Embed Size (px)
Citation preview

Cytotherapy, 2013; 15: 280e291
Mesenchymal stromal cells isolated from children with systemicjuvenile idiopathic arthritis suppress innate and adaptive immuneresponses
FRISO G. J. CALKOEN, DANIELLE M. C. BRINKMAN, CARLY VERVAT,MONIQUE M. VAN OSTAIJEN-TEN DAM, REBECCA TEN CATE,MAARTEN J. D. VAN TOL & LYNNE M. BALL
Department of Paediatrics, Leiden University Medical Centre, Leiden, The Netherlands
AbstractBackground aims. Infusion of mesenchymal stromal cells (MSCs) has been reported to be an effective treatment modality foracute graft-versus-host disease, and MSCs have been considered for use in the treatment of patients with autoimmunediseases. Before contemplating clinical studies with MSCs in patients with systemic juvenile idiopathic arthritis (sJIA), theimmunomodulatory capacity of MSCs in this setting needs to be explored. A comparative analysis of bone marrow-derivedMSCs from children with sJIA and healthy pediatric controls was performed. Methods. MSCs were successfully expandedfrom 11 patients with sJIA and 10 controls. The phenotype, differentiation and immunomodulatory capacity of these MSCswere compared. The effect of immunosuppressive drugs onMSC function was also investigated. Results.MSCs from patientswith sJIA and controls showed no differences in their suppressive effect using control peripheral blood mononuclear cells.Furthermore, the suppression of the response of peripheral blood mononuclear cells from patients with sJIA by autologoussJIA MSCs and allogeneic control MSCs was comparable. The immunosuppressive effect of both groups of MSCs wasdiminished in the presence of indomethacin (P < 0.05). MSCs from patients with sJIA and controls suppressed interleukin-2-induced natural killer cell activation to a similar extent. In addition, MSCs of patients with sJIA and controls inhibited thedifferentiation of monocytes to dendritic cells. Conclusions. This is the first explorative study in a significant cohort of patientswith sJIA to evaluate the effect of MSCs on adaptive and innate immune responses. The comparable immunosuppressivecharacteristics of MSCs derived from patients with sJIA to age-matched controls support the potential use of patient-derivedMSCs in the treatment of sJIA.
Key Words: dendritic cells, indomethacin, juvenile idiopathic arthritis, mesenchymal stromal cells, natural killer cells, T lymphocytes
Introduction
Systemic juvenile idiopathic arthritis (sJIA) is anautoinflammatory disease of unknown etiology that ischaracterized by spiking fever, exanthema, anemia,hepatosplenomegaly and arthritis (1). The outcomeof the disease and response to therapy are unpredict-able. Improvements in treatment outcome with theuse of diverse biological therapies, as anti-interleukin(IL)-1 therapy, have been reported (2e9). Despitethese observations, the disease progresses and/orrequires additional therapy in 27% of children treatedwith etanercept (10). Moreover, patients treated withthese new drugs may experience adverse effects.Recent data relate the use of tumor necrosis factor(TNF)-blocking agents in patients with juvenileidiopathic arthritis (JIA) and other inflammatory
Correspondence: Lynne M. Ball, Department of Paediatrics, Leiden University [email protected]
(Received 10 July 2012; accepted 23 October 2012)
ISSN 1465-3249 Copyright � 2013, International Society for Cellular Therapy. Phttp://dx.doi.org/10.1016/j.jcyt.2012.10.017
diseases to the development of lymphoma and othercancers in children (11e13).
Previous studies in children with refractory JIAundergoing autologous hematopoietic stem celltransplantation following chemotherapy and T celldepletion show a progression-free survival of 52%after 5 years (14). However, high morbidity due tomacrophage activation syndrome, viral infections andtransplant-related mortality occurred (15). There-fore, new treatment modalities need to be exploredfor therapy-resistant patients.
Mesenchymal stromal cells (MSCs) are multi-potent non-hematopoietic stromal cells with anti-inflammatory and anti-proliferative capacities (16,17).MSCs can be expanded easily from different tissues,including bone marrow (BM), and remain genetically
edical Centre, PO Box 9600, 2300 RC Leiden, The Netherlands. E-mail:
ublished by Elsevier Inc. All rights reserved.

Mesenchymal stromal cells from patients with sJIA 281
stable even after numerous passages (18). No uniquemarker has been identified for the isolation of MSCs;however, specific criteria to identify MSC have beenaccepted (19,20).
T cell and natural killer (NK) cell proliferation,immunoglobulin production by B cells and dendriticcell (DC) maturation are suppressed by MSCs inin vitro experiments (21e23). The mechanismsunderlying these immunomodulatory functions arenot fully understood. However, experimental datasuggest that cell-cell contact and soluble factors mayboth be involved in an overall inhibition of theinduction of effector functions (16,24e29).
The profound immunomodulatory characteristicsof MSCmake them potential candidates for use in thetreatment of patients with inflammatory diseases.Clinical trials using allogeneic MSCs in the treatmentof corticosteroid-resistant acute graft-versus-hostdisease have shown promising results (30). Moreover,the feasibility and safety of autologous MSC infusionshave been shown in phase 1 clinical trials (31e35).However, the efficacy and safety of MSCs treatmenthave yet to be determined in larger cohort studies (36).
An aberrant suppressive capacity of MSCs frompatients with severe aplastic anemia was previouslyreported (37). In addition, functional aberrationsin synovium-derived MSCs from adults with rheu-matoid arthritis have been described, suggestingthat MSCs from patients with inflammatory diseasesmay not be comparable to those derived from healthydonors (38). Therefore, before contemplating clin-ical studies of autologous MSC infusions in childrenwith treatment-resistant sJIA, the immunomod-ulatory properties of the MSCs of these patientsneed to be characterized. To our knowledge, this isthe first study describing comparable characteristicsof MSCs from a relatively large cohort of childrenwith sJIA compared with age-matched healthycontrols (HC).
Methods
Expansion of MSCs
BM of children with sJIA (n ¼ 13) was used toinitiate MSC cultures according to a protocolapproved by a local ethical committee. Fresh BMwas harvested from five patients at the time of diag-nosis. To expand the study population, frozen bonemarrow mononuclear cells (BMMC) from patientswith sJIA (n ¼ 8) were used for MSC cultures.Classification of sJIA was according to guidelinesfrom the International League of Associations forRheumatology (1). Fresh BM of 10 pediatric donorsof an allogeneic hematopoietic stem cell graft wasused as HC. Parental and patient informed consentforms were signed for each patient and control.
After Ficoll separation, BMMC were plated ata density of 0.16� 106 cells/cm2 in polystyrene cultureflasks. Cells were cultured in Dulbecco’s modifiedEagle medium with GlutaMAX (Invitrogen, Paisley,UK) supplemented with 100 U/mL penicillin, 100 mg/mL streptomycin (P/S; Invitrogen) and 10% fetalbovine serum (VWR International, Bridgeport, NJ,USA). Medium was refreshed every 3e4 days. Cul-tures were harvested at 80% confluency by treatmentwith trypsin (Invitrogen), re-plated and maintainedfor a maximum of six passages at 37�C and 5% CO2.
Characterization of MSCs
All MSC cell lines were phenotypically characterizedat their second or third passage using antibodiesagainst CD3,CD45,CD86,HLA-DR,CD31,CD34,CD73 and CD90 (all Becton Dickinson Biosciences[BD], San Diego, CA, USA). CD105 was obtainedfrom Ancell Corporation (Bayport, MN, USA). Cellswere analyzed on a FACSCalibur flow cytometer(BD). Mean fluorescence intensity (MFI) was com-pared with cells stained with isotype-matched negativecontrol antibodies (BD) and with unstained cells.
The osteoblast and adipocyte differentiationpotential was evaluated on cells at passage 4e6 asdescribed previously (39). After 3 weeks, fat vacuolesin adipocytes and calcified depositions in osteoblastswere stained with Oil Red O (Sigma, St Louis, MO,USA) or Alizarin Red (MP Biomedicals, Solon, OH,USA), respectively.
Peripheral blood mononuclear cell cultures
To investigate the immunosuppressive effect ofMSCs on proliferation of peripheral blood mono-nuclear cells (PBMC), irradiated (30 Gy)MSCs frompatients with sJIA or HC were seeded at 2-fold serialdilutions in flat-bottom 96-well plates. PBMC, iso-lated by Ficoll separation from buffy coats of healthyblood bank donors (Sanquin, Amsterdam, TheNetherlands), were added (100 000 cells/well) after4 hours. Cells were stimulated with 2 mg/mL phyto-hemagglutinin (PHA; Murex, Châtillon, France) andcultured at 37�C and 5% CO2 for 5 days. Fiftymicroliters of culture supernatant was harvested atday 4 for cytokine analysis. The concentrations ofinterferon (IFN)-g and TNF-a in supernatants weremeasured using standard protocols of enzyme-linkedimmunosorbent assay kits (Sanquin). Cells werepulsed with 3H-thymidine (1 mCi/well; Perkin Elmer,Wellesley, MA, USA) for the final 16 hours andcounted with a b-counter (Perkin Elmer). All cultureswere performed in triplicate in RPMI 1640 (Invi-trogen) medium with 10% pooled human AB serum(Sanquin) and P/S (total volume of 200 mL/well).

282 F. G. J. Calkoen et al.
To investigate the influence of drugs used in thetreatment of sJIA, indomethacin (5 mmol/L; Sigma),dexamethasone (100 ng/mL; Sigma) and the solubleTNF-a receptor adalimumab (10 mg/mL; AbbottLaboratories, Abbott Park, IL, USA) were added atinitiation of culture.
NK cell cultures
NK cells were isolated from PBMC derived fromhealthy blood bank donors by negative selectionusing an NK isolation kit (Miltenyi Biotec, BergischGladbach, Germany). An NK cell purity greater than95% was reached. NK cells (1.0 � 106/well) werestimulated with 30 IU/mL IL-2 (Chiron Corpora-tion, Emeryville, CA, USA). NK cells were co-cultured with or without irradiated MSCs in 24-wellplates at an MSC:NK ratio of 1:5 or 1:40 in RPMI1640 supplemented with 10% human AB serumand P/S.
At day 5, NK cells were harvested and countedand cell marker expression and cytotoxicity wereassessed. The expression of CD69 (BD) andNKG2D (Beckman Coulter, Brea, CA, USA) wasdetermined by flow cytometry. Cytotoxicity wasdetermined in a standard 4-hour chromium releaseassay using various target cells (2500 cells/well)(i.e., Daudi cells, which are only sensitive to killingmediated by activated NK cells) and Epstein-Barrvirus transformed B cells (EBV-BLCL) coated withantibodies through pre-incubation with anti-thymocyte globulins (ATG; Genzyme, Cambridge,MA; 50 mg/mL for 20 min) to measure antibody-dependent cell-mediated cytotoxicity (ADCC).Target cells were labeled with 100 mCi sodium-51-chromate (Perkin Elmer) for 1 hour. Effectorcells were incubated with target cells at ratiosranging from 40:1 to 0.3:1 in triplicate. Sponta-neous and maximum release were determined byincubating target cells with medium or TritonX-100 (5%; Merck Chemicals, Darmstadt, Germany),respectively. Subsequently, chromium release in thesupernatant was assessed in a b-counter. Specific lysiswas determined as follows: (experimental release �spontaneous release)/(maximumrelease� spontaneousrelease) � 100%.
DC cultures
Monocytes were isolated from PBMC derived fromhealthy blood bank donors with a positive CD14selection kit (Miltenyi Biotec). Cell purity greater than90% was reached in all isolations. Monocytes (0.5 �106)were cultured in 24-well plates for 5days inRPMI1640, 10% FCS and 1% P/S supplemented with 800IU/mL granulocyte-macrophage colony-stimulating
factor (GM-CSF) and 40 ng/mL IL-4 (both fromTebu-Bio, Le Perray en Yvelines, France) to induceDC differentiation. At day 5, cells were harvested orsimilar concentrations of IL-4 and GM-CSF, andCD40 ligand (0.25 mg/mL; Beckman Coulter,Marseille, France) and IFN-g (500 U/mL; Boeh-ringer, Mannheim, Germany), were added tomature the DC for 2 extra days. All cells harvestedon days 5 and 7 were analyzed using flow cytometryfor CD14, CD1a, CD80, CD86 and CD163expression (antibodies from BD). MSCs were co-cultured with DC at MSC:DC ratios of 1:5, 1:40 or1:100.
Statistics
Data were analyzed using GraphPad Prism (La Jolla,CA). For statistical analysis, paired and unpairedt-tests were used. P values of <0.05 were consideredsignificant.
Results
Characterization of MSCs
MSCs were successfully expanded from BM of 11 of13 patients with sJIA and 10 of 10 HCs. Age did notsignificantly differ between the patients with sJIA andHCs (mean, 7.1 years [range, 1e12 years] versus 8.9years [range, 1e19 years], respectively); 45% of thepatients with sJIA and 40% of the HCs were female.The characteristics of the patients included in thestudy are summarized in Table I. Samples from twopatients, both frozen BMMC, failed to expand. Thecryopreservation time of these samples was 10.2 and12.3 years, respectively, versus a mean storage timeof 7.5 years for the successfully expanded samples(range, 2.4e11.1 years). In addition, cultures ofthese two samples were initiated with relatively lownumbers of BMMC (0.26 � 106 and 2.8 � 106). ForMSC cultures from frozen BMMC, significantlyfewer cells were available compared with culturesfrom freshly isolated BMMC (mean, 3.9 � 106
[range, 0.26e7.56 � 106] versus 33.1 � 106 [range,16e60 � 106] BMMC, respectively).
MSCs were characterized by adhesion to plasticand a fibroblast-like appearance. In addition, allMSCs fulfilled the phenotypic criteria (i.e., expres-sion of CD73, CD90 and CD105 and no expressionof CD34, CD31 and CD45) at passage 2 or 3(Figure 1A). All MSCs had the ability to differentiateinto adipocytes and osteoblasts on stimulation(Figure 1BeI). However, MSC lines derived fromtwo patients (UPN 2 and 5 in Table I), which wereobtained from frozen material, failed to differenti-ate into the osteoblastic lineage. These two MSC
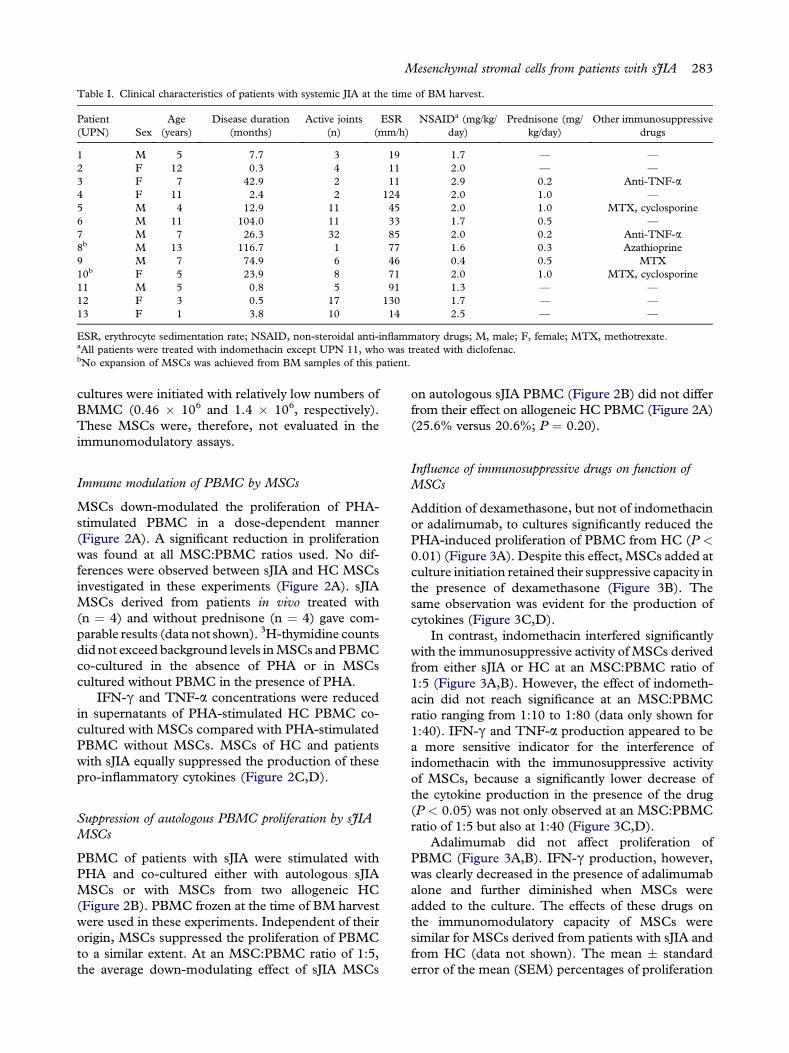
Table I. Clinical characteristics of patients with systemic JIA at the time of BM harvest.
Patient(UPN) Sex
Age(years)
Disease duration(months)
Active joints(n)
ESR(mm/h)
NSAIDa (mg/kg/day)
Prednisone (mg/kg/day)
Other immunosuppressivedrugs
1 M 5 7.7 3 19 1.7 — —
2 F 12 0.3 4 11 2.0 — —
3 F 7 42.9 2 11 2.9 0.2 Anti-TNF-a4 F 11 2.4 2 124 2.0 1.0 —
5 M 4 12.9 11 45 2.0 1.0 MTX, cyclosporine6 M 11 104.0 11 33 1.7 0.5 —
7 M 7 26.3 32 85 2.0 0.2 Anti-TNF-a8b M 13 116.7 1 77 1.6 0.3 Azathioprine9 M 7 74.9 6 46 0.4 0.5 MTX10b F 5 23.9 8 71 2.0 1.0 MTX, cyclosporine11 M 5 0.8 5 91 1.3 — —
12 F 3 0.5 17 130 1.7 — —
13 F 1 3.8 10 14 2.5 — —
ESR, erythrocyte sedimentation rate; NSAID, non-steroidal anti-inflammatory drugs; M, male; F, female; MTX, methotrexate.aAll patients were treated with indomethacin except UPN 11, who was treated with diclofenac.bNo expansion of MSCs was achieved from BM samples of this patient.
Mesenchymal stromal cells from patients with sJIA 283
cultures were initiated with relatively low numbers ofBMMC (0.46 � 106 and 1.4 � 106, respectively).These MSCs were, therefore, not evaluated in theimmunomodulatory assays.
Immune modulation of PBMC by MSCs
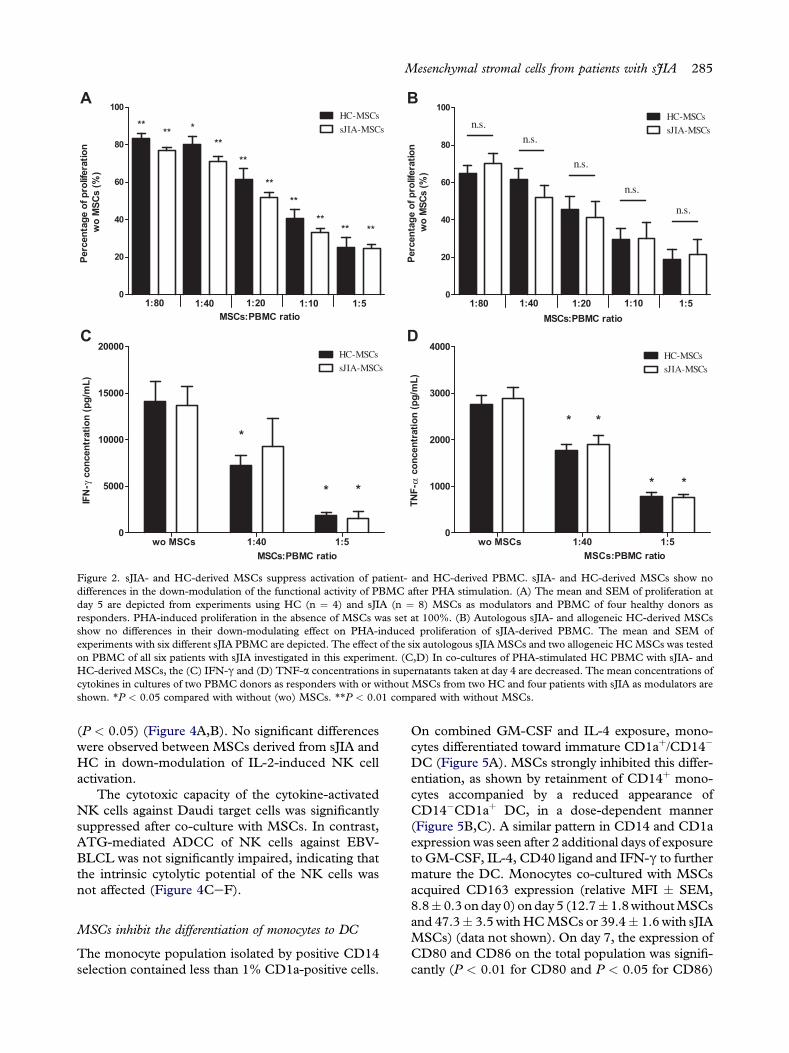
MSCs down-modulated the proliferation of PHA-stimulated PBMC in a dose-dependent manner(Figure 2A). A significant reduction in proliferationwas found at all MSC:PBMC ratios used. No dif-ferences were observed between sJIA and HC MSCsinvestigated in these experiments (Figure 2A). sJIAMSCs derived from patients in vivo treated with(n ¼ 4) and without prednisone (n ¼ 4) gave com-parable results (data not shown). 3H-thymidine countsdid not exceed background levels inMSCs andPBMCco-cultured in the absence of PHA or in MSCscultured without PBMC in the presence of PHA.
IFN-g and TNF-a concentrations were reducedin supernatants of PHA-stimulated HC PBMC co-cultured with MSCs compared with PHA-stimulatedPBMC without MSCs. MSCs of HC and patientswith sJIA equally suppressed the production of thesepro-inflammatory cytokines (Figure 2C,D).
Suppression of autologous PBMC proliferation by sJIAMSCs
PBMC of patients with sJIA were stimulated withPHA and co-cultured either with autologous sJIAMSCs or with MSCs from two allogeneic HC(Figure 2B). PBMC frozen at the time of BM harvestwere used in these experiments. Independent of theirorigin, MSCs suppressed the proliferation of PBMCto a similar extent. At an MSC:PBMC ratio of 1:5,the average down-modulating effect of sJIA MSCs
on autologous sJIA PBMC (Figure 2B) did not differfrom their effect on allogeneic HC PBMC (Figure 2A)(25.6% versus 20.6%; P ¼ 0.20).
Influence of immunosuppressive drugs on function ofMSCs
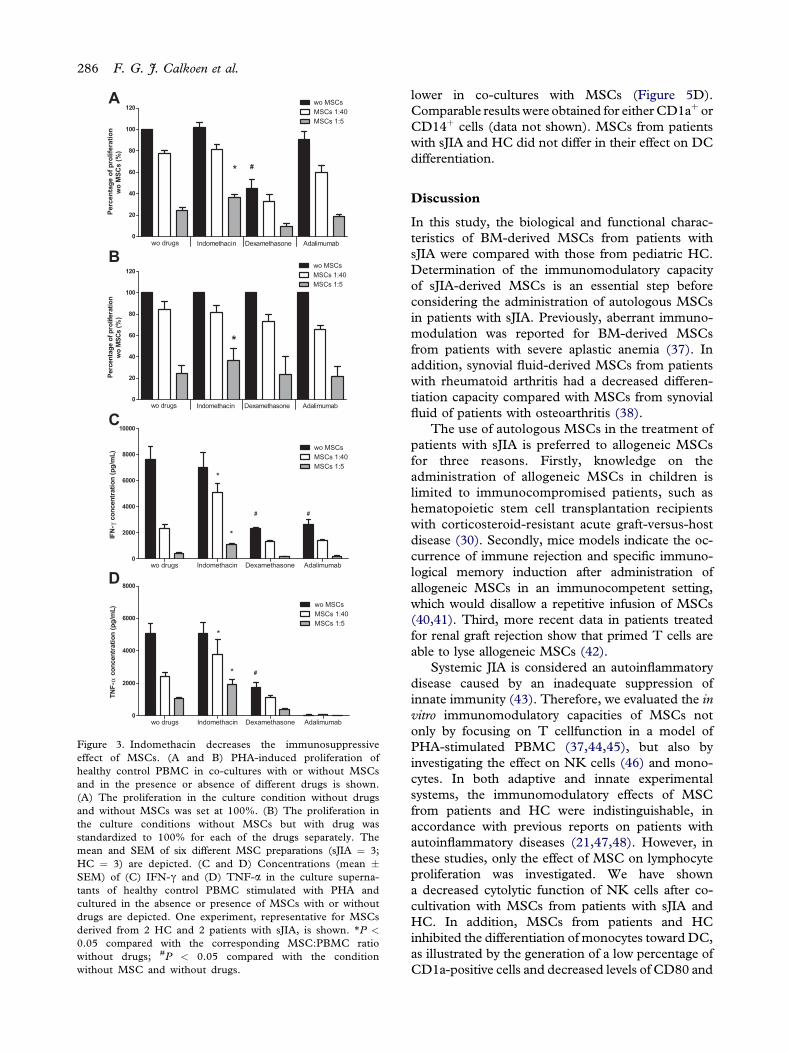
Addition of dexamethasone, but not of indomethacinor adalimumab, to cultures significantly reduced thePHA-induced proliferation of PBMC from HC (P <0.01) (Figure 3A). Despite this effect, MSCs added atculture initiation retained their suppressive capacity inthe presence of dexamethasone (Figure 3B). Thesame observation was evident for the production ofcytokines (Figure 3C,D).
In contrast, indomethacin interfered significantlywith the immunosuppressive activity of MSCs derivedfrom either sJIA or HC at an MSC:PBMC ratio of1:5 (Figure 3A,B). However, the effect of indometh-acin did not reach significance at an MSC:PBMCratio ranging from 1:10 to 1:80 (data only shown for1:40). IFN-g and TNF-a production appeared to bea more sensitive indicator for the interference ofindomethacin with the immunosuppressive activityof MSCs, because a significantly lower decrease ofthe cytokine production in the presence of the drug(P < 0.05) was not only observed at an MSC:PBMCratio of 1:5 but also at 1:40 (Figure 3C,D).
Adalimumab did not affect proliferation ofPBMC (Figure 3A,B). IFN-g production, however,was clearly decreased in the presence of adalimumabalone and further diminished when MSCs wereadded to the culture. The effects of these drugs onthe immunomodulatory capacity of MSCs weresimilar for MSCs derived from patients with sJIA andfrom HC (data not shown). The mean � standarderror of the mean (SEM) percentages of proliferation
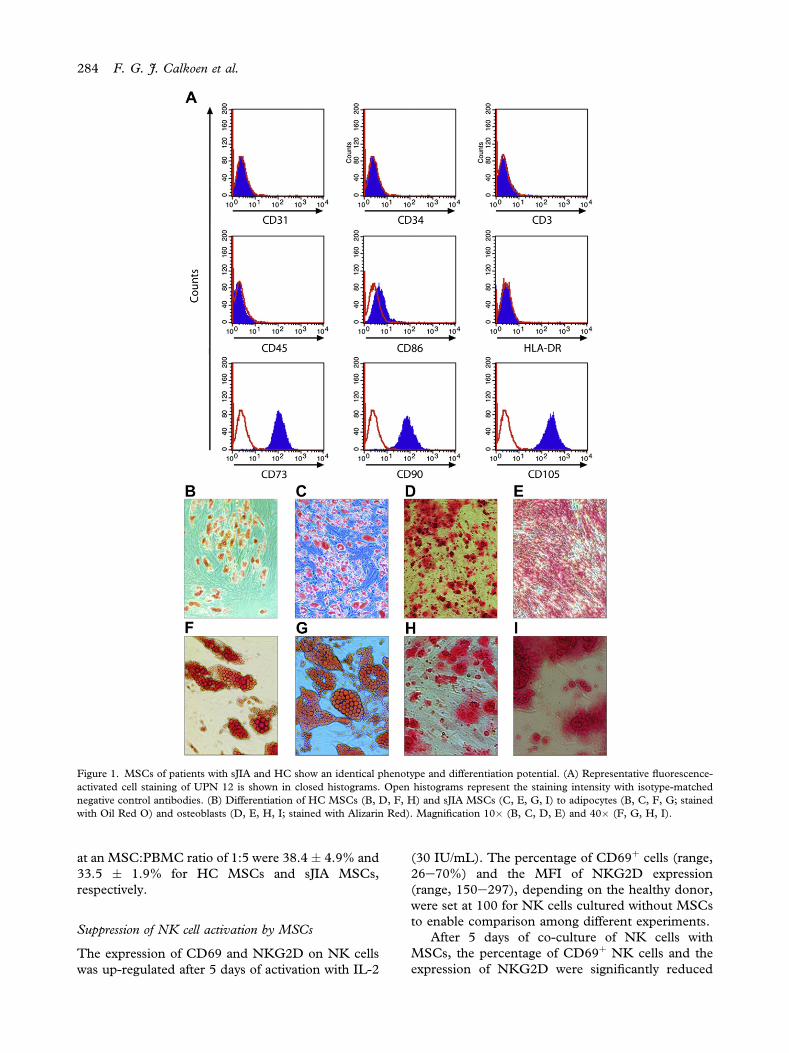
Figure 1. MSCs of patients with sJIA and HC show an identical phenotype and differentiation potential. (A) Representative fluorescence-activated cell staining of UPN 12 is shown in closed histograms. Open histograms represent the staining intensity with isotype-matchednegative control antibodies. (B) Differentiation of HC MSCs (B, D, F, H) and sJIA MSCs (C, E, G, I) to adipocytes (B, C, F, G; stainedwith Oil Red O) and osteoblasts (D, E, H, I; stained with Alizarin Red). Magnification 10� (B, C, D, E) and 40� (F, G, H, I).
284 F. G. J. Calkoen et al.
at an MSC:PBMC ratio of 1:5 were 38.4 � 4.9% and33.5 � 1.9% for HC MSCs and sJIA MSCs,respectively.
Suppression of NK cell activation by MSCs
The expression of CD69 and NKG2D on NK cellswas up-regulated after 5 days of activation with IL-2
(30 IU/mL). The percentage of CD69þ cells (range,26e70%) and the MFI of NKG2D expression(range, 150e297), depending on the healthy donor,were set at 100 for NK cells cultured without MSCsto enable comparison among different experiments.
After 5 days of co-culture of NK cells withMSCs, the percentage of CD69þ NK cells and theexpression of NKG2D were significantly reduced
A
C
0
20
40
60
80
100
Percen
tag
e o
f p
ro
liferatio
n
wo
M
SC
s (%
)
1:80 1:40 1:20 1:10 1:5
HC-MSCs
sJIA-MSCs**
**
**
**
**
**
**
**
**
*
MSCs:PBMC ratio
0
20
40
60
80
100
MSCs:PBMC ratio
Percen
tag
e o
f p
ro
liferatio
n
wo
M
SC
s (%
)
1:80 1:40 1:20 1:10 1:5
HC-MSCs
sJIA-MSCsn.s.
n.s.
n.s.
n.s.
n.s.
MSCs:PBMC ratio
IFN
-γ
co
ncen
tratio
n (p
g/m
L)
wo MSCs 1:40 1:5
0
5000
10000
15000
20000
HC-MSCs
sJIA-MSCs
*
**
MSCs:PBMC ratio
TN
F-α
co
ncen
tratio
n (p
g/m
L)
wo MSCs 1:40 1:5
0
1000
2000
3000
4000
HC-MSCssJIA-MSCs
* *
* *
B
D
Figure 2. sJIA- and HC-derived MSCs suppress activation of patient- and HC-derived PBMC. sJIA- and HC-derived MSCs show nodifferences in the down-modulation of the functional activity of PBMC after PHA stimulation. (A) The mean and SEM of proliferation atday 5 are depicted from experiments using HC (n ¼ 4) and sJIA (n ¼ 8) MSCs as modulators and PBMC of four healthy donors asresponders. PHA-induced proliferation in the absence of MSCs was set at 100%. (B) Autologous sJIA- and allogeneic HC-derived MSCsshow no differences in their down-modulating effect on PHA-induced proliferation of sJIA-derived PBMC. The mean and SEM ofexperiments with six different sJIA PBMC are depicted. The effect of the six autologous sJIAMSCs and two allogeneic HCMSCs was testedon PBMC of all six patients with sJIA investigated in this experiment. (C,D) In co-cultures of PHA-stimulated HC PBMC with sJIA- andHC-derived MSCs, the (C) IFN-g and (D) TNF-a concentrations in supernatants taken at day 4 are decreased. The mean concentrations ofcytokines in cultures of two PBMC donors as responders with or without MSCs from two HC and four patients with sJIA as modulators areshown. *P < 0.05 compared with without (wo) MSCs. **P < 0.01 compared with without MSCs.
Mesenchymal stromal cells from patients with sJIA 285
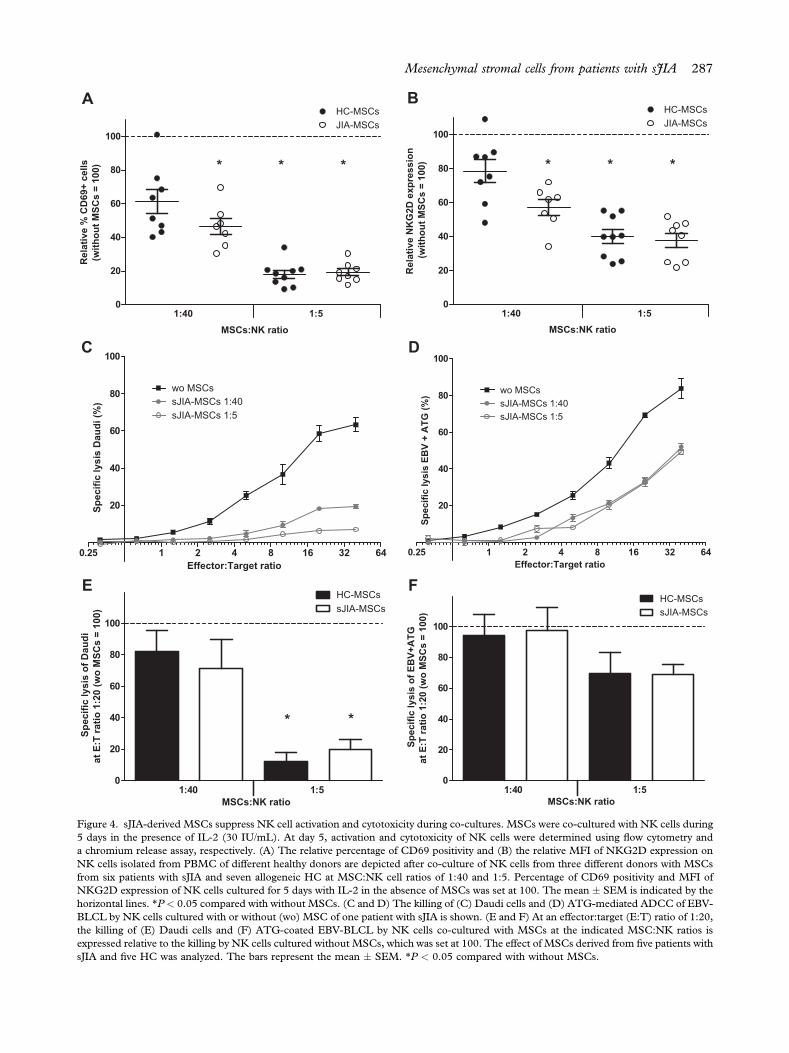
(P < 0.05) (Figure 4A,B). No significant differenceswere observed between MSCs derived from sJIA andHC in down-modulation of IL-2-induced NK cellactivation.
The cytotoxic capacity of the cytokine-activatedNK cells against Daudi target cells was significantlysuppressed after co-culture with MSCs. In contrast,ATG-mediated ADCC of NK cells against EBV-BLCL was not significantly impaired, indicating thatthe intrinsic cytolytic potential of the NK cells wasnot affected (Figure 4CeF).
MSCs inhibit the differentiation of monocytes to DC
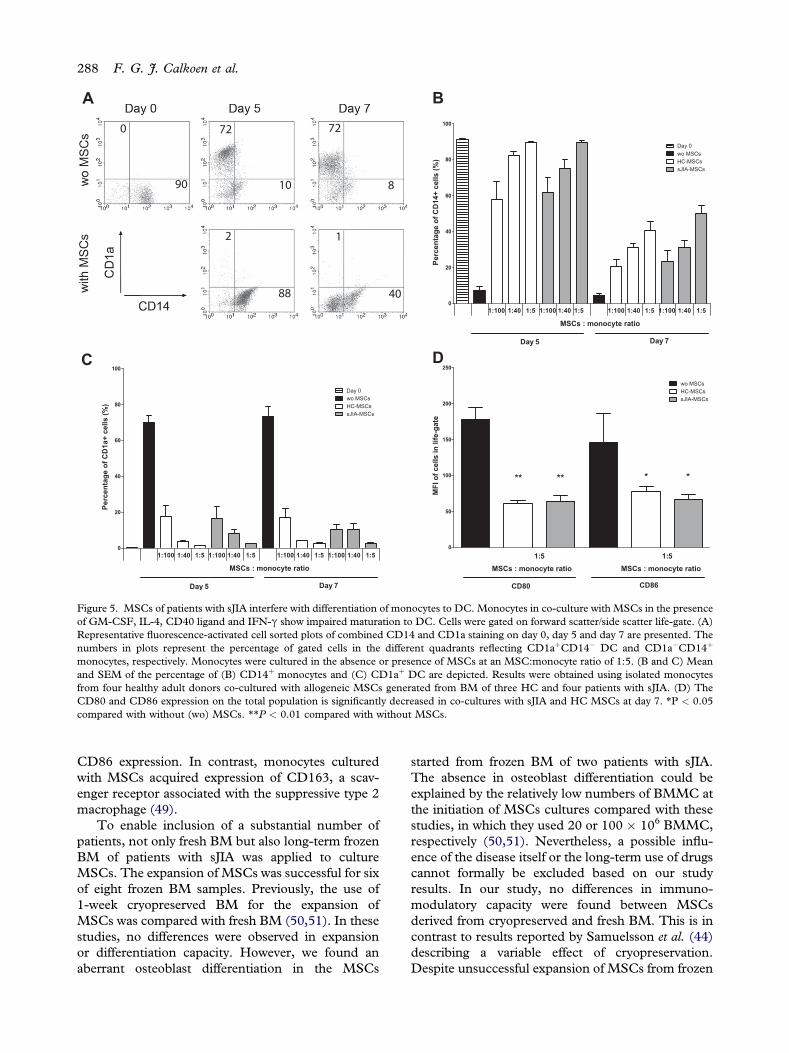
The monocyte population isolated by positive CD14selection contained less than 1% CD1a-positive cells.
On combined GM-CSF and IL-4 exposure, mono-cytes differentiated toward immature CD1aþ/CD14�
DC (Figure 5A). MSCs strongly inhibited this differ-entiation, as shown by retainment of CD14þ mono-cytes accompanied by a reduced appearance ofCD14�CD1aþ DC, in a dose-dependent manner(Figure 5B,C). A similar pattern in CD14 and CD1aexpression was seen after 2 additional days of exposureto GM-CSF, IL-4, CD40 ligand and IFN-g to furthermature the DC. Monocytes co-cultured with MSCsacquired CD163 expression (relative MFI � SEM,8.8� 0.3 onday 0) on day 5 (12.7� 1.8withoutMSCsand 47.3� 3.5 withHCMSCs or 39.4� 1.6 with sJIAMSCs) (data not shown). On day 7, the expression ofCD80 and CD86 on the total population was signifi-cantly (P < 0.01 for CD80 and P < 0.05 for CD86)
Pe
rc
en
ta
ge
o
f p
ro
life
ra
tio
n
wo
M
SC
s (%
)
0
20
40
60
80
100
120
wo drugs Indomethacin Dexamethasone Adalimumab
*
wo MSCsMSCs 1:40MSCs 1:5
#
Percen
tag
e o
f p
ro
liferatio
n
wo
M
SC
s (%
)
0
20
40
60
80
100
120
wo drugs Indomethacin Dexamethasone Adalimumab
*
wo MSCsMSCs 1:40MSCs 1:5
IFN
-γ
co
ncen
tratio
n (p
g/m
L)
wo drugs Indomethacin Dexamethasone Adalimumab0
2000
4000
6000
8000
10000
wo MSCsMSCs 1:40MSCs 1:5
*
*
# #
TN
F-α
co
ncen
tratio
n (p
g/m
L)
wo drugs Indomethacin Dexamethasone Adalimumab0
2000
4000
6000
8000
wo MSCsMSCs 1:40MSCs 1:5
*
* #
A
B
C
D
Figure 3. Indomethacin decreases the immunosuppressiveeffect of MSCs. (A and B) PHA-induced proliferation ofhealthy control PBMC in co-cultures with or without MSCsand in the presence or absence of different drugs is shown.(A) The proliferation in the culture condition without drugsand without MSCs was set at 100%. (B) The proliferation inthe culture conditions without MSCs but with drug wasstandardized to 100% for each of the drugs separately. Themean and SEM of six different MSC preparations (sJIA ¼ 3;HC ¼ 3) are depicted. (C and D) Concentrations (mean �SEM) of (C) IFN-g and (D) TNF-a in the culture superna-tants of healthy control PBMC stimulated with PHA andcultured in the absence or presence of MSCs with or withoutdrugs are depicted. One experiment, representative for MSCsderived from 2 HC and 2 patients with sJIA, is shown. *P <
0.05 compared with the corresponding MSC:PBMC ratiowithout drugs; #P < 0.05 compared with the conditionwithout MSC and without drugs.
286 F. G. J. Calkoen et al.
lower in co-cultures with MSCs (Figure 5D).Comparable results were obtained for eitherCD1aþ orCD14þ cells (data not shown). MSCs from patientswith sJIA and HC did not differ in their effect on DCdifferentiation.
Discussion
In this study, the biological and functional charac-teristics of BM-derived MSCs from patients withsJIA were compared with those from pediatric HC.Determination of the immunomodulatory capacityof sJIA-derived MSCs is an essential step beforeconsidering the administration of autologous MSCsin patients with sJIA. Previously, aberrant immuno-modulation was reported for BM-derived MSCsfrom patients with severe aplastic anemia (37). Inaddition, synovial fluid-derived MSCs from patientswith rheumatoid arthritis had a decreased differen-tiation capacity compared with MSCs from synovialfluid of patients with osteoarthritis (38).
The use of autologous MSCs in the treatment ofpatients with sJIA is preferred to allogeneic MSCsfor three reasons. Firstly, knowledge on theadministration of allogeneic MSCs in children islimited to immunocompromised patients, such ashematopoietic stem cell transplantation recipientswith corticosteroid-resistant acute graft-versus-hostdisease (30). Secondly, mice models indicate the oc-currence of immune rejection and specific immuno-logical memory induction after administration ofallogeneic MSCs in an immunocompetent setting,which would disallow a repetitive infusion of MSCs(40,41). Third, more recent data in patients treatedfor renal graft rejection show that primed T cells areable to lyse allogeneic MSCs (42).
Systemic JIA is considered an autoinflammatorydisease caused by an inadequate suppression ofinnate immunity (43). Therefore, we evaluated the invitro immunomodulatory capacities of MSCs notonly by focusing on T cellfunction in a model ofPHA-stimulated PBMC (37,44,45), but also byinvestigating the effect on NK cells (46) and mono-cytes. In both adaptive and innate experimentalsystems, the immunomodulatory effects of MSCfrom patients and HC were indistinguishable, inaccordance with previous reports on patients withautoinflammatory diseases (21,47,48). However, inthese studies, only the effect of MSC on lymphocyteproliferation was investigated. We have showna decreased cytolytic function of NK cells after co-cultivation with MSCs from patients with sJIA andHC. In addition, MSCs from patients and HCinhibited the differentiation of monocytes towardDC,as illustrated by the generation of a low percentage ofCD1a-positive cells and decreased levels of CD80 and
MSCs:NK ratio
Relative N
KG
2D
exp
ressio
n
(w
ith
ou
t M
SC
s =
100)
1:40 1:5
0
20
40
60
80
100
* **
HC-MSCsJIA-MSCs
MSCs:NK ratio
Relative %
C
D69+
cells
(w
ith
ou
t M
SC
s =
100)
1:40 1:5
0
20
40
60
80
100
* **
HC-MSCsJIA-MSCs
Effector:Target ratio
Sp
ec
ific
ly
sis
D
au
di (%
)
0.25 1 2 4 8 16 32 64
20
40
60
80
100
wo MSCssJIA-MSCs 1:40sJIA-MSCs 1:5
Effector:Target ratio
Sp
ecific lysis E
BV
+
A
TG
(%
)
0.25 1 2 4 8 16 32 64
20
40
60
80
100
wo MSCssJIA-MSCs 1:40sJIA-MSCs 1:5
MSCs:NK ratio
Sp
ec
ific
ly
sis
o
f D
au
di
at E
:T
ra
tio
1
:2
0 (w
o M
SC
s =
1
00
)
0
20
40
60
80
100
* *
HC-MSCssJIA-MSCs
5:104:1
MSCs:NK ratio
Sp
ecific lysis o
f E
BV
+A
TG
at E
:T
ratio
1:20 (w
o M
SC
s =
100)
0
20
40
60
80
100
HC-MSCssJIA-MSCs
5:104:1
BA
DC
FE
Figure 4. sJIA-derived MSCs suppress NK cell activation and cytotoxicity during co-cultures. MSCs were co-cultured with NK cells during5 days in the presence of IL-2 (30 IU/mL). At day 5, activation and cytotoxicity of NK cells were determined using flow cytometry anda chromium release assay, respectively. (A) The relative percentage of CD69 positivity and (B) the relative MFI of NKG2D expression onNK cells isolated from PBMC of different healthy donors are depicted after co-culture of NK cells from three different donors with MSCsfrom six patients with sJIA and seven allogeneic HC at MSC:NK cell ratios of 1:40 and 1:5. Percentage of CD69 positivity and MFI ofNKG2D expression of NK cells cultured for 5 days with IL-2 in the absence of MSCs was set at 100. The mean � SEM is indicated by thehorizontal lines. *P < 0.05 compared with without MSCs. (C and D) The killing of (C) Daudi cells and (D) ATG-mediated ADCC of EBV-BLCL by NK cells cultured with or without (wo) MSC of one patient with sJIA is shown. (E and F) At an effector:target (E:T) ratio of 1:20,the killing of (E) Daudi cells and (F) ATG-coated EBV-BLCL by NK cells co-cultured with MSCs at the indicated MSC:NK ratios isexpressed relative to the killing by NK cells cultured without MSCs, which was set at 100. The effect of MSCs derived from five patients withsJIA and five HC was analyzed. The bars represent the mean � SEM. *P < 0.05 compared with without MSCs.
Mesenchymal stromal cells from patients with sJIA 287
Percen
tag
e o
f C
D14+
cells (%
)
0
20
40
60
80
100
Day 5 Day 7
Day 0wo MSCsHC-MSCssJIA-MSCs
1:100 1:40 1:5 1:100 1:40 1:5 1:100 1:40 1:5 1:100 1:40 1:5
MSCs : monocyte ratio
Percen
tag
e o
f C
D1a+
cells (%
)
0
20
40
60
80
100
Day 0wo MSCs
Day 5 Day 7
1:100 1:40 1:5 1:100 1:40 1:5 1:100 1:40 1:5 1:100 1:40 1:5
MSCs : monocyte ratio
HC-MSCssJIA-MSCs
MF
I o
f cells in
life-g
ate
0
50
100
150
200
250
CD80 CD86
wo MSCs
sJIA-MSCs
** ** * *
MSCs : monocyte ratio MSCs : monocyte ratio
5:15:1
HC-MSCs
BA
DC
Figure 5. MSCs of patients with sJIA interfere with differentiation of monocytes to DC. Monocytes in co-culture with MSCs in the presenceof GM-CSF, IL-4, CD40 ligand and IFN-g show impaired maturation to DC. Cells were gated on forward scatter/side scatter life-gate. (A)Representative fluorescence-activated cell sorted plots of combined CD14 and CD1a staining on day 0, day 5 and day 7 are presented. Thenumbers in plots represent the percentage of gated cells in the different quadrants reflecting CD1aþCD14� DC and CD1a�CD14þ
monocytes, respectively. Monocytes were cultured in the absence or presence of MSCs at an MSC:monocyte ratio of 1:5. (B and C) Meanand SEM of the percentage of (B) CD14þ monocytes and (C) CD1aþ DC are depicted. Results were obtained using isolated monocytesfrom four healthy adult donors co-cultured with allogeneic MSCs generated from BM of three HC and four patients with sJIA. (D) TheCD80 and CD86 expression on the total population is significantly decreased in co-cultures with sJIA and HC MSCs at day 7. *P < 0.05compared with without (wo) MSCs. **P < 0.01 compared with without MSCs.
288 F. G. J. Calkoen et al.
CD86 expression. In contrast, monocytes culturedwith MSCs acquired expression of CD163, a scav-enger receptor associated with the suppressive type 2macrophage (49).
To enable inclusion of a substantial number ofpatients, not only fresh BM but also long-term frozenBM of patients with sJIA was applied to cultureMSCs. The expansion of MSCs was successful for sixof eight frozen BM samples. Previously, the use of1-week cryopreserved BM for the expansion ofMSCs was compared with fresh BM (50,51). In thesestudies, no differences were observed in expansionor differentiation capacity. However, we found anaberrant osteoblast differentiation in the MSCs
started from frozen BM of two patients with sJIA.The absence in osteoblast differentiation could beexplained by the relatively low numbers of BMMC atthe initiation of MSCs cultures compared with thesestudies, in which they used 20 or 100 � 106 BMMC,respectively (50,51). Nevertheless, a possible influ-ence of the disease itself or the long-term use of drugscannot formally be excluded based on our studyresults. In our study, no differences in immuno-modulatory capacity were found between MSCsderived from cryopreserved and fresh BM. This is incontrast to results reported by Samuelsson et al. (44)describing a variable effect of cryopreservation.Despite unsuccessful expansion of MSCs from frozen

Mesenchymal stromal cells from patients with sJIA 289
BM in two patients, the expansion of MSCs fromfrozen material is a promising tool for further researchon rare pediatric diseases.
An important issue in administration of MSCs topatients is the continuation of disease-specific drugs.Previously, calcineurin inhibitors and indomethacinwere shown to suppress the immunomodulatoryeffect of MSCs, whereas mycophenolic acid hasa synergistic effect with MSCs (45,52). Consistentwith previous data (27), our findings indicate thatindomethacin suppresses the immunomodulatoryfunction of MSCs. All of our patients were treatedwith non-steroidal anti-inflammatory drugs beforeaspiration of BM. Apparently, in vivo exposure ofMSCs to non-steroidal anti-inflammatory drugs doesnot interfere with the suppressive effect of MSCsafter ex vivo expansion. Dexamethasone, on the otherhand, although having a suppressive effect on PBMCproliferation by itself, did not affect the suppressiveeffect of MSCs, consistent with a previous report(52). Adalimumab, a TNF-a antagonist, did notaffect proliferation of PBMC but significantlydecreased PHA-induced production of IFN-g.Interestingly, despite the low production of IFN-g incultures containing adalimumab, MSCS retain theircapacity to reduce proliferation and IFN-g secretionin this condition.
It is relevant to evaluate whether patient-derivedcells, independent of the course and actual activity ofthe disease and past or present medication, aresusceptible to suppression by MSCs. Therefore, wedetermined the effect of patient-derived MSCs onPHA-driven T cell proliferation using autologousPBMC. Patient-derived PBMC were equally sup-pressed by MSCs compared with HC-derivedPBMC. Our data further show a comparablesuppression of T cellresponses by allogeneic andautologous MSCs, implying a comparable suscepti-bility of patient and control PBMC to MSCs.
Our in vitro study did not address the route ofadministration of these expanded MSCs, andoptimal use remains to be determined. Most re-ported clinical studies of MSCs treatment to datehave used intravenous infusions to control systemicinflammatory diseases, and this would also seem tobe a logical choice for future use in children withsJIA. In contrast, however, oligo-articular disease ortarget joints might benefit from local intra-articularadministration. Future prospective clinical trialsshould address these issues.
In conclusion, this is the first study to report thatMSCs derived from a cohort of patients with sJIAhave similar immunosuppressive capacities com-pared with age-matched HC in assays focused onadaptive (T cell) and innate (NK cell and monocyte)immunity. Therefore, the results of this study
support the use of autologous MSCs in clinical trialsof patients with sJIA.
Acknowledgments
This work was funded by a grant (NR 07-01-201)from the Dutch Arthritis Association and by theDutch Children Cancer Free Foundation (KiKa) forthe appointment of CV and FC.
Disclosure of interest: The author has nocommercial, proprietary or financial interest in theproducts or companies described in this article.
References
1. Petty RE, Southwood TR, Manners P, Baum J, Glass DN,Goldenberg J, et al. International League of Associations forRheumatology classification of juvenile idiopathic arthritis:second revision, Edmonton, 2001. J Rheumatol. 2004;31:390e2.
2. Lequerre T, Quartier P, Rosellini D, Alaoui F, De BM,Mejjad O, et al. Interleukin-1 receptor antagonist (anakinra)treatment in patients with systemic-onset juvenile idiopathicarthritis or adult onset Still disease: preliminary experience inFrance. Ann Rheum Dis. 2008;67:302e8.
3. De Benedetti F. Targeting interleukin-6 in pediatric rheu-matic diseases. Curr Opin Rheumatol. 2009;21:533e7.
4. Woo P. Anakinra treatment for systemic juvenile idiopathicarthritis and adult onset Still disease. Ann Rheum Dis. 2008;67:281e2.
5. Martini G, Zulian F. Juvenile idiopathic arthritis: current andfuture treatment options. Expert Opin Pharmacother. 2006;7:387e99.
6. Beukelman T, Patkar NM, Saag KG, Tolleson-Rinehart S,Cron RQ, DeWitt EM, et al. 2011 American College ofRheumatology recommendations for the treatment of juvenileidiopathic arthritis: initiation and safety monitoring of thera-peutic agents for the treatment of arthritis and systemicfeatures. Arthritis Care Res (Hoboken). 2011;63:465e82.
7. Quartier P, Allantaz F, Cimaz R, Pillet P, Messiaen C,Bardin C, et al. Extended report: a multicentre, randomised,double-blind, placebo-controlled trial with the interleukin-1receptor antagonist anakinra in patients with systemic-onsetjuvenile idiopathic arthritis (ANAJIS trial). Ann Rheum Dis.2011;70:747e54.
8. Nigrovic PA, Mannion M, Prince FH, Zeft A, Rabinovich CE,van Rossum MA, et al. Anakinra as first-line disease-modi-fying therapy in systemic juvenile idiopathic arthritis: report offorty-six patients from an international multicenter series.Arthritis Rheum. 2011;63:545e55.
9. Yokota S, Imagawa T, Mori M, Miyamae T, Aihara Y,Takei S, et al. Efficacy and safety of tocilizumab in patientswith systemic-onset juvenile idiopathic arthritis: a randomised,double-blind, placebo-controlled, withdrawal phase III trial.Lancet. 2008;371:998e1006.
10. Prince FH, de Bekker-Grob EW, Twilt M, van Rossum MA,Hoppenreijs EP, ten Cate R, et al. An analysis of the costs andtreatment success of etanercept in juvenile idiopathic arthritis:results from the Dutch Arthritis and Biologicals in Childrenregister. Rheumatology (Oxford). 2011;50:1131e6.
11. Horneff G, Foeldvari I, Minden K, Moebius D, Hospach T.Report on malignancies in the German juvenile idiopathicarthritis registry. Rheumatology (Oxford). 2011;50:230e6.

290 F. G. J. Calkoen et al.
12. Kwon JH, Farrell RJ. The risk of lymphoma in the treatmentof inflammatory bowel disease with immunosuppressiveagents. Crit Rev Oncol Hematol. 2005;56:169e78.
13. Diak P, Siegel J, La Grenade L, Choi L, Lemery S,McMahon A. Tumor necrosis factor alpha blockers andmalignancy in children: forty-eight cases reported to the Foodand Drug Administration. Arthritis Rheum. 2010;62:2517e24.
14. Farge D, Labopin M, Tyndall A, Fassas A, Mancardi GL,Van Laar J, et al. Autologous hematopoietic stem cell trans-plantation for autoimmune diseases: an observational study on12 years’ experience from the European Group for Blood andMarrow Transplantation Working Party on AutoimmuneDiseases. Haematologica. 2010;95:284e92.
15. Brinkman DM, de Kleer IM, ten Cate R, van Rossum MA,Bekkering WP, Fasth A, et al. Autologous stem cell trans-plantation in children with severe progressive systemic or pol-yarticular juvenile idiopathic arthritis: long-term follow-up ofa prospective clinical trial. Arthritis Rheum. 2007;56:2410e21.
16. Krampera M, Glennie S, Dyson J, Scott D, Laylor R,Simpson E, et al. Bone marrow mesenchymal stem cells inhibitthe response of naive and memory antigen-specific T cells totheir cognate peptide. Blood. 2003;101:3722e9.
17. Bocelli-Tyndall C, Bracci L, Schaeren S, Feder-Mengus C,Barbero A, Tyndall A, et al. Human bone marrow mesen-chymal stem cells and chondrocytes promote and/or suppressthe in vitro proliferation of lymphocytes stimulated by inter-leukins 2, 7 and 15. Ann Rheum Dis. 2009;68:1352e9.
18. Bernardo ME, Zaffaroni N, Novara F, Cometa AM,Avanzini MA, Moretta A, et al. Human bone marrow derivedmesenchymal stem cells do not undergo transformation afterlong-term in vitro culture and do not exhibit telomere main-tenance mechanisms. Cancer Res. 2007;67:9142e9.
19. Dominici M, Le Blanc K, Mueller I, Slaper-Cortenbach I,Marini F, Krause D, et al. Minimal criteria for definingmultipotent mesenchymal stromal cells. The InternationalSociety for Cellular Therapy position statement. Cytotherapy.2006;8:315e7.
20. Horwitz EM, Le BK, Dominici M, Mueller I, Slaper-Cortenbach I, Marini FC, et al. Clarification of the nomen-clature for MSC: The International Society for CellularTherapy position statement. Cytotherapy. 2005;7:393e5.
21. Bocelli-Tyndall C, Bracci L, Spagnoli G, Braccini A,Bouchenaki M, Ceredig R, et al. Bone marrow mesenchymalstromal cells (BM-MSCs) from healthy donors and auto-immune disease patients reduce the proliferation of autologous-and allogeneic-stimulated lymphocytes in vitro. Rheumatology(Oxford). 2007;46:403e8.
22. Sotiropoulou PA, Perez SA, Gritzapis AD, Baxevanis CN,Papamichail M. Interactions between human mesenchymalstem cells and natural killer cells. Stem Cells. 2006;24:74e85.
23. Traggiai E, Volpi S, Schena F, Gattorno M, Ferlito F,Moretta L, et al. Bone marrow-derived mesenchymal stemcells induce both polyclonal expansion and differentiation of Bcells isolated from healthy donors and systemic lupus eryth-ematosus patients. Stem Cells. 2008;26:562e9.
24. Beyth S, Borovsky Z, Mevorach D, Liebergall M, Gazit Z,Aslan H, et al. Human mesenchymal stem cells alter antigen-presenting cell maturation and induce T-cell unresponsive-ness. Blood. 2005;105:2214e9.
25. Gieseke F, Bohringer J, Bussolari R, Dominici M,Handgretinger R, Muller I. Human multipotent mesenchymalstromal cells use galectin-1 to inhibit immune effector cells.Blood. 2010;116:3770e9.
26. Meisel R, Zibert A, Laryea M, Gobel U, Daubener W,Dilloo D. Human bone marrow stromal cells inhibit alloge-neic T-cell responses by indoleamine 2,3-dioxygenase-medi-ated tryptophan degradation. Blood. 2004;103:4619e21.
27. Rasmusson I, Ringden O, Sundberg B, Le Blanc K. Mesen-chymal stem cells inhibit lymphocyte proliferation by mito-gens and alloantigens by different mechanisms. Exp Cell Res.2005;305:33e41.
28. Selmani Z, Naji A, Zidi I, Favier B, Gaiffe E, Obert L, et al.Human leukocyte antigen-G5 secretion by human mesen-chymal stem cells is required to suppress T lymphocyte andnatural killer function and to induceCD4þCD25highFOXP3þregulatory T cells. Stem Cells. 2008;26:212e22.
29. Uccelli A, Moretta L, Pistoia V. Mesenchymal stem cells inhealth and disease. Nat Rev Immunol. 2008;8:726e36.
30. Le Blanc K, Frassoni F, Ball L, Locatelli F, Roelofs H,Lewis I, et al. Mesenchymal stem cells for treatment ofsteroid-resistant, severe, acute graft-versus-host disease:a phase II study. Lancet. 2008;371:1579e86.
31. Garcia-Olmo D, Garcia-Arranz M, Herreros D, Pascual I,Peiro C, Rodriguez-Montes JA. A phase I clinical trial ofthe treatment of Crohn’s fistula by adipose mesenchymalstem cell transplantation. Dis Colon Rectum. 2005;48:1416e23.
32. Duijvestein M, Vos AC, Roelofs H, Wildenberg ME,Wendrich BB, Verspaget HW, et al. Autologous bonemarrow-derived mesenchymal stromal cell treatment forrefractory luminal Crohn’s disease: results of a phase I study.Gut. 2010;59:1662e9.
33. Sun L, Akiyama K, Zhang H, Yamaza T, Hou Y, Zhao S,et al. Mesenchymal stem cell transplantation reverses multi-organ dysfunction in systemic lupus erythematosus mice andhumans. Stem Cells. 2009;27:1421e32.
34. Sun L, Wang D, Liang J, Zhang H, Feng X, Wang H, et al.Umbilical cord mesenchymal stem cell transplantation insevere and refractory systemic lupus erythematosus. ArthritisRheum. 2010;62:2467e75.
35. Karussis D, Karageorgiou C, Vaknin-Dembinsky A, Gowda-Kurkalli B, Gomori JM, Kassis I, et al. Safety and immuno-logical effects of mesenchymal stem cell transplantation inpatients with multiple sclerosis and amyotrophic lateral scle-rosis. Arch Neurol. 2010;67:1187e94.
36. Dazzi F, Krampera M. Mesenchymal stem cells and autoim-mune diseases. Best Pract ResClinHaematol. 2011;24:49e57.
37. Bacigalupo A, Valle M, Podesta M, Pitto A, Zocchi E, DeFlora A, et al. T-cell suppression mediated by mesenchymalstem cells is deficient in patients with severe aplastic anemia.Exp Hematol. 2005;33:819e27.
38. Jones E, Churchman SM, English A, Buch MH, Horner EA,Burgoyne CH, et al. Mesenchymal stem cells in rheumatoidsynovium: enumeration and functional assessment in relationto synovial inflammation level. Ann Rheum Dis. 2010;69:450e7.
39. Bernardo ME, Emons JA, Karperien M, Nauta AJ,Willemze R, Roelofs H, et al. Human mesenchymal stem cellsderived from bone marrow display a better chondrogenicdifferentiation compared with other sources. Connect TissueRes. 2007;48:132e40.
40. Nauta AJ, Westerhuis G, Kruisselbrink AB, Lurvink EG,Willemze R, Fibbe WE. Donor-derived mesenchymal stemcells are immunogenic in an allogeneic host and stimulatedonor graft rejection in a nonmyeloablative setting. Blood.2006;108:2114e20.
41. Zangi L, Margalit R, Reich-Zeliger S, Bachar-Lustig E,Beilhack A, Negrin R, et al. Direct imaging of immunerejection and memory induction by allogeneic mesenchymalstromal cells. Stem Cells. 2009;27:2865e74.
42. Crop MJ, Korevaar SS, de KR, Ijzermans JN, vanBesouw NM, Baan CC, et al. Human mesenchymal stem cellsare susceptible to lysis by CD8þ T-cells and NK cells. CellTransplant. 2011;20:1547e59.

Mesenchymal stromal cells from patients with sJIA 291
43. Vastert SJ, Kuis W, Grom AA. Systemic JIA: new develop-ments in the understanding of the pathophysiology andtherapy. Best Pract Res Clin Rheumatol. 2009;23:655e64.
44. Samuelsson H, Ringden O, Lonnies H, Le Blanc K. Opti-mizing in vitro conditions for immunomodulation andexpansion of mesenchymal stromal cells. Cytotherapy.2009;11:129e36.
45. Aggarwal S, Pittenger MF. Human mesenchymal stem cellsmodulate allogeneic immune cell responses. Blood. 2005;105:1815e22.
46. Spaggiari GM, Capobianco A, Becchetti S, Mingari MC,Moretta L. Mesenchymal stem cell-natural killer cell inter-actions: evidence that activated NK cells are capable of killingMSCs, whereas MSCs can inhibit IL-2-induced NK-cellproliferation. Blood. 2006;107:1484e90.
47. Bernardo ME, Avanzini MA, Ciccocioppo R, Perotti C,Cometa AM,Moretta A, et al. Phenotypical/functional charac-terization of in vitro-expanded mesenchymal stromal cells frompatients with Crohn’s disease. Cytotherapy. 2009;11:825e36.
48. Larghero J, Farge D, Braccini A, Lecourt S, Scherberich A,Fois E, et al. Phenotypical and functional characteristics of in
vitro expanded bone marrow mesenchymal stem cells frompatients with systemic sclerosis. Ann Rheum Dis. 2008;67:443e9.
49. Sica A, Schioppa T, Mantovani A, Allavena P. Tumour-associated macrophages are a distinct M2 polarised pop-ulation promoting tumour progression: potential targets ofanti-cancer therapy. Eur J Cancer. 2006;42:717e27.
50. Haack-Sorensen M, Bindslev L, Mortensen S, Friis T,Kastrup J. The influence of freezing and storage on thecharacteristics and functions of human mesenchymal stromalcells isolated for clinical use. Cytotherapy. 2007;9:328e37.
51. Casado-Diaz A, Santiago-Mora R, Jimenez R, Caballero-Villarraso J, Herrera C, Torres A, et al. Cryopreserved humanbone marrow mononuclear cells as a source of mesenchymalstromal cells: application in osteoporosis research. Cytother-apy. 2008;10:460e8.
52. Buron F, Perrin H, Malcus C, Hequet O, Thaunat O,Kholopp-Sarda MN, et al. Human mesenchymal stem cellsand immunosuppressive drug interactions in allogeneicresponses: an in vitro study using human cells. TransplantProc. 2009;41:3347e52.





























