Embed Size (px)
Citation preview

Food & Function
PAPER
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article OnlineView Journal | View Issue
aInstituto en Formacion de Nutricion Animal
Camino del Jueves s/n, 18100, Armilla, Gran
Fax: +34 958 572753; Tel: +34 958 572757bInstitut Polytechnique LaSalle Beauvais, BecCase Western Research University, Clevelan
Cite this: Food Funct., 2013, 4, 1032
Received 30th November 2012Accepted 3rd February 2013
DOI: 10.1039/c3fo30351a
www.rsc.org/foodfunction
1032 | Food Funct., 2013, 4, 1032–10
Metabolic transit of N3-carboxymethyl-lysine afterconsumption of AGEs from bread crust
Irene Roncero-Ramos,a Cristina Delgado-Andrade,*a Frederic J. Tessier,b
Celine Niquet-Leridon,b Christopher Strauch,c Vincent M. Monnierc
and Marıa Pilar Navarroa
Our aim was to investigate carboxymethyl-lysine (CML) intake and excretion after feeding rats with diets
containing advanced glycation end-products (AGEs) from bread crust (BC) or its soluble or insoluble
fractions, and to identify the factors responsible for the effects observed. CML in serum and different
tissues was measured to detect possible accumulations. For 88 days, weanling rats were fed with either
a control diet or one containing BC, or its soluble low molecular weight (LMW), soluble high molecular
weight (HMW) or insoluble fractions. In the last week of the assay, faeces and urine were collected daily
and stored as a 1 week pool. After sacrifice, blood was drawn to obtain serum and some organs were
removed. CML analysis was performed by HPLC/MS/MS in diets, faeces, urines, serum and tissues. Faecal
excretion of CML was strongly influenced by dietary CML levels and represents the major route of
excretion (i.e. 33.2%). However, the urinary elimination of CML was probably limited or saturated,
especially when more complex compounds were present in the diet. BC consumption increased CML in
the cardiac tissue (170 � 18 vs. 97 � 3 mmol per mol lysine for BC and control groups), which correlated
with the CML intake. The levels of this AGE in bone were unaffected by the dietary treatment, but in tail
tendons CML was greatly increased in the animals that consumed the BC diet (102 � 13 vs. 51 � 8 mmol
per mol lysine for BC and control groups, P ¼ 0.006), which was associated with the intake of soluble
LMW compounds present in BC. Despite the CML accumulation detected in different tissues, serum
levels of protein-bound CML were unchanged, indicating the importance of measuring the free CML in
this fluid as a real index of dietary CML.
Introduction
The Maillard reaction is one of the most important chemicalreactions taking place during thermal processing of food byfrying, roasting or baking. The compounds generated in thisreaction, called Maillard reaction products (MRPs), are largelyresponsible for the characteristic sensory attributes of thermallyprocessed food.1
The Maillard reaction, also referred to as “glycation”, occursin biological systems by the nonenzymatic reaction of reducingsugar and other a-carbonylic compounds with proteins.Advanced glycation end-products (AGEs) are a heterogeneousgroup of compounds which have been associated with pro-oxidative and pro-inammatory effects and with the progres-sion of diseases such as atherosclerosis, diabetes and chronicrenal insufficiency.2
, Estacion Experimental del Zaidın, CSIC,ada, Spain. E-mail: [email protected];
auvais, France
d, Ohio, USA
39
Although nutrient composition and the method and condi-tions of food preparation dramatically affect the nal MRPs,3 itis well-established that MRPs are widely consumed as part ofthe human diet,4,5 and particularly the early glycationcompounds, such as Amadori products.6 Several studies haveshown that dietary MRPs, or AGEs, can be partially absorbed,increasing their circulating level and contributing to theendogenous load of AGEs.7 According to Faist and Erbersdo-bler8 and Koschinsky et al.4 approximately 10–30% of ingestedMRPs are absorbed and transported to the bloodstream andonly one-third of the absorbed AGEs are eliminated in the urine.
Carboxymethyl-lysine (CML) was the rst AGE identied infoods and it is oen used as a marker of both dietary AGEs andtheir in vivo levels.9 The absorption, distribution and clearanceof dietary CML from different food matrices are still unclear,despite several studies carried out in recent years to betterunderstand its bioavailability and metabolic transit.10–12
Different food groups have been analysed to estimate theircontribution to dietary CML intake10,13 and it has been reportedthat bakery products are among the major sources of dietaryCML.14 In the study by Somoza et al.15 in which rats were feddiets containing 25% bread crust, the daily intake of CML was
This journal is ª The Royal Society of Chemistry 2013

Paper Food & Function
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
calculated to be 11 mg kg�1 body weight/day and urinaryexcretion of CML was greater than in the group fed with the low-AGE diet. In another study by the same authors, CML elimina-tion of 15–22% in faeces and 26–29% in urine was recordedaer the administration to rats of dietary CML at doses of 110 or310 mg kg�1 body weight.12 A direct relationship between CMLdietary intake and its urinary and faecal excretion has beenestablished in various studies. Thus, Delgado-Andrade et al.11 intheir trial using low and high MRPs diets consumed by healthyadolescents stated that CML elimination was greater in theadolescents who consumed the high MRPs diet. The total CMLexcretion was around 47%, and so an important dietary fractioncould be absorbed and accumulated in different tissues ormetabolized by the intestinal microbiota. However, this studydid not further examine plasma or tissue CML levels.
In fact, little is known about dietary CML and its accumu-lation in different tissues and plasma, where most of the studiesdid not describe if free or protein-bound CML was measured. Inthe above-mentioned study by Somoza et al.12 higher levels ofCML in plasma and kidney were found in rats that consumed adiet with 25% bread crust. Uribarri et al.16 also reported a highcorrelation of dietary AGE intake with serum CML in healthyadults. Serum CML seems to be dependent on the intake,although it is unclear whether the compound comes from directabsorption or is formed in vivo from the dietary intake of highlyreactive carbonyl compounds.10
The purpose of the present study is to investigate CML intakeand excretion aer long-term feeding of rats with diets con-taining commonly consumed AGEs obtained from bread crust.Special attention was paid to the soluble and insoluble fractionsfrom bread crust, differing in their molecular weights. Theywere isolated and studied separately to identify the factorsresponsible for possible effects. Protein-bound CML in serumand in different tissues was measured to detect the targetorgans of accumulation.
Materials and methodsExtraction of bread crust and its soluble and insolublefractions
The bread crust (BC) was supplied by a Spanish manufacturer ofcereal-derived food products. The attached bread crumbs wereremoved manually and the sample was then weighed, lyophi-lised, powdered and homogenised. A fraction of the BC was thenstored at �20 �C until diet formulation. In order to access theMRPs and melanoproteins, another considerable fraction wasisolated, via enzymatic hydrolysis with pronase E. The resultsobtained from previous studies by our research group indicatedthe most appropriate pronase E concentration and incubationtime.17 Briey, 125 g of bread crust were digested with 750 mL ofa 0.100 mg mL�1 pronase E solution (400 U mL�1 in 1 M phos-phate buffer, pH 8.2) in stoppered test recipients at 37 �C for 72 hin a water bath under shaking. The digestion process has beenmore extensively described in a study by Roncero-Ramos et al.18
The insoluble fraction obtained was then weighed, lyophilized,homogenised and stored at �20 �C until used for diet formula-tion. It comprised the 57.52% w/w from the initial bread crust.
This journal is ª The Royal Society of Chemistry 2013
The soluble fraction (the reminder 42.48%w/w) was subjected toultraltration as described in the above-mentioned study. Thefraction constituted of compounds with a molecular masshigher than 5 kDa was retained (57.14% w/w, the retentate, highmolecular weight, HMW) and the fraction containingcompounds with a mass of less than 5 kDa was ltered (42.86%w/w, the ltrate, low molecular weight, LMW). Both fractions(retentate and ltrate) were lyophilized, powdered and homo-genised, and stored at �20 �C until used for diet formulation.
Preparation of diets
The AIN-93G puried diet for laboratory rodents (Dyets Inc,Bethlehem, PA) was used as the control diet.19 The bread crustwas added to the AIN-93G diet to reach a nal concentration of10% w/w. This diet was named Bread Crust (BC). In order todetermine the factors responsible for the effects observed in thetrial, the LMW, HMW and Insoluble fractions were also indi-vidually added to the diet in the same proportion as they werepresent in 10% of the bread crust, which was calculated basedon the recovery of each fraction aer pronase E digestion. Thesediets were named LMW, HMW and Insoluble, respectively.Bread crust is a sodium source, and so, to maintain theconcentration of this element at adequate levels, these dietswere prepared by mixing appropriate proportions of AIN-93Gand low-sodium AIN-93G diets, respectively. Calcium carbonatewas added when necessary to reach the values present originallyin the AIN-93G diet.
The individual analysis of the different diets revealed nomodication of the overall nutrient composition, comparedwith the control diet (AIN-93G). The mean � SD content of themain nutrient of the diets was: moisture (%) 7.9 � 0.4; protein(g kg�1) 168.4 � 4.0; fat (g kg�1) 77.9 � 1.6 and sodium (g kg�1)1.3 � 0.1.
Biological assays
Thirty weanling Wistar rats weighing 40.16 � 0.90 g (mean �SE) were randomly distributed into ve groups (n ¼ 6) and eachgroup was assigned to one of the dietary treatments. Theanimals were individually housed in metabolic cages in anenvironmentally controlled room under standard conditions(temperature: 20–22 �C with a 12 h light–dark cycle and 55–70%humidity). The rats had ad libitum access to their diets anddemineralised water (Milli-Q Ultrapure Water System, MilliporeCorps., Bedford, MA, USA).
The duration of the experiment was selected with the aim toreach the adult age of the rats, when major effects would start tohappen due to the chronic consumption of test diets. Thisprolonged consumption ensured the excretion and depositionof CML in different tissues. The experiment involved a prelim-inary 81-day period during which solid food intake and bodyweight changes were monitored weekly, followed by a 7-dayperiod in which faeces and urine from each animal werecollected daily and stored separately as a 1-week pool. Thefaeces were weighed, lyophilised, powdered and then homoge-nised. The urine was collected in 0.5% HCl (v/v), ltered(Whatman Filter Paper no. 40, ashless, Whatman, England) and
Food Funct., 2013, 4, 1032–1039 | 1033

Food & Function Paper
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
diluted to an appropriate volume. On day 88, aer an overnightfast, the animals were anaesthetised with sodium pentobarbital(5 mg per 100 g of body weight) (Abbott Laboratories, Granada,Spain) and terminal exsanguination was performed by a can-nulation of the carotid artery. Blood was drawn to obtain serum,and the hearts, the right pelvic bones and the tail tendons wereremoved, weighed and frozen at �80 �C until CML analysis.
All management and experimental procedures carried out inthis study were in strict accordance with the current Europeanregulations (86/609 E.E.C.) regarding laboratory animals. TheBioethics Committee for Animal Experimentation at our insti-tution (EEZ-CSIC) approved the study protocol.
Sample preparation for CML analysis
For CML determination, faeces, urines and hearts werelyophilised and powdered, while diets, serum and tail tendonswere directly processed. Bones were powdered and homoge-nised. In all samples, a weight equivalent to 10 mg proteins wasused for the analytical CML determination. Total nitrogen indiets, urine and faeces was analysed by using a Kjeldahlprocedure with demineralization (Block Digestor Selecta S-509;J.P. Selecta, Barcelona, Spain), distillation units (Buchi Labo-ratoriums Technik AG, Flawil, Switzerland), and titration units(MetromAG, Herisau, Switzerland). In the rest of the lyophilizedsamples, the nitrogen content was measured by combustionusing a LECO FP528 nitrogen analyzer according to the Dumasmethod. The nitrogen values were converted to protein bymultiplication by a factor of 6.25.
For CML determination in serum, a step prior to the acidhydrolysis was carried out for protein precipitation using tri-chloroacetic acid (TCA).
Analytical techniques
The CML analysis in extracts of diets, faeces, urine, hearts andbones, as well as protein-boundCML in serum, was performed atthe Institut Polytechnique LaSalle Beauvais. All the determina-tions were done in triplicate, when possible using the methodrecently developed byNiquet-Leridon andTessier.20Each samplewas treated with sodium borohydride to stabilize the Amadoriproducts and to prevent their conversion to CML during the acidhydrolysis. A quantity of reduced sample, equivalent to 10 mgprotein, was dissolved in 5 mL of 6 M HCl and incubated at110 �C for 20 h. Three hundred microlitres of each acid hydro-lysate was dried under vacuum and reconstituted in 300 mL ofinternal standard containing 0.15 mg of (D2)-CML and 12.5 mg of(15N2)-lysine (dissolved in 20 mM nonauoropentanoic acid(NFPA)) prior to analysis by LC-MS/MS. Since these internalstandards are isotopes of CML and lysine they follow the sameionization as CML and lysine, which increases the precision andaccuracy of the measurements compared with previousstudies.21 Liquid chromatography coupled to linear ion traptandem mass spectrometry was used for the analysis of CML.The following instrumentation and criteria were used: SurveyorHPLC system coupled to an LTQ mass spectrometer working inits tandem operation mode (ThermoFisher Scientic, Courta-boeuf, France); thermostat, 10 �C; columnHypercarb, 100mm�
1034 | Food Funct., 2013, 4, 1032–1039
2.1 mm, 5 mm with a guard column Hypercarb, 10 mm �2.1 mm, 5 mm; injection volume, 10 mL; ow rate, 0.2 mL min�1;mobile phase, 20 mM NFPA in a water–acetonitrile gradient asfollows: linear increase of acetonitrile from 0 to 50%over 20min;ion source: electrospray ionization in positive mode andmultiple reaction monitoring with the specic transitions m/z205.0/130.0 and m/z 207.0/130.0 for CML and (D2)-CML,respectively, with a normalized collision energy of 37%. Lysineand its isotope were detected with the specic transitions m/z147.0/130.0 and m/z 149.0/131.0, respectively.
For reasons of project development, the CML analysis for tailtendons was carried out at Case Western Reserve University(Cleveland, OH). CML was determined in acid hydrolysates ofprocessed collagen tendon samples and derivatised as its tri-uoroacetyl methyl esters by selected ion monitoring gaschromatography GC/MS as previously described by Sell et al.22
Despite these data being obtained by a different analyticaltechnique they are valid since they were only used to comparetotal CML content in tail tendons between groups aerconsumption of different diets.
Statistical analysis
All data were statistically tested by one-way analysis of thevariance (ANOVA), followed by Duncan's test to compare meansthat showed a signicant variation (P < 0.05). Analyses wereperformed using Statgraphics Plus, version 5.1, 2001. Evalua-tion of the relationship between the different variables wascarried out by computing the relevant correlation coefficient(Pearson's linear correlation) at the P < 0.05 condence level.
Results and discussionCML intake and excretion
The total CML content (mean � S.D.) in the different diets wasas follows: 2.18 � 0.03; 3.99 � 0.06; 2.63 � 0.03; 2.65 � 0.06 and3.96 � 0.04 mg g�1 for the control, BC, LMW, HMW and Insol-uble diets, respectively. As expected, the control diet (AIN-93G)had a certain basal level of CML, as previously documented.23
All the experimental diets had a signicantly higher CMLcontent than the control diet (P < 0.05), especially the BC andInsoluble diets, which moreover had a higher content than didthe diets containing the soluble fractions of BC (P < 0.05).
The food consumption during the last week balance was14.2 � 0.5, 14.1 � 0.3, 13.9 � 0.3, 13.6 � 0.3 and 12.9 � 0.4 gday�1 for the control, BC, LMW, HMW and Insoluble groups,respectively.18 Although the food consumption was similar inthe BC, LMW and HMW groups and signicantly lower in theInsoluble group, compared with the control group, the abun-dance of CML in the BC-derived diets led to a higher CML dailyintake in these animals (Table 1). By feeding 8 week old rats witha diet containing 25% bread crust aromaMRPs, Sebekova et al.24
observed that food consumption increased when CML contentwas higher than in the standard diet. In our study, the highestvalues of CML consumption were recorded for the BC andInsoluble groups, suggesting that the insoluble form of protein-bound CML was probably the main form present in these diets.
This journal is ª The Royal Society of Chemistry 2013
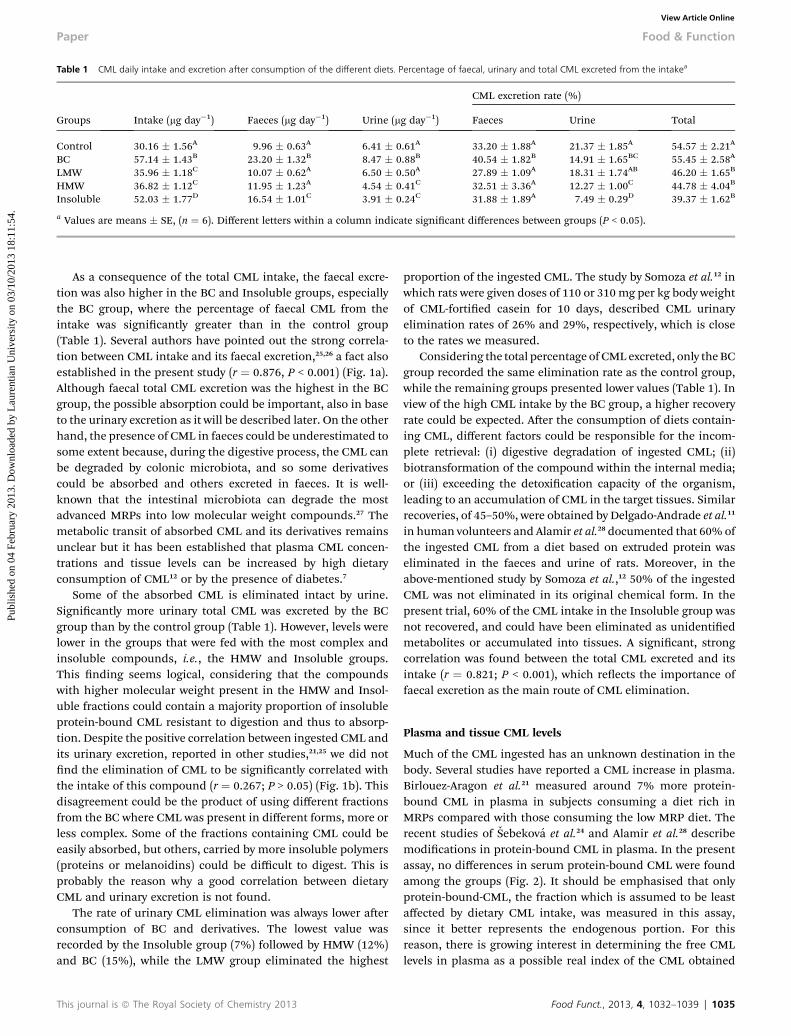
Table 1 CML daily intake and excretion after consumption of the different diets. Percentage of faecal, urinary and total CML excreted from the intakea
Groups Intake (mg day�1) Faeces (mg day�1) Urine (mg day�1)
CML excretion rate (%)
Faeces Urine Total
Control 30.16 � 1.56A 9.96 � 0.63A 6.41 � 0.61A 33.20 � 1.88A 21.37 � 1.85A 54.57 � 2.21A
BC 57.14 � 1.43B 23.20 � 1.32B 8.47 � 0.88B 40.54 � 1.82B 14.91 � 1.65BC 55.45 � 2.58A
LMW 35.96 � 1.18C 10.07 � 0.62A 6.50 � 0.50A 27.89 � 1.09A 18.31 � 1.74AB 46.20 � 1.65B
HMW 36.82 � 1.12C 11.95 � 1.23A 4.54 � 0.41C 32.51 � 3.36A 12.27 � 1.00C 44.78 � 4.04B
Insoluble 52.03 � 1.77D 16.54 � 1.01C 3.91 � 0.24C 31.88 � 1.89A 7.49 � 0.29D 39.37 � 1.62B
a Values are means � SE, (n ¼ 6). Different letters within a column indicate signicant differences between groups (P < 0.05).
Paper Food & Function
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
As a consequence of the total CML intake, the faecal excre-tion was also higher in the BC and Insoluble groups, especiallythe BC group, where the percentage of faecal CML from theintake was signicantly greater than in the control group(Table 1). Several authors have pointed out the strong correla-tion between CML intake and its faecal excretion,25,26 a fact alsoestablished in the present study (r ¼ 0.876, P < 0.001) (Fig. 1a).Although faecal total CML excretion was the highest in the BCgroup, the possible absorption could be important, also in baseto the urinary excretion as it will be described later. On the otherhand, the presence of CML in faeces could be underestimated tosome extent because, during the digestive process, the CML canbe degraded by colonic microbiota, and so some derivativescould be absorbed and others excreted in faeces. It is well-known that the intestinal microbiota can degrade the mostadvanced MRPs into low molecular weight compounds.27 Themetabolic transit of absorbed CML and its derivatives remainsunclear but it has been established that plasma CML concen-trations and tissue levels can be increased by high dietaryconsumption of CML12 or by the presence of diabetes.7
Some of the absorbed CML is eliminated intact by urine.Signicantly more urinary total CML was excreted by the BCgroup than by the control group (Table 1). However, levels werelower in the groups that were fed with the most complex andinsoluble compounds, i.e., the HMW and Insoluble groups.This nding seems logical, considering that the compoundswith higher molecular weight present in the HMW and Insol-uble fractions could contain a majority proportion of insolubleprotein-bound CML resistant to digestion and thus to absorp-tion. Despite the positive correlation between ingested CML andits urinary excretion, reported in other studies,21,25 we did notnd the elimination of CML to be signicantly correlated withthe intake of this compound (r ¼ 0.267; P > 0.05) (Fig. 1b). Thisdisagreement could be the product of using different fractionsfrom the BC where CML was present in different forms, more orless complex. Some of the fractions containing CML could beeasily absorbed, but others, carried by more insoluble polymers(proteins or melanoidins) could be difficult to digest. This isprobably the reason why a good correlation between dietaryCML and urinary excretion is not found.
The rate of urinary CML elimination was always lower aerconsumption of BC and derivatives. The lowest value wasrecorded by the Insoluble group (7%) followed by HMW (12%)and BC (15%), while the LMW group eliminated the highest
This journal is ª The Royal Society of Chemistry 2013
proportion of the ingested CML. The study by Somoza et al.12 inwhich rats were given doses of 110 or 310 mg per kg body weightof CML-fortied casein for 10 days, described CML urinaryelimination rates of 26% and 29%, respectively, which is closeto the rates we measured.
Considering the total percentage of CML excreted, only theBCgroup recorded the same elimination rate as the control group,while the remaining groups presented lower values (Table 1). Inview of the high CML intake by the BC group, a higher recoveryrate could be expected. Aer the consumption of diets contain-ing CML, different factors could be responsible for the incom-plete retrieval: (i) digestive degradation of ingested CML; (ii)biotransformation of the compound within the internal media;or (iii) exceeding the detoxication capacity of the organism,leading to an accumulation of CML in the target tissues. Similarrecoveries, of 45–50%, were obtained by Delgado-Andrade et al.11
in human volunteers and Alamir et al.28 documented that 60% ofthe ingested CML from a diet based on extruded protein waseliminated in the faeces and urine of rats. Moreover, in theabove-mentioned study by Somoza et al.,12 50% of the ingestedCML was not eliminated in its original chemical form. In thepresent trial, 60% of the CML intake in the Insoluble group wasnot recovered, and could have been eliminated as unidentiedmetabolites or accumulated into tissues. A signicant, strongcorrelation was found between the total CML excreted and itsintake (r ¼ 0.821; P < 0.001), which reects the importance offaecal excretion as the main route of CML elimination.
Plasma and tissue CML levels
Much of the CML ingested has an unknown destination in thebody. Several studies have reported a CML increase in plasma.Birlouez-Aragon et al.21 measured around 7% more protein-bound CML in plasma in subjects consuming a diet rich inMRPs compared with those consuming the low MRP diet. Therecent studies of Sebekova et al.24 and Alamir et al.28 describemodications in protein-bound CML in plasma. In the presentassay, no differences in serum protein-bound CML were foundamong the groups (Fig. 2). It should be emphasised that onlyprotein-bound-CML, the fraction which is assumed to be leastaffected by dietary CML intake, was measured in this assay,since it better represents the endogenous portion. For thisreason, there is growing interest in determining the free CMLlevels in plasma as a possible real index of the CML obtained
Food Funct., 2013, 4, 1032–1039 | 1035
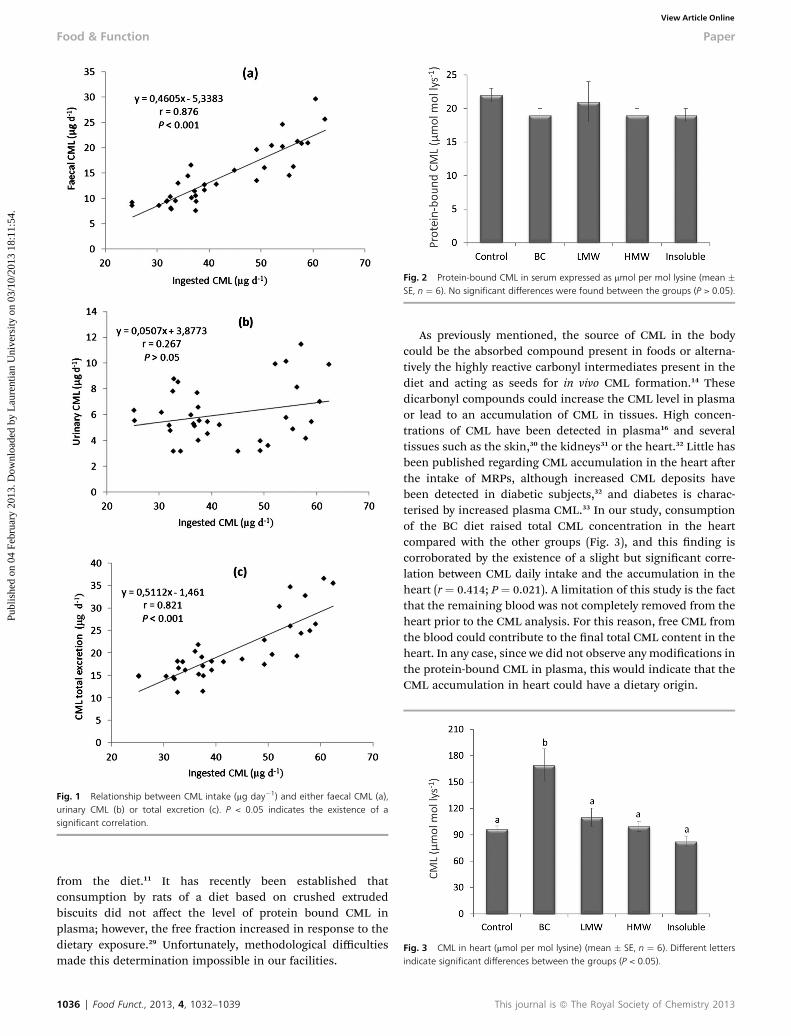
Fig. 1 Relationship between CML intake (mg day�1) and either faecal CML (a),urinary CML (b) or total excretion (c). P < 0.05 indicates the existence of asignificant correlation.
Fig. 2 Protein-bound CML in serum expressed as mmol per mol lysine (mean �SE, n ¼ 6). No significant differences were found between the groups (P > 0.05).
Fig. 3 CML in heart (mmol per mol lysine) (mean � SE, n ¼ 6). Different lettersindicate significant differences between the groups (P < 0.05).
Food & Function Paper
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
from the diet.11 It has recently been established thatconsumption by rats of a diet based on crushed extrudedbiscuits did not affect the level of protein bound CML inplasma; however, the free fraction increased in response to thedietary exposure.29 Unfortunately, methodological difficultiesmade this determination impossible in our facilities.
1036 | Food Funct., 2013, 4, 1032–1039
As previously mentioned, the source of CML in the bodycould be the absorbed compound present in foods or alterna-tively the highly reactive carbonyl intermediates present in thediet and acting as seeds for in vivo CML formation.14 Thesedicarbonyl compounds could increase the CML level in plasmaor lead to an accumulation of CML in tissues. High concen-trations of CML have been detected in plasma16 and severaltissues such as the skin,30 the kidneys31 or the heart.32 Little hasbeen published regarding CML accumulation in the heart aerthe intake of MRPs, although increased CML deposits havebeen detected in diabetic subjects,32 and diabetes is charac-terised by increased plasma CML.33 In our study, consumptionof the BC diet raised total CML concentration in the heartcompared with the other groups (Fig. 3), and this nding iscorroborated by the existence of a slight but signicant corre-lation between CML daily intake and the accumulation in theheart (r ¼ 0.414; P ¼ 0.021). A limitation of this study is the factthat the remaining blood was not completely removed from theheart prior to the CML analysis. For this reason, free CML fromthe blood could contribute to the nal total CML content in theheart. In any case, since we did not observe any modications inthe protein-bound CML in plasma, this would indicate that theCML accumulation in heart could have a dietary origin.
This journal is ª The Royal Society of Chemistry 2013

Fig. 4 CML in bone (a) and in tail tendon (b) (mmol per mol lysine) (mean � SE, n ¼ 6). Different letters indicate significant differences between the groups (P < 0.05).
Paper Food & Function
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
Regarding the total CML detected in the bones, no modi-cations were observed (Fig. 4a). Nevertheless, the total CML inthe tail tendons was signicantly increased in the BC and LMWgroups (Fig. 4b), which suggests that the low molecular weightcompounds of the BC group were possibly responsible for theeffect, since in the HMW and Insoluble groups no changes weredetected. Moreover, reactive carbonyls, expected to be presentin the BC and LMW fractions, could act as promoters for the invivo formation of CML. It has been established that tendoncollagen is more susceptible to be glycated than is bonecollagen,34 possibly because the mineral phase provides someprotection against glycation.35 Moreover, collagen type I turn-over is faster in bone than in other connective tissues such ascartilage or ligaments,36 and hence the greater CML accumu-lation in tendons could also be explained by its slower turnover.The deposition or formation of AGEs in bones has beendescribed,37 as well as in tendons,38 usually aer in vitro glyca-tion, during the advance of diabetes or even as an age-relatedaccumulation.39 However, to the best of our knowledge CMLdeposition in tendons related to simple dietary exposure hasnot been described previously. The only comparable study isthat carried out by Mikulikova et al.,40 using a diet with a highfructose content, which demonstrated a dietary-provokedincrease in pentosidine levels, another important AGE, incollagen-rich tissues. The origin of CML in bone and tendons isprobably different. In the case of tendons, there is the possi-bility of the direct deposition of the ingested CML, but this isnot the case with bone, where its special constitution andphysiology would prevent dietary CML deposition, and so itsin situ formation in a dicarbonyl compound environmentwould be more plausible.
In summary, it can be concluded that the faecal excretion ofBC-derived CML is highly inuenced by the dietary level andrepresents the major route of excretion. However, the elimina-tion rate in urine is probably saturated, with stronger effectswhen more complex compounds are present. In our study,surprisingly, BC consumption induced an increase in totalCML in the cardiac tissue, a fact that could be related to CMLintake. Levels of this AGE in bone were unaffected by dietaryfactors, but in tail tendons total CML was greatly increasedaer BC consumption, probably caused by the soluble LMW
This journal is ª The Royal Society of Chemistry 2013
compounds. In spite of the total CML accumulation detected indifferent tissues, protein-bound CML levels in serum wereunchanged, indicating the importance of measuring the freeCML in this uid as a possible real index of the CML of dietaryorigin. Likely the most signicant result of the present study isthe demonstration that the continuous consumption ofcommon dietary AGEs leads to accumulations of CML in thetarget tissues of healthy individuals, with special relevance inthe case of the heart tissue which represent the major origi-nality. The functional effects of this fact need to be consideredin depth in future studies.
Acknowledgements
This work was supported by a project of the Spanish Ministry ofScience and Innovation. The authors thank Grupo Siro, aSpanish manufacturer of cereal-derived food products, forsupplying the bread crust samples. The authors declare there isno conict of interest.
References
1 J. M. Ames, Control of the Maillard reaction in food systems,Trends Food Sci. Technol., 1990, 1, 150–154.
2 K. Sebekova and V. Somoza, Dietary advanced glycationendproducts (AGEs) and their health effects – PRO, Mol.Nutr. Food Res., 2007, 51, 1079–1084.
3 J. Uribarri, S. Woodruff, S. Goodman, W. Cai, X. Chen,R. Pyzik, A. Yong, G. E. Striker and H. Vlassara, Advancedglycation end products in foods and a practical guide totheir reduction in the diet, J. Am. Diet. Assoc., 2010, 110,911–916.e12.
4 T. Koschinsky, C.-J. He, T. Mitsuhashi, R. Bucala, C. Liu,C. Buenting, K. Heitmann and H. Vlassara, Orally absorbedreactive glycation products (glycotoxins): an environmentalrisk factor in diabetic nephropathy, Proc. Natl. Acad. Sci.U. S. A., 1997, 94, 6474–6479.
5 T. Goldberg, W. Cai, M. Peppa, V. Dardaine, B. S. Baliga,J. Uribarri and H. Vlassara, Advanced glycoxidation endproducts in commonly consumed foods, J. Am. Diet. Assoc.,2004, 104, 1287–1291.
Food Funct., 2013, 4, 1032–1039 | 1037

Food & Function Paper
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
6 C. Delgado-Andrade, I. Seiquer, A. Haro, R. Castellano andM. P. Navarro, Development of the Maillard reaction infoods cooked by different techniques. Intake of Maillard-derived compounds, Food Chem., 2010, 122, 145–153.
7 H. Vlassara, W. Cai, J. Crandall, T. Goldberg, R. Oberstein,V. Dardaine, M. Peppa and E. J. Rayeld, Inammatorymediators are induced by dietary glycotoxins, a major riskfactor for diabetic angiopathy, Proc. Natl. Acad. Sci. U. S. A.,2002, 99, 15596–15601.
8 V. Faist and H. F. Erbersdobler, Metabolic transit and in vivoeffects of melanoidins and precursor compounds derivingfrom the Maillard reaction, Ann. Nutr. Metab., 2001, 45, 1–12.
9 J. M. Ames, Determination of N3-(Carboxymethyl)lysine inFoods and Related Systems, Ann. N. Y. Acad. Sci., 2008,1126, 20–24.
10 F. Tessier and I. Birlouez-Aragon, Health effects of dietaryMaillard reaction products: the results of ICARE and otherstudies, Amino Acids, 2012, 42, 1119–1131.
11 C. Delgado-Andrade, F. J. Tessier, C. Niquet-Leridon,I. Seiquer and M. P. Navarro, Study of the urinary andfaecal excretion of N3-carboxymethyllysine in young humanvolunteers, Amino Acids, 2012, 43, 595–602.
12 V. Somoza, E. Wenzel, C. Weiss, I. Clawin-Radecker,N. Grubel and H. F. Erbersdobler, Dose-dependentutilisation of casein-linked lysinoalanine, N(epsilon)-fructoselysine and N(epsilon)-carboxymethyllysine in rats,Mol. Nutr. Food Res., 2006, 50, 833–841.
13 S. Assar, C. Moloney, M. Lima, R. Magee and J. Ames,Determination of N3-(carboxymethyl)lysine in food systemsby ultra performance liquid chromatography-massspectrometry, Amino Acids, 2009, 36, 317–326.
14 T. Henle, AGEs in foods: do they play a role in uremia?,Kidney Int., 2003, 63, S145–S147.
15 V. Somoza, M. Lindenmeier, T. Hofmann, O. Frank,H. F. Erbersdobler, J. W. Baynes, S. R. Thorpe, A. Heidland,H. Zill, S. Bek, J. Huber, T. Weigle, S. Scheidler, A. E. Buschand K. Sebekova, Dietary Bread Crust Advanced GlycationEnd Products Bind to the Receptor for AGEs in HEK-293Kidney Cells but Are Rapidly Excreted aer OralAdministration to Healthy and Subtotally NephrectomizedRats, Ann. N. Y. Acad. Sci., 2005, 1043, 492–500.
16 J. Uribarri, W. Cai, M. Peppa, S. Goodman, L. Ferrucci,G. Striker and H. Vlassara, Circulating glycotoxins anddietary advanced glyation endproducts: two links toinammatory response, oxidative stress, and aging,J. Gerontol., Ser. A, 2007, 72, 427–433.
17 C. Delgado-Andrade, J. A. Ruan-Henares and F. J. Morales,Procedure to analyse Maillard reaction-associateduorescence in cereal-based products, Czech J. Food Sci.,2008, 26, 339–346.
18 I. Roncero-Ramos, C. Delgado-Andrade, A. Haro, B. Ruiz-Roca, F. Morales and M. Navarro, Effects of dietary breadcrust Maillard reaction products on calcium and bonemetabolism in rats, Amino Acids, 2012, DOI: 10.1007/s00726-011-1160-3.
19 P. G. Reeves, F. H. Nielsen and G. C. Fahey, AIN-93 purieddiets for laboratory rodents: nal report of the American
1038 | Food Funct., 2013, 4, 1032–1039
Institute of Nutrition ad hoc writing committee on thereformulation of the AIN-76A rodent diet, J. Nutr., 1993,123, 1939–1951.
20 C. Niquet-Leridon and F. J. Tessier, Quantication ofN(epsilon)-carboxymethyl-lysine in selected chocolate-avoured drink mixes using high-performance liquidchromatography-linear ion trap tandem massspectrometry, Food Chem., 2011, 126, 655–663.
21 I. Birlouez-Aragon, G. Saavedra, F. J. Tessier, A. Galinier,L. Ait-Ameur, F. Lacoste, C.-N. Niamba, N. Alt, V. Somozaand J.-M. Lecerf, A diet based on high-heat-treated foodspromotes risk factors for diabetes mellitus andcardiovascular diseases, Am. J. Clin. Nutr., 2010, 91, 1220–1226.
22 D. R. Sell, C. M. Strauch, W. Shen and V. M. Monnier, 2-Aminoadipic acid is a marker of protein carbonyl oxidationin the aging human skin: effects of diabetes, renal failureand sepsis, Biochem. J., 2007, 404, 269–277.
23 S. Y. Tang and D. Vashishth, Non-enzymatic glycation altersmicrodamage formation in human cancellous bone, Bone,2010, 46, 148–154.
24 K. Sebekova, K. S. Klenovics, P. Boor, P. Celec, M. Behuliak,P. Schieberle, A. Heidland, M. Palkovits and V. Somoza,Behaviour and hormonal status in healthy rats on a dietrich in Maillard reaction products with or without solventextractable aroma compounds, Physiol. Behav., 2012, 105,693–701.
25 R. Liardon, D. De Weck-Gaudard, G. Philippossian andP. A. Finot, Identication of N.epsilon.-carboxymethylly-sine: a new Maillard reaction product in rat urine, J. Agric.Food Chem., 1987, 35, 427–431.
26 F. J. Tessier, C. Niquet, L. Rhaz, K. Hedhili, I. Seiquer,M. P. Navarro and C. Delgado-Andrade, in MaillardReaction Interface between Aging, Nutrition and Metabolism,ed. M. C. Thomas and J. M. Forbes, RSC Publishing,Cambridge, 2010, vol. 332, pp. 144–150.
27 V. Somoza, Five years of research on health risks andbenets of Maillard reaction products: an update, Mol.Nutr. Food Res., 2005, 49, 663–672.
28 I. Alamir, C. Niquet-Leridon, P. Jacolot, C. Rodriguez,M. Orosco, P. M. Anton and F. J. Tessier, Digestibility ofextruded proteins and metabolic transit of N(epsilon)-carboxymethyllysine in rats, Amino Acids, 2012, 16, 16.
29 C. Niquet-Leridon, I. Alamir, M. Orosco, P. M. Anton andF. J. Tessier, Digestibility of caseins and N3-carboxymethyl-lysine from extruded biscuits in rats, Book of abstracts ofthe 11� International Symposium on the Maillard reaction,2012, p. 66.
30 D. R. Sell and V. M. Monnier, Isolation, purication andpartial characterization of novel uorophores from aginghuman insoluble collagen-rich tissue, Connect. Tissue Res.,1989, 19, 77–92.
31 Y.-C. Li, S.-H. Tsai, S.-M. Chen, Y.-M. Chang, T.-C. Huang,Y.-P. Huang, C.-T. Chang and J.-A. Lee, Aristolochic acid-induced accumulation of methylglyoxal and N3-(carboxymethyl)lysine: An important and novel pathway inthe pathogenic mechanism for aristolochic acid
This journal is ª The Royal Society of Chemistry 2013

Paper Food & Function
Publ
ishe
d on
04
Febr
uary
201
3. D
ownl
oade
d by
Lau
rent
ian
Uni
vers
ity o
n 03
/10/
2013
18:
11:5
4.
View Article Online
nephropathy, Biochem. Biophys. Res. Commun., 2012, 423,832–837.
32 C. G. Schalkwijk, A. Baidoshvili, C. D. A. Stehouwer,V. W. M. van Hinsbergh and H. W. M. Niessen, Increasedaccumulation of the glycoxidation product N3-(carboxymethyl)lysine in hearts of diabetic patients:generation and characterisation of a monoclonal anti-CMLantibody, Biochim. Biophys. Acta, 2004, 1636, 82–89.
33 N. Jara, M. J. Leal, D. Bunout, S. Hirsch, G. Barrera, L. Leivaand M. P. de la Maza, Dietary intake increases serum levelsof carboxymethyl-lysine (CML) in diabetic patients, Nutr.Hosp., 2012, 27, 1272–1278.
34 G. K. Reddy, Glucose-mediated in vitro glycation modulatesbiomechanical integrity of the so tissues but not hardtissues, J. Orthop. Res., 2003, 21, 738–743.
35 D. Vashishth, G. J. Gibson, J. I. Khoury, M. B. Schaffler,J. Kimura and D. P. Fyhrie, Inuence of nonenzymaticglycation on biomechanical properties of cortical bone,Bone, 2001, 28, 195–201.
36 R. Eastell, A. Colwell, L. Hampton and J. Reeve, Biochemicalmarkers of bone resorption compared with estimates of
This journal is ª The Royal Society of Chemistry 2013
bone resorption from radiotracer kinetic studies inosteoporosis, J. Bone Miner. Res., 1997, 12, 59–65.
37 M. J. Silva, M. D. Brodt, M. A. Lynch, J. A. McKenzie,K. M. Tanouye, J. S. Nyman and X. Wang, Type 1 Diabetesin Young Rats Leads to Progressive Trabecular Bone Loss,Cessation of Cortical Bone Growth, and Diminished WholeBone Strength and Fatigue Life, J. Bone Miner. Res., 2009,24, 1618–1627.
38 D. R. Sell and V. M. Monnier, Age-related association of tailtendon break time with tissue pentosidine in DBA/2 vs.C57BL/6 mice: the effect of dietary restriction, J. Gerontol.,Ser. A, 1997, 52, B277–B284.
39 N. Verzijl, J. DeGroot, E. Oldehinkel, R. A. Bank, S. R. Thorpe,J. W. Baynes, M. T. Bayliss, J. W. Bijlsma, F. P. Lafeber andJ. M. Tekoppele, Age-related accumulation of Maillardreaction products in human articular cartilage collagen,Biochem. J., 2000, 2, 381–387.
40 K. Mikulikova, A. Eckhardt, J. Kunes, J. Zicha and I. Miksik,Advanced glycation end-product pentosidine accumulates invarious tissues of rats with high fructose intake, Physiol. Res.,2008, 57, 89–94.
Food Funct., 2013, 4, 1032–1039 | 1039





























