Embed Size (px)
Citation preview

HAL Id: hal-00929275https://hal.archives-ouvertes.fr/hal-00929275
Submitted on 1 Jan 1992
HAL is a multi-disciplinary open accessarchive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come fromteaching and research institutions in France orabroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, estdestinée au dépôt et à la diffusion de documentsscientifiques de niveau recherche, publiés ou non,émanant des établissements d’enseignement et derecherche français ou étrangers, des laboratoirespublics ou privés.
Metabolism and biochemical characteristics of yogurtbacteria. A review
A Zourari, Jp Accolas, Mj Desmazeaud
To cite this version:A Zourari, Jp Accolas, Mj Desmazeaud. Metabolism and biochemical characteristics of yogurt bacte-ria. A review. Le Lait, INRA Editions, 1992, 72 (1), pp.1-34. <hal-00929275>

Lait (1992) 72,1-34© Elsevier/INRA
Review article
Metabolism and biochemical characteristicsof yogurt bacteria. A review
A Zourari *, JP Accolas, MJ Desmazeaud
Station de Recherches Laitières, INRA, 78352 Jouy-en-Josas Cedex, France
(Received 5 July 1991; accepted 1st October 1991)
Summary - This review reports recent data regarding the metabolism and biochemistry ofStreptococcus salivarius subsp thermophilus and Lactobacillus delbrueckii subsp bulgaricus relatedto yogurt manufacture. The taxonomy of these bacteria is presented. Different proposed pathwaysfor carbohydrate metabolism are then discussed, as weil as recent molecular and genetic studies ofthe enzymes involved. Acetaldehyde is the major aromatic compound in yogurt, and so the differentpathways of its formation are briefly described. Recent studies have concerned threonine aldolasewhich catalyzes acetaldehyde synthesis by yogurt bacteria. Exocellular polysaccharides producedby lactic acid bacteria improve the texture of stirred and liquid yogurts. Some of the polysaccharidesof yogurt bacteria are currently known and particular aspects of their production are discussed.Some other properties, le proteolysis, Iipolysis, urease, oxygen metabolism, are also brieflypresented. Interactions between streptococci and lactobacilli are weil established, but more data arerequired for the complete characterization and control of mixed populations. In particular, Iittle isknown about antimicrobial compounds produced by these microorganisms. Bacteriophages ofyogurt bacteria are now weIl characterized, but liltle is known about Iysogeny in thermophilicstreptococci. Finally, progress in genetics (on both plasmid and chromosomal DNA) is brieflydiscussed.
Streptoeoeeus sal/varlus subsp thermophllus 1 Laetobaclllus delbrueekli subsp bulgarleus 1yogurt 1 metabollsm 1 Interaction
Résumé - Métabolisme et caractéristiques biochimiques des bactéries du yaourt. Unerevue. Dans le présent article, la position taxonomique de Streptococcus salivarius subsp thermo-philus et de Lactobacillus delbrueckii subsp bulgaricus est d'abord décrite. Les différentes voies dumétabolisme des glucides sont discutées ainsi que les études récentes des enzymes impliquées,sur le plan moléculaire et génétique. L'acétaldehyde est le composé caractéristique de l'arôme typi-que du yaourt. Les différentes voies métaboliques de sa synthèse sont brièvement décrites, en parti-culier celle qui met en jeu la thréonine aldolase. Les bactéries lactiques produisent des polysaccha-rides exocellulaires qui influencent la texture des yaourts brassés et des yaourts liquides. Despolysaccharides produits par les bactéries du yaourt sont maintenant connus et quelques aspects in-téressants de leur production sont actuellement étudiés. D'autres activités (protéolyse, lipolyse,uréese, métabolisme de l'oxygène) sont aussi brièvement présentées, suite à plusieurs études ré-centes. Les interactions entre streptocoques et lactobacilles sont bien établies, mais davantage dedonnées sont nécessaires pour la caractérisation complète et la msttnse des populations mixtes. Enparticulier, les connaissances des substances antimicrobiennes produites par les bactéries du
• Presentaddress: Boil-GroupeChr Hansen, Le Moulin d'Aulnay, BP 64, 91292 Arpajon Cedex, France

2 A Zourari et al
yaourt sont encore limitées. Les bactériophages de ces bactéries sont maintenant relativement biencaractérisés mais les données sur la Iysogénie du streptocoque demeurent encore incomplètes.Enfin, le progrès des études génétiques (ADN plasmidique et chromosomique) est brièvement discuté.
Streptococcus sallvarlus subsp thermophllus / Lactobaclllus delbrueckll subsp bulgarlcus /yaourt / métabolisme / interaction
CONTENTS
Introduction 2
TaxonomyS salivarius subsp thermophilusL delbrueckii subsp bulgaricus
223
Important metabolic activities for yogurt technologyAcid production - Carbohydrate metabolism
Carbohydrate metabolism of S thermophilus- Lactose and galactose transport- Metabolic pathways of lactose, glucose and galactose- Sucrose utilization
Carbohydrate metabolism of L bulgaricus
Lactic acid isomers in yogurt
44556688
9
9
99
10101111
Formation of f1avour compounds
Flavour compounds of yogurt
Metabolic pathways of acetaldehyde synthesis- Formation of acetaldehyde from glucose- Formation of acetaldehyde from threonine- Formation of acetaldehyde from DNA components- Alcohol dehydrogenaseProduction of polysaccharides
Other metabolic activitiesProteolytic activity
Amino acids in milk and yogurt
Utilization of proteins
Utilization of peptidesLipolytic activityVrease activityOxygen and metabolism
1212
12
12
13131414

Metabolism and biochemistry of yogurt bacteria 3
Particular aspects of yogurt fermentation 15
Interactions between yogurt bacteriaProduction of antimicrobial compounds by yogurt bacteriaBacteriophages of L bulgaricus and S thermophilusGenetics of S thermophilus and L bulgaricus
Conclusion
References
INTRODUCTION
Yogurt and similar fermented milk productshave been very popular for a long time inMediterranean countries (the Balkans,North Africa), in central and southwestAsia (Mongolia, Turkey, Iraq, Iran, Syria)and in central Europe. In many of thesecountries, yogurt is still manufactured us-ing traditional procedures. Since the lastworld war, yogurt consumption has beensteadily increasing not only in Europeancountries, but also in the United States, en-hancing its industrial-scale production. Atpresent, new types of fermented milk areavailable, prepared by adding fruits or fla-vouring, enriched with vitamins or contain-ing selected intestinal bacteria such asLactobacillus acidophilus and severalBifidobacterium species (Kurmann, 1984;Puhan, 1988).
ln France, the term "yogurt" can beused legally only to designate the productresulting from milk fermentation broughtabout exclusively with 2 thermophilic lacticacid bacteria, Streptococcus ssilvsriussubsp thermophi/us and Lactobacillus del-brueckii subsp bulgaricus, which must befound alive in the final product (== 10 million
15161721
2324
cfu.g-1 of yogurt). It is noteworthy thatthese 2 conditions are seldom specified byexisting legislation in most other yogurt-producing countries (Anonymous, 1989).
We review here recent data on some ofthe metabolic and biochemical aspects' ofthese starter bacteria in relation to yogurtmanufacture. Tamime and Deeth (1980)have published an excellent review con-cerning ail the technological and biochemi-cal aspects of yogurt, followed by a secondarticle dealing with its nutritional and thera-peutic properties (Deeth and Tamime,1981). These reviews include a detailedpresentation of S salivarius subsp thermo-phi/us and L delbrueckii subsp bulgaricus.Since 1981, both microorganisms havebeen extensively studied and reviews deal-ing with particular aspects of their metabo-lism and genetics have been published(Hutkins. and Morris, 1987; Mercenier andLemoine, 1989; De Vos, 1990; Mercenier,1990). The present review is an update inthis field and will give the technologicaland biochemical context of yogurt manu-facture. Emphasis is put on bacteria andnot on yogurt and only limited informationconcerning yogurt manufacture will be re-ported.

4 A Zourariet al
TAXONOMY Of S SALIVARIUS SUBSPTHERMOPHILUS AND L DELBRUECKIISUBSP BULGARICUS
S salivarius subsp thermophllus
S salivarius subsp thermophilus is the newname proposed to designate Streptoeoe-eus thermophilus which was originally de-scribed by Orla-Jensen (1919) and whichstands apart from the other streptococciand especially lactic streptococci, atpresent designated as lactococci. It is ex-clusively isolated from the dairy environ-ment, ferments only few carbohydrates, ielactose, sucrose, glucose and sometimesgalactose, and is characterized by its ther-moresistance and a rather high growthtemperature which may reach 50-52 "C.No group-specific antigen has been found(Hardie, 1986).
DNA-DNA homology studies might sup-ply more information on this bacterium andmay question its classification. The guano-sine plus cytosine content (mol G + C%) ofDNA ranges from 37.2-40.3% accordingto Farrow and Collins (1984). These au-thors obtained 61-78% homology betweenDNA of several S thermophi/us strains anda DNA probe of Streptoeoccus salivarius(the homology between DNA of S salivari-us strains and a DNA probe of S thermo-phi/us ranged from 67-91%). They thusconfirmed the high DNA homology (70-100%) observed in an earlier study byKilpper-Bëlz et al (1982) for 2 strains ofeach species. On the basis of these re-sults, Farrow and Collins (1984) proposedthat S thermophi/us should be reclassifiedas a subspecies of S salivarius despite thelarge phenotypic differences betweenthese 2 bacteria. In fact, lactate dehydro-genases of the type-strains of both strepto-cocci have different properties (Garvie,
1978). Furthermore, the therrnoresistanceof their fructose-diphosphate aldolases isdifferent, as is their level of homology withEnteroccoeus faeealis aldolase assessedby immunodiffusion assays, in spite of veryclose migration distances (London andKline, 1973).
ln addition, Garvie and Farrow (1981)had previously placed these streptococcalstrains in the same group, based on ho-mology between their DNA and ribosomalRNA of the type-strain of Streptoeoccusbovis (this strain now belongs to the Strep-toeoccus equinus species; Schleifer andKilpper-Bâlz, 1987), but in different c1us-ters. Besides, DNA-DNA molecular hybrid-ization between several strains of S ther-mophi/us and S salivarius have onlyyielded a 60% homology in optimum condi-tions and 30% under stringent hybridiza-tion conditions (Schleifer and Klllper-Bâlz,1987). Based on recent hybridization data,phenotypic differences and occurrence incompletely different biotopes (mouth/milk),Schleifer and Kilpper-Bâlz (1987) suggest-ed maintaining S thermophi/us and S sali-varius as 2 separate species. It is obviousthat the taxonomy of S thermophi/us re-quires further discussion and that moredata are required before reaching a defini-tive decision.
Colmin et al (1991) developed a DNAprobe which specifically hybridized to 25strains of subspecies thermophi/us and to2 strains of subspecies salivarius of S sali-varius. This cross-hybridization suggeststhat both species share common DNA se-quences, but more data are needed tosupport a possible close relationship be-tween these 2 streptococci. In addition, theprobe easily detects RFLP (restriction frag-ment length polymorphism) in S thermophi-lus and accordingly can be used for straincharacterization.

Metabolism and biochemistry of yogurt bacteria
L delbrueckii subsp bulgaricus
This lactobacillus was first described byOrla-Jensen (1919) and named Thermo-baeterium bulgarieum. It is presently con-sidered as one of the subspecies of Lae-tobaeillus delbrueekii. Two other sub-species, subsp delbrueekii and subsp lae-tis, also belong to this group. The old ter-minology Laetobaeillus leiehmannii is nolonger in use.
L delbrueekii subsp bulgarieus is homo-fermentative, ferments few carbohydrates,ie glucose, lactose, fructose, and some-times galactose or mannose, and has ahigh growth temperature (up to 48 or 50OC).Its DNA mol G + C% ranges from 49to 51% (Kandler and Weiss, 1986).
Simonds et al (1971) obtained a DNAhomology of 86% between the formerL bulgarieus and L laetis species, but only4.8% with Laetobaeillus helvetieus and nohomology with L helvetieus var jugurti. Thiswas confirmed by Dellaglio et al (1973)who obtained < 7% DNA homology be-tween L bulgarieus and L helvetieus orL helvetieus var jugurti. The study of Weisset al (1983) showed 80 to 100% homology(90-100% for type-strains) between theformer L bulgarieus, L leiehmannii, L laetisand L delbrueekii species. DNA homologyled to the reclassification of these 4 lactob-acilli as subspecies of L delbrueekii in themost recent classification of Bergey's Man-ual (Kandler and Weiss, 1986).
A specifc DNA probe for L delbrueekiihas been developed recently, whichmakes it possible to differentiate this spe-cies from other lactobacilli, lactococci andpropionibacteria (Delley et al, 1990).
ln the present article, we shall use theformer names of the yogurt organisms, ieS thermophilus and L bulgarieus for pur-poses of convenience.
5
IMPORTANT METABOLIC ACTIVITIESFOR YOGURT TECHNOLOGY
The role of streptococci and lactobacilli inyogurt manufacture can be summarized asfollows: milk acidification, synthesis of aro-matic compounds, development of textureand viscosity. The latter aspect is requiredmainly for stirred and liquid yogurts. Thus,for industrial yogurt manufacture, starterselection takes into account these 3 prop-erties. Evaluation of acidification propertiesis difficult because of the high buffering ca-pacity of milk and the lack of a standardprocedure. Efforts have been made to de-velop new methods for an objective evalu-ation of strains in replacement of formerprocedures (eg that of Accolas et al, 1977)based on the measurement of titratableacidity in milk. For example, the methodpatented by Corrieu et al (1989), and de-scribed by Spinnler and Corrieu (1989),uses continuous automatic recording of pHduring bacterial growth in milk and thestrains are compared on the basis of maxi-mum acidification rates and the corre-sponding time and pH. In the field of indus-trial preparation of mixed starter cultures,efforts have been made to control thegrowth of streptococcus and lactobacilluspopulations by culture conditions, pH andtemperature (Béai et al, 1989). Chambaand Prost (1989) evaluated the acidifyingproperties of strains used for cheesemak-ing, based on pH measurements duringgrowth following the temperature profile ofa standard cheesemaking operation.
The evaluation of aroma formation isgenerally based on the production of ace-taldehyde, a major aromatic compound ofyogurt, whereas the thickening character isbased on measurements of milk viscosity(Bouillanne and Desmazeaud, 1980, 1981;Zourari et al, 1991; Zourari and Des-mazeaud,1991).

6
ln the first part of this review, wepresent the metabolic activities responsi-ble for the acidification, f1avouring andthickening properties of yogurt bacteria.We then describe some metabolic activi-ties of lesser importance for yogurt manu-facture, but which determine the growth ofbacteria in milk and their interactions, ieproteolysis, lipolysis, urease activity, oxy-gen effect.
Acid production.Carbohydrate metabolism
The main role of S thermophilus and L bul-garicus in yogurt manufacture is to acidifymilk by producing a large amount of lacticacid from lactose. Lactic acid reduces thepH of the milk and leads to a progressivesolubilization of micellar calcium phos-phate. This causes the demineralization ofcasein micelles and their destabilization,which generates the complete precipitationof casein in a pH range of 4.6-4.7 (Fox,1989). In addition, lactic acid provides yog-urt with its sharp, acid taste and contrib-utes to flavour.
Lactic acid production may also occurduring yogurt storage at low temperature.lt may lead to an excessive acidificationwhich affects the organoleptic propertiesof the product. This activity depends onthe strains used and especially on the lac-tobacilli (Accolas et al, 1977; Bouillanneand Desmazeaud, 1980, 1981).
Carbohydrate metabolismof S thermophilus
Lactose and galactose transport
The mechanism of lactose transport inS thermophilus differs from that of lacto-cocci, which possess a specific system forlactose transport, the phosphoenolpyru-vate (PEP)-dependent phosphotransfe-
A Zourariet al
rase system (PTS). Lactose-6-phosphateformed during transport is hydrolyzed byan intracellular phospho-B-qalactosidaseinto glucose and galactose-6-phosphatewhich are then metabolized into lactic acid(Thomspon, 1987). S thermophilus doesnot possess lactose PTS or phospho-û-galactosidase (Tinson et al, 1982a; Thom-as and Crow, 1984), but has a lactose per-mease system including a proton-dependent membrane-Iocated permeaseand an intracellular ~-galactosidase (Pool-man et al, 1989).
The lactose permease gene of S ther-mophilus (lacS) was cloned and expressedin Escherichia coli (Poolman et al, 1989).The comparison of amino acid sequencesshowed that the streptococcal lactosetransport protein (LacS) differs from thelactose permease of E coli (LacY) and ap-pears to be a hybrid protein of 69 kDa. TheNH2-terminal region is homologous withthe melibiose carrier (MeIB) of E coli (23%overail similarity) whereas the COOH-terminal end is partially homologous (34-41%) with enzyme III (E III) of 3 differentPEP-PTS systems.
Galactose seems to be directly involvedin lactose transport. In fact, the presenceof a large amount of galactose in thegrowth medium inhibits lactose transport(Somkuti and Steinberg, 1979). Biochemi-cal studies have shown that galactose hasa considerable affinity _for lactose per-mease and acts as a competitive inhibitorof the lactose transport system (Hutkinsand Morris, 1987), but more recent datasupply another explanation. In membranevesicles of E coli, lacS permease not onlycatalyzes a proton motive force-Iinkedtransport system, but also exchange of ~-galactosides. This means that the lactosetransport reaction proceeds as a lactose-galactose antiport independent of the pro-ton motive force (Poolman et al, 1989; Hut-kins and Ponne, 1991).

Metabolism and biochemistry of yogurt bacteria 7
Galactose transport requires an exoqe-nous energy source and involves the ac-tion of a galactose permease. A PEP-PTS-like system has not been detected inS thermophilus. Glucose and galactosetransport is slower than that of lactose(Hutkins et al, 1985a). The preferentialutilization of lactose could be due tolimitations in monosaccharide transport(Somkuti and Steinberg, 1979). In light ofthe involvement of galactose in lactosetransport, however, these data should bere-evaluated.
Metabolic pathways of lactose,glucose and galactose utilization
Figure 1 summarizes the proposed path-ways for lactose and galactose transportand their utilization by S thermophilus. In-tracellular ~-galactosidase (~-gal) hydrolyz-es lactose to glucose and galactose. Glu-cose represses synthesis of this enzyme(Tinson et al, 1982a). Streptococcal ~-galhas bsen purified and characterized (Ra-mana Rao and Dutta, 1981; Greenbergand Mahoney, 1982; Smart et al, 1985).The ~-gal-encoding gene has been clonedand expressed in E coli (Herman andMcKay, 1986) and in Saccharomyces ce-revisiae (Lee et al, 1990). Streptococcal ~-gal is a protein of about 105 kDa and the~-gal-encoding gene is located in a 3.85-kbregion of the chromosomal DNA (Hermanand McKay, 1986). This enzyme is usefulas a potential selective marker in food-grade cloninq vectors, and also for its pos-sible industrial production because of itshigher thermostability compared tocurrent-Iy produced yeast enzymes.
The glucose moiety of lactose is directlyused via the Embden-Meyerhof-Parnaspathway (fig 1). S thermophilus possesses2 fructose-1,6-diphosphate-independent(FDP-independent) lactate dehydrogenas-es (LDH) which reduce pyruvate to lacticacid (Garvie, 1978).
The galactose moiety is released intothe extracellular medium by most S ther-mophilus strains which are not able to useit, ie Gal- phenotype (Hutkins and Morris,1987). Galt strains have been isolated(Somkuti and Steinberg, 1979; Thomasand Crow, 1984), however, in which galac-tose is metabolized via the Leloir pathwayinvolving the enzymes galactokinase,galactose-1-phosphate-uridyl transferaseand uridine-5-diphospho-glucose-4-epime-rase. The 2 latter enzymes are constitutiveand present in Gal- as weil as in Gal!strains (Thomas and Crown, 1984). Thedifference between the 2 phenotypesseems to be due to a difference in galac-tokinase and/or galactose permease activi-ty. Both enzymes are induced by galactoseand repressed by lactose (Hutkins et al,1985a, b). Nucleotide sequences of thegenes encoding the 2 constitutive en-zymes are homologous with the analogousgenes of other bacteria and yeasts. Theirexpression and regulation have been stud-ied (Poolman et al, 1990). Until the presenttime, the use of galactose via the tagatosepathway has not been reported for eitherS thermophilus or L bulgaricus.
It is possible to obtain Gal+ derivativesof S thermophilus strains after growth ofGal- cultures in a chemostat under lactoselimitation (Thomas and Crow, 1984). Theuse of Gal+ strains in cheesemaking de-creases residual galactose in Cheddar,Mozzarella, and Emmental cheeses andthus avoids sorne serious technologicalproblems. As mentioned by Hutkins et al(1986), high residual galactose in cheesecurd: (i) may enable a heterofermentativemetabolism of galactose during ripening,with concomitant carbon dioxide produc-tion, and may also lead to the developmentof a harmful flora generating off-flavoursand texture defects and (ii) has been impli-cated in browning reactions which occurduring processing or cooking at high tem-

8 A Zourari et al
Lactose GalactoseLactose Galactose•1
ADP ATP
2H:.-------H+
Galactase-I-P
~
UDP-gIUCaSeATP Gal-I-P uridyl trans1erasee
Hexokinase Glucose-I-phosphoteADP UDP-goloctose
~UDP-glucose -4-eplmeras e
GIUCOse-6-p_G1UCOSe-l-p"7"\ UDP-glucose
~ UTP PPi. Fructose-6-P
ATP~ 1 LELOIR PATHWAY 1~P~ .
Fructose-I.6-DiP
~Aldolase
Oihydroxyacetone C p Glycerqldehyde-3-P11
PhosPholno,pyruvate
ADP ----)1ATP.....- {yruvate kinase
Pyruvate
NADH + H+ ------JLactate dehydrogenaseNAD+~
1 LAGTIG AGIO 1
1 EMBDEN-MEYERHOF-PARNAS PATHWAY 1
Fig 1. Proposed pathways for lactose (lac) and galactose (gal) utilization in S thermophilus. From Hut-kins and Morris (1987), Thompson (1987) and De Vos (1990). P: phosphate. PPi: inorganic pyrophos-phate, ATP: adenosine triphosphate, ADP: adenosine diphosphate, UTP: uridine triphosphate, UDP:uridine diphosphate.Voies métaboliques proposées pour l'utilisation du lactose (lac) et du galactose (gal) par S thermophi-lus. D'après Hutkins et Morris (1987), Thompson (1987) et De Vos (1990). P: phosphate, PPi : pyro-phosphate inorganique, A TP : adénosine triphosphate, ADP : adénosine diphosphate, UTP : uridinetriphosphate, UDP : uridine diphosphate.

Metabolism and biochemistry of yogurt bacteria 9
peratures of cheeses such as Cheddar orMozzarella.
Sucrose utilization
Sucrose metabolism has been poorly doc-umented, especially at the molecular andgenetic levels. During the growth of S ther-mophilus on sucrose, both glucose andfructose are used simultaneously. Fructoseaccumulates in the growth medium, how-ever, even when the strain can use it(Thomas and Craw, 1983). The higher rateof sucrose utilization compared to those ofglucose and fructose might be due to a dif-ferent uptake rate (Hutkins and Morris,1987).
An inhibitory effect of high sucrose con-tent in milk (10-12%), on the growth ofyogurt bacteria has often been reported. Itis due to both an adverse osmotic effect ofthe solutes in milk and a low water activity(Tamime and Robinson, 1985).
Carbohydrate metabolismof L bulgarieus
L bulgaricus is characterized by the utiliza-tion of a small number of carbohydrates,but their metabolism has not been studiedenough. This lactobacillus uses only theglucose moiety of lactose and releases ga-lactose into the growth medium. Sornestrains can use galactose in a growth me-dium containing Iimiting concentrations oflactose. Glucose transport occurs via aPTS-like system, while lactose and galac-tose uptake occurs by means of per-meases (Hickey et al, 1986). Recent find-ings suggest that intracellular galactose isexchanged with lactose, enabling thetransport of lactose as in S thermophilus.ln fact, preliminary results indicate that ga-lactose efflux from the cells is stimulatedalmost 100-fold by the presence of lactosein the external medium (Poolman et al,1990). The lactose genes and gene organi-
zation are weil conserved in S thermophi-lus and L bulgaricus. Their lactose per-meases and 13-galactosidasesshare 60and 48% cornmon amino acids, respective-Iy (Mercenier, 1990).
Once inside the cell, lactose is hydro-Iyzed by a 13-gal.Glucose represses lac-tose transport and 13-galsynthesis. Purified13-galof an L bulgaricus strain has a maxi-mum activity at 50 "C and pH 7 (Itoh et al,1980). The 13-gal-encodinggene has beencloned in E coli and expressed using itsown promoter. The molecular mass of theenzyme is about 114 kDa, deduced fromthe nucleotide sequence of the corre-sponding gene (Schmidt et al, 1989). Ga-lactose is metabolized by L bulgaricus viathe Leloir pathway. A galactokinase activityhas been detected in Gal" strains (Hickeyet al, 1986).
Lactic acid isomers in yogurt
S thermophilus possesses 2 FDP-independent L-LDH (Garvie, 1978; Hemmeet al, 1981a) and produces L(+) lactic acid,while L bulgaricus produces OH lacticacid and possesses an NAD-dependentstereospecific LDH (Gasser, 1970). Thus,both lactic acid isomers are simultaneouslyproduced in yogurt.
The D-LDH of L bulgaricus purified byLe Bras and Garel (1991) is a dimer corn-posed of 2 identical 37-kDa chains. Theamino-terminal sequence determined on50 residues is the first D-LDH sequenceavailable and does not show significant ho-mology with any known LDH. This sug-gests that the evolutionary process of 0-LDH from L bulgaricus is not identical tothat of the NAD-dependent L-LDH family.
0(-) lactic acid is metabolized only veryslowly in man compared to the L(+) isomerand may cause metabolic disorders if in-gested in excess. The World Health Or-ganization therefore recommends a Iimited

10 A Zourariet al
daily consumption of D(-) lactic acid to100 mg.kg-1 bodyweight and some coun-tries, eg Germany, have decided to mini-mize the D(-) lactic acid content of fer-mented dairy products (Kondratenko et al,1989). In yogurt, this can only be obtainedin the absence of L bu/garieus producingthe D(-) isomer, which accounts for 30-50% of the total lactic acid content of yo-gurt (Kunath and Kandler, 1980; Alm,1982a). Nevertheless, the D(-) lactic acidcontent may reach critical concentrationsthat can lead to disturbances especiallywhen the diet is extremely unbalanced, butthis finding has not yet been reported(Gurr, 1986). Rare cases of D(-)-Iactic aci-dosis have been reported in children andadults, in association with altered bowelphysiology (short gut syndrome) and pro-liferation of active D-Iactate bacterial pro-ducers (Szylit and Nugon-Baudon, 1985).
Detection of a weak L-LDH activity inthe type-strain of L bu/garieus grown in achemostat is therefore very interesting(Ragout et al, 1989). The identification ofits mechanism of regulation could contrib-ute to increase L(+) lactic acid synthesisby this species and to reduce the D(-) lac-tic acid content of yogurt.
Formation of flavour compounds
Flavour compounds of yogurt
The typical flavour of yogurt is due to lacticacid and various carbonyl compounds, ieacetaldehyde, acetone and diacetyl, pro-duced by S thermophilus and L bu/garieus.ln addition to carbonyl substances, manyvolatile compounds have also been identi-fied in yogurt, ie volatile fatty acids (Turcicet al, 1969; Dumont and Adda, 1973) andseveral compounds derived from the ther-mal degradation of lipids, lactose and pro-teins during the heat treatment of milk be-
fore yogurt manufacture eg aldehydes, ke-tones, alcohols, lactones, sulfur corn-pounds (Tamime and Deeth, 1980).
Acetaldehyde is considered as the ma-jor flavour component of yogurt (Pette andLolkema, 1950c; Dumont and Adda, 1973;Law, 1981) while high concentrations ofthis compound in other dairy products(cheese or cream) lead to flavour defectsdescribed as "green" or "yogurt-Iike" (Lind-say et al, 1965). Bottazzi and Vescovo(1969), however, underlined the impor-tance of the acetaldehyde to acetone ratio.
Diacetyl contributes to the delicate, fullflavour of yogurt and seems to be impor-tant when the acetaldehyde content is low(Groux, 1973a). According to Rasic andKurmann (1978) S thermophilus is the onlymicroorganism responsible for diacetyl pro-duction in yogurt, whereas Dutta et a/(1973) reported that diacetyl is also largelyproduced by L bu/garieus (12-13 ppm forboth species in pasteurized milk). Rysstadand Abrahamsen (1987) detected diacetylin goat's milk yogurt with a low acetalde-hyde content and suggested that diacetylmight be very important for the flavour ofgoat's milk yogurt.
Even though the importance of acetal-dehyde, acetone and diacetyl for yogurtflavour is weil established, it is very difficultto evaluate the contribution of several iden-tified volatile compounds because theirsensory perception thresholds vary consid-erably (Dumont and Adda, 1973).
ln spite of important progress in chro-matographic methods such as head-spacetechniques which increase the detectionsensitivity of volatile compounds (Degorce-Dumas et al, 1986), only few recent stud-ies are available in this field and none ofthem has involved a large number ofstrains examined using the recently devel-oped techniques. Thus, it ls difficult tocompare results and conflicting findings

Metabolism and biochemistry of yogurt bacteria
may be due to the different sensitivity ofthe methods previously used.
Metabolic pathwaysof acetaldehyde synthesis
Formation of acetaldehyde from glucose
Two possible pathways exist:- The Embden-Meyerhof-Parnas pathwaygenerates pyruvate. An a-carboxylase cat-alyzes the formation of acetaldehyde frompyruvate. An aldehyde dehydrogenasemay also generate acetaldehyde from ace-tyl-CoA which is formed from pyruvate bythe action of a pyruvate dehydrogenase.Neither significant a-carboxylase nor alde-hyde dehydrogenase activities have beendetected by Raya et al (1986) in 2 strainsof S thermophilus and 2 strains of L bulgar-icus. Nevertheless, Lees and Jago (1976a)found an aldehyde dehydrogenase activityin 4 strains of each species. Therefore it isdifficult to conclude whether this pathwayoccurs rarely or frequently in yogurt bacte-ria.- The hexose monophosphate (HMP)pathway generates acetylphosphate. Aphosphotransacetylase catalyzes the for-mation of acetyl-CoA from acetylphos-phate and an acetate kinase leads to theformation of acetate. Although Raya et al(1986) detected these 2 activities in bothyogurt bacteria, acetaldehyde cannot beformed by this pathway since their strainsdid not possess aldehyde dehydrogenasewhich catalyzes the biosynthesis of acetal-dehyde from acetyl-CoA or acetate.
Formation of acetaldehyde from threonine
Threonine aldolase catalyzes the c1eavageof threonine to acetaldehyde and glycineand appears to be the most importantpathway for acetaldehyde production in yo-gurt. In tact, Raya et al (1986) detected
11
this enzyme in 2 strains of L bulgaricus,but not in the 2 strains of S thermophilusinvestigated. In other studies, however,this enzyme was detected in both species(Lees and Jago, 1976b; Wilkins et al,1986a; Marranzini et al, 1989). Streptococ-cal threonine aldolase activity decreasessignificantly when growth temperature in-creases from 30 to 37 oC (Lees and Jago,1976b) or from 30 to 42 -c (Wilkins et al,1986b), but that of L bulgaricus remains al-most identical. Thus, at 37 and 42 oCthre-onine aldolase activity of the streptococciis significantly lower than that of the lactob-acilli, whereas it is identical at 30 oC.Sinceyogurt is generally manufactured at hightemperatures (43· OC), acetaldehyde isprobably.produced mainly by L bulgaricus.
The addition of threonine (0.6%) or gly-cine (20/0)to the growth medium does notsignificantly affect the specific activity ofthreonine aldolase in cell extracts (Leesand Jago, 1976b). High threonine concen-trations combined with low glycine concen-trations in the growth medium increase thebiosynthesis of the enzyme. When this ra-tio is inverted, a decrease is observed.However, the threonine concentration dur-ing growth may have a greater effect on in-creasing threonine aldolase activity thanthe glycine concentration may have on in-hibiting it. Streptococcal threonine aldolaseis more affected by hi~ glycine concentra-tions than the lactobacillus enzyme (Mar-ranzini et al, 1989). The threonine aldolaseof a L bulgaricus strain has been partiailypurified and studied by Manca de Nadra etal (1987). Maximum enzyme activity is ob-served at 40 oC and pH 6.5 as weil as astrong allosteric pH-dependent inhibitionby glycine. Inhibition of threonine aldolaseby glycine could explain the low acetalde-hyde content of yogurt from goat's rnilk,which is richer in glycine than cow's milk(2.4 versus 0.84 mg.100 ml-1; Rysstad etal,1990).

12
Formation ot acetaldehydetrom DNA components
Two strains of S thermophilus and oneL bulgaricus strain studied by Raya et al(1986) possess a deoxyriboaldolase whichcatalyzes the synthesis of acetaldehydefrom 2-deoxyribose-5-phosphate. This en-zyme could be involved in the degradationrather than the synthesis of DNA. The 4lactobacilli strains and 3 out of the 4 strep-tococcal strains examined by Lees andJago (1977) did not contain this enzyme.
A~oholdehydrogenase
Alcohol dehydrogenase reduces acetalde-hyde to ethanol. This enzyme has notbeen detected in L bulgaricus, but hasbeen found in 2 of 4 strains of S thermo-philus (Lees and Jago, 1976a; Marshalland Cole, 1983; Raya et al, 1986).
Production of polysaccharides
Several Gram-negative and Gram-positivebacteria, including lactic acid bacteria, pro-duce exocellular polysaccharides. The"ropy" character is often required for themanufacture of many fermented milk prod-ucts. Ropy strains of lactococci have beenisolated from Scandinavian fermenteddairy products (Makura and Townsley,1984; Nakajima et al, 1990). In stirred yo-gurt, yogurt beverages and low milk solidsyogurts, production of polysaccharides canimprove viscosity and texture, increase re-sistance to mechanical handling and de-crease susceptibility to syneresis. The useof ropy strains is particularly important inFrance and in the Netherlands where theaddition of stabilizers in yogurt is prohibit-ed. In fact, some strains of S thermophilusand L bulgaricus produce neutral exopoly-saccharides. Only few studies are availa-ble on the characterization and productionof these polymers (Cerning, 1990).
A Zourariet al
Schellhaass (1983) isolated ropy strainsof some thermophilic lactic acid bacteria.She observed higher exopolysaccharideproduction in milk at suboptimal growthtemperatures, as measured by an increaseof relative viscosity. The polymer materialobtained contained galactose and glucosein the ratio 2:1. The slime secreted by astrain of L bulgaricus studied by Groux(1973b) contained mainly galactose andsome glucose, mannose and arabinose.Cerning et al (1988) studied an exocellularpolysaccharideproduced by S thermophilus,which contains galactose (55%), glucose(25%), mannose (15%) and small amountsof rhamnose, xylose and arabinose. The Lbulgaricus exopolysaccharide studied byCerning et al (1986) contains galactose,glucose and rhamnose in an approximatemolar ratio of 4:1:1 with a molecular massof about 500 kDa and intrinsic viscosity of4.7 dl.g-1. Doco et al (1990) studied an ex-opolysaccharide produced by S thermophi-lus after 3-4 h of incubation and obtaineda polymer composed of galactose, glucoseand N-acetylgalactosamine in a ratio of2:1:1. The molecular mass of the polymeris 1 x 103 kDa and intrinsic viscosity 1.54dl.g-1. Rhamnose, present mostly in theexopolysaccharides of L bulgaricus, hasnot been identified in the exocellular poly-mer produced by 3 lactobacillus strainsisolated from traditional Greek yogurts(Zourari, 1991). These molecules are com-posed of galactose (5O-70%), glucose(2D-40%) and some mannose and arabi-nose.
The quantities of polymer formed byropy strains of both species vary consider-ably even under identical experimentalconditions (Giraffa and Bergère, 1987;Cerning et al, 1990; Zourari, 1991). It is dif-ficult to establish a good correlation be-tween the quantity of polysaccharide pro-duced and the corresponding viscosity.This difficulty may be due to changes in

the 3-dimensional configuration of poly-mers and to their interactions with somemilk compounds, mainly caseins that areprecipitated at low pH (Olsen, 1989). In ad-dition, viscosity measurements are difficultto interpret for non-Newtonian solutionssuch as milk and fermented milk products.Bottazzi and Bianchi (1986) studied byscanning electron microscopy (SEM) thestructure of milk fermented with a ropyL bulgaricus strain. It was shown that apart of the polysaccharide covers the cellswith an uniform layer, and the rest of thepolymer binds cells together and to milkcasein via a dense network of visible fila-ments. Recently, Teggatz and Morris(1990) used SEM to explain changes oc-curing in the microstructure of ropy yogurtwhen it is subjected to a shear force. Theiraim was to understand the interactions ofexopolysaccharide material with its sur-roundings, and how they may influenceviscosity. It was observed that an increaseof shear rate first disrupts the attachmentof polymer to the bacterial surface, but thepolysaccharide material remains incorpo-rated with the casein where it continues toinfluence viscosity.
The ropy character of S thermophilus isoften unstable (Cerning et al, 1988). Thismay be partially due to the presence of gly-co hydrolase enzymes capable of hydrolyz-ing the polysaccharide material. There isno evidence of a possible loss of plasmidsencoding this character as in Lactococcuslactis subsp cremoris (Vedamuthu and Ne-ville, 1986) or in Lactobacillus casei subspcasei (Vescovo et al, 1989). In fact, ropystrains of yogurt bacteria studied by Cern-ing et al (1986, 1988, 1990) and Zourari(1991) are plasmid-free.
Metabolism and biochemistry of yogurt bacteria 13
OTHER METABOLIC ACTIVITIES
Proteo/ytie aetivity
Amino acids in milk and yogurt
ln yogurt, proteolysis is not determinant fororganoleptic properties, but it is an impor-tant factor for the selection of strains forcheesemaking. On the other hand, proteo-Iytic activity is greatly involved in both nutri-tion and interactions of yogurt bacteria,since lactic acid bacteria cannot synthe-size essential amino acids. Therefore, theyrequire an exogenous nitrogen source andutilize peptides and proteins in their growthmedium by more or less complete enzymesystems.
S thermophilus primarily requires glu-tamic acid, histidine and methionine, asweil as cystine, valine, leucine, isoleucine,tryptophan, arginine and tyrosine forgrowth (Shankar and Davies, 1977; Brac-quart et al, 1978). The uptake of branched-chain amine acids has been studied. lt isan active transport which requires an exog-enous energy source, depends on temper-ature and pH and is inhibited by L-cysteine(Akpemado and Bracquart, 1983).
The free amine acid content of cow'smilk generally does not exceed 10 mg·100ml-1 (Rasic and Kurmann, 1978; Miller etal, 1964; Alm; 1982c). In yogurt, the freeamine acid pattern depends on the type ofmilk (animal species, season), its heattreatment, manufacturing techniques, bac-terial strains used and storage conditions(Rasic et al, 1971 a, b). The free aminoacid content generally increases in yogurtcompared to that of milk. L bulgaricus ap-

14
pears to be the main species responsiblefor these changes (Miller et al, 1964; Alm,1982c; Feller et al, 1990).
Utilization of proteins
Milk proteins (caseins, whey proteins) arethe main nitrogen source for lactic acidbacteria, which utilize them with the actionof exocellularproteinases, membrane-bound aminopeptidases and intracellularexopeptidases and proteinases.
Proteinase activity has been detected inseveral strains of lactobacilli and strepto-cocci by Ezzat et al (1985) and Kalantzo-poulos et al (1990). L bulgaricus possess-es a firmly cell-bound proteinase withoptimum activity between 45 and 50 "Cand pH values ranging from5.2 to 5.8(Argyle et al, 1976). The partiaily purifiedcell-wall-associated proteinase studied byEzzat et al (1987) has maximum activity at35 "C and pH 5.5. The proteinase of L bul-garicus is more active on ~-casein than onwhey proteins (Chandan et al, 1982). ElSoda and Desrnazeaud (1982) and Laloi(1989) revealed a preferential but partialhydrolysis of ~- and us1-caseins. Laloi(1989) also observed that after growth inmilk, caseinolytic activity is 3 times higherthan that measured after growth in a com-plex medium rich in short peptides.
Desmazeaud (1974) studied an intracel-lular metalloproteinase of S· thermophiluswhich was more active on the carboxy-methylated B-chain of insulin or on gluca-gon than on caseins. A weak caseinolyticactivity was detected in both the cell enve-lopes and cytoplasm of S thermophilus. Asthe 2 corresponding enzymes had thesame electrophoretic mobility they wereconsidered to be identical (Meyer et al,1989). A high cell-wall-associated protei-nase activity characterizes 2 strains ofS thermophilus studied by Shahbal et al
A Zourari et al
(1991) who also obtained a proteinase-negative mutant. Strains which possessthis proteolytic activity have a significantlyhigher acidification rate in milk comparedwith 13 other randomly chosen strains.
Utilization of peptides
The low molecular weight peptide fractionof milk is an important nitrogen source foryogurt bacteria. The importance of pep-tides for their growth stimulation and theiracidification is now weil established, espe-cially for S thermophilus (Desmazeaud andDevoyod, 1970; Desmazeaud and Her-mier, 1973; Bracquart and Lorient, 1979;Hernrne etai, 1981b).
S thermophilus generally possesses aleucine-aminopeptidase activity (Bouillanneand Desmazeaud, 1980). Some strainspossess an arginine-amino-peptidase ac-tivity which is usually inactive against di-peptides. Two intracellular metalloen-zymes of this species have been purifiedand characterized, ie an amino-peptidasewith broad specificity and a dipeptidasewhich hydrolyzes dipeptides with a hydro-phobie and bulky residue at the NH2-terminal end (Rabier and Desma-zeaud,1973). A non specific prolyl-dipeptidase hy-drolyzing severai dipeptides has also beendetected '(Desmazeaud and Jugé, 1976).An X-prolyl-dipeptidylamino-peptidase (X-pro-DPAP) of a S thermophilus strain hasbeen purified, characterized and comparedwith that of a L bulgaricus strain. Thestreptococcal enzyme is composed of 2subunits, has a molecular mass of 165küa, an isoelectric point of 4.5 and opti-mum pH values ranging from 6.5 to 8.2(Meyer and Jordi, 1987).
Two intracellular exopeptidases of L bul-garicus have been characterized by ElSoda and Desmazeaud (1982), an amino-peptidase with specificity limited to argi-

Metabolism and biochemistry of yogurt bacteria
nine-~-naphthylamide, and a dipeptidasewith broad specificity. Laloi (1989) re-vealed the presence of 4 aminopeptidases(API, Il, III, IV) and an X-pro-DPAP in thesame species. API and APIII (cytoplasmicenzymes) are induced after growth in acomplex medium, while APII and APIV areconstitutive. Mutants partially or totally freeof APII or X-pro-DPAP have been isolatedafter chemical mutagenesis. Deficiency ofthese 2 enzymes significantly increasesthe caseinolytic activity of mutants corn-pared to the parental strain (Atlan et al,1989, 1990).
Lipolytic activity
Lipolysis is generally low in yogurt and is. therefore not significant in terrns of flaveur.
The free fatty acid content of yogurt differsonly slightly from that of milk (Rasic andVucurovic, 1973; Alm, 1982b). Therefore,only few studies with these enzyme sys-tems have been reported in yogurt bacteria.
Tamime and Deeth (1980) have sum-marized the main lipolytic activities as fol-lows: the hydrolytic activity of streptococcion tributyrin and triolein is high and, in-versely, is low on milk fat and on Tween 40or 60. The hydrolytic activity of lactobacillion tributyrin is weaker, while esterases hy-drolyzing several soluble substrates, eg n-naphthylacetate, are active in both bacte-ria.
L bulgaricus possesses intracellular es-terases active on ortho- and para-nitrophenyl derivatives of short-chain fattyacids and activity levels vary only slightlybetween different strains (El Soda et al,1986). Many strains of S thermophilus (30out of 32 tested) possess a lipase more ac-tive towards tributyrin than towards naturallipids (De Moraes and Chandan, 1982)and maximum activity was observed at45 -c and pH 9. Kalantzopoulos et al
15
(1990) detected esterase-1 and esterase-2activities in both yogurt bacteria, using 2-and 4-nitrophenylbutyrate, respectively.
Urease activity
ln milk, S thermophilus produces a largeamount of carbon dioxide (C02) which isnot formed from lactose metabolism sincethis microorganism is a strictly homofer-mentative species (Driessen et al, 1982). Italso produces a large quantity of ammonia(NH3) which was initially explained as re-sulting from the deamination of some ami-no acids (Groux, 1973a). CO2 productionis due to the activity of urease, whichbreaks down milk urea (about 250 mg.I-1)into CO2 and NH3 (Miller and Kandler,1967b; Tinson et al, 1982b; Juillard et al,1988). This leads to the alkalinization ofthe growth medium and directly affectsacidification rate measurements in milk(Spinnler and Corrieu, 1989; Famelart andMaubois, 1988; Zourari et al, 1991).
Urease activity is of technological inter-est for 3 reasons: (i) it enables streptococ-cal numbers in mixed cultures with L bul-garicus (this lactobacillus has no ureaseactivity) to be followed by measuring theamount of CO2 produced (Spinnler et al,1987; Miller and Kandler, 1967a); (ii) NH3production affects the evaluation of strep-tococcal acidifying properties by pH meas-urements; (iii) this activity may also be in-volved in the stimulation of lactobacilli byCO2 produced by the streptococci (see dis-cussion below).
Besides its technological interest, thepresence of urease activity in S thermophi-lus also has taxonomie relevance. AI-though Farrow and Collins (1984) reportedthat S thermophilus does not degradeurea, the presence of an urease activityseems to characterize most strains exam-ined until now. It also characterizes oral

16 A Zourariet al
S salivarius strains (Sissons et al, 1989)but lactococci do not possess this activity(Miller and Kandler, 1967b). The absenceof urease activity in the 6 strains of S ther-mophilus studied by Bridge and Sneath(1983) could be due to the prolonged incu-bation time (5 days at 37 OC) beforeurease assays were carried out, since Juil-lard et al (1988) demonstrated that ureaseactivity dropped during advanced station-ary phase.
Oxygen and metabolism
Lactic acid bacteria are facultative anaer-obes with a preference for anaerobic con-ditions (Whittenbury, 1978). They cannotsynthesize porphyrins and consequentlythey do not synthesize cytochromes or cat-alase. Oxygen is sometimes used for theformation of hydrogen peroxide (H202)which is toxic for lactic acid bacteria whichdo not contain catalase to break it down.Some species possess protection mecha-nisms, eg a high content of manganesewhich is oxidized by O2- radicals in Lac-tobacillus plantarum (Archibald and Frido-vich, 1981).
L bulgaricus is among the least oxygen-tolerant lactic acid bacteria (Archibald andFridovich, 1981). lt can produce a verylarge amount of H202 which activates theLPS system of milk (Iactoperoxidase-hydrogen peroxide-thiocyanate system)and inhibits its own growth as weil as thegrowth of some other bacteria such asLactobacillus acidophilus, associated inthe manufacture of some fermented milkproducts (Gilliland and Speck, 1977).S thermophilus does not produce enoughH202 to activate the LPS system andtherefore inhibition occurs only when exog-enous H202 is added to milk (Premi andBottazzi, 1972; Guirguis and Hickey,1987). On the other hand, some strepto-
coccal strains can degrade H202, eg strainTS2 studied by Smart and Thomas (1987).Nevertheless, no further experiments havebeen performed and the c1eavingactivity ofthis strain was attributed by the authors toa mechanism independent of active metab-olism.
The differences generally observed be-tween strains regarding their tolerance toand utilization of oxygen are due to varia-bilities in their enzymatic systems. Ritcheyand Seeley (1976) detected no NADH-oxidase activity in 3 S thermophilusstrains, while many lactococcal strainspossess this enzyme. Nevertheless, an-other weil studied strain possesses this en-zyme as weil as NADH-peroxidase, super-oxide dismutase, pyruvate and lactatedehydrogenases (Smart and Thomas,1987).
Teraguchi et al (1987) compared theS thermophilus STH 450 strain which hasa high oxygen consumption, with the type-strain, ATCC 19258. Strain STH 450 hasNADH-oxidase, but no pyruvate oxidase,and very low NADH-peroxidase activity.These enzyme activities are much lower inthe type-strain. Strain STH 450 accumu-lates a-acetolactate, acetoin and diacetylas end-products of aerobic glucose metab-olism.
PARTICULAR ASPECTSOF YOGURT FERMENTATION
Interactions between yogurt bacterla
A positive interaction is generally observedbetween S thermophilus and L bulgaricusin mixed culture, leading to the stimulationof growth and acid production of both bac-teria compared to their single-strain cul-tures (Pette and Lolkema, 1950a; Bautistaet al, 1966; Accolas et al, 1977). In addi-

Metabolism and biochemistry of yogurt bacteria 17
tion, total proteolysis in mixed culture (ex-pressed in Jlg of released tyrosine per mlof culture) exceeds the sum of the valuesobtained by each strain alone (Rajagopaland Sandine, 1990). Mixed yogurt culturesmay also stimulate the production of somemetabolites such as acetaldehyde (Ham-dan et al, 1971; Bottazzi et al, 1973) andinfluence carbohydrate utilization. For in-stance, one L bulgaricus strain studiedwhich cannot use galactose in pure culturemetabolizes this sugar when it is associat-ed with one strain of S thermophi/us (Amo-roso et al, 1988, 1989). Contrary behaviorwas observed with one streptococcal strain(Oner and Erickson, 1986) indicating thatinteractions between yogurt bacteria arevery complex and are greatly dependenton the strains involved.
S thermophi/us does not possess sub-stantial extracellular proteolytic activity andthe amino acid and free peptide content ofmilk is not high enough to promote its fullgrowth. Lactobacillus proteases breakdown caseins and supply the streptococ-cus with amino acids and peptides (Petteand Lolkema, 1950b, Bautista et al, 1966;Shankar and Davies, 1978; Radke-Mitchelland Sandine, 1984).
The growth of L bulgaricus is stimulatedby a compound produced by S thermophi-lus, which seems to be formic acid (Gales-loot et al, 1968; Veringa et al, 1968; Acco-las et al, 1971; Shankar and Davies,1978). Higashio et al (1978) reported acombined stimulating effect of formic andpyruvic acids. Suzuki et al (1986) observedthat addition of formic acid to boiled milkprevents abnormal cell elongation in L bul-garicus (filamentous forms). Formic acidsynthesis from pyruvate is a limiting step inpurine synthesis and this explains the com-bined action of the 2 acids. When formateis lacking, ribonucleic acid (RNA) synthesisis depressed. Elongated cells contain lessRNA than normal cells while DNA contents
of both cells are almost equal. The com-bined growth of L bulgaricus and S thermo-phi/us has the same effect as the additionof formic acid. A better knowledge of path-ways and kinetics of formic acid productionby S thermophi/us is required to definitivelyestablish the stimulating effect of this acid,questioned by Juillard et al (1987).
Driessen et al (1982) observed thatstimulation of lactobacilli growth could alsoresult from an increased CO2 content inthe growth medium, during continuous cul-ture at constant pH. S thermophilus pro-duces a large quantity of CO2 from milkurea (Tinson et al, 1982b). In this way, itcan stimulate lactobaciiii, since the quanti-ty of CO2 dissolved in milk decreases afterheat treatment and so remaining CO2 istoo low to meet the requirements of thelactobacillus (Driessen et al, 1982). Car-bon dioxide might be involved in the syn-thesis of aspartic acid (Reiter and Oram,1962).
ln conclusion, interaction between yo-gurt bacteria is a good example of integrat-ed metabolism in a mixed culture of lacticacid bacteria, but our knowledge on thestimulation of L bulgaricus by S thermophi-lus is still incomplete.
Production of antimicrobial compoundsby yogurt bacteria
As mentioned above, there is generally asymbiotic relationship between yogurt bac-teria, but growth inhibition is sometimesobserved (Moon and Reinbold, 1974;Suzuki et al, 1982; Pereira Martins andLuchese, 1988). This should be taken intoaccount when selecting starters. Inhibitionmay be due to competition for one or morenutrients of the growth medium (Moon andReinbold, 1976) or to inhibitory compoundsproduced by the strains, such as bacterio-cins and inhibitory peptides (Pereira Mar-tins and Luchese, 1988).

18 A Zourariet al
Pulusani et al (1979) extracted at least3 fractions of '" 700 Da from milk culturedwith S thermophilus, which inhibited thegrowth of Pseudomonas, Bacil/us, E coli,Flavobacterium, Shigella, Salmonella andLactococcus strains. These fractions aremost likely aromatic amines released inthe growth medium since cells are free ofany antimicrobial activity. Glucose and lac-tose, but not sucrose, are essential for pro-duction of these compounds which arealso obtained in milk-free media, eg "soy-milk" (Rao and Pulusani, 1981).
Smaczny and Krârner (1984a) studied2 bacteriocins of 10-20 kDa produced by2 S thermophilus strains which inhibitedthe growth of strains of the same speciesand to a lesser extent of enterobacteria.These molecules are sensitive to severalproteases and to a lipase, suggesting thata Iipid component participates in their ac-tive sites.
An antimicrobial substance named "bul-garican" produced by an L bulgaricus strainhas been partially purified (Reddy and Sha-hani, 1971; Reddy et al, 1983). At neutral oracid pH, this heat-stable substance was ac-tive against several strains of Bacillus,Streptococcus, Staphylococcus, Sarcina,Pseudomonas, Escherichia and Serratiaspecies. Protonated carboxyl groups maybe important for the activity of this mole-cule. Spillmann et al (1978) could not con-firm, however, bulgarican production.
A newly isolated antibacterial substancefrom L bulgaricus, active against a Pseu-domonas tragi and a Staphylococcus aure-us strains has optimum pH values close to4. It is a di- or a tripeptide (molecular mass< 700 Da) which probably contains an aro-matie group (Abdel-Bar et al, 1987).
Knowledge of antibacterial substancesproduced by yogurt bacterla is still frag-mentary and the results are often contra-dictory. Available studies are few and give
only little information on the nature of thecompounds involved, their spectrum andmode of action. These antibacterial sub-stances often seem to be peptides. Moreextensive study is required for a better un-derstanding of interactions of yogurt bacte-ria with other lactic acid bacteria and withnon-Iactic cheese flora.
Bacteriophages of L bulgaricusand S thermophilus
Knowledge of the specifie bacteriophages(phages) of thermophilic lactic acid bacte-ria has been weil documented, primarily onand after 1980. Specifie phages may at-tack S thermophilus and L bulgaricusstrains during yogurt or cheese manufac-ture and seriously affect product quality.Moreover, even if phage attacks do not de-lay acidification during yogurt manufacture,they can lead to an important decrease inthe streptococci and to a lower flavourscore of the resulting yogurt (Stadhouderset al, 1988). Phage outbreaks are less fre-quent in modern yogurt production unitsthan in cheese factories because: (a) thehigh heat treatment of milk destroys phag-es present in raw milk; (b) aseptic process-ing and packaging may prevent contamina-tions by airborne bacteriophages, bacteria,yeasts or moulds; and (c) according toLawrence and Heap (1986), an increasingviscosity of the yogurt milk below pH 5.2and the fact that no whey is drained offduring yogurt manufacture could efficientlyreduce the spreading of phages in yogurtplants. Thus, phage attacks are due eitherto accidentai contaminations (Stadhouderset al, 1984) or to the presence of Iysogenicstarter bacteria which might be the majorsource of phages by two possible routes:(i) and temperate phage particles derivedfrom a Iysogenic strain may infect a sensi-tive (indicator) strain of the same starter;

Metabolism and biochemistry of yogurt bacteria 19
(ii) temperate phages may give rise to viru-lent mutants able to superinfect and lysethe parental Iysogenic and other relatedstrains. A typical example of the latter inci-dence has been weil documented for a Iy-sogenic strain of Lactobacillus casei subspcasei used in Japan for the industrial man-ufacture of a fermented milk beverage, Va-kult (Shimizu-Kadota and Sakurai, 1982;Shimizu-Kadota et al, 1985). Substitutionof a prophage-free strain for the Iysogenicstrain immediately stopped ail phage infec-tions in the different Vakult productionunits.
Bacteriophages of L bulgaricus are veryclosely related to those of L delbrueckiisubsp lactis (L lactis) but they are corn-pletely different from those of L helveticus,which is usually used in cheesemaking(Séchaud, 1990). Two surveys of L bulgari-eus and L lactis phages (Cluzel, 1986;Mata et al, 1986) led to the description of4 groups named a, b, c and d. Ali thesephages belong to the Siphoviridae familyof Matthews (1982) which corresponds toBradley's group B (Bradley, 1967). Thehead of these phages is isometric or pro-late, collars are frequent, and the tails of-ten have diverse terminal structures(pronged or plain plates, fibers) and occa-sionally harbour regularly spaced cross-bars. Figures 2 and 3 iIIustrate some ofthese typical morphologies. Group a is thelargest phage group (20 of the 26 phagesstudied by Cluzel, 1986) and includes tem-perate phages of both subspecies (8 of the20 phages of Cluzel's group a) along withIytic phages isolated in cheese plants oryogurt production units (12 of the 20 phag-es of Cluzel's group a). Ali phages of groupa share common molecular characteristics,serological properties, Iytic spectra and arespecifie towards both subspecies (typicalmorphology in figure 2). The close relation-ship between temperate and Iytic phagesof group a suggests that Iysogeny may be
15 _
_Iv
Fig 2. Morphology of bacteriophages which be-long to group a. Their host-ranges include L bul-garicus and L lactis strains. Phage 15possessesfragile cross-bars on its tail which may be easilyremoved during phage preparation. Coll ars arevisible at the head-tail junctions of phage 0448(triple collar) and phage Iv (single collar). Fibersare present at the tail tip of phages Iv and 0448.Bars = 100 nm; the magnification factor is thesa me for ail the pictures (micrographs fromINRA, Jouy-en-Josas, France).Morphologie de bactériophages du groupe a.Ces phages attaquent indifféremment un certainnombre de souches de L bulgaricus et L lactis.Le phage 15possède des structures transver-sales fragiles tout le long de la queue, qui peu-vent se détacher facilement au cours de la pré-paration du phage. Des colliers sont visibles à lajonction de la tête et de la queue des phages0448 (triple collier) et Iv (collier simple). Desfibres sont présentes à l'extrémité distale de laqueue des phages Iv et 0448. Barres = 100 nm;l'agrandissement est le même pour toutes lesimages (micrographies de /'INRA, Jouy-en-Josas, France).

20 A Zourari et al
0235, c
Fig 3. Morphology of bacteriophages which be-long to groups b, c and d. Phages of group b,such as c31 and c5, have a host-range restric-ted to L bulgaricus strains whereas phages0235 (group c) and 0252 (group d) have host-ranges similar to those of group a phages.Phages c31 and c5 (groupe b) possess a non-contractile tail fitted with a pronçed- base plateand at least one fiber. Tail tip of prolate-headedtemperate phage 0235 (group c) exhibits asmall base plate and a fiber with 2 swellings.Isometric-headed temperate phage 0252 (groupd) possasses a fairly long tail with saveral fibers.Bars = 100 nm; the magnification factor is thesame for ail the pictures (phages c31, 0235 and0252: micrographs from INRA; phage c5: micro-graphs from INRNETH-Zürich).Morphologie des phages des groupes b, c et d.Les phages du groupe b, tels que c31 et c5, ontun spectre d'hôtes limité à quelques souches deL bulgaricus, tandis que les phages 0235(groupe c) et 0252 (groupe d) ont des spectresd'hôtes similaires à ceux des phages du groupea. Les phages c31 et c5 (groupe b) possèdentune queue non contractile pourvue d'une plaquebasale dentée et d'au moins une fibre. L'extrémi-té distale de la queue du phage tempéré à têteallongée 0235 (groupe c) porte une petite pla-que basale et une fibre avec deux renflements.
a determining factor in the occurrence andspread of these phages. Each of thegroups c and d contains a single represen-tative temperate phage of L lactis. These 2phages have Iytic spectra similar to thoseof group a phages and consequently caninfect some L lactis as weil as L bulgaricusstrains. However, no morphological (fig 3),molecular or serological similarities be-tween them or with group a phages havebeen observed. Their low incidence maylimit their importance in technology.
Group b is composed of 4 closely relat-ed virulent phages. They seem .less wide-spread than group a phages and attackonly L bulgaricus strains. They have differ-ent Iytic spectra and do not share molecu-lar and serological relationships with phag-es of the 3 other groups. SomeL bulgaricus strains sensitive to group bphages are Iysogenic and harbour pro-phages that belong to group a. Prophage-free (cured) bacterial clones become sen-sitive to the corresponding temperatephage and to ail other phages of groups a,c and d but are unexpectedly resistant togroup b phages. Inversion of phage sensi-tivity occurs along with changes in mor-phology of bacterial colonies (rough formfor cured clones/smooth form the parentalIysogenic strains; CIuzel et al, 1987). Thissuggests that these strains couId possessspecifie receptors for phages of ail fourgroups. In the presence of prophage, onlyreceptors for group b phages are function-al while those for groups a, c and d phag-es are masked or non-functional. It couldbe a Iysogenic conversion.
Le phage à tête isométrique 0252 (groupe d)possède une queue assez longue, munie deplusieurs fibres terminales. Barres = 100 nm;l'agrandissement est le même pour toutes lesimages (phages c31, 0235 et 0252 : microgra-phies de l'INRA; phage c5 : micrographies de/'INRA1ETH-Zürich).

Metabolism and biochemistry of yogurt bacteria 21
The large number of group a phagesunderlines the risk resulting from the pres-ence of Iysogenic bacteria in lactic starters.Therefore, the use of cured strains ap-pears to be a satisfactory solution, but re-quires strictly aseptic conditions duringstarter preparation and fermentation in or-der to prevent attacks by the widespreadgroup a phages.
Among phages of L bulgaricus andL lactis, only phage LL-H, isolated in Fin-land and active against L lactis strain LL23, has been extensively studied (seeAlatossava, 1987). Its morphology is repre-sentative of group a, and is closely relatedto that of phage Iv iIIustrated in figure 2.The average burst size reaches 102 phag-es per infected cell under optimal condi-tions. Phage LL-H contains 2 major struc-turai proteins (34 kDa for the head and 19kDa for the tail) and no less than 5 minorproteins. The DNA molecules are linear(34 kbp) with constant ends. A restrictionmap of LL-H DNA has been establishedand a library of restriction fragments hasbeen prepared by molecular cloninq into Ecoli. This had led to the localization of 5genes encoding structural proteins andphage Iysin. Phage LL-H DNA contains 2polycistronic c1usters of genes ie "early"genes which are involved inphaqe replica-tion and "late" genes which encode structu-rai proteins and phage Iysin. Bivalent cat-ions (Ca2+ or Mg2+) stabilize the coiledDNA into the capsid, improve adsorptionrate and control penetration efficiency ofphage DNA into the bacterial cells. A mo-lecular study of the LysA-encoding gene ofL bulgaricus temperate phage mv1 c1assi-fied in group a (Boizet et al, 1990), re-vealed a close relationship between ail Iy-sins of group a phages (including LL-H andmv1 enzymes). The lysA gene has a nu-c1eotide sequence of 585 bp correspond-ing to a 21,120 Da protein. Significant ho-mology with the N-terminal domain ofknown muramidases has been observed.
Recent studies in Germany and Francehave demonstrated the existence of an ap-parent homogeneity of the specifie phagesof S thermophilus at both the morphologi-cal and molecular levels (Krush et al,1987; Neve et al, 1989; Prévots et al,1989; Benbadis et al, 1990; Larbi et al,1990). These phages have been isolatedas Iytic and belong to the Siphoviridae fam-ily of Matthews (1982) or to Bradley'sgroup B1 (Bradley, 1967), as shown in fig-ure 4. They contain linear double-strandedDNA (34-44 kb) with complementary cohe-sive ends. Grouping of these phages hasbeen attempted on the basis of pro-tein patterns and DNA restriction profiles. Itis likely that most S thermophilus phagesstudied until now were derived from a corn-mon ancestor. As indicated by Mercenier(1990), their genomes have non-homolo-gous sequences flanked by homologousregions. It is consistent to apply the hy-pothesis of Botstein (1980) who postulatedthat the observed diversity of phages mightresult from recombination and exchange of"modules" ie genome fragments encodingfor particular biological functions.
The existence of Iysogenic strains inS thermophilus has been reported severaltimes since 1970 (Ciblis, 1970; Kurmann,1979, 1983; Smaczny and Krârner,1984b), but these studies remained incom-piete. Neve et al (1990) detected prophag-es in 2 strains by probing chromosomalDNA of 17 strains with genomic DNA of vir-ulent phages. Subsequently, the same au-thors demonstrated that these 2 strainswere inducible by mitomycin C (0.2-1Ilg.ml-1) and released mostly defectivephage particles. However, these phageswere able to relysogenize cured clonesand formed turbid plaques on agar medi-um. Presence of prophage led to a modifi-cation of sorne phenotypic traits of thestrains, ie homogeneous non-c1umpingcul-tures in broth instead of c1umpingin cured

22 A Zourari et al
strains, and autolysis at 45 oC probablydue to the thermoinducibility of prophageat elevated temperature.
A survey of S thermophilus phages iscurrently being carried out at the INRA la-boratory at Jouy-en-Josas (8 Fayard andM Haefliger, unpublished data). Ten of the120 strains examined were shown to be Iy-sogenic. Induction by mitomycin C gavetemperate phages active on one or severalindicator strains for 7 of them. Temperatephage DNA exhibited 5 different restrictionpatterns but shared homology. DNA ho-mology also existed between temperatephages and 52 phages isolated as Iytic fol-lowing acidification failures. Preliminary re-sults suggest that prophages might repre-sent a phage source for S thermophilusand the impact of Iysogeny in currentphage outbreaks should not be underesti-mated.
Starter suppliers and yogurt producersstill Jackbasic information about the originof disturbing phages and phage-bacteriarelationships. This explains why they oftenadopt a pragmatic approach to the phageproblem. Thus, an infected strain is re-placed by an insensitive strain, sometimesby a spontaneously resistant mutant, withsimilar technological properties. This is thecase for Australian yogurt manufacture(Hull, 1983). In day-te-day practice, phagecontrol requires experience and "know-how", but progress in the study of yogurtbacteria phages will certainly help to pre-vent phage attacks. For instance, improve-ment of phage resistance by genetic engi-neering might be the solution to phageproblems in the future (Sanders, 1989).
Genet/cs ofS thermophilusand L bulgaricus
Only few studies have been performed onthe genetics of S thermophilus, reviewed
by Mercenier and Lemoine (1989) andMercenier (1990). This is mainly due to im-portant methodological problems, but alsoto the fact that the major functions are car-ried by chromosomal genes making stud-ies more difficult. For this reason, many ef-forts have been made to adapt DNAtransfer methods to this species. The or-ganization and expression of genes in-volved in lactose and galactose metabo-lism were then studied by Herman andMcKay (1986), Poolman et al (1989,1990), and Lee et al (1990).
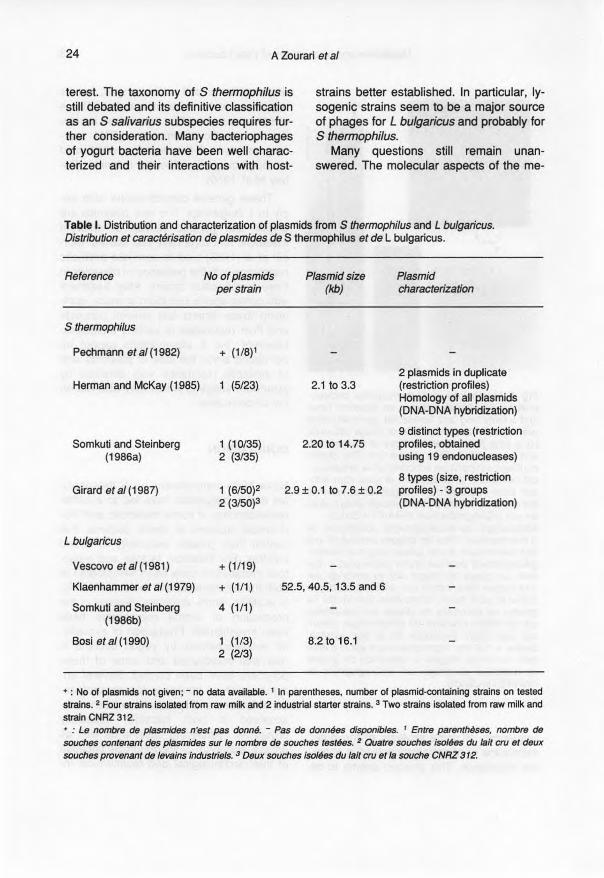
The plasmids of lactic acid bacteriaencode important characters for dairytechnology, eg carbohydrate metabolism,proteolytic activity, citrate utilization, pro-duction of antimicrobial compounds, bacte-riophage and antibiotic resistance, poly-saccharide production (McKay, 1983;Vedamuthu and Neville, 1986). S thermo-philus and L bulgaricus seem to be natural-Iy poor in plasmids (table 1).
Ali plasmids mentioned in table 1 areconsidered as being cryptic. Most of therepJicons isolated from S thermophiluswould be too small to carry a phenotypictrait (Mercenier, 1990). Until the present,the largest extrachromosomal element de-scribed is the 22.5-kb plasmid harbouredby strain IP6631, along with 2 other smallreplicons (Gavoille, 1989).
Only one endogenous plasmid of S ther-mophilus has been fully sequenced, ieplasmid pA33 (6.9 kb) from strain A033(Mercenier, 1990). Preliminary results indi-cate that this plasmid integrates and re-excises from chromosomal DNA. Its pres-ence or absence as an autonomously repli-cating extrachromosomal element providesthe host-cella with phenotypes which differby their colony morphology, cell chainlength, growth and acidification rates inmilk, antibiotic resistance and phage sensi-tivity. The nucleotide sequence of this plas-mid showed Iimited similarity with coding

Metabolism and biochemistry of yogurt bacteria
Fig 4. Morphology of S thermophilus bacterio-phages. Ali phages possess an isometric headand a fairly long and flexible tail, generally fittedwith a small base plate. Tails of phage c20 exhi-bit a long fiber, whereas those of phages schand sc are fitted with a shorter one. The clusterof phage sch particles adsorbed on a streptococ-cal cell debris is reminiscent of Iysis from with-out. Bars = 100 nm; the magnification factor isthe same for ail the pictures except phage elus-ter sch (micrographs from INRAIETH-Zürich).Morphologie de bactériophages spécifiques deS thermophilus. Tous les phages possèdent unetête isométrique et une queue longue et flexible,généralement pourvue d'une petite plaque ba-sale. La queue du phage c20 se prolonge parune longue fibre, tandis que celles des phagessch et sc sont munies d'une fibre plus courte. Legroupe de particules du phage sch adsorbéessur un débris cellulaire du streptocoque consti-tue une image évocatrice de la lyse externe.Barres = 100 nm; l'agrandissement est le mêmepour toutes les images, à l'exception du groupede particules du phage sch (micrographies de/'INRAIETH-Zür~ch).
sequences of sorne proteins of the outermembrane or of proteins involved in antibi-otic resistance. This plasmid seems to be
23
responsible for cell wall changes, but itsrole remains to be demonstrated. Theseobservations should be considered in rela-tion to specifie chromosomal DNA rear-rangements in S thermophilus, which leadto morphologically different colonies (Pé-bay et al, 1989).
These general considerations also ap-ply to L bulgaricus. The rare plasmids arecryptic and genetic studies of differentmetabolic functions have just started. Mor-elli et al (1983) tried to correlate antlblotlcresistance with the presence of plasmids inthree lactobacillus strains. After treatmentwith curing agents (ethidium bromide, acrif-lavin) these strains lost several plasmidsand their resistance to various antibiotics.However, the 2 observations cannot becorrelated since the loss of plasmids andof antibiotic resistance was obtained bypowerful mutagens which may also act onthe chromosome.
CONCLUSION
Since 1980 many studies on S thermophi-lus and L bulgaricus have led to a betterunderstanding of sorne metabolic and bio-chemical aspects of these bacteria thatcontrol their growth, especially in mixedcultures. For instance, lactose and galac-tose metabolism have been weil studied atboth the biochemical and molecular levels.ln addition, sorne enzymes involved in theproduction of aroma compounds havebeen investigated. Production of exocellu-lar polysaccharides by yogurt bacteria isnow weil established and sorne of thesepolymers have been studied. Several en-zymes such as aminopeptidases, proteas-es, esterases and lipases have been char-acterized in both bacteria, especiallyproteases hydrolyzing caseins as weil asurease activity of S thermophilus becauseof their technological and taxonomical in-
24 A Zourari et al
terest. The taxonomy of S thermophilus isstill debated and its definitive classificationas an S salivarius subspecies requires fur-ther consideration. Many bacteriophagesof yogurt bacteria have been weil charac-terized and their interactions with host-
strains better established. In particular, Iy-sogenic strains seem to be a major sourceof phages for L bulgaricus and probably forS thermophilus.
Many questions still remain unan-swered. The molecular aspects of the me-
Table 1. Distribution and characterization of plasmids from S thermophilus and L bulgaricus.Distribution et caractérisation de plasmides de S thermophilus et de L bulgaricus.
Reference No of plasmids Plasmid size Plasmidper strain (kb) characterization
S thermophilus
Pechmann et al (1982) + (1/8)1
2 plasmids in duplicateHerman and McKay (1985) 1 (5/23) 2.1 to 3.3 (restriction profiles)
Homology of ail plasmids(DNA-DNA hybridization)
9 distinct types (restrictionSomkuti and Steinberg 1 (10/35) 2.20 to 14.75 profiles, obtained
(1986a) 2 (3/35) using 19 endonucleases)
8 types (size, restrictionGirard et al (1987) 1 (6/50)2 2.9 ± 0.1 to 7.6 ± 0.2 profiles) - 3 groups
2 (3/50)3 (DNA-DNA hybridization)
L bulgaricus
Vescovo et al (1981) + (1/19)
Klaenhammer et al (1979) + (1/1) 52.5, 40.5, 13.5 and 6
Somkuti and Steinberg 4 (1/1)(1986b)
Sosi et al (1990) 1 (1/3) 8.2 to 16.12 (213)
+ : No of plasmids not 9iven; - no data available. 1 ln parentheses, number of plasmid-containing strains on testedstrains. 2 Four strains isolated from raw milk and 2 industrial starter strains. 3 Two strains isolated from raw milk andstrain CNRZ 312.+ : Le nombre de plasmides n'est pas donné. - Pas de données disponibles. t Entre perentnëses, nombre desouches contenant des plasmides sur le nombre de souches testées. 2 Quatre souches isolées du lait cru et deuxsouches provenant de levains industriels. 3 Deux souches isolées du lait cru et la souche CNRZ 312.

Metabolism and biochemistry of yogurt bacteria 25
tabolism of sucrose and other carbohy-drates should be better studied. This isparticularly true for L bu/garicus, sincemetabolic studies are less advanced thanin S thermophi/us. Volatile compounds pro-duced by S thermophilus and L bu/garicusremain to be identified because of their im-portance for the organoleptic properties ofyogurt. The study of the different biosyn-thesis pathways of acetaldehyde and othervolatile compounds, especially at the mo-lecular level, is also necessary. Further re-search is needed in the field of exopoly-saccharide production. In particular thegenetics of the metabolic pathways (en-zymes involved, and their regulation) re-quire investigation in order to control andstabilize polysaccharide production in milk.The nutritional and possible therapeuticroles of these polymers remain to be con-firmed.
ln mixed cultures, the growth of L bu/-garicus and S thermophilus is not yet fullycontrollable. Even if the interactions ofthese bacteria are weil established, manyaspects are still unknown. In particular, thereal influence of associated growth on sec-ondary metabolic activities has not beenstudied sufficiently. In addition, the charac-terization of inhibitory substances whichmay be produced would be very useful (a)for a better understanding of the relation-ship between the yogurt bacteria, and (b)for their action on other micro-organismsassociated with yogurt bacteria for· themanufacture of several fermented milkproducts.
Finally, progress in genetic studiesshould contribute to better knowledge onthe yogurt bacteria and their growth andactivity in milk.
ACKNOWLEDGMENTS
The authors are grateful to K Rérat (INRA, Ser-vice de Traduction UCD-CRJ, Jouy-en-Josas,France) who translated this paper into Englishand to J Cerning for helpful suggestions and dis-cussions during preparation of the English ver-sion of this manuscript.
REFERENCES
Abdel-Bar N, Harris ND, Rili RL (1987) Purifica-tion and properties of an antimicrobialsubstance produced by Laetobaeillus bulgari-eus. J Food Sei 52,411-415
Accolas JP, Veaux M, Auclair J (1971) Étudedes interactions entre diverses bactéries lac-tiques thermophiles et mésophiles, en rela-tion avec la fabrication des fromages à pâtecuite. Lait 51, 249-272
Accolas JP, Bloquel R, Didienne R, Régnier J(1977) Propriétés acidifiantes des bactérieslactiques thermophiles en relation avec lafabrication du yoghourt. Lait 57, 1-23
Akpemado KM, Bracquart PA (1983) Uptake ofbranched-chain amino acids by Streptococ-eus thermophilus. Appl Environ Mierobiol 45,136-140
Alatossava T (1987) Molecular biology of Lae-tobaeillus laetis bacteriophage LL-H. ActaUniv Ouluensis Ser A 191, 1-65
Alm L (1982a) Effect of fermentation on L(+) andD(-) lactic acid in milk. J Dairy Sci 65, 515-520
Alm L (1982b) Effect of fermentation on milk fatof Swedish fermented milk products. J DairySei 65, 521-530
Alm L (1982c) Effect of fermentation on proteinsof Swedish fermented milk products. J DairySei 65, 1696-1704
Amoroso MJ, Manca de Nadra MC, Oliver G(1988) Glucose, galactose, fructose, lactoseand sucrose utilization by Lactobaeillus bul-

26 A Zourari et al
garicus and Streptococcus thermophilus iso-lated from commercial yoghurt. Milchwis-senschatt 43, 626-631
Amoroso MJ, Manca de Nadra MC, Oliver G(1989) The growth and sugar utilization byLactobacillus delbrueckii ssp bulgaricus andStreptococcus salivarius ssp thermophilusisolated from market yogurt. Lait 69, 519-528
Anonymous (1989) Technologie des yaourts etlaits fermentés: les conséquences de la nou-velle réglementation. Tech Lait 1038, 22, 25-26
Archibald FS, Fridovich 1 (1981) Manganeseand defenses against oxygen toxicity in Lac-tobacillus plantarum. J Bacteriol 145, 442-451
Argyle PJ, Mathison GE, Chandan AC (1976)Production of cell-bound proteinase by Lac-tobacillus bulgaricus and its location in thebacterial cell. J Appl Bacterio/41 , 175-184
Atlan D, Laloi P, Portalier A (1989) Isolation andcharacterization of aminopeptidase-deficientLactobacillus bulgaricus mutants. Appl Envi-ron Microbio/55, 1717-1723
Atlan D, Laloi P, Portalier A (1990) X-prolyl-dipeptidyl aminopeptidase of Lactobacillusdelbrueckii subsp bulgaricus: characteriza-tion of the enzyme and isolation of deficientmutants. Appl Environ Microbiol 56, 2174-2179
Bautista ES, Dahiya AS, Speck ML (1966) Iden-tification of compounds causing symbioticgrowth of Streptococcus thermophilus andLactobacillus bulgaricus in milk. J Dairy Res33,299-307
Béai C, Louvet P, Corrieu G (1989) Influence ofcontrolled pH and temperature on the growthand acidification of pure cultures of Strepto-coccus thermophilus (CNAZ] 404 and Lac-tobacillus bulgaricus (CNAZ] 398. Appl Mi-crobiol Biotechno/32, 148-154
Benbadis L, Faelen M, Sios P, Fazel A, Merce-nier A (1990) Characterization and compari-son of virulent bacteriophages of Streptococ-cus thermophilus isolated from yogurt. Bio-chimie 72, 855-862
Boizet B, Lahbib-Mansais Y, Dupont L, Aitzen-thaler P, Mata M (1990) Cloning, expressionand sequence analysis of an endolysin-encoding gene of Lactobacillus bulgaricusbacteriophage mv1. Gene 94, 61-67
Bosi F, Bottazzi V, Vescovo M, Scolari GL, Bat·tistotti B, Dellaglio F (1990) Lactic acid bacte-ria for Grana cheese production. Part 1: Tech-nological characterization of thermophilic rodlactic acid bacteria. Sci Tecn Latt-Casearia41,105-136
Botstein D (1980) A theory of modular evolutionfor bacteriophages. Ann NY Acad Sci 354,484-491
Bottazzi V, Vescovo M (1969) Carbonyl corn-pounds produced by yYoghurt bacteria. NethMilk DairyJ23, 71-78
Bottazzi V, Bianchi F (1986) Types of microcol-onies of lactic acid bacteria, formation of voidspaces and polysaccharides in yoghurt. SciTecn Latt-Casearia 37,297-315
Bottazzi V, Battistotti B, Montescani G (1973) ln-f1uence des souches seules et associées deL bulgaricus et Str thermophilus ainsi quedes traitements du lait sur la productiond'aldehyde acétique dans le yaourt. Lait 53,295-308
Bouillanne C, Desmazeaud MJ (1980) Étude dequelques caractères de souches de Strepto-coccus thermophilus utilisées en fabricationde yoghourt et proposition d'une méthode declassement. Lait 60, 458-473
Bouillanne C, Desmazeaud MJ (1981) Classe-ment de souches de Lactobacillus bulgaricusselon quelques caractères utilisés en fabrica-tion du yoghourt. Association avec Strepto-coccus thermophilus. Sci Aliments 1, 7-17
Bracquart P, Lorient D (1979) Effet des acidesaminés et peptides sur la croissance deStreptococcus thermophilus. III. Peptidescomportant Glu, His et Met. Milchwissens-chatt 34, 676-679
Bracquart P, Lorient D, Alais C (1978) Effet desacides aminés sur la croissance de Strepto-coccus thermophilus. II. Étude sur cinqsouches. Milchwissenschatt 33, 341-344
Bradley DE (1967) A review: ultrastructure ofbacteriophages and bacteriocins. BacteriolRev 33, 230-314
Bridge PD, Sneath PHA (1983) Numerical tax-onomy of Streptococcus. J Gen Microbiol129, 565-597
Cerning J (1990) Exocellular polysaccharidesproduced by lactic acid bacteria. FEMS Mi·crobiol Reva7, 113-130 .

Metabolism and biochemistry of yogurt bacteria 27
Cerning J, Bouillanne C, Desmazeaud MJ,Landon M (1986) Isolation and characteriza-tion of exocellular polysaccharide producedby Lactobacillus bulgaricus. Biotechnol Lett8,625-628
Cerning J, Bouillanne C, Desmazeaud MJ,Landon M (1988) Exocellular polysaccharideproduction by Streptococcus thermophilus.Biotechnol Lett 10, 255-260
Cerning J, Bouillanne C, Landon M, Desma-zeaud MJ (1990) Comparison of exocellularpolysaccharide production by thermophiliclactic acid bacteria. Sci Aliments 10,443-451
Chamba JF, Prost F (1989) Mesure de l'activitéacidifiante des bactéries lactiques thermo-philes utilisées pour la fabrication des fro-mages à pâte cuite. Lait 69, 417-431
Chandan RC, Argyle PJ, Mathison GE (1982)Action of Lactobacillus bulgaricus proteinasepreparations on milk proteins. J Dairy Sci 65,1408-1413
Ciblis E (1970) Characterization of a bacterio-phage of Streptococcus thermophilus. Zen-tralbl Bakteriol Parasitenkd Infektionskr Hyg2 Abt 125, 541-554
Cluzel PJ (1986) Classification de 26 bactério-phages de Lactobacillus bulgaricus et deLactobacillus lactis. Thèse Docteur-Ingénieur, INA-PG, Paris, France
Cluzel PJ, Sério J, Accolas JP (1987) Interac-tions of Lactobacillus bulgaricus temperatebacteriophage 0448 with host strains. ApplEnviron Microbio/53, 1850-1854
Colmin C, Pébay M, Simonet JM, Decaris B(1991) A species-specific DNA probe ofStreptococcus salivarius subsp thermophilusdetects strain restriction polymorphism.
. FEMS Microbiol Lett 81, 123-128Corrieu G, Spinnier HE, Picque D, Jomier Y
(1989) Procédé de mise en évidence et decontrôle de l'activité acidifiante d'agents defermentation dans des bains de fermentationet dispositif pour sa mise en œuvre. Pat FrNo 2629612, 6.10.1989
Deeth HC, Tamime AY (1981) Yogurt: nutritiveand therapeutic aspects. J Food Prot 44, 78-86
Degorce-Dumas R, Goursaud J, Leveau JY(1986) Analyse de composés volatils duyaourt par chromatographie en phase ga-
zeuse-espace de tête (headspace). Ind Agro-Alimentaires 8, 805-808
Dellaglio F, Bottazzi V, Trovatelli LD (1973) De-oxyribonucleic acid homology and base com-position in some thermophilic lactobacilli.J Gen Microbio174, 289-297
Delley M, Mollet B, Hottinger H (1990) DNAprobe for Lactobacillus delbrueckii. Appl En-viron Microbio/56, 1967-1970
De Moraes J, Chandan RC (1982) Factors influ-encing the production and activity of a Strep-tococcus thermophilùs lipase. J Food Sci 47,1579-1583
Desmazeaud MJ (1974) Propriétés générales etspécificité d'action d'une endopeptidaseneutre intracellulaire de Streptococcus ther-mophilus. Biochimie 56, 1173-1181
Desmazeaud MJ, Devoyod JJ (1970) Actionstimulante des microcoques caséolytiquessur les bactéries lactiques thermophiles.Mise en évidence de la nature peptidique dessubstances stimulantes. Ann Biol Anim Bio-chim Biophys 10, 413-430
Desmazeaud MJ, Hermier JH (1973) Effet defragments peptidiques du glucagon vis-à-visde la croissance de Streptococcus thermo-philus. Biochimie 55, 679-684
Desmazeaud MJ, Jugé M (1976) Caractérisa-tion de l'activité protéolytique et fractionne-ment des dipeptidases et des aminopeptida-ses de Streptococcus thermophilus. Lait 56,241-260
De Vos WM (1990) Disaccharide utilization inlactic acid bacteria. In: 6th Int Symp on Ge-ne tics of Industrial Microorganisms Proc, Vol1. Soc Fr Microbiologie, 447-457
Doco T, Wièruszeski JM, Fournet B, Carcano D,Ramos P, Loones A (1990) Structure of anexocellular polysaccharide produced byStreptococcus thermophilus. Carbohydr Res198,313-322
Driessen FM, Kingma F, Stadhouders J (1982)Evidence that Lactobacillus bulgaricus in yo-gurt is stimulated by carbon dioxide producedby Streptococcus thermophilus. Neth MilkDairy J 36, 135-144
Dumont JP, Adda J (1973) Méthode rapide[d'étude] des composés très. volatils del'arôme des produits laitiers. Application auyoghourt. Lait 53, 12-22

28 A Zourari et al
Dutta SM, Kuila RK, Ranganathan B (1973) Ef-fect of different heat treatments of milk onacid and flavour production by five singlestrain cultures. Milchwissenschaft 28, 231-233
El Soda M, Desmazeaud MJ (1982) Les pep-tide-hydrolases des lactobacilles du groupeThermobacterium. 1. Mise en évidence deces activités chez Lactobacillus helveticus, Lacidophilus, L lactis et L bulgaricus. Gan JMicrobio/28, 1181-1188
El Soda M, Abd El Wahab H, Ezzat N, Desma-zeaud MJ, Ismail A (1986) The esterolyticand Iipolytic activities of the lactobacilli. II.Detection of the esterase system of Lactoba-cillus helveticus, Lactobacillus bulgaricus,Lactobacillus lactis and Lactobacillus aci-dophilus. Lait 66, 431-443
Ezzat N, El Soda M, Bouillanne C, Zevaco C,Blanchard P (1985) Cell wall associated pro-teinases in Lactobacillus helveticus, Lactoba-cillus bulgaricus and Lactobacillus lactis.Milchwissenschaft 40, 140-143
Ezzat N, Zevaco C, El Soda M, Gripon JC(1987) Partial purification and characteriza-tion of a cell wall associated proteinase fromLactobacillus bulgaricus. Milchwissenchaft42,95-97
Famelart MH, Maubois JL (1988) Comparaisonde l'évolution de l'indice de réfraction et de laviscosité au cours de /a gélification lactiquedu lait. Lait 68, 1-12
Farrow JAE, Collins MD (1984) DNA base com-position, DNA-DNA homology and long-chain fatty acid studies on Streptococcusthermophilus and Streptococcus salivarius. JGen Microbio/130, 357-362
Feller E, Cescatti G, Seppi A, Avaneini A, Gia-comelli F, Bossi MG (1990) Yoghurt. Nutri-tional, microbiological and biochemical char-acteristics. Latte 15, 674-683
Fox PF (1989) The milk protein system. In: De-velopments in Dairy Ghemistry - 4. Function-al Milk Proteins (Fox PF, ed) Elsevier ApplSei, London, 1-53
Gales/oot TE, Hassing F, Veringa HA (1968)Symbiosis in yoghurt (1). Stimulation of Lac-tobacillus bulgaricus by a factor produced byStreptococcus thermophilus. Neth Milk DairyJ22,50-63
Garvie El (1978) Lactate dehydrogenases ofStreptococcus thermophilus. J Dairy Res 45,515-518
Garvie El, Farrow JAE (1981) Sub-divisionswithin the genus Streptococcus using deoxy-ribonucleic aeid / ribosomal ribonucleic acidhybridization. Zentralbl Bakteriol MikrobiolHyg 1Abt Orig G 2, 299-310
Gasser F (1970) Electrophoretic characteriza-tion of lactic dehydrogenases in the genusLactobacillus. J Gen Microbiol62, 223-239
Gavoille S (1989) Analyse et caractérisation del'ADN plasmidique de Streptococcus salivari-us subsp thermophilus souche IP6631. Mé-moire DEA, Univ Nancy l, France
Gilliland SE, Speck ML (1977) Instability of Lac-tobacillus acidophilus in yogurt. J Dairy Sci60,1394-1398
Giraffa G, Bergère JL (1987) Nature du ca-ractère épaississant de certaines souches deStreptococcus thermophilus: étude prélimi-naire. Lait 67, 285-298
Girard F, Lautier M, Novel G (1987) DNA-DNAhomology between plasmids from Strepto-coccus thermophilus. Lait 67, 537-544
Greenberg NA, Mahoney RR (1982) Productionand characterization of ~-galactosidase fromStreptococcus thermophilus. J Food Sci 47,1824-1828,1835
Groux M (1973a) Étude des composants de laflaveur du yoghourt. Lait 53, 146-153
Groux M (1973b) Kritische Betrachtung des heu-tigen Joghurt-Herstellung mit Berücksichtungdes Protein-Abbaues. Schweiz Milchztg 4,2-8
Guirguis N, Hickey MW (1987) Factors affectingthe performance of thermophilic starters. 2.Sensitivity to the lactoperoxidase system.Aust J Dairy Techno/42, 14-16
Gurr MI (1986) Nutritional aspects of fermentedmilk products. In: 22th Int Dairy Gongr E,T Le Hague, 641-655
Hamdam IY, Kunsman JE Jr, Deane DO (1971)Acetaldehyde production by combined yogurtcultures. J Dairy Sci 54, 1080-1082
Hardie JM (1986) Genus Streptocccus. In: Ber-gey's Manual of Systematic Bacteriology, Vol2 (Sneath PHA, Mair NS, Sharpe ME, HoltJG, eds) Williams & Wilkins, Baltimore, 1043-1070

Metabolism and biochemistry of yogurt bacteria 29
Hemme D, Nardi M, Wahl D (1981a) Propriétésdes lacticodéshydrogénases de Streptococ-cus thermophilus indépendantes du fructose1,6-diphosphate. Lait 61 , 1-18
Hemme DH, Schmal V, Auclair JE (1981 b) Ef-fect of the addition of extracts of thermophiliclactobacilli on acid production by Streptococ-eus thermophilus in milk. J Dairy Res 48,139-148
Herman RE, McKay LL (1985) Isolation and par-tial characterization of plasmid DNA fromStreptococcus thermophilus. Appl EnvironMicrobio/50, 1103-1106
Herman RE, McKay LL (1986) Cloning and ex-pression of the l3-o-galactosidase gene fromStreptococcus thermophilus in Escherichiacoli. Appl Environ Microbio/52, 45-50
Hickey MW, Hillier AJ, Jago GR (1986) Trans-port and metabolism of lactose, glucose andgalactose in homofermentative lactobacilli.Appl Environ Microbio/51 , 825-831
Higashio K, Kikuchi T, Furuichi E (1978) Symbi-ose entre Lactobacillus bulgaricus et Strepto-coccus thermophilus dans le yoghourt. 20thGongr Int Lait F, 522-523
Hull RR (1983) Factory-derived starter culturesfor the control of bacteriophage in cheesemanufacture. Aust J Dairy Techno/38, 149-154
Hutkins R, Morris HA, McKay LL (1985a) Galac-tose transport in Streptococcus thermophilus.Appl Environ Microbio/50, 772-776
Hutkins R, Morris HA, McKay LL (1985b) Galac-tokinase activity in Streptococcus thermophi-lus. Appl Environ Microbio/50, 777-780
Hutkins R, Halambeck SM, Morris HA (1986)Use of galactose-fermenting Streptococcusthermophilus in the manufacture of Swiss,Mozzarella and short-method Cheddarcheese. J Dairy Sci 69, 1-8
Hutkins RW, Morris HA (1987) Carbohydratemetabolism by Streptococcus thermophilus:a review. J Food Prot 50, 876-884
Hutkins RW, Ponne C (1991) Lactose uptakedriven by galactose efflux in Streptococcusthermophilus: evidence for a galactose-lactose antiporter. Appl Environ Microbio/57,941-944
Itoh T, Ohhashi M, Toba T, Adachi S (1980) Pur-ification and properties of l3-galactosidase
from Lactobacillus bulgaricus. Milchwissens-chatt 35, 593-597
Juillard V, Spinnler HE, Desmazeaud MJ, Boqui-en CY (1987) Phénomènes de coopération etd'inhibition entre les bactéries lactiques utili-sées en industrie laitière. Lait 67, 149-172
Juillard V, Desmazeaud MJ, Spin nier HE (1988)Mise en évidence d'une activité uréasiquechez Streptococcus thermophilus. Gan J Mi-crobio/34, 818-822
Kalantzopoulos G, Tsakalidou E, ManolopoulouE (1990) Proteinase, peptidase and esteraseactivities of cell-free extracts from wild strainsof Lactobacillus delbrueckii subsp bulgaricusand Streptococcus salivarius subsp thermo-phi/us isolated from Greek YOgurt' J DairyRes 57,593-601
Kandler 0, Weiss N (1986) Genus Lactobacil-lus. In: Bergey's Manual of Systematic Bacte-riology, Vol 2 (Sneath PHA, Mair NS, SharpeME, Holt JG, eds) Williams and Wilkins, Balti-more, MD 1209-1234
Kilpper-Bâlz R, Fischer G, Schleifer KH (1982)Nucleic acid hybridization of Group N andGroup D streptococci. Gurr Microbiol7, 245-250
Klaenhammer TR, Scott LF, Sutherland SM,Speck ML (1979) Plasmid DNA isolation fromLactobacillus acidophilus and Lactobacillusbulgaricus. Abstr Annu Meet Am Soc Microbi-0179, 132 (Dairy Sci Abstr 42, 1677)
Kondratenko MS, Saeva S8, Iwanova PD, Ban-kova N (1989) Neue Sâurewecker für bulga-rische 8auermilch. Dtsch Molk-Ztg 31, 978-980
Krusch U, Neve H, Luschei B, Teuber M (1987)Characterization of virulent bacteriophages ofStreptococcus salivarius subsp thermophi/usby host specificity and electron microscopy.Kiel Milchwirtsch Forschungsber 39, 155-167
Kunath P, Kandler 0 (1980) Der Gehalt an L(+)-und D(-)-Milchsaure in Joghurtprodukten.Milchwissenschatt 35, 470-473
Kurmann JA (1984) Aspects of the production offermented milks. Bull Fed Int Lait 179, 16-28
Kurmann JL (1979) Bakteriophagenbedingte8auerungsstërungen in Hankâsereine.Schweiz Mi/chwirtsch Forsch 8, 71-76
Kurmann JL (1983) Freisetzen von Bakterio-phagen bei fhermophllen Milchsaurebakterien

30 A Zourari et al
durch Penicillin, eine verbogene Ursachefür phagenbedingte Sauerungsstôrungen.Schweiz Milchwirtsch Forsch 12, 39-43
Laloi P (1989) Analyse biochimique, physiolo-gique et génétique de l'activité caséinoly-tique et des aminopeptidases de Lactobacil-lus bulgaricus. Thèse Doctorat, Univ Lyon l,France
Larbi D, Colmin C, Rousselle L, Decaris B, Si-monet JM (1990) Genetic and biologicalcharacterization of nine Streptococcus sali-varius subsp thermophilus bacteriophages.Lait 70, 107-116
Law BA (1981) The formation of aroma and üa-vour compounds in fermented dairy prod-ucts. Dairy Sci Abstr 43, 143-154
Lawrence RC, Heap HA (1986) The New Zea-land starter system. Bull Fed Int Lait 199, 14·20
Le Bras G, Garel JR (1991) Properties of D·lactate dehydrogenase from Lactobacillusbulgaricus: a possible different evolutionaryorigin for the D- and L-lactate dehydrogenas-es. FEMS Microbiol Lett79, 89-94
Lee BH, Robert N, Jacques C, Ricard L (1990)Cloning and expression of l3-galactosidasegene from Streptococcus thermophilus inSaccharomyces cerevisiae. Biotechnol Lett12,499-504
Lees GJ, Jago GR (1976a) Acetaldehyde: an in-termediate in the formation of ethanol fromglucose by lactic acid bacteria. J Dairy Res43,63-73
Lees GJ, Jago GR (1976b) Formation of acetal-dehyde from threonine by lactic acid bacte-ria. J Dairy Res 43, 75-83
Lees GJ, Jago GR (1977) Formation of acetal-dehyde from 2-deoxy-D-ribose-5-phosphatein lactic acid bacteria. J Dairy Res 44, 139-144
Lindsay RC, Day EA, Sandine WE (1965)Green f1avourdefect in lactic starter cultures.J Dairy Sci 48, 863-869
London J, Kline K (1973) Aldolase of lactic acidbacteria: a case history in the use of an en"zyme as an evolutionary marker. BacteriolRev37,453-478
Macura D, Townsley PM (1984) Scandinavianropy milk-Identification and characterization
of endogenous ropy lactic streptococci andtheir extracellular excretion. J Dairy Sci 67,735-744
Manca de Nadra MC, Raya RR, Pesce de RuizHoigado A, Olivier G (1987) Isolation andproperties of threonine aldolase of Lactoba-cillus bulgaricus YOP12. Milchwissenschaft42,92-94
Marranzini RM, Schmidt RH, Shireman RB, Mar-shall MR, Comell JA (1989) Effect of threo-nine and glycine concentrations on threoninealdolase activity of yogurt microorganismsduring growth in modified milk prepared byultrafiltration. J Dairy Sci 72, 1142-1148
Marshall VM, Cole WM (1983) Threonine aldo-lase and alcohol dehydrogenase activities inLactobacillus bulgaricus and Lactobacillusacidophilus and their contribution to f1avourproduction in fermented milks. J Dairy Res50,375-379
Mata M, Trautwetter A, Luthaud G, RitzenthalerP (1986) Thirteen virulent and temperatebacteriophages of Lactobacillus bulgaricusand Lactobacillus lactis belong to a singleDNA homology group. Appl Environ Microbiol52,812-818
Matthews REF (1982) Classification and nomen-clature of viruses. Fourth report of the Inter-national Committee on Taxonomy of Viruses.Intervirology 17, 1-200
McKay LL (1983) Functional properties of plas-mids in lactic streptococci. Antonie van Leeu-wenhoek 49, 259-274
Mercenier A (1990) Molecular genetics of Strep-tococcus thermophilus. FEMS Microbiol Rev87,61-78
Mercenier A, Lemoine Y (1989) Genetics ofStreptococcus thermophilus: a review.J Dairy Sci 72, 3444-3454
Meyer J, Jordi R (1987) Purification andcharacterization of X-prolyl-dipeptidyl-amino-peptidase from Lactobacillus lactis and fromStreptococcus thermophilus. J Dairy Sci 70,738-745
Meyer J, Howard D, Jordi R, Fürst M (1989) Lo-cation of proteolytic enzymes in Lactobacilluslactis and Streptococcus thermophilus andtheir influence on cheese ripening. Milchwis-senschaft 44, 678-681

Metabolism and biochemistry of yogurt bacteria 31
Miller l, Kandler 0 (1967a) EiweiBabbau undAnreicherung freier Aminosaueren durchMilchsaurebakterien in Milch. 1. DieVerânderunq des Stickstoff-Fraktionen.Milchwissenschaft 22, 150-159
Miller l, Kandler 0 (1967b) EiweiBabbau und An-reicherung freier Aminosaueren durchMilchsaurebakterien in Milch. III. Die Anrei-cherung von freien Aminosauren durchStreptobakterien und Streptokokken. Milch-wissenschaft 22, 608-615
Miller l, Martin H, Kandler 0 (1964) Das Ami-nosaurespektrum von Joghurt. Milchwissens-chaft 19, 18-25
Moon NJ, Reinbold GW (1974) Selection of ac-tive and compatible starters for yogurt. CultDairyProdJ9, 10 & 12
Moon NJ, Reinbold GW (1976) Commensalismand competition in mixed cultures of Lactoba-cillus bulgaricus and Streptococcus thermo-philus. J Milk Food Techno/39, 337-341
Morelli L, Vescovo M, Bottazzi V (1983) Plas-mids and antibiotic resistances in Lactobacil-lus helveticus and Lactobacillus bulgaricusisolated from natural whey cultures. Microbio-logica6,145-154
Nakajima H, Toyoda S, Toba T, Itoh T, Mukai T,Kitazawa H, Adachi S (1990) A novel phos-phopolysaccharide from slime-forming Lacto-coccus lactis subsp cremoris SBT 0495.J Dairy Sci73, 1472-1477
Neve H, Krusch U, Teuber M (1989) Classifica-tion of virulent bacteriophages of Streptococ-eus salivarius subsp thermophilus isolatedfrom yoghurt and Swiss-type cheese. ApplMicrobiol Biotechno/30, 624-629
Neve H, Krusch U, Teuber M (1990) Virulentand temperate bacteriophages of thermophil-ic lactic acid streptococci. 23th Congr Int Lait,Vol Il,363
Olsen RL (1989) Effects of polysaccharides onrennet coagulation of skim milk proteins.J Dairy Sci 72, 1695-1700
Oner MD, Erickson LE (1986) Anaerobic fer-mentation of Lactobacillus bulgaricus andStreptococcus thermophilus on 3% non-fatdry milk with pure and mixed culture. Biotech-nol Bioeng 28, 883-894
Orla-Jensen S (1919) The Lactic Acid Bacteria.AF Host and Son, eds, Copenhagen
Pébay M, Colmin C, Decaris B, Simonet JM(1989) Variations phénotypiques et réar-rangements génomiques chez Streptococcusthermophilus. 2nd Congr Soc Fr Micrabiol,127
Pechmann H, Geis A, Teuber M (1982) Plas-mides des streptocoques lactiques. 21stCongr Int Lait F, vol 1, book 2,352-353
Pereira Martins JF, Luchese RH (1988) The as-sessment of growth compatibility betweenstrains of Lactobacillus bulgaricus and Strep-tococcus thermophilus. Rev Inst LaticCândido Tostes (Brasil) 43,11-13
Pette JW, Lolkema H (1950a) Yoghurt. 1. Symbi-osis and antibiosis in mixed cultures of Lbbulgaricus and Se thermophilus. Neth MilkDairy J 4, 197-208
Pette JW, Lolkema H (1950b) Yoghurt. II.Grawth stimulating factors for Sc thermophi-lus. Neth Milk Dairy J 4, 209-224
Pette JW, Lolkema H (1950c) Yoghurt. III. Acidproduction and aroma formation in yoghurt.Neth Milk DairyJ4, 261-273
Poolman B, Royer TJ, Mainzer SE, Schmidt BF(1989) Lactose transport system of Strepto-coccus thermophilus: a hybrid protein withhomology to the melibiose carrier and en-zyme III of phosphoenolpyruvate-dependentphosphotransferase systems. J Bacteriol171, 244-253
Poolman B, Royer TJ, Mainzer SE, Schmidt BF(1990) Carbohydrate utilization in Streptococ-cus thermophilus: characterization of thegenes for aldolase 1-epimerase (mutarotase)and UDPglucose 4-epimerase. J Bacteriol172, 4037-4047
Premi L, Bottazzi V (1972) Hydrogen peroxideformation and hydrogen peroxide splitting ac-tivity in lactic acid bacteria. Milchwissens-chaft27,762-765
Prevots F, Relano P, Mata M, Ritzenthaler P(1989) Close relationship of virulent bacterio-phages of Streptococcus salivarius subspthermophilus at both the protein and the DNAlevels. J Gen Microbio/135, 3337-3344
Puhan Z (1988) Results of the questionnaire1785B "Fermented milks". Bull Fed Int Lait227,138-164
Pulusani SR, Rao DR, Sunki GR (1979) Antimi-crobial activity of lactic cultures: partial purifi-

32 A Zourari et al
cation and characterization of antimicrobialcompound(s) produced by Streptococcusthermophilus. J Food Sci 44, 575-578
Rabier D, Desmazeaud MJ (1973) Inventairedes différentes activités peptidasiques intra-cellulaires de Streptococcus thermophi/us.Purification et propriétés d'une dipeptide-hydrolase et d'une aminopeptidase. Biochim-ie 55, 389-404
Radke-Mitchell L, Sandine WE (1984) Associa-tive growth and differential enumeration ofStreptococcus thermophilus and Lactobacillusbu/garicus: a review. J Food Prot 47, 245-248
Ragout A, Pesee de Ruiz Hoigado A, Oliver G,Sineriz F (1989) Presence of an L(+)-Iactatedehydrogenase in ceIls of Lactobacillus de/-brueckii ssp bu/garicus. Biochimie 71, 639-644
Rajagopal SN, Sandine WE (1990) Associativegrowth and proteolysis of Streptococcus ther-mophilus and Lactobacillus bu/garicus inskim milk. J Dairy Sci73, 894-899
Ramana Rao MV, Dutta SM (1981) Purificationand properties of beta-galactosidase fromStreptococcus thermophilus. J Food Sci 46,1419-1423
Rao DR, Pulusani SR (1981) Effect of culturalconditions and media on the antimicrobial ac-tivity of Streptococcus thermophilus. J FoodSci 46, 630-632
Rasic J, Vucurovic N (1973) Untersuchung derfreien Fettsâuren in Joghurt aus Kuh-, Schaf-und Ziegenmilch. Milchwissenschaft 28, 220-222
Rasic J, Kurmann JA (1978) Yoghurt: ScientificGrounds, Techn%gy, Manufacture andPreparations. Rasic J, Kurmann JA, publ,Tech Dairy Publ House distributors, Copen-hagen, Denmark
Rasic J, Stojsavljevic T, Curcic R (1971a) Astudy on the amino acids of yoghurt. II. Ami-no acids content and biological value of theproteins of different kinds of yoghurt. Milch-wissenschaft 26, 219-224
Rasic J, Curcic R, Stojsavljevic T, Obradovic B(1971b) A study on the amino acids of yo-ghurt. III. Amino acids content and biologicalvalue of the proteins of yoghurt made fromgoat's milk. Milchwissenschaft 26, 496-499
Raya RR, Manca De Nadra MC, Pesee De RuizHoigado A, Oliver G (1986) Acetaldehydemetabolism in lactic acid bacteria. Milchwis-senschaft 41, 397-399
Reddy GV, Shahani KM (1971) Isolation of anantibiotic from Lactobacillus bu/garicus.J Dairy Sci 54,748
Reddy GV, Shahani KM, Friend BE, ChandanRC (1983) Natural antibiotic activity of Lac-tobacillus acidophilus and bu/garicus. III. Pro-duction and partial purification of bulgaricanfrom Lactobacillus bulgaricus. Cult DairyProd J 18,15-19
Reiter B, Oram JD (1962) Nutritional studies oncheese starters. 1. Vitamin and amino acid re-quirements of single strain starters. J DairyRes 29,63-77
Ritchey TW, Seeley HW Jr (1976) Distribution ofcytochrome-Iike respiration in streptococci.J Gen Microbio/93, 195-203
Rysstad G, Abrahamsen RK (1987) Formationof volatile aroma compounds and carbon di-oxide in yogurt starter grown in cows' andgoats' milk. J Dairy Res 54, 257-266
Rysstad G, Knutsen WJ, Abrahamsen RK(1990) Effect of threonine and glycine on ac-etaldehyde formation in goats' milk yogurt.J Dairy Res 57,401-411
Sanders ME (1989) Bacteriophage resistanceand its applications to yogurt flora. In: Yogurt:Nutritiona/ and Hea/th Properties (ChandanRC, ed) Nat Yogurt Assoc, USA, 57-67
Schellhaass SM (1983) Characterization of exo-cellular slime produced by bacterial startercultures used in the manufacture of ferment-ed dairy products. PhD Thesis, Univ Minne-sota, MN, USA
Schleifer KH, Kilpper-Balz R (1987) Molecularand chemotaxonomic approaches to the clas-sification of streptococci, enterococci and lac-tococci: a review. Syst Appl Microbio/10, 1-19
Schmidt BF, Adams RM, Requadt C, Power S,Mainzer SE (1989) Expression and nucleo-tide sequence of the Lactobacillus bu/garicus~-galactosidase gene cloned in Escherichiacoli. J Bacterio/171, 625-635
Séchaud L (1990) Caractérisation de 35 bactéri-ophages de Lactobacillus he/veticus. Thèsedoctorat, Univ Paris VII et XI - ENSIAAMassy, France

Metabolism and biochemistry of yogurt bacteria 33
Shahbal S, Hemme D, Desmazeaud M (1991)High cell wall-associated proteinase activityof sorne Streptococcus thermophilus strains(H-strains) correlated with a high acidificationrate in milk. Lait 71, 351-357
Shankar PA, Davies FL (1977) Amino aeid andpeptide utilization by Streptococcus thermo-phi/us in relation to yoghurt manufacture.J Appl Bacterio/43, viii
Shankar PA, Davies FL (1978) Relation entreStreptococcus thermophilus et Lactobacillusbulgaricus dans les levains du yoghourt. 20eCongr Int Lait F, 521-522
Shimizu-Kadota M, Sakurai T (1982) Prophagecuring in Lactobacillus casei by isolation of athermoinducible mutant. Appl Environ Micro-bio/43, 1284-1287
Shimizu-Kadota M, Kiwaki M, Hirokawa H, Tsu-chida N (1985) ISL 1: a new transposableelement in Lactobacillus casei. Mol Gen Gen-et 200, 193-198
Simonds J, Hansen PA, Lakshmanan S (1971)Deoxyribonucleic aeid hybridization amongstrains of lactobacilli. J Bacteriol 107, 382-384
Sissons CH, Loong PC, Hancock EM, CutressTW (1989) Electrophoretic analysis of ureas-es in Streptococcus salivarius and in saliva.Oral Microbiollmmuno/4, 211-218
Smaczny T, Krâmer J (1984a) Sâuer-ungsstërungen in der Joghurt-, Bioghurt- undBiogarde- Produktion, bedingt durch Bacteri-oeine und Bakteriophagen von Streptococcusthermophilus. 1. Verarbeitung und Charakteri-sierung der Bacteriocine. Dtsch Molk-Ztg105,460-461,464
Smaczny T, Krâmer J (1984b) Sâue-rungsstërungen in der Joghurt-, Bioghurt-und Biogarde- Produktion, bedingt durchBacterioeine und Bakteriophagen von Strep-tococcus thermophilus. II. Verbreitung undCharakterisierung der Bacteriophagen. DtschMolk-Ztg105,614-618
Smart JB, Thomas TD (1987) Effect of oxygenon lactose metabolism in lactic streptococci.Appl Environ Microbio/53, 533-541
Smart JB, Crow VL, Thomas TD (1985) Lactosehydrolysis in milk and whey using ~-galactosidase from Streptococcus thermophi-lus. N Z J Dairy Sci Techno/20, 43-56
Somkuti GA, Steinberg DH (1979) Adaptabilityof Streptococcus thermophi/us to lactose,glucose-and galactose. J Food Prot42, 881-882,887 -
Somkuti GA, Steinberg DH (1986a) Distributionand analysis of plasmids in Streptococcusthermophilus. J Ind Microbio/1, 157-163
Somkuti GA, Steinberg DH (1986b) Generalmethod for plasmid DNA isolation from ther-mophilic lactic aeid bacteria. J Biotechnol 3,323-332
Spillmann H, Puhan Z, Banhegyi M (1978) Anti-mikrobielle Aktivitât thermophiler Laktobazil-len. Milchwissenschaft 33, 148-153
Spinnier HE, Corrieu G (1989) Automatic meth-od to quantify starter activity based on pHmeasurement. J Dairy Res 56,755-764
Spinnler HE, Bouillanne C, Desmazeaud MJ,Corrieu G (1987) Measurement of the partialpressure of dissolved CO2 for estimating theconcentration of Streptococcus thermophilusin coculture with Lactobacillus bulgaricus.Appl Microbiol Biotechno/25, 464-470
Stadhouders J, Hassing F, Leenders GJM,Driessen FM (1984) Disturbance of acid pro-duction by bacteriophages in the manufac-ture of yogurt. Zuivelzicht 2, 40-43
Stadhouders J, Smalbrink L, Maessen-DamsmaG, Klompmaker T (1988) Phage preventionat the manufacture of yogurt. Voedingsmid-delentechno/ogie 21, 21·24
Suzuki l, Ando T, Fujita Y, Kitada T, Morichi T(1982) Symbiotic and antagonistic relation-ships in mixed cultures of Lactobacillus bul-garicus and Streptococcus thermophilus. JpnJ Zootech Sci 53, 161-169 (Dairy Sci Abstr44, W6246)
Suzuki l, Kato S, Kitada T, Yano N, Morichi T(1986) Growth of Lactobacillus bulgaricus inmilk. 1. Cell elongation and the role of formicaeid in boiled milk. J Dairy Sci 69, 311-320
Szylit 0, Nugon-Baudon L (1985) Lactobacil/uscontribution to the nutritional physiopathologyof the host. A review. Sci Aliments 5 (supplV),257-262
Tamime AY, Deeth HC (1980) Yogurt: technolo-gy and biochemistry. J Food Prot43, 939-977
Tamime AY, Robinson RK (1985) Yoghurt. Sci-ence and Techno/ogy. Pergamon Press, Ox-ford

34 A Zourari et al
Teggatz JA, Morris HA (1990) Changes in therheology and microstructure of ropy yogurtduring shearing. Food Struct9, 133-138
Teraguchi S, Ono J, Kiyosawa l, Okonogi S(1987) Oxygen uptake activity and aerobicmetabolism of Streptococcus thermophilusSTH450. J Dairy Sci 70, 514-523
Thomas TD, Crow VL (1983) Lactose and su-crose utilization by Streptococcus thermophi-lus. FEMS Microbiol Lett 17, 13-17
Thomas TD, Crow VL (1984) Selection of galac-tose-fermenting Streptococcus thermophilusin lactose-Iimited chemostat cultures. ApplEnviron Microbio/48, 186-191
Thompson J (1987) Sugar transport in the lacticacid bacteria. In: Sugar Transport and Me-tabolism in Gram-positive Bacteria (Reizèr J,Peterkofski A, eds) Halsted Press, John Wi-ley, New York, 13-38
Tinson W, Hillier AJ, Jago GR (1982a) Metabo-Iism of Streptococcus thermophilus. 1. Utili-zation of lactose, glucose and galactose.AustJ Dairy Techno/37, 8-13
Tinson W, Broome MC, Hillier AJ, Jago GR(1982b) Metabolism of Streptococcus ther-mophilus. 2. Production of CO2 and NH3from urea. Aust J Dairy Techno/37, 14-16
Turcic M, Rasic J, Canic V (1969) Influence ofStr thermophilus and Lb bulgaricus cultureon volatile acids content in the f1avourcom-ponents of yoghurt. Milchwissenschaft 24,277-281
Vedamuthu ER, Neville JM (1986) Involvementof a plasmid in production of ropiness (mu-coidness) in milk cultures by Streptococcuscremoris MS. Appl Environ Microbiol 51,677-682
Veringa HA, Galesloot TE, Davelaar H (1968)Symbiosis in yoghurt. (II). Isolation and iden-tification of a growth factor for Lactobacillusbulgaricus produced by Streptococcus ther-mophilus. Neth Milk Dairy J 22, 114-120
Vescovo M, Bottazzi V, Sarra PG, Dellaglio F(1981) Evidence of plasmid deoxyribonucleic
acid in Lactobacillus. Microbiologica 4, 413-419
Vescovo M, Scolari GL, Bottazzi V (1989) Plas-mid-encoded ropiness production in Lactoba-cil/us casei ssp casei. Biotechnol Lett 11,709-712
Weiss N, Schillinger U, KandJer0 (1983) Lac-tobacillus lactis, Lactobacillus leichmanii andLactobacillus bulgaricus, subjective syno-nyms of Lactobacillus delbrueckii, and de-scription of Lactobacillus delbrueckii subsplactis comb nov and Lactobacillus delbrueckiisubsp bulgaricus comb nov. Syst Appl Micro-biol4, 552-557
Whittenbury R (1978) Biochemical characteris-tics of Streptococcus species. In: Streptococ-ci (Skinner FA, Quesnel LB, eds) AcademiePress, New York, 51-69
Wilkins DW, Schmidt RH, Kennedy LB (1986a)Threonine aldolase activity in yogurt bacteriaas determined by headspace gas chromatog-raphy. J Agric Food Chem 34, 150-152
Wilkins DW, Schmidt RH, Shireman RB, SmithKL, Jezeski JJ (1986b) Evaluating acetalde-hyde synthesis from L-[14C(U))threonine byStreptococcus thermophilus and Lactobacil-lus bulgaricus. J Dairy Sci 69, 1219-1224
Zourari A (1991) Caractérisation de bactérieslactiques thermophiles isolées à partir deyaourts artisanaux grecs. Thèse DoctoratINA-PG, Paris-Grignon, France
Zourari A, Desmazeaud MJ (1991) Caractérisa-tion de bactéries lactiques thermophiles iso-lées de yaourts artisanaux grecs. II. Souchesde Lactobacillus delbrueckii subsp bulgaricuset cultures mixtes avec Streptococcus sali-varius subsp thermophilus. Lait 71, 463-482
Zourari A, Roger S, Chabanet C, DesmazeaudMJ (1991) Caractérisation de bactéries lac-tiques thermophiles isolées de yaourts artisa-naux grecs. 1. Souches de Streptococcus sal-ivarius subsp thermophilus. Lait 71, 445-461