Embed Size (px)
Citation preview

BioEssays Vol. 7, No. 6 - December 1987 255
REVIEW ARTICLES
Memories of fos Tom Curran and James I. Morgan
' . . .he who, seeing fossil shells on a mountain, conceived the theory of the deluge' Samuel Butler
Summary
Induction of c-fos expression occurs following treatment of diverse cell types with agents that trigger mitogenesis, digerentiation or membrane depolar- ization. We suggest that c-fos may be regarded as a marker for a set of rapidly induced genes (termed cellular immed- iate-early genes) whose function is to couple extracellular stimulation to long- term responses. In the brain, these genes muy contribute to the adaptive alterations inidred in neuronol plusticitji.
Introduction
A cell responds to signal molecules and cues in its environment with an inte- grated response that results from the coupling of extracellular stimuli to intracellular effector pathways. In gen- eral. extracellular stimuli evoke two types of responses in target cells. The first are rapid, transcription-indepen- dent, events that constitute the cell's immediate reaction to the stimulus. Such responses occur in the millisecond to minute time-frame. The second type of response involves longer-term, tran- scription-dependent, alterations that may represent, at one extreme, a stable change in the differentiated state of a cell or, in less dramatic circumstances, a persistent modification of the cell's im- mediate reaction to the primary stimu- lus. These alterations occur within hours and days and, depending upon the circumstances, may be termed dif- ferentiation or adaptation. While there is a wealth of knowledge regarding the molecular and biophysical mechanisms that link cell stimulation to short-term responses (e.g. second messengers such as cyclic A M P and calcium ions), little is known regarding the pathways that underlie stimulus-transcription coup- ling. This has led to a quest both for the molecules that couple cell stimulation to transcriptional activation and for the genes implicated in, and responsible for, long-term responses.
Fos and Stimulus-Transcription Coupling
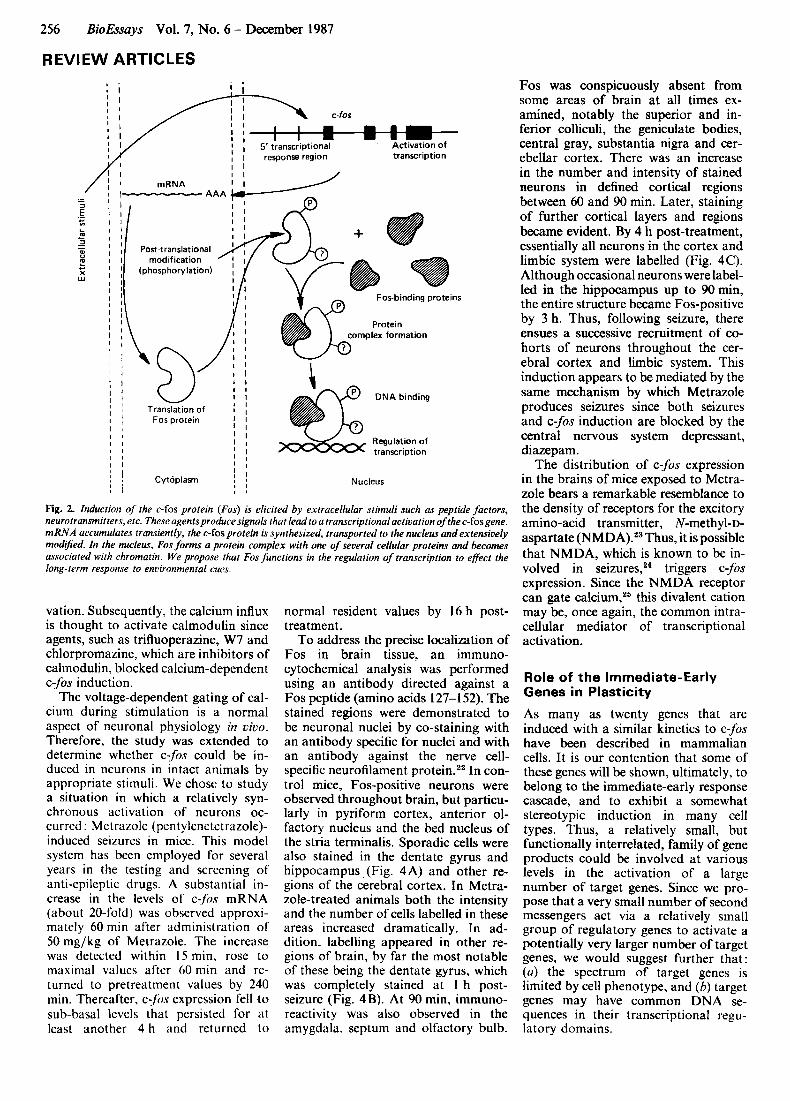
A general model for the coupling of stimuli to both long- and short-term responses is depicted in Fig. 1. In this scheme the stimulus brings about alter- ations in the level or activity of second messenger molecules. The second mes- sengers then elicit short-term responses directly and, by way of adaptive regu- lator molecules, bring about the long- term response. In molecular terms, one can envisage the adaptive regulators as proteins that modulate the expression of genes involved in the long-term response. The discovery that several proto-oncogenes provide key functions in signal transduction processes' has provided candidate genes for the role of adaptive regulators. In particular, the proto-oncogenes encoding nuclear pro- teins, such as c-fos, have many appro- priate characteristics. Induction of c-fos occurs rapidly and transiently following treatment of cells with polypeptide growth factors and other agents.'.'' The c:fos protein (Fos) undergoes extensive post-translational m~dification,~ forms complexes with several cellular pro- teins5 and binds to DNA6 (Fig. 2). The v-f0.s gene has been shown to have the ability to stimulate transcription from selected promoters in trans.' Most compelling is the observation of Spiegel- man and colleagues that Fos can bind to a specific D N A sequence in the 5' region of a gene that undergoes tran- scriptional regulation during adipocyte differentiation." A number of laboratories have iso-
lated as many as twenty genes that, like cji)s, are rapidly induced even in the presence of protein synthesis inhibit- ors.!' I ' We propose that c:JOS and the other inducible genes are components of a poorly-defined, signal-transcription
Short-term
Stimulus -Second messengers\
Adaptive- Long-term regulators response
Fig. 1. Prcipcised rnrchunism ,for .v~imulu.c-re.vpon.vc coupling.
coupling cascade that links extracellular stimuli to long-term adaptive responses (Fig. 3). The immediate-early genes of bacteriophage and eukaryotic viruses provide an interesting parallel : they are rapidly synthesized following infection of a host cell, their transcription does not require protein synthesis, and they generally function in regulating ex- pression of other viral By analogy to this situation we and others have referred to the rapidly induced genes as cellular immediate-early genes (Fig. 3).
Fos Induction in Brain
One of the most notable examples of a long-term alteration in cellular response is that of learning. Indeed, this phen- omenon points to many of the proper- ties required by an adaptive mechanism, as discussed cogently by Kandel and colleague^.'^ A critical finding in a learning paradigm in Aplysia has been the discovery that transient inhibition of protein synthesis during, and im- mediately after, stimulation specifically eliminates long-term facilitation.lS This observation predicts a requirement for the synthesis of certain gene products for long- but not short-term responses.
Initial insights into the coupling of stimulation to cyfos expression in neur- ons were made using PC12 cells in cul- ture. PC12 cells have been utilized as an in ilitro model for neurodevelopment as they differentiate in response to the neurotrophic agent, nerve growth factor (NGF) and possess many charac- teristics of neurons.'' Thus, PC12 cells provide an attractive system for investi- gating stimulus-transcription coupling by non-mitogenic stimuli. Our stud- ies,". '" and those of others,'!' " demon- strated a rapid induction of c-f0.s by NFG, neurotransmitters and substances that cause membrane depolarization. The agents that elicited ~ ~ J 0 . s expression by depolarization all required extra- cellular calcium and were blocked by antagonists of voltage-dependent cal- cium channe1s.l' Since agonists of this channel could stimulate cTfi)s expression directly in a calcium-dependent manner, i t was concluded that the ingress of calcium evoked transcriptional acti-
256 BioEssays Vol. 7, No. 6 - December 1987
REVIEW ARTICLES I .
I I c-fos I '
transcription , I
1
I
mRNA 2 / AAA I- fa
Fos-binding proteins
(phosphorylation)
Protein complex formation
DNA binding
i Fos protein
Regulation of I transcription I
Fig. 2. Induction of the c-fos protein (Fos) is elicited by extracellular stimuli such as peptide factors, neurotransmitters, etc. These agentsproducesignals that leadto a transcriptional activation of the c- fosgene. mRNA accumulates transiently, the c-fos protein is synthesized, iransported to the nucleus and extensively modified. In the nucleus. Fos forms a protein complex with one of several cellular proteins and becomes associated with chromatin. We propose that Fos functions in the regulation of transcription to effect the long-term response to environmental cues.
vation. Subsequently, the calcium influx is thought to activate calmodulin since agents, such as trifluoperazine, W7 and chlorpromazine, which are inhibitors of calmodulin, blocked calcium-dependent c-fos induction.
The voltage-dependent gating of cal- cium during stimulation is a normal aspect of neuronal physiology in vivo. Therefore. the study was extended to determine whether c$os could be in- duced in neurons in intact animals by appropriate stimuli. We chose to study a situation in which a relatively syn- chronous activation of neurons oc- curred : Metrazole (pentylenetetrazo1e)- induced seizures in mice. This model system has been employed for several years in the testing and screening of anti-epileptic drugs. A substantial in- crease in the levels of c+.s mRNA (about 20-fold) was observed approxi- mately 60 min after administration of 50 mg/kg of Metrazole. The increase was detected within 15 min. rose to maximal values after 60 min and rc- turned to pretreatment values by 240 min. Thereafter. c+.s expression fell to sub-basal levels that persisted for at least another 4 h and returned to
normal resident values by 16 h post- treatment.
To address the precise localization of Fos in brain tissue, an immuno- cytochemical analysis was performed using an antibody directed against a Fos peptide (amino acids 127-1 52). The stained regions were demonstrated to be neuronal nuclei by co-staining with an antibody specific for nuclei and with an antibody against the nerve cell- specific neurofilament protein.22 In con- trol mice, Fos-positive neurons were observed throughout brain, but particu- larly in pyriform cortex, anterior ol- factory nucleus and the bed nucleus of the stria terminalis. Sporadic cells were also stained in the dentate gyrus and hippocampus, (Fig. 4A) and other re- gions of the cerebral cortex. In Metra- zole-treated animals both the intensity and the number of cells labelled in these areas increased dramatically. In ad- dition, labelling appeared in other re- gions of brain, by far the most notable of these being the dentate gyrus, which was completely stained at 1 h post- seizure (Fig. 4B). At 90 min, immuno- reactivity was also observed in the amygdala. septum and olfactory bulb.
Fos was conspicuously absent from some areas of brain at all times ex- amined, notably the superior and in- ferior colliculi, the geniculate bodies, central gray, substantia nigra and cer- ebellar cortex. There was an increase in the number and intensity of stained neurons in defined cortical regions between 60 and 90 min. Later, staining of further cortical layers and regions became evident. By 4 ti post-treatment, essentially all neurons in the cortex and limbic system were labelled (Fig. 4C). Although occasional neurons were label- led in the hippocampus up to 90min, the entire structure became Fos-positive by 3 h. Thus, following seizure, there ensues a successive recruitment of co- horts of neurons throughout the cer- ebral cortex and limbic system. This induction appears to be mediated by the same mechanism by which Metrazole produces seizures since both seizures and c-fos induction are blocked by the central nervous system depressant, diazepam.
The distribution of c-fos expression in the brains of mice exposed to Metra- zole bears a remarkable resemblance to the density of receptors for the excitory amino-acid transmitter, N-methyl+- aspartate (NMDA).23 Thus, it is possible that NMDA, which is known to be in- volved in seizures,24 triggers c-fus expression. Since the NMDA receptor can gate calcium,25 this divalent cation may be, once again, the common intra- cellular mediator of transcriptional activation.
Role of the Immediate-Early Genes in Plasticity
As many as twenty genes that are induced with a similar kinetics to c-fos have been described in mammalian cells. It is our contention that some of these genes will be shown, ultimately, to belong to the immediate-early response cascade, and to exhibit a somewhat stereotypic induction in many cell types. Thus, a relatively small, but functionally interrelated, family of gene products could be involved at various levels in the activation of a large number of target genes. Since we pro- pose that a very small number of second messengers act via a relatively small group of regulatory genes to activate a potentially very larger number of target genes, we would suggest further that : (a) the spectrum of target genes is limited by cell phenotype, and (h) target genes may have common DNA se- quences in their transcriptional regu- latory domains.

Bio Essays Vol. 7, No. 6 - December 1987 257
REVIEW ARTICLES
Cellular consequences 4
Long-term n responses
A
Cytoplasm I Nucleus
Fig. 3. Proposed model for the role of cellular immediate-early genes in stimulus-response coupling. Extracellular ligands (L) interact with receptors (R) and activate secondmessenger systems via membrane- transducing components (T). Subsequently, second messengers elicit rapid alterations that constitute a short-term response. In addition, the same second messengers, directly or indirectly, act to indurn transcription of the cellular immediate-early genes. We have used c-fos as a marker for the immediate- early cellular response; however, we propose that a set of such genes exists (nominally designated A-C here). This transcriptional activation could have a number of consequences. First, immediate-early gene products may be components or regulators of the signal transduction cascade (for example, receptors, G proteins, kinases, phosphatases, etc.). This would permit the cell to modify its response to repeated stimulation. Second, immediate-early genes may promote the transcriptional activation of further target genes (designated 1-7) that cause changes in long-term responses. In the nervous system these target genes might encode ion channels, neuropeptide precursors, synaptic proteins or extracellular matrix proteins. Potential regulation of immediate-early genes by negative feedback control is indicated by dotted lines.
In studying c-fos induction in PC12 cells and fibroblasts, we have detected a set of inducible gene products that share a conserved epitope with c-fos.'6 The pattern of proteins obtained fol- lowing stimulation seemed to be de- pendent on the inducing agent used. This led to the notion that the precise pattern of immediate-early genes seen following stimulation would be a func- tion both of cell type and inducing agent. The integration of the effects of these gene products would then deter- mine the cellular response. Further levels of complexity are possible. Fos undergoes differential post-trans- lational modification depending on the inducing agent,27 which could affect its function, and it binds to cellular pro- teins that differ between cell types.26 In this way, a small number of immediate- early genes could, by complexing to different cellular proteins and under- going various post-translational modifi- cations, generate an enormous reper- toire of responses.
In terms of neuronal plasticity, the data obtained in our seizure studies lead to a number of specific embellishments of the model. It was noted that, fol- lowing the administration of Metrazole, a subsequent dose was relatively in-
effective at re-inducing c - j i ~ s . ~ ~ This situation persisted for as long as the Fos protein was present in neurons, suggesting that Fos or one of the immediate-early genes might repress induction. This would not be without precedent amongst viral immediate- early genes and would provide a mech- anism for regulating Fos levels strin- gently. Indeed, one postulate for any regulatory mechanism is that the signal should be relatively short-lived. We have observed that c-fos is transiently activated, subject to negative feedback regulation and its mRNA and protein have short half-lives. In addition, Fos and the Fos-related antigens are syn- thesized during the same time period identified as that in which protein synthesis is obligatory for the establish- ment of long-term alterations in neural function.
In the dentate gyrus, a prime location for c-fos induction following seizure, repeated induction of seizure causes alterations in the level of expression of several neurotransmitters and their receptors." Specific examples are the y-aminobutyric acid (GABA) and the muscarinic acetylcholine receptors that are candidate target genes subject to regulation by the immediate-early re-
Fig. 4. Fos is synthesized in particular subsets of neurons following seizure. Mice received a single injection of Metrazole (50 mglkg). Animals were anesthetized immediately (A), I h later ( B ) or 3 h later (C). They were then perfused with para- formaldehyde and tissues were processedfor immune cytochemistry asdescribedpreviously.~2 The coronal sections shown here were maak through similar regions of the hippocampus. The control section (A) shows sporadic hbellmg of neuronal nuclei in the dentate gyms (dg). One hour after administration of Metrazole essentially all neurons in the dentate gyrus (dg) are positive andsome staining is detected in the hippocampus (hc) (B). A@er 4 h, the dentate gyrus (dg), hippocampus (hc), and several layers of the cortex (c) are Fos-positive (C).
sponse genes in neurons. In a more general sense, we would predict that other genes would include those en- coding proteins involved in synaptic structure and efficiency, neurotrans- mitter synthesis and ion conduct- ances. Thus, in this model, learning and memory, and other persistent changes in the central nervous system would have their basis in the regulation of gene expression.
With the demonstration of several inducible genes in fibroblasts, we would predict that, given the complexity of the central nervous system, a plethora of neuronal immediate-early genes await discovery. The seizure model described for c-fos induction provides an experi-

258 BioEssays Vol. 7 , No. 6 - December 1987
REVIEW ARTICLES
mental paradigm for their identification and isolation.
REFER EN CES 1 HELDIN, C.-H. & WESTERMARK, B. (1984). Growth factors: mechanism of action and relation to oncogenes. Cell 3l, 9-20. 2 CURRAN, T., BRAVO, R. & MULLER, R. (1985). Transient induction of c-fos and c-myc is an immediate consequence of growth factor stimulation. Cancer Sur- ueys 4, 655681. 3 CURRAN, T. (1987). fos. In The Oncogene Handbook (ed. E. P. Reddy, A. M. Skalka and T. Curran), Elsevier, Amsterdam (in press). 4 C ~ R A N , T., MILLER, A. D., Zorc~?,, L. & VERMA, I. M. (1984). Viral and cellularfos proteins: a comparative analysis. Cell 36, 25%268. 5 CURRAN, T., VAN BEVEREN, C., LING, N. & VERMA, I. M. (1985). Viral and cellularfos proteins are complexed with a 39,000 dalton protein. Mol. Cell. Bid . 5,167-172. 6 SAMBUCETTI, L.C. & CURRAN, T. (1986). The Fos protein complex is associated with DNA in isolated nuclei and binds to DNA cellulose. Science 234, 1417-1419. 7 SETOYAMA, C., FRUNZIO, R., LIAN, G., MUDRYII, M. & DE CROMBRUGGHE, B. (1986). Transcriptional activation encoded by the v-fos gene. Proc. Natl. Acad. Sci. USA 83,3213-3217. 8 DISTEL, R. J., Ro, H.-S., ROSEN, B. S., GROW, D. L. & SPIEGELMAN, B. M. (1 987). Nucleoprotein complexes that regulate gene expression in adipocyte differentiation : direct participation of c-fos. Cell 49, 835-844. 9 COCHRAN, B. H., REFFEL, A. C. &STILES, C. D. (1983). Molecular cloning of gene sequences regulated by platelet-derived growth factor. Cell 33,939-947. 10 LAU, L. F. & NATHANS, D. (1987). Expression of a set of growth-related immediate-early genes in BALB/c
3T3 cells: coordinate regulation with c-Jos and c-myc. Proc. Natl. Acad. Sci. USA 84, 1 182-1 186. 11 LIM, R. W., VARNUM, 9. C. & HERSCHMAN, H. R. (1987). Cloning of tetradecanoyl phorbol ester induced 'primary response' sequences and their expression in density-arrested Swiss 3T3 cells and a TPA non- proliferative variant. Oncogene (in press). 12 GRASSO, R. J. & BUCHANAN, J. M. (1969). Synthesis of early RNA in bacteriophage TCinfected E. coli 9. Nature 224, 882-886. 13 RAKUSANOVA, T., BEN-PORAT, T., HIMENO, M. & KAPLAN, A. S. (1971). Early functions of the genome of herpesviruses. I. Characterization of the RNA syn- thesized in cycloheximide-treated infected cells. Virology 46, 877-889. 14 GOELET, P., CASTELUCCI, V. F., SCHACHER, S. & KANDEL, E. R. (1985). The long and the short of long- term memory - a molecular framework. Nature 322, 4 19-422. 15 MONTAROLO, P. G., GOELET, P., CASTELLUCCI, V. F., MORGAN, J., KANDEL, E. R. & SCHACHER, S. (1987). A critical period for macromolecular synthesis in long-term heterosynaptic facilitation in Aplysia. Science 239, 12494255. 16 GREENE, L. A. & TISCHLER, A. S. (1982). PC12 pheochromocytoma cultures in neurobiological research. Aduanc. Cell. Neurobiol. 3, 373414. 17 CURRAN, T. & MORGAN, J . I. (1985). Superinduction of c-fos by nerve growth factor in the presence of peripherally active benzodiazepines. Science 229, 1265- 1268. 18 MORGAN, J. I. & CURRAN, T. (1985). Role of ion flux in the control of c-fos expression. Nature 322, 552-555. 19 GREENBERG, M. E., GREENE, L. A. & ZIFF, E. B. (1985). Nerve growth factor and epidermal growth factor induce rapid changes in proto-oncogene transcription in PC12 cells. J . Biol. Chem. 260, 14101-14110.
(1986). Stimulation of neuronal acetylcholine receptors induces rapid gene transcription. Science 234, 80-83.
20 GREENBERG, M. E., ZIFF, E. B. & GENE, L. A.
21 KRUIIER, W., SCHUBERT, D. & VERMA, 1. M. (1985). Induction of the proto-oncogene fos by nerve growth factor. Proc. Natl. Acad. Sci. USA 82, 733C7334. 22 MORGAN, J. I., &HEN, D. R., HEMPSTEAD. J. L. & CURRAN, T. (1987). Mapping patterns of c-fos expression in the central nervous system after seizure. Science 237, 192-197. 23 WONG, E. H. F., KEMP, J. A., PRIESTLY, T., KNIGHT, A. R., WOODRUFF, G. N. & IVERSEN, L. L. (1986). The anticonvulsant MK-861 is a potent N-methyl-o-aspartate antagonist. Proc. Natl. Acad. Sci. USA 83, 7 104-7108. 24 MODY, I. & HEINEMANN, U. (1987). NMDA receptors of dentate gyrus granule cells participate in synaptic transmission following kindling. Nature 326, 701-704. 25 ASCHER, P. & NOWAK, L. (1987). Electrophysio- logical studies of NMDA receptors. Trends in Neurosci. 10, 284288. 26 FRANZA, B. R., JR., SAMBUCETTI, L. C., Corn, D. R. & CURRAN, T. (1987). Analysis of Fos protein complexes and Fos-related antigens by high-resolution two-dimensional gel electrophoresis. Oncogene 1, 213- 221. 27 CURRAN, T. & MORGAN, J. I. (1986). Barium modu- lates c-fos expression and post-translational modifi- cation. Proc. Natl. Acad. Sci. USA 83, 8521-8524. 28 SHIN, C., F'EDERSON, H. B. & MCNAMARA, J.O. (1985). y-aminobutyric acid and benzodiazepine re- ceptors in the kindling model of epilepsy: a quantitative radiohistochemical study. J. Neurosci. 5, 26962701
T O M C U R R A N and JAMES I . M O R G A N are at the Roche Institute of Molecular Biology, Nutley, NJ 07110. USA
Ribosomal Protein Autoantibodies in Systemic Lupus Erythematosus Keith Elkon, Eloisa Bonfa, Susan Skelly, Herbert Weissbach and Nathan Brot
Summary
Autoantibodies to three eukaryotic 60s ribosomal phosphoproteins PO, PI and P2 have been found in the sera of 10-20 yo ofpatients with systemic lupus erythema- tosus (SLE). These three proteins share a common epitope contained within the carboxyl terminal 22 amino acids of each protein. Because central nervous system disturbances, with major behavioural disorders, occur in a signiJicant fraction of SLE patients, the antiribosomal auto- antibodies were measured in this subset of SLE individuals to determine whether or not there was an association. This anti- body is present in 90 yo of SLE patients who were diagnosed as having psychosis, secondary to the disease.
Introduction
A large number of diseases are thought to be autoimmune in origin. These
diseases include thyroiditis, rheumatoid arthritis, juvenile diabetes, myasthenia gravis, scleroderma, polymyositis and systemic lupus erythematosus (SLE). SLE is often considered a prototype of autoimmune disorders and is generally characterized by the presence of serum autoantibodies directed against nucleo- protein complexes and DNA.l In ad- dition, it has been known for some time that about 1&20~0 of SLE patients' sera contain antibodies against cyto- plasmic constituents.'S3 Although one of these cytoplasmic constituents was known to be the ribo~ome,~-ll only recently have the specific antigenic ribosomal components been identi- fied.12313 In this article we will describe the specific epitope present on three 60s ribosomal phosphoproteins called PO, PI and P212.13 that SLE anti-ribosome antibodies recognize.
Reactivity of SLE Sera with PO, P1 and P2
It was initially observed that sera from SLE patients with antibodies against cytoplasmic components reacted specifi- cally with three ribosomal proteins.", l3
Fig. 1A shows, by immunoblot analy- sis, the reactivity of five of these patients' sera with three ribosomal proteins of 38, 19 and 17 kDa. These three proteins have been identified as 60s ribosomal proteins PO, P1 and P2, respectively. This identification was based on their migration on two-dimensional gel elec- trophoresis (Fig. 1 B) which is identical to that of the authentic proteins, as well as by the changes in migration that were observed when the proteins were treated with alkaline phosphatase.12 Finally, the pattern of the two-dimensional immunoblot using human antisera was identical to that observed with a mouse monoclonal anti-PO, P1, and P2 (anti-





























