Embed Size (px)
Citation preview

1
Membrane permeable C-terminal dopamine transporter peptides attenuate amphetamine-evoked dopamine release*
Mattias Rickhag1, William A. Owens2, Marie-Therese Winkler3, Kristine Nørgaard Strandfelt1, Mette Rathje1, Gunnar Sørensen1,4, Bjørn Andresen1, Kenneth L. Madsen1, Trine Nygaard Jørgensen1, Gitta
Wörtwein4, David P.D. Woldbye4, Harald Sitte3, Lynette C. Daws2 and Ulrik Gether1
1Molecular Neuropharmacology Laboratory, Lundbeck Foundation Center for Biomembranes in Nanomedicine, Department of Neuroscience and Pharmacology, Faculty of Health and Medical Sciences,
University of Copenhagen, Copenhagen, Denmark, 2Department of Physiology, University of Texas Health Science Center at San Antonio, San Antonio, Texas, USA, 3Institute of Pharmacology, Center for
Physiology and Pharmacology, Medical University Vienna, Vienna, Austria, 4Laboratory of Neuropsychiatry, Faculty of Health and Medical Sciences, Department of Neuroscience and
Pharmacology, University of Copenhagen, Copenhagen, Denmark
*Running title: DAT C-terminus interactions and AMPH-evoked DA release
To whom correspondence should be addressed: Ulrik Gether, Department of Neuroscience and Pharmacology, Panum Institute 18.6, Blegdamsvej 3, DK-2200 Copenhagen N, Denmark. Tel: +45 23840089. Fax: +45 35327610. E-mail: [email protected] Background: The significance of dopamine transporter (DAT) C-terminal protein-protein interactions for amphetamine-induced dopamine efflux is unsettled. Results: Cell-permeable C-terminal DAT peptides attenuate amphetamine-induced dopamine efflux and locomotor activity in mice. Conclusion: DAT C-terminal protein-protein interactions are critical for the effects of amphetamine in vivo. Significance: Targeting protein-protein interactions might be a way of inhibiting the effects of psychostimulants. SUMMARY The dopamine transporter (DAT) is responsible for sequestration of extracellular dopamine (DA). The psychostimulant amphetamine (AMPH) is a DAT substrate, which is actively transported into the nerve terminus, eliciting vesicular depletion and reversal of DA transport via DAT. Here, we investigate the role of the DAT C-terminus in AMPH-evoked
DA efflux using cell-permeant dominant-negative peptides. A peptide, which corresponded to the last 24 C-terminal residues of DAT (TAT-C24 DAT) and thereby contained the CaMKIIα (Ca2+-calmodulin dependent protein kinase IIα) binding domain and the PDZ (PSD-95/Discs-large/ZO-1) binding sequence of DAT, was made membrane-permeable by fusing it to the cell-membrane transduction domain of the HIV-1 Tat protein (TAT-C24WT). The ability of TAT-C24WT but not a scrambled peptide (TAT-C24Scr) to block the CaMKIIα-DAT interaction was supported by co-immunoprecipitation experiments in heterologous cells. In heterologous cells, we also found that TAT-C24WT, but not TAT-C24Scr, decreased AMPH-evoked MPP+ efflux. Moreover, chronoamperometric recordings in striatum revealed diminished AMPH-evoked DA efflux in mice preinjected with TAT-C24WT. Both in heterologous cells and in striatum, the peptide did not further inhibit efflux upon KN-93-mediated inhibition of
http://www.jbc.org/cgi/doi/10.1074/jbc.M112.441295The latest version is at JBC Papers in Press. Published on July 24, 2013 as Manuscript M112.441295
Copyright 2013 by The American Society for Biochemistry and Molecular Biology, Inc.
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

2
CaMKIIα activity, consistent with a dominant-negative action preventing binding of CaMKIIα to the DAT C-terminus. This was further supported by the ability of a peptide with perturbed PDZ-binding sequence, but preserved CaMKIIα binding (TAT-C24AAA), to diminish AMPH-evoked DA efflux in vivo to the same extent as TAT-C24WT. Finally, AMPH-induced locomotor hyperactivity was attenuated following systemic administration of TAT-C24WT but not TAT-C24Scr. Summarized, our findings substantiate that DAT C-terminal protein-protein interactions are critical for AMPH-evoked DA efflux and suggest that it may be possible to target protein-protein interactions to modulate transporter function and interfere with psychostimulant effects. Dopamine (DA) plays a pivotal role as a modulatory neurotransmitter controlling motor function, cognitive processes, reward mechanisms as well as neuroendocrine secretion. Aberrant DA signaling contributes to the development of neuropsychiatric diseases such as schizophrenia, drug addiction and Parkinson’s disease (1,2). DA homeostasis in the brain is regulated by the presynaptic DA transporter (DAT), which removes extracellular DA from the synaptic cleft and thereby maintains homeostatic transmitter levels. DAT belongs to the solute carrier 6 (SLC6) gene family of sodium/chloride-coupled transporters (also referred to as the family of Neurotransmitter:Sodium:Symporters) character-ized by 12 transmembrane spanning domains with both the N- and C-termini located within the cytosol (3,4). Deletion of DAT in genetically modified mouse models has shown that DAT is fundamental for regulating spatio-temporal availability of DA (1,5). DAT is also a molecular substrate for the psychostimulants cocaine and amphetamine (AMPH). Whereas cocaine acts as a competitive DAT inhibitor (6), AMPH is a substrate, which is actively transported into the presynaptic nerve terminal, leading to vesicular depletion of DA and reverse transport of DA via DAT (7,8). This DA efflux dramatically elevates extracellular DA levels and is believed to contribute substantially to the addictive properties
of AMPH (9). AMPH also causes DAT to move away from the plasma membrane to the intracellular compartment, thereby decreasing DAT surface levels (10-12). Interestingly, AMPH-induced endocytosis of DAT has been suggested to occur through a mechanism requiring stimulation of Ca2+-calmodulin dependent protein kinase IIα (CaMKIIα) (13).
Several DAT-interacting proteins have been identified and suggested to be involved in regulating trafficking or the transport activity (3,4,14,15). Both CaMKIIα and the PDZ (PSD-95/Discs-large/ZO-1 homology) domain protein PICK1 (protein interacting with C kinase 1) have been shown to associate with the distal C-terminus of DAT (16). Whereas the last 3-4 C-terminal residues of DAT constitute a prototypical PDZ binding sequence that interacts with the PDZ domain of PICK1, the CaMKIIα binding domain consists of the last 24 C-terminal residues of DAT (17). Although it has been proposed to mediate surface targeting of DAT (16), the functional significance of PICK1 binding to DAT remains uncertain (18,19). CaMKIIα, however, has been implicated in phosphorylation of serines in the DAT N-terminus (17,20,21). Interestingly, phoshorylation of these serines is critical for AMPH-induced DAT-mediated DA efflux by shifting the transporter from a “reluctant” to a permissive state for efflux without influencing uptake activity (22). We previously showed that this phosphorylation is facilitated by a direct association of CaMKIIα to DAT. Hence, we obtained evidence for a model in which CaMKIIα binding to a domain corresponding to the last 24 C-terminal residues facilitates N-terminal phosphorylation and thereby AMPH-induced DA efflux (17). The model concurs with data suggesting that CaMKIIα is activated upon AMPH administration (23,24) possibly as a consequence of elevated intracellular calcium levels through yet poorly understood mechanisms (25). Impaired AMPH-induced efflux in synaptosomes from mice with ablated CaMKIIα activity has provided further indirect support for the model (26). Moreover, the model suggests, in agreement with several earlier observations (8,27), that AMPH-induced DA efflux is not directly coupled to
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

3
uptake and that inward and outward transport represent different functional modes of the transporter.
Despite the presently available data, the significance of CaMKIIα association to the DAT C-terminus remains uncertain. Moreover, it remains to be determined whether disrupting such C-terminal protein-protein interactions might represent a possible pharmacological target for blunting the psychostimulatory effects of AMPH. To address these questions, we designed cell-permeable peptides corresponding to the DAT C-terminus and tested their effect in DAT expressing heterologous cells as well as in vivo. Our data demonstrate that a peptide corresponding to the last 24 C-terminal residues of DAT, which constitutes the CaMKIIα binding domain, can act in a dominant-negative fashion causing impaired AMPH-induced MPP+ efflux in heterologous cells and decreased AMPH-induced DA efflux in the striatum of living mice. A similar effect was observed for a peptide with perturbed PDZ binding sequence suggesting that binding of PDZ domain protein to DAT is less important for efflux than CaMKIIα. Co-immunoprecipitation experiments validated the interaction between DAT and CaMKIIα and we further showed that the cell permeable DAT C-terminus peptide disrupted the interaction in heterologous cells. Furthermore, systemic administration of the C-terminal WT DAT peptide, but not a scrambled peptide, attenuated AMPH-stimulated locomotor activity in mice.
EXPERIMENTAL PROCEDURES Immunocytochemistry - Synthetic peptides were purchased from Schafer N (Copenhagen, Denmark). The amino acid sequence was H-YGRKKRRQRRRPEKDRELVDRGEVRQFTLRHWLKV-OH for TAT-C24WT, H-YGRKKRRQRRRPEKDRELVDRGEVRQFTLRHWAAA-OH for TAT-C24AAA and H-CYGRKKRRQRRRKRTPFLLFVQEVDWRDLKVGHRER-OH for TAT-C24Scr peptide. Biotinylated TAT-peptide variants (TAT-C24WT and TAT-C24Scr) were dissolved in 0.9 % saline and immediately frozen in aliquots (10 mg/ml; corresponds to TAT-C24WT: 2.1 mM and for TAT-C24Scr: 2.2 mM). The TAT-peptide aliquots
were thawed on ice right before use. HEK293 cells were seeded in six-well plates and biotinylated peptides (TAT-C24WT and TAT-C24Scr; final concentration 1 µM) were added to the respective wells. Following 1h incubation with TAT-peptides at 37˚C, cells were fixed in 4% paraformaldehyde and rinsed in phosphate buffered saline (PBS). Fluorescein streptavidin (1:50, Vector Laboratories Inc., USA) was incubated with cells for 3h in PBS containing 0.1 % Triton X-100 at room temperature. This was followed by additional rinsing in PBS and cells were finally mounted on glass slides with ProlongGold antifade reagent (Molecular Probes, Invitrogen). TAT-peptides were visualized using a Zeiss LSM 510 confocal laser-scanning microscope with an oil immersion 63×1.4 numerical aperture objective (Carl Zeiss). Streptavidin fluorescein was excited with a 488nm laserline from an argon-krypton laser, and detection of the emitted light was done using a 505-530 nm bandpass filter. Images were analyzed using the IMAGEJ software. Co-immunoprecipitation – T-Rex HEK293 cells (human embryonic kidney) transfected to stably express CaMKIIα in a tetracycline-dependent manner (based on stable expression of Tet repressor from pcDNA6/TR) and with stabile expression of human DAT (hDAT, N-terminally tagged with YFP) were cultivated as previously described (T-Rex 293 CaMKIIα YFP-hDAT cells) (17). Cells were maintained at 37˚C in a humidified 5% CO2 atmosphere. Confluent cells in 75 cm2 flasks were preincubated with TAT-C24WT (1 µM), TAT-C24Scr (1 µM) or vehicle for 1h before the cells were rinsed in Tris-buffered saline (TBS) followed by solubilization in ice cold lysis buffer containing 1% (v/v) Triton X-100, 20 mM Tris-HCl pH 8.0, 150 mM NaCl, 1 mM sodium orthovanadate, 5 mM NaF, 5 mM sodium pyrophosphate and a protease inhibitor cocktail (Roche Diagnostics). Cell lysates were centrifuged at 16,000 x g for 15 min at 4°C to remove cell debris and supernatant was then diluted to 1 mg/ml. The supernatant was incubated with rat anti-DAT antibody (MAB369 1:1000, Millipore) for 1h at 4°C during constant rotation. 25 µl Protein-G agarose beads (Roche Diagnostics) were
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

4
washed once with lysis buffer and then incubated with supernatant for 2h at 4°C during constant rotation. As control for these experiments, pull-down without primary antibody and cell lysates without TAT-C24WT/Scr preincubation were performed in parallel. Subsequently, beads were washed twice in TBS containing 0.1% (v/v) Triton X-100 and then eluted using 5X loading buffer. Proteins were separated by SDS-PAGE and transferred to Immobilon-P membranes. Membranes were blocked in PBS containing 0.05% Tween-20 and 5% dry milk, and incubated overnight with antibodies against rabbit phosphorylated CaMKIIα (Thr 286, 1:1000, Santa Cruz Biotechnology, USA). Following incubation with HRP-conjugated anti-rabbit antibodies, the blots were visualized by chemiluminescence (Ecl-kit, Amersham). To normalize pull-down of pCaMKIIα, blots were stripped and re-probed for rat anti-DAT (MAB369, 1:1000, Millipore) to determine DAT immunoprecipitation for each condition. CaMKIIα co-IP were normalized to TAT-C24Scr for each experiment and then one- sample t test was used for statistical analysis (p<0.05). Biotinylation of surface proteins - T-Rex 293 CaMKIIα YFP-hDAT cells were seeded two days before experiment in pre-coated 6-well plates at a density of 500,000 cells/well. On the day of experiment, cells were preincubated with 1 µM TAT-C24WT, TAT-C24Scr or vehicle (control) (without peptide treatment) for 1h. Cells were rinsed twice in cold PBS and then incubated with sulfo-NHS-biotin (Thermo Scientific) for 40 min on ice. Rinsing twice in 100 mM glycine was used to quench excess biotin and this was followed by additional washing. Cells were lysed in 500 µl solubilization buffer containing 1% (v/v) Triton X-100, 25 mM Tris-HCl, 150 mM NaCl, 1 mM EDTA, 0.2 mM PMSF, 5 mM NEM and a protease inhibitor cocktail (Roche Diagnostics). Cell lysates were centrifuged at 16,000 x g for 15 min at 4°C to remove cell debris and supernatant was collected. An aliquot was removed to determine protein concentration using BCA assay. 175 µl avidin beads (Thermo Scientific) were washed once in 0.8 ml lysis buffer and 300 µl
protein extracts were then incubated with the avidin beads overnight at 4°C during constant rotation. The beads were washed four times in lysis buffer and then eluted using SDS sample buffer containing 100 mM DTT. Total protein extracts and eluted proteins were separated on a SDS-PAGE and transferred to Immobilon-P membranes. Membranes were blocked in PBS containing 0.05% Tween-20 and 5% dry milk, and incubated overnight with a monoclonal rat anti-DAT antibody (MAB369, Millipore). Following incubation with an anti-rat horseradish peroxidase-conjugated secondary antibody, the blots were visualized by chemiluminescence (Ecl-kit, Amersham) and quantified using ImageJ. DA uptake - T-Rex 293 CaMKIIα YFP-hDAT cells were seeded in 24-well plates at 100,000 cells/well on the day prior to the assay. On the day of the experiment, cells were preincubated with TAT-C24WT and Tat-C24Scr at a final concentration of 1 µM for 30 min and then rinsed once in uptake buffer (25 mM HEPES, 120 mM NaCl, 5 mM KCl, 1.2 mM CaCl2, 1.2 mM MgSO4, 1 mM L-ascorbic acid, 5 mM D-glucose and 10 nM cathechol-O-methyl-transferase inhibitor (RO-41-0960), pH 7.4). After addition of pre-warmed uptake buffer and unlabeled DA or AMPH at given concentrations, uptake was initiated by addition of labeled DA (91.1 Ci/mmol) (Perkin Elmer Life Sciences, USA). Following 5 min of incubation at 37˚C, reaction was terminated by washing the cells twice with ice-cold uptake buffer followed by cell lysis in 1% SDS. All samples were transferred to 24-well counting plates, followed by addition of Optiphase Hisafe scintillation fluid (Perkin Elmer). Samples were finally counted in a Wallac Tri-Lux β-scintillation counter (Perkin Elmer Life Sciences). Uptake data were analyzed by non-linear regression analysis using Graph Pad Prism. The IC50 values used in the estimation of Km and Vmax for uptake were calculated from means of pIC50 values and the S.E. interval from the pIC50±S.E. Cell culture and MPP+ efflux - In vitro experiments were carried out using the T-Rex 293 cell with stabile expression of human DAT and
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

5
CaMKIIα (in a tetracycline-dependent manner) (T-Rex 293 CaMKIIα FLAG-hDAT cells) as previously described (17). Efflux experiments were performed in a superfusion system as described using MPP+ as substrate (27). Cells (20 x 105) were seeded on glass coverslips (5 mm in diameter), coated with poly-D-lysine (Sigma), and at this time point (1 day before the experiment) tetracycline was added to induce CaMKIIα expression (17). Prior to the experiment, 0.3 µM of the radioactive labeled DAT substrate [3H]-MPP+ (Perkin Elmer Life Sciences, USA) was added to the cells and incubated for 30 min at 37°C. Some coverslips received freshly dissolved TAT-peptides (TAT-C24WT, TAT-C24Scr and TAT-C24AAA) at a final concentration of 50 nM. The coverslips were transferred to small superfusion chambers (volume = 0.2 ml) and superfused with Krebs-Ringer Hepes buffer at room temperature (25°C) at a rate of 0.7 ml/min for 45 min to establish a stable efflux line (=basal efflux). After basal efflux was stabilized, the experiment was initiated with the collection of fractions every 2 min. After 6 min, 15 µM KN-93 (Calbiochem), a CaMKII-inhibitor, was added to the buffer of the channels indicated; the remaining channels received buffer. After 10 min, AMPH (3 µM, Sigma) was added to all buffers. Finally, the remaining radioactivity was recovered by superfusing the cells for 6 min with 1 % SDS. AMPH-induced release of radioactivity was calculated as percentage of radioactivity present in the cell at any given time point. Chronoamperometry - In vivo high-speed chronoamperometry was conducted using the FAST-12 system (Quanteon LLC, USA) as previously described with some modification (28,29). Recording electrode/micropipette assemblies were constructed using a single carbon-fiber (30 µm diameter (Specialty Materials, USA), which was sealed inside fused silica tubing (Schott, North America). The exposed tip of the carbon fiber (150 µm in length) was coated with 5% Nafion (Aldrich Chemical Co); 3–4 coats baked at 200 °C for 5 min per coat) to provide a 1,000-fold selectivity of DA over its metabolite dihydroxyphenylacetic acid (DOPAC). Under
these conditions, microelectrodes displayed linear amperometric responses to 0.25–10 µM DA during in vitro calibration in 100 mM phosphate-buffered saline (pH 7.4). Male C57Bl/6 mice weighing between 25 and 30 g were anesthetized by intraperitoneal injection (10 ml/kg body weight) of a mixture of urethane (70 mg/ml) and α-chloralose (7 mg/ml), then fitted with an endotracheal tube to facilitate breathing, and placed into a stereotaxic frame (David Kopf Instruments, USA). Body temperature was maintained by a water circulated heating pad. To locally deliver test compounds (see below) close to the recording site, a glass multi-barrel micropipette (FHC, USA) was positioned adjacent to the microelectrode using sticky wax (Moyco, USA). The center-to-center distance between the microelectrode and the micropipette ejector was ~200 µm. The micropipette was filled with AMPH (800 µM, Sigma), the peptide of interest (20 µM; TAT-C24WT, TAT-C24Scr, TAT-C24AAA, or vehicle (aCSF). Artificial CSF and AMPH solutions were prepared fresh each day. The peptides were initially dissolved in DMSO to make 20 mM stock solutions and then frozen. On the day of the experiment, peptide stocks were diluted to 20 µM (0.1% DMSO) in fresh aCSF. The electrode/micropipette assembly was lowered into the striatum at the following coordinates (in mm from bregma [Franklin and Paxinos, 1997): A/P, +1.1; M/L, ±1.4; D/V, −2.25. The application of drug solutions was accomplished using a Picospritzer II (General Valve Corporation, USA) in an ejection volume of 125 nl for AMPH to deliver 100 pmol, and 50 or 100 nl for peptides to deliver 1 or 2 pmol (5–25 psi for 0.25–3 s). After ejection of test agents, there is an estimated 10–200-fold dilution caused by diffusion through the extracellular matrix to reach a concentration of 4–80 µM (AMPH) or 0.1-2 µM (peptide of interest) at the recording electrode (30). It is worth noting that the estimated concentration of AMPH reaching the recording electrode approximates those reported in striatum (2-10 µM), as measured by microdialysis and HPLC, after a single, behaviourally effective systemic injection (31). To record the AMPH-evoked
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

6
efflux of DA at the active electrode, oxidation potentials consisting of 100-ms pulses of 550 mV, each separated by a 1-s interval during which the resting potential was maintained at 0 mV were applied with respect to a Ag/AgCl reference electrode implanted into the contralateral superficial cortex. Oxidation and reduction currents were digitally integrated during the last 80 ms of each 100-ms voltage pulse. For each recording session, DA was identified by its reduction/oxidation current ratio: 0.55–0.80. AMPH and peptides were ejected according to the following sequence. AMPH was pressure- ejected intrastriatally to evoke the release of DA. As soon as DA was cleared from extracellular fluid (ECF); typically 5-30 min after AMPH-application, the peptide of interest or equivalent volume of aCSF was locally applied to striatum, and 45 min and 90 min later the same amount of AMPH was again pressure-ejected. At the termination of each experiment, an electrolytic lesion was made to mark the placement of the recording electrode tip. Mice were then decapitated while still anesthetized, and their brains were removed, frozen on dry ice, and stored at −80°C until sectioned (20 µm) for histological verification of electrode location within the striatum. Data were analyzed using 1-way ANOVA with Dunnett’s multiple comparison test (GraphPad Prism) using the maximal signal amplitude of the released DA (in µM) to define AMPH-evoked DA release. Locomotor activity assessment - Adult mice (C57Bl/6J Charles River Laboratories, USA) were housed under diurnal light conditions with free access to food and water. Mice received 100 µg TAT-C24WT, TAT-C24Scr) or 0.9% saline (NaCl) intravenously. Following 45 min, animals received either 2 mg/kg AMPH or isotonic saline intraperitoneally and were placed in an open-field apparatus. AMPH was obtained from HS Pharmacy (Copenhagen, Denmark) and dissolved in 0.9% saline. The drug was prepared immediately before use. TAT-peptides were dissolved in 0.9 % saline and immediately frozen in aliquots (1 mg/ml). Locomotor activity was assessed using the open-field paradigm consisting of white boxes (40x40x80 cm). Recording of
locomotor activity was performed for 60 min and a video-tracking program (Ethovision 3.1, Noldus Information Technology) was used to analyze distance travelled by the animals. AMPH-induced locomotor hyperactivity was analysed using a repeated measures 2-way ANOVA followed by Bonferroni’s multiple comparison test. Significance level was set to p<0.05.
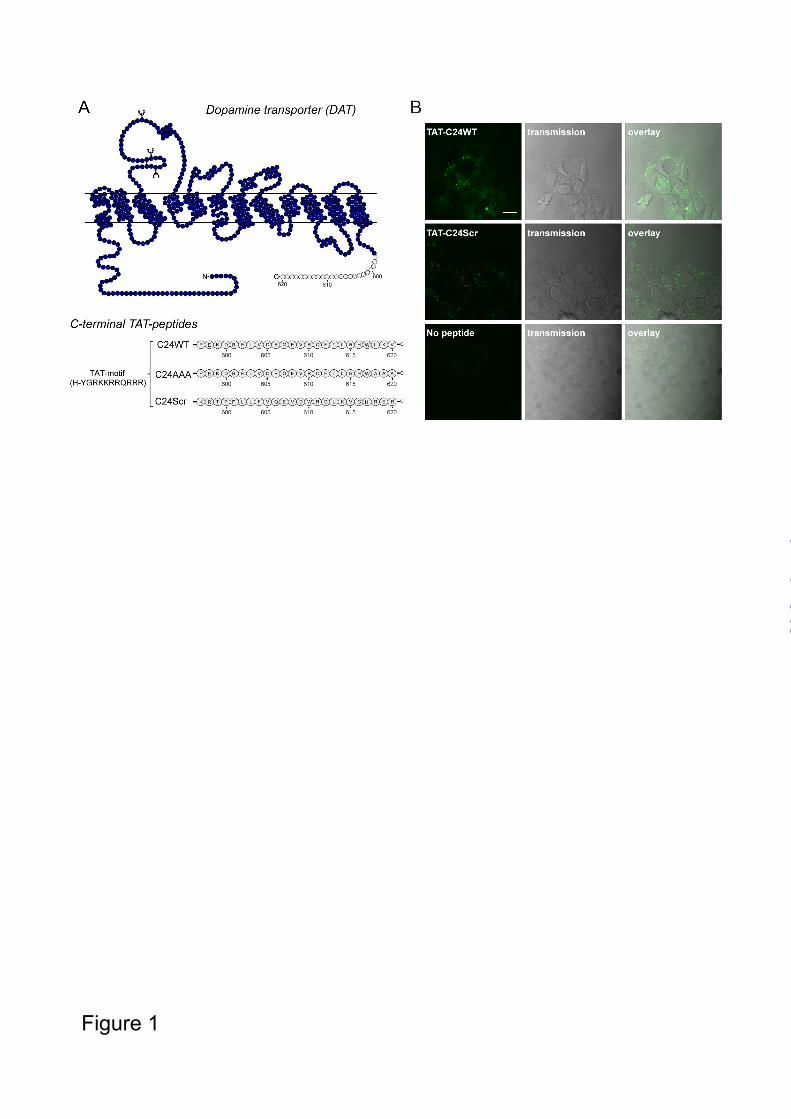
RESULTS Intracellular localization of cell-permeable DAT C-terminus peptides in HEK293 cells - We have previously shown evidence for a direct interaction between CaMKIIα and the distal 24 residues of the DAT C-terminus, and that this interaction might be critical for AMPH-induced DA efflux in cells (17). To further assess the significance of the DAT C-terminus for AMPH-induced DA efflux, we decided to investigate whether a C-terminal peptide corresponding to the CaMKIIα binding domain would be able to act in a dominant-negative fashion and blunt the response to AMPH by impairing CaMKIIα binding. The peptide was made membrane-permeable by fusing to the N-terminus of the cell-membrane transduction domain of the human immunodeficiency virus-type 1 (HIV-1) Tat protein (Tyr-Gly-Arg-Lys-Lys-Arg-Arg-Gln-Arg-Arg-Arg) (32,33) (Fig. 1A). Previous reports have successfully used TAT-conjugated dominant-negative peptides to efficiently perturb particular protein-protein interactions (32-36). In addition to a TAT-conjugated WT peptide (TAT-C24WT), we generated an equivalent scrambled peptide (TAT-C24Scr) (Fig. 1A). Moreover, we generated biotinylated variants to assess cell membrane permeability. Importantly, both peptides accumulated efficiently in HEK293 cells as determined by visualizing the peptides with fluorescently tagged streptavidin (fluorescein streptavidin) and confocal microscopy (Fig. 1B). TAT-C24WT peptide blocks CaMKIIα interaction with DAT - CaMKIIα associates with the DAT C-terminus as shown in pull-down assay using C-terminal glutathione-S-transferase fusion proteins containing the last 24 residues (C24) of DAT (17). The interaction has been further supported by co-
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

7
immunoprecipitation in both heterologous cells and in synaptosomal preparations (17,26). Here, we show that pCaMKIIα is observed in immunoprecipitates from T-Rex 293 CaMKIIα YFP-hDAT cells obtained with rat anti-DAT antibody, thus substantiating an association of phosphorylated CaMKIIα with the DAT C-terminus (Fig. 2). Pretreatment of the cells with TAT-C24WT strongly inhibited the amount of co-immunoprecipitated pCaMKIIα whereas TAT-C24Scr showed significantly less efficacy (* p<0.05, one-sample t-test, Fig. 2). These data support, in agreement with our prediction, that TAT-C24WT, but not the scrambled TAT-C24WT peptide, is capable of blocking the CaMKIIα/DAT interaction in living cells. AMPH-induced MPP+ efflux in heterologous cells co-expressing DAT and CaMKIIα is attenuated by TAT-peptides mimicking the DAT C - We then assessed the effect of the TAT-C24 peptides on AMPH-induced [3H]-MPP+ efflux in T-Rex 293 CaMKIIα FLAG-hDAT cells induced with tetracycline (Fig. 3A-B). We have previously shown by immunoblotting that CaMKIIα is robustly induced in these cells following tetracycline treatment without influencing DA uptake (17). Cells were preloaded with [³H]-MPP+, a hydrophilic DAT substrate resistant to enzymatic degradation and with minimal non-specific diffusion (27). Cells were then superfused and the experiment was initiated with collection of 2-min fractions, AMPH (3 µM) was added after 10 min. Cells were treated with TAT-C24WT (50 nM), TAT-C24Scr (50 nM) or vehicle during the preloading sequence. AMPH-evoked [³H]-MPP+
efflux was significantly attenuated by preincubation with the TAT-C24 WT peptide (* p<0.05, Fig. 3A). Importantly, TAT-C24Scr had no inhibitory effect on efflux, validating the efficacy of the WT peptide (Fig. 3A). These results were compared with experiments employing the CaMKII-inhibitor, KN-93 (15 µM), which markedly decreased AMPH-induced efflux (****p<0.0001, Fig. 3B). KN93 served as a control to show that AMPH-induced efflux of [³H]-MPP+ could be dramatically decreased as previously shown (17). Furthermore,
preincubation with both TAT-C24WT and KN-93 did not produce an additive inhibitory effect compared to KN-93 alone, indicating that their effects likely are exerted via the same pathway (Fig. 3B).
We also tested the effect of the peptides on [3H]-DA uptake in the T-Rex 293 CaMKIIα YFP-hDAT cells. These experiments showed that both DA uptake and the apparent affinity of DA for DAT are not affected by application of cell-permeable peptides mimicking the DAT C-terminus; TAT-C24WT and TAT-C24Scr peptides were applied 30 min prior DA uptake and there was no significant effect on uptake capacity (Vmax) and the Km value for DA uptake (Fig. 3C-D). This excludes acute effects of the peptides both on DAT catalytic activity and implies that TAT-C24WT selectively impaired AMPH-induced [3H]-MPP+ efflux in DAT expressing cells consistent with a dominant-negative effect on the DAT/CaMKIIα interaction. To exclude any effect of the peptides on AMPH interaction with the transporter we also performed competition [3H]-DA uptake experiments with AMPH. These experiments suggested unaltered AMPH affinity for DAT upon preincubation with TAT-peptides (Ki for AMPH after TAT-C24WT pretreatment, 1.8 µM; Ki for AMPH after TAT-C24Scr pretreatment, 0.8 µM; Ki control, 1.6 µM, n=2).
Finally, we wanted to exclude that TAT-C24WT or TAT-C24Scr directly affected trafficking and surface expression of the transporter. Cells were treated for 1h with TAT-C24WT, TAT-C24Scr or vehicle before assessing DAT surface expression using a surface biotinylation protocol. As shown in Fig. 3E, immunoblots for surface-expressed DAT and total DAT levels revealed no effect of the TAT-peptides. TAT-peptides mimicking the DAT C-terminus inhibit DA efflux in the striatum - We utilized high-speed chronoamperometric recordings in striatum to assess kinetics of DA release in vivo following administration of TAT-peptides mimicking DAT C-terminus. AMPH was pressure-ejected intrastriatally to evoke the release of DA. As soon as DA was cleared from extracellular fluid, typically 5-30 min after
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

8
AMPH-application, the peptide of interest or equivalent volume of aCSF (vehicle) was locally applied to striatum, and 45 min and 90 min later the same amount of AMPH was again pressure-ejected. Representative traces of DA efflux in mouse striatum upon AMPH stimulation are shown in Fig. 4A-C. Application of TAT-C24Scr did not influence DA release while TAT-C24WT inhibited release (Fig. 4A, B). In agreement with our previous findings (17), administration of the CaMKII inhibitor KN-93 reduced DA release (Fig. 4D). The inhibition was similar to that seen in response to TAT-C24WT and, importantly, combined administration of TAT-C24WT and KN-93 resulted in a similar attenuation of DA efflux compared to KN-93 alone suggesting no additive effect (Fig. 4C-D). Data are presented as fractional AMPH-induced DA release to illustrate a significant decrease in DA efflux upon WT peptide (TAT-C24WT) and KN-93 administration while vehicle (aCSF) and scrambled peptide (TAT-C24Scr) did not have any effect (Fig. 4D). This is consistent with data from the heterologous cells and shows that disrupting interactions with DAT C-terminus by cell-permeant peptides significantly decreases DA efflux in mouse striatum upon AMPH challenge. Because the DAT C-terminus contains a prototypic PDZ binding sequence known to interact with the PDZ-domain protein PICK1 (16,18), the TAT-C24WT peptide might also compete for binding of PDZ domain proteins to DAT in vivo. To address this issue, we generated an additional peptide, TAT-C24AAA, where the C-terminal PDZ-target sequence (-LLV) was substituted for three alanines (-AAA) resulting in abolished PDZ-mediated protein-protein interactions (18). The triple alanine substitution does not, however, affect CaMKIIα binding (17). The peptide should therefore only associate with CaMKIIα and not PICK1 since the PDZ-motif has been disrupted. First, we investigated the effect of the peptide in T-Rex 293 CaMKIIα FLAG-hDAT cells. The cells were preloaded with [³H]-MPP+ before superfusion and initiation of the experiment with collection of 2 min fractions. During the preloading sequence, cells were treated with either TAT-C24AAA (50 nM), TAT-C24Scr (50 nM) or vehicle and AMPH was added after 10 min.
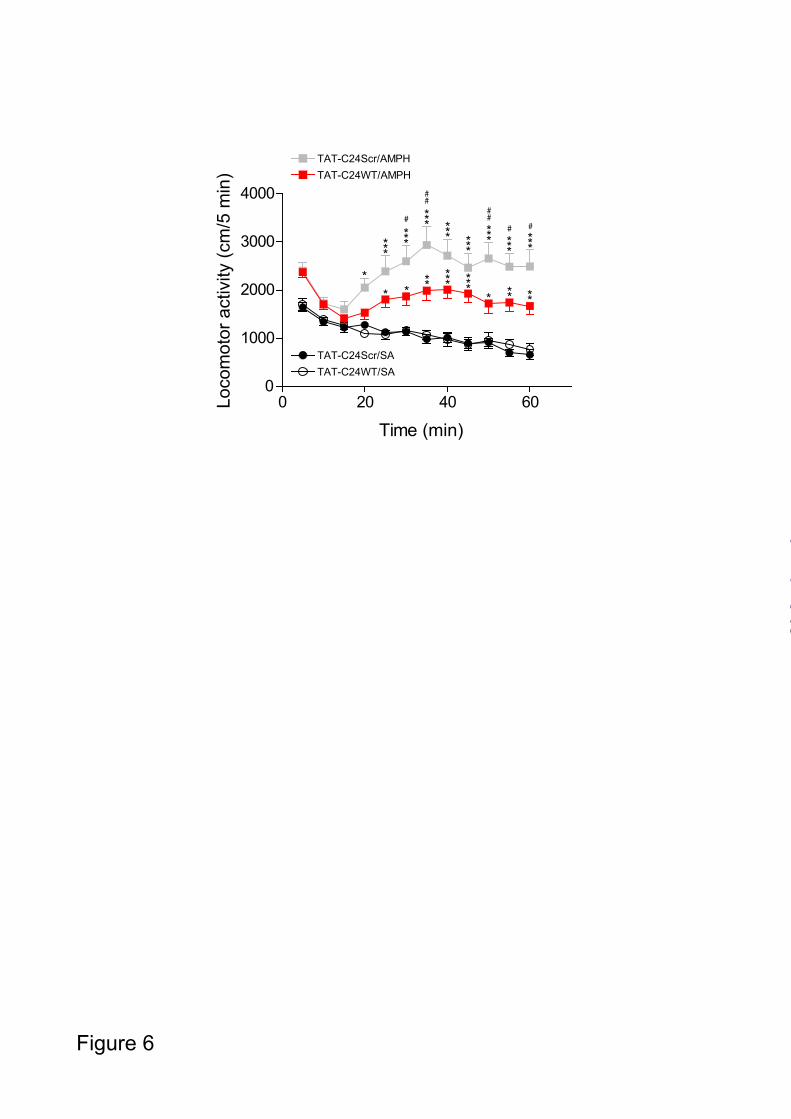
Intriguingly, AMPH-induced [³H]-MPP+ efflux was significantly reduced following preincubation with the TAT-C24AAA peptide compared to vehicle and TAT-C24Scr (control peptide) (*** p<0.001, Fig. 5A). Chronoamperometric recordings were then performed to assess the effect of TAT-C24AAA on DA release in response to AMPH-challenge. Representative traces for DA release in striatum before (baseline) and 90 min following TAT-C24AAA peptide administration demonstrate a decrease in DA efflux (Fig. 5B). Fractional AMPH-induced DA release showed a more pronounced attenuation 90 min following TAT-C24AAA administration compared to 45 min while no effect was seen for vehicle (* p<0.05, Fig. 5C). Hence, pre-administration of the TAT-C24AAA peptide markedly reduces AMPH-evoked DA release in striatum as determined by in vivo high-speed chrono-amperometry. TAT-C24WT peptide attenuates AMPH-induced hyperactivity - Several previous reports have shown efficient delivery of TAT-conjugated peptides across blood brain barrier and into the brain tissue upon systemic administration (32,34,35). We wished therefore to investigate whether in vivo administration of TAT-C24WT would be able to impair AMPH-induced locomotor hyperactivity in mice. To investigate effect of TAT-C24WT and TAT-C24Scr peptides on basal locomotion per se, mice were administered 100 µg of TAT-C24WT or TAT-C24Scr in the tail vein (intravenously). Saline (SA) solution or AMPH (2 mg/kg) was administered i.p. To allow sufficient time for peptide accumulation in brain tissue 45 min was permitted to elapse before beginning a 60 minute period of locomotor activity recordings. This time window has been shown to be sufficient to allow peptide accumulation in brain tissue (35). Our data showed that the membrane-permeable peptides did not affect locomotor activity per se (C24WT/SA, C24Scr/SA, Fig. 6). However, mice pretreated with TAT-C24Scr and subsequently challenged with AMPH showed a robust increase in locomotor activity that peaked 30 min after AMPH injection. Administration of TAT-C24WT peptide significantly attenuated the AMPH-
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

9
induced locomotor hyperactivity compared with TAT-C24Scr (# p<0.05, ## p<0.01 TAT-C24Scr/AMPH versus TAT-C24WT/AMPH, * p<0.05, ** p<0.01, *** p<0.001 versus own saline group, Bonferroni post-hoc test after repeated measures 2-way-ANOVA, Fig. 6). Behaviorally, psychostimulant-induced locomotor hyperactivity was attenuated in mice preinjected with TAT-C24WT peptide. This indicates that also in vivo disruption of the DAT/CaMKIIα interaction interferes with AMPH effects. DISCUSSION DAT-mediated reverse transport of DA is critical for the addictive and rewarding properties of AMPH. AMPH promotes DA efflux by inhibiting loading of cytoplasmic DA into vesicles causing an accumulation of cytosolic DA and subsequent reverse transport of DA via DAT into the synaptic cleft (9). We have previously shown evidence that CaMKIIα plays a role in AMPH-induced DA efflux (17). CaMKIIα was found to bind the DAT C-terminus, which in turn was suggested to facilitate phosphorylation of the DAT N-terminus and mediate AMPH-induced DA efflux (17). Recently, it was observed that AMPH-induced efflux is impaired in synaptosomes from mice with ablated CaMKIIα activity including a mouse model of the neurogenetic disease known as Angelman syndrome (26). In addition, studies in Drosophila have supported a role of DAT N-terminal phoshorylation in AMPH-induced DA efflux in vivo (37). To further characterize the role of the DAT C-terminus and CaMKIIα in AMPH-evoked DA efflux, we employ here dominant-negative peptides to disrupt DAT C-terminal protein-protein interactions. In agreement with our hypothesis, we find that the cell-permeant peptides mimicking the DAT C-terminus can reduce AMPH-evoked MPP+ and DA release both in vitro and in vivo. By use of co-immunoprecipitation experiments we were also able to validate that indeed the WT, but not a scrambled peptide, was able to block the association between CaMKIIα and DAT.
In heterologous cells, preloaded with [³H]-MPP+ and expressing both DAT and CaMKIIα, we found that the TAT-C24WT peptide, but not
the control peptide (TAT-C24Scr), efficiently attenuated AMPH-induced efflux. This is consistent with a dominant-negative action of the peptide preventing binding of CaMKIIα to the endogenous DAT C-terminus. The inability of TAT-C24WT to further inhibit [³H]-MPP+ efflux upon KN-93-mediated blockade of CaMKIIα activity further supports this conclusion, i.e. no additive effect was seen. The peptides had no effect on DA uptake and DAT surface levels, according to uptake and surface biotinylation experiments, respectively, consistent with no effect of the peptides on transporter turn-over rate and transporter trafficking. A recent study analyzed the effect of a corresponding DAT peptide and also this peptide showed no effect on DA uptake in the WT transporter (38).
To assess the effects of the peptides on DA release in vivo, we utilized high-speed chronoamperometry providing a high-time resolution on release kinetics compared to microdialysis (39). AMPH-evoked DA efflux in striatum was significantly reduced following application of TAT-C24WT while efflux was not altered upon administration of vehicle or TAT-C24Scr. Equivalently, AMPH-induced locomotor hyperactivity in mice was substantially lower upon in vivo administration of TAT-C24WT as compared to TAT-C24Scr. Altogether, the data are consistent with a critical role of C-terminal protein-protein interactions for AMPH-induced efflux and AMPH-induced locomotor hyperactivity.
Interestingly, the PDZ domain of PICK1 can bind both the DAT C-terminus and the C-terminus of PKCα (protein kinase Cα), and thus PICK1 could potentially act as a scaffold to bring PKCα in close proximity of DAT N-terminus. Notably, such a function of PICK1 has been proposed in relation to its binding to ASIC1/2 (Acid Sensing Ion Channels 1/2) and the GluR2 subunit of the AMPA-receptor (40,41). It has also previously been suggested that PKC isoforms, including PKCα and PKCβ, might play a role in AMPH-mediated DA efflux (17,23,42-44). Furthermore, the dominant-negative TAT-C24WT peptide would be predicted to abolish not only the association of CaMKIIα to endogenous DAT but also the association of the PDZ domain protein
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

10
PICK1 (17). It is therefore important that we assessed the effect of a TAT-peptide containing a disrupted PDZ-binding motif (TAT-C24AAA) and found that this attenuated [³H]-MPP+ release to a similar extent as TAT-C24WT. This finding in heterologous cells was further supported by chronoamperometric recordings in striatum preinjected with dominant-negative TAT-C24AAA peptide, i.e. the chrono-amperometric recordings showed markedly impaired DA release upon AMPH challenge. Although a role in efflux of interactions with PDZ domain proteins like PICK1 still cannot be excluded, the data do suggest that PDZ domain interactions are less important for DA efflux compared to the association of CaMKIIα to the DAT C-terminus.
Protein-protein interactions involving the DAT N-terminus have also been shown to play an important role in AMPH-evoked DA efflux. A recent study demonstrated that AMPH stimulates binding of syntaxin1A to the DAT N-terminus in a CaMKII-dependent manner in both heterologous cells and synaptosomal preparations. Inhibition of CaMKIIα activity using a membrane-permeable CaMKIIα inhibitor decreased the interaction between DAT and syntaxin1A. Importantly, similar effects were obtained by inhibiting CaMKIIα with KN-93 and it was hypothesized that AMPH-mediated activation of CaMKIIα increases DAT-syntaxin1A interactions resulting in reversed operation of DAT (45). This is in accordance with our data showing attenuated DA release upon TAT-peptides abolishing DAT C-terminus interactions, and it emphasizes the importance of CaMKIIα for AMPH-induced DA efflux. However, the mechanisms by which N-terminal phosphorylation and syntaxin1A binding mechanistically can alter the function of the transporter and cause reverse transport is still not understood. It was suggested that N-terminal phosphorylations shift the transporter from a reluctant to a willing state for AMPH-induced DA efflux (22). It has been speculated that this involves a conformational change in the transporter that might favour substrate efflux by increasing the intracellular affinity of DA and/or sodium for DAT (46). Alternatively, the phosphorylations may promote a channel-like mode of the transporter leading to rapid bursts of
DA efflux (47). The conformational flexibility of the N-terminus for AMPH-induced efflux might also be important; hence, tethering the N-terminus of SERT to the single transmembrane domain Tac (α-subunit of the interleukin-2 receptor) was found to strongly impair AMPH-induced efflux (48).
Intriguingly, the present data suggest that the effects of psychostimulants can be modulated by targeting DAT protein-protein interactions. Our data also corroborate the use of the TAT sequence as a simple way of solving the challenging problem of targeting peptides to the intracellular compartment. The TAT sequence is derived from the human immunodeficiency virus-type 1 (HIV-1) Tat protein and is capable of promoting intracellular accumulation of even larger proteins such as the β-galactosidase protein (32). There are now several examples substantiating the usefulness of the TAT sequence in relation to both studies of protein-protein interactions and in brain disease. Of specific interest for the present study, a TAT peptide containing an internal sequence from the C-terminus of the GluR2 subunit of the AMPA-type ionotropic glutamate receptor was shown to partially blunt behavioral sensitization to AMPH. The synthetic GluR2-derived peptide blocked regulated AMPA (α-amino-3-hydroxy-5-methyl-isoxazole-4-propionic acid) receptor endocytosis (35). Also, TAT fusion peptides with sequences from the C-terminus of the NMDA-type ionotropic glutamate receptor were shown to perturb NMDA (n-methyl-D-aspartate) receptor-PSD-95 interaction and thereby attenuate ischemic brain damage (33). Targeting the PDZ domain(s) of PSD-95 represents a pharmacological strategy for treatment of acute stroke (33,36).
Several DAT-interacting proteins have been identified implying that DAT is associated with a multi-protein network within the presynaptic terminal that control critical aspects of DAT function (4,14). A recent investigation showed that the Ras-like GTPase Rin interacts with the DAT and is required for PKC-mediated DAT internalization (49). The authors speculate that Rin may be involved in CaMKII activation, thereby representing a putative link between DAT, CaMKII and MAP kinase signalling. Altogether, it is becoming increasingly clear that DAT is subject to complex and delicate regulatory mechanisms
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

11
that govern trafficking and catalytic activity of the protein via complex protein-protein interactions and posttranslational modifications. Some interactions and pathways might even serve dual roles such as e.g. CaMKIIα and protein kinase C that both have been suggested to be involved in controlling trafficking and DA efflux (13,17,23,42-44). Further studies are required to dissect this complicated interplay. In this study, we have focused on one particular protein-protein
interaction and shown how its modulation is likely to change reverse transport and therefore might represent a strategy for therapeutic intervention in relation to abuse of AMPHs. It is interesting to consider how future detailed characterization of other DAT protein-protein interactions might identify new peptide targets and thereby new putative strategies for treating diseases characterized by dopaminergic dysfunction.
REFERENCES
1. Gainetdinov, R. R., and Caron, M. G. (2003) Monoamine transporters: from genes to behavior. Annu Rev Pharmacol Toxicol 43, 261-284
2. Torres, G. E., and Amara, S. G. (2007) Glutamate and monoamine transporters: new visions of form and function. Curr Opin Neurobiol 17, 304-312
3. Kristensen, A. S., Andersen, J., Jorgensen, T. N., Sorensen, L., Eriksen, J., Loland, C. J., Stromgaard, K., and Gether, U. (2011) SLC6 neurotransmitter transporters: structure, function, and regulation. Pharmacol Rev 63, 585-640
4. Sager, J. J., and Torres, G. E. (2011) Proteins interacting with monoamine transporters: current state and future challenges. Biochemistry 50, 7295-7310
5. Giros, B., Jaber, M., Jones, S. R., Wightman, R. M., and Caron, M. G. (1996) Hyperlocomotion and indifference to cocaine and amphetamine in mice lacking the dopamine transporter. Nature 379, 606-612
6. Beuming, T., Kniazeff, J., Bergmann, M. L., Shi, L., Gracia, L., Raniszewska, K., Newman, A. H., Javitch, J. A., Weinstein, H., Gether, U., and Loland, C. J. (2008) The binding sites for cocaine and dopamine in the dopamine transporter overlap. Nat Neurosci 11, 780-789
7. Jones, S. R., Gainetdinov, R. R., Jaber, M., Giros, B., Wightman, R. M., and Caron, M. G. (1998) Profound neuronal plasticity in response to inactivation of the dopamine transporter. Proc Natl Acad Sci U S A 95, 4029-4034
8. Sitte, H. H., Huck, S., Reither, H., Boehm, S., Singer, E. A., and Pifl, C. (1998) Carrier-mediated release, transport rates, and charge transfer induced by amphetamine, tyramine, and dopamine in mammalian cells transfected with the human dopamine transporter. J Neurochem 71, 1289-1297
9. Sulzer, D., Sonders, M. S., Poulsen, N. W., and Galli, A. (2005) Mechanisms of neurotransmitter release by amphetamines: a review. Prog Neurobiol 75, 406-433
10. Daniels, G. M., and Amara, S. G. (1999) Regulated trafficking of the human dopamine transporter. Clathrin-mediated internalization and lysosomal degradation in response to phorbol esters. J Biol Chem 274, 35794-35801
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

12
11. Kahlig, K. M., Javitch, J. A., and Galli, A. (2004) Amphetamine regulation of dopamine transport. Combined measurements of transporter currents and transporter imaging support the endocytosis of an active carrier. J Biol Chem 279, 8966-8975
12. Saunders, C., Ferrer, J. V., Shi, L., Chen, J., Merrill, G., Lamb, M. E., Leeb-Lundberg, L. M., Carvelli, L., Javitch, J. A., and Galli, A. (2000) Amphetamine-induced loss of human dopamine transporter activity: an internalization-dependent and cocaine-sensitive mechanism. Proc Natl Acad Sci U S A 97, 6850-6855
13. Wei, Y., Williams, J. M., Dipace, C., Sung, U., Javitch, J. A., Galli, A., and Saunders, C. (2007) Dopamine transporter activity mediates amphetamine-induced inhibition of Akt through a Ca2+/calmodulin-dependent kinase II-dependent mechanism. Mol Pharmacol 71, 835-842
14. Eriksen, J., Jorgensen, T. N., and Gether, U. (2010) Regulation of dopamine transporter function by protein-protein interactions: new discoveries and methodological challenges. J Neurochem 113, 27-41
15. Torres, G. E. (2006) The dopamine transporter proteome. J Neurochem 97 Suppl 1, 3-10 16. Torres, G. E., Yao, W. D., Mohn, A. R., Quan, H., Kim, K. M., Levey, A. I., Staudinger,
J., and Caron, M. G. (2001) Functional interaction between monoamine plasma membrane transporters and the synaptic PDZ domain-containing protein PICK1. Neuron 30, 121-134
17. Fog, J. U., Khoshbouei, H., Holy, M., Owens, W. A., Vaegter, C. B., Sen, N., Nikandrova, Y., Bowton, E., McMahon, D. G., Colbran, R. J., Daws, L. C., Sitte, H. H., Javitch, J. A., Galli, A., and Gether, U. (2006) Calmodulin kinase II interacts with the dopamine transporter C terminus to regulate amphetamine-induced reverse transport. Neuron 51, 417-429
18. Bjerggaard, C., Fog, J. U., Hastrup, H., Madsen, K., Loland, C. J., Javitch, J. A., and Gether, U. (2004) Surface targeting of the dopamine transporter involves discrete epitopes in the distal C terminus but does not require canonical PDZ domain interactions. J Neurosci 24, 7024-7036
19. Rickhag, M., Hansen, F. H., Sorensen, G., Strandfelt, K. N., Andresen, B., Gotfryd, K., Madsen, K. L., Vestergaard-Klewe, I., Ammendrup-Johnsen, I., Eriksen, J., Newman, A. H., Fuchtbauer, E. M., Gomeza, J., Woldbye, D. P., Wortwein, G., and Gether, U. (2013) A C-terminal PDZ domain-binding sequence is required for striatal distribution of the dopamine transporter. Nature communications 4, 1580
20. Foster, J. D., Pananusorn, B., and Vaughan, R. A. (2002) Dopamine transporters are phosphorylated on N-terminal serines in rat striatum. J Biol Chem 277, 25178-25186
21. Vaughan, R. A., Huff, R. A., Uhl, G. R., and Kuhar, M. J. (1997) Protein kinase C-mediated phosphorylation and functional regulation of dopamine transporters in striatal synaptosomes. J Biol Chem 272, 15541-15546
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

13
22. Khoshbouei, H., Sen, N., Guptaroy, B., Johnson, L., Lund, D., Gnegy, M. E., Galli, A., and Javitch, J. A. (2004) N-terminal phosphorylation of the dopamine transporter is required for amphetamine-induced efflux. PLoS Biol 2, E78
23. Kantor, L., Hewlett, G. H., and Gnegy, M. E. (1999) Enhanced amphetamine- and K+-mediated dopamine release in rat striatum after repeated amphetamine: differential requirements for Ca2+- and calmodulin-dependent phosphorylation and synaptic vesicles. J Neurosci 19, 3801-3808
24. Pierce, R. C., and Kalivas, P. W. (1997) Repeated cocaine modifies the mechanism by which amphetamine releases dopamine. J Neurosci 17, 3254-3261
25. Gnegy, M. E., Khoshbouei, H., Berg, K. A., Javitch, J. A., Clarke, W. P., Zhang, M., and Galli, A. (2004) Intracellular Ca2+ regulates amphetamine-induced dopamine efflux and currents mediated by the human dopamine transporter. Mol Pharmacol 66, 137-143
26. Steinkellner, T., Yang, J. W., Montgomery, T. R., Chen, W. Q., Winkler, M. T., Sucic, S., Lubec, G., Freissmuth, M., Elgersma, Y., Sitte, H. H., and Kudlacek, O. (2012) Ca(2+)/calmodulin-dependent protein kinase IIalpha (alphaCaMKII) controls the activity of the dopamine transporter: implications for Angelman syndrome. J Biol Chem 287, 29627-29635
27. Scholze, P., Norregaard, L., Singer, E. A., Freissmuth, M., Gether, U., and Sitte, H. H. (2002) The role of zinc ions in reverse transport mediated by monoamine transporters. J Biol Chem 277, 21505-21513
28. Owens, W. A., Sevak, R. J., Galici, R., Chang, X., Javors, M. A., Galli, A., France, C. P., and Daws, L. C. (2005) Deficits in dopamine clearance and locomotion in hypoinsulinemic rats unmask novel modulation of dopamine transporters by amphetamine. J Neurochem 94, 1402-1410
29. Williams, J. M., Owens, W. A., Turner, G. H., Saunders, C., Dipace, C., Blakely, R. D., France, C. P., Gore, J. C., Daws, L. C., Avison, M. J., and Galli, A. (2007) Hypoinsulinemia regulates amphetamine-induced reverse transport of dopamine. PLoS Biol 5, e274
30. Callaghan, P. D., Irvine, R. J., and Daws, L. C. (2005) Differences in the in vivo dynamics of neurotransmitter release and serotonin uptake after acute para-methoxyamphetamine and 3,4-methylenedioxymethamphetamine revealed by chronoamperometry. Neurochem Int 47, 350-361
31. Clausing, P., Gough, B., Holson, R. R., Slikker, W., Jr., and Bowyer, J. F. (1995) Amphetamine levels in brain microdialysate, caudate/putamen, substantia nigra and plasma after dosage that produces either behavioral or neurotoxic effects. J Pharmacol Exp Ther 274, 614-621
32. Schwarze, S. R., Ho, A., Vocero-Akbani, A., and Dowdy, S. F. (1999) In vivo protein transduction: delivery of a biologically active protein into the mouse. Science 285, 1569-1572
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

14
33. Aarts, M., Liu, Y., Liu, L., Besshoh, S., Arundine, M., Gurd, J. W., Wang, Y. T., Salter, M. W., and Tymianski, M. (2002) Treatment of ischemic brain damage by perturbing NMDA receptor- PSD-95 protein interactions. Science 298, 846-850
34. Bertaso, F., Zhang, C., Scheschonka, A., de Bock, F., Fontanaud, P., Marin, P., Huganir, R. L., Betz, H., Bockaert, J., Fagni, L., and Lerner-Natoli, M. (2008) PICK1 uncoupling from mGluR7a causes absence-like seizures. Nat Neurosci 11, 940-948
35. Brebner, K., Wong, T. P., Liu, L., Liu, Y., Campsall, P., Gray, S., Phelps, L., Phillips, A. G., and Wang, Y. T. (2005) Nucleus accumbens long-term depression and the expression of behavioral sensitization. Science 310, 1340-1343
36. Bach, A., Clausen, B. H., Moller, M., Vestergaard, B., Chi, C. N., Round, A., Sorensen, P. L., Nissen, K. B., Kastrup, J. S., Gajhede, M., Jemth, P., Kristensen, A. S., Lundstrom, P., Lambertsen, K. L., and Stromgaard, K. (2012) A high-affinity, dimeric inhibitor of PSD-95 bivalently interacts with PDZ1-2 and protects against ischemic brain damage. Proc Natl Acad Sci U S A 109, 3317-3322
37. Pizzo, A. B., Karam, C. S., Zhang, Y., Yano, H., Freyberg, R. J., Karam, D. S., Freyberg, Z., Yamamoto, A., McCabe, B. D., and Javitch, J. A. (2012) The membrane raft protein Flotillin-1 is essential in dopamine neurons for amphetamine-induced behavior in Drosophila. Mol Psychiatry
38. Sakrikar, D., Mazei-Robison, M. S., Mergy, M. A., Richtand, N. W., Han, Q., Hamilton, P. J., Bowton, E., Galli, A., Veenstra-Vanderweele, J., Gill, M., and Blakely, R. D. (2012) Attention deficit/hyperactivity disorder-derived coding variation in the dopamine transporter disrupts microdomain targeting and trafficking regulation. J Neurosci 32, 5385-5397
39. Daws, L. C., Montanez, S., Owens, W. A., Gould, G. G., Frazer, A., Toney, G. M., and Gerhardt, G. A. (2005) Transport mechanisms governing serotonin clearance in vivo revealed by high-speed chronoamperometry. J Neurosci Methods 143, 49-62
40. Hruska-Hageman, A. M., Wemmie, J. A., Price, M. P., and Welsh, M. J. (2002) Interaction of the synaptic protein PICK1 (protein interacting with C kinase 1) with the non-voltage gated sodium channels BNC1 (brain Na+ channel 1) and ASIC (acid-sensing ion channel). Biochem J 361, 443-450
41. Xia, J., Chung, H. J., Wihler, C., Huganir, R. L., and Linden, D. J. (2000) Cerebellar long-term depression requires PKC-regulated interactions between GluR2/3 and PDZ domain-containing proteins. Neuron 28, 499-510
42. Cowell, R. M., Kantor, L., Hewlett, G. H., Frey, K. A., and Gnegy, M. E. (2000) Dopamine transporter antagonists block phorbol ester-induced dopamine release and dopamine transporter phosphorylation in striatal synaptosomes. Eur J Pharmacol 389, 59-65
43. Johnson, L. A., Guptaroy, B., Lund, D., Shamban, S., and Gnegy, M. E. (2005) Regulation of amphetamine-stimulated dopamine efflux by protein kinase C beta. J Biol Chem 280, 10914-10919
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

15
44. Cervinski, M. A., Foster, J. D., and Vaughan, R. A. (2005) Psychoactive substrates stimulate dopamine transporter phosphorylation and down-regulation by cocaine-sensitive and protein kinase C-dependent mechanisms. J Biol Chem 280, 40442-40449
45. Binda, F., Dipace, C., Bowton, E., Robertson, S. D., Lute, B. J., Fog, J. U., Zhang, M., Sen, N., Colbran, R. J., Gnegy, M. E., Gether, U., Javitch, J. A., Erreger, K., and Galli, A. (2008) Syntaxin 1A interaction with the dopamine transporter promotes amphetamine-induced dopamine efflux. Mol Pharmacol 74, 1101-1108
46. Bowton, E., Saunders, C., Erreger, K., Sakrikar, D., Matthies, H. J., Sen, N., Jessen, T., Colbran, R. J., Caron, M. G., Javitch, J. A., Blakely, R. D., and Galli, A. (2010) Dysregulation of dopamine transporters via dopamine D2 autoreceptors triggers anomalous dopamine efflux associated with attention-deficit hyperactivity disorder. J Neurosci 30, 6048-6057
47. Kahlig, K. M., Binda, F., Khoshbouei, H., Blakely, R. D., McMahon, D. G., Javitch, J. A., and Galli, A. (2005) Amphetamine induces dopamine efflux through a dopamine transporter channel. Proc Natl Acad Sci U S A 102, 3495-3500
48. Sucic, S., Dallinger, S., Zdrazil, B., Weissensteiner, R., Jorgensen, T. N., Holy, M., Kudlacek, O., Seidel, S., Cha, J. H., Gether, U., Newman, A. H., Ecker, G. F., Freissmuth, M., and Sitte, H. H. (2010) The N terminus of monoamine transporters is a lever required for the action of amphetamines. J Biol Chem 285, 10924-10938
49. Navaroli, D. M., Stevens, Z. H., Uzelac, Z., Gabriel, L., King, M. J., Lifshitz, L. M., Sitte, H. H., and Melikian, H. E. (2011) The plasma membrane-associated GTPase Rin interacts with the dopamine transporter and is required for protein kinase C-regulated dopamine transporter trafficking. J Neurosci 31, 13758-13770
FOOTNOTES The work was supported by the National Institute of Health Grants P01 DA 12408 (UG), the Danish Health Medical Research Council (MR, UG), The Novo Nordisk Foundation (‘Fabrikant Vilhelm Pedersen og Hustrus Mindelegat’) (UG), The Lundbeck foundation (MR, UG) and University of Copenhagen BioScaRT Program of Excellence (UG). We thank Pia Elsman for excellent technical assistance.
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

16
FIGURE LEGENDS FIGURE 1. Intracellular localization of TAT-peptides mimicking DAT C-terminus in HEK293 cells. A. Schematic illustration of DAT topology with the TAT-peptide targeting sequence in grey (24 C-terminal residues). The sequence of the synthesized TAT-pepides are indicated (TAT-C24WT, TAT-C24Scr and TAT-C24AAA). B. Biotin-conjugated TAT-C24 peptides (WT and Scr) were visualized with streptavidin fluorescein in untransfected HEK293 cells and show specific localization to intracellular compartments. Control experiments included application of physiological saline solution (which was used for dissolving TAT-C24WT and TAT-C24Scr biotin-conjugated peptides) to the cells and showed absence of labelled peptides. Scale bar= 10 µm FIGURE 2. TAT-C24WT peptide blocks CaMKIIα /DAT interaction in T-Rex 293 CaMKIIα YFP-hDAT cells. DAT was immunoprecipitated (IP) with rat anti-DAT antibody in cell lysates from T-Rex 293 cells co-expressing DAT and CaMKIIα. Cells were pretreated with either 1 µM TAT-C24WT or TAT-C24Scr peptide. As controls, cell lysates without peptide and pull-down without rat anti-DAT antibody were performed in parallel. Left panel, A representative immunoblot with rabbit phosphorylated anti-pCaMKIIα antibody (IB: pCaMKIIα) shows that TAT-C24WT peptide (WT) disrupts CaMKIIα-DAT interaction while TAT-C24Scr (Scr) shows less efficacy. Cell lysates without peptide show robust co-immunoprecipitation (co-IP) of pCaMKIIα. A representative immunoblot for DAT immunoprecipitation demonstrates comparable DAT pull-down for all conditions (IB: DAT). A representative immunoblot for total cell lysates shows robust induction of pCaMKIIα (Ctl=control, WT=TAT-C24WT, Scr=TAT-C24Scr, no ab= no antibody, left panel). Right panel, Normalized pCaMKIIα co-IP with pretreatment of either TAT-C24WT or TAT-C24Scr shows significant reduced co-IP with TAT-C24WT compared to TAT-C24Scr, mean ± S.E., n=4, * P <0.05, one-sample t test. FIGURE 3. Effect of TAT-C24WT and TAT-C24Scr peptide on AMPH-induced MPP+ efflux in T-Rex 293 CaMKIIα FLAG hDAT cells. A. Cells were preloaded with [³H]-MPP+, superfused and the experiment was initiated with collection of 2 min fractions; during the preloading sequence, cells were treated with TAT-C24WT (50 nM), TAT-C24Scr (50 nM) or vehicle. AMPH (3 µM) was added after 10 min. Data are fractional release per 2 min in % (means ± S.E. of 12 observations from 3 experimental days), **** P<0.0001, two-way ANOVA with Bonferroni’s multiple comparisons test. B. AMPH-induced [³H]-MPP efflux upon co-application of TAT-C24WT/Scr and the CaMKII-inhibitor KN-93. Data are fractional release per 2 min in % (means ± S.E. of 12 observations from 3 experimental days), **** p<0.0001, two-way ANOVA with Bonferroni’s multiple comparisons test. C. Preincubation with TAT-peptides (TAT-C24WT and TAT-C24Scr; 1 µM) 30 min prior to measurement of [3H]-DA uptake in T-Rex 293 CaMKIIα FLAG hDAT cells showed no effect on DA transport (Vmax). D. The Km value for DA was also unchanged in cells pretreated with TAT-peptides (Km for DA after TAT-C24WT treatment, 3.27 ± 1.77 µM; Km for DA after TAT-C24Scr treatment, 2.10 ± 0.55 µM; Km for DA, control, 1.50 ± 0.19 µM. Data are means ± SE of experiments (n=4-5) performed in triplicate. E. Pretreatment with TAT-C24WT or TAT-C24Scr for 1h does not affect DAT surface expression according to a surface biotinylation assay. A representative immunoblot for surface-expressed DAT and total DAT levels in T-Rex 293 CaMKIIα YFP-hDAT cells show unaltered DAT levels after preincubation with TAT-C24WT (WT), TAT-C24Scr (Scr) or vehicle (Ctl). DAT surface levels were normalized to total DAT extracts and then expressed relative to control group without peptide treatment. Normalized data are expressed as means ± S.E., n=3 (WT and Ctl) or n=2 (Scr) and compared with respective controls by 1-way ANOVA, p>0.05.
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

17
FIGURE 4. AMPH-evoked DA efflux in striatum following administration of TAT-C24 DAT peptides mimicking DAT C-terminus. Representative oxidation currents (converted to micromolar values) produced by pressure ejection of AMPH into the striatum. A. Oxidation currents before and 90 min following application of TAT-C24Scr show no effect on DA release. B. Oxidation currents before and 90 min following application of TAT-C24WT demonstrate substantial decrease in DA efflux. C. Oxidation currents before and 90 min following co-application of TAT-C24WT and KN-93. D. Data are presented as fractional AMPH-induced DA efflux after local application of vehicle (aCSF) (n=10), TAT-C24Scr (n=11), TAT-C24WT (n=14), KN-93 (n=9) and C24WT+KN-93 (n=3), * p<0.05, 1-way ANOVA with Dunnett’s multiple comparison test. FIGURE 5. The role of DAT PDZ-domain interactions for AMPH-evoked DA efflux. A. T-Rex 293/ CaMKIIα FLAG hDAT cells were preloaded with [³H]-MPP+, superfused and the experiment was initiated with collection of 2 min fractions; during the preloading sequence, cells were treated with either TAT-C24Scr (50 nM), TAT-C24AAA (50 nM) or vehicle. AMPH (3 µM) was added after 10 min. Data are presented as area-under-curve (AUC, arbitrary units) and show significant reduction in efflux upon TAT-C24AAA treatment (means ± S.E. of 12 observations from 3 experimental days), *** p<0.0001 1-way ANOVA with Bonferroni’s multiple comparison test. B. Representative oxidation currents (converted to micromolar values) produced by intrastriatal pressure ejection of AMPH before and 90 min following application of TAT-C24AAA C. Data are presented as fractional AMPH-induced DA efflux after local application of vehicle (aCSF) (n=10), and TAT-C24AAA (n=8), * p<0.05, 1-way ANOVA with Dunnett’s multiple comparison test. FIGURE 6. Effect of TAT-C24 DAT peptides mimicking DAT C-terminus on AMPH-induced locomotor hyperactivity. Intravenous administration of TAT-C24WT peptide prior to AMPH administration significantly attenuates the AMPH-induced hyperactivity compared to treatment with TAT-C24Scr. Data are means ± S.E, # p<0.05, ## p<0.01 TAT-C24Scr/AMPH versus TAT-C24WT/AMPH; * p<0.05, ** p<0.01, *** p<0.001 versus own saline group, Bonferroni post-hoc tests after repeated measures 2-way ANOVA (nC24WT/AMPH=16, nC24Scr/AMPH=10). Note that administration of TAT-peptides prior to saline administration did not affect locomotor activity per se (nC24WT/SA=11, nC24Scr/SA=11).
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from
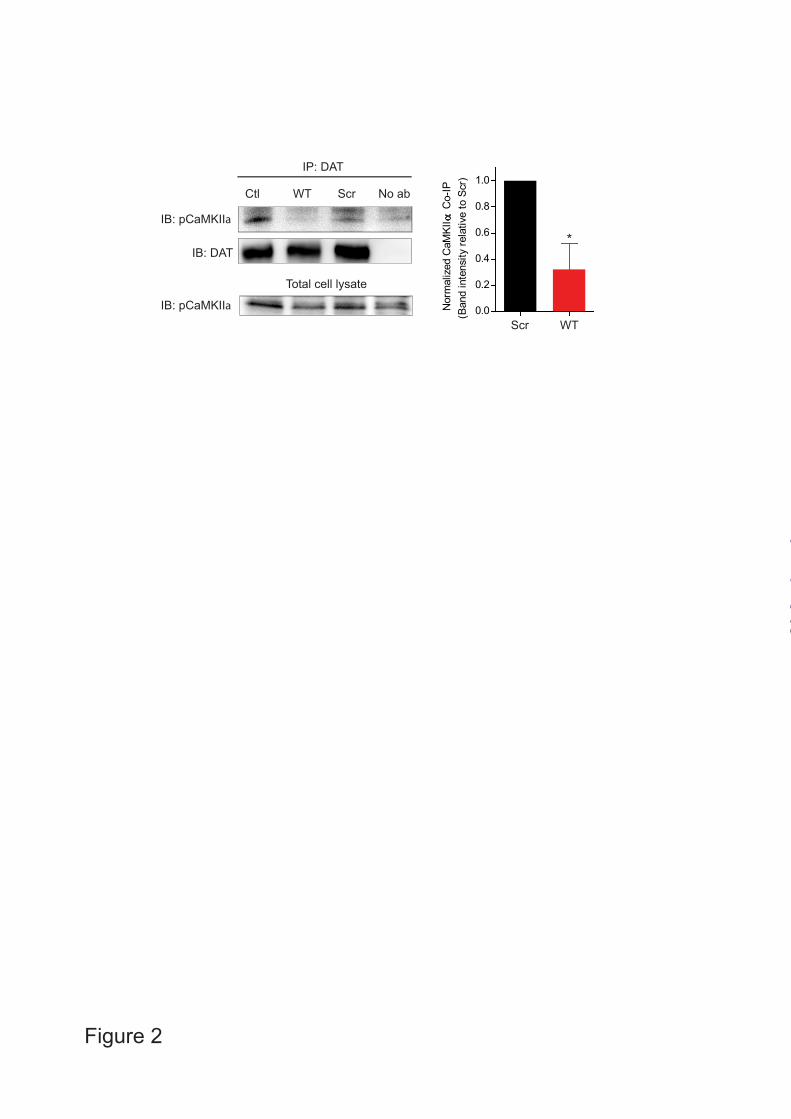
Figure 2
IB: pCaMKIIa
IB: DAT
Ctl WT Scr No ab
IP: DAT
IB: pCaMKIIa
Total cell lysate
0.0
0.2
0.4
0.6
0.8
1.0
Nor
mal
ized
CaM
KII α
Co-
IP(B
and
inte
nsity
rela
tive
to S
cr)
*
WTScr
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from
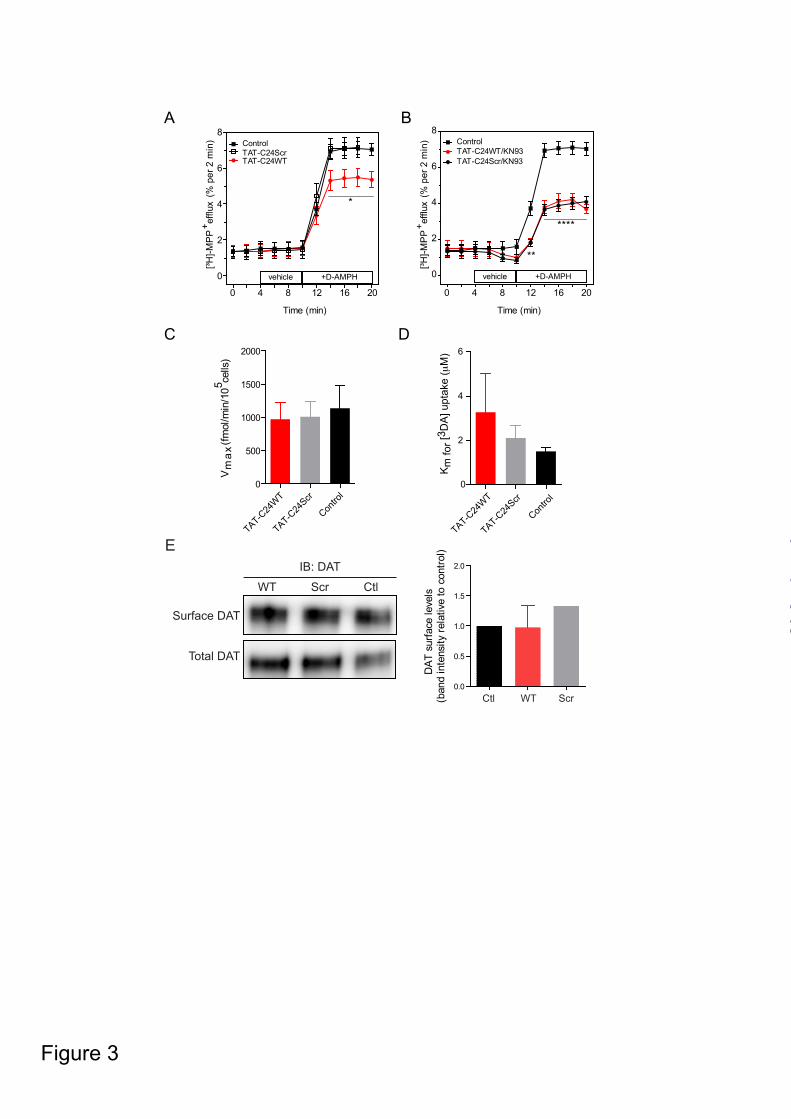
Figure 3
A B
Time (min)
[³H]-M
PP
+ effl
ux(%
per
2 m
in)
0 4 8 12 16 20
0
2
4
6
8
TAT-C24ScrTAT-C24WT
Control
+D-AMPHvehicle
Time (min)
[³H]-M
PP
+ effl
ux
(% p
er 2
min
)
+D-AMPHvehicle
0 4 8 12 16 20
0
2
4
6
8
TAT-C24Scr/KN93
ControlTAT-C24WT/KN93
*
**
****
C
Vm
ax(fm
ol/m
in/1
05 cells
)
TAT-C24
WT
TAT-C24
Scr
Contro
l0
500
1000
1500
2000D
TAT-C24
WT
TAT-C24
Scr
Contro
l0
2
4
6
Km
for [
3 DA
] upt
ake
(μM
)
E
Surface DAT
Total DAT
WT Scr Ctl
IB: DAT
0.0
0.5
1.0
1.5
2.0
DAT
sur
face
leve
ls(b
and
inte
nsity
rela
tive
to c
ontro
l)
Ctl WT Scr
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from
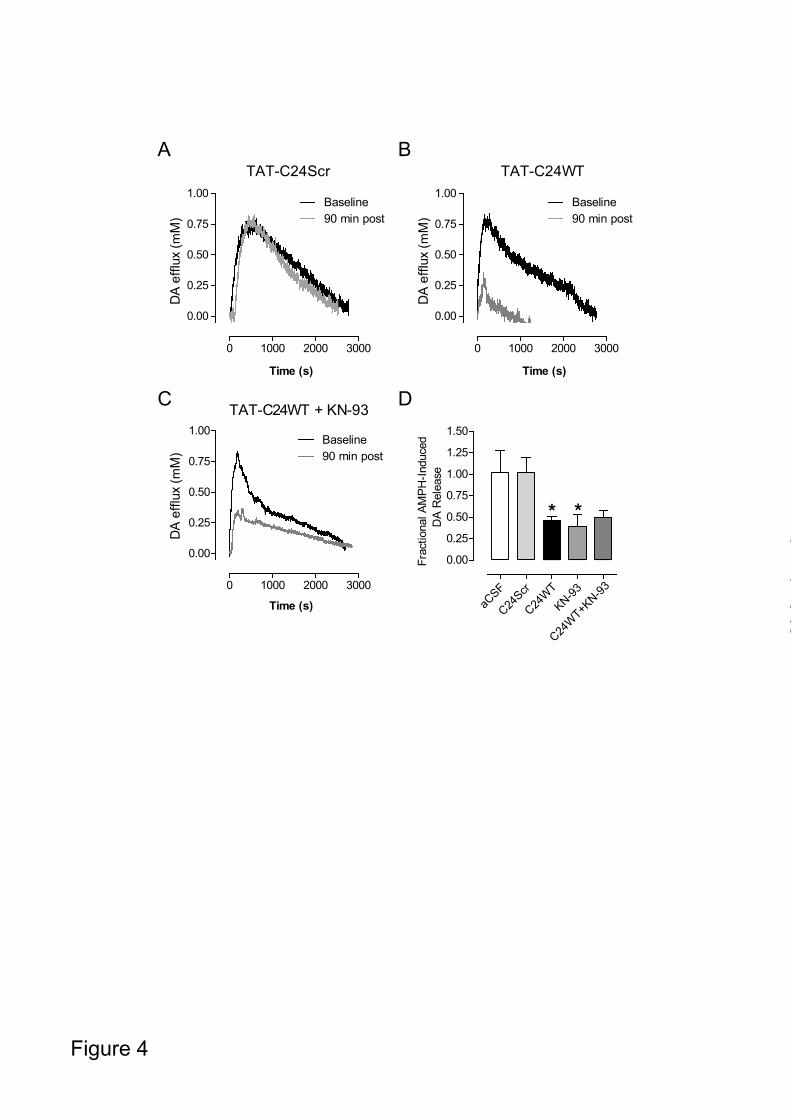
Figure 4
TAT-C24Scr
Time (s)
DA
efflu
x (m
M)
DA
efflu
x (m
M)
Time (s)
TAT-C24WT
DA
efflu
x (m
M)
A B
C D
0 1000 2000 3000
0.00
0.25
0.50
0.75
1.00
Time (s)
0 1000 2000 3000
0.00
0.25
0.50
0.75
1.00Baseline90 min post
TAT-C24WT + KN-93
0.00
0.25
0.50
0.75
1.00Baseline90 min post
aCSF
C24Scr
C24W
TKN-93
C24W
T+KN-93
0.00
0.25
0.50
0.75
1.00
1.25
1.50Fr
actio
nal A
MPH
-Indu
ced
DA
Rel
ease
Baseline90 min post
* *
0 1000 2000 3000
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from
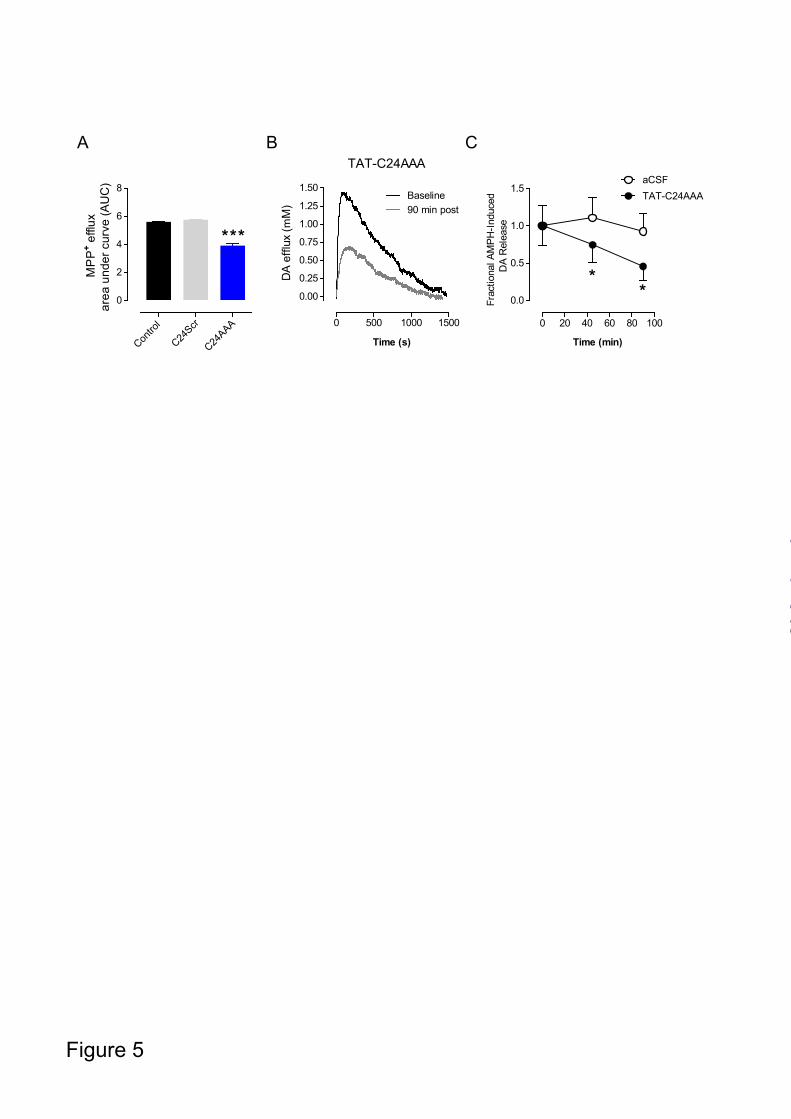
Figure 5
ATAT-C24AAA
DA
efflu
x (m
M)
B C
0 500 1000 1500
0.00
0.25
0.50
0.75
1.00
1.25
1.50Baseline90 min post
Time (s)
0 20 40 60 80 100
0.0
0.5
1.0
1.5aCSFTAT-C24AAA
Time (min)
Frac
tiona
l AM
PH-In
duce
dD
A R
elea
se
Contro
l
C24Scr
C24AAA
0
2
4
6
8
MP
P+ e
fflux
area
und
er c
urve
(AU
C)
**
** *
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from
0 20 40 600
1000
2000
3000
4000
Time (min)
Loco
mot
or a
ctiv
ity (c
m/5
min
)
Figure 6
TAT-C24Scr/SATAT-C24WT/SA
TAT-C24Scr/AMPHTAT-C24WT/AMPH
** * *
***
******
*** *** ****** ***
***
***** ** **
###
##
# #
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from

GetherJørgensen, Gitta Wörthwein, David P. D. Woldbye, Harald Sitte, Lynette C. Daws and Ulrik
Mette Rathje, Gunnar Sørensen, Bjørn Andresen, Kenneth L. Madsen, Trine Nygaard Mattias Rickhag, William A. Owens, Marie-Therese Winkler, Kristine Nørgaard Strandfelt,
amphetamine-evoked dopamine releaseMembrane permeable C-terminal dopamine transporter peptides attenuate
published online July 24, 2013J. Biol. Chem.
10.1074/jbc.M112.441295Access the most updated version of this article at doi:
Alerts:
When a correction for this article is posted•
When this article is cited•
to choose from all of JBC's e-mail alertsClick here
by guest on March 21, 2020
http://ww
w.jbc.org/
Dow
nloaded from