Embed Size (px)
Citation preview

Comp. Biochem. Physiol.. 1975, Vol. 52A, pp. 355 to 357. Pergamon Press. Printed in Great Britain
METABOLIC RATES OF SMALL HOMEOTHERMS IN A WATERBATH
J. EMIL MORHARDT 1., THEODORE H. FLEMING ;z, J. A. MCCRUM l, P. MOLT l AND C. MILLER 1
t Department of Biology, Washington University, St. Louis, MO, U.S.A. ZDepartment of Biology, University of Missouri, St. Louis, MO, U.S.A.
(Received 27 August 1974)
Abstract--l. The rates of heat loss of small birds (Junco hyemalis, Passer domesticus) and rodents (Rattus norvegicus albino, Spermophilus beldingi, Heteromys desmarestianus) immersed to the neck in water were 5 to 10 times as great as those in air.
2. Maximum metabolic rates were elicited at waterbath temperatures of 28 to 30°C. 3. Minimum metabolic rates were comparable to those from the literature of the same species in
air but maximum rates for R. norvegicus and P. domesticus were lower than those attained in air or in a helium-oxygen atmosphere.
INTRODUCTION
WHEN ANALYZING metabolic capacity of birds and mammals, it is often useful to know the maximum and minimum metabolic rates that these animals can achieve under standard conditions in the absence of activity--for example, when monitoring the time course of temperature acclimation. Increased meta- bolic rates can be induced in homeotherms by lower- ing the ambient temperatures, b u t many small homeotherms have sufficiently good fur or feather in- sulation that very low air temperatures are required to elicit maximum metabolic rates (cf. Hart, 1962). Energy transfer between animals and the environment can be increased by several methods such as physi- cally removing the fur or feathers, or diminishing in- sulation either by increasing convection or by increas- ing the conductivity of the surrounding medium. In some experiments it is advantageous to leave the fur or feathers intact, so that increased convection and conductance are often the method of choice. Musac- chia & Jacobs (1973) and Rosenmann & Morrison (1974) have recently reviewed the utility of more highly conductive gas mixtures such as He-O2 in place of air for increasing heat transfer from animals to the environment. Wetting animals with cold water is also a useful technique for increasing heat transfer (Giaja, 1925). In this study we have trained indivi- duals of several species of small birds and rodents to rest quietly, submerged to the neck in a waterbath, in order to increase heat loss. The rate of heat transfer was much greater in the waterbath than in air. We have shown that water temperatures higher than usual laboratory ambient temperatures are low enough to elicit maximum resting metabolic rates un- der these conditions, although the rates are somewhat lower than the maximum rates reported in the litera- ture from animals of the same species in air or in He--O 2 mixtures at lower temperatures.
We think this technique may have utility for moni- toring the time course of metabolic acclimation where it is inconvenient to provide low ambient tempera- tures.
* Present address: HDR Ecosciences, 804 Anacapa, Santa Barbara, CA 93101, U.S.A.
METHODS
The animals used included 7 Rattus norvegicus (albino rats, 100-300 g) from the National Laboratory Animal Co., St. Louis, Missouri, USA; 8 Spermophilus beldingi (B¢lding ground squirrels, 200-380 g) trapped near Bishop, Califor- nia, USA; 7 Heteromys desmarestianus (Desmarest's spiny pocket mouse, 84--88g) trapped in Costa Rica; 9 Junco hyemalis (slate-colored juncos, 20-21 g); and 3 Passer domesticus (house sparrows, 27-29 g), the latter two species trapped in St. Louis, Missouri, USA. The squirrels, rats and mice had been kept in an animal rogm at ~ 23°C for at least several months prior to experimentation. The experiments on sparrows and juncos "were performed within a few weeks of capture in the spring (March-April).
Core temperatures (To) were measured by introducing thermocouples through the rectum into the colons (near the diaphragm) of the mammals and through the cloacas into the large intestines of the birds. Waterbath tempera- tures (T,,b) and T, were recorded on a multipoint potenti- ometer. The animal holder (Fig. 1) consisted of a hardware cloth cylinder just large enough to accommodate the ani- mal comfortably with the head protruding through a hole in a plastic plate attached to one end of the cylinder. In early experiments the animals were held in place by rubber bands across the bottom of the cylinder that kept them from retreating. An improved design utilized a second plas- tic plate with a neck-sized hole in it. This second plate was split at the hole so that it could be slid into place around the neck and clamped loosely, securely restraining the animal. Most animals rapidly became accustomed to the holder and did not struggle.
Oatyglln Analyzer
a .
Fig. 1. The waterbath, and recording systems for tempera- ture and oxygen consumption.
355
356 J. EMIL MORHARDT et al.
° " ' - co . . r " " . . . .
¢r- 40 ~ . .
bolic r ~ . ~ .~._.,"" rn zo~-~-- - , . . . ,~ . ~ . "
laJ ~; I I I I
0 30 60 90 120
TIME, minules
Fig. 2. A typical experiment. Waterbath temperature (T,b) was controlled and core temperature T~) in the cloaca and metabolic rate were allowed to come to equilibrium at
each Twb.
A hood was lowered over the animal's head and air was pulled past at rates from 1 to 31/min depending on the size of the animal. Metabolic rate was measured as oxygen consumption by diverting a portion of this air stream to an oxygen analyzer (Beckman, E-2). Oxygen con- sumption was converted to Watts by assuming 5.6 W per I Oz consumed. Waterbath temperature was regulated to +0.1°C with a 400W immersion heater controlled by an on-off thermoregulator. Changes of waterbath temperature were accomplished by circulating hot (70°C) or cold (5°C) water through a heat-exchange coil in the bath.
RESULTS
A typical experiment is shown in Fig. 2. The Twb was initially adjusted to a level which resulted in minimum metabolic rate (Mm~,) and was sequentially lowered in a series of steps of 1-2°C each until a maximum metabolic rate ( M m J was reached and further decreases of T~b did not cause further in- creases of M. After each drop of Twb, T¢ and M were allowed to stabilize and M was recorded for 8--10 min. Figure 3 shows summary plots of the metabolic rates thus obtained as functions of T,b. Similar data in the
30~ Rots
HS -0 2 watorbath
I 0 ~ ( Rosenm~lnn ~ \ I R' MorrJs°n)-tJ~X \
/ I ] I I I I -20 0 20 40
~ - ~1 ~ Squirrels
2 0 - QIr
(Morhardt~ Jo - a ~ , . ~ _ ~ \
I I I I I I -20 0 20 40
Sparrows walorbat~
, o \ I '
2O
I I I i i I I I 1 -60 "40 -:~0 0 20 40
TEMPERATURE, *C
Fig. 3. Metabolic rates plotted against ambient tempera- tures. Data from the waterbath are from this study. The sources of data from animals in the air and in helium- oxygen mixtures are identified on the figure. Average
regressions are shown.
• B. norve,u!ratt - 0 S. bsldiJJ
~_~orfiolly defealhered • ~, desmoreMianus 10 - A Mus musculvl ~
_ D ~ domestlcu~ • d, h.yemellll
,~
I e l . 0 I00 200 300 4 0 0
BODY WEIGHT, grams
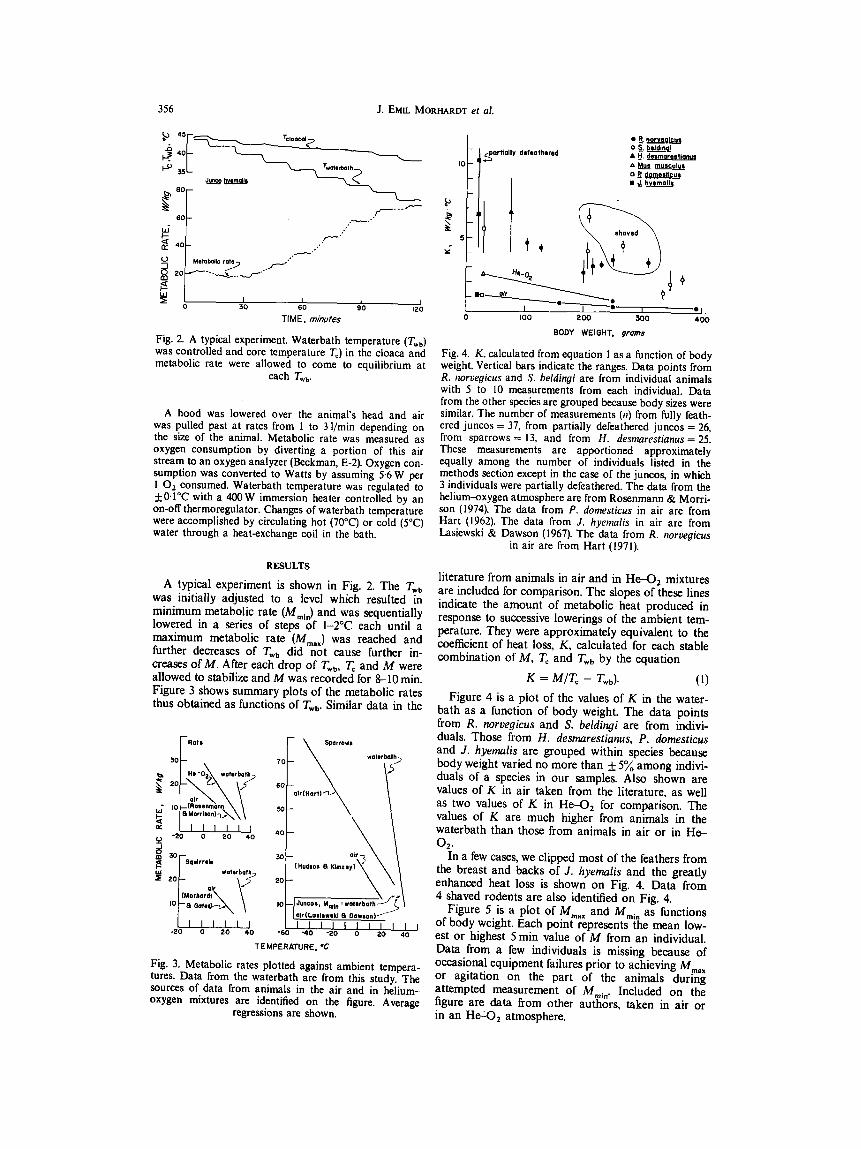
Fig, 4. K, calculated from equation 1 as a function of body weight. Vertical bars indicate the ranges. Data points from R. norvegicus and S. beldingi are from individual animals with 5 to 10 measurements from each individual. Data from the other species are grouped because body sizes were similar. The number of measurements (n) from fully feath- ered juncos = 37, from partially defeatbered juncos = 26, from sparrows = 13, and from H. desmarestianus= 25. These measurements are apportioned approximately equally among the number of individuals listed in the methods section except in the case of the juncos, in which 3 individuals were partially defeathered. The data from the helium-oxygen atmosphere are from Rosenmann & Morri- son (1974). The data from P. domesticus in air are from Hart (1962). The data from J. hyemalis in air are from Lasiewski & Dawson (1967). The data from R. norvegicus
in air are from Hart (1971).
literature from animals in air and in He-4)2 mixtures are included for comparison. The slopes of these lines indicate the amount of metabolic heat produced in response to successive lowerings of the ambient tem- perature. They were approximately equivalent to the coefficient of heat loss, K, calculated for each stable combination of M, Tc and T,b by the equation
K = M/T~ - Twb). (1)
Figure 4 is a plot of the values of K in the water- bath as a function of body weight. The data points from R. noroegicus and S. beldingi are from indivi- duals. Those from H. desmarestianus, P. domesticus and J. hyemalis are grouped within species because body weight varied no more than +_ 5 ~ among indivi- duals of a species in our samples. Also shown are values of K in air taken from the literature, as well as two values of K in He-O 2 for comparison. The values of K are much higher from animals in the waterbath than those from animals in air or in He- 02,
In a few cases, we clipped most of the feathers from the breast and backs of J. hyemalis and the greatly enhanced heat loss is shown on Fig. 4. Data from 4 shaved rodents are also identified on Fig. 4.
Figure 5 is a plot of M,..~ and Mmj" as functions of body weight. Each point represents the mean low- est or highest 5 min value of M from an individual. Data from a few individuals is missing because of occasional equipment failures prior to achieving M~, x or agitation on the part of the animals during attempted measurement of Mm~ .. Included on the figure are data from other authors, taken in air or in an HerO 2 atmosphere.

Small homeotherms in a waterbath 357
70 •
, 6O
50 "
IE • 0 4 0 .3 0 • m 3 0
.
20 -
,G ~
Mmox Mmln • v sparrows • o juncos • ~. mice
• o rate • o squirrels
• ¢ ( ~ H e - O 2 ® •
f . . ® • oo •
~ o % .o °
I00 2 0 0 300
BODY WEIGHT, groins
. . % ®
o¢+ [ 4OO
Fig. 5. Maximum and minimum metabolic rates of indivi- duals as functions of body weight. The circled data points are from other studies, all in air except the one on rats identified as being in a helium-oxygen atmosphere (Rosen- mann & Morrison, 1974). The data on 125-150g rats are from Giaja (1925); those on 380g rats from HCroux et al. (1959), and those on 390g rats, from Depocas et al. (1957). The M,m" data point from J. hyemalis is from Lasiewski & Dawson (1967) and that from P. domesticus is from Hudson & Kimzey (1966). The M=, x value for P.
domesticus is from Hart (1962).
DISCUSSION
Partial immersion is clearly a good technique for increasing rate of heat lo.ss and raising the metabolic rates of homeotherms at moderate ambient tempera- tures. The coefficient of heat loss, K, as used here, includes as its major component, heat transferred across the body wall (and fur or feathers) and lost to the waterbath by conduction and convection. However, K also includes heat loss from the head, ears and respiratory tract including heat lost by eva- poration. These routes of heat loss have not been par- titioned, but we think that they are likely to be quan- titatively similar whether the posterior part of the ani- mal is in the waterbath or air. In any case, the values of K do describe the increased rate of heat transfer in the waterbath over that in air or He--O2 whatever the actual route of heat loss.
The minimum metabolic rates achieved by this technique are comparable to those from other studies (Fig. 5) and are achieved much more rapidly than is usually the case in air. The maximum metabolic rates from some other studies have been substantially higher than those we observed, however. Values from Hart (1962) on sparrows and Rosenmann & Morrison (1974) on albino rats in particular are both much higher than we observed in the waterbath (Fig. 5). There are at least two possible reasons for the discre- pancies. To be sure that the differences are related to technique and not to physiological variation, it would be necessary to perform experiments in air,
He--O 2 and water on the same individuals, something that has yet to be done. The lower Mm, x values we observed could also result from a lowering of body temperature that usually accompanied 'a lowering of waterbath temperature. These experiments were deli- berately done at equilibrium values of T,b and Tc and it may be speculated that if Tc remained high, as it may have in Hart's (1962) and Rosenmann & Morrison's (1974) experiments, that higher values of M.~,x would have been obtained. This may be a moot point in partial-immersion experiments because in rats, at least, metabolic rate is inversely related to core temperature as well as being influenced by skin temperature. If core temperature were to remain high (~ 40°C) as can be arranged by suddenly lowering skin temperature, there would be no increase in M until T¢ fell (Morhardt et al., 1975).
In conclusion, partial immersion in a waterbath is a good method of increasing rates of heat transfer to achieve high, perhaps maximal, metabolic rates from homeotherms that will tolerate restraint. It has the added advantage, not explored here, of unambi- guous control and knowledge of skin temperatures if fur or feathers are removed prior to testing.
REFERENCES
DEPOCAS F., HART J. S, ~' HEROUX O. (1957) Energy meta- bolism of the white rat after acclimation to warm and cold environments. J. appl. Physiol. 10, 393-397.
GIAJA J. 0925) Le metabolisme de sommet et le quotient metabolique. Ann. physiol. Physicochim. Biol. 1, 596--627.
HART J. S. (1962) Seasonal acclimatization in four species of small wild birds. Physiol. Zool. 35, 224-236.
HART J. S. (1971) Rodents. In Comparative Physiology of Thermoregulation (Edited by WHITTOW G. C.), Vol. 2, Mammals pp. 1-149. Academic Press, New York.
HI~ROUX O., DEPOCAS F. 'g' HART J. S. (I959) Comparison between seasonal and thermal acclimation in white rats--I. Metabolic and insulative changes. Can. J. BiD- chem. Physiol. 37, 473-478.
HUDSON J. W. & KIMZEY S. L. (1966) Temperature regula- tion and metabolic rhythms in populations of the house sparrow, Passer domesticus. Camp. Biochem. Physiol. 17, 203-217.
LASlEWSK1 R. C. & DAWSON W. R. (1967) A re-examination of the relation between standard metabolic rate and body weight in birds. Condor 69, 13-23.
MORHARDT J. E., LATTANZI D. & MILLER C. (1975) Metabolic responses of unanesthetized rats to manipula- tion of skin temperature. Am. J. Physiol. 228. 575-580.
MORHARDT S. S. & GATES [). M. (1974) Energy-exchange analysis of the Belding squirrel and its habitat. Ecol. Monogr. 44, 17-44.
MUSACCHIA X. J. & JACOBS M. (1973) Helium-cold induced hypothermia in the white rat. Proc. Sac. exp. Biol. Med. 142, 734-739.
ROSENMANN M. & MORRISON P. 0974) Maximum oxygen consumption and heat loss facilitation in small homeo- therms by He-Or Am. J. Physiol. 226. 490-495.
Key Word Index--Heteromys desmarestianus; Junco hye- malis; Passer domesticus; oxygen consumption; metabolic rates; coefficient of heat loss.


















![rcin.org.pl · ACTA THERÍOLOGICA . VOL. XVI, 1: 1—21. BIAŁOWIEŻApril1971 ,A Piiotr POCZOPKO . Metabolic Level in Aduls t Homeotherms [With 5 Tables & 1 Fig.] Metabolic level](https://img.pdfslide.us/doc/110x75/60ad6556fa180c3dd80926bf/rcinorgpl-acta-therologica-vol-xvi-1-1a21-biaowieapril1971-a-piiotr.jpg)










