Embed Size (px)
Citation preview

Melanocyte-Associated T Cell Epitopes Can Function asAutoantigens for Transfer of Alopecia Areata to Human ScalpExplants on Prkdcscid Mice
Amos Gilhar, Marina Landau,* Bedia Assy, Raya Shalaginov, Sima Sera®movich, and Richard S. Kalish²Skin Research Laboratories, Flieman Medical Center and Rappaport Faculty of Medicine, Technion Institute of Technology, Haifa, Israel;
*Department of Dermatology, Elias Sourasky Medical Center, Tel Aviv, Israel; ²Department of Dermatology, State University of New York at Stony
Brook, U.S.A.
Alopecia areata is a tissue restricted autoimmunecondition affecting the hair follicle, resulting in hairloss. The goal of this study was to test the hypothesisthat the autoantigen of alopecia areata is melanocyteassociated. Potential autoantigens were tested in thehuman scalp explant/Prkdscid CB-17 mouse transfersystem. Scalp T cells from lesional (bald) alopeciaareata scalp were cultured with antigen-presentingcells, and antigen, along with interleukin-2. TheT cells were then injected into autologous lesionalscalp grafts on SCID mice, and hair regrowth wasmeasured. Hair follicle homogenate was used as anautoantigen control. T cells cultured with melanomahomogenate induced statistically signi®cant reduc-tion in hair growth (p < 0.01 by ANOVA). HLA-A2-
restricted melanocyte peptide epitopes were thentested with lesional scalp T cells from HLA-A2-posi-tive alopecia areata patients. Melanocyte-peptide-activated T cells signi®cantly reduced the number ofhairs regrowing in two experiments with six patients(p < 0.001 by ANOVA). Injected scalp grafts showedhistologic and immunochemical changes of alopeciaareata. The most consistent peptide autoantigenswere the Gp100-derived G9-209 and G9-280peptides, as well as MART-1 (27±35). Melanocytepeptide epitopes can function as autoantigens foralopecia areata. Multiple peptides were recognized,suggesting epitope spreading. Key words: autoimmu-nity/human/peptides/skin. J Invest Dermatol 117:1357±1362, 2001
Alopecia areata is a tissue restricted autoimmunecondition directed at the hair follicle, resulting in hairloss. Patients frequently suffer severe psychiatricconsequences (Colon et al, 1991; Koo et al, 1994).This is especially true of girls and young women who
become bald. The incidence of alopecia areata in the U.S.A.(Minnesota) is 20.2 per 100,000 person-years, with a lifetime risk ofapproximately 1.7% (Safavi et al, 1995). There is no signi®cantgender difference. The disease is often chronic with a remitting,relapsing course. Although it responds to immunosuppression,generalized immunosuppression has signi®cant morbidity andtreatment is frequently frustrating and not successful. Newtreatment options are essential. With evidence that alopecia areatais a T-lymphocyte-mediated autoimmune condition, it has becomea model system for the study of pathogenesis and treatment ofT-cell-mediated autoimmunity, and as such is a model for a hostof additional T-cell-mediated autoimmune conditions.
Hair loss in alopecia areata is associated with a perifollicularlymphocytic in®ltrate made up primarily of CD4+ cells, with aCD8+ intrafollicular in®ltrate (Todes-Taylor et al, 1984), alongwith expression of both HLA-DR and intercellular adhesionmolecule 1 (ICAM-1) on the follicular epithelium (Messenger andBleehen, 1985; Khoury et al, 1988). It is presumed that the
follicular expression of DR and ICAM-1 is induced by interferon-g(INF-g). Lesional scalp from alopecia areata patients grafted ontonude mice regrows hair coincident with a loss of in®ltratinglymphocytes from the graft (Gilhar and Krueger, 1987). Alopeciaareata responds to immunosuppressive doses of systemic steroids.The condition also has an association with additional autoimmuneprocesses such as autoimmune thyroiditis (Milgraum et al, 1987)and vitiligo (Shellow et al, 1992). Circulating autoantibodies tofollicular structures are present in alopecia areata, but they are alsoreported in normal controls (Tobin et al, 1997), and there is noconsistent pattern of reactivity of these antibodies. Furthermore, itis not possible to transfer alopecia areata by injection of IgG intohuman skin explants on nude mice (Gilhar et al, 1992).
Severe combined immunode®ciency (SCID) mice do not rejecthuman lymphocytes or skin. SCID mice grafted with human skinand injected with human lymphocytes can be used to study theimmunopathogenesis of human skin conditions, and have beenused to demonstrate the role of lymphocytes in induction(Boehncke et al, 1996; Wrone-Smith and Nickoloff, 1996) andmaintenance (Gilhar et al, 1997) of psoriasis lesions. We have sinceextended this model to the use of human scalp explants on SCIDmice to study alopecia areata.
We have shown that it is possible to transfer alopecia areata tohuman scalp explants on SCID mice by the injection of scalp-in®ltrating T lymphocytes (Gilhar et al, 1998). It was necessary to®rst activate the T lymphocytes in vitro by culture with hair folliclehomogenate in the presence of antigen-presenting cells (APC).Culture with nonfollicular scalp homogenate did not induce hairloss (Gilhar et al, 1999). Thus, alopecia areata is mediated by
Manuscript received May 31, 2001; revised August 2, 2001; accepted forpublication August 14, 2001.
Reprint requests to: Dr. Amos Gilhar, Laboratory for Skin Research,Rappaport Building, Technion Faculty of Medicine, P.O.B. 9649, Bat-Galim, Haifa 31096, Israel.
0022-202X/01/$15.00 ´ Copyright # 2001 by The Society for Investigative Dermatology, Inc.
1357

T lymphocytes that recognize a hair follicle antigen. We have sincefound that optimal transfer of hair loss requires both CD4+ andCD8+ T cells (Gilhar et al, in press). Depletion of either CD8+ Tcells (McElwee et al, 1996) or CD4+ T cells (McElwee et al, 1999)can reverse alopecia areata in the Dundee experimental bald rat,supporting a synergy or cooperation between CD8+ and CD4+ Tcells. In¯ammatory T cells of alopecia areata are cytotoxic andpossess both the Fas/Fas ligand and granzyme B cytotoxicmechanisms (Bodemer et al, 2000). It is proposed that the CD4+T cells provide help for the effector function of the CD8+ T cells.
It is hypothesized that the autoantigen of alopecia areata ismelanocyte derived (Paus et al, 1994). The initial basis for this is thefrequent clinical observation that with disease activity pigmentedhairs are lost preferentially to nonpigmented (e.g., white) hairs.Furthermore, with regrowth, there is a tendency for the initialregrowing hairs to be white. Melanocytes are a signi®cantcomponent of the hair bulb, which is the site of immunologicattack. Furthermore, there is an association of alopecia areata withvitiligo (Shong and Kim, 1991; Shellow et al, 1992), and hair bulbmelanocytes in alopecia areata show both histologic and ultra-structural abnormalities (Tobin et al, 1990). It is possible to inducelymphocyte-mediated alopecia in mice immunized against mela-nocyte-associated (e.g., melanoma) antigens (Becker et al, 1996).
There is a large literature on melanocyte-associated T cellepitopes generated by the search for melanoma-associated antigens(Bakker et al, 1995; Rivoltini et al, 1995; Parkhurst et al, 1996;Salgaller et al, 1996; Fleischhauer et al, 1997; Salazar-Onfray et al,1997; Bettinotti et al, 1998; Kittlesen et al, 1998). Most interest hasfocused on peptides presented by HLA-A2, which is present inapproximately half the population1. HLA-A2 may also be associatedwith alopecia areata (Hordinsky et al, 1984). Many of the antigenicpeptides are derived from melanosome-associated proteins such asgp100, MART-1/Melan-A, melanocortin 1 receptor (MC1R), ortyrosinase, and are present in both normal melanocytes andmelanomas. MAGE antigens tend to be preferentially expressedon melanoma cells, and are less likely to function as autoantigens foralopecia areata.
The goal of this study was to test the hypothesis that melanocyte-associated antigens can function as autoantigens to induce hair lossin alopecia areata. For this purpose, HLA-A2-positive patients withalopecia areata were selected for studies of HLA-A2-restrictedmelanocyte peptide epitopes. Scalp T cells were cultured withautologous APC and either hair follicle homogenate (positivecontrol) or melanocyte T cell epitopes. The cells were thentransferred to autologous scalp explants on SCID mice, inducinghair loss.
MATERIALS AND METHODS
Patients Ten patients with either alopecia totalis or severe alopeciaareata were studied. Specimens from four patients were used forexperiments with melanoma homogenate, and specimens from sixpatients were used for experiments with melanocyte-associated peptides.HLA-A2-positive alopecia areata patients were selected for the peptidecomponent of this study. The four donors used for the melanomahomogenate experiment were not HLA typed. Severe alopecia areatawas de®ned as large areas of alopecia with small residual areas of hair.These patients would be categorized as S4 (76%±99% hair loss) by thealopecia areata investigational assessment guidelines (Olsen et al, 1999).Duration, age, sex, and clinical characteristics of all donors are listed inTable I. There were no other preselection criteria, and data on these 10patients were previously unpublished. Informed consent was obtainedafter the nature and possible consequences of the studies were explained.Protocols were approved by the Institutional Review Board. None ofthe patients had any therapy in the 60 d before obtaining the scalpbiopsy specimens. HLA-A2 expression was determined bymicrolymphocytotoxicity assay (Immunology Allergy and AIDS Institute,Tissue Typing Laboratory, B. Rappaport Faculty of Medicine, Technionand Rambam Medical Center, Haifa).
Punch biopsies (2 mm) from involved scalp were obtained from eachpatient. Four biopsies from each patient were used for the isolation ofcutaneous T cells. Two biopsies were snap-frozen in liquid nitrogen forimmunoperoxidase staining. One frozen biopsy was sectioned vertically,and the other was sectioned horizontally. An additional biopsy from eachpatient was submitted for routine histologic examination with hematox-ylin and eosin and horizontally sectioned in its entirety. Punch biopsies(2 mm) were grafted to SCID mice in sets of three biopsies per mouse,one mouse per group per subject.
Animals CB-17 Prkdcscid (SCID) mice (Charles River, U.K.), 2±3 mo of age, were used in this study. The mice were raised in thepathogen-free animal facility of the B. Rappaport Faculty of Medicine,Technion-Israel Institute of Technology. Animal care and researchprotocols were in accordance with institutional guidelines.
Skin grafting Graft transplantation to SCID mice was performed aspreviously described (Gilhar and Krueger, 1987; Gilhar et al, 1988,1998). Each 2 mm graft was inserted through an incision in the skin intothe subcutaneous tissue over the lateral thoracic cage of each mouse, andcovered with a standard Band Aid dressing. The dressing was removedon day 7.
Isolation of T lymphocytes from scalp punch biopsies Fourpunch biopsies from each patient were used for T cell isolation. Tissue-in®ltrating lymphocytes were isolated from scalp punch biopsies with theuse of collagenase (Sigma, St Louis, MO), as described previously (Gilharet al, 1998).
Isolation of human peripheral blood mononuclear cells(PBMC) PBMC were isolated from heparinized whole blood bycentrifugation over Hypaque 1077 (Amersham Pharmacia Biotech,Uppsala, Sweden).
Hair follicle homogenate preparation Anagen hair follicles wereisolated under a stereodissecting microscope from normal scalp biopsiesobtained from healthy subjects who underwent hair transplantationprocedures. The follicles were homogenized and processed as previouslydescribed (Gilhar et al, 1998).
Peptides Peptides were selected based on identi®cation in theliterature as HLA-A2-restricted melanocyte (e.g., melanoma) associated Tcell epitopes. The following peptides were commercially synthesized(Chiron Technologies, Raleigh, NC) with free amino and carboxylicacid termini: Gp100/G9-154, KTWGQYWQV (Bakker et al, 1995;Parkhurst et al, 1996; Salgaller et al, 1996); Gp100/G9-209,ITDQVPFSV (Bakker et al, 1995; Parkhurst et al, 1996; Salgaller et al,1996); Gp100/G9-280, YLEPGPVTA (Bakker et al, 1995; Parkhurst etal, 1996; Salgaller et al, 1996); MC1R 291, AIIDPLIYA (Salazar-Onfrayet al, 1997); MC1R 244, TILLGIFFL (Salazar-Onfray et al, 1997);MC1R 283, FLALIICNA (Salazar-Onfray et al, 1997); MART-1 (27±35), AAGIGILTV (Bettinotti et al, 1998; Rivoltini et al, 1995); andtyrosinase, AFLPWHRFL (Kang et al, 1995). Peptides were negative forendotoxin by the Limulus Amebocyte Lysate method (Bactochem, NesZiona, Israel).
Human melanoma cells MeWo cells were obtained from Dr. Z.Smetana (Central Virology Laboratory, Chaim Sheba Medical Center,
Table I. Clinical summary of alopecia areata patients
Patient Age Sex Alopecia subtype Duration
A 31 F Severea 8 yB 32 M Severe 4 yC 26 F Severe 10 yD 26 F Severe 10 y1 23 M Severe 7 y2 39 F Severe 2 y3 36 M Severe 2 y4 32 M Totalis 6 y5 22 F Severe 10 y6 31 M Totalis 10 y
aSevere alopecia areata is de®ned as large areas of alopecia with small residualareas of hair, or nearly complete alopecia totalis (76%±99% hair loss). Donors A±Dwere studied in the melanoma homogenate experiment. Donors 1±6 are HLA-A2positive and were studied in the melanomo peptide experiments.
1Antigen frequencies from Histocompatibility Testing 1980. Los Angeles:UCLA Tissue Typing Laboratory, 1980.
1358 GILHAR ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY

Tel Hashomer, Israel) (Sauvaigo et al, 1986). They were propagated inRPMI-1640 supplemented with 10% fetal bovine serum (FBS), 1%glutamine, 1% sodium pyruvate, plus penicillin/streptomycin.
Culture of T lymphocytes Lymphocytes were plated at 1 3 105
cells per well in RPMI-FCS, along with irradiated (5000 R) PBMC(1 3 105 cells per well) in 24 plate wells (Greiner, Lake Mary, FL), asfeeders. After 3 d of culture, recombinant interleukin-2 (rIL-2, 10 U perml; Genzyme Diagnostics, San Carlos, CA) was added. Every 7 d the Tcells were re-stimulated with feeders, for a total of 21 d culture. Asindicated for experimental groups, lymphocytes were stimulated witheither follicular homogenate (10 mg per ml) or peptide (10 mg per ml),along with the feeder cells at each stimulation.
Phenotypic characterization of cultured Tlymphocytes Phenotype of the cultured T cell lines was determinedby immunocytochemistry on cytospin preparations, using procedures wehave previously published (Gilhar et al, 1997). This approach waspreferred over cyto¯uorograph analysis because of the small numbers ofcells available. The following monoclonal antibodies were used: anti-CD3 FITC (clone SK 7, Becton Dickinson), anti-CD4 (IgG2a, S3.5),anti-CD8 (IgG2a, 3B5) (Caltag, Burlingame, CA), anti-HLA-DR(IgG2a, DK22, Dako, Glostrup, Denmark). Subtype matched mouseIgG-FITC (Becton Dickinson) was used as a negative control.
Injection of cultured T lymphocytes into grafts on SCIDmice The SCID mice bearing grafts of lesional scalp were divided intogroups as indicated for each experiment. Between the twenty-third andthirtieth days, the grafts were injected intradermally (0.1 ml) withautologous lymphocytes as noted for each experiment. On day 90biopsies were obtained for immunohistochemical and histologic analysis.Both the percentage of grafts with hair and the number of hairs per graftwere recorded.
Immunohistochemical staining Staining was performed as reportedpreviously (Gilhar et al, 1998). Monoclonal antibodies to human antigensused were as follows: anti-HLA-DR, anti-CD54 (ICAM-1) (Biodesign,Carmel, NY), anti-CD3, anti-CD4, anti-CD8, anti-CD25 (Dako), andanti-HLA-class 1 (Dako). Each specimen was coded at the time ofbiopsy and evaluated by two observers who were blind to the coding.
Statistical analysis Statistical analysis was carried out using ANOVAfor multiple comparisons.
RESULTS
Melanocyte-associated antigens can function asautoantigens in alopecia areata Lesional scalp T cells werecultured with human melanoma cell homogenates (MeWo cells) intwo concentrations (10 mg per ml and 30 mg per ml), along withautologous APC and IL-2 for 24 d. The positive control wasprovided by incubation of scalp T cells with hair folliclehomogenate as a source of autoantigen. Scalp T lymphocytesrecovered from the cultures with melanoma homogenate (30 mgper ml) were then injected into autologous scalp grafts on SCIDmice, as were scalp T cells incubated with hair follicle homogenate,or APC alone with IL-2. Signi®cant (p < 0.01 by ANOVA) hairloss was noted in grafts injected with T cells incubated either withhuman melanoma homogenate (MeWo cells) or human hair folliclehomogenate. Figure 1 shows results from four patients. Thisdemonstrates that it was possible to transfer alopecia areata to scalpexplants on SCID mice by activation of lesional T cells withmelanoma antigens.
HLA-2-restricted melanoma peptide epitopes can act asautoantigens for transfer of hair loss in the human scalpexplant/SCID mouse system HLA-A2-restrictedmelanocyte peptide epitopes were tested as potential autoantigensfor alopecia areata. HLA-A2-positive alopecia areata patients wereselected for this study. In a preliminary study, PBMC from thesepatients were incubated with a panel of melanoma peptides at10 mg per ml, and then cultured with IL-2. Fresh APC and peptidewere added to the cultures every 7 d. At the end of 24 d of culturethe viable cell recovery was determined by trypan blue count. Theresults from this preliminary experiment were used to determinethe choice of peptide for each donor in the subsequent experimentusing lesional scalp T cells. It would be expected that the
autoreactive T cells would be enriched in lesional skin relative toperipheral blood (Kalish and Johnson, 1990).
For transfer experiments, lesional scalp lymphocytes from sixHLA-A2-positive donors were cultured with autologous APC, pluseither IL-2 medium alone, hair follicle homogenate (positivecontrol), or one or two melanocyte T cell epitope peptides.Table II shows the lymphocyte recovery after 24 d. The initialnumber of lymphocytes per group was 5 3 105. Cell recoveryfrom peptide and follicular homogenate groups varied between5.8 3 106 and 19 3 106. The responding T cells were phenotypedby immunohistochemistry analysis of cytospin preparations(Table III). The T cells were shown to be activated as judgedby expression of CD25 (19%±30% positive). CD4+ T cellspredominated for all donors.
Lesional scalp T cells from the above cultures (Table II) wereinjected into autologous scalp grafts on SCID mice as previouslydescribed. Six donors, all HLA-A2 positive, were used in twoexperiments. The negative control scalp grafts were not injectedwith T cells. Additional negative control scalp grafts were injectedwith T cells cultured with APC and IL-2 in the absence ofexogenous antigen, or PBMC activated with IL-2 alone. Positivecontrol grafts were injected with T cells cultured with follicularhomogenate. Melanoma peptide was added, along with APC, tothe T cells cultured in the ®nal group as indicated in Table II.
Melanoma-peptide-activated T cells were able to signi®cantly (p< 0.001 by ANOVA) reduce the numbers of hairs regrowing inboth experiments (Table IV). T cells incubated with follicularhomogenate also resulted in signi®cant reduction in hair numbers.There was no reduction of hairs in grafts injected with T cellscultured with IL-2 and APC, or PBMC. For all six donors, no hairswere present at day 90 in any of the three grafts per donor fromgrafts injected with T cells activated by either melanocyte peptides,or hair follicle homogenate. This demonstrates that melanocytepeptide epitopes are capable of activating T cells to induce hair lossin alopecia areata.
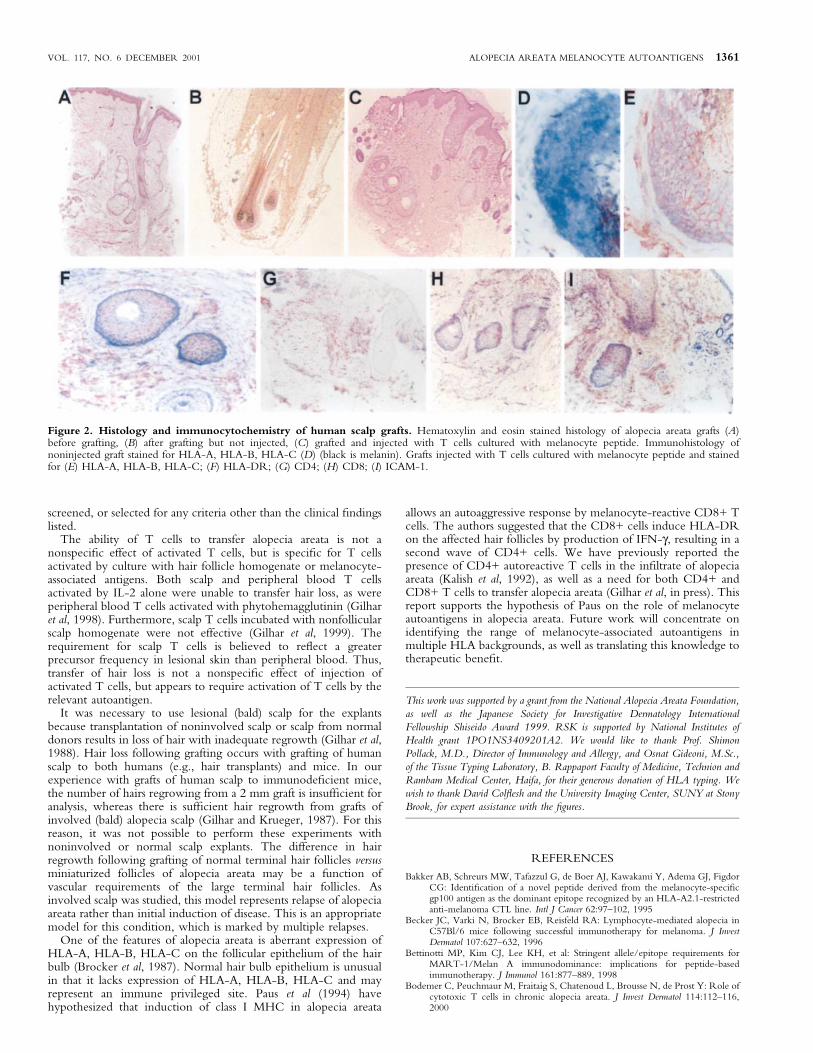
Melanocyte-peptide-induced transfer of alopecia areatainduces the histologic and immunologic features ofalopecia areata Biopsies were taken from the mice in theabove experiment after 90 d and were analyzed by both histologyand immunohistochemistry. Noninjected grafts showed growth ofnormal terminal anagen hairs. Injected grafts, by contrast, showeddystrophic hair follicles with a dense in®ltrate of CD4+ T cells, aswell as an intrafollicular in®ltrate of CD8+ T cells (Fig 2). Incontrast to noninjected grafts, the follicular epithelium of theinjected grafts expressed HLA-A, HLA-B, HLA-C, HLA-DR, andICAM-1 (Fig 2). These are known features of active alopeciaareata.
Figure 1. Hair loss induced by melanoma antigen. Signi®cant(p < 0.01 by ANOVA) hair loss was noted in grafts injected with T cellsincubated either with human melanoma homogenate (MeWo cells)30 mg per ml or human hair follicle homogenate. Data represent fourdonors.
VOL. 117, NO. 6 DECEMBER 2001 ALOPECIA AREATA MELANOCYTE AUTOANTIGENS 1359

DISCUSSION
The goal of this study was to test the hypothesis that melanocyte-associated antigens can function as autoantigens in alopecia areata.This was tested by isolating T cells from lesional scalp of alopeciaareata patients and culturing them with autologous APC, IL-2, andpotential autoantigens. The T cells were then injected intoautologous scalp explants on SCID mice. Hair regrows on thesescalp explants unless they are injected with autologous T cells thathave been appropriately activated. Previously we have demon-strated that homogenized hair follicle provides an autoantigen thatcan activate T cells for the transfer of alopecia areata. It was foundthat melanoma homogenate and melanocyte T cell epitopes canlikewise activate scalp T cells to prevent hair growth and inducehair loss.
The melanocyte-associated T cell epitopes used for this studywere originally identi®ed as HLA-A2-restricted melanoma-associ-ated T cell epitopes recognized by CD8+ T cells. We havepreviously demonstrated that transfer of alopecia areata requires aninteraction between CD8+ and CD4+ T cells, which was thereason for our choice of these peptides. HLA-A2 may be weaklyassociated with alopecia areata (Hordinsky et al, 1984). In our
screening, many alopecia areata patients were HLA-A2 negative,however. It is likely that other melanocyte-associated T cellepitopes, restricted by additional HLA alleles, can function asautoantigens in alopecia areata.
Five HLA-A2-restricted melanocyte peptides were identi®edwith the ability to act as autoantigens in alopecia areata (Gp100/G9-154, Gp100/G9-280, Gp100/G9-209, MC1R 291, MART-127±35). The initial choice of peptides tested was biased by thosepreviously reported in the melanoma literature. It is likely that themost immunodominant epitopes were preferentially identi®ed,however. Obviously, there is no single melanocyte antigen, orepitope, which is generally associated with alopecia areata. Epitopespreading is likely to have a role in the broad nature of theantimelanocyte response.
The observation that alopecia areata can be transferred by scalp Tcells activated by hair follicle homogenate has proven to be highlyreproducible. The effect has been observed to date in a total of 27patients. This includes the initial published series of six patients(Gilhar et al, 1998), 11 patients studied in CD4/CD8 separationexperiments (Gilhar et al, in press), as well as a positive control forthe 10 patients in this report. Aside from the six patients in thisstudy selected for HLA-A2 positivity, the patients were not
Table III. Phenotype of scalp T cells from HLA-A2+ donors responding to peptide epitopesa
Antigen P-1 P-2 P-3 P-4 P-6
Peptide Gp100/G9-280 MC1R 291 Gp100/G9-280 MART-1 27±35 Gp100\G9-154CD4+ 54% 85% 39% 54% 55%CD8+ 36% 15% 26% 40% 36%CD25+ 25% 21% 19% 30% 25%
aPhenotype was determined by immunohistochemistry on cytospin preparations.
Table IV. Effects of melanocyte peptide stimulation on hairs per grafta
Group Antigen stimulus
Experiment 1Donors 1, 2, 3Hairs per graft (STD)
Experiment 2Donors 4, 5, 6Hairs per graft (STD)
1 No T cells injected 3.8 6 0.5 3.5 6 1.72 Hair follicle homogenate 0.0 6 0 0.0 6 03 Autologous PBMC 3.2 6 0.5 3.4 6 1.64 Scalp T cells in IL-2, no homogenate 3.5 6 0.5 ND5 Peptide A 0.0 6 0 0.0 6 06 Peptide B ND 0.0 6 0
aScalp T cells or PBMC were incubated with either irradiated autologous PBMC and IL-2 alone or antigen provided by either hair follicle homogenate or melanocyticpeptides for 24 d. The T cells were then injected into autologous scalp explants on SCID mice, and hair growth was observed. The above data were obtained at 90 d. Thepatients and peptides studied above correspond to numbers in Tables I±III. The following comparisons are all signi®cant for both experiments (p < 0.001 by ANOVA),peptide A versus PBMC, peptide B versus PBMC, hair follicle homogenate versus PBMC, hair follicle homogenate versus no T cells.
Table II. Recovery of scalp T cells after culture with melanocyte peptide epitopes, and hair follicle homogenate HLA-A2alopecia areata patients
Antigen P-1 P-2 P-3 P-4 P-5 P-6
APC + IL-2, no antigen 3.9a 5.2 3.5 4.5 11.0 3.0Follicle homogenate 6.9 8.1 5.8 6.9 14.0 6.9Peptide A 10.0 11.0 6.9 9.4 12.0 7.0Peptide A Gp100/G9-280 MC1R 291 Gp100/G9-280 Gp100/G9-280 Gp100/G9-280 Gp100\G9-154Peptide B NA NA NA 6.7 19.0 6.5Peptide B NA NA NA MART-1 27±35 MART-1 27±35 Gp100/G9-209
aCell recovery3106 after 24 d culture with APC, and antigens as shown. Data are shown for a total of six patients. Initial cell number was 5 3104 for all groups. All pa-tients were HLA-A2 positive. Donors 1±6 correspond to the donor numbers in Table I.
1360 GILHAR ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY
screened, or selected for any criteria other than the clinical ®ndingslisted.
The ability of T cells to transfer alopecia areata is not anonspeci®c effect of activated T cells, but is speci®c for T cellsactivated by culture with hair follicle homogenate or melanocyte-associated antigens. Both scalp and peripheral blood T cellsactivated by IL-2 alone were unable to transfer hair loss, as wereperipheral blood T cells activated with phytohemagglutinin (Gilharet al, 1998). Furthermore, scalp T cells incubated with nonfollicularscalp homogenate were not effective (Gilhar et al, 1999). Therequirement for scalp T cells is believed to re¯ect a greaterprecursor frequency in lesional skin than peripheral blood. Thus,transfer of hair loss is not a nonspeci®c effect of injection ofactivated T cells, but appears to require activation of T cells by therelevant autoantigen.
It was necessary to use lesional (bald) scalp for the explantsbecause transplantation of noninvolved scalp or scalp from normaldonors results in loss of hair with inadequate regrowth (Gilhar et al,1988). Hair loss following grafting occurs with grafting of humanscalp to both humans (e.g., hair transplants) and mice. In ourexperience with grafts of human scalp to immunode®cient mice,the number of hairs regrowing from a 2 mm graft is insuf®cient foranalysis, whereas there is suf®cient hair regrowth from grafts ofinvolved (bald) alopecia scalp (Gilhar and Krueger, 1987). For thisreason, it was not possible to perform these experiments withnoninvolved or normal scalp explants. The difference in hairregrowth following grafting of normal terminal hair follicles versusminiaturized follicles of alopecia areata may be a function ofvascular requirements of the large terminal hair follicles. Asinvolved scalp was studied, this model represents relapse of alopeciaareata rather than initial induction of disease. This is an appropriatemodel for this condition, which is marked by multiple relapses.
One of the features of alopecia areata is aberrant expression ofHLA-A, HLA-B, HLA-C on the follicular epithelium of the hairbulb (Brocker et al, 1987). Normal hair bulb epithelium is unusualin that it lacks expression of HLA-A, HLA-B, HLA-C and mayrepresent an immune privileged site. Paus et al (1994) havehypothesized that induction of class I MHC in alopecia areata
allows an autoaggressive response by melanocyte-reactive CD8+ Tcells. The authors suggested that the CD8+ cells induce HLA-DRon the affected hair follicles by production of IFN-g, resulting in asecond wave of CD4+ cells. We have previously reported thepresence of CD4+ autoreactive T cells in the in®ltrate of alopeciaareata (Kalish et al, 1992), as well as a need for both CD4+ andCD8+ T cells to transfer alopecia areata (Gilhar et al, in press). Thisreport supports the hypothesis of Paus on the role of melanocyteautoantigens in alopecia areata. Future work will concentrate onidentifying the range of melanocyte-associated autoantigens inmultiple HLA backgrounds, as well as translating this knowledge totherapeutic bene®t.
This work was supported by a grant from the National Alopecia Areata Foundation,
as well as the Japanese Society for Investigative Dermatology International
Fellowship Shiseido Award 1999. RSK is supported by National Institutes of
Health grant 1PO1NS3409201A2. We would like to thank Prof. Shimon
Pollack, M.D., Director of Immunology and Allergy, and Osnat Gideoni, M.Sc.,
of the Tissue Typing Laboratory, B. Rappaport Faculty of Medicine, Technion and
Rambam Medical Center, Haifa, for their generous donation of HLA typing. We
wish to thank David Col¯esh and the University Imaging Center, SUNY at Stony
Brook, for expert assistance with the ®gures.
REFERENCES
Bakker AB, Schreurs MW, Tafazzul G, de Boer AJ, Kawakami Y, Adema GJ, FigdorCG: Identi®cation of a novel peptide derived from the melanocyte-speci®cgp100 antigen as the dominant epitope recognized by an HLA-A2.1-restrictedanti-melanoma CTL line. Intl J Cancer 62:97±102, 1995
Becker JC, Varki N, Brocker EB, Reisfeld RA: Lymphocyte-mediated alopecia inC57Bl/6 mice following successful immunotherapy for melanoma. J InvestDermatol 107:627±632, 1996
Bettinotti MP, Kim CJ, Lee KH, et al: Stringent allele/epitope requirements forMART-1/Melan A immunodominance: implications for peptide-basedimmunotherapy. J Immunol 161:877±889, 1998
Bodemer C, Peuchmaur M, Fraitaig S, Chatenoud L, Brousse N, de Prost Y: Role ofcytotoxic T cells in chronic alopecia areata. J Invest Dermatol 114:112±116,2000
Figure 2. Histology and immunocytochemistry of human scalp grafts. Hematoxylin and eosin stained histology of alopecia areata grafts (A)before grafting, (B) after grafting but not injected, (C) grafted and injected with T cells cultured with melanocyte peptide. Immunohistology ofnoninjected graft stained for HLA-A, HLA-B, HLA-C (D) (black is melanin). Grafts injected with T cells cultured with melanocyte peptide and stainedfor (E) HLA-A, HLA-B, HLA-C; (F) HLA-DR; (G) CD4; (H) CD8; (I) ICAM-1.
VOL. 117, NO. 6 DECEMBER 2001 ALOPECIA AREATA MELANOCYTE AUTOANTIGENS 1361

Boehncke WH, Dressel D, Zollner TM, Kaufmann R: Pulling the trigger onpsoriasis. Nature 379:777, 1996
Brocker EB, Echternach-Happle K, Hamm H, Happel R: Abnormal expression ofclass I and class II major histocompatibility antigens in alopecia areata:modulation by topical immunotherapy. J Invest Dermatol 88:564±568, 1987
Colon EA, Popkin MK, Callies AL, Dessert NJ, Hordinsky MK: Lifetime prevalenceof psychiatric disorders in patients with alopecia areata. Compr Psychiatry32:245±251, 1991
Fleischhauer K, Tanzarella S, Russo V, Sensi ML, van der Bruggen P, Bordignon C,Traversari C: Functional heterogeneity of HLA-A*02 subtypes revealed bypresentation of a MAGE-3-encoded peptide to cytotoxic T cell clones. JImmunol 159:2513±2521, 1997
Gilhar A, Krueger GG: Hair growth in scalp grafts from patients with alopecia areataand alopecia universalis grafted onto nude mice. Arch Dermatol 123:44±50, 1987
Gilhar A, Pillar T, Etzioni A: The effect of topical cyclosporin on the immediateshedding of human scalp hair grafted onto nude mice. Br J Dermatol 119:767±770, 1988
Gilhar A, Pillar T, Assy B, David M: Failure of passive transfer of serum from patientswith alopecia areata and alopecia universalis to inhibit hair growth in transplantsof human scalp skin grafted on to nude mice. Br J Dermatol 126:166±171, 1992
Gilhar A, David M, Ullman Y, Berkutski T, Kalish RS: T-Lymphocyte dependenceof psoriatic pathology in human psoriatic skin grafted to SCID mice. J InvestDermatol 109:283±288, 1997
Gilhar A, Ullmann Y, Berkutzki T, Assy B, Kalish RS: Alopecia areata transferred tohuman scalp explants on SCID mice with T-lymphocyte injections. J ClinInvest 101:62±67, 1998
Gilhar A, Shalaginov R, Assy B, Sera®movich S, Kalish RS: Alopecia areata is a T-lymphocyte mediated autoimmune disease: lesional human T-lymphocytestransfer alopecia areata to human skin grafts on SCID mice. J Invest DermatolSymp Proc 4:207±210, 1999
Gilhar A, Landau M, Assy B, Shalaginov R, Sera®movich S, Kalish RS: Alopeciaareata is mediated by cooperation between CD4+ and CD8+ T-lymphocytes:transfer to human scalp explants on Prkdcscid mice. Arch Dermatol, in press
Hordinsky MK, Hallgren H, Nelson D, Filipovich AH: Familial alopecia areata. HLAantigens and autoantibody formation in an American family. Arch Dermatol120:464±468, 1984
Kalish RS, Johnson KL: Enrichment and function of urushiol (poison ivy) speci®c T-lymphocytes in lesions of allergic contact dermatitis to urushiol. J Immunol145:3706±3713, 1990
Kalish RS, Johnson KL, Hordinsky MK: Autoreactive T-cells are variably enrichedrelative to peripheral blood in the scalp lesions of alopecia areata. Arch Dermatol128:1072±1077, 1992
Kang X, Kawakami Y, el-Gamil M, et al: Identi®cation of a tyrosinase epitoperecognized by HLA-A24-restricted, tumor-in®ltrating lymphocytes. J Immunol155:1343±1348, 1995
Khoury EL, Price VH, Greenspan JS: HLA-DR expression by hair folliclekeratinocytes in alopecia areata: evidence that it is secondary to thelymphoid in®ltration. J Invest Dermatol 90:193±200, 1988
Kittlesen DJ, Thompson LW, Gulden PH, et al: Human melanoma patients recognizean HLA-A1-restricted CTL epitope from tyrosinase containing two cysteineresidues: implications for tumor vaccine development. J Immunol 160:2099±2106, 1998
Koo JY, Shellow WV, Hallman CP, Edwards JE: Alopecia areata and increasedprevalence of psychiatric disorders. Intl J Dermatol 33:849±850, 1994
McElwee KJ, Spiers EM, Oliver RF: In vivo depletion of CD8+ T-cells restores hairgrowth in the DEBR model for alopecia areata. Br J Dermatol 135:211±217,1996
McElwee KJ, Spiers EM, Oliver RF: Partial restoration of hair growth in the DEBRmodel for alopecia areata after in vivo depletion of CD4+ T cells. Br J Dermatol140:432±437, 1999
Messenger AG, Bleehen SS: Expression of HLA-DR by anagen hair follicles inalopecia areata. J Invest Dermatol 85:569±572, 1985
Milgraum SS, Mitchell AJ, Bacon GE, Rasmussen JE: Alopecia areata, endocrinefunction, and autoantibodies in patients 16 years of age or younger. J Am AcadDermatol 17:57±61, 1987
Olsen E, Hordinsky M, McDonald-Hull S, Price V, Roberts J, Shapiro J, Stenn K:Alopecia areata investigational assessment guidelines. J Am Acad Dermatol40:242±246, 1999
Parkhurst MR, Salgaller ML, Southwood S, Robbins PF, Sette A, Rosenberg SA,Kawakami Y: Improved induction of melanoma-reactive CTL with peptidesfrom the melanoma antigen gp100 modi®ed at HLA-A*0201-binding residues.J Immunol 157:2539±2548, 1996
Paus R, Slominski A, Czarnetzki BM: Is alopecia areata an autoimmune-responseagainst melanogenesis-related proteins, exposed by abnormal MHC class Iexpression in the anagen hair bulb? Yale J Biol Med 66:541±554, 1994
Rivoltini L, Kawakami Y, Sakaguchi K, et al: Induction of tumor-reactive CTL fromperipheral blood and tumor-in®ltrating lymphocytes of melanoma patients byin vitro stimulation with an immunodominant peptide of the human melanomaantigen MART-1. J Immunol 154:2257±2265, 1995
Safavi KH, Muller SA, Suman VJ, Moshell AN, Melton LJ 3rd: Incidence of alopeciaareata in Olmsted County, Minnesota, 1975 through 1989. Mayo Clinic Proc70:628±633, 1995
Salazar-Onfray F, Nakazawa T, Chhajlani V, et al: Synthetic peptides derived fromthe melanocyte-stimulating hormone receptor MC1R can stimulate HLA-A2-restricted cytotoxic T lymphocytes that recognize naturally processed peptideson human melanoma cells. Cancer Res 57:4348±4355, 1997
Salgaller ML, Marincola FM, Cormier JN, Rosenberg SA: Immunization againstepitopes in the human melanoma antigen gp100 following patientimmunization with synthetic peptides. Cancer Res 56:4749±4757, 1996
Sauvaigo S, Fretts RE, Riopelle RJ, Lagarde AE: Autonomous proliferation ofMeWo human melanoma cell lines in serum-free medium: secretion ofgrowth-stimulating activities. Int J Cancer 37:123±132, 1986
Shellow WV, Edwards JE, Koo JY: Pro®le of alopecia areata: a questionnaire analysisof patient and family. Intl J Dermatol 31:186±189, 1992
Shong YK, Kim JA: Vitiligo in autoimmune thyroid disease. Thyroidology 3:89±91,1991
Tobin DJ, Fenton DA, Kendall MD: Ultrastructural observations on the hair bulbmelanocytes and melanosomes in acute alopecia areata. J Invest Dermatol94:803±807, 1990
Tobin DJ, Hann SK, Song MS, Bystryn JC: Hair follicle structures targeted byantibodies in patients with alopecia areata. Arch Dermatol 133:57±61, 1997
Todes-Taylor N, Turner R, Wood GS, Stratte PT, Morhenn VB: T cellsubpopulations in alopecia areata. J Am Acad Dermatol 11:216±223, 1984
Wrone-Smith T, Nickoloff BJ: Dermal injection of immunocytes induces psoriasis. JClin Invest 98:1878±1887, 1996
1362 GILHAR ET AL THE JOURNAL OF INVESTIGATIVE DERMATOLOGY





























