Angiogenesis is a well-recognized processfavoring tumor progression and invasion (Weis and Cheresh, 2011). GOfunction analysis and KEGG pathway enrichment analysis in this studyfurther indicate that tumorigenic actions of both REG3A and IL6 involveactivation of RAS protein signal transduction, negative regulation of ap-optosis, negative regulation of programmed cell death, negative regula-tion of cell death, and anti-apoptosis.
Beyond our microarray results, we uncovered genes regulated byREG3A on tumor growth using interaction network analysis. Weconstructed an interaction network for DEGs of REG3A and IL6 onSW1990 cells to explore the functional mechanism of REG3A on prolif-eration of pancreatic cancer cells (Fig. 6). Beyond ourmicroarray results,we uncovered genes regulated by REG3A on tumor growth. It is appar-ent that genes on the center of the network can bedivided into two clus-ters for both REG3A and IL6 processed SW1990 cells. One clusterconsists almost entirely of transcription factors such as genes of PAXfamily, BACH family and GATA family. Genes of the PAX family play

Fig. 7. Interaction network for DEGs of REG3A (A) or IL6 (B) treated SW1990 cells. The red nodes stand for genes in our microarray data, the green and yellow nodes represent for genesbeyond our microarray results. The yellow nodes are identical genes between REG3A and IL6 treated SW1990 cells, while the green nodes represent for different genes.
271Q. Xu et al. / Gene 578 (2016) 263–273
critical roles during fetal development and cancer growth (Frost et al.,2008). PAX4 participates in the growth of pancreas islet and in the dif-ferentiation of islet β cells (SosaPineda et al., 1997). BACH1 is closely re-lated structurally to BACH2 (Itoh-Nakadai et al., 2014a). BACH 1 andBACH2 interact to regulate B cell development (Itoh-Nakadai et al.,2014b). In light of results of this study, we can speculate that REG3Amay interact with BACH2 to regulate both humoral and cellular immu-nity. GATA1 and GATA2 are transcription factors which play a pivotalrole during erythropoiesis (Moriguchi and Yamamoto, 2014). In addi-tion, both are indispensable to the activation and development ofmast cell (Masuda et al., 2007; Ohmori et al., 2012).Mast cells arewide-ly regarded as pivotal effector cells in allergic disorders, and they canalso be important as initiators and effective promoters of innate immu-nity (Galli et al., 2005). In addition, REG3A apparently regulates thetranscription factor STAT5A, which possesses high sequence homologywith STAT3, whose expression is impacted by IL6 (Fig. 6). The othercluster revealed by interaction network analysis includes genes relatedto tumor growth such as TP53, c-fos, c-jun. c-fos and c-jun bond todipolymer called AP-1, a key regulator of major biological events suchas cell proliferation and apoptosis death (Salvat et al., 1999). Besides,
REG3A can regulate transcription factor STAT5Awhich possesses highlyhomology with the STAT3 impacted by IL6 (Fig. 6). Thus we speculatethat REG3A and IL6may be important regulators of both tumor develop-ment and immunity.
In summary, results presented here implicate at least eight specificgenes and four pathways involved in the transformation of the pancre-atic cancer SW1990 cell line. They also show that REG3A cannot onlyimmunoregulate through activation of transcription factors BACH,GATA and STAT families and Th2 cytokines – the last implicating thepromotion of angiogenesis – but can also alter p53 signaling and PDGFand RAS pathways to promote tumor growth. In a number of respects,actions of REG3A on SW1990 cells are similar to those of IL6. The regu-lation network constructed here for REG3A made it possible for us toidentify hub nodes for REG3A-mediated tumor growth. Applications ofthis methodology may prove to be useful in future efforts to find newtargets for treatment of pancreatic cancer.
Conflict of interest
The authors declare no conflict of interest.
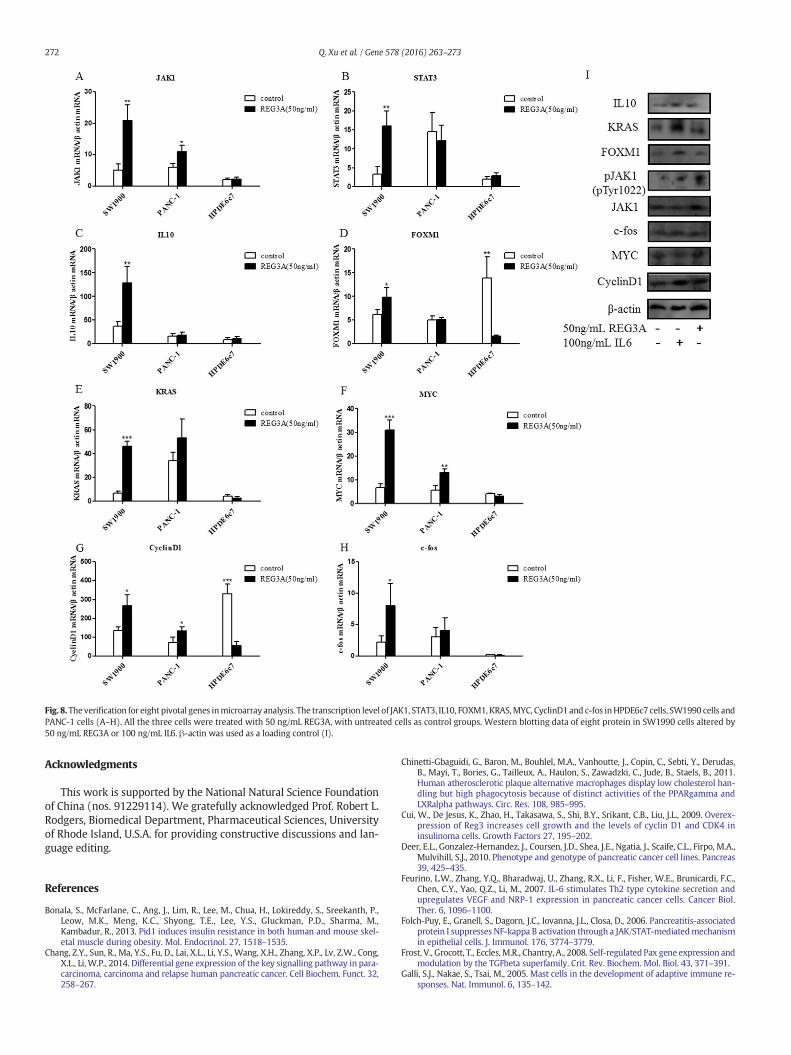
Fig. 8.Theverification for eight pivotal genes inmicroarray analysis. The transcription level of JAK1, STAT3, IL10, FOXM1, KRAS,MYC, CyclinD1and c-fos inHPDE6c7 cells, SW1990 cells andPANC-1 cells (A–H). All the three cells were treated with 50 ng/mL REG3A, with untreated cells as control groups. Western blotting data of eight protein in SW1990 cells altered by50 ng/mL REG3A or 100 ng/mL IL6. β-actin was used as a loading control (I).
272 Q. Xu et al. / Gene 578 (2016) 263–273
Acknowledgments
This work is supported by the National Natural Science Foundationof China (nos. 91229114). We gratefully acknowledged Prof. Robert L.Rodgers, Biomedical Department, Pharmaceutical Sciences, Universityof Rhode Island, U.S.A. for providing constructive discussions and lan-guage editing.
References
Bonala, S., McFarlane, C., Ang, J., Lim, R., Lee, M., Chua, H., Lokireddy, S., Sreekanth, P.,Leow, M.K., Meng, K.C., Shyong, T.E., Lee, Y.S., Gluckman, P.D., Sharma, M.,Kambadur, R., 2013. Pid1 induces insulin resistance in both human and mouse skel-etal muscle during obesity. Mol. Endocrinol. 27, 1518–1535.
Chang, Z.Y., Sun, R., Ma, Y.S., Fu, D., Lai, X.L., Li, Y.S., Wang, X.H., Zhang, X.P., Lv, Z.W., Cong,X.L., Li, W.P., 2014. Differential gene expression of the key signalling pathway in para-carcinoma, carcinoma and relapse human pancreatic cancer. Cell Biochem. Funct. 32,258–267.
Chinetti-Gbaguidi, G., Baron, M., Bouhlel, M.A., Vanhoutte, J., Copin, C., Sebti, Y., Derudas,B., Mayi, T., Bories, G., Tailleux, A., Haulon, S., Zawadzki, C., Jude, B., Staels, B., 2011.Human atherosclerotic plaque alternative macrophages display low cholesterol han-dling but high phagocytosis because of distinct activities of the PPARgamma andLXRalpha pathways. Circ. Res. 108, 985–995.
Cui, W., De Jesus, K., Zhao, H., Takasawa, S., Shi, B.Y., Srikant, C.B., Liu, J.L., 2009. Overex-pression of Reg3 increases cell growth and the levels of cyclin D1 and CDK4 ininsulinoma cells. Growth Factors 27, 195–202.
Deer, E.L., Gonzalez-Hernandez, J., Coursen, J.D., Shea, J.E., Ngatia, J., Scaife, C.L., Firpo, M.A.,Mulvihill, S.J., 2010. Phenotype and genotype of pancreatic cancer cell lines. Pancreas39, 425–435.
Feurino, L.W., Zhang, Y.Q., Bharadwaj, U., Zhang, R.X., Li, F., Fisher, W.E., Brunicardi, F.C.,Chen, C.Y., Yao, Q.Z., Li, M., 2007. IL-6 stimulates Th2 type cytokine secretion andupregulates VEGF and NRP-1 expression in pancreatic cancer cells. Cancer Biol.Ther. 6, 1096–1100.
Folch-Puy, E., Granell, S., Dagorn, J.C., Iovanna, J.L., Closa, D., 2006. Pancreatitis-associatedprotein I suppresses NF-kappa B activation through a JAK/STAT-mediatedmechanismin epithelial cells. J. Immunol. 176, 3774–3779.
Frost, V., Grocott, T., Eccles,M.R., Chantry, A., 2008. Self-regulated Pax gene expression andmodulation by the TGFbeta superfamily. Crit. Rev. Biochem. Mol. Biol. 43, 371–391.
Galli, S.J., Nakae, S., Tsai, M., 2005. Mast cells in the development of adaptive immune re-sponses. Nat. Immunol. 6, 135–142.

273Q. Xu et al. / Gene 578 (2016) 263–273
Galliher-Beckley, A.J., Schiemann, W.P., 2008. Grb2 binding to Tyr284 in TbetaR-II isessential for mammary tumor growth and metastasis stimulated by TGF-beta.Carcinogenesis 29, 244–251.
Goumas, F.A., Holmer, R., Egberts, J.H., Gontarewicz, A., Heneweer, C., Geisen, U., Hauser,C., Mende, M.M., Legler, K., Rocken, C., Becker, T., Waetzig, G.H., Rose-John, S.,Kalthoff, H., 2015. Inhibition of IL-6 signaling significantly reduces primary tumorgrowth and recurrencies in orthotopic xenograft models of pancreatic cancer. Int.J. Cancer.
Hayashi, N., Kataoka, H., Yano, S., Tanaka, M., Moriwaki, K., Akashi, H., Suzuki, S., Mori, Y.,Kubota, E., Tanida, S., Takahashi, S., Joh, T., 2015. A novel photodynamic therapytargeting cancer cells and tumor-associated macrophages. Mol. Cancer Ther. 14,452–460.
Heilmann, A.M., Perera, R.M., Ecker, V., Nicolay, B.N., Bardeesy, N., Benes, C.H., Dyson, N.J.,2014. CDK4/6 and IGF1 receptor inhibitors synergize to suppress the growth ofp16(INK4A)-deficient pancreatic cancers. Cancer Res. 74, 3947–3958.
Heldin, C.H., 2013. Targeting the PDGF signaling pathway in tumor treatment. Cell Com-munication And Signaling 11.
Hidalgo, M., 2010. Pancreatic cancer. N. Engl. J. Med. 362, 1605–1617.Hou, W.R., Xie, S.N., Wang, H.J., Su, Y.Y., Lu, J.L., Li, L.L., Zhang, S.S., Xiang, M., 2011. Intra-
muscular delivery of a naked DNA plasmid encoding proinsulin and pancreaticregenerating III protein ameliorates type 1 diabetes mellitus. Pharmacol. Res. 63,320–327.
Itoh-Nakadai, A., Hikota, R., Muto, A., Kometani, K., Watanabe-Matsui, M., Sato, Y.,Kobayashi, M., Nakamura, A., Miura, Y., Yano, Y., Tashiro, S., Sun, J., Ikawa, T., Ochiai,K., Kurosaki, T., Igarashi, K., 2014a. The transcription repressors Bach2 and Bach1 pro-mote B cell development by repressing the myeloid program. Nat. Immunol. 15,1171–1180.
Itoh-Nakadai, A., Hikota, R., Muto, A., Kometani, K., Watanabe-Matsui, M., Sato, Y.,Kobayashi, M., Nakamura, A., Miura, Y., Yano, Y., Tashiro, S., Sun, J.Y., Ikawa, T.,Ochiai, K., Kurosaki, T., Igarashi, K., 2014b. The transcription repressors Bach2 andBach1 promote B cell development by repressing the myeloid program. Nat.Immunol. 15, 1171–1180.
Ji, Z., Mei, F.C., Xie, J., Cheng, X., 2007. Oncogenic KRAS activates hedgehog signaling path-way in pancreatic cancer cells. J Biol Chem 282, 14048–14055.
Leahy, M., Lyons, A., Krause, D., O'Connor, R., 2004. Impaired Shc, Ras, and MAPK activa-tion but normal Akt activation in FL5.12 cells expressing an insulin-like growth factorI receptor mutated at tyrosines 1250 and 1251. J. Biol. Chem. 279, 18306–18313.
Legendre, O., Sookdeo, A., Foster, D.A., 2014. BxPC3 pancreatic cancer cells express a trun-cated Smad4 protein upon PI3K and mTOR inhibition. Oncol Lett 7, 1165–1168.
Lesina, M., Kurkowski, M.U., Ludes, K., Rose-John, S., Treiber, M., Kloppel, G., Yoshimura,A., Reindl, W., Sipos, B., Akira, S., Schmid, R.M., Algul, H., 2011. Stat3/Socs3 activationby IL-6 transsignaling promotes progression of pancreatic intraepithelial neoplasiaand development of pancreatic cancer. Cancer Cell 19, 456–469.
Liu, X., Wang, J., Wang, H., Yin, G., Liu, Y., Lei, X., Xiang, M., 2015. REG3A acceleratespancreatic cancer cell growth under IL-6-associated inflammatory condition: in-volvement of a REG3A-JAK2/STAT3 positive feedback loop. Cancer Lett. 362, 45–60.
Mantovani, A., Sozzani, S., Locati, M., Allavena, P., Sica, A., 2002. Macrophage polarization:tumor-associated macrophages as a paradigm for polarized M2mononuclear phago-cytes. Trends Immunol. 23, 549–555.
Masuda, A., Hashimoto, K., Yokoi, T., Doi, T., Kodama, T., Kume, H., Ohno, K., Matsuguchi,T., 2007. Essential role of GATA transcriptional factors in the activation of mast cells.J. Immunol. 178, 360–368.
Matallanas, D., Crespo, P., 2010. New druggable targets in the Ras pathway? Curr. Opin.Mol. Ther. 12, 674–683.
Mohamadkhani, A., Naderi, E., Sharafkhah, M., Fazli, H.R., Moradzadeh, M., Pourshams, A.,2013. Detection of TP53 R249 mutation in Iranian patients with pancreatic cancer.J. Oncol. 2013, 738915.
Moriguchi, T., Yamamoto, M., 2014. A regulatory network governing Gata1 and Gata2gene transcription orchestrates erythroid lineage differentiation. Int. J. Hematol.100, 417–424.
Nam, S., Wen,W., Schroeder, A., Herrmann, A., Yu, H., Cheng, X., Merz, K.H., Eisenbrand, G.,Li, H., Yuan, Y.C., Jove, R., 2013. Dual inhibition of Janus and Src family kinases bynovel indirubin derivative blocks constitutively-activated Stat3 signaling associatedwith apoptosis of human pancreatic cancer cells. Mol. Oncol. 7, 369–378.
Ohmori, S., Takai, J., Ishijima, Y., Suzuki, M., Moriguchi, T., Philipsen, S., Yamamoto, M.,Ohneda, K., 2012. Regulation of GATA factor expression is distinct between erythroidand mast cell lineages. Mol. Cell. Biol. 32, 4742–4755.
Okochi, S., Stanek, A., Xu, M., Seiden, L., Mueller, C.M., Ou, P., Iovanna, J.L., Sugiyama, G.,Alfonso, A.E., Huan, C., 2014. Regenerating islet-derived 3α (Reg3α) is required foracinar cell NFκB activation and tissue protection in experimental acute pancreatitis(AP). J. Am. Coll. Surg. 219, S26.
Oshima, M., Okano, K., Muraki, S., Haba, R., Maeba, T., Suzuki, Y., Yachida, S., 2013.Immunohistochemically detected expression of 3 major genes (CDKN2A/p16, TP53,and SMAD4/DPC4) strongly predicts survival in patients with resectable pancreaticcancer. Ann. Surg. 258, 336–346.
Ouyang, H., Mou, L., Luk, C., Liu, N., Karaskova, J., Squire, J., Tsao, M.S., 2000. Immortalhuman pancreatic duct epithelial cell lines with near normal genotype and pheno-type. Am. J. Pathol. 157, 1623–1631.
Salvat, C., Aquaviva, C., Jariel-Encontre, I., Ferrara, P., Pariat, M., Steff, A.M., Carillo, S.,Piechaczyk, M., 1999. Are there multiple proteolytic pathways contributing to c-Fos,c-Jun and p53 protein degradation in vivo? Mol. Biol. Rep. 26, 45–51.
Sarkar, S., Han, J., Sinsimer, K.S., Liao, B., Foster, R.L., Brewer, G., Pestka, S., 2011. RNA-binding protein AUF1 regulates lipopolysaccharide-induced IL10 expression byactivating IkappaB kinase complex in monocytes. Mol. Cell. Biol. 31, 602–615.
Sheng,W.W., Chen, C.P., Dong, M., Zhou, J.P., Liu, Q.F., Dong, Q., Li, F., 2014. Overexpressionof calreticulin contributes to the development and progression of pancreatic cancer.J. Cell. Physiol. 229, 887–897.
Siegel, R.L., Miller, K.D., Jemal, A., 2015. Cancer statistics, 2015. CA Cancer J. Clin. 65, 5–29.SosaPineda, B., Chowdhury, K., Torres, M., Oliver, G., Gruss, P., 1997. The Pax4 gene is es-
sential for differentiation of insulin-producing beta cells in the mammalian pancreas.Nature 386, 399–402.
Stegh, A.H., 2012. Targeting the p53 signaling pathway in cancer therapy— the promises,challenges and perils. Expert Opin. Ther. Targets 16, 67–83.
Talar, B., Czyz, M., 2013. TGF-beta signaling pathways in cancers. Postepy Hig. Med. Dosw.67, 1008–1017.
Wagner, E.F., Nebreda, A.R., 2009. Signal integration by JNK and p38 MAPK pathways incancer development. Nat. Rev. Cancer 9, 537–549.
Wang, J., Zhou, H., Han, Y., Liu, X., Wang, M., Wang, X., Yin, G., Li, X., Xiang, M., 2014.SOCS3 methylation in synergy with Reg3A overexpression promotes cell growth inpancreatic cancer. J Mol Med (Berl) 92, 1257–1269.
Wee, S., Jagani, Z., Xiang, K.X.Q., Loo, A., Dorsch, M., Yao, Y.M., Sellers, W.R., Lengauer, C.,Stegmeier, F., 2009. PI3K pathway activation mediates resistance to MEK inhibitorsin KRAS mutant cancers. Cancer Res. 69, 4286–4293.
Weis, S.M., Cheresh, D.A., 2011. Tumor angiogenesis: molecular pathways and therapeutictargets. Nat. Med. 17, 1359–1370.
Xie, Z., Yin, X., Gong, B., Nie, W., Wu, B., Zhang, X., Huang, J., Zhang, P., Zhou, Z., Li, Z., 2015.Salivary microRNAs show potential as a noninvasive biomarker for detecting resect-able pancreatic cancer. Cancer Prev. Res. (Phila.) 8, 165–173.
Yu, S., Lu, Z., Liu, C., Meng, Y., Ma, Y., Zhao, W., Liu, J., Yu, J., Chen, J., 2010. miRNA-96 sup-presses KRAS and functions as a tumor suppressor gene in pancreatic cancer. CancerRes. 70, 6015–6025.
Zhou, G.X., Ding, X.L., Wu, S.B., Zhang, H.F., Cao, W., Qu, L.S., Zhang, H., 2015. Inhibition of5-lipoxygenase triggers apoptosis in pancreatic cancer cells. Oncol. Rep. 33, 661–668.

本文献由“学霸图书馆-文献云下载”收集自网络,仅供学习交流使用。
学霸图书馆(www.xuebalib.com)是一个“整合众多图书馆数据库资源,
提供一站式文献检索和下载服务”的24 小时在线不限IP
图书馆。
图书馆致力于便利、促进学习与科研,提供最强文献下载服务。
图书馆导航:
图书馆首页 文献云下载 图书馆入口 外文数据库大全 疑难文献辅助工具



![Clustering of Microarray Data Reveals Transcript Patterns · Clustering of Microarray Data Reveals Transcript Patterns Associated with Somatic Embryogenesis in Soybean1[w] Franc¸oise](https://img.pdfslide.us/doc/110x75/5ed2c7ccae2cb511b17808ae/clustering-of-microarray-data-reveals-transcript-clustering-of-microarray-data-reveals.jpg)

























