Embed Size (px)
Citation preview

Limnologica 29 (1999)407424 L I M N O L O G I C A http://www.urbanfiscber.de/j ournals/limno
© by Urban & Fischer Verlag
1) Institut ftir Zoologie der Freien Universitiit Berlin, AG Protozoologie, Berlin, Germany; 2) Institut ftir 0kologie der Friedrich-Schiller-Universit~it Jena, AG Limnologie, Jena, Germany
Periphyton and Sphagnicolous Protists of Dystrophic Bog Lakes (Brandenburg, Germany)
II. Characteristic Species and Trophy of the Lakes
MICHAELA C. STRUDER-KYPKE 1) • WILFRIED SCHONBORN 2)
With 8 Figures and one Table
Key words: Bog lakes; artificial substrates; ciliates; flagellates; testate amoebae; eutrophication
Abstract
Glass-slides were exposed in tour different bog lakes located in great forest areas in Northern Germany, about 100 km north of Berlin, in order to examine the protist community of these dystrophic, acid lakes. Species composition and succession of the periphyton were studied during one year. The four investigated bog lakes exhibit simi- lar species compositions, most of the species were present in each of the four lakes. The periphyton was composed of a great variety of pro- tist species. Specimens of all major flagellated protists and of most of the ciliate taxa were found on the slides. Many of these species are ubiquitous. Some species, however, like the flagellates Spongomonas sacculus, Rhipidodendron huxIeyi, Cyathobodo sp., and Pseudoden- dromonas vlkii as well as the ciliates Ophrydium hyaIinum, Lep- topharynx costatus, Uroleptus caudatus, and Platyophrya sphagni are found more frequently in acid waters. A regular and common occur- rence of these species partly characterizes the acid Sphagnum bogs. Besides that, common species like Bodo spp., Cryptomonas spp., Go- niomonas truncata, Chilodonella uncinata, Vorticella sp., and Cyr- tolophosis mucicola are represented in the periphyton. Examinations of the testate amoebae in the Sphagnum mats show a very different species composition compared to earlier studies.
The investigated bog lakes revealed mesotrophic conditions. In the upper regions of the Sphagnum mats and in the center of the larg- er bog lakes, oligotrophic conditions still prevail. The lower regions of the Sphagnum mat and the smaller lakes, however, are influenced by the partly degradation of plants and the run-off water from sur- rounding land. In comparison to earlier studies, the species composi- tion indicates a beginning eutrophication of the bog lakes. A direct anthropogenic impact can be excluded.
Introduction
The bog lakes were formed by the melting of ice blocks which were broken from the withdrawing glaciers of the
Weichselian stage of the last glaciation and deposited in the glacial drift or outwash plain. Those ice blocks often took several hundred years to melt completely, finally resulting in the formation of kettle lakes (Fig. 1). About 11,000 years ago, the paludification began and mesotrophic-subneutral circumstances prevailed. This process stopped approximate- ly 9,000 years ago. The ice blocks were melted, the climate changed and it became dry and warmer. Approximately 4,000-2,000 years ago, a secondary turf layer started to grow on these paludificated lakes. Water was supplied only by precipitation and the run-off from surrounding land (CASPER & SCHEFFLER 1990; SUCCOW 1987; SUCCOW & JESCHKE 1990).
The larger bogs possess a residual lake which shows al- most no further terrestrialization, but reveals the excess water of the bog. Its edges are surrounded by Sphagnum mats that regularly tear off. In the center, kettle hole bogs are oligotrophic. The bog is surrounded by laggs (small water- filled ditches) which collect the run-off water and are more mesotrophic (StJccow & JESCHKE 1990).
With exception of the testate amoebae, protists of bogs have always been neglegted in studies dealing with this habi- tat (e.g. HARNISCH 1925, 1929). Only few investigations were carried out concerning the protist community of acid bog lakes. The flagellates of bogs have not been examined or presented in detail up to now. SKUJA'S works (1948, 1956, 1964) described many planktonic flagellates, but from acid lakes. The ciliates of acid bogs were comprehensively stud- ied and many new species were described by GROLII~RE (1975a, 1975b, 1977a, 1977b, 1978) and WENZEL (1953). Besides that, only older literature exists about this habitat
0075-9511/99/29/04-407 $ 12.00/0 407

i-
~ 15,000 b. Chr. Deposited ice blocks in the glacial drift
~ 11,000 b. Chr. (late glacial) Melting of the ice blocks and formation of kettle lakes, begin of the paludification
8,000 b. Chr. (post glacial) End of the paludification and temporary stagnation in the peat formation
Fig. 1. Development of the bog lakes (after JESCHKE 1987).
(e.g. LEVANDER 1900; MERMOD 1914; SCHLENKER 1908; THIEBAUD &; FAVRE 1906).
Testate amoebae are named tyrphobionts or sphagno- bionts and are considered as characteristic organisms of bogs. Therefore, extensive studies exist concerning their dis- tribution, zoning, ecology, and systematics (e.g. HARNISCH 1925, 1927, 1929; MEISTERFELD 1977, 1979a, 1979b; SCHONBORN 1962a, 1962b, 1963, 1990). Each region of the Sphagnum mats comprises its own community of testate amoebae. The horizontal distribution of these protists de- pends on the degree of humidity; the vertical zoning is de- pendent on the light impact and the trophy (SCHONBORN 1962b, 1963). Characteristic genera of testate amoebae with- in the wet Sphagnum are: Hyalosphenia, Difflugia, Arcella, Centropyxis, Nebela, Euglypha, and Heleopera. The species Hyalosphenia papilio, Amphitrema flavum, A. stenostoma, Heleopera sphagni, and Placocista spinosa harbour zoo- chlorellae and are very sensitive indicators for oligotrophic conditions (ScHONBORN 1963). Reasons why the group of testate amoebae in general are typical bog inhabitors, are the relative low competitive pressure with other protists and the oligotrophic conditions in bogs (GOTTLICH 1990; SCHON- BORN 1962a, 1962b).
Our study deals with the species composition of the bog lakes, with main emphasis on the periphyton in the Sphag- num mat. Indicator species of acid, dystrophic lakes are fig- ured out and the environmental conditions (especially the trophy) of the lakes are compared to earlier studies (SCHON- BORN et al. 1965) and discussed.
The investigated bog lakes Himmelreichsee, Teufels-See, Groger Barschsee and Kleiner Barschsee were described in detail in STRODER-KYPKE (1999, this volume), showing the seasonal changes in the periphyton community and the peri- phyton structures of different habitats within the lakes.
Material and Methods
Sampling The sampling of the periphyton with glass slides as artificial sub- strates according to WmBERT (1969) was performed as described in STRI~DER-KYPKE (1999, this volume). Also the live counting of the protists is decribed there in detail.
The sampling and counting of the testate amoebae of the Sphag- num mats were done as described by SCn0NBORN (1963). To differ- entiate the communities of the upper and lower horizons, the Sphag- num cushion was cut - starting from the surface - into short pieces of approximately 1 cm length and examined separately.
Light microscopy Species determination was performed by means of an Axiophot mi- croscope with high magnification and - if necessary - with phase contrast or differential interference contrast. Additionally, protargol impregnation of whole slides was carried out in order to confirm species determinations of the ciliates.
Species determination was made mainly with the help of the fol- lowing literature:
Flagellates: ETTL (1983), ETTL & GARTNER (1988), HUBER- PESTALOZZI (1941, 1955, 1968), PATTERSON & LARSEN (1991), POPOVSKY & PFIESTER (1990), SKUJA (1948, 1956, 1964), and STAR- MACH (1985).
Ciliates: FOISSNER et al. (1991, 1992, 1994, 1995), KAnL (1930, 1931, 1932, 1935).
Protargol impregnation of whole slides The protargol staining of the exposed slides followed the procedure described by WILBERT (1976). The cells were fixed in vapour of a 2% aqueous solution of Osmium tetroxide for 1-2 minutes. Subse- quently the liquid remaining on the slides was mixed with a drop of egg-albumen and the slides were allowed to dry at room temperature
408 Limnologica 29 (1999) 4
for 24 hours. The periphyton was post-fixed in Bouin's solution for 25 minutes and then transferred to 70% ethanol for 20 minutes, in order to remove the fixative. At this stage, the staining can be inter- rupted, e. g. to collect the slides of several sampling dates. The slides were washed for 5 minutes in running tap water. Subsequently, oxi- dation in a 5% Potassiumpermanganate solution for 1-2 seconds, fol- lowed by bleaching with 5% oxalic acid, also for 1-2 seconds, took place. Both steps were followed by a 10 minute washing step in run- ning tap water. Then the slides were rinsed in Aqua dest. and placed in a 0.5% Protargol solution (= Albumosesilver of the Fa. Fluka, Buchs/Switzerland) for 45 minutes at 45-50 °C. After this impregna- tion, the slides were rinsed in Aqua dest. and transferred into a 1% Hydroquinone solution for 5 minutes. The slides were washed for 10 minutes in running tap water and finally fixed in a 5% Sodiumthio-
sulfate solution for 1-2 minutes. After a last washing step in running tap water for 5 minutes, the slides were dehydrated in ethanol series (30, 50, 70, 95, 100%) with Xylene as final medium (5 minutes each) and mounted with Entellan (Fa. Merck, Darmstadt).
Results and Discussion
Species list of the flagellates and ciliates in the four bog lakes
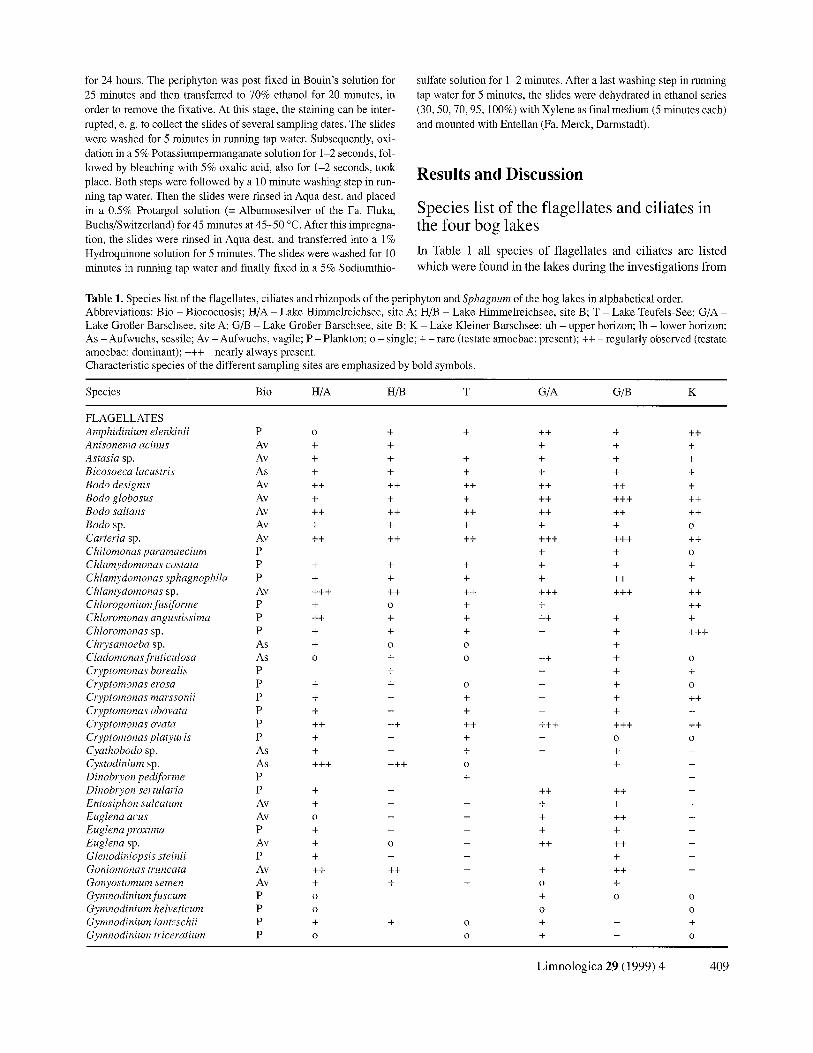
In Table 1 all spec ies o f f lage l lates and c i l iates are l i s ted
w h i c h w e r e f o u n d in the lakes during the inves t iga t ions from
Table 1. Species list of the flagellates, ciliates and rhizopods of the periphyton and Sphagnum of the bog lakes in alphabetical order. Abbreviations: Bio - Biocoenosis; H/A - Lake Himmelreichsee, site A; H/B - Lake Himmelreichsee, site B; T - Lake Teufels-See; G/A - Lake Groger Barschsee, site A; G/B - Lake GroBer Barschsee, site B; K - Lake Kleiner Barschsee; uh - upper horizon; lh - lower horizon; As - Aufwuchs, sessile; Av - Aufwuchs, vagile; P - Plankton; o - single; + - rare (testate amoebae: present); ++ - regularly observed (testate amoebae: dominant); +++ - nearly always present. Characteristic species of the different sampling sites are emphasized by bold symbols.
Species Bio H/A H/B T G/A G/B K
FLAGELLATES Amphidinium elenkinii P o + + Anisonema acinus Av + + Astasia sp. Av + + + Bicosoeca lacustris As + + + Bodo designis Av ++ ++ ++ Bodo globosus Av + + + Bodo saltans Av ++ ++ ++ Bodo sp. Av + + + Carteria sp. Av ++ ++ ++ Chilomonas paramaecium P Chlamydomonas costata P + + + Chlamydomonas sphagnophila P + + + Chlamydomonas sp. Av + + + + + + +
ChloJvgonium fusi forme P + o + Chloromonas angustissima P ++ + + Chloromonas sp. P + + + Chrysamoeba sp. As + o o
Cladomonasfi 'uticulosa As o + o ClTptomonas borealis P + Cryptomonas erosa P + + o Cryptomonas marssonii P + + + Cryptomonas obovata P + + + Cryptomonas ovata P ++ ++ ++ Cryptomonas platyuris P + + + Cyathobodo sp. As + + + Cystodinium sp. As + + + + + + o Dinobryon pediforme P + Dinobryon sertularia P + + Entosiphon sulcatum A v + + +
Euglena acus A v o + +
Euglena proxima P + + + Euglena sp. Av + o + Glenodiniopsis steinii P + + + Goniomonas truncata A v + + + + +
Gonyostomum semen Av + + +
Gymnodinium fi~scum P o Gymnodinium helveticum P o Gymnodinium lantzschii P + + o Gymnodinium triceratium P o o
+ + +
+ +
+ +
+ +
+ + + +
+ + + + +
+ + + +
+ +
+ + + + + +
+ +
+ +
+ + +
+ + + + + +
+
+ + +
+ +
+
+ + +
+ +
+ +
+ +
+ +
+ + + + + +
+ 0
+ +
+
+ + + +
+ +
+ + +
+ +
+ + + +
+
+ + +
0 +
+ 0
0
+ +
+ +
+ +
+
+
+
+
+ +
+ +
0
+ +
0
+
+
+ +
+ +
+
+ + +
o
+
o
+ +
+
+ +
o
+
+
+
+
+
+
+
+
+
+
L i m n o l o g i c a 29 (1999 ) 4 4 0 9
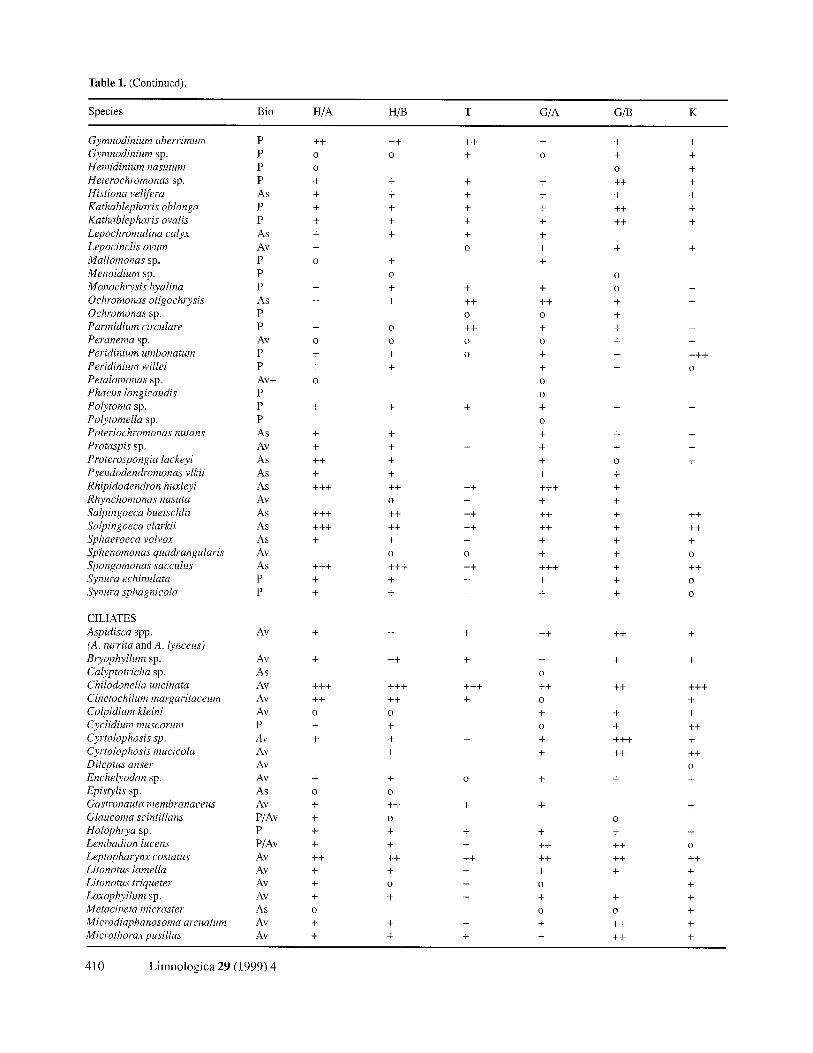
Table 1. (Continued).
Species Bio H/A H/B T G/A G/B K
Gymnodinium uberrimum P ++ ++ ++ + + + Gymnodinium sp. P o o + o + +
Hemidinium nasutum P o o + Heterochromonas sp. P + + + + + + +
Histiona velifera A s + + + + + +
Kathablepharis obIonga P + + + + ++ + Kathablepharis ovalis P + + + + ++ + LepochTvmulina calyx A s + + + +
Lepocinclis ovum A v + o + + +
Mallomonas sp. P o + +
Menoidium sp. P o o
Monochrysis hyalina P + + + + o + Ochtvmonas oligochrysis A s + + + + + + + +
Ochromonas sp. P o o +
Parmidium circulare P + o ++ + + + Peranema sp. A v o o o o + +
Peridinium umbonatum P + + o + + +++ Peridinium willei P + + + + o Petalomonas sp. A v + o o
Phacus longicaudis P o Polytoma sp. P + + + + + +
Polytomella sp. P o
Poteriochromonas nutans A s + + + + +
P~vtaspis sp. A v + + + + + +
Protetvspongia lackeyi A s + + + + o +
Pseudodendromonas vlkii A s + + + +
Rhipidodendron huxleyi A s + + + + + + + + + + +
Rhynchomonas nasuta A v o + + +
Salpingoeca buetschlii A s + + + + + + + + + + + +
Salpingoeca clarkii A s + + + + + + + + + + + +
Sphaeroeca volvox A s + + + + + +
Sphenomonas quadrangularis A v o o + + o
Spongomonas sacculus A s + + + + + + + + + + + + + +
Synura echinulata P + + + + + o Synura sphagnicola P + + + + + o
C I L I A T E S
Aspidisca spp . A v + + +
(A. turrita a n d A. lynceus) Bryophyllum sp. A v + + + +
Calyptotricha sp. A s
Chilodonella uncinata A v + + + + + + + + +
Cinetochilum margaritaceum A v + + + + +
Colpidium kleini A v o o
Cyclidium muscorum P + + Cyrtolophosis sp. Av + + + Cyrtolophosis mucicola A v +
Dileptus anser A v
Enchelyodon sp. A v + + o
Epistylis sp. A s o o
Gastronauta membranaceus A v + + + +
Glaucoma scintillans P / A v + o
Holophrya sp. P + + +
Lembadion lucens P / A v + + +
Lepwpharynx costatus A v + + + + + +
Litonotus lamella A v + + +
Litonotus triqueter A v + o +
Loxophyllum sp. A v + + +
Metacineta micraster A s o
Microdiaphanosoma arcuatum A v + + +
Microthorax pusillus A v + + +
+ + + + +
+ +
0
+ + + +
0
+ +
0 +
+ + + +
+ + +
+ +
+
+ + +
+
+
+ +
+
+ +
0
+
0
+ + +
+ + + + 0
+ + + + + +
+ + +
0 +
+ + +
0 0 +
+ + + +
+ + + +
4 1 0 L i m n o l o g i c a 2 9 ( 1 9 9 9 ) 4
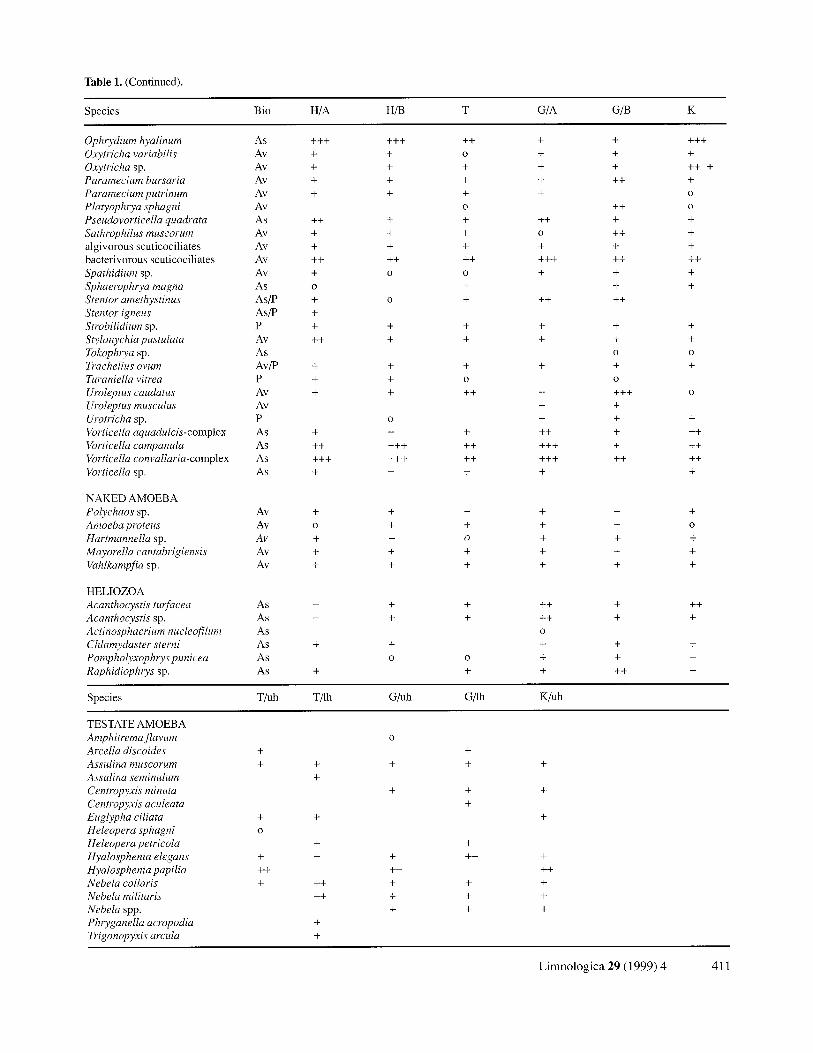
Table 1. (Continued).
Species Bio H / A H/B T G / A G/B K
Ophrydium hyalinum As + + + + + + ++ + +
Oxytricha variabilis Av + + o + +
Oxytricha sp. Av + + + + +
Paramecium bursaria Av + + + + ++
Paramecium putrinum A v + + + +
Platyophrya sphagni Av o ++
Pseudovorticella quadrata As ++ + + ++ +
Sathrophilus muscorum A v + + + o ++
algivorous scuticociliates Av + + + + + b a c t e r i v o r o u s scu t icoc i l i a tes Av ++ ++ ++ + + + ++
Spathidium sp. Av + o o + +
Sphaerophrya magna As o + +
Stentor amethystinus A s / P + o + ++ ++
Stentor igneus A s / P +
Strobilidium sp. P + + + + +
Stylonychia pustulata Av ++ + + + +
Tokophrya sp. A s o
Trachelius ovum A v / P + + + + +
Turaniella vitrea P + + o o Uroleptus caudatus A v + + ++ + + + +
Uroleptus museulus Av + +
Urotricha sp. P o + +
Vorticella aquadulcis-complex A s + + + ++ +
Vorticella campanula As ++ + + + ++ + + + +
Vorticella convallaria-complex As + + + + + + ++ + + + ++
Vorticella sp. As + + + +
N A K E D A M O E B A
Polychaos sp. Av + + + + +
Amoeba proteus Av o + + + +
Hartmannella sp. Av + + o + + Mayorella cantabrigiensis Av + + + + +
Vahlkampfia sp. Av + + + + +
H E L I O Z O A
Acanthocystis turfacea A s + + + ++ +
Acanthocystis sp. As + + + ++ +
Actinosphaerium nucleofilum As o
Chlamydaster sterni As + + + +
Pompholyxophrys punicea A s o o + +
Raphidiophrys sp. A s + + + ++
Species T/uh T/ lh G / u h G/ lh K / u h
T E S T A T E A M O E B A
Amphitrema flavum o Arcella discoides + + Assulina muscorum + + + + + Assulina seminulum + Centropyxis minuta + + + Centropyxis aculeata + Euglypha ciliata + + + Heleopera sphagni o Heleopera petricola + + Hyalosphenia elegans + + + ++ + Hyalosphenia papilio ++ ++ ++ Nebela collaris + ++ + + + Nebela militaris ++ + + + Nebela spp. + + +
Phryganella acropodia + Trigonopyxis arcula +
+ + +
+
+ + q-
+
O
O
+
+
+
+ +
+
+
+
+
O
+
O
+
+ +
+ +
+-I-
+
+ +
+
+
+
+
L i m n o l o g i c a 2 9 ( 1 9 9 9 ) 4 4 1 1

September 1992 until September 1993. The list shows a great variety of protists and comprises only the most impor- tant species on the slides and some easy determinable rare species. Only few naked amoebae and heliozoans are listed. The testate amoebae were found in 1996 by squeezing water out of the Sphagnum plants and therefore a different listing is applied. They were partly observed on the slides in the earlier years. Metazoans were not taken into account, neither many of the heterotrophic nanoflagellates. Determination was often only possible to the genus level.
A general evaluation of the colonization of the exposed slides shows some differences concerning the species com- position in the four investigated bog lakes. There are many species that occurred in all four lakes. Every lake, however, also possesses some characteristic species. Especially in the upper region of the Sphagnum mat in Lake Grol3er Barschsee a totally different species composition of the periphyton was observed. The flagellates Chlamydomonas sphagnophila Chloromonas angustissima, Euglena acus, Heterochro- monas sp., Katablepharis ovalis, K. oblonga, and the ciliates Cyrtolophosis sp., C. mucicola, Microdiaphanosoma arcua- turn, Microthorax pusillus, Oxytricha sp., Paramecium bur- saria, Platyophrya sphagni, Sathrophilus muscorum, and Uroleptus caudatus occurred rarely or never on the other sampling sites. Some of these observed species are consid- ered as characteristic soil protists (e.g. Chloromonas an- gustissima, Microdiaphanosoma arcuatum). This refers to the intermediate position of the Sphagnum mat, in which the inner regions are generally submerged, but the outer regions may fall dry during water shortage. The milieu in the upper Sphagnum region is very different from the other habitats of the bogs: Due to the permanent new formation of Sphagnum shoots and the lacking degradation of old plant parts, the conditions are rather oligotrophic in this upper horizon. Fur- thermore, the pH within the Sphagnum mat is more acidic than in the water body because the Sphagnum plants function as a cation exchanger. Adaptations to the low concentration of nutrients are vagility, a broad food spectrum, or the sym- biosis with zoochlorellae. The latter is realized by many pro- tists that live in the upper region of the Sphagnum mat (e.g. Paramecium bursaria, Platyophrya sphagni, Uroleptus cau- datus, Mayorella viridis, Acanthocystis tutfacea, Hyalosphe- nia papilio, Heleopera sphagni) and was proofed in earlier studies (HARNISCH 1925; WENZEL 1953). The lower horizon (brown, with dead Sphagmun stems) is dark and has a higher nutrient concentration.
Only few sessile species colonized the slides. The most abundant sessile ciliates were Vorticella campanula, Ophry- dium hyalinum and specimens of the Vorticella convallaria- complex. Sessile flagellates were represented by Cyathobo- do sp., Salpingoeca spp., Spongomonas sacculus, Rhipido- dendlvn huxleyi, Protelvspongia lackeyi and few numbers of Bicosoeca lacustris, Cladomonas fruticulosa and Pseudo- dendlvmonas vlkii. Most species were vagile and a large number of planktonic protists was also observed on the
slides. This is striking since slide methods generally prefer sessile species, whereas other artificial substrates favour the colonization of vagile species (BAMFORTH 1982; WEITZEL et al. 1979). The ratio of sessile to vagile flagellate and ciliate species is 27 : 47 = 0.57 and therefore indicates mesotrophic conditions within the bog lakes (ZOLOTAREV 1995).
The saprobical index of the species listed here is also mainly oligo- or mesosaprobic. Dinobryon and Salpingoeca, for example, avoid saprobic habitats (LEMMERMANN 1914); also SLADECEK (1973) defined Salpingoeca as indicator species for oligosaprobic conditions. ZOLOTAREV (1995) showed that in oligosaprobic waters Salpingoecidae domi- nate, whereas in mesosaprobic waters the colonial choanoflagellates and in polysaprobic environments vagile bodonids and heterotrophic euglenids are more important. Bodonid and heterotrophic euglenid species had only minor impact concerning species number and abundance in the pe- riphyton of the bog lakes (compare STRUDER-KYPKE 1999, this volume). GROL~gRE (1977b, 1978) mostly found alpha- to beta-mesosaprobic ciliate species in the Sphagnum mats of acid bog lakes. None of the species observed by him had an oligosaprobic index, but many of them were not classi- fied. Dystrophic lakes contain much organic material, but predominantly in a form that cannot be used by the organ- isms. The mineralization of the plant parts is stopped in an early stage, therefore the density of bacteria in the bogs which serve as food for protozoa is too low for high abun- dances e.g. of large peritrich filter feeders (GROLIERE 1977b). Bacterivorous organisms of the lakes are mostly hetero- trophic flagellates like Salpingoeca spp., Cyathobodo sp., Bodo spp., and Goniomonas truncata (compare STRUDER- KYPKE 1999, this volume).
A great part of the observed ciliates shows a wide food spectrum which comprises bacteria, blue-green algae, di- atoms, green algae, small flagellates, and even small ciliates (e.g. Uroleptus caudatus, Litonotus lamella, Holophrya sp., or Prorodon sp.) in order to cope with the low availability of nutrients. Osmotrophy probably is important in the lakes in spite of the little dissolved organic contents. SrtERR (1988) noted that heterotrophic flagellates are able to ingest and use high molecular compounds. In the bog lakes with a high per- centage of little decayed material, this seems to be a imagin- able strategy for energy and food metabolism. In contrary to oligotrophic lakes, dystrophic lakes show a different energy flux: The heterotrophic plankton often has a higher biomass production than the phytoplankton, since it is able to use de- tritus particles or dissolved organic matter (DOM) as energy source (HESSEN et al. 1990). Mixotrophy is not very common in the bog lakes. Also in other oligotrophic waters, mixotrophic protists are not as abundant as might be as- sumed considering the benefit of this growth mode (ARNDT & BERNIN6ER 1995). Chrysomonads and dinoflagellates composed only a small percentage of the periphyton com- munity, cryptomonads are almost obligate autotrophic (SANDERS & PORTER 1990).
412 Limnologica 29 (1999) 4

Indicator species of bog lakes
Most of the species that occurred regularly in the lakes are ubiquitous and common in limnetic habitats (e.g. Bodo, Chilodonella, Cryptomonas, Gymnodinium, Vorticella). Therefore they cannot be regarded as indicator species of bog lakes. Compared to eutrophic habitats they reach higher abundances, since the competition with typical bacterivores (e g. scuticociliates that can reach high population densities) is less important in oligotrophic or dystrophic environments. Furthermore, many of the ubiquitous species are characteris- tic pioneers of newly exposed substrates and are repressed in the course of succession (CAIRNS & HENEBRY 1982).
As for other animal taxa (e.g. insects and vertebrates), a characterization of the protist fauna in bogs is hard to define by certain species. Many of the flagellates and ciliates show high tolerances concerning oxygen, pH range, availability of food, and DOM in the water, therefore they are not strictly bound to certain habitats. Often an exact species determina- tion is not possible, since adaptations to extreme environ- ments occur. One example is the rather common symbiosis with zoochlorellae in Sphagnum mats (or the chloroplast re- tention, respectively), and WENZEL (1953) reported from mosses often stunted forms that were smaller than specimens examined from other waters. An additional problem is the great variety of habitats in bogs that ranges from dry soil en- vironments to planktonic distribution in the residue lakes. The species composition of dry mosses, wet Sphagnum mats and the free water-body differs considerably from each other. This makes a comparison to other reports difficult, since dif- ferent methods of sampling lead to very different results. WENZEL (1953) compared the ciliate community and species of different bogs. Since he mainly investigated the mosses of dry habitats, he found many soil ciliates (e.g. Colpoda). In spite of the observation of some soil ciliates, the greatest per- centage of the species we noted were aquatic, and therefore only few species described by WENZEI~ (1953) are in accor- dance to our reports. Cinetochilum margaritaceum, Cyrtolo- phosis mucicola, Leptopharynx costatus, Litonotus lamella, Microdiaphanosoma arcuatum, Sathrophilus muscorum, and Uroleptus caudatus were species of his samples that oc- curred in the investigated bog lakes.
Also the sphagnophil species described by GROLIgRE (1978), were not observed in the periphyton (except Lep- topharynx costatus). This may be due to the applied methods (squeezing of the Sphagnum versus exposure of slides), to different investigated habitats or it may represent the fact that no characteristic protist species of bogs exist, but that these most abundant species are generally common. Another reason may be a beginning eutrophication of the investigated bog lakes which is also indicated by our investigation of the testate amoeba community in the Sphagnum of the lakes (see below).
Nevertheless, some species seem to be more common in dystrophic bogs than in other habitats. A joint occurrence of
the following ciliates and flagellates together with other ubiquitous protists and, of course, testate amoebae marks a typical bog lake periphyton community.
Among the flagellates, Spongomonas sacculus, Rhipido- dendron huxleyi, Cyathobodo, and Pseudodendlvmonas species can be assumed as characteristic bog protists. They were reported to appear almost exclusively in bog lakes. Al- though Spongomonas sacculus and Rhipidodend~vn huxleyi, e.g., are relatively common, their main distribution was noted in Sphagnum bogs (HIm3ERD 1976a, 1983). KENT'S (1880) original description of both species is based on sam- ples from bog lakes. They form their matrix from iron and manganese compounds. Both elements are sufficiently avail- able in the bog lakes (STR~DER-KYPKE & HAUSMANN 1998), maybe this is the reason for the common occurrence of these species in bogs. Also the Cyathobodo and Pseudoden- dlvmonas species were mostly described from Sphagnum bogs (HIBBERD 1976b, 1985; MIGNOT 1974).
Leptopharynx costatus, Platyophrya sphagni, and Ulv- leptus caudatus are ciliate species which were mostly found in bogs or small waters (FoISSNER 1980; GROLI~RE 1975b, 1977a, 1978; KAHL 1931, 1932; MERMOD 1914; PENARD 1922; WENZEL 1953). GROI, IERE (1977b, 1978) defined sev- eral characteristic ciliate species for acid bogs in France, but only Leptopharynx costatus, Uroleptus caudatus, Urotricha ovata and Platyophrya sphagni (described as P. viridis and synonymized by FOISSNER 1993) were observed during our study. Cyclidium sphagnetorum was regarded by GROLIERE (1978) to be a characteristic species of the bogs with relative abundances of up to 95-99%. This species was not observed in our investigated bogs. Neither did MERMOD (1914) and WENZEL (1953) find C. sphagnetorum during their studies. GROHERE (1975, 1977a, 1977b, 1978), however, investigat- ed very acid habitats with a pH ranging from 3-5, this may be one reason for the lacking of his characteristic species in our subneutral environments. He noted that the most abun- dant ciliates of this upper region in the bogs harboured zoochloreUae in their cytoplasm. This was confirmed by the study of STR~DER-KYeKE (1999, this volume), but also many colourless ciliates and heterotrophic flagellates occurred in this upper region.
Testate amoebae The upper, moist horizons of the Sphagnum mats in Lake Teufels-See were dominated by Hyalosphenia papilio, a characteristic species of oligotrophic bog lakes that harbours zoochlorellae. Other typical zoochlorellae-bearing species like Heleopera sphagni, Placocista spinosa, and especially Amphitrema spp. were much less abundant or did not appear at all. Colourless species like Hyalosphenia elegans, Nebela ssp., and Assulina muscorum, however, that also occur in habitats of higher trophy, e.g. in swamps and in the lower horizons of the Sphagnum mats, were observed in this upper region. The lower and dry region of the Sphagnum revealed a
Limnologica 29 (1999) 4 413

typical community of testate amoebae with dominance of two Nebela species. Besides that, many other colourless but no zoochlorellae-harbouring species occurred. The density of testate amoebae was comparable to other bog lakes for both horizons in Lake Teufels-See.
In Lake Grof3er Barschsee similar results were obtained: The upper horizon was dominated by Hyalosphenia papilio, but apart from this and rare observations of Amphitrema flavum, no other characteristic species with zoochlorellae were noted. Instead of this, colourless species like Centropy- xis minuta, Nebela spp., and Hyalosphenia elegans occurred. The latter dominated the lower humid horizon, where a strik- ing low density of testate amoebae was recognizable. Mostly species of the genera Arcella, Nebela, and Centropyxis were found.
The upper, moist horizon of Lake Kleiner Barschsee re- vealed a very high density of testate amoebae. Dominant was Hyalosphenia papilio, all other species were colourless, e.g. Nebela spp., Centropyxis minuta, Hyalosphenia elegans, and Euglypha ciliata.
As on the slides of the upper region, the squeezed @hag- num samples showed relative high abundances of other pro- tists with zoochlorellae (naked amoebae, heliozoans and cili- ates), photoautotrophic flagellates of the genus Euglena and
heterotrophic flagellates, but also nematodes, bdelloid ro- tifers, and mites. The water of the squeezed Sphagnum showed a distinct sediment of detritus which had an C:N- ratio of 18 and therefore must be regarded as an easy degrad- able and favourized source of nutrients. The pH of the squeezed water revealed subneutral conditions that ranged around 6.3, and once it even reached 6.8.
The lack of typical testacean indicators and the occur- rence of species of the lower horizon in the upper region may be due to the eutrophication of the bogs which definitely could be proofed by the comparison of the distribution of tes- tate amoebae in the years 1963 and 1992-1998. The commu- nity structures of testate amoebae that were as described in 1963 (SCHONBORN 1963) could be confirmed until 1972. After that, no studies were made until 1992. Investigations of the years 1992, 1994 and 1996-1998 showed that the com- munity structure had changed distinctly. Amphitrema species like A.flavum and A. stenostoma which are very sensitive in- dicator species for oligotrophic bogs and are restricted to the upper horizon of the Sphagnum, were rarely found. Besides that, Placocista spinosa could not be observed. Most species of the lower region are also distributed in swamps (e.g. Hyalosphenia elegans, Nebela spp., Centropyxis spp., Trigonopyxis spp., and Assulina spp.) (ScHONBORN 1962b,
Upper horizon:
green, living Sphagnum
stems
Lower horizon:
brown, dead Sphagnum
stems
Testate amoebae in moist Sphagnum mats
k
1963 and before it Recorded since 1992
Decreasing density row: Decreasing density row:
Hyalosphenia papilio*
Amphitrema flavum*
Heleopera sphagni*
Amphitrema stenostoma *
Centropyxis minuta
Ptacocista spinosa*
Hyalosphenia papilio*
Hyalosphenia elegans
Nebela tincta
Euglypha ciliata
Assulina muscorum
Centropyxis minuta
Nebela collaris
Nebela tincta
Assulina muscorum
Hyalosphenia etegans
Nebela collaris
Nebela minor
Nebela militaris
Centropyxis minuta
Hyalosphenia elegans
Nebela tincta
Euglypha ciliata
Nebela collaris
Nebela militaris
Centropyxis minuta
Fig. 2. Stratification of testate amoebae in the moist zone of Sphagnum mats. Summarized results from Lake Teufels-See, Lake Groger Barschsee and Lake Kleiner Barschsee. Explanation: Species marked with an asterisks = with zoochlorellae.
414 Limnologica 29 (1999) 4
1963). Apart from the changes in species composition, the stratification of the Sphagnum mats is mixed up: Colourless species of the lower horizons also occur in the upper region of the mat, showing that this horizon has become more nutri- ent rich, due to an eutrophication of the bogs. In earlier stud- ies, only zoochlorellae-harbouring species were found in this upper zone (Fig. 2).
Characteristic flagellate and ciliate species of the lakes and their seasonal distribution
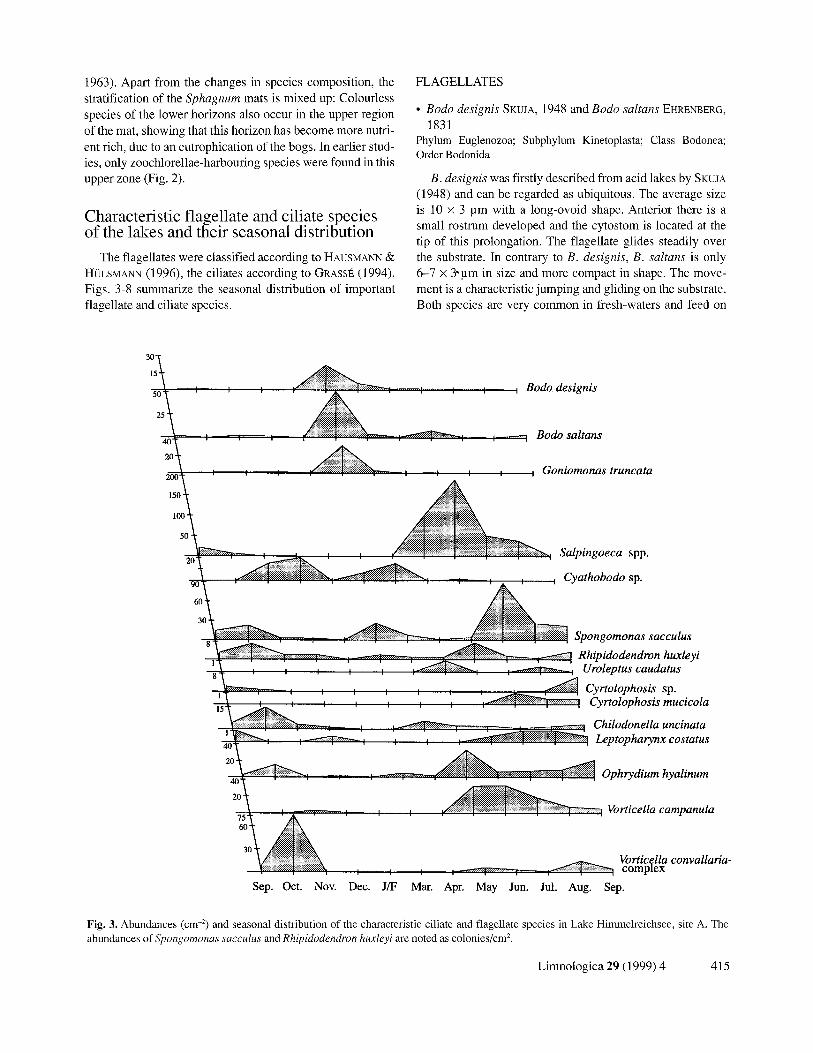
The flagellates were classified according to HAUSMANN ~; Ht2LSMANN (1996), the ciliates according to GRASSt~ (1994). Figs. 3-8 summarize the seasonal distribution of important flagellate and ciliate species.
FLAGELLATES
• Bodo designis SKUJA, 1948 and Bodo saltans EHRENBERG, 1831
Phylum Euglenozoa; Subphylum Kinetoplasta; Class Bodonea; Order Bodonida
B. designis was firstly described from acid lakes by SKUJA (1948) and can be regarded as ubiquitous. The average size is 10 x 3 ~m with a long-ovoid shape. Anterior there is a small rostrum developed and the cytostom is located at the tip of this prolongation. The flagellate glides steadily over the substrate. In contrary to B. designis, B. saltans is only 6-7 x 3 ~ m in size and more compact in shape. The move- ment is a characteristic jumping and gliding on the substrate. Both species are very common in fresh-waters and feed on
30 ~
5:, t 40"
20
15o ~
20'
Bodo designis
Bodo saltans
Goniomonas truncata
Salpingoeca spp.
Cyathobodo sp.
30~
8"
11
:-:,x,:,:::
20
40 "
3T
Spongomonas sacculus
Rhipidodendron huxleyi Uroleptus caudatus
Cyrtolophosis sp. Cyrtolophosis mucicola
Chilodonella uncinata Leptopharynx costatus
Ophrydium hyalinum
Vorticella campanula
30
Sep. Oct. Nov. Dec. J/F Mar. Apr. May Jun. Jul. Aug.
Vortic¢lla convallaria- complex
Sep.
Fig. 3. Abundances (cm -2) and seasonal distribution of the characteristic ciliate and flagellate species in Lake Himmelreichsee, site A. The abundances of Spongomonas sacculus and Rhipidodendron huxleyi are noted as colonies/cm 2.
Limnologica 29 (1999) 4 415
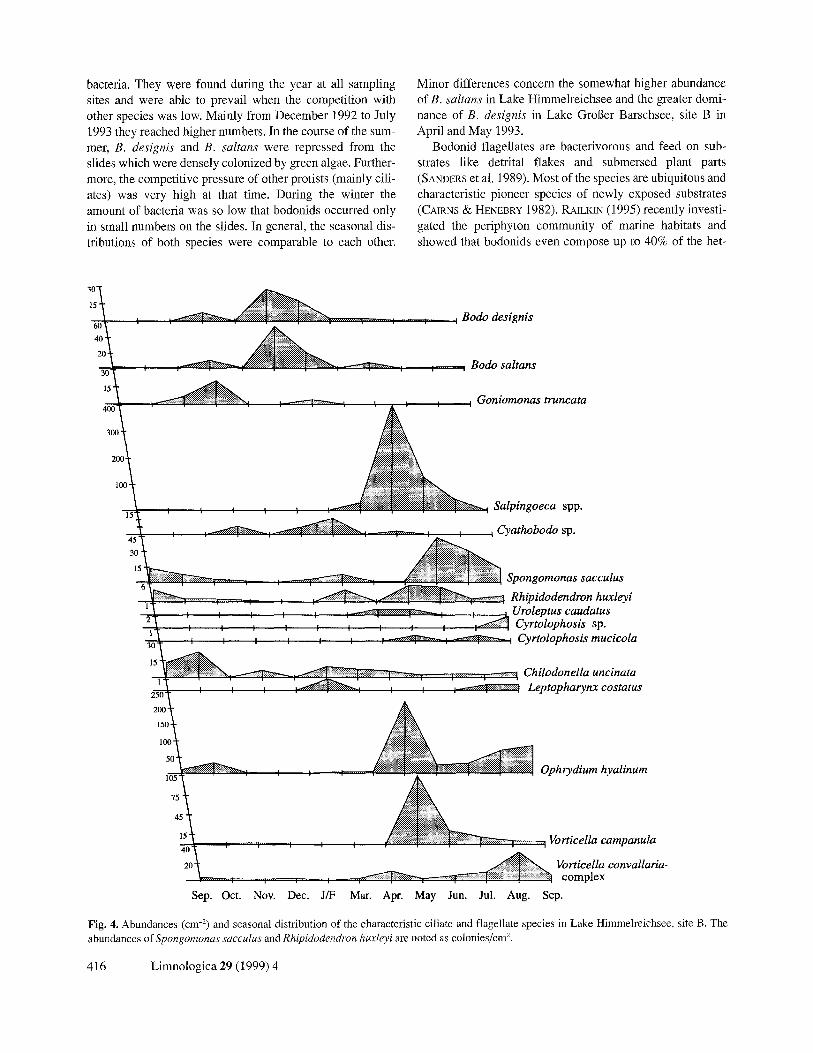
bacteria. They were found during the year at all sampling sites and were able to prevail when the competition with other species was low. Mainly from December 1992 to July 1993 they reached higher numbers. In the course of the sum- mer, B. designis and B. saltans were repressed from the slides which were densely colonized by green algae. Further- more, the competitive pressure of other protists (mainly cili- ates) was very high at that time. During the winter the amount of bacteria was so low that bodonids occurred only in small numbers on the slides. In general, the seasonal dis- tributions of both species were comparable to each other.
Minor differences concern the somewhat higher abundance of B. saltans in Lake Himmelreichsee and the greater domi- nance of B. designis in Lake GroBer Barschsee, site B in April and May 1993.
Bodonid flagellates are bacterivorous and feed on sub- strates like detrital flakes and submersed plant parts (SANDERS et al. 1989). Most of the species are ubiquitous and characteristic pioneer species of newly exposed substrates (CAIRNS & HENEBRY 1982). RAILKIN (1995) recently investi- gated the periphyton community of marine habitats and showed that bodonids even compose up to 40% of the her-
307
40 I 2O
3-"6" " 15
400"
Bodo designis
Bodo saltans
, Goniomonas truncata
30£
20(
45 "i 30" 15
Salpingoeca spp.
Cyathobodo sp.
15
-? 250" 200'
150 i 100 50
105"
Spongomonas sacculus
Rhipidodendron huxleyi Uroleptus caudatus
t Cyrtolophosis sp. • ~ Cyrtolophosis mucicola
Chilodonella uncinata Leptopharynx costatus
~ Ophrydium hyalinum 75 __ 45
15 7ff
Sep. Oct. Nov. Dec. J/F Mar. Apr. May Jun. Jul. Aug.
Vorticella campanula
~ Vorticella convallaria- complex
Sep.
Fig. 4. Abundances (cm-:) and seasonal distribution of the characteristic ciliate and flagellate species in Lake Himmelreichsee, site B. The abundances of Spongomonas sacculus and Rhipidodendron huxleyi are noted as colonies/cm 2.
416 Limnologica 29 (1999) 4
erotrophic flagellates in these communities. Bodonids reach highest abundances in polysaprobic waters; in oligo- and mesosaprobic habitats their species and individual number is considerably lower (ZOLOTAREV 1995).
• Goniomonas truncata (FRESENIUS, 1858) STEIN, 1878 Phylum Chromista; Subphylum Cryptomonada; Order Cryptomona- dida
This primary colourless cryptomonad is ubiquitous pre- sent in fresh-waters. The flagellate is 10-15 pm long and re- veals a rigid ovoid shape. Obvious are the ejectisomes and the two flagella at the truncated anterior cell pole. G. trunca- ta is mainly found in the periphyton and is osmo- and phagotrophic. It was observed throughout the year in low abundances on the periphyton, only in Lake Himmelreichsee extreme peaks were reached in December 1992 and March 1993. The low individual number per cm 2 indicates a perma- nent occurrence of G. truncata that was not always noted. At site B of Lake Grof3er Barschsee autotrophic cryptomonad
species were present in high numbers, suggesting a plank- tonic distribution in surface near regions.
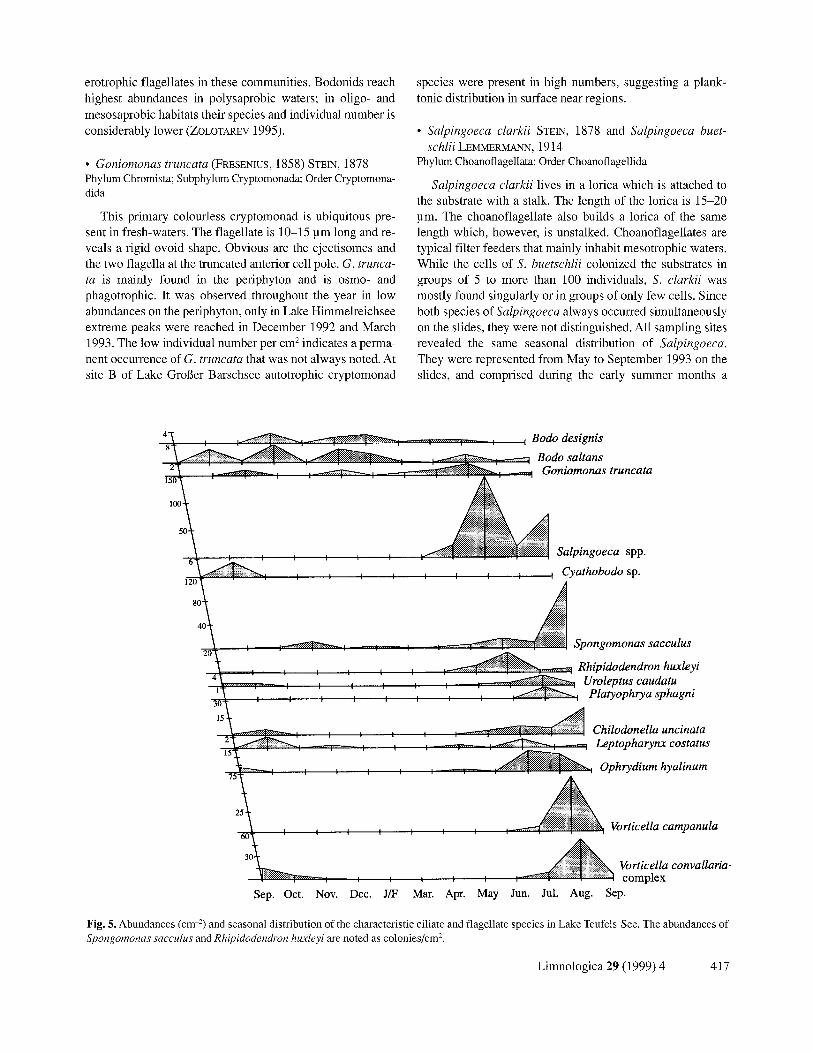
• Salpingoeca clarkii STEIN, 1878 and Salpingoeca buet- schIii LEMMERMANN, 1914
Phylum Choanoflagellata; Order Choanoflagellida
Salpingoeca clarkii lives in a lorica which is attached to the substrate with a stalk. The length of the lorica is 15-20 pro. The choanoflagellate also builds a lorica of the same length which, however, is unstalked. Choanoflagellates are typical filter feeders that mainly inhabit mesotrophic waters. While the cells of S. buetschlii colonized the substrates in groups of 5 to more than 100 individuals, S. clarkii was mostly found singularly or in groups of only few cells. Since both species of Salpingoeca always occurred simultaneously on the slides, they were not distinguished. All sampling sites revealed the same seasonal distribution of Salpingoeca. They were represented from May to September 1993 on the slides, and comprised during the early summer months a
4"
150"
I(R
120 "!
8(-
4~
'r 30" 15
Bodo designis
Bodo saltans , Goniomonas truncata
Salpingoeca spp.
Cyathobodo sp.
Spongomonas sacculus
I Rhipidodendron huxleyi Uroleptus caudatu
,~ Platyophrya sphagni
Chilodonella uncinata Leptopharynx costatus
Ophrydium hyalinum
Vorticella campanula
Sep. Oct. Nov. Dec. J/F Mar. Apr. May Jun. Jul. Aug.
Vorticella convallaria- complex
Sep.
Fig. 5. Abundances (cm "-) and seasonal distribution of the characteristic ciliate and flagellate species in Lake Teufels-See. The abundances of Spongomonas sacculus and Rhipidodendron huxleyi are noted as colonies/cm 2.
Limnologica 29 (1999) 4 417
dominating part of the periphyton. They preferably colonize filamentous green algae, zygnemales of the genus Spirogyra or diatoms (preferably of the genus Tabellaria) that were most abundant at this time. In the further course of the sum- mer, their number decreased and in the winter months rarely species of this genus were observed. Also in the plankton of other lakes salpingoecids are found more frequently during late spring and summer (SANDERS et al. 1989). Their de- crease in the course of the summer may be due to the stronger occurrence of peritich ciliates and the competition with these larger protists. A comparison of the seasonal dis- tribution of Vorticella and Salpingoeca reveals an alternation of the species of the V. convallaria complex and the flagel- lates. In Lake Kleiner Barschsee, where OphlTdium hyal- inure was a significant component of the periphyton, the
salpingoecids appeared generally only in low abundances. At site B of Lake Groger Barschsee the bacterial density was not high enough for filter feeders and additionally almost no filamentous green algae grew on the slides.
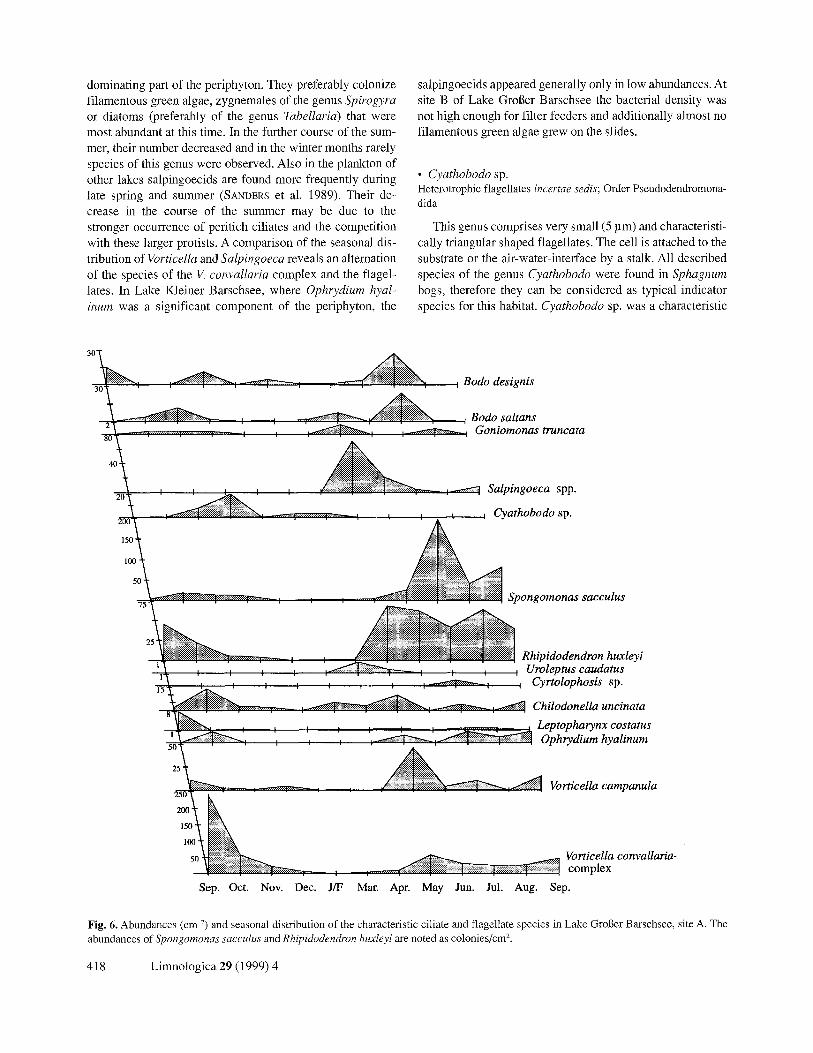
• Cyathobodo sp. Heterotrophic flagellates incertae sedis; Order Pseudodendromona- dida
This genus comprises very small (5 ~m) and characteristi- cally triangular shaped flagellates. The cell is attached to the substrate or the air-water-interface by a stalk. All described species of the genus Cyathobodo were found in Sphagnum bogs, therefore they can be considered as typical indicator species for this habitat. Cyathobodo sp. was a characteristic
30"1
30"
20"
150 t 100
50
75
Bodo designis
Bodo saltans .~ Goniomonas truncata
Salpingoeca spp.
Cyathobodo sp.
Spongomonas sacculus
25
150 100"
50
Sep. Oct. Nov. Dec. J/F Mar. Apr. May Jun. Jul. Aug.
Rhipidodendron huxleyi Uroleptus caudatus Cyrtolophosis sp.
Chilodonella uncinata
Leptopharynx costatus Ophrydium hyalinum
Vorticella campanula
Vorticella convaUaria- complex
Sep.
Fig. 6. Abundances (cm ~) and seasonal distribution of the characteristic ciliate and flagellate species in Lake Groger Barschsee, site A. The abundances of Spongomonas saccuhts and Rhipidodendron huxleyi are noted as colonies/cm 2.
418 Limnologica 29 (1999) 4
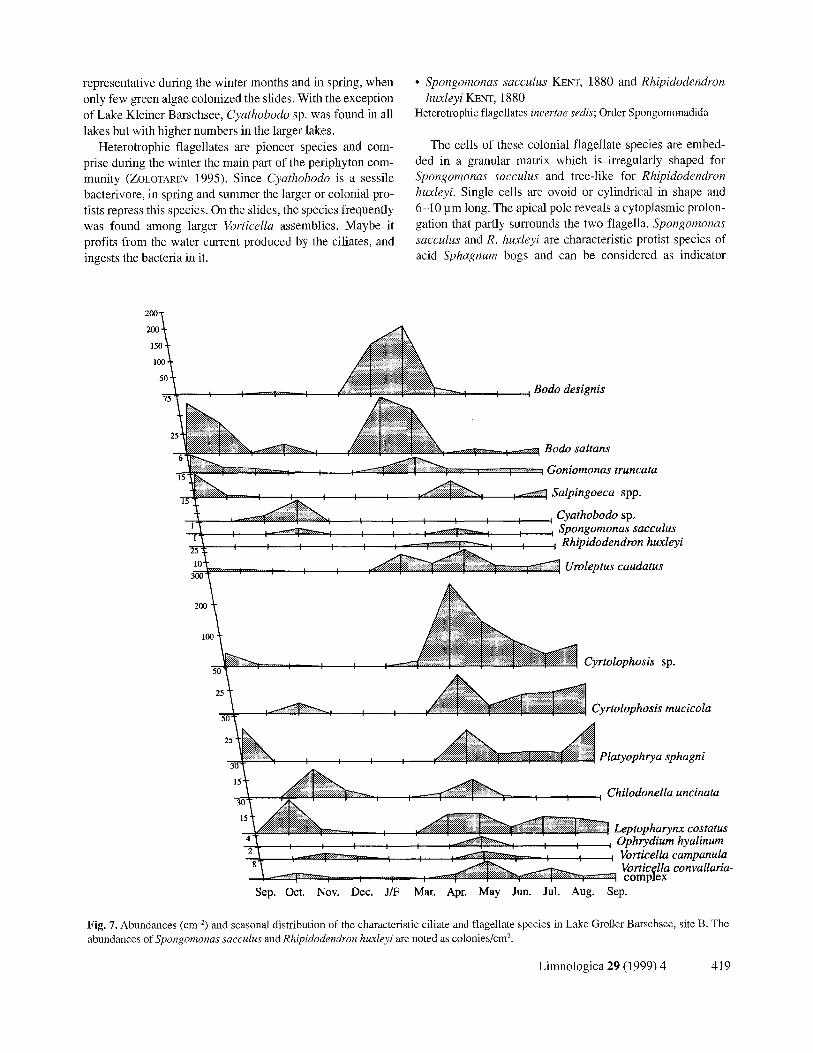
representative during the winter months and in spring, when only few green algae colonized the slides. With the exception of Lake Kleiner Barschsee, Cyathobodo sp. was found in all lakes but with higher numbers in the larger lakes.
Heterotrophic flagellates are pioneer species and com- prise during the winter the main part of the periphyton com- munity (ZOLOTAREV 1995). Since Cyathobodo is a sessile bacterivore, in spring and summer the larger or colonial pro- tists repress this species. On the slides, the species frequently was found among larger Vorticella assemblies. Maybe it profits from the water current produced by the ciliates, and ingests the bacteria in it.
• Spongomonas sacculus KENT, 1880 and Rhipidodendmn huxleyi KENT, 1880
Heterotrophic flagellates incertae sedis; Order Spongomonadida
The cells of these colonial flagellate species are embed- ded in a granular matrix which is irregularly shaped for Spongomonas sacculus and tree-like for Rhipidodendron huxleyi. Single cells are ovoid or cylindrical in shape and 6-10 ,um long. The apical pole reveals a cytoplasmic prolon- gation that partly surrounds the two flagella. Spongomonas sacculus and R. huxleyi are characteristic protist species of acid Sphagnum bogs and can be considered as indicator
. . ^ .
des~n~
to saltans
niomonas truncata
tlpingoeca spp.
7yathobodo sp. ~pongomonas sacculus Rhipidodendron huxleyi
Uroleptus caudatus
9phosis sp.
,lophosis mucicola
(yophrya sphagni
iilodonella uncinata
Sep. Oct. Nov. Dec. J/F Mar. Apr. May Jun. Jul. Aug.
.eptopharynx costatus Ophrydium hyalinum Vorticella campanula Vortic¢lla convaUaria- complex
Sep.
Fig. 7. Abundances (cm -2) and seasonal distribution of the characteristic ciliate and flagellate species in Lake GroBer Barschsee, site B. The abundances of Spongomonas sacculus and Rhipidodendron hualeyi are noted as colonies/cm 2.
Limnologica 29 (1999) 4 419
gnis !ans ,nas truncata
~eca spp. monas sacculus
9phosis sp.
donella uncinata
opharynx costatus
Sep. Oct. Nov. Dec. J/F Mar. Apr. May Jun. Jul. Aug.
9phrydium hyalinum Vorticella campanula Vorticella convallaria- complex
Sep.
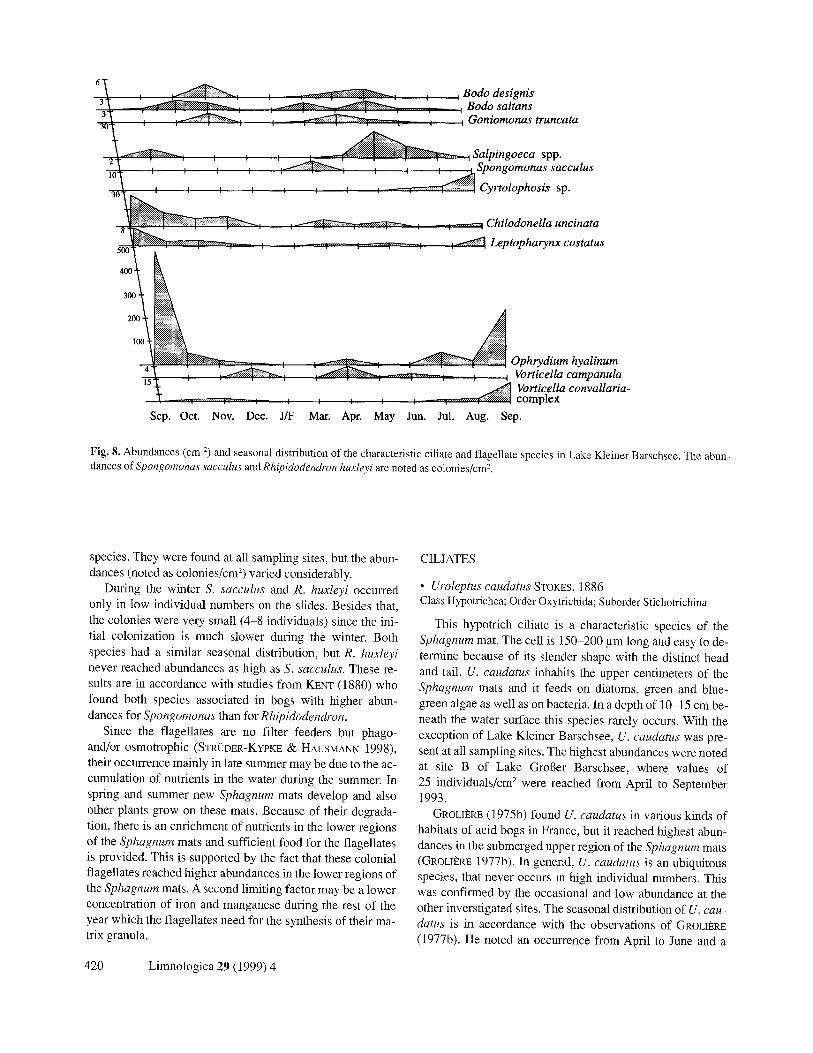
Fig. 8. Abundances (cm 4) and seasonal distribution of the characteristic ciliate and flagellate species in Lake Kleiner Barschsee. The abun- dances of Spongomonas sacculus and Rhipidodendron huxleyi are noted as colonies/cm 2.
species. They were found at all sampling sites, but the abun- dances (noted as colonies/cm 2) varied considerably.
During the winter S. sacculus and R. huxleyi occurred only in low individual numbers on the slides. Besides that, the colonies were very small (4-8 individuals) since the ini- tial colonization is much slower during the winter. Both species had a similar seasonal distribution, but R. huxleyi never reached abundances as high as S. sacculus. These re- sults are in accordance with studies from KENT (1880) who found both species associated in bogs with higher abun- dances for Spongomonas than for Rhipidodendron.
Since the flagellates are no filter feeders but phago- and/or osmotrophic (STR~DER-KYPKE & HAUSMANN 1998), their occurrence mainly in late summer may be due to the ac- cumulation of nutrients in the water during the summer. In spring and summer new Sphagnum mats develop and also other plants grow on these mats. Because of their degrada- tion, there is an enrichment of nutrients in the lower regions of the Sphagnum mats and sufficient food for the flagellates is provided. This is supported by the fact that these colonial flagellates reached higher abundances in the lower regions of the Sphagnum mats. A second limiting factor may be a lower concentration of iron and manganese during the rest of the year which the flagellates need for the synthesis of their ma- trix granula.
CILIATES
• Uroleptus caudatus STOKES, 1886 Class Hypotrichea; Order Oxytrichida; Suborder Stichotrichina
This hypotrich ciliate is a characteristic species of the Sphagnum mat. The cell is 150-200 pm long and easy to de- termine because of its slender shape with the distinct head and tail. U. caudatus inhabits the upper centimeters of the Sphagnum mats and it feeds on diatoms, green and blue- green algae as well as on bacteria. In a depth of 10-15 cm be- neath the water surface this species rarely occurs. With the exception of Lake Kleiner Barschsee, U. caudatus was pre- sent at all sampling sites. The highest abundances were noted at site B of Lake Grol3er Barschsee, where values of 25 individuals/cm 2 were reached from April to September 1993.
GROLIE~E (1975b) found U. caudatus in various kinds of habitats of acid bogs in France, but it reached highest abun- dances in the submerged upper region of the Sphagnum mats (GROLIERE 1977b). In general, U. caudatus is an ubiquitous species, that never occurs in high individual numbers. This was confirmed by the occasional and low abundance at the other inverstigated sites. The seasonal distribution of U. cau- datus is in accordance with the observations of GROLIERE (1977b). He noted an occurrence from April to June and a
420 Limnologica 29 (1999) 4

peak in May 1993. The observed high individual numbers from April to September 1993 correlate with the generally high abundances of ciliates in the periphyton. At this time, sufficient coccoid green algae colonized the slides and pre- sented the main food source of U. caudatus.
Striking was the presence of two morphotypes: a colour- less, slender form and a green, more rounded form with zoochlorellae. Also FO~SSNER (1980) did not exclude the oc- currence of zoochlorellae-harbouring specimens within this ciliate species. This chloroplast retention or facultative sym- biosis with zoochlorellae represents an adaptation to rather oligotrophic biotops (HARNISCH 1929).
• Cyrtolophosis sp. and Cyrtolophosis mucicola STOKES, 1885 Class Colpodea; Subclass Colpodia; Order Cyrtolophosidida
These small, only 20-28 pm long ciliate species are vagile and glide over the substrate. C. mucicola builds small hyaline loricas, which could not be definitely identified for Cyrtolophosis sp., although there are some evidences that they may exist for both species. The infraciliature is com- prised of few, slightly spirally running kineties. Anterior is the buccal region with 3-5 adoral membranelles located. The distinguishing character of the two species are few (10-15) spherical symbiontic green algae in the cytoplasm of Cyr- tolophosis sp. which may be zoochlorellae. Zoochlorellae- harbouring species of Cyrtolophosis haven't been described yet. The investigated species, however, showed a high con- stancy in number and position throughout the cytoplasm of the internal green algae. Apart from that, no digestion stages of them were found, suggesting a symbiosis with zoochlorel- lae. It remains to be solved, if this symbiosis with zoo- chlorellae is only an adaption of C. mucicola to oligotrophic environments (comparable to U. caudatus), or if the erection of a new species is justified. C. mucicola is abundant in most fresh-waters and can be considered as ubiquitous. FOISSNER et al. (1991) and FOISSNER (1993) noted the saprobity as oligo- to polysaprobic. Cyrtolophosis sp. and C. mucicola are species that inhabited mainly the upper region of the Sphagnum mat. They were not represented in the periphyton of Lake Teufels-See. Both species reached highest abun- dances in May 1993 at site B of Lake GroBer Barschsee. During the winter and at the other sampling sites, Cyr- tolophosis occurred only occassionally.
The milieu spectrum of C. mucicola is widespread, there- fore high abundances are not related to certain abiotic fac- tors, but to the lack of competitors (FoISSNER et al. 1991; FOISSNER 1993). This explains the low individual numbers, since the slides were colonized by many different protists. GROLIERE (1977b) noted the highest abundances of C. muci- cola at water temperatures below 10 °C. This observation could not be confirmed by our investigations, since the species also occurred during the summer months at higher temperatures, up to 20 °C.
• Platyophtya sphagni (PENARD, 19221) FOISSNER, 1993 Class Colpodea; Subclass Colpodia; Order Cyrtolophosidida
The third colpodid ciliate is metabolic, 50-70 pm long and harbours numerous zoochlorellae in the cytoplasm. P. sphagni was repeatedly found in Sphagnum bogs and there- fore it can be considered as indicator species for this habitat. The species occurred in September 1992 and from May to September 1993 on the slides of site B in Lake Groger Barschsee. The abundances ranged from 10-50 individu- als/cm 2. During the winter it was not present on the slides.
Platyophrya sphagni was described from Sphagnum bogs and small ponds (FoISSNER 1993; GROLII~RE 1977a). Its re- striction to the upper regions of the Sphagnum mat is in ac- cordance to observations of HARNISCH (1929) and GROL~ERE (1977b).
• Chilodonella uncinata EHRENBERG, 1838 Class Phyllopharyngea; Subclass Cyrtophoria; Order Chilodonellida
This species is ubiquitous and colonizes many different habitats. C. uncinata is 30-50 pm long and several kineties of varying length are running over the ventral side. Dorsally, there are no kineties but 2 dorsal brushes. The cytopharynx is supported by a basket of microtubules. C. uncinata was one of the few species that occurred throughout the year in the periphyton and was observed at all sampling sites. The abun- dances were relative low, peaks were reached in the fall of 1992 and in the spring of 1993.
As a typical pioneer species on newly exposed substrates, Chilodonella uncinata grazes on bacteria of the surface of the substrates and is most frequently found during the initial phase of the colonization. KAHL (1931) described C. uncina- ta from bog lakes.
• Leptopharynx costatus MERMOD, 1914 Class Nassophorea; Subclass Nassulia; Order Microthoracida
This species is very small (20-40 pm long) and vagile. Few kineties are curved on the ventral side of the cell. Ante- rior to the oral region, oblique running ciliary rows form a dense tuft of cilia. Most reports of L. costatus are from ter- restrial biotopes and from mires. Together with other species it can be considered as an indicator species for acid, dys- trophic habitats. L. costatus was found throughout the year at all sampling sites. The abundances were very low within the range of 1-8 individuals/cm 2.
GROLI~RE (1977b) observed high abundances of Lep- topharynx costatus in various bogs in France, mainly in September and October, as well as partly in May. During our investigations, however, L. costatus occurred regularly but in low abundances (often as single observation) on the slides. Only in the upper regions of the Sphagnum mats, a compara- ble seasonal distribution was observed. FOISSNER et al. (1994) ranked this species as oligo- to beta-mesosaprobic. Because of the regular occurrence almost exclusively in
Limnologica 29 (1999) 4 421

Sphagnum bogs, L. costatus can be considered as indicator species of this habitat (GROLIgRE 1978). Its generally low in- dividual numbers are in accordance with the results of BAM- FORTH (1982) who compared different artificial substrates: only approximately 50% of all heterotrophic vagile species were observed on glass-slides.
• Ophrydium hyalinum WRZESNIOWSKI, 1877 Class Oligohymenophorea; Subclass Peritrichia; Order Sessilida
This sessile, colonial peritrich ciliate resembles the better known species O. versatile. In contrary to this species, how- ever, it harbours no zoochlorellae and its spherical colonies reach only maximum sizes of a few millimeters in diameter. The extended cell is approximately 200 pm long. At the pos- terior ends the cells are linked by dichotomously branched strands. Originally described by WRZESN~OWSKI (1877) from a small pond, O. hyalinum can be considered as an indicator species of acid, dystrophic waters. It was found in all lakes, but in varying abundances. Maximal individual numbers were reached in September 1992 as well as in May and September 1993.
Among the ciliates, the lower regions of the Sphagnum mats were inhabited with a high percentage by sessile species. Vagile and free-swimming species were only found in low abundances. As mentioned before, the use of of glass- slides as artificial substrates prefers sessile species (BAM- FORTH 1982; WEITZEL et al. 1979; WILBERT 1969). Apart from that, large sessile filter feeders like peritrich ciliates find more food in this region of the Sphagnum mats than in the upper, green horizon. Ophrydium hyalinum reached highest abundances in Lake Kleiner Barschsee, in which only rarely vorticellids were observed. Explanations for the repression of the peritrichs during the summer may be the predation pressure by metazoans like rotifers and cladocer- ans (ARNDT 1993; WmBERT 1969) or the lack of nutrients and insufficient bacterial densities during this time.
• Vorticella campanula EHRENBERG, 1831 and Vorticella convallaria-complex
Class Oligohymenophorea; Subclass Peritrichia; Order Sessilida
V. campanula is 50-150 pm long and the cytoplasm ap- pears dark due to lipid droplets. Therefore it is easy to distin- guish from other Vorticella species. The species of the V. convallaria complex, however, are hard to determine by means of live observations. They are bell-shaped, 40-95 pm long and 35-45 pm broad. The length of the stalk is very variable. The macronucleus of J-shaped and the rim of the peristom distinctly surpasses the body. Both Vorticella species - as other peritrichs - are ubiquitous and very com- mon in many fresh-waters. The species were found in all lakes with varying abundances.
Vorticella campanula does not only feed on bacteria, but also on algae and detritus. The species is very common and indicates mesosaprobic habitats. The annual cycle reached a
peak in early spring. In contrary, the species of the V. conva- llaria complex showed highest abundances in early summer. Since the nutritional needs are approximately the same for both species, the change in dominance may be due to abiotic factors. Eventually the acidification of the water by the growing Sphagnum plants during the summer is one reason for the decrease in the individual numbers of V. campanula, since this species avoids acid habitats (FoISSNER et al. 1992).
Trophy of the investigated bog lakes The investigations of the periphyton community and the tes- tate amoebae, as well as general observations point to a eu- trophication of the bog lakes during the last 20 years.
The Sphagnum mat became 25-50% smaller, Ledum palustre advanced towards the lake, many other herbs grow on the Sphagnum mat that were not abundant before. Al- though Drosera plants are still optimally distributed within the bogs, the other plants can be considered as a source for the formation of detritus and for the raising of the pH (from 5.2-5.7 in 1963 to 6.3-6.8 in 1996). This, however, enables a better degradation of Sphagnum and other plant parts and leads to an increasing trophy of the lakes. Also the low C:N- ratio of the sediment may be due to the invasion of different herbs and resulted in a further fertilization of the bogs. Nor- mally, Sphagnum residues reveal a C:N-ratio of >30 and are therefore difficult to decay. Subsequently, bacteria, het- erotrophic flagellates and higher organisms colonized the Sphagnum and repressed the original community. The changes in the distribution and occurrence of some species of testate amoebae as well as the protist species composition in the periphyton also indicate more mesotrophic conditions in the investigated bog lakes. Although still a vertical zoning exists, more and more heterotrophic species and such with- out zoochlorellae are found in the upper region of the Sphag- num mats. The lower regions are definitely mesotrophic. Also the comparison with other studies (e.g. GROLI~RE) shows distinct differences in the characteristic species which may in part be due to the eutrophication of the investigated bogs. Since the bog lakes are located within large forest areas, they show no direct anthropogenic sources which may have caused this eutrophication. Therefore, the fertilization only could have happened through the air (entry approxi- mately 20 kg N/ha. a) or climatic changes. In the past in- creasing dryness frequently has been resulted in a rapid eu- trophication of Sphagnum bogs, because herbs and trees could colonize the mats.
Acknowledgements: The study was carried out at the Institute of Zoology, Free University of Berlin, Germany. It was supported by grants from the State of Berlin ("Nachwuchsf6rdertmgsgesetz" and "Sonderprogramm zur F/Srderung von Nachwuchswissenschaft- lerinnen") to M. C. STRUDER-KYPKE. We wish to thank Prof. Dr. K. HAUSMANN who sustained this study on a large scale and in whose laboratory most of the work was done. Permissions to take samples
422 Limnologica 29 (1999) 4

from the protected lakes were given by the "Ministerium ftir Umwelt, Naturschutz und Raumordnung des Landes Brandenburg - Abt. Naturschutz - " and the "Amt ftir Forstwirtschaft Fttrstenberg". The support given by the "Forst~imter" in Menz, Schgnhom and Zechlinerhtitte is appreciated. Prof. Dr. R. KOSCnEL and Mr. W. SCHEFFLER from the Institute of Freshwater Ecology and Inland Fisheries, Department of Limnology in Neuglobsow, helped with valuable informations concerning the investigated area.
References
ARNDT, H. (1993): Rotifers as predators on components of the microbial web (bacteria, heterotrophic flagellates, ciliates) - a re- view. Hydrobiologia 285/286:231-246.
- & BEftNINGER, U.-G. (1995): Protists in aquatic food webs - com- plex interactions. In: G. BRUGEROLLE & J.-R MIGNOT (eds), Pro- tistological Actualities (Proceed. 2nd Europ. Congr. Protistol., Clermond-Ferrand 1995), pp. 224-232.
BAMFORTH, S. S. (1982): The variety of artificial substrates used for microfauna. In: J. Jr. CAIRNS (ed.), Artificial substrates, pp. 1 1 5 - 1 3 0 . Ann Arbor, Michigan.
CAIRNS, J. Jr. & HENEBRY, M. S. (1982): Interactive and noninterac- rive protozoan colonization processes. In: J. CAIRNS Jr. (ed.), Arti- ficial substrates, pp. 27-30. Ann Arbor, Michigan.
CASPER, S. J. & SCHEFFLER, W. (1990): Diatom analysis, late-glacial and post-glacial development of Lake Kleiner Barsch-See (GDR) - a preliminary note. Limnologica 21 (1): 147-155.
ETTL, H. (1983): Chlorophyta I. Phytomonadina. In: H. EarL, J. GERLOFF, H. HEYNIG & D. MOLLENHAUER (eds.), Stigwasserflora von Mitteleuropa Vol. 9. Stuttgart.
- & GARTNEN, G. (1988): Chlorophyta II. Tetrasporales, Chlorococ- cales, Gloeodendrales. In: H. ETTL, J. OERLOFF & D. MOLLEN- HAUER (eds.), Stigwasserflora von Mitteleuropa Vol. 10. Stuttgart.
FOISSNER, W. (1980): Taxonomische Studien fiber die Ciliaten des Grogglocknergebietes (Hohe Tauem, (3sterreich). IX. Ordnungen Heterotrichida und Hypotrichida. Ber. Nat.-Med. Ver. Salzburg 5 : 71-117.
- (1993): Colpodea (Ciliophora). In: D. MAArHES (ed.), Protozoen- fauna Vol. 4/1. Stuttgart.
- BERGEN, H. & KOHMANN, F. (1992): Taxonomische und 6kologi- sche Revision der Ciliaten des Saprobiensystems. II. Peritrichia, Heterotrichida, Odontostomatida. Informationsberichte des Bayer. Landesamtes ftir Wasserwirtschaft, Miinchen, 5/92.
- BERGEN, H. & KOHMANN, F. (1994): Taxonomische und 6kologi- sche Revision der Ciliaten des Saprobiensystems. III. Hymeno- stomata, Prostomatida, Nassulida. Informationsberichte des Bayer. Landesamtes fiir Wasserwirtschaft, Mtinchen, 1/94.
-- BERGEN, H., BLATTERER, H. & KOHMANN, E (1995): Taxonomische und 6kologische Revision der Ciliaten des Saprobiensystems. IV. Gymnostomatea, Loxodes, Suctoria. Informationsberichte des Bayer. Landesamtes fiJr Wasserwirtschaft, Mtinchen, 1/95.
- BLATTERER, H., BERGER, H. & KOI-IMANN, E (1991): Taxono- mische und /Skologische Revision der Ciliaten des Saprobien- systems. I. Cyrtophorida, Oligotrichida, Hypotrichia, Colpodea. Informationsberichte des Bayer. Landesamtes ftir Wasserwirt- schaft, Mtinchen, 1/91.
G~3TTLICn, K. (ed.) (1990): Moor- und Torfkunde (3. Aufl.). Stuttgart. GRASSI~, R-R (ed.) (1994): Trait6 de Zoologie. Infusoires Cilids.
Tome II, Fasc. 2. Paris.
GROLII~RE, C.-A. (1975a): 12tude de quelques cilids hymdnostomes des eaux acides de la region de Besse-en-Chandesse. Ann. Stat. Biol. Besse-en-Chandesse 9: 79-109.
- (1975b): Descriptions de quelques ciligs hypotriches des tour- bi~res a sphaignes et des 6tendues d'eau acides. Protistologica 11 (4): 481-498.
- (1977a): Contribution a l'6tude des ciligs des sphaignes et des 6tendues d'ean acides. I. Description de quelques esp~ces de gymnostomes, hypostomes, hymgnostomes et hgtgrotriches. Ann. Stat. Biol. Besse-en-Chandesse 10: 265-297.
- (1977b): Contribution a l'gtude des ciligs des sphaignes: II. Dy- namique des populations. Protistologica 13 (3): 335-352.
- (1978): Contribution a l'6tude des ciligs des sphaignes: III. t~tude mathgmatique des r6sultats. Protistologica 14 (3): 295-311.
HARNISCH, O. (1925): Studien zur C)kologie und Tiergeographie der Moore. Zool. Jb. (Syst) 51: 1-166.
- (1927): Einige Daten zur rezenten und fossilen testaceen Rhi- zopodenfauna der Sphagnen. Arch. Hydrobiol. 18: 343-360. (1929): Die Biologic der Moore. In: A. THIENEMANN (ed.), Die Binnengew~sser 7, pp. 1-146. Stuttgart.
HAUSMANN, K. & HOLSMANN, N. (1996): Protozoology (2nd ed.), Stuttgart.
HESSEN, D. O., ANDERSEN, T. & LYCHE, A. (1990): Carbon meta- bolism in a humic lake: pool sizes and cycling through zooplank- ton. Limnol. Oceanogr. 35 (1): 84-99.
HIBBERD, D. J. (1976a): The fine structure of the colonial colorless flagellates Rhipidodendron splendidum STEIN and Spongomonas uvella STEIN with special reference to the flagellar apparatus. J. Protozool. 23 (3): 374-385.
- (1976b): Observations on the ultrastructure of three new species of Cyathobodo PETERSEN and HANSEN (C. salpinx, C. intricatus and C. simplex) and on the external morphology of Pseudoden- dromonas vlkii BOURRELLY. Protistologica 12 (2): 249-261.
- (1983): Ultrastructum of the colonial colourless zooflagellates Phalansterium digitatum STEIN (Phalansteriida ord. nov.) and Spongomonas uvella STEIN (Spongomonadida ord. nov.). Protis- tologica 14 (4): 523-535.
- (1985): Observations on the ultrastructure of new species of Pseu- dodendromonas BOURRELLY ( P. operculifera and P . insignis) and Cyarhobodo PETERSEN and HANSEN (C. peltams and C. gemma- tus), Pseudodendromonadida ord. nov. Arch. Protistenkd. 129: 3-11.
HUBER-PESTALOZZl, G. (1941): Das Phytoplankton des Siigwassers. Chrysophyceen, farblose Flagellaten, Heterokonten. In: A. THIE- NEMANN (ed.), Die Binnengew~isser 16 (2/1), pp 1-247. Stuttgart.
- (1955): Das Phytoplankton des Siigwassers. Euglenophyceen. In: A. THIENEMANN (ed.), Die Binnengewfisser 16 (4), pp 1-606. Stuttgart.
- (1968) : Das Phytoplankton des Stigwassers. Cryptophyceae, Chloromonadophyceae, Dinophyceae (2nd edition). In: H.-J. EL- STER & W. OHLE (eds), Die Binnengewfisser 16 (3), pp 1-322. Stuttgart.
JESCHKE, L. (1987): Unsere Ackersglle und ihre Funktion in der Landschaft. Naturschutzarbeit in Mecklenburg 30: 29-33.
KAHL, A. (1930): Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infusoria) 1. Allgemeiner Teil und Prostomata. In: E DAHL (ed.), Die Tierwelt Deutschlands 18, pp 1-180. Jena
- (1931): Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infuso- ria) 2. Holotricha auger den im ersten Teil behandelten Prostoma- ta. In: E DAI4L (ed.), Die Tierwelt Deutschlands 21, pp 181-398. Jena
Limnologica 29 (1999) 4 423

- (1932): Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infuso- ria) 3. Spirotricha. In: E DAI-IL (ed.), Die Tierwelt Deutschlands 25, pp 399-650. Jena
- (1935): Urtiere oder Protozoa I: Wimpertiere oder Ciliata (Infuso- ria) 4. Peritricha und Chonotricha. In: F. DAHL (ed.), Die Tierwelt Deutschlands 30, pp 651-886. Jena
KENT, W. S. (1880-1882): A manual of the infusoria. London. LEMMERMANN, E. (1914): Flagellatae I. Pantostomatinae, Proto-
mastiginae, Distomatinae. In: A. PASCHER (ed.), Die SiiBwasser- flora Deutschlands, 0sterreichs und der Schweiz 1, pp 28-137. Jena
LEVANDEI~, K. M. (1900): Zur Kenntnis des Lebens in den stehenden Kleingew~issern auf den Sk~eninseln. Acta soc. Fauna Flora Fen- nica 18: 3-107.
MEISTERFELD, R. (1977): Die horizontale und vertikale Verteilung der Testaceen (Rhizopoda, Testacea) in Sphagnum. Arch. Hydro- biol. 79 (3): 319-356.
- (1979a): Zur Systematik der Testaceen (Rhizopoda, Testacea) in Sphagnum. Eine REM-Untersuchung. Arch. Protistenkd. 121: 246-269.
-(1979b): Clusteranalytische Differenzierung der Testaceen- zi3nosen (Rhizopoda, Testacea) in Sphagnum. Arch. Protistenkd. 121: 270-307.
MERMOD, G. (1914): Recherches sur la fanne infusorienne des tour- bi5res et des eaux voisines de Sainte-Croix (Jura vaudois). Rev. Suisse Zool. 22: 31-114.
MIGNOT, J. R (1974): l~tude ultrastructurale d'un protiste flagell6 in- colore: Pseudodendromonas vlkii BOURRELLY. Protistologica 10 (3): 397-412.
PATTERSON, D. J. & LA~SEN, J. (1991): The biology of free-living heterotrophic flagellates. The Systematic Association, Special Vol. 45. Oxford.
PENARD, E. (1922): l~tudes sur les infusoires d'eau douce. Gen6ve. POPOVSKY, J. & PFIESTER, L. A. (1990): Dinophyceae. In: H. ETTL, J.
GERLOFF, H. HEYNIG & D. MOLLENttAUER (eds.), Sfigwasserfora yon Mitteleuropa 6, pp 1-288. Stuttgart.
RAILKIN, A. I. (1995): Heterotrophic flagellates on artificial sub- strates in the White Sea. Cytology 37 (11): 951-957.
SANDERS, R. W. & PORTER, K. G. (1990): Relationship between pho- totrophy and phagotrophy in the mixotrophic chrysophyte Poteri- oochromonas malhamensis. Microbiol. Ecol. 19: 97-109.
- - BENNETT, S. J. & DEBIASE, A. E. (1989): Seasonal patterns of bacterivory by flagellates, ciliates, rotifers, and cladocerans in a freshwater planktonic community. Limnol. Oceanogr. 34 (4): 673-687.
SCHLENKER, G. (1908): Das Schwenninger Zwischenmoor und zwei Schwarzwald-Hochmoore im Bezug auf ihre Entstehung. Pflan- zen und Tierwelt. Geol.-biol. Untersuch. von Torfmooren 3: 1-279.
SCHONBORN, W. (1962a): Neue Testaceen aus dem Grogen Stechlin- see und dessen Umgebung. Limnologica 1 (1): 83-91.
- (1962b): Zur Okologie der sphagnikolen, bryokolen und terri- kolen Testaceen. Limnologica 1 (3): 231-254.
- (1963): Die Stratigraphie lebender Testaceen im Sphagnetum der Hochrnoore. Limnologica 1 (4): 315-321.
- (1990): Analyse subfossiler Protozoenschalen der Sedimente eines kleinen sauren Waldsees (Kleiner Barsch-See, n6rdliche DDR). Limnologica 21 (1): 137-145.
- FLOSSNER, D. & PROFT, G. (1965): Die limnologische Charakter- isierung des Profundals einiger norddeutscher Seen mit Hilfe von Testaceen-Gemeinschaften. Limnologica 3:371-380.
SHERR, E. B. (1988): Direct use of high molecular weight poly- saccharide by heterotrophic fagellates. Nature 335:348-351.
SKUJA, H. (1948): Taxonomie des Phytoplanktons einiger Seen in Uppland, Schweden. Symb. Bot. Upsalensis 9: 1-399.
- (1956): Taxonomische und biologische Studien fiber das Phyto- plankton schwedischer Binnengew~sser. Nova Acta Reg. Soc. Sci. Upsalensis (Ser. 4) 16: 1-404.
- (1964): Grundzfige der Algenflora und Algenvegetation der Fjeldgegenden um Abisko in Schwedisch-Lappland. Nova Acta Reg. Soc. Sci. Upsalensis (Ser. 4) 18: 1-465.
SLADECEK, V. (1973): System of water quality from the biological point of view. Arch. Hydrobiol., Beth. Ergebn. Limnol. Plank- tonkd. 7: IV and 1-218.
STARMACH, K. (1985): Chrysophyceae nnd Haptophyceae. In: H. ETTL, J. GERLOFF, H. HEYNIG & D. MOLLENHAUER (eds.), StifS- wasserflora von Mitteleuropa 1, pp 1-515. Stuttgart.
STRf2TDER-KYPKE, M. C. (1999): Periphyton and sphagnicolous pro- tists of dystrophic bog lakes. I. Annual cycles, distribution and comparison to other lakes. Limnologica 29: 393-406. & HAUSMANN, K. (1998): Ultrastructure of the heterotrophic flagellates Cyathobodo sp., Rhipidodendron huxleyi KENT, 1880, Spongomonas sacculus KENT, 1880, and Spongomonas sp. Europ. J. Protistol. 34: 376-390.
Succow, M. (1987): Zur Entstehung und Entwicklung der Moore der DDR. Z. geol. Wiss. 15: 373-387.
- & JESCHKE, L. (1990): Moore in der Landschaft (2. Aufl.). Leip- zig, Jena, Berlin.
THIEBAUD, M. & FAVRE, J. (1906): Sur la faune invert6br6e des mares de Pouilleret. Zool. Anz. 30: 155-163.
WEITZEL, R. L., SANOCKI, S. L. & HOLECEK, H. (1979): Sample replication of periphyton collected from artificial substrates. In: R. L. WEITZEL, (ed.), Methods and measurements of periphyton communities: A review. American Society for Testing and Mate- rials, pp. 90-115.
WENZEL, F. (1953): Die Ciliaten der Moosrasen trockener Standorte. Arch. Protistenkd. 99: 70-141.
WILI3ERT, N. (1969): Okologische Untersuchung der Aufwuchs- und Planktonciliaten eines eutrophen Weihers. Arch. Hydrobiol. / Suppl. 35: 411-518.
- (1976): A standardized method for identifying and counting the vagile and sessile periphyton. Oecologia 24: 343-347.
W~ZESN~OWSK~, A. (1877): BeitrSge zur Naturgeschichte der Infu- sorien. Z. wiss. Zool. 29: 267-323.
ZOLOTAREV, V. (1995): Microperiphyton flagellate communities, their characteristics and functional groups. Cytology 37 (11): 958-963.
Received: September 2, 1998
Authors' addresses: Dr. MICHAELA STRISDER-KYPKE, College of Biological Science, Department of Zoology, University of Guelph, Guelph, Ontario N1G 2W1, Canada. e-mail: [email protected] Dr. habil. WILFRIED SCHONBORN, Friedrich-Schiller-Universitfit Jena, Institut ffir 0kologie, AG Limnologie, Winzerlaer Str. 10, D-07745 Jena, Germany.
424 Limnologica 29 (1999) 4