Embed Size (px)
Citation preview

TOXICOLOGY AND APPLIED PHARMACOLOGY 81,5 17-524 ( 1985)
Methyl Ethyl Ketone Peroxide Damage to Cytochrome P-450 Peroxidase Activities’
MITSURU ANDO AND AL L. TAPPEL
Department of Food Science and Technology, University of California, Davis, California 95616
Received May 6, 1985; accepted August 10, 1985
Methyl Ethyl Ketone Peroxide Damage to Cytochrome P-450 Peroxidase Activities. ANDO, M., AND TAPPEL, A. L. (1985). Toxicol. Appl. Pharmacol. 81, 517-524. Some new aspects of damage to cytochrome P-450 pcroxidase activities by a toxic peroxide were investigated. Methyl ethyl ketone peroxide (MEKP) damage to cytochrome P-450 and inhibition of cytochrome P- 450-mediated pe.roxidase activities were studied. In vivo, hepatic microsomal NADH- and NADPH- peroxidase activities were induced in rats by phenobarbital and 3-methylcholanthrene. Pheno- barbital treatment induced more NADH- and NADPH-peroxidase activity than did 3-methyl- cholanthrene treatment. In vitro, microsomal cytochrome P-450 from rat liver was more sensitive to damage by MEKP than was this protein from rat kidney. Destruction of cytochrome P-450 hemoprotein and inhibition of its associated peroxidase activities by MEKP increased as a function of time of exposure to the peroxide. The addition of MEKP to microsomes resulted in a progressive increase in spectral absorbance at 433 nm. MEKP was an irreversible inhibitor of NADH- and NADPH-peroxidase, and NADH-peroxidase was more sensitive to damage than was NADPH- peroxidase. At 0 time and low concentration, MEKP was immediately bound to microsomal cytochrome P-450 and it exhibited competitive inhibition of NADH- and NADPH-peroxidase. At high concentration, MEKP was a mixed inhibitor of NADH- and NADPH-peroxidase. This study also showed that MEKP is a substrate for microsomal P-450 peroxidase activities and suggests that these peroxidase activities may play an important role in metabolism of toxic lipophilic organic peroxides that enter the endoplasmic reticulum. o 1985 Academic RSS. I~C.
Methyl ethyl ketone peroxide (MEKP), a li- pophilic organic peroxide, is used industrially to initiate polymerization reactions in the manufacture of plastics and fabrics. MEKP decomposes to free radicals in the presence of transition metal ions at room temperature (Sheppard and Mageli, 1982). The routes of entry of MEKP into the body are via inhala- tion, ingestion, and skin and eye absorption. When given orally or intraperitoneally to mice or rats, MEKP causes gross hemorrhage, lung injury, and mild liver damage; mice given MEKP develop malignant tumors (U.S. En-
’ This research was supported by Grant AM-09933 from the National Institute of Arthritis, Diabetes, Digestive and Kidney Diseases.
vironmental Protection Agency, 1979; Na- tional Institute of Occupational Safety and Health, 1977; Sitting, 198 1; SPI Bulletin, 1982). Litov et al. (1981) showed that MEKP injected into vitamin E-deficient rats caused severe lipid peroxidation, as determined by measurement of the immediate, high produc- tion of pentane. Expired volatile hydrocar- bons, including pentane, are used as an index of in vivo lipid peroxidation (Dillard et al., 1977; Hafeman and Hoekstra, 1977).
The peroxidation of lipids in biological membranes is a destructive phenomenon as- sociated with a variety of cellular damage. Lipid peroxides damage endoplasmic reticu- lum and inhibit microsomal enzyme activities (Hruszkewycz et al., 1978; Cadenas and Sies,
517 004i-OOSX/85 $3.00 Copyright 8 1985 by Ac&temic Pms, Inc. Au rights of rcpmductiom in my form nxmed.

518 ANDG AND TAPPEL
1982; O’Brien, 1982). Cytochrome P-450, the terminal ox&se of the hepatic microsomal monooxygenase system, is one of the enzymes most sensitive to damage by lipid peroxides (Jeffery et al., 1977). In addition to functioning as a monooxygenase, cytochrome P-450 cat- alyzes hydroperoxide-dependent substrate hy- droxylation, and it has peroxidase activity against lipid hydroperoxides (Hrycay and O’Brien, 1972, 1973, 1974; Nordblom et al., 1976; Rahimtula and O’Brien, 1974, 1977). Via its peroxidase activities, cytochrome P-450 may protect other microsomal components from the damaging effect of various or- ganic hydroperoxides. Therefore, investiga- tions of the relationship between the enzy- matic activities of cytochrome P-450 perox- idases on organic peroxides and inhibition of these activities by the same peroxides are desirable. Since different forms of cytochrome P-450 have been purified and characterized from phenobarbital- and 3-methylcholan- threne-treated animals (Imai and Sato, 1974; Guengerich, 1978; Coon et al., 1979; Ryan et al., 1979; Harada and Omura, 1981) this study investigated the in vivo induction of NADH- and NADPH-peroxidase by these two drugs. The destruction by MEKP of the in- duced and noninduced microsomal cyto- chrome P-450 hemoprotein was examined by spectral studies, and the in vitro inhibition of tetramethylphenylenediamine- (TMPD-), NADH-, and NADPH-peroxidase activities was studied.
METHODS
Animals. Six-week-old, male Sprague-Dawley rat? were fed ad Iibitum a commercial rat diet.’ Phenobarbital4 was dissolved in 1 N NaOH, adjusted to pH 9.5, and admin- istered ip to tats at a dose of 80 mg& body wt on three successive days. 3-Methylcholanthrene4 was dissolved in purified olive oil and administered ip at a dose of 80 mg/kg body wt two times, with a l-day interval between
2 Simonsen Laboratories, Gihoy, Calif. 3 Purina Rat Chow, Ralston Purina Co., St. Louis, MO. ’ Sigma Chemical Co., St. Louis, MO.
injections. The rats were fasted for 24 hr, anesthetized with ether, and killed by severing the main abdominal artery.
Preparation of microsomes. Microsomes from liver and kidney were prepared according to the modified method of Noshiro and Omura (1978). Livers and kidneys were thoroughly perfused in situ, from the heart, with cold 0.9% NaCl. The perfused tissues were homogenized with 9 vol of0.25 M sucrose/l0 mM Tris-HCl buffer, pH 7.5/l mM EDTA in a Potter-Elvehjem glass-Teflon homogenizer. The homogenates were centrifuged at 900g for 10 min, and then at 10,OOOg for 15 min. The supernatant fraction was centrifuged at 92,OOOg for 70 min. The microsomes were suspended in 0.15 M KCl/ 1 mM EDTA and were centrifuged at 92,000g for 70 min. The washed microsomes were suspended in 0.15 M KCl/lO mM Tris-HCl, pH 7.5/l mM EDTA, and were placed immediately in liquid nitrogen. Measurements of cytochrome P-450 and cyto- chrome b5 hemoprotein were made the same day.
Analytical methods. The concentration of microsomes for assay of cytochromes was 2 mg protein/ml. Protein was measured by a Coomassie blue dye-binding assay (Bradford, 1976) with ovalbumin as a standard. Cyto- chromes P-450 and b5 in liver microsomes were deter- mined according to the methods described by Omura and Sato (1964a), and Estabrook and Werringloer ( 1978) with extinction coefficients of 9 1 mM-‘Cm-’ (between 490 and 450 nm) and 185 mM&m-’ (between 426 and 409 nm), respectively. Cytochrome P-450 in kidney microsomes was determined according to the method of Grrenius et al. (1973).
Difference spectra showing the binding of MEKP to microsomal P-450 were measured according to the pro- cedure of Jefcoate (1978). Repetitive scanning at 50 nm/min with a scanning spectrophotomete? was used to produce a difference spectrum between 350 and 500 nm. Microsomes from control rats, suspended at a concentra- tion of 2 mg protein/ml, and 0.98 nmol cytochrome P- 45O/mJ 100 mM potassium phosphate, pH 7.5, were placed in both reference and sample cuvettes at 22°C so that a baseline correction could be made. Microsomes incubated in 100 mM potassium phosphate, pH 7.5/5 mM ME@ were then placed in the sample cuvette, and the difference spectrum was recorded.
Enzyme assays. The induced forms of cytochrome P- 450 and the noninduced forms from control rats were defined by measurement of aminopyrine N-demethylase (Orrenius, 1965) and benzo(cy)pyrene hydroxylase (Nebert and Gelboin, 1968). All incubation media contained 0.625 mM S-AMP to prevent breakdown of NADPH by micro- somal pyrophosphatase (Werringloer, 1978). The concen- tration of microsomes in the reaction mixtures was 2 mg protein/ml. In the assays of aminopyrine oxidation, 2 mM
5 Varian DMSlOO, Varian, Palo Alto, calif. 6 PolySciences, Warrington, Pa.

DAMAGE TO CYTGCHROME P-450 ACTIVITIES 519
NADPH/S mM MgC&/5 mM aminopyrene/2 mM NADH by transition metal-catalyzed decomposition of MEKP. were used. In assays of benzo(&pyrene hydroxylase, 0.4 The incubation mixtures were maintained at 22°C for 0 mu NADPH/O. 1 mM benzo(a)pyrene/ 1 mu NADH were to 30 min, after which, equal volumes of the mixtures used. Aminopyrine Ndemethylase (Werringloer, 1978) were placed into sample and reference cuvettes, and the and benzo(a)pyrene hydroxylase (Nebert and Gelboin, peroxidase activity was measured as described above. The 1968) assays were done at 37°C for 5 and 10 min, re- reaction rates were corrected for absorbance change in the spectively. absence of cumene hydroperoxide.
Assay of TMPD oxidation activity was performed as described by O’Brien and RahimtuIa (1978). The 0.8-ml reaction mixtures consisted of 80 mM Tris-HCl, pH 7.5/l mM EDTA/l.S mM TMPD/0.3 mhi sodium azide/ 0.4 mg microsomal protein per ml/O.5 mM cumene hy- droperoxide substrate. Reaction rates were corrected for TMPD oxidation in the absence of cumene hydroperoxide and were measured by following the rate of Wurster’s blue free-radical formation at 6 10 nm, with an extinction coef- ficient of 11.6 mM-‘cm-‘.
To measure the destruction of cytochromes f-450 and b5, MEKP was added to the microsomal suspensions at concentrations of 6 to 6000 pM. The incubation mixtures consisted of 0.15 M KCl/lO mM Tris-HCl, pH 7.5/l mM EDTA/6 mg microsomal protein/ml. After being held at 22°C for 0 to 30 min, glutathione was added at a final concentration of 2 to 20 mM to reduce unreacted MEKP. Equal volumes of the mixtures were placed into sample and reference cuvettes, and cytochromes P-450 and b5 were measured as described above.
NADPH- and NADH-peroxidase activities were deter- mined according to the methods of Hrycay and O’Brien (1973,1974). Unless otherwise indicated, reaction mixtures consisted of 80 mM sodium phosphate buffer, pH 7.5/l mg microsomal protein per ml/O. 15 mM NADPH or NADH/O.S mM cumene hydroperoxide substrate. Reac- tion rates were cormcted for NADPH or NADH oxidation in the absence of cumene hydroperoxide and were cal- culated with an extinction coefficient for NADH and NADPH of 6.22 m&cm-‘. To estimate the reaction ki- netics of NADPH- and NADH-peroxidase, the reaction mixtures contained varying amounts of cumene hydro- peroxide as substrate.
Statistical analysis. Differences between groups were analyzed by Student’s t test, and p < 0.05 was considered significant.
RESULTS
The inhibitory effects of MEKP on the enzymatic ac- tivities of cytochrome P-450 were assessed by measurement of TMPD, NADH-, and NADPH-peroxidase with varying concentrations of cumene hydroperoxide as substrate. The incubation mixtures consisted of 0.15 M KCl/ 10 mM Tris- HCl, pH 7.5/l mM EDTA/6 mg microsomal protein/ml. MEKP was added at concentrations of 6 to 2600 WM.
EDTA was included to prevent formation of free radicals
Hemoprotein, monooxygenase, and perox- idase activities of liver microsomes. To study the in vivo induction of cytochrome P-450 peroxidase activities and their destruction by MEKP, the drug oxidation activities of the microsomes obtained from phenobarbital- and 3-methylcloranthrene-treated rats were inves- tigated. Phenobarbital and 3-methylcholan- threne treatments of rats increased cytochrome P-450 and cytochrome b5 contents in liver microsomes (Table 1). On a protein basis, the microsomal aminopyrine N-demethylation
TABLE 1
HEMOPROTEIN CONTENTS AND NADPH-SUPPORTED OXIDATION OF CHEMICAL COMPOUNDS BY RAT LIVER MICRO.WME~
Treatment group
Cytochrome Cytochrome P-450 b5
(nmol mg protein-‘)
Aminopyrine Benzo(cu)pyrene
[nmol product min -’ mg protein -’ and (nmol P-450-‘)]
Control
Phenobarbital
3-Methylcholanthrene
0.49 * 0.04 0.33 + 0.03 4.1 + 0.9 0.21 rl: 0.03 (9.8 + 4.9) (0.45 f 0.05)
1.02 + 0.05’ 0.43 + 0.05’ 8.8 + 0.5” 0.17 f 0.03 (8.8 + 0.9) (0.17 f 0.02)”
1.18 + 0.25” 0.45 f 0.04” 3.8 + 1.3 1.18 k 0.27” (3.5 f 1.9)0 (1.12 + 0.31)”
’ Significantly different from control group with no treatment (p < 0.05, Student’s t test).

520 AND0 AND TAPPEL
activity of phenobarbital-treated rats was 2 15% greater than that of control rats. On the basis of cytochrome P-450, aminopyrine Ndemethylation activity was 36% lower in 3-methylcholanthrene-treated rats than in control rats. On the basis of protein and cy- tochrome P-450, respectively, 3-methylchol- anthrene treatment increased benzo(a)pyrene hydroxylation activity 562 and 249% over these activities in controls. On the basis of cy- tochrome P-450, benzo(a)pyrene hydroxyl- ation was 38% lower in phenobarbital-treated rats than in controls.
K,,, and V,,, values for cytochrome P-450- mediated NADH- and NADPH-peroxidase in liver microsomes are shown in Table 2. NADH- and NADPH-peroxidase were each induced by phenobarbital and 3-methylchol- anthrene treatments. The NADH-peroxidase activity of phenobarbital- and 3-methylchol- anthrene-treated rats was 463 and 305%, re- spectively, greater than that of control rats. The NADPH-peroxidase activity of pheno- barbital- and 3-methylcholanthrene-treated rats was 5 18 and 265%, respectively, greater than that of control rats. Phenobarbital in- duced more NADH- and NADPH-peroxidase activity than did 3-methylcholanthrene.
Efict of MEKP on cytochrome P-450. The in vitro destructive effect of MEKP on cyto- chrome P-450 increased as a function of in- cubation time up to 20 min. The addition of MEKP to microsomal suspensions resulted in
-0.253- 470 500
WAVELENGTH (nm)
FIG. 1. Change in the difference spectrum induced by addition of 5 mM MEKP to a pooled sample of liver mi- crosomes prepared from three control rats. Experimental procedures are described under Methods. a, 0 time; b, 1 min; c, 6 min; d, 11 min; e, 16 min; and f, 21 min.
an absorbance change in the difference spec- trum that was characterized by a trough at 4 17 nm. Repetitive scanning of the samples re- vealed a progressive increase in absorbance at 433 nm that continued to increase for up to 16 min (Fig. 1). The destructive effect of MEKP on cytochrome P-450 and the asso- ciated TMPD- and NADH-peroxidase activ- ities increased up to 20 min, as shown in Fig. 2.
The destruction of cytochrome P-450 and cytochrome b5 in liver and kidney microsomes after 20-min incubation with MEKP is shown in Fig. 3. Destruction of cytochrome P-450 was biphasic in liver microsomes. Cytochrome P-450 of liver microsomes from phenobarbi-
TABLE 2
K,,, AND V,, VALUES FOR LIVER MICRO~~MAL NADH- AND NADPH-PEROXIDASE
WITH CUMENE HYDROPEROXIDE AS SUBSTRATE
Treatment group
Control Phenobarbital 3-Methylcholanthrene
NADH-peroxidase NADPH-peroxidase
VIII VUi (nmol NADH oxidized (nmol NADPH oxidized
(2) min-’ mg protein-‘) (2) min-’ mg protein -‘)
0.88’ 40 0.34 17 0.20 185 0.20 88 0.36 122 0.67 45
’ Bach value was obtained with a pooled sample of three microsomal preparations.

DAMAGE TO CYTOCHROME P-450 ACTIVITIES 521
FIG. 2. Destruction of cytochrome P-450 (O), TMPD
TlMEtMIN)
peroxidase (m), and NADH-peroxidase (A) by 300 pM MEKP as a function of time. Each value was obtained with a pooled sample of liver microsomes prepared from three control rats.
ml-treated rats was relatively resistant to MEKP, and biphasic destruction was not ev- ident. The extent of MEKP damage to cyto- chrome P-450 prepared from 3-methylchol- anthrene- and phenobarbital-treated rats was similar. Microsomal cytochrome P-450 from kidney was relatively more resistant to de- struction by MEKP than was cytochrome P- 450 from liver. Cytochrome b5 was not af- fected by concentrations of MEKP below 6 mM.
Effect of MEKP on NADH- and NADPH- peroxidase activities. Since liver microsomal cytochrome P-450 was very sensitive to MEKP, the effects of various concentrations of MEKP on cytochrome P-450-mediated
120 *
zs E!" 80 8% CLW of- 40 e ?"
0 r I00 IO' I02 IO’ lo4
MEKP (PM)
FIG. 3. Destruction of hemoprotein in liver and kidney microsomes after 20 min incubation with MEKP. Cyto- chrome P-450 from control rat liver microsomes, (0); liver cytochrome P-450 from phenobarbital-treated rats, (v); liver cytochrome P-450 from 3-methylcholanthrene- treated rats, (A); kidney cytochrome P-450 from control rats, (m); and liver cytochrome b5 from control rats, (e). Each value was obtained with a pooled sample of three microsomal preparations. Experimental procedures are described under Methods.
g! 7100 I- 5 80
560
MEKPtpM)
FIG. 4. Inhibition of liver microsomal NADH- and NADPH-peroxidase activity by MEKP. NADH-peroxi- dase activities with cumene hydroperoxide as substrate, at 0 time, (0); NADH-peroxidase activities after 20 min in- cubation, (A); and NADPH-peroxidase activities after 20 min of incubation, (+). Each value was obtained with a pooled sample of three microsomal preparations from control rats. Assay procedures are described under Meth- OdS.
NADH- and NADPH-peroxidase activities were examined. Cytochrome P-450 peroxidase activities were more sensitive to destruction by MEKP than was the cytochrome P-450 protein (Fig. 2), and NADH-peroxidase was less resistant to damage than was NADPH- peroxidase (Fig. 4). The inhibitory effect of MEKP on NADPH- and NADH-peroxidase
MEKP,,uM
[CUMENE HYDROPEROXIDE] (mM)“
FIG. 5. Double-reciprocal plot of liver microsomal NADH-peroxidase activities with cumene hydroperoxide as substrate. Experimental procedures are described under Methods. (A) 0 time, and (B) after 20 min incubation with MEKP.
522 ANDO AND TAPPEL
0.6 DISCUSSION
[CUMENE HYDKQPEROXIDE] (mM)“
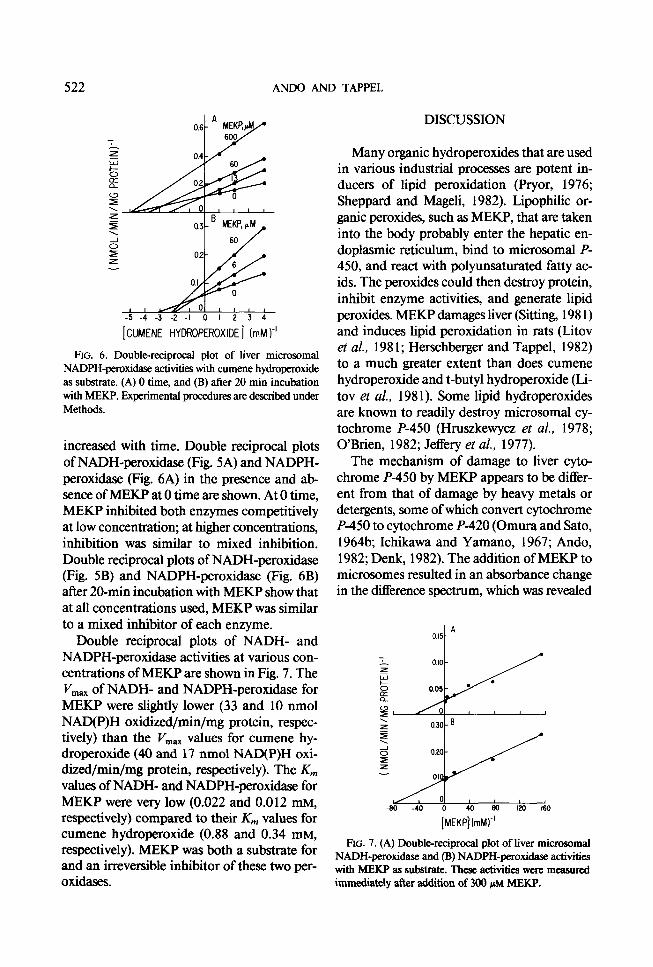
FIG. 6. Double-reciprocal plot of liver microsomal NADPH-peroxidase activities with cumene hydroperoxide as substrate. (A) 0 time, and (B) after 20 min incubation with MEKP. Experimental procedures are described under Methods.
increased with time. Double reciprocal plots of NADH-peroxidase (Fig. 5A) and NADPH- peroxidase (Fig. 6A) in the presence and ab- sence of MEKP at 0 time are shown. At 0 time, MEKP inhibited both enzymes competitively at low concentration; at higher concentrations, inhibition was similar to mixed inhibition. Double reciprocal plots of NADH-peroxidase (Fig. 5B) and NADPH-peroxidase (Fig. 6B) after 20-min incubation witb MEKP show that at all concentrations used, MEKP was similar to a mixed inhibitor of each enzyme.
Many organic hydroperoxides that are used in various industrial processes are potent in- ducers of lipid per-oxidation (Pryor, 1976; Sheppard and Mageli, 1982). Lipophilic or- ganic peroxides, such as MEKP, that are taken into the body probably enter the hepatic en- doplasmic reticulum, bind to microsomal P- 450, and react with polyunsaturated fatty ac- ids. The peroxides could then destroy protein, inhibit enzyme activities, and generate lipid peroxides. MEKP damages liver (Sitting, 198 1) and induces lipid peroxidation in rats (Litov et al., 198 1; Herschberger and Tappel, 1982) to a much greater extent than does cumene hydroperoxide and t-butyl hydroperoxide (Li- tov et al., 198 1). Some lipid hydroperoxides are known to readily destroy microsomal cy- tochrome P-450 (Hruszkewycz et al., 1978; O’Brien, 1982; Jeffery et al., 1977).
The mechanism of damage to liver cyto- chrome P-450 by MEKP appears to be differ- ent from that of damage by heavy metals or detergents, some of which convert cytochrome P-450 to cytochrome P-420 (Omura and Sato, 1964b; Ichikawa and Yamano, 1967; Ando, 1982; Denk, 1982). The addition of MEKP to microsomes resulted in an absorbance change in the difference spectrum, which was revealed
A OIS- Double reciprocal plots of NADH- and NADPH-peroxidase activities at various con- centrations of MEKP are shown in Fig. 7. The V,,,, of NADH- and NADPH-peroxidase for MEKP were slightly lower (33 and 10 nmol NAD(P)H oxidized/min/mg protein, respec- tively) than the V,, values for cumene hy- droperoxide (40 and 17 nmol NAD(P)H oxi- dized/min/mg protein, respectively). The K,,, values of NADH- and NADPH-peroxidase for MEKP were very low (0.022 and 0.012 mM, 1
-SO -40 0 40 so I20 160
respectively) compared to their Km values for cumene hydroperoxide (0.88 and 0.34 mrvi,
[MEKP](mlk
respectively). MEKP was both a substrate for FIG. 7. (A) Double-reciprocal plot of liver microsomal
and an irreversible inhibitor of these two per- NADH-peroxidase and (B) NADPH-peroxidase activities with MEKP as substrate. These activities were measured
oxidases. immediately after addition of 300 PM MEKP.

DAMAGE TO CYTGCHROME P-450 ACTIVITIES 523
as a progressive increase in absorbance at 433 tective roles need to be evaluated in future re- nm. MEKP appeared to bind to cytochrome search. P-450, and progressively to destroy it. MEKP, together with any lipid peroxides that may REFERENCES have been formed by MEKP, was responsible for irreversible inhibition of NADH- and
ANDO, M. (1982). Cadmium effect on microsomal drug
NADPH-peroxidase activities as a function of metabolizing enzyme activity in rat livers with respect to differences in age and sex. Environ. Res. 27, 446-
time. Cytochrome P-450 from kidney was rel- 456. atively more resistant to destruction by MEKP BRADFORD, M. ( 1976). A rapid and sensitive method for
than that from liver. the quantitation of microgram quantities of protein uti-
MEKP was both a substrate for an irre- lizing the principle of protein-dye binding. Anal.
versible inhibitor of NADH- and NADPH- Biochem. 72,248-2X
CADENAS, E., AND SIES, H. (1982). Low level chemilu- peroxidase. It is reasonable that at 0 time, and minescence of liver microsomal fractions initiated by
at low concentration, MEKP is a competitive tert-butyl hydroperoxide. Eur. J. B&hem. 124, 349-
inhibitor of NADH- and NADPH-peroxidase. 356.
At high concentration, excess MEKP can bind COON, M. J., CHIANG, Y. L., AND FRENCH, J. (1979).
to other sites on cytochrome P-450, damage Chemical characterization of the enzymes involved in drug metabolism. In The Induction ofDrug Metabolism
the protein, and/or initiate lipid peroxidation; (R. W. Estabrook, and L. Linderlaub, eds.), pp. 201-
therefore, MEKP behaves like a mixed inhib- 2 11. F. K. Schattauer-Verlag, Stuttgart.
itor of these peroxidase activities. The sensi- DENK, H. (1982). Effect of detergents on the mixed func-
tivities of cytochrome P-450 peroxidase activ- tion oxidase and other microsomal enzymes. In Inter-
ities to MEKP were different; NADPH-per- national Encyclopedia of Pharmacology and Therapeu- tics, Section 108: Hepatic Cytochrome P-450 Monoox-
oxidase was more resistant to the damage by ygenase System (J. B. Schenkman, and D. Kupfer, eds.),
MEKP than was NADH-peroxidase. pp. 469-495, Pergamon, New York.
Hepatic microsomal monooxygenase is vital DILLARD, C. J., DUMELIN, E. E., AND TAPPEL, A. L.
for the metabolism of lipophilic organic per- ( 1977). Effect of dietary vitamin E on expiration of pen-
oxides. Cytochrome P-450 peroxidase activi- tane and ethane by the rat. Lipids 12, 109-l 14.
ESTABROOK, R. W., AND WERRINGLOER, J. (1978). The ties catalyze the reduction of organic hydro- measurement of difference spectra: application to cy-
peroxides to their inactive metabolites (Hrycay tochromes of microsomes. In Methods in Enzymology
and O’Brien, 1972, 1973, 1974). NADH- (S. Fleischer, and L. Packer, eds.), Vol. LB, pp. 212-
and NADPH-peroxidases require NADH, 220, Academic Press, New York.
NADPH, and microsomal electron-transport GUENGERICH, F. P. (1978). Separation and purification
of multiple forms of microsomal cytochrome P-450. J. components (Hrycay et al., 1975). Hepatic Biol. Chem. 253,793 1-7939.
NADH- and NADPH-peroxidase activities HAFEMAN, D. Cl., AND HOEKSTRA, W. G. (1977). Pro-
were stimulated by phenobarbital and 3- tection against carbon tetrachloride-induced lipid per-
methylcholanthrene, which are typical micro- oxidation in the rat by dietary vitamin E, selenium, and
somal inducing agents. The induction of more methionine as measured by ethane evolution. J. Nutr. 107,656-665.
hepatic NADH- and NADPH-peroxidase ac- HARADA, N., AND OMURA, T. (198 1). Selective induction
tivity by phenobarbital treatment than by 3- of two different molecular species of cytochrome
methylcholanthrene may result from differ- P-450 by phenobarbital and 3-methylcholantbrene. J.
ences in the types of cytochrome P-450 sub- B&hem. 89,237-248,
species induced by each treatment. Since HERSCHBERGER, L. A., AND TAPPEL, A. L. (1982). Effect
of vitamin E on pentane exhaled by rats treated with NADH and NADPH are important elec- methyl ethyl ketone peroxide. Lipids 17, 686-69 1.
tron donors under physiological conditions, HRUSZKE~YCZ, A. M., GLENDE, E. A., JR., AND RECK-
NADH- and NADPH-peroxidase may serve NAGEL, R. 0. (1978). Destruction of microsomal cy-
important protective roles within the endo- tochrome P-450 and glucose-6-phosphatase by lipid ex-
plasmic reticulum via metabolism of toxic or- tracted from peroxidized microsomes. Toxicol. Appl. Pharmacol. 46,695-702.
ganic peroxides, such as MEKP. These pro- HRYCAY, E. G., AND G’BRIEN, P. J. (1972). Cytochrome

524 AND0 AND TAPPEL
P-450 as a microsomal peroxidase in steroid hydroper- oxide reduction. Arch. Biochem. Biophys. 153,480-494.
HRYCAY, E. G., AND O’BRIEN, P. J. (1973). Microsomal electron transport. I. Reduced nicotinamide adenine dinucleotide phosphate-cytochrome c reductase and cytochrome P-450 as electron carriers in microsomal NADPH-peroxidase activity. Arch. Biochem. Biophys. 157,7-22.
HRYCAY, E. G., AND O’BRIEN, P. J. (1974). Microsomal electron transport. II. Reduced nicotinamide adenine dinucleotide-cytochrome b5 reductase and cytochrome P-450 as electron carriers in microsomal NADH-per- oxidase activity. Arch. B&hem. Biophys. 160,230-245.
HRYCAY, E. G., JONEN, H. G., Lu, A. Y. H., AND LEVIN, W. (1975). Reconstitution of reduced nicotinamide ad- enine dinucleotide phosphate- and reduced nicotin- amide adenine dinucleotide-peroxidase activities from solubilized components of rat liver. Arch. Biochem. Biophys. 166, 145-151.
ICHIKAWA, Y., AND YAMANO, T. (1967). Reconversion of detergent- and sullhydryl reagent-produced P-420 to P-450 by polyols and glutathione. Biochim. Biophys. Acta 131, 490-497.
IMAI, Y., AND SATO, R. (1974). A gel-electrophoretically homogeneous preparation of cytochrome P-450 from liver microsomes of phenobarbital-treated rabbits. Biochem. Biophys. Res. Commun. 60, 8-14.
JEFCOATE, C. R. (1978). Measurement of substrate and inhibitor binding to microsomal cytochrome P-450 by optical-difference spectroscopy. In Methods in Enzy- mology (S. Fleischer, and L. Packer, eds.), Vol. LB, pp. 258-279, Academic Press, New York.
JEFFERY, L. H., NERLAND, D., EL-AZBARY, R., AND
MANNERING, G. J. (1977). Destruction of cytochrome P-450 by linoleic acid hydroperoxide. In Microsomes and Drug Oxidations (V. Ullrich, I. Roots, A. Hilde- brand& R. W. Estabrook, and A. H. Conney, eds.), pp. 323-330, Pergamon, New York.
LITOV, R. E., MAI-IXEWS, L. C., AND TAPPEL, A. L. ( 198 1). Vitamin E protection against in vivo lipid per- oxidation initiated in rats by methyl ethyl ketone per- oxide as monitored by pentane. Toxicol. Appl. Phar- macol. 59, 96- 106.
National Institute for Occupational Safety and Health (1977). Information profiles on potential occupational hazards: MEKP. pp. 37-41, Report PB-276, 678, Rockville, Md.
NEBERT, D. W., AND GELBOIN, H. V. (1968). Substrate- inducible microsomal aryl hydroxylase in mammalian cell culture. J. Biol. Chem. 243,6242-6249.
NORDBLOM, G. D., WHITE, R. E., AND COON, M. J. (1976). Studies on hydroperoxidedependent substrate hydroxylation by purified liver microsomal cytochrome P-450. Arch. B&hem. Biophys. 175, 524-533.
NOSHIRO, M., AND OMURA, T. (1978). Immunochemical study on the electron pathway from NADH to cyto- chrome P-450 of liver microsomes. J. B&hem. 83,6 l- 77.
O’BRIEN, P. J., AND RAHIMTULA, A. D. (1978). A per- oxidase assay for cytochrome P-450. In Methods in En- zymology (S. Pleischer, and L. Packer, eds.), Vol. LB, pp. 407-4 12, Academic Press, New York.
O’BRIEN, P. J. (1982). Hydroperoxides and superoxides in microsomal oxidations. In International Encyclopedia of Pharmacology and Therapeutics, Section 108: Hepatic Cytochrome P-450 Monooxygenase System (J. B. Schenkman, and D. Kupfer, eds.), pp. 567-586, Per- gamon, New York.
OMURA, T., AND SATO, R. (1964a). The carbon monoxide- binding pigment of liver microsomes. I. Evidence for its hemoprotein nature. J. Biol. Chem. 239,2370-2378.
OMURA, T., AND SATO, R. (1964b). The carbon monoxide- binding pigment of liver microsomes. II. Solubiiition, purification, and properties. J. Biol. Chem. 239,2379- 2385.
ORRENIUS, S. ( 1965). On the mechanism of drug hydrox- ylation in rat liver microsomes. J. Cell Biol. 26, 713- 723.
ORRENIUS, S., ELLIN, A., JAKO~SSON, S., THOR, H., m, D. L., SCHENKMAN, J. B., AND ESTABROOK, R. W. (1973). The cytochrome P-450 containing monoxygen- ase system of rat kidney cortex microsomes. Drug Me- tab. Disp. 1, 350-357.
PRYOR, W. A. (1976). Free Radicals in Biology, Vol. 3, Academic Press, New York.
RAHIMTULA, A. D., AND O’BRIEN, P. J. (1974). Hydro- peroxide catalyzed liver microsomal aromatic hydrox- ylation reactions involving cytochrome P-450. Biochem. Biophys. Res. Commun. 60,440-447.
RAHIMTULA, A. D., AND O’BRIEN, P. J. (1977). The role of cytochrome P-450 in the hydroperoxide-catalyzed oxidation of alcohols by rat-liver microsomes. Eur. J. Biochem. 77,20 l-208.
RYAN, D. E., THOMAS, P. E., KOREZENIOWSKI, D., AND LEVIN, W. (1979). Separation and characterization of highly purified forms of liver microsomal cytochrome P-450 from rats treated with polychlorinated biphenyls, phenobarbital, and 3-methylcholanthrene. J. Biol. Chem. 254, 1365- 1374.
SHEPPARD, C. S., AND MAGELI, 0. L. (1982). Peroxides and peroxy compounds, organic. In Encyclopedia of Chemical Technology (K. Othmer, ed.), Vol. 17, pp. 27-89, Wiley-Interscience, New York.
SITTING, M. ( 198 1). Handbook of Toxic and Hazardous Chemicals. Noyes Publications, Park Ridge, N.J.
SPI BULLETIN ( 1982). Commercial Organic Peroxide Toxicological Data. The Society of the Plastics Industry, Inc., New York.
U.S. Environmental Protection Agency (1979). Chemical Hazard Information Profile: MEKP, Washington, D.C.
WERRINGLOER, J. (1978). Assay of formaldehyde gener- ated during microsomal oxidation reactions. In Methori in Enzymology (S. Pleischer, and L. Packer, eds.), Vol. LB, pp. 297-302, Academic Press, New York.





























