Embed Size (px)
Citation preview

[CANCER RESEARCH 46, 1617-1622, April 1986]
Methods of Immunosuppression for Study of Growth and Lung Colony Formationby Human Tumor Cells in Mice1
Asya Kovnat, Kong Khoo, Peter Selby, and Ian Tannock2
Departments of Medicine and Medical Biophysics, Ontario Cancer Institute and University of Toronto, 500 Sherbourne Street, Toronto, Ontario, Canada M4X1K9
ABSTRACT
Mice that are immune-suppressed by thymectomy and by sequentialtreatment with 1-0-D-arabinofuranosylcytosine and whole body irradiation may be used as hosts for generation of human tumor xenografts. Wehave studied the effect of various additional methods of immune suppression on the formation of tumors after i.m. injection and on the formationof lung colonies after i.v. injection with the human MGH-U1 bladdercancer cell line. Success of transplantation was improved by treatment ofimmune-suppressed animals with either heterologous antilymphocyteserum or a monoclonal anti-Thy-1.2 antibody. Success of lung colonyformation was also improved by antilymphocyte serum but not by monoclonal anti-Thy-1.2 antibody. Admixture of heavily irradiated cells (10')
to the viable inoculum of tumor cells in addition to antilymphocyte serumtreatment improved the success of i.m. transplantation but not that oflung colony formation. Treatment with corticosteroids or treatment withcarrageenan to suppress macrophage activity added toxicity and did notimprove the success of xenografting. Immune suppression decreased thenatural killer cell activity of normal mice and treatment with antiinter-feron to further suppress natural killer cells may also enhance xenograftformation. Administration of cyclosporin A to normal mice allowed thegrowth of a single xenograft but was not a useful method for immuno-suppression. The success of xenografting into immune-deprived mice wassuperior to that for two strains of nude mice maintained in our laboratory,and i.v. injection of tumor cells did not lead to lung colonies in these nudemice. Immune-deprived mice are a useful alternative to nude mice for thestudy of xenografts derived from human tumor cell lines and may allowthe study of experimental lung métastases.
INTRODUCTION
Xenogenic transplantation of human tumors into rodents,and particularly into congenitally athyrnic "nude" mice, is a
widely used technique in experimental cancer research.Mice that are immune suppressed by thymectomy and by
sequential treatment with ara-C3 and WBI may be used as
alternative hosts to nude mice for generation of human tumorxenografts (1). Pretreatment with ara-C or with some otherchemotherapeutic agents prevents lethal effects of WBI forreasons that are poorly understood (2). Immune-deprived micehave the advantages of being sturdy, readily available, andinexpensive compared to nude mice. In addition, immune-deprived mice may allow a low incidence of spontaneous andexperimental métastasesfrom some human tumor cell lines (1,3), whereas métastasesoccur rarely in adult nude mice, althoughthey may occur in newborn nude mice (4).
Unfortunately, both nude and immune-deprived mice areimperfect hosts for transplantation of human tumors. Severaltypes of host defense mechanisms may limit the success of
Received 8/9/85; revised 12/6/85; accepted 12/27/85.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by a National Cancer Institute of Canada Fellowship to A.K. andby N1H Grant CA 29526 awarded through the National Bladder Cancer Project.
2To whom requests for reprints should be addressed.3The abbreviations used are: ara-C, l-|8-i>arabinofuranosylcytosine; WBI,
whole body irradiation; TAX (mice), mice treated by thymectomy, 1-ß-D-arabi-nofuranosylcytosine, and whole body irradiation; ALS, antilymphocyte serum;MATS, monoclonal antithymocyte serum; anti-asGMI, antiasialo GM1 serum;Cy-A, cyclosporin A; NK, natural killen PCS, fetal calf serum; TD-50, transplantation dose of 50%, i.e., the number of cells required to produce progressivelygrowing xenografts in 50% of mice; HR, heavily radiated.
transplantation. The possibility of immunological recovery inimmune-deprived mice may be a potential disadvantage in long-term experiments (1). The mechanisms which limit the successof xenografting in immune-deprived mice are complex butinclude the following factors: (a) residual and recovering T-cellfunction. Recovery of T-cell immunity in immune-deprivedmice (5) has sometimes led to the spontaneous regression ofxenografts (1). Residual T-cell immunity might be suppressedby ALS (6-9) or by administration of CY-A, a new immuno-suppressive agent which has a relatively specific action directedagainst T-lymphocytes (10, 11); (b) macrophages. Macrophagefunction has been implicated in resistance to xenografts (12)and to métastasesin syngeneic mice (13). Carageenans depressmacrophage function and have been reported to increase thesuccess of xenografting (14); (c) natural killer cells. NK cellshave been implicated in host defenses against transplantabletumors and in prevention of métastasesin nude mice (15-17).Agents that inhibit NK cell activity such as sheep antiinterferon,estrogens, or anti-asGMI have been reported to increase theincidence of xenogeneic tumors and métastases(18-20); and(d) local factors. The admixture of large numbers of heavilyradiated nonviable cells or even of inert plastic microsphereshas been found to increase the success of local transplantationand formation of lung colonies after i.v. injection of viable cellsin syngeneic mice (21, 22).
Optimization of xenografting requires recognition of theabove factors and the development of methods for overcominghost defenses against the xenografted tumor. In the presentstudy we have assessed the influence of several modificationsto immune-deprived mice on the success of transplantation ofa human tumor cell line. We have studied both local tumorgrowth after i.m. implantation and formation of lung coloniesafter i.v. injection of human tumor cells.
MATERIALS AND METHODS
Mice. Male CBA/CaJ mice (Jackson Laboratories) underwent thymectomy at 4-5 wk of age. Three to 6 wk later they received an i.p.injection of ara-C (200 mg/kg) followed 48 h later by 8.5 Gy WBI asdescribed previously (23). These animals will be referred to as TAXmice. In some experiments mice received cyclophosphamide instead ofara-C. A small number of NK cell deficient beige mice (C57BL/6J Bg/Bg) were obtained from the National Cancer Institute, and were immunedeprived as described above. Athymic RNC nu/nu and C57BL/6 nulnu mice were bred in the animal colony at Ontario Cancer Institute.The nude mice were maintained in shoe box cages in laminar air-flowrooms and received sterilized food.
Cell Lines. The established human bladder cancer cell lines used inthese experiments were kindly supplied by Dr. G. Prout and his colleagues (Urology Research Laboratories, Massachusetts General Hospital). Most experiments were performed with the MGH-U1 cell linewhich is of common origin with cell lines designated 1.1 and T24 (24,25). Characteristics of the cell line have been confirmed by cytogeneticexamination and isoenzyme analysis performed by the National CancerInstitute culture identification service (Dr. W. D. Peterson Jr., Children's Hospital of Michigan, Detroit). A second cell line that we havereferred to previously as RT-4 (23,26) has been identified by isoenzymeand karyotypic analysis as a tetraploid subline of MGH-U I. To avoidconfusion this cell line will be referred to as MGH-U 1/TP. The lines
1617on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

XENOGRAFTS IN IMMUNE-DEPRIVED MICE
were found to be free of mycoplasma and electron microscopic examination has failed to reveal the presence of viruses.
The cell lines were maintained as monolayers in plastic flasks in .. -
medium supplemented with antibiotics and 10% PCS (27); they werereestablished from frozen stock at about 3-mo intervals.
Tumor Implantation and Growth. Cells were harvested with 0.05%trypsin and 0.02% EDTA. Cells were counted in a hemocytometer, andviability assessed by trypan blue exclusion was always greater than 90%.Appropriate dilutions of cells were prepared in media with 10% FCSand, if required, HR cells of the same cell type (100 Gy by l37Cs-y-rays)were mixed with viable cells to provide Id1' HR cells in the 0.1 -ml
inoculum volume. Cells were injected into the left thigh muscle of micefor generation of tumors.
The diameter of tumors growing in the leg after i.m. implantationwas assessed as described previously (23). Any animals which died priorto 3 mo after transplantation without tumors (or with tumors less than9 mm) were removed from analysis. In most experiments, mortality ofTAX mice was less than 10%, and it was found to be independent ofthe size of the tumor cell inoculum. A few methods that causedincreased early mortality in mice will be discussed later.
In several experiments graded numbers of tumor cells were injectedin groups of mice to explore the relationship between success oftransplantation and size of the inoculum. We attempted to analyzethese experiments and to calculate the value of 11)5(1 and its SE byfitting the data to a modified Poisson model. A maximum likelihoodmethod was applied to try to obtain the best fit of the expected curveto the data (28).
Experimental Metastasis Assay. Suspensions of single cells wereprepared from cell cultures as described above. In many experiments10* HR cells and/or IO6 plastic microspheres (mean diameter 15 ±5
(SE) ittn; 3M Company, St. Paul, MM) were added to provide aninoculum volume of 0.2 ml/mouse. The inoculum was agitated prior toinjection to minimize clumping and was injected into one of the lateraltail veins of the mice. Cells were injected 24 h after WBI was given toCBA mice as part of the schedule of immune suppression, since severalinvestigators have demonstrated increased lung colony formation whentumor cells have been injected into syngeneic mice at 24-48 h after
thoracic irradiation (29, 30).Four to 6 wk after injection the mice were killed, and lungs were
removed, washed, and fixed in Bouin's solution and 24 h later trans
ferred into 95% ethanol. Tumor nodules were counted with the aid ofa dissecting microscope.
Modifications of Immune Suppression
Antilymphocyte Serum. Heterologous rabbit anti-mouse ALS (Microbiological Associates, Bethesda, MD) was diluted 1:1 with sterile0.9% saline solution and stored at -20*C. Mice given ALS received
their initial injections of 0.2 ml on the day of tumor transplantationand then twice weekly for 4-6 wk.
Monoclonal Antithymocyte Serum. The lini antigen is common tol cd K of all murine strains, including CBA mice, and also appears on
some NK cells (31, 32). A hybridoma which secretes complement-dependent cytotoxic anti-Thy-1.2 antibodies was developed at the Ontario Cancer Institute by Dr. R. A. Phillips and colleagues, usingimmunization procedures similar to those described by Lake and Douglas (33). The hybridoma, designated P3/NSl/l-Ag4-l was maintainedin ..medium plus 10% FCS with periodic reestablishment from frozenstock. MATS used in current experiments was prepared from ascitesin BALB/c mice. These mice received two i.p. injections of pristane (2,6,10,14-tetramethylpentadecane; Aldrich Chemical Co., Milwaukee,WI) I wk apart and I wk later were treated with ara-C (200 mg/kg i.p.)followed after 48 h by 4 Gy WBI. The hybridoma (2 x 10* cells) wasinjected i.p. 1 wk later and produced ascites in 7-10 days. The micewere killed and the ascites were aspirated and centrifuged at 3000 rpmfor 20 min. The supernatant (MATS) was stored at -20°C after
measurement of the antibody titer as described below.In most experiments, immune-deprived mice that received MATS
were given injections i.p. with 0.2 ml twice weekly for 4-6 wk, startingon the day of tumor implantation.
Carrageenan. i-Carrageenan (Type V; Sigma Chemical Co., St. Louis,
MO) was injected into some animals in an attempt to suppress macrophage function. Carrageenan was dissolved in physiological saline at80"C, and 5 mg in 0.2 ml were injected i.p. into mice on the day prior
to tumor transplantation (14).Corticosteroids. Some mice received methylprednisolone (50 mg/kg)
injected i.p. twice weekly for 6 wk.Cyclosporin-A. Cy-A, a cyclic endecapeptide of fungal origin (34)
was kindly supplied by Drs. J. F. Borei and E. Wiskott (Sandoz Ltd.,Basel, Switzerland). It was dissolved in ethanol and Tween 80 plussaline for i.p. injections. For p.o. administration the drug was given ina solution of olive oil. Daily doses ranged from 20-100 mg/kg.
Estrogen. Some mice received 17/3-estradiol. The hormone was implanted s.c. in silastic tubing (Dow-Corning, Midland, MI) to allowslow release of the hormone as described by others (35). Briefly, a 2-cm piece of silastic tubing containing 5-15 mg of 17/3-estradiol wasimplanted s.c. into the flanks of 4- to 5-wk-old thymectomized CBA/CaJ mice using anesthesia (tribromoethanol, 300 mg/kg). Mice receivedfurther immunosuppression with ara-C and WBI 3 wk later.
Antiinterferon. Sheep antiserum to mouse L-cell interferon (NIH,Bethesda, MD) was dissolved in saline and stored at —20'C.One ml ofthe resulting solution has a neutralizing titer of 3 x 10" against 8 IU
interferon. Mice receiving antiinterferon were given injections i.v. of0.1 ml of a 1:3 dilution in saline on the day of xenograft implantationand weekly for the next 3 wk.
Antiasialo GM1 Serum. Lyophilized rabbit anti-asGMl, (WakoChemicals, USA Inc., Dallas, TX), was reconstituted with distilledwater, diluted 1:15 with physiological saline, and kept at —20°C.Mice
were treated i.v. with 0.2 ml of antiserum on the day of tumor transplantation.
Assessment of Immunological Activity
Determination of Antithymocyte Titer. A microcytotoxicity test wasused to measure the titer of ALS, MATS, or of blood from animalsthat had received these antisera. Repeated samples of blood (50 uI)were drawn from tail veins without killing the mice; these samples werecentrifuged and the pooled sera from identically treated animals werekept frozen. For determination of titer the serum was diluted withCedarlane cytotoxicity media (RPMI 1640 with 25 HIM4-(2-hydroxy-ethyl)-l-piperazineethanesulfonic acid buffer and 0.3% bovine serumalbumin; Cedarlane, Hornby, Ontario, Canada). A suspension of freshthymocytes was prepared by mechanical dissociation of thymuses fromCBA/CaJ mice, washed twice, and diluted to 3 x Id" cells/ml. Aliquots
of 25 n\ were added to individual wells of a round bottomed 96-wellflexible titration plate (Fisher Scientific, Toronto, Ontario, Canada)and serial dilutions of sera were added and mixed. Positive (i.e., knowntiter) and negative control sera were included in all experiments. Aftera 45 min incubation at 4 ( . the plates were centrifuged to pellet thecells, and the supernatant was discarded. Fifty /il guinea pig complement(Cedarlane) were added to each well and the plates were incubated fora further 60 min at 37*C. Viable and damaged cells were scored in a
hemocytometer using an inverted phase-contrast microscope withaqueous eosin used as an indicator of cell damage. The cytotoxicantithymocyte titer was determined as that dilution of serum whichkilled 50% of the thymocytes.
Determination of NK Cell Activity. The NK cell activity in immune-deprived mice was assayed by a direct 6-h s'Cr release assay using NKsensitive YAC-1 and NK-resistant P815 cells as targets (36, 37). Asingle cell suspension of spleen cells from identically treated mice wasprepared, and 0.1-ml vol of different cell dilutions were placed in V-bottomed microtiter trays (Linbro Flowlabs, Mississauga, Ontario,Canada). Target cells were labeled by incubation in 0.2 ml of a 10 mCi/ml solution of s'N.i.-( 'rOj (New England Nuclear, Boston, MA),
washed, and added to the wells in 0.1-ml vol to give effectortarget cellratios of 100:1, 50:1, or 25:1. After 6 h incubation at 37'C, 0.1 ml ofsupernatant was withdrawn from each well, and s'Cr release (R) wasmeasured using a gamma counter. Spontaneous release (S) of s'Cr was
measured from target cells incubated alone, and the total amount ofs'Cr that could be released ('/")was estimated from target cells incubated
in 1% acetic acid. NK cell activity was then expressed as the specific
1618on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
XENOGRAFTS IN IMMUNE-DEPRIVED MICE
lysis given by
RESULTS
Specific lysis =R-ST-S'
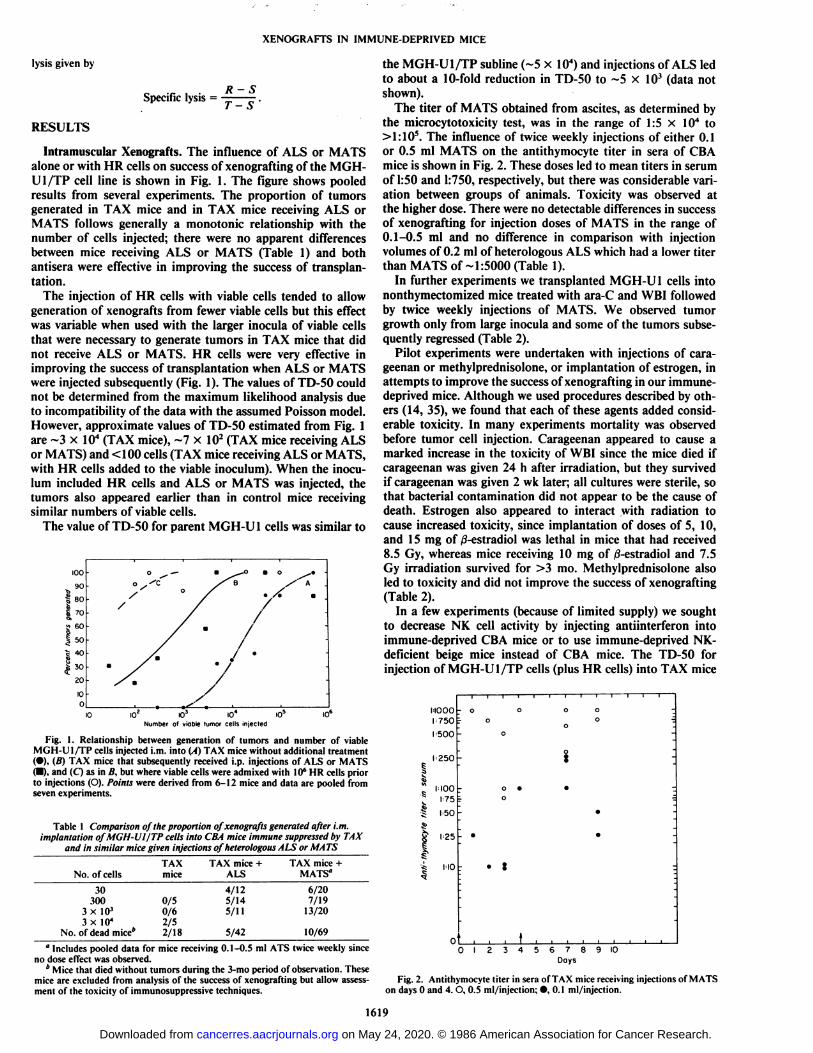
Intramuscular Xenografts. The influence of ALS or MATSalone or with HR cells on success of xenografting of the MGH-Ul/TP cell line is shown in Fig. 1. The figure shows pooledresults from several experiments. The proportion of tumorsgenerated in TAX mice and in TAX mice receiving ALS orMATS follows generally a monotonie relationship with thenumber of cells injected; there were no apparent differencesbetween mice receiving ALS or MATS (Table 1) and bothantisera were effective in improving the success of transplantation.
The injection of HR cells with viable cells tended to allowgeneration of xenografts from fewer viable cells but this effectwas variable when used with the larger inocula of viable cellsthat were necessary to generate tumors in TAX mice that didnot receive ALS or MATS. HR cells were very effective inimproving the success of transplantation when ALS or MATSwere injected subsequently (Fig. 1). The values of TD-50 couldnot be determined from the maximum likelihood analysis dueto incompatibility of the data with the assumed Poisson model.However, approximate values of TD-50 estimated from Fig. 1are ~3 x IO4(TAX mice), ~7 X IO2(TAX mice receiving ALS
or MATS) and <100 cells (TAX mice receiving ALS or MATS,with HR cells added to the viable inoculum). When the inoculum included HR cells and ALS or MATS was injected, thetumors also appeared earlier than in control mice receivingsimilar numbers of viable cells.
The value of TD-50 for parent MGH-U1 cells was similar to
10° 10"
Number of viable tumor cells injected
10s
Fig. I. Relationship between generation of tumors and number of viableMGH-U I/TP cells injected i.m. into (A) TAX mice without additional treatment(•),(B) TAX mice that subsequently received i.p. injections of ALS or MATS(•),and (O as in li. but where viable cells were admixed with 10*HR cells priorto injections (O). Points were derived from 6-12 mice and data are pooled fromseven experiments.
Table 1 Comparison of the proportion of xenografts generated after i.m.implantation ofMGH-Ul/TP cells into CBA mice immune suppressed by TAX
and in similar mice given injections of heterologous ALS or MATS
No. ofcells30
3003x IO33x IO4
No. of dead mice*TAX
mice0/5
0/62/52/18TAX
mice+ALS4/12
5/145/115/42TAX
mice +MATS"6/20
7/1913/2010/69
°Includes pooled data for mice receiving 0.1-0.5 ml ATS twice weekly since
no dose effect was observed.* Mice that died without tumors during the 3-mo period of observation. These
mice are excluded from analysis of the success of xenografting but allow assessment of the toxicity of immunosuppressive techniques.
the MGH-U 1/TP subline (~5 x 10") and injections of ALS ledto about a 10-fold reduction in TD-50 to ~5 X IO3 (data not
shown).The titer of MATS obtained from ascites, as determined by
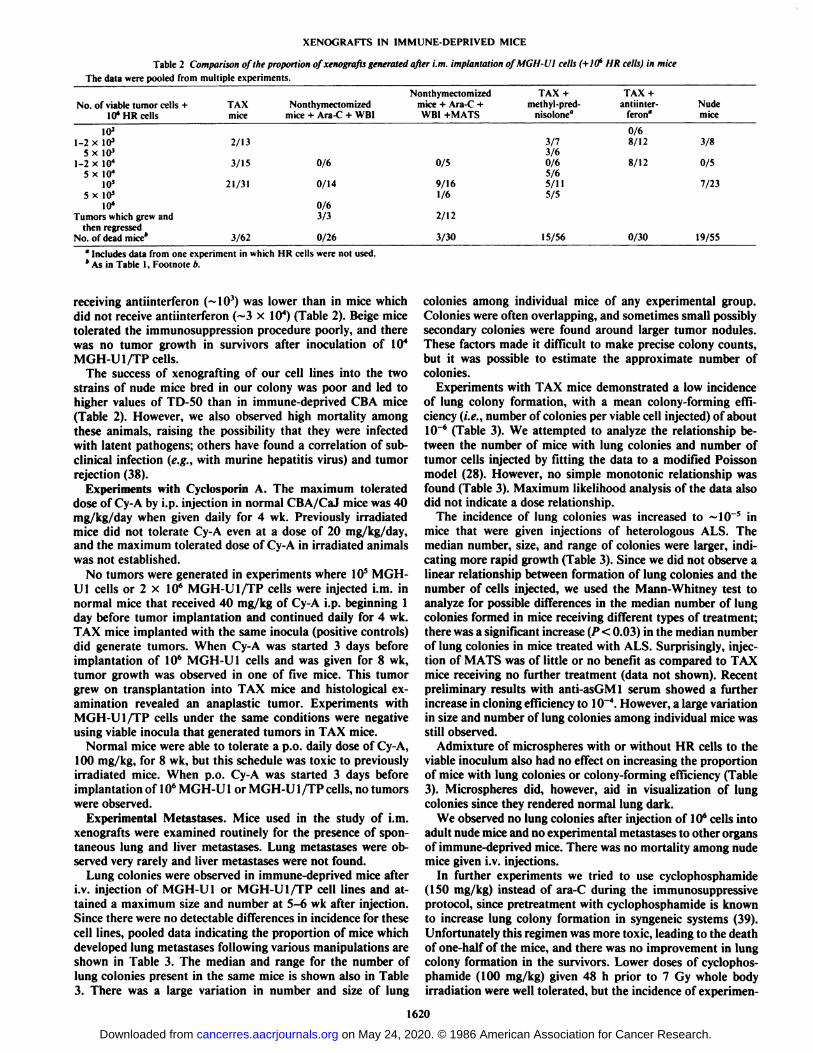
the microcytotoxicity test, was in the range of 1:5 x IO4 to>1:105. The influence of twice weekly injections of either 0.1
or 0.5 ml MATS on the antithymocyte titer in sera of CBAmice is shown in Fig. 2. These doses led to mean titers in serumof 1:50 and 1:750, respectively, but there was considerable variation between groups of animals. Toxicity was observed atthe higher dose. There were no detectable differences in successof xenografting for injection doses of MATS in the range of0.1-0.5 ml and no difference in comparison with injectionvolumes of 0.2 ml of heterologous ALS which had a lower titerthan MATS of -1:5000 (Table 1).
In further experiments we transplanted MGH-U 1 cells intononthymectomized mice treated with ara-C and WBI followedby twice weekly injections of MATS. We observed tumorgrowth only from large inocula and some of the tumors subsequently regressed (Table 2).
Pilot experiments were undertaken with injections of cara-geenan or methylprednisolone, or implantation of estrogen, inattempts to improve the success of xenografting in our immune-deprived mice. Although we used procedures described by others (14, 35), we found that each of these agents added considerable toxicity. In many experiments mortality was observedbefore tumor cell injection. Carageenan appeared to cause amarked increase in the toxicity of WBI since the mice died ifcarageenan was given 24 h after irradiation, but they survivedif carageenan was given 2 wk later; all cultures were sterile, sothat bacterial contamination did not appear to be the cause ofdeath. Estrogen also appeared to interact with radiation tocause increased toxicity, since implantation of doses of 5, 10,and 15 mg of /3-estradiol was lethal in mice that had received8.5 Gy, whereas mice receiving 10 mg of ß-estradioland 7.5Gy irradiation survived for >3 mo. Methylprednisolone alsoled to toxicity and did not improve the success of xenografting(Table 2).
In a few experiments (because of limited supply) we soughtto decrease NK cell activity by injecting antiinterferon intoimmune-deprived CBA mice or to use immune-deprived NK-deficient beige mice instead of CBA mice. The TD-50 forinjection of MGH-U 1/TP cells (plus HR cells) into TAX mice
NOOOr 750I'500
I'250
1 MOO
175
h50
1-25
NO • t
0123456789 IODays
Fig. 2. Antithymocyte titer in sera of TAX mice receiving injections of MATSon days 0 and 4. O, 0.5 ml/injection; •.0.1 ml/injection.
1619
on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
XENOGRAFTS IN IMMUNE-DEPRIVED MICE
Table 2 Comparison of the proportion ofxenografts generated after i.m. implantation ofMGH-UI cells (+l(f HR cells) in mice
The data were pooled from multiple experiments.
No. of viable tumor cells -10* HRcellsIO21-2
xIO35xIO11-2
xIO4SxIO4IO55x
IO5IO4Tumors
which grewandthenregressedNo.
of dead mice*TAX
mice2/133/1521/313/62Nonthymectomizedmice + Ara-C +WBI0/60/140/63/30/26Nonthymectomizedmice + Ara-C +WBI+MATS0/59/161/62/123/30TAX
+methyl-pred-nisolone"3/73/60/65/65/115/515/56TAX
+antiinter
feron*0/68/128/120/30Nudemice3/80/57/2319/55
' Includes data from one experiment in which HR cells were not used.* As in Table 1, Footnote b.
receiving antiinterferon (~103) was lower than in mice whichdid not receive antiinterferon (~3 x 10") (Table 2). Beige mice
tolerated the immunosuppression procedure poorly, and therewas no tumor growth in survivors after inoculation of 10'
MGH-U1/TP cells.The success of xenografting of our cell lines into the two
strains of nude mice bred in our colony was poor and led tohigher values of TD-50 than in immune-deprived CBA mice(Table 2). However, we also observed high mortality amongthese animals, raising the possibility that they were infectedwith latent pathogens; others have found a correlation of sub-clinical infection (e.g., with murine hepatitis virus) and tumorrejection (38).
Experiments with Cyclosporin A. The maximum tolerateddose of Cy-A by i.p. injection in normal CBA/CaJ mice was 40mg/kg/day when given daily for 4 wk. Previously irradiatedmice did not tolerate Cy-A even at a dose of 20 mg/kg/day,and the maximum tolerated dose of Cy-A in irradiated animalswas not established.
No tumors were generated in experiments where IO5 MGH-Ul cells or 2 x IO6 MGH-U1/TP cells were injected i.m. innormal mice that received 40 mg/kg of Cy-A i.p. beginning 1day before tumor implantation and continued daily for 4 wk.TAX mice implanted with the same inocula (positive controls)did generate tumors. When Cy-A was started 3 days beforeimplantation of IO6 MGH-U1 cells and was given for 8 wk,
tumor growth was observed in one of five mice. This tumorgrew on transplantation into TAX mice and histológica! examination revealed an anaplastic tumor. Experiments withMGH-U1/TP cells under the same conditions were negativeusing viable inocula that generated tumors in TAX mice.
Normal mice were able to tolerate a p.o. daily dose of Cy-A,100 mg/kg, for 8 wk, but this schedule was toxic to previouslyirradiated mice. When p.o. Cy-A was started 3 days beforeimplantation of 106MGH-U1 or MGH-U1 /TP cells, no tumors
were observed.Experimental Métastases. Mice used in the study of i.m.
xenografts were examined routinely for the presence of spontaneous lung and liver métastases.Lung métastaseswere observed very rarely and liver métastaseswere not found.
Lung colonies were observed in immune-deprived mice afteri.v. injection of MGH-U 1 or MGH-U 1/TP cell lines and attained a maximum size and number at 5-6 wk after injection.Since there were no detectable differences in incidence for thesecell lines, pooled data indicating the proportion of mice whichdeveloped lung métastasesfollowing various manipulations areshown in Table 3. The median and range for the number oflung colonies present in the same mice is shown also in Table3. There was a large variation in number and size of lung
colonies among individual mice of any experimental group.Colonies were often overlapping, and sometimes small possiblysecondary colonies were found around larger tumor nodules.These factors made it difficult to make precise colony counts,but it was possible to estimate the approximate number ofcolonies.
Experiments with TAX mice demonstrated a low incidenceof lung colony formation, with a mean colony-forming efficiency (i.e., number of colonies per viable cell injected) of about10" (Table 3). We attempted to analyze the relationship be
tween the number of mice with lung colonies and number oftumor cells injected by fitting the data to a modified Poissonmodel (28). However, no simple monotonie relationship wasfound (Table 3). Maximum likelihood analysis of the data alsodid not indicate a dose relationship.
The incidence of lung colonies was increased to ~10~5 in
mice that were given injections of heterologous ALS. Themedian number, size, and range of colonies were larger, indicating more rapid growth (Table 3). Since we did not observe alinear relationship between formation of lung colonies and thenumber of cells injected, we used the Mann-Whitney test toanalyze for possible differences in the median number of lungcolonies formed in mice receiving different types of treatment;there was a significant increase (P< 0.03) in the median numberof lung colonies in mice treated with ALS. Surprisingly, injection of MATS was of little or no benefit as compared to TAXmice receiving no further treatment (data not shown). Recentpreliminary results with anti-asGMl serum showed a furtherincrease in cloning efficiency to 1o ', However, a large variation
in size and number of lung colonies among individual mice wasstill observed.
Admixture of microspheres with or without HR cells to theviable inoculum also had no effect on increasing the proportionof mice with lung colonies or colony-forming efficiency (Table3). Microspheres did, however, aid in visualization of lungcolonies since they rendered normal lung dark.
We observed no lung colonies after injection of 10' cells into
adult nude mice and no experimental métastasesto other organsof immune-deprived mice. There was no mortality among nudemice given i.v. injections.
In further experiments we tried to use cyclophosphamide(ISO mg/kg) instead of ara-C during the immunosuppressiveprotocol, since pretreatment with cyclophosphamide is knownto increase lung colony formation in syngeneic systems (39).Unfortunately this regimen was more toxic, leading to the deathof one-half of the mice, and there was no improvement in lungcolony formation in the survivors. Lower doses of cyclophosphamide (100 mg/kg) given 48 h prior to 7 Gy whole bodyirradiation were well tolerated, but the incidence of experimen-
1620
on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

XENOGRAFTS IN IMMUNE-DEPRIVED MICE
Table 3 Lung colony formation after i.v. injection ofMGH-Ul or MGH-VÌ/TPceli lines in TAX miceMice received inocula or viable cells alone or admixed with HR cells and microspheres." Some groups of mice received twice weekly i.p. injections of ALS.
TAX mice TAX mice (HR + microspheres) TAX mice (ALS) TAX mice(HR + microspheres/ALS)
No. of tumorcells3-5
x Iff11-3 x 10*5-6 x 10'
0.9-1 x 10«2-3 x 10«
10'%
(proportion)of mice with
colonies57
(4/7)77 (7/9)40 (2/5)
100(4/4)Median
(range)no. ofcolonies5
(5-13)4(1-10)2(1-3)4.5 (2-29)%
(proportion)of mice with
colonies33
(3/9)31 (4/13)25 (1/4)50(7/14)Median
(range)no. ofcolonies1.5(1-9)
3(1-11)
KD3 (1-34)%
(proportion)of mice withcolonies71
(12/17)83(10/12)60 (27/45)77 (7/9)Median
no.ofcolonies8
(1/66)120(3->200)32(1->200)42(3-151)%
(proportion)of mice withcolonies77(10/13)
62 (18/29)86(21/24)68 (39/57)94(16/17)Median
(range)no. ofcolonies4(1-71)
18(2-180)63(1->200)30(1->200)60(1->200)
'' In some experiments we compared lung colony formation for viable cells admixed with microspheres alone or for microspheres with HR cells. The results were
similar and for simplicity these results have been analyzed together.
tal lung métastasesin such mice was low (data not shown).Natural Killer Cell Activity. Estimates of NK cell activity of
spleen cells from different mice are summarized in Table 4.The level of NK cell activity of spleen cells from normal CBAmice was similar to that in spleen cells from RNC nu/nu miceand this activity decreased with the age of the mice after a peakat 5-8 wk (data not shown). The NK cell activity of spleen cellsfrom immune-deprived TAX mice was lower than in normalmice but additional treatment with heterologous ALS did notlower NK cell activity further. Treatment of normal CBA micewith ALS alone also had no effect on NK cell activity.
DISCUSSION
Our results demonstrate that conventional CBA mice, whichhave been immune deprived by thymectomy, followed by treatment with ara-C and WBI may allow the growth of xenograftsfrom a human bladder cancer cell line. These results are inagreement with the work of Steel et al.(l) and Floersheim (40).We have shown that the success of transplantation is improvedby treating mice with either heterologous ALS or MATS aloneor by combining these treatments with an admixture of HRcells to viable cells. We have found also that treatment withantiinterferon may improve the success of xenografting, but wecould not investigate this further because of a limited supply ofthis agent. TAX mice demonstrate lung colonies after i.v.injection of the human tumor cell line, as previously reportedby Thomas (3) for two human melanoma cell lines that weremaintained by transplantation as xenografts. The efficiency oflung colony formation was improved by giving animals injections of heterologous ALS but not by injecting MATS.
The antithymocyte titer of MATS, as measured in a cytotox-icity test using fresh thymocytes as targets, was much higher
Table 4 Summary ofNK cell activity assessed for spleen cells from 7.5-9-wk-otdRNC nu/nu mice, normal CBA/CaJ mice, 6-9 wk old with or without injections
of ALS, and TAX mice with or without ALSData represent the percentage of specific cytotoxicity for YAC-1 cells after 6
h incubation at an effectortarget cell ratio of 100:1. Mean ±SE is for pooledgroups of 3-4 spleens.
CBAmiceExperiment
Nudemice12345'6789I8.0±32.0
±11.0±21.0
±1.01.51.81.2Nontreated22.0
±1.910.0±1.120.0±1.826.0±2.08±
1.524±2.114
+0.94±0.85±
1.0ALS9±2
3511TAX±0.04±0.6±0.2±
1.1TAX
+ALS6±
1.57±0.52
±0.3
" Effectorto target cell ratio = 30:1 in this experiment.
than that of heterologous ALS. The greater efficiency of heterologous ALS in permitting experimental lung métastaseswastherefore surprising. Heterologous ALS has a broad immuno-suppressive effect and reacts with various types of lymphocytesas well as RBC, platelets, and macrophages (7, 41); however, itdid not lower NK cell activity in our mice. MATS is morespecific for I -cells, although it may also suppress NK cellssince about 50% of murine NK cells express the Thy-1 antigen(42). The similar efficiency of the two antisera in facilitatingi.m. xenografts suggests that the dominant host defenses againstlocal implantation may be different from those which are responsible for suppressing lung colony formation. It is alsopossible that the schedule or route of administration of theMATS was not optimal.
Several studies have implicated NK cells in the destructionof circulating tumor cells, thus inhibiting lung colony formationin both syngeneic and nude mice (16). CBA/CaJ mice used incurrent experiments had high levels of NK cell activity, similarto those recorded in nude mice, but the method used forimmunosuppression was effective in decreasing this activity.The difference in NK cell activity may be a factor in allowingimmune-suppressed mice but not nude mice to develop lungcolonies after i.v. injection of tumor cells. The further benefitof injecting heterologous ALS in permitting lung métastasesinimmune-deprived mice is probably due to activity against othertypes of lymphoid cells, since it did not further depress NK cellactivity (Table 4). Preliminary results with anti-asGM 1 serumsuggest that it may be superior to heterologous ALS treatment.Its effect is probably due to suppression of residual NK activityin immune-deprived mice.
Doses of carrageenan, 0-estradiol, and glucocorticoids recommended by others for further immunosuppression of micewere found to be toxic to immune-deprived CBA mice. It isprobable that each of these agents interacts with WBI to increase its toxicity, since carrageenan was tolerated 2 wk afterirradiation, while lower doses of /3-estradiol and irradiationcould be combined. The use of ara-C 48 h prior to WBI protectsthe mouse from radiation toxicity through mechanisms that arepoorly understood. Other anticancer drugs have been reportedto have a similar radiation-sparing effect (2), but in the presentexperiments the combination of cyclophosphamide and irradiation showed increased toxicity compared to the use of ara-C.Both cyclophosphamide and radiation are known to exert lungdamage, and each can increase lung colony formation in syngeneic mice. However, the increased toxicity of these agentswhen used together prevented this from being a useful approach.
It was possible on one occasion to observe the growth of ahuman tumor xenograft using high doses of Cy-A alone inotherwise normal mice; this result attests to the potent iminimesuppressive activity of the drug. It appears difficult to combineCy-A with other methods of immune suppression since Cy-A
1621on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

XENOGRAFTS IN IMMUNE-DEPRIVED MICE
was found to be toxic to previously irradiated mice. Others havereported that maximum tolerated dosages of Cy-A were required in order to prolong the survival of xenografts (40, 43).
We attempted to develop and improve lung colony formationby xenogeneic human tumor cells for two reasons: (a) to providea possible alternative to in vitro clonogenic assays for studyingeffects of treatment; and (/>) to provide a model for study ofsome aspects of the metastatic phenotype of human malignantcells. Unfortunately, the variation in lung colony formationamong individual mice (Table 3) was too great to allow use ofxenogeneic lung colonies from this cell line as a method forassessing cell survival after treatment. Additional treatment ofTAX mice with ALS improved the metastatic efficiency but didnot decrease the variability in colony formation (Table 3). Wewere not able to improve the magnitude and uniformity ofcolony formation by admixing microspheres with or withoutHR cells, although this has been effective in syngeneic mice(22). Our results are consistent with those published by Thomas(3) who found a possible negative effect of adding HR cellswhen each of two human melanoma cell lines were injectedwith microspheres and HR cells into immune-deprived mice.
The present results confirm that immune-suppressed CBA/CaJ mice may provide an alternative to nude mice for the studyof human tumor xenografts, and especially for the study ofexperimental metastasis. These mice are relatively inexpensive,quite sturdy, and may allow the study of biological and therapeutic properties of primary human tumors as well as cell lines(44,45). The success of xenografting may be improved by addedtreatment with ALS.
REFERENCES
1. Steel, G. G., Courtenay, V. D., and Rostom, A. Y. Improved immune-suppression techniques for the xenografting of human tumours. Br. J. Cancer,37:224-230, 1978.
2. Millar, J. t., Blackett, N. \l . and Hudspith, B. N. Enhanced post-irradiationrecovery of the haemopoietic system in animals pretreated with a variety ofcytotoxic agents. Cell Tissue Kinet., //: 543-553, 1978.
3. Thomas, J. M. A lung colony clonogenic cell assay for human malignantmelanoma in immune-suppressed mice and its use to determine chemosen-sitivity, radiosensitivity, and the relationship between tumour size and response to therapy. Br. J. Surg., 66:696-700, 1979.
4. Kozlowski, J. M., Hart, I. R., Fidler, 1.J., and Hanna, N. A human melanomaline heterogeneous with respect to metastatic capacity in athymic nude mice.J. Nati. Cancer Insl., 72:913-917, 1984.
5. Miller, J. F. A. P., Doak, S. M. A., and Cross, A. M. Role of the thymus inrecovery of the immune mechanism in the irradiated adult mouse. Proc. Soc.Exp. Biol. Med., 112: 785-792, 1963.
6. Phillips, 11. and Gazet, J.-C. Transplantation of primary expiants of humantumour to mice treated with antilymphocyte serum. Br. J. Cancer, 24: 92-95. 1970.
7. Gershwin, M. £.,Ikeda. R. M., Erickson, K., and Owens, R. Enhancementof heterotransplanted human tumor graft survival in nude mice treated withantilymphocyte serum and in congenitally athymic-asplenic (lasat) mice. J.Nati. Cancer Inst., 61:245-248, 1978.
8. Michaelides, M., Hogarth, P. M., and Mackenzie, 1. F. C. The immunosup-pressive effect of monoclonal anti-Lyt-l.l antibodies in vivo. Eur. J. lmmunoi., //: 1005-1012, 1981.
9. Hiraki, S., Miyoshi, I., Nakamura, K., Ohta, T., Tsubota, T., Uno, J., Tanaka,T., and Kiniura, I. Heterotransplantation of human leukemic B-cell, T-celland null-cell lines in hamsters. Acta Med. Okayama, 33:15-20, 1979.
10. Cohen, D. J., Loertscher, R., Rubin, M. F., Tilney, N. L., Carpenter, C. B.,and Strom, T. B. Cyclosporine: a new immunosuppressive agent for organtransplantation. Ann. Intern. Med.. 101:667-682,1984.
11. Morris, P. J. The impact of cyclosporin A on transplantation. Adv. Surg.,/ 7:99-127, 1984.
12. Evans, R. Macrophages and neoplasms: new insights and their implicationin tumor immunobiology. Cancer Metastasis Rev., 1: 227-239,1982.
13. Fidler, I. J. Therapy of spontaneous métastasesby intravenous injection ofliposomes containing lymphokines. Science (Wash., DC), 208: 1469-1471,1980.
14. Kopper, L., Van Hanh, T., Lapis, K., and Timar. J. Increased take rate ofhuman tumour xenografts after carrageenan treatment. Eur. J. Cancer, 16:671-678. 1980.
15. Kiessling, R., and Wigzel, H. An analysis of the murine NK cell as tostructure, function and biological relevance. Immunol. Rev., 44: 165-208,1979.
16. Hanna, N. Inhibition of experimental tumor metastasis by selective activationof natural killer cells. Cancer Res., 42: 1337-1342, 1982.
17. Roder, J., and Duwe, A. The beige mutation in the mouse selectively impairsnatural killer cell function. Nature (Lond.), 2 78:451-453, 1979.
18. Gidlund, M., Orn, A., Wigzel, H., Senik, A., and Gresser, I. Enhanced NKcell activity in mice injected with interferon and interferon inducers. Nature(Lond.), 273: 759-761, 1978.
19. Seaman, W. E.. Blackman, H. A., Gindhart, T. D., Rowbinian, J. R., Loeb,J. M., and Talal, N. Estradici reduces natural killer cells in mice. J. Immunol.,121: 2193-2198, 1978.
20. Kasai, M., Yoneda, T., Hobu, S., Maruyama, Y., Okumura, K., and Tokun-aga. T. In vivo effect of anti-asialo GM1 antibody on natural killer activity.Nature (Lond.), 291: 334-335, 1981.
21. Revesz, L. Effect of leihai I\ damaged tumor cells upon the development ofadmixed viable cells. J. Nati. Cancer Inst., 20: 1157-1186, 1958.
22. Hill, R. P., and Bush, R. S. A lung colony assay to determine the radiosensitivity of the cells of a solid tumour. Int. J. Radiât.Biol. Relat. Stud. Phys.Chem. Med., 75:435-444, 1969.
23. Kovnat, A., Armitage, M., and Tannock, I. Xenografts of human bladdercancer in immune-deprived mice. Cancer Res., 42: 3696-3703, 1982.
24. Kato, T., Irwin, R. J., Jr., and Prout, G. R., Jr. Cell cycles in two cell linesof human bladder carcinoma. Tohoku J. Exp. Med., 121: 157-164, 1977.
25. O'Toole, C. M., Povey, S., Hepburn, P., and Franks, L. M. Identity of somehuman bladder cancer cell lines. Nature (Lond.), 301:429-430, 1983.
26. Tannock, I., Choo, B., and Buick, R. The radiation response of humanbladder cancer assessed in vitro or as xenografts in immune-deprived mice.Int. J. Radiât.Oncol. Biol. Physiol., 10: 1897-1902, 1984.
27. Stanners, C. P., Eliceiri, G. L., and Green, H. Two types of ribosome inmouse-hamster hybrid cells. Nat. New Biol., 230: 52-54, 1971.
28. Porter, E. H. The statistics of dose/cure relationships for irradiated tumours.Part 1. Br. J. Radiol., 53: 210-227, 1980.
29. Withers, H. R., and Milas, L. Influence of preirradiation of lung on development of artificial pulmonary métastasesof fibrosarcoma in mice. CancerRes., 33: 1931-1936, 1973.
30. Brown, J. M., and Parker, E. T. Host treatments affecting artificial pulmonary métastases:interpretation of loss of radioactively labelled cells fromlungs. Br. J. Cancer, 40:677-688, 1979.
31. Gelfand, M. C., and Paul, W. E. Prolongation of allograft survival in miceby adminstation of ami-thy 1 serum. I. Mediation by in vivo activation ofregulatory T-cells. J. Immunol., 115: 1-4, 1975.
32. Herberman, R. B., Nunn, M. E., and Holden, H. T. Low density of thy 1antigen on mouse effector cells mediating natural cytotoxicity against tumorcells. J. Immunol., 121: 304-309,1978.
33. Lake, P., and Douglas, T. C. Recognition and genetic control of helperdeterminants for cell surface antigen thy-1. Nature (Lond.), 275: 220-222,1978.
34. Borei, J. F., Feurer, C., Magnee, C., and Stahelin, H. Effects of the new anti-lymphocytic peptide cyclosporin A in animals. Immunology, .*-'.•1017-1025,1977.
35. Seaman, W. E., Merigan, T. C., and Talal, N. Natural killing in estrogen-treated mice responds poorly to poly I.C despite normal stimulation ofcirculating interferon. J. Immunol., 123: 2903-2905, 1977.
36. Cikes, M., Friberg, S., Jr., and Klein, G. Progressive loss of H-2 antigenswith concomitant increase of cell-surface antigen(s) determined by Moloneyleukemia virus in cultured murine lymphomas. J. Nati. Cancer Inst., 50:347-362, 1973.
37. Kiessling, R., Klein, E., and Wigzel, H. "Natural" killer cells in the mouse.
1. Cytotoxic cells with specificity for mouse Moloney leukemia cells. Specificity and distribution according to genotype. Eur. J. Immunol., 5: 112-117,1975.
38. Akimaru, K., Stuhlmiller, G. M., and Seigier, 11. F. Influence of mousehepatitis virus on the growth of human melanoma in the peritoneal cavity ofthe athymic mouse. J. Surg. Oncol., 17: 327-339, 1981.
39. Steel, G. G., and Adams, K. Enhancement by cytotoxic agents of artificialpulmonary metastasis. Br. J. Cancer, 36:653-658, 1977.
40. Floersheim, G. L. Comparative growth of human tumors in pharmacologically immunosuppressed immune-deprived, cyclosporin A-treated and nudemice. Eur. J. Cancer Clin. Oncol., 18: 589-594, 1982.
41. Lance, E. M., Medawar, P. B., and Taub, R. N. Anti lymphocyte serum.Adv. Immunology, 17:1-92, 1973.
42. Mattes, M. J., Sharrow, S. ().. Herberman, R. B., and Holden, II I.Identification and separation of thy-1 positive mouse spleen cells active innatural cytotoxicity and antibody-dependent cell-mediated cytotoxicity. J.Immunol., 123: 2851-2860, 1979.
43. Homan, W. P., Williams, K. A., Fabre, J. W., Millard, P. R., and Morris, P.J. Prolongation of cardiac xenograft survival in rats receiving cyclosporin A.Transplantation (Baltimore), 31: 164-166, 1981.
44. Selby. P. J.. Thomas, J. M., Monaghan. P., Sloane, J., and Peckham. M. J.Human tumour xenografts established and serially transplanted in miceimmunologically deprived by thymectomy, cytosine arabinoside and wholebody irradiation. Br. J. Cancer, 41: 52-61, 1980.
45. Kovnat, A., Buick, R. N., Connolly, J. G., Jewett, M. A., Keresteci, A. G.,and Tannock, I. F. Comparison of growth of human bladder cancer in tissueculture or as xenografts with clinical and pathological characteristics. CancerRes., 44: 2530-2533, 1984.
1622
on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1986;46:1617-1622. Cancer Res Asya Kovnat, Kong Khoo, Peter Selby, et al. Colony Formation by Human Tumor Cells in MiceMethods of Immunosuppression for Study of Growth and Lung
Updated version
http://cancerres.aacrjournals.org/content/46/4_Part_1/1617
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/46/4_Part_1/1617To request permission to re-use all or part of this article, use this link
on May 24, 2020. © 1986 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from





























