Embed Size (px)
Citation preview

MHP1, an Essential G-ene in Saccharomyces cerevisiae Required for Microtubule Function Irmgard Irminger-Finger,* Ed Hurt , ~ Aline Roebuck,* Martine A. Collart*, and Stuart J. Edelstein*
*Department of Biochemistry, *Department of Medical Biochemistry, University of Geneva, 1211 Geneva 4, Switzerland; and ~EMBL, 69012 Heidelberg, Germany
Abstract. The gene for a microtubule-associated pro- tein (MAP), termed MHP1 (MAP-Homologous Pro- tein 1), was isolated from Saccharomyces cerevisiae by expression cloning using antibodies specific for the Drosophila 205K MAP. MHP1 encodes an essential protein of 1,398 amino acids that contains near its COOH-terminal end a sequence homologous to the mi- crotubule-binding domain of MAP2, MAP4, and tau. While total disruptions are lethal, NH2-terminal dele- tion mutations of MHP1 are viable, and the expression of the COOH-terminal two-thirds of the protein is suf- ficient for vegetative growth. Nonviable deletion-dis- ruption mutations of MHP1 can be partially comple- mented by the expression of the Drosophila 205K MAP. Mhplp binds to microtubules in vitro, and it is the COOH-terminal region containing the tau-homolo-
gous motif that mediates microtubule binding. Anti- bodies directed against a COOH-terminal peptide of Mhplp decorate cytoplasmic microtubules and mitotic spindles as revealed by immunofluorescence micros- copy. The overexpression of an NH2-terminal deletion mutation of MHP1 results in an accumulation of large- budded cells with short spindles and disturbed nuclear migration. In asynchronously growing cells that overex- press MHP1 from a multicopy plasmid, the length and number of cytoplasmic microtubules is increased and the proportion of mitotic cells is decreased, while hap- loid cells in which the expression of MHP1 has been si- lenced exhibit few microtubules. These results suggest that MHP1 is essential for the formation and/or stabili- zation of microtubules.
M ICROTUBULES (MTs) 1 are involved in a number of essential cellular processes such as cell mor- phogenesis, intracellular transport, cell motility,
and mitosis. Important potential regulators of MT dynam- ics are the MT-associated proteins (MAPs) (Lee, 1993; Wiche et al., 1991). Initially MAPs were isolated from neu- ral tissues rich in MTs by means of copurification with poly- merized MTs. These neural MAPs, including MAP 1A (Schoenfeld et al., 1989), MAP 1B (Noble et al., 1989), MAP2 (Lewis et al., 1988), and tau (Lee et al., 1988), were shown to stimulate MT assembly from purified tubulin in vitro (Cleveland et al., 1977; Murphy et al., 1977; Bulinski and Borisy, 1979). Although these MAPs have been char- acterized biochemically, their specific functions have not been determined unambiguously.
Address all correspondence to Stuart J. Edelstein, Department of Bin- chemistry, University of Geneva, 30 quai Ernest-Ansermet, 1211 Geneva 4, Switzerland. Tel.: (41) 22 702 64 86. Fax: (41) 22 702 64 76. e-mail: Stuart. [email protected]
1. Abbreviations used in this paper. ADH, alcohol dehydrogenase; AP, as- sembly promoting; DAPI, 4',6-diamidino-2-phenylindole; MAP, microtu- ble-assoeiated protein; MT, mierotubnie; NLS, nuclear localization signal; ORF, open reading frame; SPB, spindle pole body.
A group of ubiquitously expressed, thermostable MAPs has also been described as phosphorylated proteins of ,-.~200 kD (MAP4-type proteins) comprising (as in the case of MAP1, MAP2, and tau) multiple protein isoforms. Im- munocytochemistry has shown that these MAPs colocalize with mitotic spindle and interphase MTs (Bulinski and Borisy, 1980; Izant et al., 1983). Cloning and sequencing of mouse, human (Chapin and Bulinski, 1991; West et al., 1991), and bovine (Aizawa et al., 1990) MAP4 have re- vealed that the MAP4 proteins share an MT-binding do- main homologous to the MT-binding motif first discov- ered in tau and MAP2 (Lee et al., 1988; Lewis et al., 1988). The common sequence motif, the assembly promoting (AP) sequence (Aizawa et al., 1989), consists of three or four basic repeats of 31 amino acid residues within the COOH-terminal domain of the proteins.
In several species MAPs of ,~200 kD have been discov- ered that are immunologically related to the Drosophila 205K MAP but not to MAP4-type proteins (Kimble et al., 1992). The Drosophila 205K MAP was analyzed and found to contain no sequence homologies to other MAPs. Moreover, it possesses an MT-binding region without ho- mology to tau, MAP2, or MAP4 (Irminger-Finger et al., 1990). However, the 205K MAP is similar to MAP4 and MAP2 in terms of an extensive NH2-terminal acidic region
© The Rockefeller University Press, 0021-9525/96/11/1323/17 $2.00 The Journal of Cell Biology, Volume 135, Number 5, December 1996 1323-1339 1323
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

and, as in the case of tau, a short, acidic COOH-terminal region that is preceded by a basic region containing the MT-binding domain.
The in vivo functions of all the MAPs identified so far remain uncertain, mainly because of the absence of ge- netic studies. In the isolated case of the Drosophila 205K MAP where such studies were possible, the protein was found not to be essential (Pereira et al., 1992). A powerful tool to more fully investigate the functions of MAPs is the yeast Saccharomyces cerevisiae, since it is possible to es- tablish genetic interactions between MAPs and tubulin or other cytoskeletal components. A number of laboratories have attempted to identify MAPs in yeast. Their ap- proaches have involved purifying proteins by MT-affinity columns (Barnes et al., 1992) or from cellular structures comprising MTs such as the spindle pole body (SPB) (Kil- martin et al., 1993) and the centromere-binding complex (Jiang et al., 1993), identifying yeast proteins that cross- react with antibodies against mammalian MAPs (Hasek et al., 1991), cloning genes complementing cellular defects in- volving MTs (Hoyt et al., 1990; Stearns et al., 1990; Mc- Millan and Tatchell, 1994), and cloning yeast sequences using PCR with degenerated primers derived from homol- ogous sequences of different species (Hyman et al., 1992). These strategies led to the identification of CPF5p, a com- ponent of the yeast centromere-binding complex, a homo- logue of mammalian MAP1A and B (Jiang et al., 1993), and a number of genes encoding proteins that share simi- larities with motor proteins (Berlin et al., 1990; Meluh and Rose, 1990; Hoyt et al., 1992; Lillie and Brown, 1992; Roof et al., 1992; Li et al., 1993). In a biochemical approach, Sepl has been identified as an accessory protein in MT function in yeast (Interthal et al., 1995). A genetic screen for complementation of a temperature-sensitive allele of the 13-tubulin gene led to the identification of STU1, a gene encoding a protein with a predicted size of 174 kD and no significant homology to known proteins (Pasqua- lone and Huffaker, 1994).
We chose to search for yeast proteins that cross-react with antibodies directed against the Drosophila 205K MAP, leading to the discovery of a novel essential MAP gene, MHP1 (MAP Homologous Protein 1). We present evidence that the product of this gene, Mhplp, interacts with MTs and that mutations in MHP1 influence MT structure and function during interphase and mitosis.
Materials and Methods
Strains and Microbiological Techniques All yeast strains used in this study are listed in Table I. Genetic manipula- tions and growth media were standard (Sherman et al., 1986; Rose et al., 1990).
Cloning of MHP1 A partial sequence of the MHP1 gene was cloned by screening a kgtl l eDNA expression library from S. cerevisiae (kindly provided by Dr. B. Altman, University of Bern, Switzerland). 20,000 plaques were screened and plated in Y1090r- host cells to a plaque density of 1,000-2,000 plaques per plate. Plates were incubated at 42"C for 4 h, or until very small plaques were visible, then overlaid with nitrocellulose filters previously soaked in 10 mM isopropy113-t~-thiogalactopyranoside (IPTG), and incubated for 4 h at 37°C. Filters were blocked with 3% BSA in TBS for 30 min, washed
with TBS, and incubated with a polyclonal anti-205K MAP antibody (Goldstein et al., 1986) in a 1:500 dilution for 2 h at room temperature. Af- ter washing with TBS, the secondary antibody, a mouse anti-IgG coupled to alkaline phosphatase (Sigma Chemical Co., St. Louis, MO), was ap- plied in a 1:5,000 dilution. Positive phages were visualized with 5-bromo- 4-chloru-3-indolylphosphate disodinm salt (BCIP) and nitro blue tetrazo- lium (NBT) (Sigma Chemical Co.). Plaques in the region of the signal were resuspended in phage buffer (10 mM Tris, pH 7.5, 10 mM MgC12), replated, and rescreened with the same procedures. Three positive plaques were obtained and tested for the expression of the immunoreac- tive protein on Western blots. The insert of one phage was subcloned into the Bluescript-vector (pBS) and sequenced, the 740-bp insert was used as radioactive probe to screen a library of genomic DNA partially digested by San3A (kindly provided by Dr. B. Dujon, Institute Pasteur, Paris, France), and the clone F2B8 was isolated and sequenced. General molec- ular cloning techniques were as described (Maniatis et al., 1989).
DNA Sequence Analysis DNA sequencing was performed with the dideoxy chain termination method (Sanger et al., 1977), using the Sequenase kit (UBS; United States Biochemical Corp., Cleveland, OH). Both strands of the genomic region containing the MHP1 gene were sequenced on average twice. Sequence data were assembled and analyzed with the Genetics Computer Group programs (Devereux et al., 1984).
Computer Analysis of the MHP1 Sequence Secondary structure analysis of the amino acid sequence of MHP1 was performed using the algorithms of Gamier et al. (1978) and Chou and Fasman (1978). The protein databases PIR and Swissprot were searched with the entire Mhplp sequence using FASTA (Pearson and Lipman, 1988). The sequence of Mhplp was compared directly to sequences of known MAPs using FASTA and PILEUP (Devereux et al., 1984).
In Vitro Transcription-Translation Coupled in vitro transcription and translation were performed with a tran- scription-translation kit (Promega Corp., Madison, WI) and [35S]methio- nine, according to the manufacturer's instructions. For each reaction 2 I~g of plasmid DNA were used. N1300 was a subclone in pBS containing the region from bp 656-3,662, and Nl121 was generated by restriction diges- tion. Construct Cl188 was generated by subcloning the PCR-amplified re- gion from bp 3,562-4,368 into pBS.
MT-binding Assay MT-binding assays were performed using porcine brain tubulin, purified by phosphocellulose chromatography. MTs were preassemblod and stabi- lized in the presence of 2 mM GTP and 20 p.m taxol as described (Vallee, 1982). To test the MT-binding activity of the in vitro-translated proteins, the reaction mixture was centrifuged to precipitate nonsoluble material, protease inhibitors were added to the assembly reaction, and 4 Ixl of the in vitro translation reactions was added to assembled MTs in a total volume of 25 p~l. When Mhplp from yeast cells was tested, protease inhibitors were added to the assembly reaction before addition of 15 ~.1 total yeast protein extract in a total of 50 Ixl. Yeast cells (100 Ixg) were lysed in 100 p,! lysis buffer (100 mM Pipes-KOH, pH 6.9, 2 mM EGTA, I mM MgCI2) by vigorous vortexing (30 s) with glass beads and immediately used for the binding assay to minimize the degradation of Mhplp. Assembly reactions were centrifuged, pellets were rinsed with assembly buffer and recentri- fuged, and supernatants and pellets were analyzed by PAGE.
Antibody Production and Purification The COOH-terminal region of MHP1 from bp 3,562--4,368 was amplified by PCR, generating terminal NdeI sites, and cloned into the Ndel site of the His-tag expression vector pET-15b (Studier et al., 1990), generating the plasmid pET-C1188, which was expressed in Escherichia coli. The ap- parent molecular mass of the His-tagged protein expressed by pET-Cl188 was 36-kD on SDS gels. The 36-kD protein, termed Cl188p, was purified on "His-bind" resin (Novagen, Madison, WI), and 100 p.g was injected subcutaneously into rabbits. The collected sera were centrifuged, and the supernatant was immunopurified on membrane-bound antigen as de- scribed (Olmsted, 1986).
The Journal of Cell Biology, Volume 135, 1996 1324
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

Table L Saccharomyces cerevisiae Strains Used in This Study
Name Genotype Sources
Y501 2n a/ct ura3-52 lys2-801 a ade2-101 o trpl -A63 P. Hieter his3-A200 leu2-Al
Y502 2n a/t~ ura3-52 lys2-801 a ade2-101 o trpl-A63 This work his3. A200 leu2- Al mhp l + /mhp l- Al : : TRP1
Y503 2n a/t~ ura3-52 lys2-801~ ade2-101 o trpl-A63 This work his 3- A2 00 leu2- A l mhp l + /mhp l - A l : : TRP1 [pADN-205]
Y5(M n a/a urn3-52 lys2-801 a ade2-101 o trpl-A63 This work his3-A200 leu2-Al mhp I-AI : : TRP1 [pADN-205]
Y505 2n a/tz ura3-52 lys2-801 a ade2-101o trpl-A63 This work his3-A200 leu2-A1 [pMAC-PsO
Y506 2n a/c~ ura3-52 lys2-801 a ade2-101 o trpl-/t63 This work his3-A200 leu2-Al mhpl+/mhpl-Al::TRPl [pMAC-Pst]
Y507 2n a/ct ura3-52 lys2-801 a ade2-101 o trpl-za63 This work his3-A200 leu2-A1 [pMACmyc-Pst]
Y508 2n a/c~ ura3-52 lys2-801 a ade2-101 o trpl-A63 This work his3-A200 leu2-A1 [pMAC-Bam]
Y509 2n a/a ura3-5 2 lys2-801 a ade2-101 o trp l-A63 This work his3-A200 leu2-A1 [Yep-MPH]
Y521 2n a/a urn3-52 lys2-801a ade2-101o trp l- A63 This work his3-A200 leu2-A1 mhp l + /mhp l- A4 : : TRP1
Y531 2n a/t~ ura3- 5 2 lys2-8O l a ade 2-101 o trp l- A63 This work his3-A200 leu2-A1 mhp l + /mhp l-A5 : : HlS3
Y105 2n a/a ura3-52 lys2-801 a ade2-101 o trp l-A63 This work his3-A200 leu2-A1 mhp l + /mhp l-A2 : : URA3
Y106 n ~ ura3-52 lys2-801~ ade2-101o trpl-A63 This work his3-A200 leu2- A1 mhp1- A2 : : URA3
Y561 2n a/a ura3-52 lys2-8Ola ade2-101o trpl-d63 This work his3-A200 leu2-A1 mhp l ÷ /mhp l-A6: :kan r
Y562 2n a/a ura3-52 lys2-801~ ade2-101 o trp1-A63 This work his3-A200 leu2- A1 mhp l + /mhp l- A6 : : kan ~ [pMAC-MHP]
Y563 2n a/cK ura3- 5 2 lys2-801~ ade2-101o trp1- A6 3 This work his3-A200 leu2-A1 mhpl +/mhpl-A6::kan r [pMAC-Pst]
Y564 n a ura3-52 lys2-801~ ade2-101 o trp1-A63 his 3-A200 This work leu2-A1 mhp l-A6: :kan r [pMAC-MI-IP]
Y565 2n a/a ura3-52 lys2-801, ade2-101 o trp1-A63 This work h is3-A200 leu2- A1 mhp l + /mhp l - A6 : : kan r [pADN-205]
Y566 n a u r a 3-52 lys2-801~ ade2-101o trp1-A63 his3-A200 This work leu2-A1 mhpl-A6::kan ~ [pADN-205]
Tubulin-binding Assay Crude bacterial lysates from 200 ml cultures of E. coli expressing Cl188p from plasmid pET-C1188 or the His-tagged NH2-terminal region of MHP1, encoding amino acids 1-440, from plasmid pET-N440, were loaded onto His-bind resin columns. PET-N440 had been generated by cloning the PCR-amplified coding region of MHP1 with terminal Ndel sites into the NdeI site of pET-15. Deletion of the 3' XhoI fragment and religation al- lowed the creation of pET-N440. The Ni-columns were washed with stan- dard His-bind binding buffer, equilibrated with 100 mM Pipes, pH 6.9, and loaded with a crude protein extract from 1 g of frozen S. cerevisiae strain Y501 prepared in 4 ml MT-binding buffer. Columns were washed exten- sively with Pipes and eluted with His-bind elution buffer. Column frac- tions were analyzed by PAGE and Western blotting.
Western Blots Blotting procedures were performed as described (Ortega Perez et al., 1994). The membranes were incubated at room temperature for 2 h with anti-MHP1 in a 1:200 dilution, or anti-205K MAP in a 1:500 dilution. The secondary antibody was alkaline phosphatase-conjugated anti-rabbit IgG (Sigma Chemical Co.), in a 1:5,000 dilution, and the reaction was visual- ized by BCIP and NBT (Sigma Chemical Co.). When a chemilumines- cence detection method was chosen, anti-MHP1 was used in a 1:1,000 dilu- tion, and goat peroxidase-coupled secondary antibody was used in a 1:5,000 dilution. Signal detection was carried out with the ECL kit (New England Nuclear, Boston, MA).
Indirect Immunofluorescence Microscopy Yeast cells were harvested during exponential growth phase and prepared for immunofluorescence staining as described (Kilmartin and Adams, 1984). The anti-ct-tubulin antibody YOLl/34 (Kilmartin et al., 1982) was used at a 1:20 dilution, and a 1:200 dilution was used for the polyclonal anti-MAP antibodies. To visualize MTs and Mhplp, TRITC-labeled sec- ondary antibodies (Socochim, Pully, Switzerland) were used for tubulin, and FITC-labeled anti-rabbit IgG (Socochim) were used for the poly- clonal anti-MHP1 antibodies. The DNA staining dye 4',6-diamidino-2- phenylindole (DAPI) and 0.1% p-phenylenediamine were added to the mounting medium to visualize the nuclear region. An alternative proce- dure was applied to better preserve the cytoplasmic localization of Mhplp; the fixation and digestion of cells was 30 min each, and the time of antibody incubation was reduced to 50-60 rain.
Epitope Tagging of Mhp lp The antigenic myc-tag (Munro and Pelham, 1984) was PCR amplified, generating terminal BamHI sites, and cloned into the BamHI site located between the GAL promoter and the MHP1 sequence on plasmid pMAC- Pst (Table II), generating pMACmyc-Pst. Constructs were sequenced to confirm correct insertion and conservation of the open reading frame (ORF). Y507 cells, transformed with plasmid pMACmyc-Pst (Tables I and II), were grown in selective medium overnight and for another 10 h in selective medium containing 1% galactose or 1% glucose. For immunoflu- orescence microscopy, the tagged epitope was detected with monoclonal
Irminger-Finger et al. Essential Yeast Microtubule-associated Protein 1325
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

Table II. Plasmids Used in This Study
Plasmid Description Source or Reference
pADNS LEU2, 2Ix Colicelli et al., 1989 pADN-205 205K MAP eDNA cloned into pADNS polylinker This study pBS-Bam 3.7-kb MHP1 fragment, containing amino acid codons 219-1,398 and 3'-untranslated region, in pBS This study pET- 15b T7 expression vector containing His-tag Studier et al., 1990 pET-C1188 820-bp MHP1 fragment, containing amino acid codons 1,188-1,398 and 3'-untranslated region, in pET This study pET-N440 1,320-bp MHP1 fragment, containing amino acid colons 1--440, in pET This study pMAC-80 GAlA promoter cloned between SacI and BamI-H site of polylinker of the pRS315 plasmid This study pMAC-Bam 3.7-kb MHP1 fragment, containing amino acid codons 219-1,398 and 3'-untranslated region, in pMAC-80 This study pMAC-MHP 4.2-kb MHP1 fragment, containing amino acid codons 1-1,398 and 3'-untranslated region, in pMAC-80 This study pMAC-PST 2.5-kb MHP1 fragment, containing amino acid codons 561-1,398 and 3'-untranslated region, in pMAC-80 This study pMACmyc-Pst Antigenic myc-tag, cloned into BamHI site ofpMAC-Pst at Y-end of MHP1 sequence This study pRS315 LEU3, CEN6 Sikorski and Hieter, 1989 Yep24 URA3, 21.~ Botstein et al., 1979 Yep-MHP 5.5 kb, containing MHP1 gene in Yep24 This study
anti-myc antibodies (generous gift of Prof. T. Kreis and Dr. L. Huber, University of Geneva, Switzerland) and visualized with a secondary anti- mouse lgG fluorescein conjugate.
Disruption of MHP1
The mhpl-Al allele (see Fig. 6) was generated by excising a l . l-kb EcoRI fragment from the MHP1 coding region, from amino acid residues 818- 1,210 (see Fig. 2 A), and inserting a 0.9-kb EcoRI fragment containing the TRP1 + marker. The MHP1-TRP1 construct was excised from the plasmid with SalI (one genomic SalI site and one site in the plasmid linker) and transformed into the diploid Trp- strain Y501 (Rothstein, 1983). Trio + transformants Y5ff2 were analyzed for correct insertion of the marker gene by PCR and Southern blot analysis. The mhpl-A2::URA3 mutation was generated by inserting the URA3 + marker between the two PstI sites at position 1667 and 1679, respectively (see Fig. 2 A). A 4.5-kb SalI-HindlII fragment was excised and used to generate the heterozygous Ura + cells Y105. Insertion of the marker gene was confn'med on Southern blots. mhpl-A4::TRP1 was generated by cloning a PvuII fragment containing the TRP1 + marker gene between two NcoI sites (positions - 2 and 1833) within the MHP1 gene. A 3-kb NotI-SalI fragment (both sites in the linker sequence of pBS) was excised and used to transform Y501 cells, and Trp + transformants Y521 were selected. The correct insertion of the marker gene was confirmed by PCR. The mhpl-A5::HIS3 mutation was generated by excising the PstI-EcoRI fragment (from position 1667 to 3626) and replacing it with a PstI-EcoRI fragment containing the HIS3 + marker gene. A 3.5-kb SalI-SalI fragment (one SalI site near the 5' end of MHP1 and one SalI site in the linker sequence at the 3' end of the MHP1 gene) was used to transform Y501 cells, and His + transformants Y531 were selected and analyzed for insertion of the marker gane by PCR.
The mhpl-A6::kan" allele was generated by PCR amplifying the kanr gene from the plasmid pFA6-kanMX4 (Wach et al., 1994), using the primers 5' CACTTAAATACGCAAAATATACAAGATAGCCCTACAACT- GCTCCATGCAGCTGAAGCITCGTACGC 3' and 5' TFAT'ITAGT- ACTACTGTTCCGCGCATCCTGGATITFGTCTAGCACGCATAGG- CCACTAGTGGAT 3'. The resulting DNA fragment contained terminal ends homologous to 45 bp flanking the respective translation start and stop sites of MHP1. Transformants were selected on plates containing rich medium supplemented with 100 mg of G418 per liter. Six independent transformants were obtained that contained the mhpl-A6::kan r allele cor- rectly substituted at the MHP1 locus as analyzed by PCR.
Complementation and Overexpression Experiments To complement the mhpl-A6::kan" allele with MHP1, we generated pMAC-MHP. The region including nucleotides 1-670, downstream from the unique SalI site, was amplified by PCR with terminal BamHI sites and cloned into the BamHI site of pMAC-80, a derivative of pRS315 (Sikorski and Hierter, 1989), into which we had inserted the GAL promoter (Table II). The remaining 3.6 kb of the MHP1 coding region from the SalI site at bp 752 to the 3' end (SalI site in the multicloning site of pBS) were in- serted into the SalI site, and correct orientation was determined by restric- tion digestion analyses. Heterozygous mhpl+/mhpl-A6::kan r cells Y561 were transformed with pMAC-MHP or pMAC-Pst (Table II), and Leu +
and G418-resistant cells were spornlated and dissected on glucose- and ga- lactose-containing plates. In 8 out of 12 tetrads, two colonies of normal size and two small colonies could be observed after 4-5 d of incubation at 30°C, with the normal-sized colonies unable to grow on (3418. All small colonies were Leu + and resistant to G418. Colonies growing from individ- ual spores were followed by microscopic inspection for 7 d. Leu + and G418-resistant cells from glucose plates were grown in liquid medium con- taining either glucose or galactose, and aliquots were taken after 4 and 20 h of growth and prepared for immunofluorescence.
Overexpression of the 3' half of MHP1, designated mhpl-A3, was per- formed with the clone pMAC-Pst. The mhpl-A3 allele was generated by cloning the region from bp 1,769-4,339 of the MHP1 gene between the PstI and SalI sites of pMAC-80 (Table II). The correct insertion was veri- fied by restriction analysis. Y501 and Y502 cells were transformed, and Leu + or Leu + Trp + lines were generated and designated Y505 and Y506, respectively, pMAC-Bam was generated by cloning a BamHI-PstI frag- ment (bp 657-1,684) of MHP1 between the BamHI and PstI sites of pMAC-Pst. Y501 cells were transformed with pMAC-Bam and Leu + transformants were designated Y508. For pheuotypic analysis, the cells were grown in selective medium overnight, and 5 to 20 h in selective me- dium and 2% galactose.
Rescue experiments of the mhpl-A1 deletion mutation were attempted with the 4.5-kb cDNA of the Drosophila 205K MAP (lrminger-Finger et al., 1990), which was cloned into the NotI site of the yeast multicopy expres- sion vector pADNS (Colicelli et al., 1989), containing the alcohol dehy- drogenase (ADH) promoter and the selectable marker LEU2 +, generat- ing plasmid pADN-205 (Table II). The heterozygous mhpl+/mhpl-A1 cells Y502 and mhpl+/mhpl-A6 cells Y561 were transformed with the re- sulting plasmid pADN-205. Leu + transformants were selected and used for tetrad analysis. Silencing of the ADH promoter was performed in me- dium containing 2% galactose instead of glucose. Cell growth was quanti- fied by growing cells for 12-20 h in glucose, and then the OD~00 was mea- sured. Cells were diluted to an OD~0 of 0.05 and growth was monitored in glucose and in galactose.
Results
Cloning of MHP1 To clone genes encoding yeast MAPs, a eDNA expression library was screened with a previously produced antibody raised against the Drosophila 205K MAP (Goldstein et al., 1986). The positive cDNA phage 205.41a was used as hy- bridization probe to isolate the genomic clone F2B8 (Fig. 1 A). DNA fragments from different regions within F2B8 were used to determine the size of the MHP1 transcripts on Northern blots (Fig. 1 A, probes 1 and 2). Both probes hybridized to an mRNA of 4,500 nucleotides. DNA se- quence analysis of the transcribed region, including probe 1 and 2, revealed an ORF coding for 1,398 amino acids
The Journal of Cell Biology, Volume 135, 1996 1326
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

A ~;YP6
'P~be'l '1000 bp
MHP1 2 0 5 . 4 1 a
' Probe 2
Nsl:,
B I C . . . . : ' t ' " . . . . . : " " " " : l " : . . . . . . . . . .
HIS EF.hand MT-blndlng NL8
+05 + 0 ~ ~ + 0.1 -0.1 * OJ -0.5
0 200 ~ I~O QO0 1000 1200 1,1~0
Figure 1. MHP1 gene structure and diagram of protein organiza- tion. (A) Organization of genomic clone F2B8 with relative local- ization of the MHP1 coding region between GYP6 and NSP1, as well as the size and relative location of hgtll eDNA clone 205.41a, are shown. (Arrows) Directions of transcription. Probe 1 and 2 were used to probe Northern blots. (B) Schematic diagram of the organization of Mhplp. (Asterisks) Potential phosphoryla- tion sites. HIS, EF-hand, MT-binding, and NLS refer to the posi- tions of motifs discussed in the text. The density of positively and negatively charged amino acids in a sliding window of 20 residues was calculated and plotted against the amino acid sequence.
preceded by a putative CAAT box and a TATA box at po- sitions -52 and -27, respectively (Fig. 2 A). The coding region of MHP1 is identical with ORF YJL042W on chro- mosome X and is separated by 344 noncoding bp from NSP1 (Hurt, 1988) and ~1,300 bp from GYP6 (Strohm et al., 1993).
D N A and Protein Sequence Analysis
No significant homologies with other proteins were found when the EMBL and Swissprot databases (release May 1996) were searched with the MHP1 sequence. From amino acid residues 128-137, nine histidines are encoded that could have a function in metal binding. With sequence analysis programs for the detection of specific motifs, a possible EF hand motif was identified between amino acid residues 615 and 629 (Fig. 2 A). When the MHP1 protein sequence was compared directly with protein sequences of known MAPs using PILEUP, MAP4, MAP2, tau, and Mhplp could be aligned and showed a significant similar- ity in a region close to the COOH terminus, including resi- dues 1,211-1,266 in Mhplp (Fig. 2 B). Within this region, Mhplp has between 23 and 32% identity or between 33 and 38% similarity with the sequences of the other MAPs. The highest similarity score was found with MAP4, and the lowest with tau. Included in this region is (from resi- dues 1,234-1,266 of Mhplp) one repeat of the AP motif common to tau, MAP2, and MAP4 (Lee et al., 1988; Lewis et al., 1988; Aizawa et al., 1990; Chapin and Bulinski, 1991; West et al., 1991). Some 50 residues downstream from the putative MT-binding motif, a nuclear localization sig- nal (NLS) consensus sequence (Roberts, 1989) is present (Fig. 2 A).
MHP1 encodes a hydrophilic protein, with a calculated
isoelectric point (pI) of 6.9, but is composed of alternating basic and acidic regions within which the calculated pI val- ues range from 10-12 and from 4.5-5, respectively (Fig. 1 B). The highest concentration of charged amino acids is observed within acidic and basic regions near the COOH- terminal end of Mhplp. The NH2-terminal and the COOH- terminal regions of Mhplp are very proline rich, typical of extended protein structures, but between residues 1,070 and 1,150 a propensity for the formation of ot helices is predicted (Chou and Fasman, 1978; Garnier et al., 1978).
A total of 52 potential phosphorylation sites for the cAMP-dependent kinase (Edelmann et al., 1987), the cal- cium-phospholipid--dependent kinase (Gonzales et al., 1989), the CaZ+-calmodulin-dependent kinase (Pearson et al., 1985), and the casein kinase II (Diaz-Nido et al., 1988) are situated within the NHz-terminal two-thirds of Mhplp, but only two potential sites are localized within the COOH- terminal domain (residues 1,201-1,398) of the protein (Figs. 1 B and 2 A).
Identtfication of Mhp lp with Anti-MHP1 Antibodies
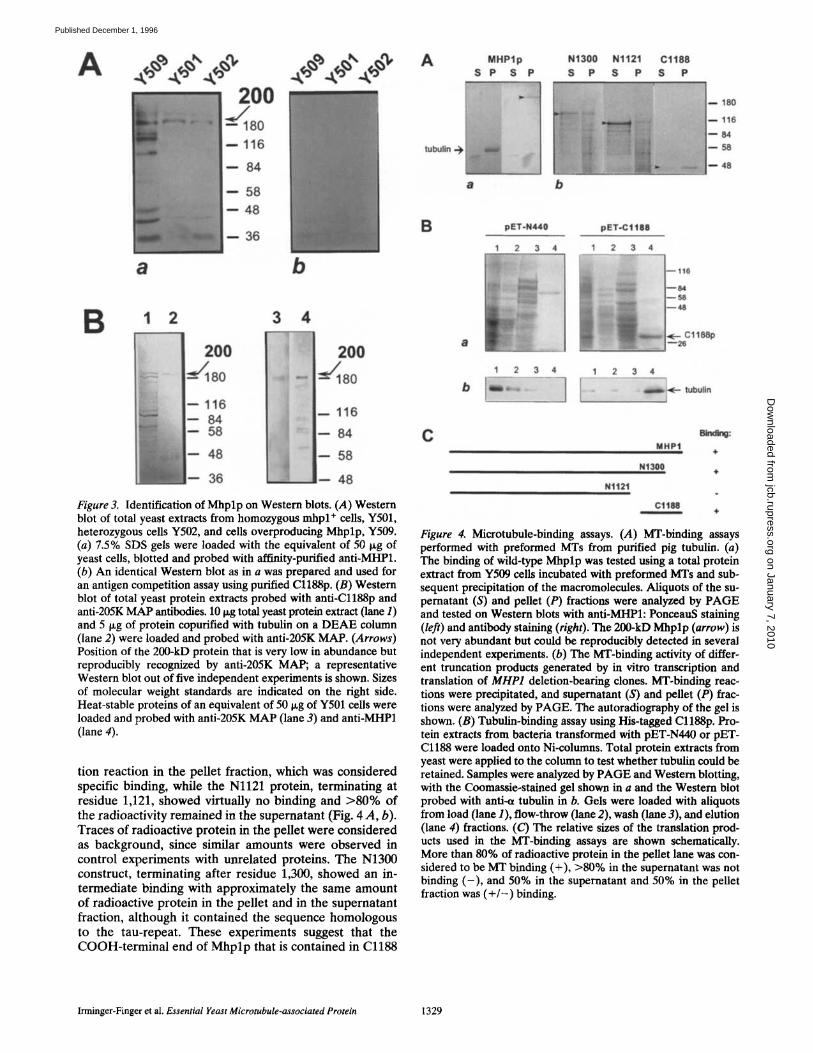
To obtain Mhplp-specific antisera, clone pET-Cl188 en- coding amino acids 1,188-1,398 was expressed in E. coli, and antibodies were generated against the His-tagged 36-kD protein C1188p. The antiserum was tested on West- ern blots and recognized the purified samples of Cl188p and Cl188p in protein lysates of induced bacterial cul- tures. On Western blots of whole cell extracts from S. cerevi- siae, a 200-kD protein as well as smaller proteins of ~<46 leD were consistently recognized by anti-MHP1 (Fig. 3 A; Y501 and Y502) when 20 i~g of total protein extract was loaded.
The amount of anti-MHP1 reactive protein is reduced in mhpl+/mhpl-A1 heterozygous cells Y502 compared with mhpl + cells Y501 (Fig. 3 A). When an equal amount of protein from Y509 cells overproducing Mhplp from a multicopy plasmid was loaded, the 200-kD protein was substantially increased on Western blots, but also a pro- tein of 150 kD could be detected. The full-length MHP1 gene expressed in E. coli also migrates at 200 kD, support- ing the conclusion that full-length Mhplp is identical with the 200-kD band (data not shown). Less intense staining was observed for proteins of 230, 90, 60, and 46 kD. After longer exposure times, the same patterns of proteins were observed in the protein extracts from Y501 and Y502 cells. To test the specificity of the antibody reaction, competi- tion experiments were performed using purified Cl188p. The addition of 1-10 ~g/ml, purified Cl188p was tested, and 5 ixg/ml was found to be sufficient to compete the anti- body recognition of Mhplp on Western blots. When 5 ixg/ml purified Cl188p was added during the antibody incuba- tion, the 200-kD as well as the other proteins observed in Y509 cells could be completely competed with Cl188p (Fig. 3 A, b). These proteins presumably contain the COOH terminus against which anti-MHP1 was raised, and are either isoforms of Mhplp and/or degradation prod- ucts, although the caveat remains that anti-MHP1 might recognize an epitope that is present on other proteins. The observed size of 200 kD for Mhplp is considerably larger than its calculated molecular mass of 154 kD, but it could result from phosphorylation and abnormal migration as a result of the acidic portions of the protein.
Imlinger-Finger et al. Essential Yeast Microtubule-associated Protein 1327
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

A
-3 TCCA~CTCCA~CACOCaAA~Aa~aCCC4~ATCCC~a~C ~. ~ D S K D r O g L L X E n R ~ P C I D
5a cr,mc.a ~:~c,a~x:aa~cc CA~sccrcc ~CC a ~ ^ ~C~CCa ¢,aCC AC,~C.aAA'CC c a 0 V G W L V R P SA S T ~ X S SR P G K S
&0l S K A N S V A p D I Q N D T A R p p V
~0 F • T 8 v M S s S ~ ~ L ~ S ~ D K G R R
238 CACTC c~vr AO~ ' A ~ C ~ A A T~ TAATCAt~ ~ ' I ~ A A ~ A C A TC C |OH S V A A S L L M D N 0 R A N^ G S m S
29~ ~cccAc~cxTTccrceccc~x~v^o~Tc~a~c^~n~cacr~ccrc 1 0 0 V p T N I P P p R G R ~ K S V V g T N L
ase ,~.r a~c¢~'rcaooc't ca crcc c.c, aca'c~ a'~c* cc AC CACC C, CCACC*C Ca'CC ̂c ̂C ~U~ 1 ~ 1 0 5 N V I~ A D S G H n H II H R H H H H m E
4 1 8 CdgI'C~IL"~'~=A CCThAOAA GG~r COG~ I~Z'LTr r aOt O A ~ CA TAGg~AC4~G 1 4 0 D A p J~ p K K V G ? le X S L l e G H R K K
4"/9 (k'~'I'CAtI~kA~GGA~CGAGA J ~ t ~ COASt x ~ o ~ O z ~ o ~ x a x a x z a s • s ~ m ~
s~$ ¢'a;~ac c ~ ~ ~aCCa~aA~Aa¢.~C.A~ C~a~ACCa Ta~c'aacc,~aA~cacc^ l|OV D n 0 * A I R I~ g a T A "~ I $ A g $ e
5911 ~ G TAC~TGCTCCA ~ A T A A C GATACTGTOGTA¢ C GTTAAC AAGA ~ e o e e ~ v s 0 ~ ~ s ~ ~ O ~ V V P L T ~
SSB TCCIkACa&CO~AA TCAGAA GrD~TA " F ~ T C ^ ~ A C T A C C A ~ l ~ l e s x T Ig s Z V y Y g N n P Q S V ¥ H O R
718 ATOC GCACATACCA CTCACCAGAAOA OGGTAAAG ~" GACCCAA CTTC C C CC~CA GACC.AT 240M R T Y N S P E EG K V Dq T S p A D D
a~o I . : N ¥ G O $ a p D e a L . D F L X y y X
a38 '1~ TAAGGA TTACAAATTA GC C GCC'FI~ AAA ~A^O3C, AA ~ATTA A G'I~ k ~ GTC T ~80 g K D y K L A A l' K E O N P I K S S A S
sg~J CCCAC.~ACCAAG~TAGA~TCA'i~ACATAAC~¢A~CCACAAC~"T ~ 0 0 F T T K K N R X k S e S ~ ~ N D K p 0 p
95S OeeAA~'~exerrC.CTeACCaaO~,rln~AC¢,erA*~.~OACCCAr,a:coeC~CATeeC ~a0a K S L A H Q K F D A K O R p Z p p H p
1018 GACGCA CCCAAA ~ ~ A T t " C A A GC AACGCATC GA T^ ~A ~40D h P K t, P S A F R K K H p S N A S I V
1078 GACAC AGIOGA TTCCAAT~ C GTTTC CTCG~C AOC~TAATAA TCAGACACCT ~|0D T V D S N S D V S £ ~ A ~ N N NQ T P
s | o s s H K F G A F L R K V T S ¥ G I~ N I~ N
1198 AAC~GCA CCAA T~-CT~ CTCL ~ A - A iq. ~ ~C AAACGTCAAT~ATCCAGACACCTC ATCGAC~ ~0~N 9 T N A S S L S A N VN N pD T S S T
1258 TCC CTA'I~ T~At 71"I~ T ~ CC C'T'I~ ~TAA C C A ~ ~ 4 ^ a s L W S S S S M E F D p S K I T T V p G
1 3 1 8 C ~ TA TAC~L~PT~CGT~ C~i'X~ T ~ ACCTAT.~AAL~GA T 4 4 0 L g N I R P L K H V S F a T N T y F N D
~37a C C ' e C C a C * A C ~ U ~ a ~ A T O c * ~ r A ~ A A a ~ a * ~ 4eeP P 0 0 i c s K N P R X a EV EV K p N
1431~ GGATCT~ A'PITATAC~TTGAC~CGCAA~AACOC~T~ATGGAAK GAt'T ~8~G ~ VV l H R L ~ • ~ ~ R K K 1 M ~ ~ T
l O e ^ .~OC, OCAa~XXAa CmATrXAA~: C CC.~ ~C.~ ¢ S 0 0 S t , O V V V t~ G T O Q L K L L N p ]g g D
ISSq C~CCOU~CC~G*~T~A~aO~O*~CCCC~e4U~a~aC~ 520D A N A K .~ R E E M A P Q K 0 N E V S A
S4~H D E E D N N S Q R R N Z V M A X A g A
167B GCTGC~%G,A~GO2 AGA ~ ~CM~C A C C'TAAC C , ^ A ~ ~ 5{0A A E A R A ~ Z A ~ N E L K R I VT NN
17~ aa.~OAAC-A.~ ~ C'A'OrCA or AA 0.~ c~',~'ca~-'cA,~r r ~ACCaVr C C~T^A~CCXaTC-A'~r 580E • g V T V S K T A S H L T I D K p M I
17~ T C ~ ¢ a ~ T ~ C C ~ C * ~ T ~ I C C ^ G T ~ T ~ C C ~ C G U T 600S R R O A S T ~ S L A g MV S ~ D T ~ G
1~58 XC~Aa CC, CA~A~a~.~TC, A*~A C,A~a ~'~r r ~CCCCa C C ' ~ O a ?XCCmC ̂ C~a C S;10T N a n D g C ?, I L e p @ S L K I p H D
1918 A T~ATA CC CGTTGTTG~ AC TTGC (~ A ~ ~F~C CC A TACCA GCCACA TTGAA G ~0 I V Y T R C C n L R E ~ L p I p A T L K
2038 'I'¢~i'A ~ q ' F r OG'~= G:I'I~:: AC~CGA'A'?I'~T TTGAGCA'~'A~C'a~C CC ~ a TGTC~Z~I " ~eos ~v ~ z w ~ v ~ D v u s z x ~v ~c ~
? o o s ~ ~ a v o ~ v ~ ~ ~ x z ~ s s L V
2~5~ ~ac AaACA~C a,v~Awaaacaccccc,t.l~eAmc, Aa ~ a
~lev p s i t T N V (} K p S K S S L K ~ K I
2~8 ~a~O~TUC~TOC~CTTCC4U~aTa~a~CC~C*TU~*CT~a~C~OeT ? | O L R M Q C N L ~ N R S D M N W D L L T A
|00g I A L M G G h g I I V I ~ G A K 14 N ~
ll0A 0 F l N F I L VA C I A T E R L ~ LA
2518 TA TAA'/~GATTATC~AAGAOCCAATGI~ATGA TI~G~CC AAGTGOA TCGTTC AATCA/U%A ~ o ~ . o ~ s ~ s ~ c ~ a X W N v ~ S X
2 5 7 8 ~i~ACAGG~A~AC GT~AACGA T'i~'i~ k^ACTTTCC 'I~'~I~ GAT s ~ o v m ~ ~ D v a ~ . m ~ x ~ S S ~ m V
~eOA V L G K X Q K A N E K NV r K ~ L S M
2758 CT~ATAAGCgI~F/~ATGTTA TIEOOAAAA'~AAAA~TATATC G~ACAATCCT ~ o L z $ v L c y s z N L X r L D 1 S S N ~
2 , : , a a c e * ~ c a c r a A , z . r a~an- r TTT~CCTWlTr r ' r w~Te ' r r ~0A ~ F P H C V pT L I D F L pV F VN L
28"/8 GI~ C G~Y~gC & TA'~'T~qCTACAA TAATTTAAGTTC~ AC GA gI~TAGTGA T ~ 9~0V R L " I D Y N N L S S T S VV M L A g
29J8 ATATTA CC ~ A J~I4~TTA T'pi'ppLT.A ~AOGAA C~T.~.^A~ s s o i L v v c s e L , y e s ~ L ~ T s ~ D L
I 0 0 0 A $ S K A I. l a A V R K $ S S L M T L O
ao~a ~ c r A ~ r A T A ~ o A C . a A c a ~ r A a ~ A a A a ~ r m e a ~ . r ^ T g e ~ c ~ r a a a A l o ~ o v D y v y M e S N ~ ~ g ~ I S L y A L ~
3116 A A T A T T C ~ G G G G A ~ TAGI~A CC.~TA~AGATA TCAAAGA TAOC~ 10~0N I Q G E L K R V N g D D K D X K D S Q
3175 a ~ . r c a.~oec.r a c x c ~ e e a ~ c r X ~ o ~ e n ' r r r ̂ T r a . ~ ' r c ,O ,~a t a e a ~ T ~ x o ~ e r s a L o D O L S ~ L ~ T Z X ^ ~ ~: S S
~080H y N K M V S N y M A K I A LA m l K I
IX00S ~ v v , o L y ~ L ~ L N O ~ L ~ ~ ~ O
a~5~ ~ UC~C~CT~a~rAO*TT*~T~a~aC~ ~ T c r o*c c~ IX~OK ~ A L I R L C Y X g A S L g R • C D L
3 4 1 8 C'~C~AAACAAAGACATAACAACA~ TCACCA ~ ~ XX&OL K Q R H N N T L K ~ P S A V g K g R K
x x s o ~ O N Q A 0 P N S g S e Q R M F I S S S
3538 A T A ~ A C ~ A ~ T A ~ ~ A ~ CA ~ A ~ T * ~ 1180Z L Q N S D S ~ A L M p ~ G S* I V E K
~5~a ~CA~ ~C CC~'mC A~Aa C,a ~CCCr COAa~n~C XC.~OAaO~'CaaC C.AC~C~r A~am 1 2 0 0 S g P D A g D A V g F R E G n O g g V N
a658 C*Cam~AC~r~C~TT~C~a*Taa~q~TaTe~UUU~a
3718 ~C~rA~CATT~U~a~* ~ ~ C C *
3"/'/8 GTGGAC AAA~qAA Tt'TTAAA ~CAATt~CTTAGATAGTGAC C AC~ATAA~GGAA I~0V D K R Y L" R A A Q S L D 8 D ~ I K E
~8~a T~CrOqTOAa~T~aC~C~T~crATcCrTCUC~.~TOA~C~C~TA~** 23e01" ~ L K N D V S T I L G V l D g L H S Q
3698 ~kc c&c~ ATCA T A ' ~ A ( I ~ ~ G A ~ C C t I ~ A GAA
395a CC*~O~Ca*C*O~:CTCeC~*TCOaT~C~T~C~CeCC~T¢C z~oe ~ ~ ~ ~ ~ s s ~ s ~ D s S A . * S ~ S
4018 ACTG ATCCG~TATCC.~CA ~ CA~ TACTTCTA GAACGAACG ATAATC ATC ATA'~CT I~40T D P I S T O S N T S R T N D N H n I P
4078 CCGA CCGATOCGt~I~ T'I~'~A~T'I~A~CAATC~Gdt t ~ T~ CA'I~C X3eOp T D a p G P D K F M N N A E E N a I D
4138 C~TGCTTAT~ATGATGTGCTAGACAAAAT~CAGGAT~CGCGGAACAGTAGTACgAAATAA ~ a s o ~ A v ~ D v ~ ~ • I o v ^ ~ N s s T x •
4198 TAAGGCA T t I~*TCTTAAT'~AAq~I~ T T ~ A T T ~ GAAA~TAT/U~T4~
4 2 5 8 GGJ~ ~:~'C CTCA C,^C A~ACG TA ~ A ' C G ^ ~A ~A^ ~ x ~ a ~
B
mn WL~.~IB a S wlE_VlMMt a v p a ~ "~10 r @ .le
Figure 2. DNA sequence and predicted protein se- quence of MHP1. (A) DNA sequence of the MHP1 gene and predicted amino acid se- quence are presented. Bold letters indicate the TATA and CAAT sequences, a polyadenylation signal, the first methionine initiation codon, and potential initia- tion codons of the mhpl-A2:: URA3 mutation (see Fig. 6). Potentially phosphorylated amino acids are indicated by an asterisk below the residue. The putative EF hand motif (beginning at residue 615), the region homologous to MAP2, MAP4, and tau (from residues 1,211-1,266), and the potential NLS (residues 1,314-1,317) are underlined. Sequence data are available from EMBL/GenBank/ DDBJ under accession num- ber X84256. (B) Protein se- quence alignment of AP se- quences of different MAPs. Protein sequences of Mhplp (1,211-1,266), human (233- 288) and mouse (222-277) tau, mouse MAP2 (1,709- 1,764), human (1,007-1,062) and mouse (980-1,035) MAP4, and bovine MAP U (944-977) are aligned. Amino acids, identical or similar (E/D, K/R, I/L/V) in at least three proteins, are boxed. The alignment corre- sponding to the AP repeat is bordered with asterisks.
A 200-kD protein plus a 180-kD protein were also reac- tive with the anti-205K M A P antiserum originally used for the screening of the kgt l l expression library (Fig. 3 B, lanes I and 2), but only the 200-kD protein copurified with yeast tubulin on a D E A E column (Bellocq et al., 1992). When heat-stable yeast proteins were tested with anti- MHP1 and anti-205K MAP, the 200-kD protein was also detected by both antisera (Fig. 3 B, lanes 3 and 4).
Mhplp Has MT-binding Activity In Vitro
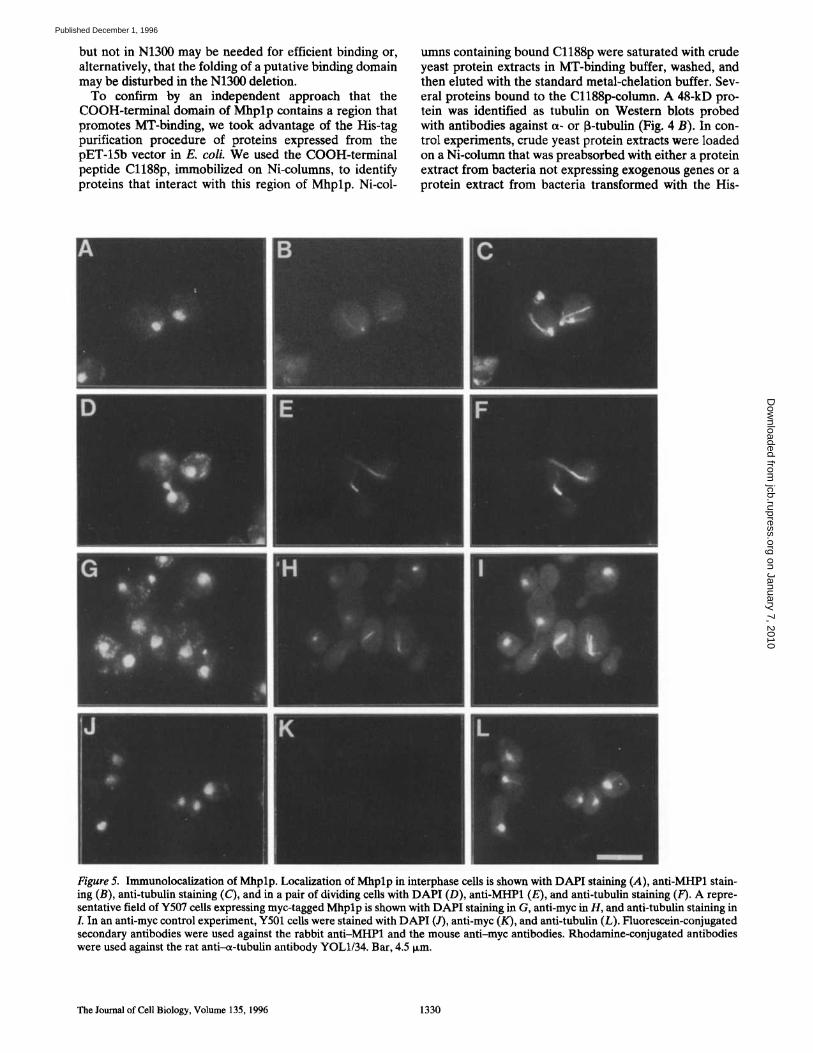
To determine whether Mhplp binds to MTs in vitro, MT-
binding and cosedimentation experiments were carried out. Total yeast proteins were incubated with preformed MTs and centrifuged, and supernatant and pellet fractions were analyzed on Western blots probed with anti-MHP1 (Fig. 4 A, a). The anti-MHPl-reactive 200-kD protein was coprecipitated with MTs and could be identified on West- ern blots• To localize the MT-binding activity within the MHP1 sequence, deletion-bearing clones were in vitro transcribed and translated, and aliquots of the in vitro translation reactions were tested in MT-binding assays. The COOH-terminal fragment Cl188 coprecipitated with preformed MTs with >80% of the counts of the transla-
The Journal of Cell Biology, Volume 135, 1996 1328
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996
A 200
--- 116
" - 84
• - 58
- - 48
- - 3 6
8 b
i 1 2 3 4 7 " . . . . . . . . . . . . . .
- . , , ,
, .
. . .
I 200 80 / 1 8 0
,6 i B4 116
58 84
48 E 58 36 _~ 48
Figure 3. Identification of Mhplp on Western blots. (A) Western blot of total yeast extracts from homozygous mhpl ÷ cells, Y501, heterozygous cells Y502, and cells overproducing Mhplp, Y509. (a) 7.5% SDS gels were loaded with the equivalent of 50 lxg of yeast cells, blotted and probed with affinity-purified anti-MHP1. (b) An identical Western blot as in a was prepared and used for an antigen competition assay using purified Cl188p. (B) Western blot of total yeast protein extracts probed with anti-Cl188p and anti-205K MAP antibodies, i0 ~g total yeast protein extract (lane 1) and 5 Ixg of protein eopurified with tubulin on a DEAE column (lane 2) were loaded and probed with anti-205K MAP. (Arrows) Position of the 200-kD protein that is very low in abundance but reproducibly recognized by anti-205K MAP; a representative Western blot out of five independent experiments is shown. Sizes of molecular weight standards are indicated on the fight side. Heat-stable proteins of an equivalent of 50 ~g of Y501 cells were loaded and probed with anti-205K MAP (lane 3) and anti-MHP1 (lane 4).
tion reaction in the pellet fraction, which was considered specific binding, while the Nl121 protein, terminating at residue 1,121, showed virtually no binding and >80% of the radioactivity remained in the supernatant (Fig. 4 A, b). Traces of radioactive protein in the pellet were considered as background, since similar amounts were observed in control experiments with unrelated proteins. The N1300 construct, terminating after residue 1,300, showed an in- termediate binding with approximately the same amount of radioactive protein in the pellet and in the supernatant fraction, although it contained the sequence homologous to the tau-repeat. These experiments suggest that the COOH-termina l end of M h p l p that is contained in Cl188
A MHPlp S P S P
N1300 N1121 8 P 8 P
Cl188 8 P
180
116 84 58
48
B 1
pET-N440
2 3 4
,,,,,
:..::.::.i:.:,.".:":."...
,,',, • ,,, ,,,
2 3 4
pET-C1188
2 3 4
:1188p
1 2 3 4
D tubulln
C
Nt121
MHP1
Nt3ee
m
Ctltl8
Figure 4. Microtubule-binding assays. (A) MT-binding assays performed with preformed MTs from purified pig tubulin. (a) The binding of wild-type Mhplp was tested using a total protein extract from Y509 cells incubated with preformed MTs and sub- sequent precipitation of the macromolecules. Aliquots of the su- pematant (S) and pellet (P) fractions were analyzed by PAGE and tested on Western blots with anti-MHPl: PonceauS staining (left) and antibody staining (right). The 200-kD Mhplp (arrow) is not very abundant but could be reproducibly detected in several independent experiments. (b) The MT-binding activity of differ- ent truncation products generated by in vitro transcription and translation of MHP1 deletion-bearing clones. MT-binding reac- tions were precipitated, and supernatant (S) and pellet (P) frac- tions were analyzed by PAGE. The autoradiography of the gel is shown. (B) Tubulin-binding assay using His-tagged Cl188p. Pro- tein extracts from bacteria transformed with pET-N440 or pET- Cl188 were loaded onto Ni-columns. Total protein extracts from yeast were applied to the column to test whether tubulin could be retained. Samples were analyzed by PAGE and Western blotting, with the Coomassie-stained gel shown in a and the Western blot probed with anti~ tubulin in b. Gels were loaded with aliquots from load (lane 1), flow-throw 0ane 2), wash (lane 3), and elution (lane 4) fractions. (C) The relative sizes of the translation prod- ucts used in the MT-binding assays are shown schematically. More than 80% of radioactive protein in the pellet lane was con- sidered to be MT binding (+), >80% in the supernatant was not binding ( - ) , and 50% in the supernatant and 50% in the pellet fraction was ( + / - ) binding.
Irminger-Finger et al. Essential Yeast Microtubule-associated Protein 1329
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996
but not in N1300 may be needed for efficient binding or, alternatively, that the folding of a putative binding domain may be disturbed in the N1300 deletion.
To confirm by an independent approach that the COOH-terminal domain of Mhplp contains a region that promotes MT-b'mding, we took advantage of the His-tag purification procedure of proteins expressed from the pET-15b vector in E. coli. We used the COOH-terminal peptide Cl188p, immobilized on Ni-columns, to identify proteins that interact with this region of Mhplp. Ni-col-
umns containing bound Cl188p were saturated with crude yeast protein extracts in MT-binding buffer, washed, and then eluted with the standard metal-chelation buffer. Sev- eral proteins bound to the Cl188p-eolumn. A 48-kD pro- tein was identified as tubulin on Western blots probed with antibodies against a- or g-tubulin (Fig. 4 B). in con- trol experiments, crude yeast protein extracts were loaded on a Ni-column that was preabsorbed with either a protein extract from bacteria not expressing exogenous genes or a protein extract from bacteria transformed with the His-
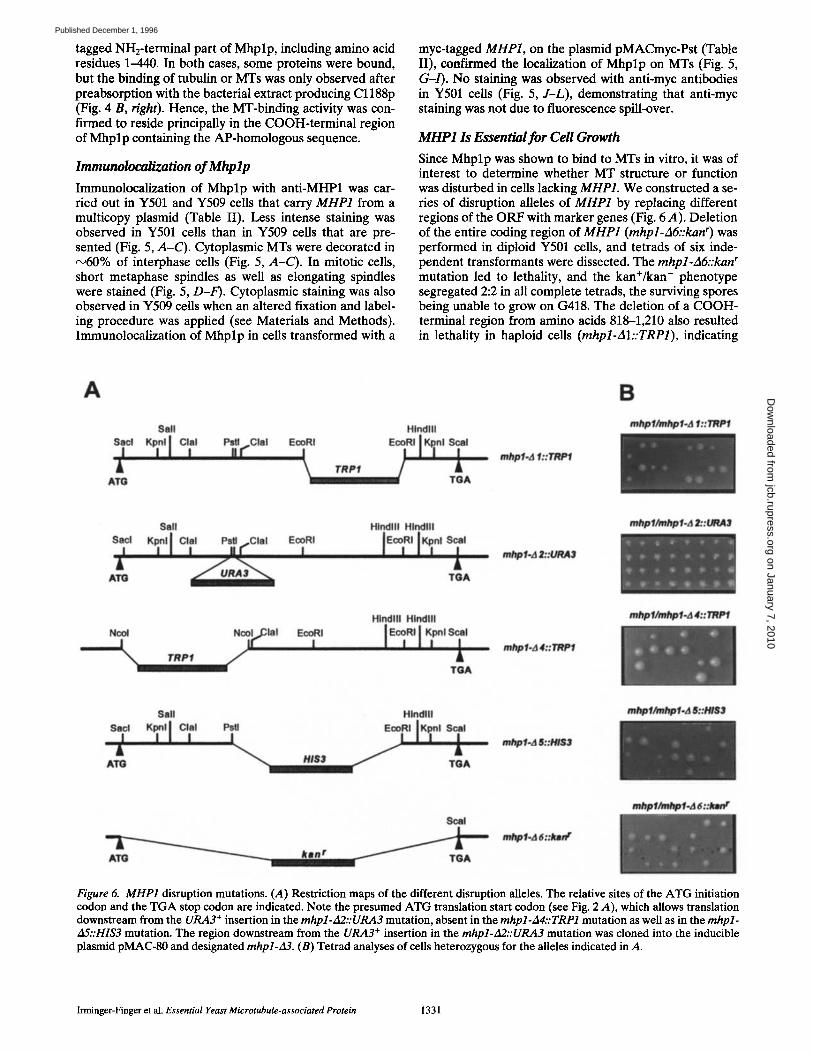
Figure 5. Immunolocalization of Mhplp. Localization of Mhplp in interphase cells is shown with DAPI staining (A), anti-MHP1 stain- ing (B), anti-tubulin staining (C), and in a pair of dividing cells with DAPI (D), anti-MHP1 (E), and anti-tubulin staining (F). A repre- sentative field of Y507 cells expressing myc-tagged Mhplp is shown with DAPI staining in G, anti-myc in H, and anti-tubulin staining in L In an anti-myc control experiment, Y501 cells were stained with DAPI (J), anti-myc (K), and anti-tubulin (L). Fluorescein-conjugated secondary antibodies were used against the rabbit anti-MHP1 and the mouse anti-myc antibodies. Rhodamine-conjugated antibodies were used against the rat anti--a-tubulin antibody YOL1/34. Bar, 4.5 i~m.
The Journal of Cell Biology, Volume 135, 1996 1330
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996
tagged NH2-terminal part of Mhplp, including amino acid residues 1--440. In both cases, some proteins were bound, but the binding of tubulin or MTs was only observed after preabsorption with the bacterial extract producing Cl188p (Fig. 4 B, right). Hence, the MT-binding activity was con- firmed to reside principally in the COOH-terminal region of Mhplp containing the AP-homologous sequence.
Immunolocalization of Mhp lp
Immunolocalization of Mhplp with anti-MHP1 was car- ried out in Y501 and Y509 cells that carry MHP1 from a multicopy plasmid (Table II). Less intense staining was observed in Y501 cells than in Y509 cells that are pre- sented (Fig. 5, A-C). Cytoplasmic MTs were decorated in ~60% of interphase cells (Fig. 5, A-C). In mitotic cells, short metaphase spindles as well as elongating spindles were stained (Fig. 5, D-F). Cytoplasmic staining was also observed in Y509 cells when an altered fixation and label- ing procedure was applied (see Materials and Methods). Immunolocalization of Mhplp in cells transformed with a
myc-tagged MHP1, on the plasmid pMACmyc-Pst (Table II), confirmed the localization of Mhplp on MTs (Fig. 5, G-/). No staining was observed with anti-myc antibodies in Y501 cells (Fig. 5, J-L), demonstrating that anti-myc staining was not due to fluorescence spill-over.
MHPI Is Essential for Cell Growth
Since Mhplp was shown to bind to MTs in vitro, it was of interest to determine whether MT structure or function was disturbed in cells lacking MHP1. We constructed a se- ries of disruption alleles of MHP1 by replacing different regions of the ORF with marker genes (Fig. 6 A). Deletion of the entire coding region of MHP1 (mhpl-A6::kan r) was performed in diploid Y501 cells, and tetrads of six inde- pendent transformants were dissected. The mhpl-A6::kan r mutation led to lethality, and the kan+/kan - phenotype segregated 2:2 in all complete tetrads, the surviving spores being unable to grow on G418. The deletion of a COOH- terminal region from amino acids 818-1,210 also resulted in lethality in haploid cells (mhpl-AI::TRP1), indicating
A
Sail Hlndlll " ' , A' . . . / ,
A'I~ ~ TGA
ml~l.A l:drwPl
B mlqJ1/a~pl.d I:: TRPf
Si l l Hlndlll Hlndlll
'r' l °'r' +.' ATG TGA
mhpl-d I:.'URAI
I mlpl /n~p l .a l . . :Ult~
Hlndlll Hlndlll
+ ~ TRP, ,,Clal I"'l+'',l TGA
m~pf.d 4:: lRPf
mlp f / m l ~ . a 4 : :111Pf
Sail Hindlll mlMl/mlMl-d Ik:lfl$3
ATG H/413 TGA
mhpl.a l l : : l f l~
aW~/,w~t.A6:..Imr Soul
mhpl,,d 6::lmn r
Figure 6. MHP1 disruption mutations. (+4) Restriction maps of the different disruption alleles. The relative sites of the ATG initiation codon and the TGA stop codon are indicated. Note the presumed ATG translation start codon (see Fig. 2 A), which allows translation downstream from the URA3 + insertion in the mhpl-A2::URA3 mutation, absent in the mhpl-A4::TRP1 mutation as well as in the mhpl- A5::HIS3 mutation. The region downstream from the URA3 + insertion in the mhpl-A2::URA3 mutation was cloned into the inducible plasmid pMAC-80 and designated mhpl-A3. (B) Tetrad analyses of cells heterozygous for the alleles indicated in A.
Irminger-Finger et al. Essential Yeast Microtubule-associated Protein 1331
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

that the NH2-terminal 818 amino acids encoded on the mhpl-AI::TRP1 allele are not sufficient for viability.
Surprisingly, the interruption of the ORF after amino acid 558 by the insertion of the URA3 ÷ marker gene led to viable haploid cells (Fig. 6). Tetrad analysis of the het- erozygous diploid strain mhpl+/mhpl-A2::URA3, Y105, resulted in four viable spores, the Ura+/Ura - phenotype segregating 2:2 and the mhpl-fl2::URA3 haploid cells Y106 growing only slightly slower than the wild-type cells. When we analyzed the MHP1 mRNA in wild-type and mhpl-Ag::URA3 disrupted cells, we found that the mutant cells expressed two transcripts (Fig. 7 A), a 1.8-kb mRNA covering the MHP1 5' sequences and a 2.7-kb mRNA cov- ering MHP1 3' sequences, while wild-type cells expressed a single 4.5-kb mRNA. This 4.5-kb transcript is not ob- served in the disrupted haploid mhpl-A2::URA3 cells. A weak hybridization of a 2.8-kb RNA is visible in Y501 cells that might be a second MHP1 transcript or a nonspecific cross-hybridization. These results suggest that MHP1 func- tion can be obtained when NH2-terminal and COOH-ter- minal sequences of MHP1 are expressed from separate transcripts. Analysis of the proteins expressed in wild-type and mhpl-A2::URA3 cells on Western blots (Fig. 7 B) shows that in wild-type cells, a 200-kD protein and bands of 150, 90, and 46 kD react with anti-MHP1, while in mhpl-A2::URA3 haploid cells, no proteins >90 kD can be detected.
To further test the hypothesis that sequences down- stream of the URA3 + insertion were expressed spontane- ously in the mhpl-A2::URA3 allele, we generated deletion alleles mhpl-A4::TRP1 (deletion of the first 613 amino ac- ids) and mhpl-A5::HIS3 (deletion of the region encoding amino acids 557-1,210) that do not contain the two pre- sumed ATG translation initiation codons at positions 1792 and 1839, respectively. Both alleles proved to be lethal.
The progeny of mhpl-AI:: TRP1, mhpl-A4::TRP1, mhpl- A5::HIS3, or mhpl-A6::kan r cells could germinate and go through a few cell divisions, but they died at stages of 8 to 32 cells. The time needed for germination was variable for different spores and explains the size differences of the colonies (Fig. 6 B). The heterozygous cells showed a re- duced growth rate in rich medium. Analysis of asynchro- nously growing Y502 and Y561 cells by DAPI and anti- tubulin staining showed 12% of the cells with short or long spindles and divided nuclei, compared with 2-3% rou- tinely observed for wild-type cells under the same condi- tions, indicating that they are arrested or slowed during mitosis (data not shown).
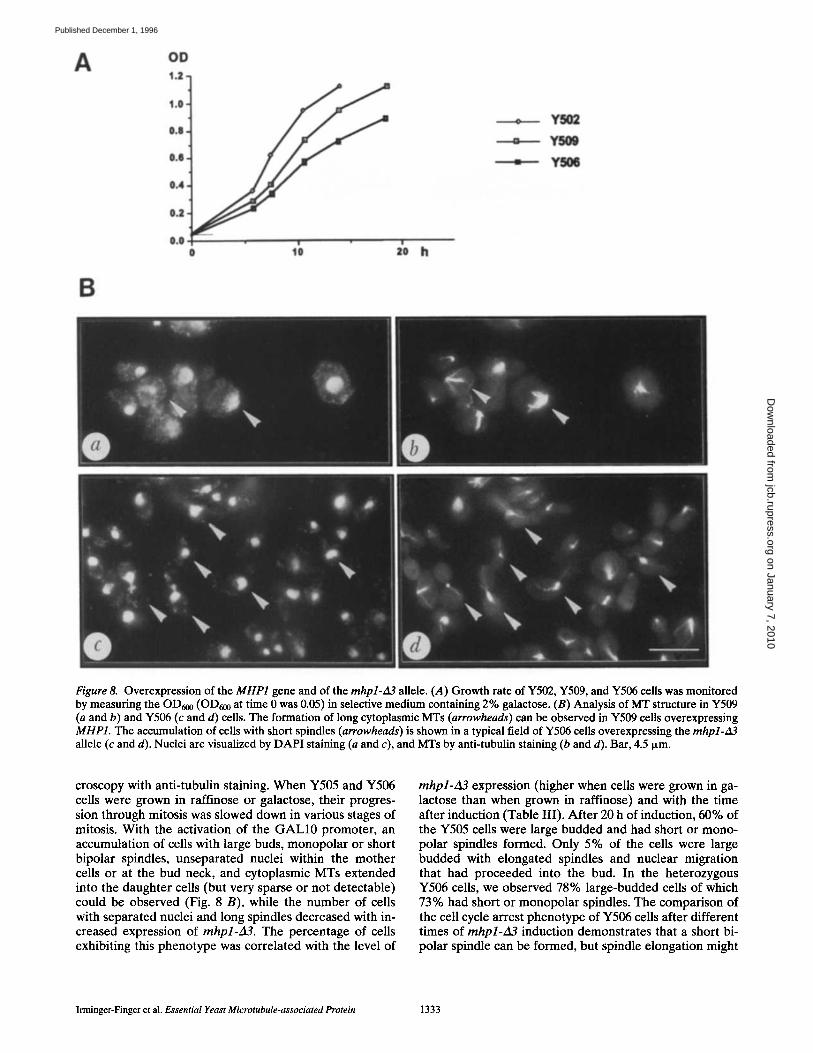
Altered MT Phenotypes Induced by MHP1 Overexpression Mutations
Since the heterozygous deletion mutations of MHP1 showed a gene dosage effect, we investigated whether overex- pressed MHP1, or deletion-bearing alleles of MHP1, would lead to a dominant phenotype. Y509 cells that express MHP1 constitutively from a multicopy plasmid (pYEP- MHP) exhibit a reduced growth rate (Fig. 8 A). The analy- sis of MT structures in asynchronously growing Y509 cells showed that the amount of cells with elongated spindles and divided nuclei was <1%, while the parental strain Y501 grown under similar conditions contained 2-3% of
180
116
84
M
48
3e
Figure 7. Analysis of the mhpl-A2 mutation. (A) Northern blots were probed with a 700-bp probe from the 5' end and a 600-bp probe from the 3' end of MHP1 (see Fig. 1 A). In mhpl-A2:: URA3 haploid cells Y106, the 5'-end probe and the 3'-end probe detected different mRNAs (arrows). In wild-type cells Y501, a 4.5-kb RNA was detected with both probes. (B) Western blot of total protein from mhpl-~2::URA3 haploid cells (Y106) and dip- loid Y509 cells probed with anti-MHP1. In mhpl-A2::URA3 hap- loid cells, anti-MHP1 reacts with a 90-kD protein compared with a 200-kD protein and smaller proteins in mhpl ÷ cells, Y509.
cells with short or long spindles and divided nuclei. In ad- dition, Y509 cells showed an increased cell volume (150- 200% of diploid wild-type cells), many abnormally long MT filaments emanating from one SPB, increased nuclear . DNA staining, and small patches of DAPI-stained mate- rial observable in the cytoplasm (Fig. 8 B, a). In 6% of the cells, bud formation occurred but no spindle was formed. The increase in length and number of MTs in Y509 cells, as a consequence of increased expression of MHP1, is compatible with the role of Mhplp as a positive regulator of MT stability.
A different phenotype could be induced by the overex- pression of the COOH-terminal portion of Mhplp. The region encoded on pMAC-Pst (Table II), designated mhpl-A3, was transformed into homozygous mhpl ÷ cells and into mhpl+/rnhpl-A1 heterozygous cells, generating Y505 and Y506 cells, respectively. Cells were grown in me- dium containing either glucose, galactose, or raffinose (a sugar derepressing the GAL promoter and leading to less expression than galactose). The induction of mhpl-A3 was monitored on Western blots, and the expression of a 90-kD protein, as had been identified in mhpl-A2 cells under conditions of unstimulated expression (see Fig. 7 B), could be observed in Y505 and Y506 when cells were grown in galactose or raffinose (data not shown).
The expression of mhpl-A3 provoked a negative effect on growth rate. While control cells Y502 reached satura- tion after ~24 h with a doubling time of 3 h in media con- taining galactose, for transformed Y505 and Y506 cells, longer times were required to reach saturation when grown in galactose-containing media, with generation times of 4.5 and 6 h, respectively. When media containing increasing amounts of galactose and decreasing amounts of glucose, or the partial inducer raffinose, were used, the growth rates of Y505 and Y506 cells were correlated with the galactose or raffinose content in the media.
To determine the consequence of the expression of mhpl-~3 on MT structure and function, we analyzed asyn- chronously growing cultures by immunofluorescence mi-
The Journal of Cell Biology, Volume 135, 1996 1332
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996
|
zo h
OD 1.2
1.0
0.8
0.0
0.,4
0.Z
0.0 0 t0
T
J
- Y S O 0
= YSM
B
A
Figure 8. Overexpression of the MHP1 gene and of the mhpl-zl3 allele. (.4) Growth rate of Y502, Y509, and Y506 cells was monitored by measuring the OD600 (OD600 at time 0 was 0.05) in selective medium containing 2% galactose. (B) Analysis of MT structure in Y509 (a and b) and Y506 (c and d) cells. The formation of long cytoplasmic MTs (arrowheads) can be observed in Y509 cells overexpressing MHP1. The accumulation of cells with short spindles (arrowheads) is shown in a typical field of Y506 cells overexpressing the mhpl-~13 allele (c and d). Nuclei are visualized by DAPI staining (a and c), and MTs by anti-tubulin staining (b and d). Bar, 4.5 Ixm.
croscopy with anti-tubulin staining. When Y505 and Y506 cells were grown in raffinose or galactose, their progres- sion through mitosis was slowed down in various stages of mitosis. With the activation of the G A L l 0 promoter, an accumulation of cells with large buds, monopolar or short bipolar spindles, unseparated nuclei within the mother cells or at the bud neck, and cytoplasmic MTs extended into the daughter cells (but very sparse or not detectable) could be observed (Fig. 8 B), while the number of cells with separated nuclei and long spindles decreased with in- creased expression of mhpl-d3. The percentage of cells exhibiting this phenotype was correlated with the level of
mhpl-Zl3 expression (higher when cells were grown in ga- lactose than when grown in raffinose) and with the time after induction (Table III). After 20 h of induction, 60% of the Y505 cells were large budded and had short or mono- polar spindles formed. Only 5% of the cells were large budded with elongated spindles and nuclear migration that had proceeded into the bud. In the heterozygous Y506 cells, we observed 78% large-budded cells of which 73% had short or monopolar spindles. The comparison of the cell cycle arrest phenotype of Y506 cells after different times of rnhpl-zl3 induction demonstrates that a short bi- polar spindle can be formed, but spindle elongation might
Irminger-Finger et aL Essential Yeast Microtubule-associated Protein 1333
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

be hampered. In control cells transformed with pMAC- MHP overexpressing the entire MHP1 gene or trans- formed with pMAC-80 (Tables I and II), no such pheno- types were observed. The NH2-terminal 561 amino acid residues of MHP1 therefore appear to be required for the function of Mhplp during progression through anaphase.
MHP1 Null Allele Complemented with MHP1 and with mhp l-A3
To further characterize the null phenotype of the MHP1 deletion alleles, we have performed rescue experiments with MHP1 on plasmids under the control of inducible promoters, pMAC-MHP contains full-length MHP1 under the GALl0 promoter, while pMAC-Pst contains the 3' re- gion of the gene designated mhpl-A3. With both plasmids, the lethal phenotype of the haploid mhpl-A6::kaff cells could be partially rescued, giving rise to small colonies (Fig. 9 A). The fact that mhpl-A3 could also partially com- pensate for the null mutation of MHP1 is consistent with the finding that the insertion mutation mhpl-A2 is viable, and confirms that the expression of the region encoding amino acids 560-1,398 is sufficient for vegetative growth. With both plasmids, rescue could only be observed when dissections were carried out on plates containing glucose, and tetrads dissected on plates containing galactose gave rise to only two colonies of normal size. Microscopic in- spection of the spores on galactose-containing plates, at in- tervals of 1 d, showed that all cells sporulated and divided initially with the same doubling times until they reached the ,'-q6-32 cell stage. Two colonies slowed down in their doubling time and finally stopped growing. We presume that the overproduction of Mhplp under the GALl0 pro- moter was toxic to the cells, and that leaky expression of MHP1 from the GALl0 promoter allowed reduced tran- scription and translation of MHP1 and mhpl-A3 in the small-sized colonies growing on glucose plates.
MHPI Expression Is Required for Sufficient Formation of Mrs To examine the MT phenotype in the MHP1 null allele, we have generated haploid mhpl-A6 cells Y564, comple- mented with inducible MHP] on the plasmid pMAC- MHP. When Y564 cells were grown under noninduced conditions, Mhplp was below detectable levels when tested on Western blots or by immunofluorescence analy-
Table IlL Cell Cycle Arrest of mhpl-A3 Mutant
sis. However, since it became apparent that leaky expres- sion of MHP1 from the GALl0 promoter was sufficient for cell growth (Fig. 9 A), we analyzed the phenotype of Y564 cells after growth in glucose and galactose. Immu- nostaining with anti-tubulin antibodies and DAPI re- vealed that the rescued haploid mhpl-A6::kan r cells con- tained few MTs (Fig. 9 B). At least 15% of the cells formed small or large buds, but only weak staining of mi- totic spindles could be observed. However, the diffuse staining of tubulin in the cytoplasm was increased. When cells were grown for several hours in galactose, which led to a reduced growth rate, long MT fibers were formed in 5% of the cells, in N10% of the cells weak staining was ob- served, and in the majority of cells neither MT nor DAPI staining was detectable. The parental diploid cells Y562 prepared under identical conditions showed intense MT staining in interphase and mitotic cells (Fig. 9 B). The comparison of the MT morphology of the null mutant, wild-type cells, and cells expressing elevated levels of MHP1 suggests that Mhplp is required for the formation or stabi- lization of MTs and that the gene dosage of MHP1 is cru- cial for correct MT function.
Partial Rescue of Lethal Alleles of MHP1 with the Drosophila 205K MAP
Since Mhplp could be considered as a MAP based on in vitro MT-binding assays, it was important to test whether the lethal phenotype of the mhpl-A1 or rnhpl-A6 alleles could be rescued by the expression of another MAP. Can- didate proteins were the Drosophila 205K MAP because of its immunological relationship with Mhplp, and the mammalian MAP4 because of its sequence homology with Mhplp. Heterozygous mhpl÷/mhpl-Al and mhpl+/rnhpl- A6 cells were transformed with the 205K MAP eDNA un- der the control of the ADH promoter, and Leu + transfor- mants were sporulated and tetrads dissected. In 8 out of 12 dissected tetrads, two colonies of normal size and two small colonies were seen after 5 d of incubation at 30°C (Fig. 10 A, inset), the two small colonies being Leu + Trp + or Leu + G418 resistant, respectively. Dissection of the Y502 and Y561 strains under identical conditions repro- ducibly showed that all complete tetrads developed only two viable spores after 5 d of incubation at 30°C. In tests of the ability of the mouse MAP4 eDNA, cloned into the same plasmid pADNS, to compensate the rnhpl-A1 muta-
Unbudded Time of
Cell line induction* Mononncleate
Large bud
Monopolar spindle Short spindle Elongated spindle
h % (n)
Y501 0 97 (118) 0 0 3 (4) Y501 4 96 (205) 0 2 (4) 2 (4) Y501 12 96 (160) 0 1 (2) 3 (5) Y501 20 97 (174) 0 1 (2) 2 (4) Y506 0 91 (153) 0 2 (3) 7 (12) Y506 4 45 (87) 14 (25) 24 (46) 18 (35) Y506 12 39 (37) 14 (13) 33 (31) 14 (13) Y506 20 22 (17) 25 (19) 48 (35) 5 (4)
*Time of induction indicates hours of growth in galactose-containing medium. Numbers in other columns correspond to percentages and numbers of ceils observed with the indicated phenotype.
The Journal of Cell Biology, Volume 135, 1996 1334
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996
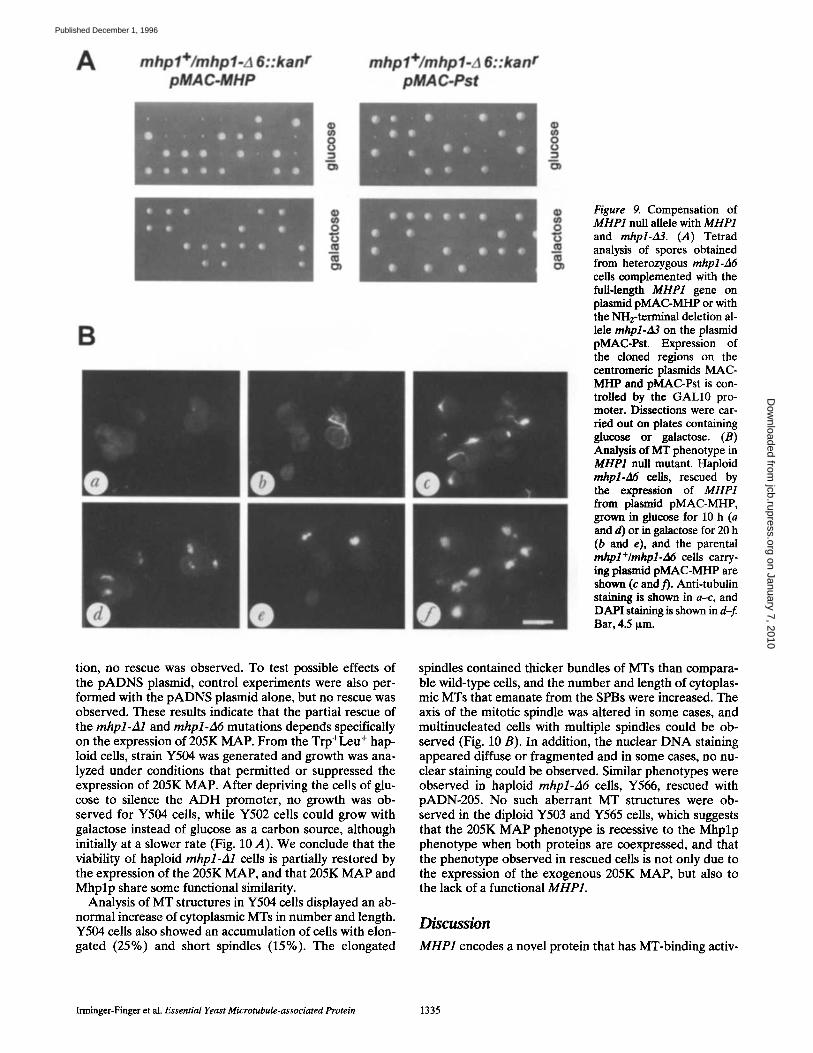
A mhpl+/mhpl-A 6::kanr pMAC-MHP
mhp l+/m hp l.Ll 6 : : ka nr pMAC-Pst
8 o w
8 m
0
B
_m m
O
Figure 9. Compensation of MHP1 null allele with MHP1 and mhpl-d3. (A) Tetrad analysis of spores obtained from heterozygous mhpl-A6 cells complemented with the full-length MHP1 gene on plasmid pMAC-MHP or with the NH2-terminal deletion al- lele mhpl-zl3 on the plasmid pMAC-Pst. Expression of the cloned regions on the centromeric plasmids MAC- MHP and pMAC-Pst is con- trolled by the GALl0 pro- moter. Dissections were car- ded out on plates containing glucose or galactose. (B) Analysis of MT phenotype in MHP1 null mutant. Haploid mhpl-A6 cells, rescued by the expression of MHP1 from plasmid pMAC-MHP, grown in glucose for 10 h (a and d) or in galactose for 20 h (b and e), and the parental mhpl+/mhpl-A6 cells carry- ing plasmid pMAC-MHP are shown (c and t). Anti-tubulin staining is shown in a-c, and DAPI staining is shown in d-f. Bar, 4.5 p,m.
tion, no rescue was observed. To test possible effects of the pADNS plasmid, control experiments were also per- formed with the pADNS plasmid alone, but no rescue was observed. These results indicate that the partial rescue of the mhpl-A1 and mhpl-A6 mutations depends specifically on the expression of 205K MAP. From the Trp+Leu + hap- loid cells, strain Y504 was generated and growth was ana- lyzed under conditions that permitted or suppressed the expression of 205K MAP. After depriving the cells of glu- cose to silence the A D H promoter, no growth was ob- served for Y504 cells, while Y502 cells could grow with galactose instead of glucose as a carbon source, although initially at a slower rate (Fig. 10 A). We conclude that the viability of haploid mhpl-A1 cells is partially restored by the expression of the 205K MAP, and that 205K MAP and Mhplp share some functional similarity.
Analysis of MT structures in Y504 cells displayed an ab- normal increase of cytoplasmic MTs in number and length. Y504 cells also showed an accumulation of cells with elon- gated (25%) and short spindles (15%). The elongated
spindles contained thicker bundles of MTs than compara- ble wild-type cells, and the number and length of cytoplas- mic MTs that emanate from the SPBs were increased. The axis of the mitotic spindle was altered in some cases, and multinucleated cells with multiple spindles could be ob- served (Fig. 10 B). In addition, the nuclear D N A staining appeared diffuse or fragmented and in some cases, no nu- clear staining could be observed. Similar phenotypes were observed in haploid mhpl-A6 cells, Y566, rescued with pADN-205. No such aberrant MT structures were ob- served in the diploid Y503 and Y565 cells, which suggests that the 205K MAP phenotype is recessive to the Mhplp phenotype when both proteins are coexpressed, and that the phenotype observed in rescued cells is not only due to the expression of the exogenous 205K MAP, but also to the lack of a functional MHP1.
Discussion
MHP1 encodes a novel protein that has MT-binding activ-
Irminger-Finger et al. Essential Yeast Microtubule-associated Protein 1335
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996
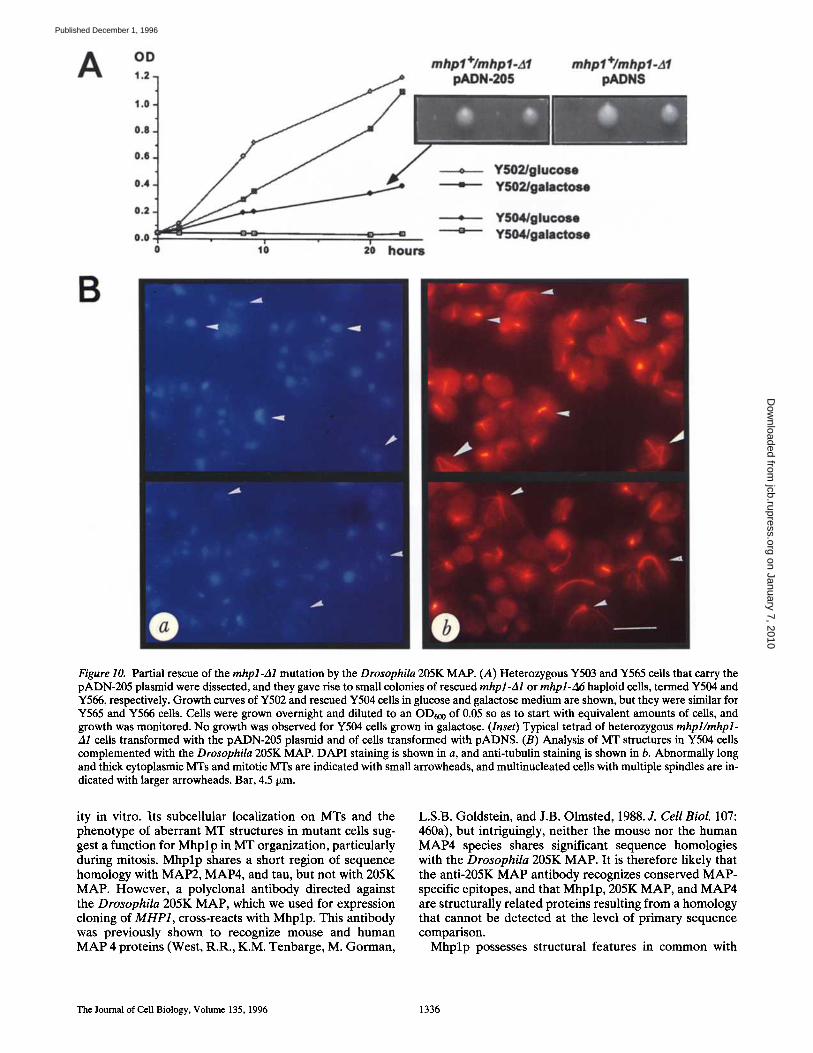
A OD 1.2
1 . 0
0 . 8
0 . 0
0.4
0.2
0.0 T - = = "~ .... - o '~ 20 ho.m
B
mhpl+lmhpl.al pN)N.20S
mhpl+/mhpl.A1 pAtoiS
• w ~ u m u ~
YfdM/oluoooo = YM4/oolNtoee
Figure 10. Partial rescue of the mhpl-zal mutation by the Drosophila 205K MAP. (A) Heterozygous Y503 and Y565 cells that carry the pADN-205 plasmid were dissected, and they gave rise to small colonies of rescued mhpl-A1 or rnhpl-A6 haploid cells, termed Y504 and Y566, respectively. Growth curves of Y502 and rescued Y504 cells in glucose and galactose medium are shown, but they were similar for Y565 and Y566 cells. Cells were grown overnight and diluted to an OD600 of 0.05 so as to start with equivalent amounts of cells, and growth was monitored. No growth was observed for Y504 ceils grown in galactose. (Inset) Typical tetrad of heterozygous mhpl/mhpl- A1 cells transformed with the pADN-205 plasmid and of cells transformed with pADNS. (B) Analysis of MT structures in Y504 cells complemented with the Drosophila 205K MAP. DAPI staining is shown in a, and anti-tubulin staining is shown in b. Abnormally long and thick cytoplasmic Mrs and mitotic MTs are indicated with small arrowheads, and multinucleated cells with multiple spindles are in- dicated with larger arrowheads. Bar, 4.5 ~m.
ity in vitro. Its subcellular localization on MTs and the phenotype of aberrant MT structures in mutant ceils sug- gest a function for Mhplp in MT organization, particularly during mitosis. Mhplp shares a short region of sequence homology with MAP2, MAP4, and tau, but not with 205K MAP. However, a polyclonal antibody directed against the Drosophila 205K MAP, which we used for expression cloning of MHP1, cross-reacts with Mhplp. This antibody was previously shown to recognize mouse and human MAP 4 proteins (West, R.R., K.M. Tenbarge, M. Gorman,
L.S.B. Goldstein, and J.B. Olmsted, 1988. J. Cell Biol. 107: 460a), but intriguingly, neither the mouse nor the human MAP4 species shares significant sequence homologies with the Drosophila 205K MAP. It is therefore likely that the anti-205K MAP antibody recognizes conserved MAP- specific epitopes, and that Mhplp, 205K MAP, and MAP4 are structurally related proteins resulting from a homology that cannot be detected at the level of primary sequence comparison.
Mhplp possesses structural features in common with
The Journal of Cell Biology, Volume 135, 1996 1336
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

other known MAPs. Its estimated size on SDS gels is 50% larger than its calculated molecular weight, an observation made for other highly charged MAPs (Irminger-Finger et al., 1990; West et al., 1991). It is possible, however, that the 230-, 200-, and 150-kD proteins that are recognized by anti-MHP1 antibodies are isoforms of Mhplp generated by posttranslational modification. Mhplp is highly charged and the putative MT-binding domain is followed by an acidic COOH-terminal region, which is similar to the organization of tau and 205K MAP (Lee et al., 1988; Irminger-Finger et al., 1990). In contrast with MAP2, tau, and 205K MAP consisting of an acidic NH2-terminal por- tion, a basic domain containing the MT-binding region, and in the case of tau and 205K MAP an acidic COOH- terminal region, Mhplp contains alternating acidic and ba- sic regions, and its acidity gradually increases from the NH2-terminal to the COOH-terminal end. However, the density of potential phosphorylation sites gradually de- creases from the NH2-terminal to the COOH-terminal end (Fig. 1 B) and could lead to more negative charges within the NH2-terminal half of the protein.
Phosphorylation has been characterized as an important mechanism for the regulation of the interaction of MAPs with MTs. In contrast to tau, MAP2, and 205K MAP, the potential kinase target sites on the amino acid sequence of Mhplp localize within the NH2-terminal two-thirds of the protein with fewer sites in the COOH-terminal portion containing the putative MT-binding region. It is therefore possible that MT binding in Mhplp is less influenced by phosphorylation than by other functions residing in the NH2-terminal portion of Mhplp. An important function for the NH2-terminal domain of Mhplp is also suggested by the phenotype induced by the rnhpl-A3 allele (Fig. 9).
We have demonstrated that the region of Mhplp suffi- cient and essential for MT binding is located within the COOH-terminal 210 amino acid residues of Mhplp. The localization of the AP homologous sequence motif within this region is consistent with its role in MT binding. How- ever, the region found to be homologous to the AP motif is less basic than other AP sequences, although flanked by sequences containing basic residues (Figs. 1 B and 2). It is conceivable that these adjacent basic sequences partici- pate in MT binding in Mhplp or that the binding is achieved without a predominantly basic charge, as was re- ported for binding motifs of other MAPs (Hemphill et al., 1992). In support of this idea is the finding that an interre- peat region of the AP repeats in tau has higher MT affinity than a single repeat (Goode and Feinstein, 1994). A signif- icant contribution of the sequences outside the AP motif, which was also reported for MAP4 (Olson et al., 1995), might explain why for Mhplp a single repeat could be suf- ficient for MT binding. The reduced binding activity of the deletion N1300, containing the AP-homologous sequence but not the entire COOH terminus of Mhplp, could be due to a participation of sequences close to the COOH terminus to MT binding. It is also possible that the protein folding of the deletion N1300 is altered when the COOH terminus is missing.
The generation of Mhplp-specific antibodies permitted the identification of Mhplp as a 200-kD protein on West- ern blots comigrating with bacterially produced Mhplp. Minor bands of 150, 90, and 60 kD also react with anti-
MHP1 and are increased in Y509 cells overproducing Mhplp. These proteins could be degradation products, isoforms, or products of differential transcription and translation. The former possibility would be consistent with the expression of a 2.8-kb mRNA observed in wild- type cells, which hybridized to the 3'-end probe of MHP1 (Fig. 7 A). The differential transcription and/or translation could be a regulatory mechanism for the expression of MHP1 gene products encoding different functions. The lo- calization of the anti-MHPl-reactive epitope to cytoplasmic and nuclear MTs is consistent with MT-binding experi- ments. However, since anti-MHP1 was generated against the COOH-terminal peptide Cl188p, we cannot exclude that the staining observed in immunofluorescence analy- ses is due to cross-reaction with isoforms of Mhplp <200 kD, as observed on Western blots.
Indirect evidence that differential transcription and translation could lead to functional products comes from the viable mhpl-A2 mutation, derived from an insertion in the NH2-terminal half of the protein-coding region, that disrupts the ORF after residue 557. It seems likely that mhpl-A2 mutant cells express both NH2- and COOH-ter- minal truncation products of MHP1, since two separate transcripts are detected. The translation of a COOH-ter- minal 90-kD protein, detected with anti-MHP antibodies in mhpl-A2 cells (Fig. 7 B), could be initiated at one of the candidate methionine initiation codons located at posi- tions 598 and 612 (Fig. 2 A). The mhpl-A4 mutation delet- ing coding sequences from residues 1--613, including the two potential translation initiation codons, is lethal in hap- loid cells. Also, the mhpl-A5 mutation, deleting residues downstream of the URA3 ÷ insertion site in the mhpl-A2 mutation, results in lethality in haploid cells, supporting the conclusion that transcription initiation can occur at po- sitions 598 or 612. Viability of the null mutant could also be restored with plasmid pMAC-Pst (Fig. 9 A) containing the region downstream from amino acid position 561, demonstrating that the COOH-terminal region is suffi- cient for viability without the NH2-terminal portion of the gene expressed separately.
The results of gene disruption experiments and in vitro MT-binding experiments suggest that Mhplp is organized in a COOH-terminal domain (residues 821-1,398) essen- tial for MT binding, and an NH2-terminal domain (resi- dues 1-561) important for proper function of Mhplp, which is absent in the mhpl-A3 mutation. Deleting the re- gion of MHP1 that includes the domain essential for MT binding leads to lethality in haploid cells (Fig. 6), but the NH2-terminal deletion allele mhpl-A3 can partially com- pensate lethality of the complete null allele, demonstrating that mhpl-A3 provides sufficient MHP1 function for via- bility (Fig. 9). The overexpression of the mhpl-A3 allele leads to reduced growth with a specific phenotype of het- erogeneously arrested cells at different stages of mitosis mostly before the onset of anaphase. The number of mutant cells is correlated with the expression level of the mutant protein. In contrast with mutants effecting SPB du- plication, such as karl and cdc31 (Vallen et al., 1994), mhpl-A3 cells arrest at various times of spindle formation, and it is the number of cells with elongated spindles and divided nuclei that becomes more reduced when the ex- pression of mhpl-A3 is increased (Table III). The hetero-
Irminger-Finger et al. Essential Yeast Microtubule-associated Protein 1337
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

geneous phenotype of inhibition of the progression through anaphase strongly suggests a structural defect compatible with a role in MT stabilization and/or formation.
A different phenotype was observed with the overex- pression of the entire MHP1. The additional production of Mhplp results in increased formation and/or stabilization of cytoplasmic MTs. It is possible that the protein encoded on rnhpl-A3 is missing a signal that retains it in the cyto- plasm or that the truncated form is more efficiently trans- ported to the nucleus interacting with nuclear MTs. Re- ports from a number of laboratories demonstrate that the interaction of MAP4-type proteins with MTs is regulated during the cell cycle by phosphorylation by specific kinases (Aizawa et al., 1991; Vandrd et al., 1991; Ookata et al., 1995). Therefore, a possible explanation for the cell cycle block observed in rnhpl-A3 cells and the phenotype in- duced by overexpression of MHP1 could be deregulated phosphorylation. Increased levels of Mhplp or the mutant protein encoded on rnhpl-A3 could expend the capacities of different specific kinases that regulate the interaction of Mhplp with MTs. Mutational analyses and biochemical approaches should provide information on the function of the NH2-terminal domain of Mhplp and help to identify regulatory factors that interact with Mhplp, such as ki- nases.
All mutations induced by gene disruption, overexpres- sion of the entire MHP1, or the COOH-terminal part of MHP1 affect the formation of MTs and the function of MTs during the progression through cell cycle. A reduced gene dosage in heterozygous mhpl+/mhpl-Al::TRP1 and mhpl+/mhpl-Al::kan r cells appears to affect cells at late mitosis, since in an exponentially growing culture, the number of cells at this stage is elevated. In cells overex- pressing the entire MHP1 gene, we observe long cytoplas- mic MTs, indicating that an increased concentration of Mhplp promotes MT polymerization and/or increases MT stability. In complete null mutants Y564, expressing mini- mal amounts of Mhplp from the "silenced" GALl0 pro- moter, the formation of MTs is reduced. In Y504 and Y566 cells rescued by the expression of 205K MAP, an excess of aberrant MT structures can be observed as a result of the MT-stabilizing effect of overexpressed 205K MAP. A sim- ilar phenotype had been observed in the fission yeast when mammalian MAP4 was overexpressed (Olmsted, J.B., and J.R. Mclntosh. 1994. Mol. Biol. Cell. 5:169a). These data suggest that Mhplp regulates the fine tuning of MT stability that cannot be achieved by the expression of a truncated form of Mhplp or by a surrogate MAP from another species. Future research will focus on the timing and nature of the essential interactions of Mhplp with MTs by using mhpl-A6 haploid cells complemented by mutagenized inducible MHP1 on expression plasmids. Overall, the system described here should also permit the determination of functions generally possessed by MAPs by domain-swapping experiments between Mhplp and other MAPs, and by the testing of hybrid proteins in MHP1 null mutants.
We are grateful to Drs. A. Pereira, R. Ortega Perez, and W.D. Heyer for helpful comments and critical reading of the manuscript; M. Princevalle for generation of anti-MHP1 antibodies; and N. Mathis for excellent tech- nical help. We are indebted to Dr. L. Huber for useful suggestions and dis-
cussions, Dr. O. Schaad for specific computer programs, and Prof. J. Olm- sted for the mouse MAP4 eDNA clone.
This work was supported by a grant from the Swiss National Science Foundation (to S.J. Edelstein).
Received for publication 1 July 1996 and in revised form 9 August 1996.
References
Aizawa, H., H. Kawasaki, H. Murofushi, S. Kotani, K. Suzuki, and H. Sakai. 1989. A common amino acid sequence in 190-kDa microtubule-associated protein and tan for the promotion of microtubule assembly. J. Biol. Chem. 264:5885-5890.
Aizawa, H., Y. Emori, H. Murofushi, H. Kawasaki, H, Sakai, and K. Suzuki. 1990. Molecular cloning of a ubiquitously distributed microtubule-associated protein with M~ 190,000. J. Biol. Chem. 265:13849-13855.
Aizawa, H., M. Kamijo, Y. Ohba, A. Mori, K. Okuhara, H. Kawasaki, H. Muro- fushi, K. Suzuki, and H. Yasuda. 1991. Microtubule destabilization by cdcd2/ H1 histone kinase: phosphorylation of a "Pro-rich region" in the microtu- bule-binding domain of MAP4. Biochem. Biophys. Res. Commun. 179:1620- 1626.
Barnes, G., A. Louie, and D. Botstein. 1992. Yeast proteins associated with mi- crotubules in vitro and in vivo. Mol. Biol. Cell. 3:29-47.
Bellocq, C., I. Andrey-Toruare, A.-M. Paunier, B. Maeder, L. Paturle, D. Job, J. Haiech, and S. Edelstein. 1992. Purification of assembly-competent tubu- lin from Saccharomyces cerevisiae. Fur. J. Biochem. 210:343-349.
Berlin, V., C.A. Styles, and G.R. Fink. 1990. BIK1, a protein required for mi- erotubule function during mating and mitosis in Saccharomyces cerevisiae, colocalizes with tubulin. J. Cell Biol. 111:2573-2586.
Botstein, D., S.C. Falco, S.E. Stewart, M. Breunan, S. Scherer, D.T. Stinch- comb, K. StruM, and R.W. Davis. 1979. Sterile host yeasts (SHY): a eukary- otic system of biological containment for recombinant DNA experiments. Gene (Amst.). 8:17-24.
Bulinski, J.C., and G.G. Borisy. 1979. Self-assembly of microtubules in extracts of cultured HeLa cells and the identification of HeLa mierotubule-associ- ated protein. Proc. Natl. Acad. Sci. USA. 76:293-297.
Bulinski, J.C., and G.G. Borisy. 1980. Widespread distribution of a 210,000 tool wt microtubule-associated protein in cells and tissues of primates. J. Cell Biol. 87:802-808.
Chapin, S.J., and J.C. Bulinski. 1991. Non-neural 210 x 10 a Mr microtubule- associated protein (MAP4) contains a domain homologous to the microtu- bule-binding domains of neural MAP2 and tan. J. Cell Sci. 98:27-36.
Chou, P., and G.D. Fasman. 1978. Prediction of the secondary structure of pro- teins from their amino acid sequence. Adv. Enzymol. 47:45-147.
Cleveland, D,W., S.Y. Hwo, and M.W. Kirschner. 1977. Physical and chemical properties of purified tan factor and the role of tan in microtubule assembly. J. Mol. Biol. 116:227-247.
Colicelli, J., C. Birchmeier, T. Michaeli, K. O'Neill, M. Riggs, and M. Wigler. 1989. Isolation and characterization of a mammalian gene encoding a high- affinity cAMP phosphodiesterase. Proc. Natl. Acad. Sci. USA. 86:3599-3603.
Devereux, J., P. Haeberli, and O. Smithies. 1984. A comprehensive set of se- quence analysis programs for the VAX. Nucleic Acids Res. 12:387-395.
Diaz-Nido, J., L. Serrano, E. Mendez, and J. Avila. 1988. A casein kinase II- related activity is involved in phosphorylation of microtubule-associated protein MAP1B during neuroblastoma cell differentiation. J. Cell Biol. 106: 2057-2065.
Edelmann, A.M., D.K. Blumenthal, and E.G. Krebs. 1987. Protein serine/thre- onine kinases. Annm Rev. Biochem. 56:567-613.
Gamier, J., D.J. Osguthorpe, and B. Robson. 1978. Analysis of the accuracy and implications of simple methods for predicting the secondary structure of globular proteins. J. Mol. Biol. 120:97-120.
Goldstein, L.S.B., R.A. Laymon, and J.R. Mclntosh. 1986. A microtubnie-asso- ciated protein in Drosophila melanogaster, identification, characterization, and isolation of coding sequences. J. Cell Biol. 107:2076-2087.
Gonzales, G.A., K.K. Yamamoto, W.H. Fischer, D. Karl P. Meuzel, I. Biggs, W.W. Vale, and M.R. Montminy. 1989. A cluster of phosphorylation sites on the cyclic AMP-regulated nuclear factor CRBE predicted by its sequence. Nature (Lond.). 337:749-752.
Goode, B.L., and S.C. Feinstein. 1994. Identification of a novel microtubule binding and assembly domain in the developmentally regulated inter-repeat region of tau. J. Cell Biol. 124:769-782.
Hasek, J., J. Jochova, P. Draber, V. Viklicky, and E. Streiblova. 1991. Localiza- tion of a 210-kD microtubule-interacting protein in the yeast Saccharomyces cerevisiae. Can. J. Microbiol. 38:149-152.
Hemphill, A., M. Affolter, and T. Seebeck. 1992. A novel microtubule-binding motif identified in a high molecular weight microtubule-assoeiated protein from Trypanosoma brucei. J. Cell Biol. 117:95-103.
Hoyt, M.A., T. Sterns, and D. Botstein. 1990. Chromosome instability mutants of Saccharomyces cerevisiae that are defective in mierotubule-mediated pro- cesses. Mol. Cell. Biol. 10:223-234.
Hoyt, A.M., L. He, K.K. Loo, and W.S. Saunders. 1992. Two Saccharomyces cerevisiae kinesin-related gene products required for mitotic spindle assem- bly. J. Cell Biol. 118:109-120.
The Journal of Cell Biology, Volume 135, 1996 1338
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996

Hurt, E.C. 1988. A novel nucleoskeletal-like protein located at the nuclear pe- riphery is required for the life cycle of the yeast Saccharomyces cerevisiae. EMBO (Eur. Mol. BioL Organ.) J. 7:4323--4334.
Hyman, A.A., K. Middleton, M. Centola, T.J. Mitchison, and J. Carbon. 1992. Microtubule-motor activity of a yeast eentromere-binding protein complex. Nature (Lond. ). 359:533-539.
Interthal, H., C. Bellocq, J. Bahler, V.1. Bashkirov, S. Edelstein, and W.D. Heyer. 1995. A role of Sepl (=Keml, Xrnl) as a mierotubule-associated protein in Saccharomyces cerevisiae. EMBO (Eur. Mol. Biol. Organ.) J. 14: 1057-1066.
Irminger-Finger, I., R.A. Laymon, and L.S.B. Goldstein. 1990. Analysis of the primary sequence and microtubule-binding region of the Drosophila 205K MAP. J. Cell Biol. 111:2563-2572.
Izant, J., J. Weatherbee, and J.R. McIntosh. 1983. A microtubule-associated protein antigen unique to mitotic spindle microtubules in PtK1 cells. J. Cell Biol. 96:424-434.
Jiang, W., K. Middleton, H. Yoon, C. Fouquet, and J. Carbon. 1993. An essen- tial yeast protein, CBF5p, binds in vitro to centromeres and microtubules. Mol. Cell. Biol. 13:4884--4893.
Kilmartin, J.V., and A.E. Adams. 1984. Structural rearrangements of tubulin and actin during the cell cycle of the yeast Saccharomyces. Z Cell Biol. 98: 922-933.
Kilmartin, J.V., B. Wright, and C. Milstein. 1982. Rat monoclonal anti-tubulin antibodies derived by using a new nonsecreting rat cell line. J. Cell Biol. 93: 576-582.
Kilmartin, J.V., S.L. Dyos, D. Kershaw, and J.T. Finch. 1993. A spacer protein in the Saccharomyces cerevisiae spindle pole body whose transcript is cell cy- cle-regulated. Z Cell Biol. 123:1175-1184.
Kimble, M., A.L. Khodjakov, and R. Kuriyama. 1992. Identification of ubiqui- tous high-molecular-mass, heat-stable microtubule-associated proteins (MAPs) that are related to the Drosophila 205-kDa MAP but are not re- lated to the mammalian MAP4. Proc. Natl. Acad. Sci. USA. 89:7693-7697.
Lee, G. 1993. Non-motor microtubule-associated proteins. Curr. Opin~ Cell Biol. 5:88-94.
Lee, G., N. Cowan, and M. Kirschner. 1988. The primary structure and hetero- geneity of tan protein from mouse brain. Science (Wash. DC). 239:265-288.
Lewis, S.A., D. Wang, and N.J. Cowan. 1988. Microtubule-associated protein MAP2 shares a microtubule-binding motif with tau protein. Science (Wash. DC). 242:936-939.
Li, Y.-Y., E. Yeh, and K. Bloom, 1993. Disruption of mitotic spindle orientation in a yeast dynein mutant. Proc. Natl. Acad. Sci. USA. 90:10096-10100.
Lillie, S.H., and S.S. Brown. 1992. Suppression of a myosin defect by a kinesin- related gene. Nature (Lond.). 356:358--361.
Maniatis, T., E.F. Fritsch, and J. Sambrook. 1989. Molecular Cloning: a Labora- tory Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 545 pp.
McMillan, J.N., and K. Tatchell. 1994. The JNM1 gene in the yeast Saccharo- myces cerevisiae is required for nuclear migration and spindle orientation during the mitotic cell cycle. J. Cell Biol. 125:143-158.
Meluh, P.B., and M.D. Rose. 1990. KAR3, a kinesin-related gene required for yeast nuclear fusion. Cell. 60:1029-1041.
Munro, S., and H.R.B, Pelham. 1984. Use of peptide tagging to detect proteins expressed from cloned genes: deletion mapping functional domains of Drosophila hsp70. EMBO (Eur. Mol. Biol. Organ.) J. 3:3087-3093.
Murphy, D.B., R.B. Vallee, and G.G. Borisy. 1977. Identity and polymeriza- tion-stimulatory activity of the non-tubulin proteins associated with microtu- bules. Biochemistry. 16:2598-2605.
Noble, M., S.A. Lewis, and N.J. Cowan. 1989. The microtubule-binding domain of microtubule-associated protein MAP1B contains a repeated sequence motif unrelated to that of MAP2 and tan. J. Cell Biol. 109:3367-3376.
Olmsted, J.B. 1986. Analysis of cytoskeletal structures using blot-purified monospecific antibodies. Methods Enzymol. 134:467--472.
Olson, K.R., J.R. Mclntosh, and J.B. Olmsted. 1995. Analysis of MAP4 func- tion in living cells using green fluorescent protein (GFP) chimeras. J. Cell
Biol. 130:639-650. Ookata, K., S. Hisanaga, J.C. Bulinski, H. Murofushi, H. Aizawa, T.J. Itoh, H.
Hotani, E. Okumura, K. Tachibana, and T. Kishimoto. 1995. Cyclin B inter- action with microtubule-associated protein 4 (MAP4) targets p34cdc2 kinase to microtubules and is a potential regulator of M-phase microtubule dynam- ics. Z Cell BioL 128:849-862.
Ortega Perez, R., I. Irminger-Finger, J.F. Arrighi, N. Capelli, D. van Tulnen, and G. Turian. 1994. Identification and partial purification of calmodulin- binding microtubule-associated proteins from Neurospora crassa. Eur. J. Biochem. 226:303-310.
Pasqualone, D., and T.C. Huffaker. 1994, STU1, a suppressor of a 13-tubulin mutation, encodes a novel and essential component of the yeast mitotic spin- dle. Z Cell Biol. 127:1973--1984.
Pearson, R.B., J.R. Woodgett, P. Cohen, and B.E. Kemp. 1985. Substrate speci- ficity of a multifunctional calmodulin-dependent protein kinase. J. Biol. Chem. 260:14471-14476.
Pearson, W.R., and D.J. Lipman. 1988. Improved tools for biological sequence comparison. Proc. Natl. Acad. Sci. USA. 85:2444-2448.
Pereira, A., J. Doshen, E. Tanaka, and L.S.B. Goldstein. 1992. Genetic analysis of a Drosophila microtubule-assoeiated protein. J. Cell Biol. 116:377-383.
Roberts, B. 1989. Nuclear location signal-mediated protein transport. Biochem. Biophys. Actor 1008:263-280.
Roof, D.M., P.M. Meluh, and M.D. Rose. 1992. Kinesin-related proteins re- quired for assembly of the mitotic spindle. J. Cell Biol. 118:95-108.
Rose, M.D., F. Winston, and P. Hierter. 1990. Methods in Yeast Genetics. A Laboratory Course Manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY. 198 pp.
Rothstein, R.J. 1983. One step gene disruption in yeast. Methods Enzymol. 101: 202-210.
Sanger, F., S. Michlen, and A.R. Coulson. 1977. DNA sequencing with chain terminating inhibitors. Proc. Natl. Acad. Sci. USA. 74:5463-5467.
Schoenfeld, T.A., L. MeKerracher, R. Obar, and R.B. Vallee. 1989. MAP 1A and MAP 1B are structurally related microtubule-associated proteins with distinct developmental patterns in the CNS. Z Neurosci. 9:1712-1730.
Sherman, F., G.R. Fink, and J.B. Hicks. 1986. Methods in Yeast Genetics. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY. 163-167.
Sikorski, R.S., and P. Hieter. 1989. A system of shuttle vectors and yeast host strains designed for efficient manipulation of DNA in Saccharomyces cerevi- siae. Genetics. 122:19-27.
Steams, T., A. Hoyt, and D. Botstein. 1990. Yeast mutants sensitive to anti- microtubule drugs define three genes that regulate microtubule function. Genetics. 124:251-262.
Strohm, M., P. Vollmer, T.J. Tan, and D. Gallwitz. 1993. A yeast GTPase-acti- vating protein that interacts specifically with a member of the YPT/RAB family. Nature ( Lond. ). 361:736-739.
Studier, W., A.H. Rosenberg, J.J. Dunn, and J.W. Dubendorff. 1990. Use of the T7 RNA polymerase to direct expression of cloned genes. Methods Enzy- tool. 185:60-89.
Vallee, R.B. 1982. A taxol-dependent procedure for the isolation of microtu- bules and microtubule-associated proteins (MAPs). J. Cell Biol. 92:435-442.
Vallen, E.A., W. Ho, M. Winey, and M.D. Rose. 1994. Genetic interactions be- tween CDC31 and KARl, two genes required for duplication of the micro- tubule organizing center in Saccharomyces cerevisiae. Genetics. 137:407--422.
Vandr6, D.D., V.E. Centonze, J. Peloquin, R.M. Tombes, and G.G. Borisy. 1991. Proteins of the mammalian mitotic spindle: phosphorylation/dephos- phorylation of MAP-4 during mitosis. Z Cell Sci. 98:577-588.
Wach, A., A. Brachat, R. P6hlmann, and P. Philippsen. 1994. New heterologous modules for classical or PCR-based gene disruptions in Saccharomyces cere- visiae. Yeast. 10:1793-1808.
West, R.R., K.M. Tenbarge, and J.B. Olmsted. 1991. A model for microtubule- associated protein 4 structure. J. Biol. Chem. 266:21886-21896.
Wiche, G., C. Oberkanins, and A. Himmler. 1991. Molecular structure and function of microtubule-associated proteins. Int. Rev. Cytol. 124:217-273.
Irminger-Finger et al. Essential Yeast Mtcrotubule-associated Protein 1339
on January 7, 2010 jcb.rupress.org
Dow
nloaded from
Published December 1, 1996





























