Embed Size (px)
Citation preview

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 245-256
MEJOR PREDICTOR LINEAL E INSESGADO FAMILIAR DE APTITUD COMBINATORIA GENERAL EN EXPERIMENTOS PARCIALES DE CRUZAS DIALÉLICAS CON EFECTOS MATERNOS*
BEST LINEAR UNBIASED FAMILIAR PREDICTOR FOR PARTIAL DIALLEL EXPERIMENTS WITH MATERNAL EFFECTS
Osval Antonio Montesinos-López1§, Ángel Agustín Mastache-Agunas2, Ignacio Luna-Spinoza3, Carlos Moisés Hernández-Suárez4 y Guadalupe Hernández-Lira5
1Facultad de Telemática, Universidad de Colima. Av. Universidad # 333. Campus Colima. C. P. 28040. 2Centro de Estudios Profesionales, Colegio Superior Agropecuario del Estado de Guerrero. 3Universidad del Istmo-Campus Ixtepec. Cd. Universitaria s/n, 70110, Ixtepec, Oaxaca. 4Facultad de Ciencias, Universidad de Colima. Av. Universidad # 333. Campus Colima. C. P. 28040. 5Instituto de Socioeconomía, Estadística e Informática, Programa en Estadística, Colegio de Postgraduados. 56230. Montecillo, Estado de México, México. §Autor para correspondencia: [email protected].
* Recibido: Noviembre, 2006
Aceptado: Marzo, 2009
RESUMEN
En el mejoramiento genético de plantas y animales se han utilizado los experimentos de cruzas dialélicas (diseños completos de Griffing) para realizar estimaciones de parámetros y pruebas de hipótesis, lo cual es importante para la toma de decisiones en programas de mejoramiento genético. Los esquemas completos de Griffing son útiles cuando el número de líneas progenitoras es pequeño, sin embargo, cuando este número es elevado es difícil preparar, establecer y conducir los trabajos de campo. Una alternativa consiste en emplear los diseños parciales de cruzas dialélicas, los cuales ensayan un subconjunto del total de cruzas que es posible formar entre los progenitores básicos. Estos experimentos pueden ser simétricos o asimétricos. Además, es frecuente que los investigadores formen grupos o familias de progenitores, lo que implica considerar la estimación de los efectos del grupo o familia. No obstante, hasta ahora no se ha realizado una investigación para obtener el Mejor Predictor Lineal Insesgado del efecto familiar o de grupo considerando los efectos maternos en experimentos parciales bajo el modelo de efectos mixtos. Por lo que en éste trabajo se derivan los MPLI familiares para aptitud combinatoria general en experimentos dialélicos parciales con efectos maternos, y se realiza un algoritmo computacional en comandos SAS-IML que permite la aplicación de la metodología propuesta.
Palabras clave: aptitud combinatoria general, cruzas dialélicas, experimentos parciales, modelo de efectos mixtos.
ABSTRACT
In plant or animal breeding the experiments with diallel crosses (complete Griffing´s designs) have been frequently used to estimate parameters and test hypothesis, which is really important for decision making in breeding programs. Nevertheless, the complete Griffing´s designs are useful when the number of parental lines is reduced. With a high number of parental lines is difficult to prepare, stablish and conduct the trials in the field. In this situation a partial diallel experiment is recommended, which can be symetric or asymetric. An option is to use the partial diallel designs, which test a subset from the total number of crosses that is possible to develop among the parental lines. These trials can either be symetric or asymetric. In addition is frequent that researchers conform groups of families or parents that implies the consideration of estimating group or family effects. Nevertheless, so far no research has been conducted to estimate the best unbiased lineal predictor of group or family effect taking into account maternal effects

Osval Antonio Montesinos-López et al.246 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
in partial designs under a mixed model. For the above reason, in this research the best unbiased lineal predictor of familias for general combining ability in parcial diallel designs, including maternal effects, are derived, and a computer algorithm in SAS-IML commands that allows for the application of the proposed methodology.
Key words: diallel crosses, general combining ability, mixed effects model, partial diallel experiments.
INTRODUCCIÓN
En el mejoramiento genético de plantas y animales se han utilizado extensamente los experimentos de cruzas dialélicas para realizar estimaciones de parámetros y pruebas de hipótesis. Los diseños de Griffing (1956a, 1956b) sirven para estimar aptitud combinatoria general (ACG), aptitud combinatoria específica (ACE), efectos maternos (EM), efectos recíprocos (ER) y componentes de varianza. Estas estimaciones son importantes en la toma de decisiones de programas de mejoramiento genético. Los esquemas completos de Griffing son útiles cuando el número de líneas progenitoras es reducido, sin embargo, cuando el número de líneas progenitoras es elevado es difícil preparar, establecer y conducir los trabajos de campo. Una solución alternativa consiste en emplear los diseños parciales de cruzas dialélicas, los cuales ensayan un subconjunto del total de cruzas que es posible formar entre los progenitores básicos; proporcionado de esta manera una herramienta más elástica al genetista.
Los experimentos parciales de cruzas dialélicas pueden ser simétricos o asimétricos. Los primeros requieren que cada progenitor se involucre con el mismo número de cruzas; los últimos requieren que al menos uno de los progenitores participe en un número diferente de cruzas. Los más útiles son los que cumplen condiciones de simetría.
Algunas soluciones de diseño cuando el número de líneas progenitoras es elevado son las propuestas por Kempthorne y Curnow (1961), Fyfe y Gilbert (1963) y Rojas (1973). Estos autores han abordado el análisis y la estimación de parámetros a partir de un modelo de efecto fijos. Para un modelo de efectos mixtos, Mastache (1999) sugiere derivar los mejores predictores lineales e insesgados (MPLI) mediante la metodología desarrollada por Herderson (1963, 1973), Harville (1976), Harville y Carriquiry (1992) para obtener estimadores insesgados y de mínima varianza.
Adicionalmente, es común que los investigadores formen grupos o familias de progenitores, como lo hacen Cervantes et al. (1999) y Ron et al. (1999), de forma tal que los miembros de cada grupo o familia tienen características similares. Esto implica considerar la estimación de los efectos del grupo o familia. No obstante, hasta ahora no se ha realizado una investigación para obtener el MPLI del efecto familiar o de grupo bajo el modelo de efectos mixtos. En este trabajo se derivan los MPLI empíricos de los efectos familiares en experimentos dialélicos parciales con efectos maternos bajo el modelo de efectos mixtos. Adicionalmente, se proporciona un algoritmo computacional en comandos SAS-IML que permiten la aplicación de la metodología propuesta.
MATERIALES Y MÉTODOS
Modelo lineal de efectos mixtos
En términos matriciales, el modelo lineal de efectos mixtos se representa mediante la forma
Y=Xα+Z θ +ε , (1)
),0( 2eGσθ~θ
,)()( 22ee RZZGVyVar σσ +′==
y ε es un vector nxl no
ε~ ).,0( 2eRσ αXyE =)(
y G = matriz diagonal tal que donde, ,)/( 22pe IG σσθ=
2θσ
2θσ
),0( 2ep GN σ ),0( 2
en RN σε
donde, Y= vector nxl de observaciones, X= matriz diseño nxf conocida, α = vector fxl de parámetros desconocidos correspondientes a los efectos fijos, Z = matriz diseño nxp conocida, θ = vector pxl no observable de los efectos aleatorios tal que observable de efectos residuales o términos de error talque Además, = varianza del término aleatorio θ, = varianza de los términos de error, Ip = matriz identidad de orden p, y =R frecuentemente es la matriz identidad de orden n . Con base en el modelo (Ec. 1), Henderson (1963, 1973) desarrolló una técnica para abordar los aspectos aleatorios, derivando para ellos los MPLI.
Método de Henderson para obtener los MPLI
El método propuesto por Henderson (1963, 1973) para obtener los MPLI sobre la base del modelo de efectos mixtos (Ec. 1) supone que en la Ec. 1, θ y ε son vectores aleatorios no observables, tales que θ ~ y ~ , donde, G y R= matrices no singulares. Henderson (1963, 1973) reporta que la densidad conjunta de θ y y es f(y, θ)= d(y/ θ)c(θ),

Mejor predictor lineal e insesgado familiar de aptitud combinatoria general en experimentos parciales de cruzas dialélicas con efectos maternos 247
donde, c(θ )= densidad marginal de θ y d(y/ θ) = densidad condicional de y dado θ En forma explicita f (y, θ) es:
(2) .
Las siguientes ecuaciones normales del modelo mixto se obtienen maximizando la densidad conjunta (Ec. 2) con respecto a α y θ.
(3)
Las soluciones para α, en las ecuaciones normales (Ec. 3), son idénticas a las de mínimos cuadrados generalizados obtenidas de la ecuación X'V-1X'α=X'V-1y, donde, V= ZGZ'+R y Var(y)= Vσ2 , de que en las ecuaciones normales (Ec. 3) no se requiere la inversa de matriz V. De las ecuaciones normales (Ec. 3) se tiene que el MPLI para el vector aleatorio θ es:
(4)
En la mayoría de las aplicaciones, los componentes de varianza involucrados en G y R son desconocidos y deben estimarse a partir de la información experimental. De acuerdo con Harville y Carriquiry (1992), el procedimiento de estimación se divide en dos etapas. En la primera se estiman los componentes de varianza y en la segunda se utilizan estos estimadores, incluyéndolos en las ecuaciones normales (Ec. 3), para obtener el MPLI, llamado MPLI empírico. En esta investigación los componentes de varianza se obtienen por el método derivado del análisis de varianza, cuyas propiedades son descritas por (Robinson, 1991).
En algunas ocasiones, al realizar la estimación de los componentes de varianza con la información correspondiente se obtienen estimaciones negativas, en cuyo caso, Robinson (1991) sugiere considerarlos iguales a cero, aunque esto provoca que los estimadores sean sesgados.
Modelo lineal en experimentos dialélicos
Si se incluyen los efectos maternos, de acuerdo con Martínez (1983 y 1988), el modelo lineal apropiado para el análisis de experimentos dialélicos establecidos en el diseño de bloques completos al azar es:
(5)
donde, yijk= valor fenotípico observado de la cruza (i, j) en el bloque k, µ = efecto común a todas las observaciones, gi = efecto de aptitud combinatoria general del progenitor i, sij = efecto de aptitud combinatoria específica de la cruza (i, j), mi= efecto materno del progenitor i , lij= efecto recíproco de la cruza (i, j); δk = efecto del bloque k, y eijk= efecto aleatorio del error correspondiente a la observación (i, j, k). Los términos gi, sij, mi, lij y eijk se consideran como variables aleatorias normales no correlacionadas entre y dentro de ellas, con media cero y varianzas y respectivamente, con sij= sji y lij= -lji. Si sólo se consideran las medias de las cruzas y se elimina el efecto de bloques, dado que éste es ortogonal con las cruzas, la representación del modelo (Ec. 5) se reduce a:
(6)
Si se ensaya un subconjunto t ≤ p2 del total de cruzas posibles a partir de p progenitores, entonces también es posible utilizar el modelo (Ec. 6), el cual se expresará de la siguiente forma:
(7)
donde, nij= 1 si la cruza (i, j) participa en el experimento y nij= 0 en caso contrario. Los términos restantes de la Ec. 7 son idénticos a como se definieron en la Ec. 8, con la única diferencia de que ahora son los promedios por bloques.
RESULTADOS
Estimación de los MPLI familiares de ACG y EM
Cuando se ensayan las p(p-1)/2 cruzas directas y las p(p-1)/2 cruzas recíprocas se genera el diseño tres de Griffing, por lo que en este caso t= p(p-1). Cuando se ensayan las cruzas directas, las recíprocas y las p autofecundaciones se tiene el diseño uno de Griffing, por lo que t= p2 . En los experimentos parciales sólo se estudia una fracción del total de cruzas, es decir, t < p(p-1) cuando no se ensayan las autofecundaciones y t < p2 cuando éstas son consideradas en el experimento. Por lo tanto, la derivación analítica de los MPLI familiares de ACG con efectos maternos es similar
∝−− ′−
−−′−−− θθ
σθαθα
σθ1
21
2 21)()(
21
),(GZXyRZXy
ee eeyf
,2eσ,2
gσ ,2sσ ,2
mσ 2lσXR-1Xα + XR-1Zθ= XR-1y,
Z'R-1Xα + [Z'R-1Z + G-1]θ= Z'R-1y.
e
θ= [Z'R-1Z+G-1]-1(Z'R-1yV-1X α).
yijk= µ+gi+gj+sij+mi+mj+lij+δk+eijk1 ≤ i, j ≤ p, k=1,2...,r
_yij.= µ+gi+gj+sij+mi-mj+lij+ēij.
_nyjyij.= nij(µ+gi+gj+sij+mi-mj+lij+ēij.),
ˆ ˆ
ˆ
ˆˆ

Osval Antonio Montesinos-López et al.248 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
en ambos casos, con y sin las autofecundaciones, tomando la precaución de generar adecuadamente los elementos matriciales correspondientes.
MPLI familiar parcial de ACG
Si en el modelo (Ec. 7) se considera las siguientes igualdades:
entonces
En notación matricial se tiene que:
(8)
donde, y*= un vector txl de observaciones que contiene el promedio de las cruzas observadas, j = vector txl de unos, t= número total de cruzas; µ= escalar que representa el efecto común a todas las cruzas, Zg= matriz diseño de orden txp correspondiente a los efectos de la ACG, g = vector pxl tal que g ~Np(0,Gg σ2), s es un vector txl tal que s ~ Nt(0,Rsσ2) y e0 es un vector txl de términos de error tal que e0 ~ Nh(0,Re σ2). Debido a que s y e0 tienen una estructura de covarianzas similar, entonces es factible utilizar la igualdad e*= s+e0, la cual nos conduce a que la Ec. 8 pueda representarse como:
(9)
Además,Var(y*)= E[(y*-jµ)(y*-jµ)']= E[(Zgg+e*)(Zgg+e0)']
(10)
donde, a la matriz identidad
de orden p, S= matriz simétrica txt formada por unos en aquellas posiciones en las que la cruza es tal que ij= ji y por ceros en las posiciones restantes, E es una matriz similar a S, diferenciándose de ésta por la presencia de un dos en los casos en que i= j . Las matrices S y E son singulares, esto implica que R* también sea singular, pero, de acuerdo con Harville (1976), si R* es singular en las ecuaciones
normales del modelo mixto, entonces R*-1 se sustituye por
alguna inversa generalizada R*. Por lo antes expresado y por la (Ec. 10) se tiene un caso particular del modelo (Ec. 1) en el que n= t, f= 1, p= p y sus componentes son: X= j, α= µ, Z= Zg, θ = g, R= R* y ε= e*. Así, las ecuaciones normales del modelo mixto son:
(11)
Si se conocen los componentes de varianza , y , entonces el MPLI parcial para ĝ se obtiene al resolver el sistema de ecuaciones simultaneas (Ec. 11). Considerando la restricción en el sistema de ecuaciones
(Ec. 11) se tiene que j'R*Zg ĝ = 0. Por lo tanto µ= ( j'R* j)-1
j'R*y* y el MPLI de ACG es:
(12)
La Ec. 12 es el MPLI parcial de ACG y tiene la siguiente forma ĝ' = [ĝ1, ĝ2,..., ĝp]. Dado que necesitamos estimarlos efectos familiares, esto implica que con los p progenitores se formaran familias o grupos de progenitores. Por ejemplo, supóngase que se cuentan con p =6 progenitores que se agrupan en tres familias: f1={p1, p2}, f2 ={p3, p4}, f3={p5, p6}esto implica que ĝ' = [ĝ1, ĝ2, ĝ3, ĝ4, ĝ5, ĝ6] en términos de los efectos familiares es igual a f ' = [ f1, f2, f3] . Por tanto, para el cálculo del efecto de familias también se utiliza la Ec. 12, pero en términos de las familias formadas. Por ello, no es posible simplificar la Ec. 12 y expresarla en términos de las familias porque este número depende del investigador y porque no todas las cruzas estas participando.
MPLI parcial de EM
Si en el modelo (Ec. 7) se consideran las siguientes igualdades:
entonces
(13)
2eσ 2
sσ 2gσ
0ˆ1
=∑=
p
iig
_ _ (yij.+yji.) (ēij.+ ēji.)nij = nij(µ+gi+gj+sij)+nij
, 2 2 _ _ (yij.+yji.) (ēij.+ ēji.)y* =
y e0 =
, 2 2
ij. ij.
nij y*= nij(µ+gi+gj+sij+e0 ).
ij.
ij.
y*= jµ+Zg g+s+e0,
ee
e
y*= jµ+Zg g+e*
σ2E= Zg(σ2Ip)Z'+σ2S+
= [ZgGgZ'+R*]σ2,
g s
2r e
e[ ]))(()( ******** ′++=
′
= emZemZEyyEyVar mm [ ]))(()( ******** ′++=
′
= emZemZEyyEyVar mm
-
- - -j'R* jµ + j'R* Zg g= j'R* y
*
- - -1 -Z'R* jµ+[Z'R* Zg+Gg ]g=Z'R* y
*
ˆ --
-
- - - ĝ= [Z'R* Zg+G-1]-1(Z'R* y
*-Z'R* jµ).
g g g g
ˆ
_ _ (yij.+yji.) (ēij.+ ēji.)nij = nij(mi-mj+lij)+nij)+nij
, 2 2
_ _
1 1 y
* *= (yij.+ yji.), y e$ = (ēij.+ ēji.), 2 2
ij. ij.
nij y**= nij(mi-mj+lij+e$ )
ij.
ij.
σ2 σ2 EGg=
Ip, R*=
S+
, Ip= σ
2 σ2 2rg s
e ep
e
ggI G
=2
2
σσ
pe
gg IG
= 2
2
σσ
pe
gg IG
= 2
2
σσ
pe
ggI G
=2
2
σσ

Mejor predictor lineal e insesgado familiar de aptitud combinatoria general en experimentos parciales de cruzas dialélicas con efectos maternos 249
En notación matricial se tiene:
(14)
donde, y** es un vector txl de observaciones, Zm= a la matriz diseño de orden txp,= a los efectos maternos, m = vector de orden pxl tal que m ~Np(0,Gm σ2), l = a vector de orden txl tal que l ~ Nt(0, Rsσ2) y e$ = a un vector txl de términos de error tal que e$ ~Nt(0, Reσ2). Como l y e$ tienen una estructura de covarianzas similar, entonces es factible hacer e*= l+ e$, lo cual nos conduce a que y** pueda representarse como:
(15)
Además, Var(Y*)= E [Y**Y**']= E [(Zmm+e*)(Zmm+e*)']
(16)
donde, es la matriz identidad
de orden p, S* y E* son similares a S y E, con la diferencia de que en S* los unos fuera de la diagonal son negativos y en la matriz E* los términos correspondientes a las autofecundaciones son iguales a cero. Debido a que Rm es singular, se sustituye por alguna inversa generalizada (Harville, 1976). Por lo antes expresado y por la (Ec. 16) se tiene un caso particular del modelo (Ec. 2) en el que n= t, p = p y sus componentes son X= j, Z= Zm, θ= m, R= Rm y ε= e*. Así, las ecuaciones normales del modelo mixto son:
(17)
Si se conocen los componentes de varianza σe, σl y σm, entonces el MPLI parcial para se obtiene al resolver el sistema de ecuaciones simultaneas (Ec. 17). Considerando la restricción en el sistema de ecuaciones (Ec. 17) se
tiene que j 'RmZmm= 0. Por lo tanto, el MPLI parcial de EM es:
(18)
La Ec. 18 es el MPLI parcial de EM y tiene la siguiente forma m'= (m1, m2,..., mp). Como se requieren los efectos
fami l ia res de EM, con los p p rogeni tores se formaran fami l ias . De igual forma supóngase que se cuentan con p =6 progeni tores que se agrupan en tres familias: f1={p1, p2}, f2 ={p3, p4}, f3={p5, p6} esto implica que m'= (m1, m2, m3, m4, m5, m6)en términos de los efectos familiares es igual a f * '= [ f 1, f 2, f 3] . Por tanto, para el cálculo del efecto de familias también se utiliza la Ec. 18, pero en términos de las familias formadas. Esto conduce a que no sea posible simplificar la Ec. 18 y expresarla en términos de familias.
Componentes de varianza
De acuerdo con Martínez (1983), los estimadores obtenidos a través del método de análisis de la varianza para los diseños parciales de cruzas dialélicas con efectos maternos en el que se ensayan t ≤ p2 cruzas o tratamientos son:
donde, CME es el cuadrado medio del error, CMER es el cuadrado medio del efecto recíproco, CMEM es elcuadrado medio del efecto materno, CMACE es elcuadrado medio de aptitud combinatoria específicay C M A C G e s e l c u a d r a d o m e d i o d e a p t i t u d combinatoria general. Sustituyendo σe, σg, σs y en Ec. 12 se obt ienen los MPLI empír icos de ACG. De manera similar, sustituyendo σe, σl y σm en la Ec. 18 seobtienen los MPLI empíricos de EM.
Aplicación
A c o n t i n u a c i ó n s e a p l i c a l a m e t o d o l o g í apropues ta para de terminar e l MPLI fami l ia r de ACG y de EM en expe r imen tos pa rc i a l e s . Los datos utilizados corresponden a un experimentop a r c i a l d e c r u z a s d i a l é l i c a s ( C u a d r o 1 ) e n donde se consideran t= 24 cruzas a lojadas endos bloques completos al azar (r= 2). La solucións e p r e s e n t a e n f o r m a m a n u a l y e n f o r m a computacional.
0ˆ1
=∑=
p
iim
y**= Zmm+l+e$
e
e
e
y**= Zmm+e*.
σ2E*= Zm(σ2 Ip)Z' +σ2S*+
= [ZmGmZ' +Rm]σ2,
m m l
2r m
e
e[ ]))(()( ******** ′++=
′
= emZemZEyyEyVar mm [ ]))(()( ******** ′++=
′
= emZemZEyyEyVar mm
σ2 σ2 E*
Gg=
Ip, Rm=
S*+
, Ip σ
2 σ2 2rm l
e ep
e
ggI G
=2
2
σσ
pe
gg IG
= 2
2
σσ
pe
gg IG
= 2
2
σσ
pe
ggI G
=2
2
σσ
- -j'RmZmm= j'Rmy**
' - -1 ˆ ' -[ZmR*Zm+Gm]m= ZmRm y**
2 2 2
- ˆ
' - -1
' -m= [ZmRmZm+Gm ]-1ZmRm y
*.
ˆ
ˆ
ˆ * * *
2 2 2
p-1σe = CME, σl = (CMER-CME)/2r, σm=
(CMEM_CMER), 2rpsˆ
2 1 2 ( p-1)σs=
[CMACE-CME] y σg=
[CMACG-CMACE], 2r 2rs( p-2)ˆ
2 2 2
2 2 2

Osval Antonio Montesinos-López et al.250 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
;
;
MPLI familiar parcial de EM
De manera similar, para los valores medios de las cruzas se tiene que:
Cruza I J Dialélo Bloque Cruza I J Dialélo BloqueI II I II
1* 1 2 1 6.6 3.7 13 4 2 6 5 4.92* 1 3 2 5.5 5.4 14 4 3 8 5 4.33* 1 5 3 8 8.2 15* 4 5 10 6.4 6.54* 1 6 4 4.2 7.1 16* 4 6 11 8.2 65 2 1 1 6.5 5.5 17 5 1 3 5.5 7.16* 2 3 5 7.2 7.9 18 5 3 9 5 4.77* 2 4 6 6.3 4.9 19 5 4 10 5.2 3.48* 2 6 7 7.3 6 20* 5 6 12 4.5 59 3 1 2 6.1 5.3 21 6 1 4 6.6 5.710 3 2 5 6.4 6.5 22 6 2 7 5.1 4.511* 3 4 8 8.2 6 23 6 4 11 4.6 5.112* 3 5 9 4.5 5 24 6 5 12 3.8 3.5
Cuadro 1. Diseño parcial generado con los datos de Carballo et al. (1999).
Solución manual
MPLI familiar parcial de ACG
A partir de los datos del Cuadro 1 se tienen los siguientes resultados para valores medios de las cruzas:
=′ −
9416.207045.218857.226217.208678.205165.22
* gg ZRZ
=′ −
5896.215896.215896.215896.215896.215896.21
* µjRZ g
=
110000101000100010
000110000011100001010001000101000011
gZ , , -
−−
−−
=
−−−−−−
=
−
3698.00479.0
6115.04510.02604.0
51775.0
5896.219416.205896.217045.215896.218857.225896.216217.205896.218678.205896.215165.22
9373.39843.09843.009843.09843.09843.09373.39843.09843.009843.09843.09843.09373.39843.09843.0009843.09843.09373.39843.09843.0
9843.009843.09843.09373.39843.09843.09843.009843.09843.09373.3
ˆˆˆˆˆˆ
1
6
5
4
3
2
1
gggggg
Sustituyendo Zg , y en la Ec. 12 se tiene que el MPLI parcial de ACG es:
gg ZRZ −′ * µ* jRZ g−′
Si los progenitores se agrupan en las siguientes tres familias: , , . Reordenando términos se tiene que:
},{ 432 ppf = },{ 653 ppf =},{ 211 ppf =
−
−−
=
−−−−−−
=
6480.01149.02961.19679.07218.0
9269.0
5896.219416.205896.217045.215896.218857.225896.216217.205896.218678.205896.215165.22
ˆˆˆˆˆˆ
9373.39843.09843.00983.09843.09843.09373.39843.09843.009843.09843.09843.09373.39843.09843.0009843.09843.09373.39843.09843.0
9843.009843.09843.09373.39843.09843.09843.009843.09843.09373.3
3
3
2
2
1
1
ffffff
Esto nos conduce a que las ecuaciones normales en términos del efecto familiar son las siguientes:
−=
5331.03262.02051.0
ˆˆˆ
8432.99529.29529.29529.28432.99529.29529.29529.28432.9
3
2
1
fff
−
=
−
5331.03262.02051.0
8432.99529.29529.29529.28432.99529.29529.29529.28432.9
ˆˆˆ 1
3
2
1
fff
Finalmente, el MPLI familiar parcial de ACG es igual a:
−
−
−−−−−−
=
332.1118.2789.0
1179.00272.00272.00272.01179.00272.00272.00272.01179.0
ˆˆˆ
3
2
1
fff
−=
077.0047.0029.0
ˆˆˆ
3
2
1
fff
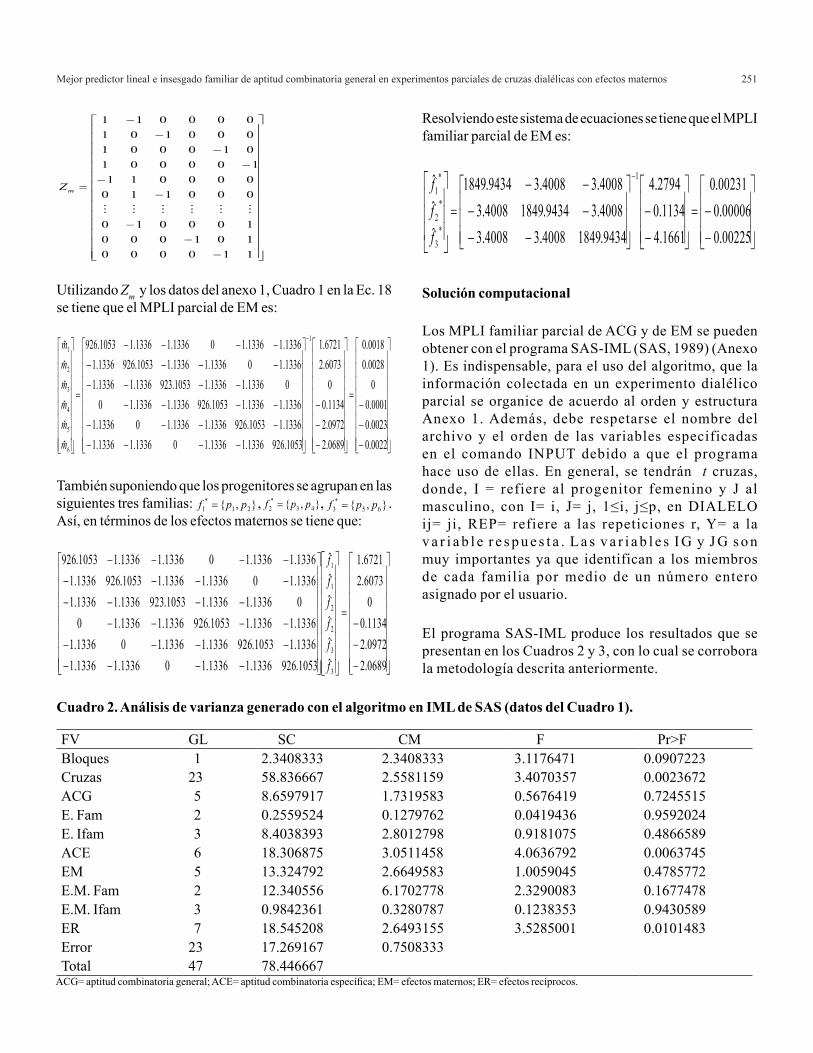
Mejor predictor lineal e insesgado familiar de aptitud combinatoria general en experimentos parciales de cruzas dialélicas con efectos maternos 251
Utilizando Zm y los datos del anexo 1, Cuadro 1 en la Ec. 18 se tiene que el MPLI parcial de EM es:
También suponiendo que los progenitores se agrupan en las siguientes tres familias: , , . Así, en términos de los efectos maternos se tiene que:
Resolviendo este sistema de ecuaciones se tiene que el MPLI familiar parcial de EM es:
Solución computacional
Los MPLI familiar parcial de ACG y de EM se pueden obtener con el programa SAS-IML (SAS, 1989) (Anexo 1). Es indispensable, para el uso del algoritmo, que la información colectada en un experimento dialélico parcial se organice de acuerdo al orden y estructura Anexo 1. Además, debe respetarse el nombre delarchivo y el orden de las variables especificadas en el comando INPUT debido a que el programa hace uso de ellas. En general, se tendrán t cruzas, donde, I = refiere al progenitor femenino y J almasculino, con I= i, J= j, 1≤i, j≤p, en DIALELOij= ji, REP= refiere a las repeticiones r, Y= a lav a r i a b l e r e s p u e s t a . L a s v a r i a b l e s I G y J G s o nmuy importantes ya que identifican a los miembrosde cada familia por medio de un número entero asignado por el usuario.
El programa SAS-IML produce los resultados que se presentan en los Cuadros 2 y 3, con lo cual se corrobora la metodología descrita anteriormente.
−−
−
−−
−−
−−
=
110000101000100010
000110000011100001
010001000101000011
mZ
−−−
=
−−−
−−−−−−−−−−−−
−−−−−−−−−−−−
=
−
0022.00023.00001.00
0028.00018.0
0689.20972.21134.00
6073.26721.1
1053.9261336.11336.101336.11336.11336.11053.9261336.11336.101336.11336.11336.11053.9261336.11336.1001336.11336.11053.9231336.11336.11336.101336.11336.11053.9261336.11336.11336.101336.11336.11053.926
ˆˆˆˆˆˆ
1
6
5
4
3
2
1
mmmmmm
},{ 21*
1 ppf = },{ 65*
3 ppf =},{ 43*
2 ppf =
−−−
=
−−−−−−−−−−−−
−−−−−−−−−−−−
0689.20972.21134.00
6073.26721.1
ˆˆˆˆˆˆ
1053.9261336.11336.101336.11336.11336.11053.9261336.11336.101336.11336.11336.11053.9261336.11336.1001336.11336.11053.9231336.11336.11336.101336.11336.11053.9261336.11336.11336.101336.11336.11053.926
3
3
2
2
1
1
ffffff
FV GL SC CM F Pr>FBloques 1 2.3408333 2.3408333 3.1176471 0.0907223Cruzas 23 58.836667 2.5581159 3.4070357 0.0023672ACG 5 8.6597917 1.7319583 0.5676419 0.7245515E. Fam 2 0.2559524 0.1279762 0.0419436 0.9592024E. Ifam 3 8.4038393 2.8012798 0.9181075 0.4866589ACE 6 18.306875 3.0511458 4.0636792 0.0063745EM 5 13.324792 2.6649583 1.0059045 0.4785772E.M. Fam 2 12.340556 6.1702778 2.3290083 0.1677478E.M. Ifam 3 0.9842361 0.3280787 0.1238353 0.9430589ER 7 18.545208 2.6493155 3.5285001 0.0101483Error 23 17.269167 0.7508333Total 47 78.446667
−−=
−−
−−−−−−
=
−
00225.000006.0
00231.0
1661.41134.0
2794.4
9434.18494008.34008.34008.39434.18494008.34008.34008.39434.1849
ˆˆˆ 1
*3
*2
*1
fff
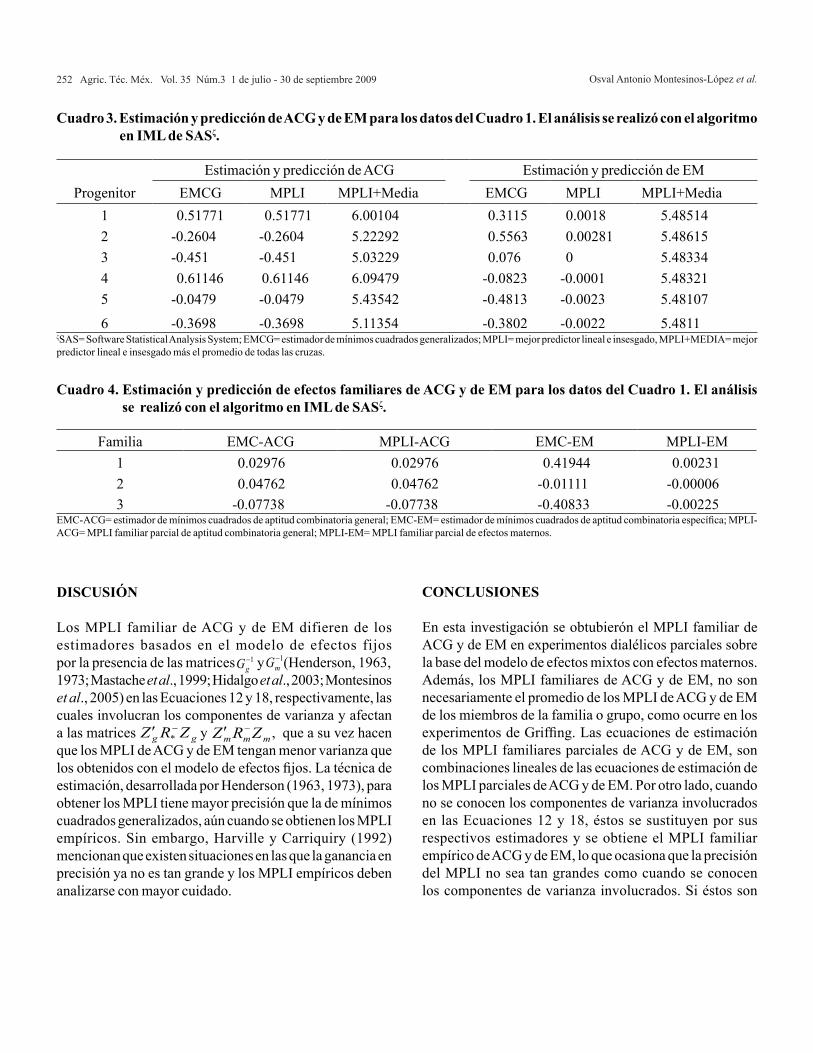
Cuadro 2. Análisis de varianza generado con el algoritmo en IML de SAS (datos del Cuadro 1).
ACG= aptitud combinatoria general; ACE= aptitud combinatoria específica; EM= efectos maternos; ER= efectos recíprocos.
Osval Antonio Montesinos-López et al.252 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
DISCUSIÓN
Los MPLI familiar de ACG y de EM difieren de los estimadores basados en el modelo de efectos fijos por la presencia de las matrices y (Henderson, 1963, 1973; Mastache et al., 1999; Hidalgo et al., 2003; Montesinos et al., 2005) en las Ecuaciones 12 y 18, respectivamente, las cuales involucran los componentes de varianza y afectan a las matrices y , que a su vez hacen que los MPLI de ACG y de EM tengan menor varianza que los obtenidos con el modelo de efectos fijos. La técnica de estimación, desarrollada por Henderson (1963, 1973), para obtener los MPLI tiene mayor precisión que la de mínimos cuadrados generalizados, aún cuando se obtienen los MPLI empíricos. Sin embargo, Harville y Carriquiry (1992) mencionan que existen situaciones en las que la ganancia en precisión ya no es tan grande y los MPLI empíricos deben analizarse con mayor cuidado.
CONCLUSIONES
En esta investigación se obtubierón el MPLI familiar de ACG y de EM en experimentos dialélicos parciales sobre la base del modelo de efectos mixtos con efectos maternos. Además, los MPLI familiares de ACG y de EM, no son necesariamente el promedio de los MPLI de ACG y de EM de los miembros de la familia o grupo, como ocurre en los experimentos de Griffing. Las ecuaciones de estimación de los MPLI familiares parciales de ACG y de EM, son combinaciones lineales de las ecuaciones de estimación de los MPLI parciales de ACG y de EM. Por otro lado, cuando no se conocen los componentes de varianza involucrados en las Ecuaciones 12 y 18, éstos se sustituyen por sus respectivos estimadores y se obtiene el MPLI familiar empírico de ACG y de EM, lo que ocasiona que la precisión del MPLI no sea tan grandes como cuando se conocen los componentes de varianza involucrados. Si éstos son
Estimación y predicción de ACG Estimación y predicción de EMProgenitor EMCG MPLI MPLI+Media EMCG MPLI MPLI+Media
1 0.51771 0.51771 6.00104 0.3115 0.0018 5.485142 -0.2604 -0.2604 5.22292 0.5563 0.00281 5.486153 -0.451 -0.451 5.03229 0.076 0 5.483344 0.61146 0.61146 6.09479 -0.0823 -0.0001 5.483215 -0.0479 -0.0479 5.43542 -0.4813 -0.0023 5.48107
6 -0.3698 -0.3698 5.11354 -0.3802 -0.0022 5.4811
Familia EMC-ACG MPLI-ACG EMC-EM MPLI-EM1 0.02976 0.02976 0.41944 0.002312 0.04762 0.04762 -0.01111 -0.000063 -0.07738 -0.07738 -0.40833 -0.00225
Cuadro 3. Estimación y predicción de ACG y de EM para los datos del Cuadro 1. El análisis se realizó con el algoritmo en IML de SASζ.
ζSAS= Software Statistical Analysis System; EMCG= estimador de mínimos cuadrados generalizados; MPLI= mejor predictor lineal e insesgado, MPLI+MEDIA= mejor predictor lineal e insesgado más el promedio de todas las cruzas.
Cuadro 4. Estimación y predicción de efectos familiares de ACG y de EM para los datos del Cuadro 1. El análisis se realizó con el algoritmo en IML de SASζ.
EMC-ACG= estimador de mínimos cuadrados de aptitud combinatoria general; EMC-EM= estimador de mínimos cuadrados de aptitud combinatoria específica; MPLI-ACG= MPLI familiar parcial de aptitud combinatoria general; MPLI-EM= MPLI familiar parcial de efectos maternos.
1−gG 1−
mG
gg ZRZ −′ * mmm ZRZ −′

Mejor predictor lineal e insesgado familiar de aptitud combinatoria general en experimentos parciales de cruzas dialélicas con efectos maternos 253
mejores que los obtenidos bajo el modelo de efectos fijos. También es necesario mencionar que los diseños uno y tres de Griffing ocurren como un caso particular de la clase de experimentos dialélicos parciales que consideran efectos maternos. Finalmente, el algoritmo computacional realizado proporciona de una forma rápida y compacta un análisis completo de estimación y predicción de efectos familiares en experimentos parciales de cruzas dialélicas, en donde se involucra la estimación de efectos maternos que considera aleatorios a los efectos de aptitud combinatoria general y específica, efectos maternos y recíprocos, con lo cual se logra un panorama completo de estimación y predicción.
LITERATURA CITADA
Cervantes, S., T; Castillo, G. F. y Domínguez, C. E. 1999. Cambios en la media y en la varianza de caracteres cuantitativos de cebada por irradiación recurrente. Agrociencia 33:251-299.
Fyfe, J. L. and Gilbert, N. 1963. Partial diallel cross. Biometrics 19:287-306.
Griffing, B. 1956a. A generalized treatment of the use of dialell crosses in quantitative inheritance. Heredity 10:31-50.
Griffing, B. 1956b. Concept of general and specific combining ability in relation to dialell crossing systems. Austr. J. Biol. Sci. 9:463-491.
Harville, D. A. 1976. Extension of the Gauss-Markov theorem to include the estimation of random effects. Ann. Statist 4:384-395.
Harville, D. A and Carriquiry, A. L. 1992. Classical and Bayesian prediction as applied to an unbalanced mixed linear model. Biometrics 48:987-1003.
Henderson, C. R. 1963. Selection index and expected genetic advance. pp: 141-163. In: Hanson, W. D. Robinson, H. F. (eds). Statistical genetics and plant breeding. Nat. Acad. Sci., Nat. Res. Council, Publication 982, Washington, D. C.
Henderson, C. R. 1973. Sire evaluation and genetics trends. pp:10-41. In: Proc. Anim. Breed. Genet. Symp. In honor of Dr. Jay L. Lush. Champaign, ILL.
Hidalgo, C. J. V; Martínez, G. A; Mastache, L. A. A y Rendón, S. G. 2003. Mejor predictor lineal e insesgado para aptitud combinatoria general y análisis combinado de los diseños dos y cuatro de Griffing. Rev. Fitotec. Mex. 26 (4):319-329.
Kempthore, O. R. and Curnow N. 1961. The partial diallel cross. Biometrics 17:229-250.
Martínez, G. A. 1983. Diseño y análisis de los experimentos de cruzas dialélicas. Centro de Estadística y Cálculo, Colegio de Postgraduados. Chapingo, Estado de México. p. 345.
Martínez, G. A. 1988. Diseños experimentales. 1a. Edición. Trillas. México.
Mastache, L. A. A; Martínez, G. A y Castillo, M. A. 1999. Los mejores predictores lineales e insesgados (MPLI) en los diseños uno y tres de Griffing. Agrociencia 33:349-359.
Montesinos, L. O. A; Martínez, G. A; Mastache, L. A. A y Rendón, S. G. 2005. Mejor predictor lineal e insesgado para aptitud combinatoria específica de los diseños dos y cuatro de Griffing. Rev. Fitotec. Mex. 28 (4):369-376.
Robinson, G. K. 1991. That BLUP is a good thing: the estimation of random effects. Statistical Science 6(1):15-51.
Ron, P. J; Ramírez, D. J. L; Valdivia, B. R. y Lozano, B. M. 1999. Comparación de tipos de varidades de maíz desarrolladas por el INIFAP en la región Centro-Occidente de México. Agrociencia 33:267-273.
Rojas, B. A. 1973. Design and analysis of diallel crosses. Department of experimental Statistics, New Mexico State University, USA.
Statistical Analysis System Institute. (SAS), Institute Inc. 1989. SAS/IML Software: Usage and Reference, Version 6. Cary, N. C.
Anexo 1. Algoritmo computacional en IML de SAS.
OPTIONS PS=60 PAGENO=1 NODATE;DATA MASTACHE;INPUT CRUZA I J DIALELO REP Y1 Y2 Y3 Y4;IF I=1 THEN IG=1; IF I=2 THEN IG=1; IF I=3 THEN IG=2; IF I=4 THEN IG=2; IF I=5 THEN IG=2; IF I=6 THEN IG=2; IF J=1 THEN JG=1; IF J=2 THEN JG=1; IF J=3 THEN JG=2; IF J=4 THEN JG=2; IF J=5 THEN JG=2; IF J=6 THEN JG=2; ANGEL=2; CARDS;

Osval Antonio Montesinos-López et al.254 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Title " Análisis familial de experimentos simétricos balanceados de cruzas dialélicas ";PROC IML;USE MASTACHE; READ ALL INTO MATRIZ; CRUZA= MATRIZ[,1];A=MATRIZ[,2]; B=MATRIZ[,3]; REP=MATRIZ[,5]; N=NROW(MATRIZ); UNO = J(N,1,1);CERO=J(N,1,0); MDIS=DESIGN(CRUZA); M00=UNO*INV(UNO`*UNO)*UNO`; X= UNO||MDIS;XX=X`*X;XXIG=GINV(XX);M=X*XXIG*X`; BLOQ= DESIGN(REP); W=X||BLOQ;WW=W`*W;WINV=GINV(WW);WWW=W*WINV*W`;IDEN=I(N); T= NCOL(MDIS);R=MAX(REP);P=MAX(B); NC=NCOL(MATRIZ);J=UNO;ANGEL=MATRIZ[,NC]; IF ANY (A=B) THEN Q=1;ELSE Q=0; IF ANY (ANGEL=1) THEN PRINT "DISEÑO COMPLETAMENTE AL AZAR";ELSE PRINT "DISEÑO DE BLOQUES AL AZAR";
Title " La matriz diseño Zp ";A0=J(N,P,.);B0=J(N,P,.);DO AAA=1 TO P;DO CCC=1 TO N;IF A[CCC,1]=AAA THEN A0[CCC,AAA]=1;ELSE A0[CCC,AAA]=0;IF B[CCC,1]=AAA THEN B0[CCC,AAA]=1;ELSE B0[CCC,AAA]=0;END; END;AB= A0+B0;Zp= AB; ZpZp= Zp`*Zp;DIAGO= DIAG(ZpZp);Zii=DIAGO*J(P,1,1); X0= UNO||AB;X0X0=X0`*X0;X0IG=GINV(X0X0); M0=X0*X0IG*X0`;
Title " Matrices diseño para familias (ACG) ";IG= MATRIZ[,NC-2];JG=MATRIZ[,NC-1];FAMILIAS= MAX(JG);A000=J(N,FAMILIAS,.);B000=J(N,FAMILIAS,.);DO AAA=1 TO FAMILIAS;DO CCC=1 TO N;IF IG[CCC,1]=AAA THEN A000[CCC,AAA]=1;ELSE A000[CCC,AAA]=0;IF JG[CCC,1]=AAA THEN B000[CCC,AAA]=1;ELSE B000[CCC,AAA]=0;END;END;FAMGDIS = A000+B000;MIG=FAMGDIS*GINV(FAMGDIS`*FAMGDIS)*FAMGDIS`;COEFIC = UNO`*FAMGDIS;COEFIC=DIAG(COEFIC); FAMCOEF= TRACE(COEFIC);FAMCOEF=COEFIC*P/FAMCOEF;
Title " Diseños con eval. de C. recíprocas ";IF ANY(B<A) THEN DO;IF ANY(Zii-Zii[1,1]) >0.1 THEN PRINT "DIALÉLICO PARCIAL ASIMÉTRICO-CON C. RECÍPROCAS";ELSE IF 2*R*(2*Q+P-1)-Zii[1,1]>0.1 THEN PRINT "DIALÉLICO PARCIAL SIMÉTRICO-CON C. RECÍPROCAS";ELSE IF Q=1 THEN PRINT "DISEÑO 1 DE GRIFFING";ELSE PRINT "DISEÑO 3 DE GRIFFING";DIAL= MATRIZ[,4];D=DESIGN(DIAL);DD=D`*D;D0=D*(GINV(DD))*D`;Zm=A0-B0;ZmZm=Zm`*Zm; MAB=Zm*GINV(ZmZm)*Zm` ;S=D*D` ;E=I (N) ;Sm=2*(MDIS*MDIS` ) -S ;TRA1= TRACE((D0-M0)*E);TRA2=TRACE((D0-M0)*S);TRA3=TRACE((M0-M00)*E);TRA4=TRACE((M0-M00)*S);TRA5= TRACE((IDEN-00)*Zp*Zp`);TRA6=TRACE((M-D0-MAB)*Sm);TRA7=TRACE(MAB*Sm);TRA8=TRACE(Zm*Zm`);PRINT N T R P;
Title " Matrices diseño para familias (EM) ";FAMMDIS = A000-B000; MJG=FAMMDIS*GINV(FAMMDIS`*FAMMDIS)*FAMMDIS`;CONTfam= J(P,FAMILIAS,.); FKF=R*T/P;DO AAA= 1 TO FAMILIAS;FKKF1=0;DO CCC=1 TO N BY FKF; FKKF1=FKKF1+1;IF IG[CCC,1]= AAA THEN CONTfam[FKKF1,AAA]=1/FAMCOEF[AAA,AAA];ELSE CONTfam[FKKF1,AAA]=0;END; END;
Title " Análisis de varianza ";GLt= ROUND(TRACE(GINV((M-M00)*(M-M00))));IF ANY(ANGEL=1) THEN DO;GLr=.;GLe=N-1-GLt;END; IF ANY(ANGEL=2) THEN DO;GLr=ROUND(TRACE(GINV((WWW-M)*(WWW-M))));GLe=N-1-GLr-GLt;END;GLacg= ROUND(TRACE(GINV((M0-M00)*(M0-M00))));GLace= ROUND(TRACE((GINV(D0-M0)*(D0-M0))));GLem= OUND(TRACE(GINV((MAB)*MAB)));GLer=GLt-GLacg-GLace-GLem; FV = J(12, 5, .);UN=J(P,1,1);PROG=J(P, 4, .);PPP=J(P,1,.);PRG=PROG;FAMILIA=J(FAMILIAS, 4, .);FAMIL=J(FAMILIAS,1,.);DO LLL= 1 TO P BY 1;PPP[LLL,1]=LLL;END;DO LLLL = 1 TO FAMILIAS BY 1;FAMIL[LLLL,1]=LLLL;END;DO F= 6 TO (NC-3) BY 1;VARIABLE= F-5;Y= MATRIZ[,F];FC= Y`*M00*Y;MEDIA=UNO`*Y/N;SCTOT=Y`*Y-FC;IF ANY(ANGEL=1) THEN DO;SCE=Y`*(IDEN-M)*Y;CME=SCE/GLe;SCB=.;CMB=.;FBLOQ=.;END;IF ANY(ANGEL=2) THEN DO;SCE=Y`*(IDEN-WWW)*Y;CME=SCE/GLe;SCB=Y`*(WWW-M)*Y;CMB=SCB/GLr;FBLOQ=CMB/CME;END;SCCRUZA = (Y`*M*Y)-C;CMCRUZA =SCCRUZA/GLt;FCRUZA=CMCRUZA/CME;SCACE =

Mejor predictor lineal e insesgado familiar de aptitud combinatoria general en experimentos parciales de cruzas dialélicas con efectos maternos 255
Y`*(D0-M0)*Y;CMACE=SCACE/GLace;FACE=CMACE/CME; SCACG= (Y`*M0*Y)-FC;CMACG=SCACG/GLacg;FACG=CMACG/CMACE;SCEM= Y`*MAB*Y;SCER=SCCRUZA-(SCACG+SCACE+SCEM);CMEM=SCEM/GLem;CMER=SCER/GLer;FEM=CMEM/CMER;FER=CMER/CME; V=(CME**.5)*100/MEDIA;GLFAMg = ROUND(TRACE(GINV((MIG-M00)*(MIG-M00))));GLIFAMg=GLacg-GLFAMg;GLFAMm = ROUND(TRACE(GINV(MJG*MJG)));GLIFAMm=GLem-GLFAMm;SCFAMg = Y`*MIG*Y-FC;CMFAMg =SCFAMg/GLFAMg; FFAMg =CMFAMg/CMACE;SCIFAMg = SCACG-SCFAMg; CMIFAMg=SCIFAMg/GLIFAMg;FIFAMg= MIFAMg/CMACE;SCFAMm = Y`*MJG*Y; CMFAMm =SCFAMm/GLFAMm;FFAMm=CMFAMm/CMER;SCIFAMm= CEM-SCFAMm; CMIFAMm=SCIFAMm/GLIFAMm;FIFAMm=CMIFAMm/CMER;PROBFAMg= 1-PROBF(FFAMg,GLFAMg,GLace);PROBIFMg=1-PROBF(FIFAMg,GLIFAMg,GLace);PROBFAMm= 1-PROBF(FFAMm,GLFAMm,GLer);PROBIFMm=1-PROBF(FIFAMm,GLIFAMm,GLer);PROBb = 1-PROBF(Fbloq,GLr,GLe);PROBt=1-PROBF(Fcruza,GLt,GLE);P R O B a c g = 1 - P R O B F ( F a c g , G L a c g , G L a c e ) ; P R O B a c e = 1 - P R O B F ( F a c e , G L a c e , G L e ) ; P R O B e m = 1-PROBF(Fem,GLem,GLer); PROBer =1-PROBF(Fer, GLer, GLe);FV[1,1] = GLr;FV[2,1]=GLt;FV[3,1]=GLacg;FV[4,1] =GLFAMg;FV[5,1] =GLIFAMg;FV[6,1]=GLace;FV[7,1]=GLem;FV[8,1]=GLFAMm;FV[9,1]=GLIFAMm;FV[10,1]=GLer;FV [11,1]=GLe;FV[12,1]=N-1;FV[1,2] = SCB;FV[2,2]=SCCRUZA;FV[3,2]=SCACG;FV[4,2]=SCFAMg;FV[5,2]=SCIFAMg;FV[6,2]=SCACE;FV[7,2]=SCEM;FV[8,2]=SCFAMm;FV[9,2]=SCIFAMm;FV[10,2]=SCER;FV[11,2]=SCE;FV[12,2]=SCTOT;FV[1,3]=CMB;FV[2,3]=CMCRUZA;FV[3,3]=CMACG;FV[4,3]=CMFAMg;FV[5,3]=CMIFAMg;FV[6,3]CMACE;FV[7,3] =CMEM;FV[8,3]=CMFAMm;FV[9,3]=CMIFAMm;FV[10,3]=CMER;FV[11,3]=CME;FV[1,4]= FBLOQ;FV[2,4]=FCRUZA; FV[3,4]=FACG;FV[4,4]=FFAMg;FV[5,4]=FIFAMg;FV[6,4]=FACE;FV[7,4]=FEM;FV[8,4]=FFAMm;FV[9,4]=FIFAMm;FV[10,4]=FER;FV[1,5] = PROBb;FV[2,5]=PROBt;FV[3,5]=PROBacg;FV[4,5]=PROBFAMg;FV[5,5]=PROBIFMg;F V [ 6 , 5 ] = P R O B A C E ; F V [ 7 , 5 ] = P R O B E M ; F V [ 8 , 5 ] = P R O B FA M m ; F V [ 9 , 5 ] = P R O B I F M m ; F V [10,5]=PROBER;CCC={"GL""SC" "CM" "F" "Pr > F"};IF ANY(ANGEL=1) THEN DDD={" . " "CRUZAS" " ACG" " E.FAM." " E.IFAM." " ACE" " EM"" E.M.FAM." " E.M.IFAM." " ER" "ERROR" "TOTAL"};ELSE DDD={"BLOQUES" "CRUZAS" " ACG" " E.FAM." " E.IFAM." " ACE" " EM"" E.M.FAM." "E.M.IFAM." " ER" "ERROR" "TOTAL"};
Title " Estimación de las componentes de varianza ";VARe = CME;VARs=(GLace*CMace-CME*TRA1)/(TRA2);VARg = (GLacg*CMacg-VARs*TRA4-CME*TRA3) /(TRA5);VARr = GLer*(CMer-CME)/TRA6;VARm=(GLem*(CMem-CME)-VARr*TRA7)/TRA8; IF VARs > 0 THEN VARs=VARs;ELSE VARs=0;IF VARg>0 THEN VARg=VARg;ELSE VARg=0;IF VARr > 0 THEN VARr=VARr;ELSE VARr=0;IF VARm>0 THEN VARm=VARm;ELSE VARm=0;RR= (VARs/VARe)*S + E;GRR=GINV(RR);RI=(VARr/VARe)*Sm + E;GRI=GINV(RI);
Title " EMC, EMCG y el MPLI de ACG y de EM ";MU= INV(J`*GRR*J)*J`*GRR*Y;MC= GINV(Zp`*Zp)*Zp`*(Y-MEDIA*J);EMCG=GINV(Zp`*GRR*Zp)*Zp`*GRR*(Y-MU*J);IF VARg > 0 THEN INVGp=(VARe/VARg)*I(P);ELSE INVGp=0*I(P);EMCGMED=EMCG+MU*UN;MPLI= GINV(Zp`*GRR*Zp+INVGp)*Zp`*GRR*(Y-MU*J);MPLIMED=MPLI+MU*UN;PROG[,1]= EMCG;PROG[,2]=MPLI; PROG[,3]=EMCGMED;PROG[,4]=MPLIMED;EEE = {"EMCG" "MPLI" "EMCG+MU""MPLI+MU"};FFF=CHAR(PPP,3,0);IF VARm > 0 THEN INVGm=(VARe/VARm)*I(P);ELSE INVGm=0*I(P);EMCm=GINV(Zm`*Zm)*Zm`*Y;EMCGm =GINV(Zm`*GRI*Zm)*Zm`*GRI*Y; MPLIm = GINV(Zm`*GRI*Zm+INVGm)*Zm`*GRI*Y;PRG[,1]= EMCGm; PRG[,2]=MPLIm;PRG[,3]=EMCGm+MU*UN;PRG[,4]=MPLIm+MU*UN;
Title " Los EMCG y los MPLI para familias ";IF VARg > 0 THEN INVGpp=(VARe/VARg)*FAMCOEF;ELSE INVGpp=0*FAMCOEF;PFAMg =GINV(FAMGDIS`*GRR*FAMGDIS+INVGpp)*FAMGDIS`*GRR*(Y-MU*J);EFAMg=GINV(FAMGDIS`*GRR*FAMGDIS) *FAMGDIS`*GRR*(Y-MU*J);IF VARm > 0 THEN INVGmm=(VARe/VARm)*FAMCOEF;ELSE INVGmm=0*FAMCOEF;

Osval Antonio Montesinos-López et al.256 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
PFAMm=GINV(FAMMDIS`*GRI*FAMMDIS+INVGmm)*FAMMDIS`*GRI*Y;EFAMm=GINV(FAMMDIS`*GRI*FAMMDIS) *FAMMDIS`*GRI*Y;FAMILIA[,1]= EFAMg;FAMILIA[,2]=PFAMg;FAMILIA[,3]=EFAMm;FAMILIA[,4]=PFAMm;EEEE={"EMCGg" "MPLIg" "EMCGm" "MPLIm"};FFFF=CHAR(FAMIL,3,0);
Title " La matriz de coeficientes: C de ACG y EM ";CC1= (UNO`*GRR*UNO)||(UNO`*GRR*Zp);CC2= (UNO`*GRR*Zp)`||((Zp`*GRR*Zp)+INVGp);CC3=CC1`||CC2`;CCCC = GINV(CC3);GAMA=2:P+1;CCCC=CCCC[GAMA, GAMA];CC4= Zm`*GRI*Zm+INVGm;CCCCC=GINV(CC4);
Title " Impresión de resultados ";PRINT VARIABLE;PRINT "CUADRO 1. ANÁLISIS DE VARIANZA.";PRINT FV[ROWNAME=DDD COLNAME=CCC];PRINT MEDIA[FORMAT= 12.5] CV[FORMAT=12.5];PRINT,;PRINT " ESTIMACIÓN DE LAS COMPONENTES DE VARIANZA";PRINT VARe[FORMAT=12.5] VARr[FORMAT=12.5] VARm[FORMAT=12.5] VARs[FORMAT=12.5] VARg[FORMAT=12.5];PRINT /;PRINT MU[FORMAT= 12.5];PRINT,;PRINT "CUADRO 2. ESTIMACIÓN Y PREDICCIÓN DE ACG.";PRINT PROG[ROWNAME=FFF COLNAME=EEE FORMAT=12.5];PRINT ,; PRINT "CUADRO 3. ESTIMACIÓN Y PREDICCIÓN DE EM.";PRINT PRG[ROWNAME=FFF COLNAME=EEE FORMAT=12.5];PRINT ,; PRINT "CUADRO 4. ESTIMACIÓN Y PREDICCIÓN FAMILIAL.";PRINT FAMILIA[ROWNAME=FFFF COLNAME=EEEE FORMAT=12.5];PRINT /;PRINT "CUADRO 5. LA MATRIZ DE COEFICIENTES C22 DE ACG.";PRINT CCCC[FORMAT=7.3];PRINT ,;PRINT "CUADRO 6. LA MATRIZ DE COEFICIENTES C DE EM.";PRINT CCCCC[FORMAT=7.3];PRINT /; END;END; QUIT;

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 257-266
CALIDAD DE SEMILLA EN COLECTAS DE CHILE DE AGUA (Capsicum annuum L.) DE LOS VALLES CENTRALES DE OAXACA, MÉXICO*
SEED QUALITY IN CHILE DE AGUA (Capsicum annuum L.) LANDRACES FROM VALLES CENTRALES, OAXACA, MEXICO
Erik Pablo Carrillo1, José Apolinar Mejía Contreras1§, Aquiles Carballo Carballo1, Gabino García de los Santos1, Víctor Heber Aguilar Rincón2 y Tarsicio Corona Torres2
1Programa de Recursos Genéticos y Productividad, Producción de Semillas y 2Genética, Colegio de Postgraduados. Km. 36.5 carretera México-Texcoco. C. P. 56230. Montecillo, Estado de México. ([email protected]), ([email protected]), ([email protected]), ([email protected]). Tel. y Fax: 01 595 95 2 02 62. §Autor para correspondencia: [email protected].
* Recibido: Mayo, 2007
Aceptado: Enero, 2009
RESUMEN
El cultivo del chile de agua es económicamente importante en los Valles Centrales de Oaxaca, debido a su consumo generalizado en esta región. La semilla utilizada para la siembra de este cultivo se obtiene de forma artesanal, debido a esto se desconocen los estándares de calidad de semilla tanto físicos como fisiológicos. El objetivo fue determinar la calidad física y fisiológica de la semilla de 14 colectas de chile de agua. Se establecieron tres ensayos bajo condiciones de campo, uno en el ciclo primavera-verano y otro en otoño-invierno de 2004 en San Sebastián Abasolo, Oaxaca, y otro bajo condiciones de invernadero en Montecillo, Estado de México. El diseño experimental fue bloques completos al azar con cuatro repeticiones. Se cosecharon cinco frutos maduros de cada una de cinco plantas por parcela. Las variables evaluadas fueron: peso de mil semillas, peso volumétrico, germinación estándar y con envejecimiento acelerado, longitud de plántula y raíz, peso seco de plántula y raíz. En un análisis de componentes principales (CP) los dos primeros explicaron 98.36% de la variabilidad observada. Las variables importantes en el CP1 fueron: peso de mil semillas, peso volumétrico y longitud de plántula; y para el CP2 germinación con y sin envejecimiento acelerado. Un análisis de conglomerados formó cuatro grupos; las
colectas de San Sebastián Abasolo y Santiaguito mostraron alta calidad física y fisiológica de semilla, reflejada en un mayor porcentaje de germinación, respuesta favorable al envejecimiento acelerado, mayor peso de mil semillas y peso volumétrico, así como mayor acumulación de materia seca en los tres ensayos.
Palabras clave: análisis discriminante, conglomerados, chile criollo, semilla artesanal.
ABSTRACT
The crop of “chile de agua” in the Central Valley of Oaxaca is economically important due to its generalized consumption in this region. The seed to establish this crop is obtained through the traditional method, for this reason the physical and physiological quality standards of those seeds is unknown. The aim of this research was to determine the physical and physiological quality in the seed of 14 landraces of “chile de agua” produced in three environments. The 14 landraces were sown at San Sebastián Abasolo, Tlacolula, Oaxaca during the 2004 spring-summer and fall-winter

Erik Pablo Carrillo et al.258 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
cycles under field condition and at Montecillo, Mexico under greenhouse conditions. The experimental design was a randomized complete block with four replications. From the three environments five ripened fruits from each of five plants per plot were taken for seed evaluation. The physical quality of the seed was evaluated through the weight of 1000 seeds and volumetric weight, whereas the physiological quality by means of standard germination and after accelerated aging, seedling and root length and dry weight. In a principal component analysis the first two components explained 98.36% of the total variation; the variables important in the first component were weight of 1 000 seeds, volumetric weight and seedling length, for the second component were germination with and without accelerated aging. A cluster analysis formed four groups of landraces; landraces from San Sebastián Abasolo and Santiaguito showed high physical and physiological seed quality reflected in a higher percentage of germination and favorable response to accelerated aging and greater volumetric and one thousand seeds weight, as well as greater accumulation of dry matter in the three environments.
Key words: artisanal seed, cluster, chili landraces, discriminant analysis.
INTRODUCCIÓN
En la producción de chile en México los requerimientos de semilla corresponden en su mayoría a variedades nativas, producidas por el propio agricultor (Montes y Martínez, 1992). En los Valles Centrales de Oaxaca el procedimiento para obtener la semilla de chile de agua es artesanal, pocos productores dejan en su parcela un área exclusiva para producción de semilla (López, 1989). Además de la diversidad genética existente entre y dentro de las poblaciones, se observa una pobre calidad genética de la semilla de los cultivares criollos, que afecta también la calidad fisiológica y física.
En la producción comercial de semillas la calidad está determinada por un conjunto de atributos, donde la calidad genética, física, sanitaria y fisiológica juegan un papel importante (Besnier, 1989; Copeland y McDonald, 1995). La calidad fisiológica implica la integridad de las estructuras y procesos fisiológicos, siendo los
principales indicadores: la viabilidad, germinación y vigor, que dependen del genotipo (Perry, 1972; Moreno et al., 1988). Entre los factores que pueden tener efecto en la calidad de la semilla están el grado de madurez y tiempo de maduración de la semilla después de la cosecha. Randle y Honma (1981) mencionan que en chile las semillas completan su madurez fisiológica en un período de reposo que varía de una a seis semanas después de que los frutos fueron cosechados, dependiendo del tipo de chile.
Los factores relacionados con la calidad física de la semilla son: contenido de humedad, peso por volumen y pureza. También se puede considerar el color, tamaño de semilla, peso de mil semillas y daño por hongos e insectos. Las semillas deben reunir ciertos estándares de calidad dependiendo de la especie para ser consideradas de buena calidad física (Moreno, 1996).
Puente y Bustamante (1991) observaron en chile habanero que al avanzar el estado de madurez del fruto hubo un incremento en el porcentaje de germinación y viabilidad de la semilla. Doijode (1991) estudió el efecto de la posición dentro del fruto (base, medio y punta) en la calidad de semilla de chile; este autor encontró que el porcentaje de germinación de las semillas de la base fue mayor en comparación con las de la parte media y de la punta del fruto. Por su parte, Cochran (1974) observó en pimiento morrón que porcentaje de germinación y emergencia de las semillas grandes fue mayor y produjeron plántulas más vigorosas, uniformes y con mayor cantidad de materia seca en comparación con las de semilla chica. Edwards y Sundstrom (1987) evaluaron el efecto de la madurez del fruto y del tiempo de cosecha y posmaduración en la germinación de chile tabasco. Estos autores obtuvieron porcentajes de germinación 81% al haber cosechado frutos rojos; además, el porcentaje de germinación se elevó 86% después de un período de posmaduración de 21 días a 25 ºC.
Los programas de mejoramiento genético prestan poca atención a la semilla y sus caracteres intrínsecos. No obstante, estos caracteres pueden facilitar la producción, mejorando la emergencia en campo y favoreciendo un mayor rendimiento (Carballo, 1992). Considerando la importancia y la gran diversidad existente de materiales criollos del chile de agua en los Valles Centrales de

Calidad de semilla en colectas de chile de agua (Capsicum annuum L.) de los Valles Centrales de Oaxaca, México 259
Oaxaca, de los cuales se desconocen los estándares de calidad de semilla, el objetivo de la presente investigación fue determinar la calidad física y fisiológica de la semilla en colectas de chile de agua producidas en diferentes ambientes.
MATERIALES Y MÉTODOS
En esta investigación se utilizaron 14 materiales nativos de chile de agua colectados en los valles Centrales del estado de Oaxaca (Cuadro 1).
Variables evaluadas:
Prueba de germinación. Para esta prueba se emplearon cuatro repeticiones de 25 semillas de acuerdo con las recomendaciones de la International Seed Testing Association (2004). Los caracteres evaluados fueron: porcentaje de germinación con base en las plántulas con raíz, hipocotilo y epicotilo bien desarrollados, sanas y sin malformaciones a los 15 días de iniciada la prueba.
Longitud de plántula. Se tomó una muestra de cinco plántulas normales al azar en cada repetición y se midió la longitud en cm del cuello de la raíz hasta el ápice de la hoja más larga.
Longitud de raíz. En cinco plántulas normales tomadas al azar, se midió la longitud de la raíz en cm desde el cuello de la raíz hasta el ápice de la misma.
Peso seco de la parte aérea. En cinco plántulas normales elegidas al azar se pesó la parte aérea, después de ser secadas a 75 ºC durante 72 h.
Registro de colecta
Localidad Municipio Altura (m) Latitud N
Longitud O
CHA1 Ejutla de Crespo (EJU) Ejutla 1 504 16º 33’ 96º 43’CHA2 San Jerónimo Tlacochahuaya (TCH) Tlacolula 1 600 17º 00’ 96º 35’CHA3 San Juan Guelavia I (SJGI) Tlacolula 1 600 16º 57’ 96º 32’CHA4 Santiago Apóstol (SAP) Ocotlán 1 477 16º 48’ 96º 43’CHA5 San Sebastián Abasolo (SSA) Tlacolula 1 575 16º 59 96º 35’CHA6 San Francisco Lachigolo (SFL) Tlacolula 1 607 17º 00’’ 97º 35’CHA7 San Pablo Huitzo (SPH) Etla 1 760 17º 16’ 96º 52’CHA8 Santiaguito (SAN) Etla 1 643 17º 12’ 96º 47’CHA9 Zimatlán de Álvarez (ZIM) Zimatlán 1 976 16º 51’ 96º 47’
CHA10 San Sebastián Teitipac (SST) Tlacolula 1 620 16º 57’ 96º 36’CHA11 Villa Sola de Vega (SV) Sola de vega 1 598 16º 30’ 97º 58’CHA12 San Juan Guelavia II (SJGII) Tlacolula 1 600 16º 57’ 96º 32’CHA13 Tlacolula de Matamoros I (TLAI) Tlacolula 1 584 16º 57’ 96º 28’CHA14 Tlacolula de Matamoros II (TLAII) Tlacolula 1 584 16º 57’ 96º 28’
Cuadro 1. Colectas de chile de agua y ubicación geográfica del sitio de colecta.
La semilla de las colectas se sembró en campo en condiciones de riego en San Sebastián Abasolo, Oaxaca, en los ciclos primavera-verano y otoño-invierno de 2004, y en condiciones de invernadero en el ciclo otoño-invierno de 2004 en Montecillo, Texcoco, Estado de México. Se utilizó un diseño experimental de bloques completos al azar con cuatro repeticiones. La parcela experimental consistió de un surco de 10 m de longitud en el que fueron transplantadas dos plantas por mata cada 30 cm. En el invernadero la parcela experimental consistió de 10 macetas con dos plantas por maceta (bolsas de polietileno negro de 30 x 30 cm), con una capacidad de 6 kg. Se utilizó tezontle rojo granulado como sustrato. Al llegar a la madurez (coloración rojo intenso) se cosecharon cinco frutos por planta y se secaron al sol antes de extraer la semilla. La semilla aprovechable se obtuvo con un separador neumático vertical de 4 cm de diámetro y 50 cm de largo. Las pruebas se realizaron en el laboratorio de análisis de semillas del programa de producción de semillas del Instituto de Recursos Genéticos y Productividad del Colegio de Postgraduados.
Erik Pablo Carrillo et al.260 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Peso seco de raíz. Peso en gramos de las raíces en cinco plántulas normales, después de ser secadas a 75 ºC durante 72 h.
Peso de mil semillas. Se pesaron 25 semillas en ocho repeticiones y se calculó la proporción correspondiente al peso de mil semillas.
Peso volumétrico. Para tomar el peso volumétrico se utilizó una probeta de 10 ml, se pesaron 2 g de c/u de las 14 muestras midiéndose posteriormente el volumen. Transformando, el valor obtenido a kg Hl.
Prueba de envejecimiento acelerado. Se utilizó la metodología propuesta por Delouche y Baskin (1996), la cual consistió en someter las semillas a una temperatura de 45 ºC ± 1 ºC y a una humedad relativa del 100%, por un período de 48 h. Se utilizaron cajas de plástico de 10 x 10 x 3.5 cm, a las que se les agregaron 40 ml de agua destilada, colocando una malla de alambre por encima del nivel del agua como medio de soporte de las semillas y así evitar el contacto de éstas con el agua. En cada caja se depositaron 100 semillas por colecta obtenidas en campo. Después del período de envejecimiento la semilla se evaluó en una prueba de germinación estándar de acuerdo con las recomendaciones de ISTA (2004), para lo cual se establecieron cuatro repeticiones de 25 semillas por tratamiento y se realizó un sólo conteo a los 15 días. Se realizó un análisis de varianza bajo un diseño de bloques completamente al azar y una comparación de medias
mediante la diferencia significativa honesta. Posteriormente se efectuó un análisis de componentes principales para determinar las variables que explican en mayor grado la variabilidad de las colectas (Pla, 1986). Se realizó un análisis de conglomerados jerárquico mediante el ligamiento promedio a partir de las distancias euclidianas. Con la matriz de distancias se construyó un dendrograma por el método de ligamiento medio (Van-Tongeres, 1997). Para verificar la certeza del agrupamiento se realizó un análisis de varianza con un criterio de clasificación (efecto de grupos) así como un análisis discriminante múltiple para comparar las diferencias entre los grupos (Mardia et al., 1979), se uso el software SAS V8.0 (1998).
RESULTADOS Y DISCUSIÓN
Para caracteres de semilla los resultados del análisis de varianza muestran que para todas las variables hubo diferencias altamente significativas (p≤0.01) entre localidades. De igual forma para colectas hubo diferencias altamente significativas en los caracteres evaluados; para la interacción ambiente x colecta se observaron diferencias altamente significativas en todas las variables excepto para PSR (Cuadro 2). Este último carácter mostró nula variación a través de ambientes. Estos resultados muestran diferencias en la calidad física y fisiológica de la semilla en las colectas, así como las diferencias entre localidades de producción.
Fuente de variación
GL GE EA PV PMS LPL LRA PSA PSR
Ambiente 2 10.49* 44.39** 267.68** 7.78** 9.12** 0.04** 3.41** 0.004**
Colecta 13 336.2** 119.4** 6.90** 1.81** 4.41** 0.02** 2.07** 0.003**
Amb x col 26 7.31** 6.78** 7.77** 0.22** 0.17** 0.0008** 0.10** 0.0001
Error 117 2.91 2.4 1.05 0.03 0.08 0.0003 0.05 0.0009
C.V (%) 2.70 3.76 3.26 3.24 4.26 4.34 4.88 5.41
R2 0.93 0.88 0.92 0.92 0.89 0.89 0.85 0.83*,** = significativo al 0.05 y 0.01 de probabilidad, respectivamente; GE= germinación; EA= envejecimiento acelerado; PV= peso volumétrico; PMS= peso de mil semillas; LPL= longitud de plántula; LRA= longitud de raíz; PSA= peso seco de parte aérea; PSR= peso seco de raíz.
Cuadro 2. Cuadrados medios para ocho caracteres de semilla determinados en 14 colectas de chile de agua (Capsicum annuum L.).

Calidad de semilla en colectas de chile de agua (Capsicum annuum L.) de los Valles Centrales de Oaxaca, México 261
En las variables germinación y envejecimiento acelerado, las colectas de San Sebastián Abasolocon 92.7 y 51.4% y Santiaguito con 89.6 y 49.3%,tuvieron mayor calidad fisiológica de semilla, reflejada
en un mayor porcentaje. Por otra parte, los porcentajesmas bajos se registraron en las colectas Zimatlán(70.0 y 33.2 %) y Sola de Vega (70.6 y 33.6)(Cuadro 3).
semilla reflejada en su peso. Las colectas de San Sebastián Abasolo, Santiaguito y San Pablo Huitzo mostraron mayor longitud de plántula y raíz con 7.76 y 5.73 cm; 7.52 y 5.29 cm; 7.74 y 5.24 cm, respectivamente; así como para el peso seco de estas variables, que para San Sebastián Abasolo fue de 0.5310 y 0.2213 mg, Santiaguito 0.5149 y 0.2034 mg, y San Pablo Huitzo 0.5105 y 0.2018 mg de materia seca producida para plántula y raíz (Cuadro 3).
Colecta GE EA PV PMS LPL LRA PSA PSR
CHA 1 74.9 de 36.3 d-f 30.17 f-h 5.15 f-h 6.25 ef 0.4257 d-f 4.483 c 0.1696 c
CHA 2 77.3 d 39.1 b-d 31.10 d-g 5.31 d-g 6.16 ef 0.4201 ef 4.525 c 0.1726 c
CHA 3 72.9 e-g 35.3 ef 29.91 gh 5.11 gh 6.27 ef 0.4282 d-f 4.583 c 0.1744 c
CHA 4 75.9 cde 37.7 c-e 29.53 hi 5.04 hi 6.35 de 0.4320d de 4.508 c 0.1696 c
CHA 5 92.7 a 51.4 a 36.57 a 6.24 a 7.76 a 0.5310 a 5.733 a 0.2213 a
CHA 6 81.3 c 40.4 bc 32.40 cd 5.53 cd 7.30 b 0.4975 b 5.183 b 0.1979 b
CHA 7 83.5 c 41.6 b 33.40 c 5.70 c 7.47 ab 0.5105 ab 5.241 b 0.2018 b
CHA 8 89.6 b 49.3 a 34.90 b 5.96 b 7.52 ab 0.5149 ab 5.291 b 0.2034 b
CHA 9 70.0 g 33.2 f 28.37 i 4.84 i 5.90 f 0.4006 f 4.358 c 0.1651 c
CHA 10 74.1 d-f 36.3 d-f 30.46 e-h 5.20 e-h 6.34 de 0.4311 de 4.575 c 0.1777 c
CHA 11 70.6 fg 33.6 f 29.08 hi 4.96 hi 6.08 ef 0.4115 ef 4.400 c 0.1666 c
CHA 12 74.4 de 36.6 d-f 31.35 d-f 5.35 d-f 6.43 c-e 0.4377 c-e 4.641 c 0.1777 c
CHA 13 83.9 c 41.8 b 31.68 de 5.41 de 6.67 cd 0.4532 cd 4.583 c 0.1755 c
CHA 14 84.21 c 42.1 b 32.03 cd 5.47 cd 6.78 c 0.4604 c 4.633 c 0.1775 c
DSH 0.05 2.39 2.05 1.435 0.244 0.398 0.0276 0.3256 0.0138
Cuadro 3. Valor medio para ocho caracteres de calidad de la semilla en 14 colectas de chile de agua producidas en tres ambientes.
GE= germinación; EA= envejecimiento acelerado; PV= peso volumétrico; PMS=peso de mil semillas; LPL= longitud de plántula; LRA= longitud de raíz; PSA= peso seco parte aérea; PSR= peso seco de raíz. Medias con la misma letra son estadísticamente iguales (Tukey 0.05).
Para los caracteres de calidad física de la semilla como peso de mil semillas y peso volumétrico, las colectas San Sebastián Abasolo con 36.57 kg hL y 6.24 g; Santiaguito con 34.90 kg hL y 5.96 g; y San Pablo Huitzo con 33.40 kg hL y 5.70 g fueron las de mayor peso. En contraste, las de Zimatlán y Sola de Vega presentaron 28.37 kg hL y 4.84 g; 29.08 kg hL y 4.96 g. Hubo diferencias significativas entre colectas para la calidad física de la

Erik Pablo Carrillo et al.262 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
El peso de la semilla estuvo correlacionado con el tamaño de plántula y producción de materia seca, ya que las colectas de San Sebastián Abasolo, Santiaguito y San Pablo Huitzo que presentaron mayor peso de semilla también produjeron plántulas de mayor tamaño y acumularon más materia seca que el resto de las colectas.
Los resultados coinciden con los obtenidos por Cochran (1974), quien al evaluar caracteres de calidad física y fisiológica de la semilla en pimiento morrón observó que porcentaje de germinación y acumulación de materia seca en la semilla grande fue mayor en relación con los de semilla chica. Carballo (1992) señaló que la calidad física
y fisiológica es un elemento esencial a considerar en la producción de semillas, así como también es importante conocerlos, para el establecimiento y producción de cultivos con altos índices de calidad.
El análisis de componentes principales mostró que los dos primeros componentes contribuyeron a explicar 98.36% de la variación total. El componente principal 1 con un valor propio de 7.25, explicó 94.50% de la variación total, donde las variables de mayor contribución fueron peso de mil semillas, peso volumétrico y longitud de plántula. Las primeras dos variables corresponden a la calidad física de la semilla, y la última está relacionada con el vigor (Cuadro 4).
condiciones de manejo en su producción. Zúñiga (1988) en chile serrano observó alta calidad de semilla reflejada en mayores valores para las variables porcentaje de germinación, peso volumétrico, velocidad de emergencia y materia seca de plántula en los primeros dos cortes, así como también mayor calidad de fruto. Al respecto, Montes y Martínez (1992) mencionaron que en los frutos de los primeros nudos el grado de cruzamiento es bajo comparado con lo que ocurre en frutos de nudos superiores, contribuyendo a una calidad uniforme de las semillas.
En chile la maduración de las semillas generalmente coincide con el inicio de cambio de coloración de los frutos. Es importante destacar que no siempre es necesario esperar la maduración completa de los frutos; Generalmente,
Variable CP1 CP2Germinación (GE) 0.3472 0.5005Envejecimiento acelerado (EA) 0.3466 0.5195Peso volumétrico (PV) 0.3590 0.1446Peso de mil semillas (PMS) 0.3592 0.1377Longitud de plántula (LPL) 0.3555 -0.2420Longitud de raíz (LRA) 0.3561 -0.2445Peso seco parte aérea (PSA) 0.3516 -0.4105Peso seco de raíz (PSR) 0.3527 -0.3906Valor propio 7.25 0.22Porcentaje de variabilidad explicada 94.50 3.86Porcentaje de variación acumulada 94.50 98.36
Cuadro 4. Vectores y valores propios de los componentes principales CP1 y CP2 de las variables analizadas en semilla de 14 colectas de chile de agua producida en tres ambientes.
El componente principal 2 explicó 3.86% de la variación, las variables germinación con y sin envejecimiento acelerado fueron las que más influyeron en este componente. Estas variables dependen de la calidad fisiológica de la semilla (Cuadro 4).
Las colectas de San Sebastián Abasolo y Santiaguito presentaron valores sobresalientes para peso de mil semillas, peso volumétrico y longitud de plántula, que difieren de las colectas provenientes de Villa Sola de Vega y Zimatlan de Álvarez graficadas en el extremo opuesto del eje (Figura 1).
El tamaño de la semilla es influenciado por el genotipo, con diferencias entre y dentro de las colectas, aunque el tamaño real depende de su posición en la planta madre y las

Calidad de semilla en colectas de chile de agua (Capsicum annuum L.) de los Valles Centrales de Oaxaca, México 263
semillas provenientes de frutos en etapa de maduración ya alcanzaron la madurez fisiológica. A este respecto, Puente y Bustamante (1991); Randle y Honma (1981) y Edwards y Sundstrom (1987) mencionaron que entre los factores que tienen efecto en la calidad de la semilla están el grado de madurez del fruto a la cosecha y el tiempo de maduración de la semilla después de cosechados los frutos. Por lo general se cosecha cuando los frutos se tornan completamente rojos, bajo el supuesto de que se obtiene semilla de buena calidad; sin embargo, la pigmentación que coincide con tal calidad puede variar con el genotipo, con las condiciones agroclimáticas y con el manejo.
En el componente principal 2 las variables más importantes fueron germinación y envejecimiento acelerado. Las colectas con mayor porcentaje de germinación con y sin envejecimiento acelerado fueron de San Sebastián Abásolo y Santiaguito, con 92.7 y 51.4% y 89.6 y 49.3%, respectivamente; mientras que las de Zimatlán (70 y 33.2%) y Sola de Vega (70.6 y 33.6%) mostraron un menor porcentaje de germinación con y sin envejecimiento acelerado. Estas últimas colectas también presentaron menor peso volumétrico y peso de mil semillas, así como menor longitud de plántula y raíz. Copeland y McDonald (1995) mencionaron que la calidad fisiológica de la semilla se refleja en su tamaño y que la condición nutrimental de la planta madre, densidad de población y etapa de maduración de lasemilla son características relacionadas directamente. Las colectas de San Sebastián Abasolo y Santiaguito fueron superiores en promedio de los tres ambientes, y la de San Sebastián Abasolo mostró mayor calidad de semilla tanto física como fisiológica.
En el análisis de conglomerados, al realizarse el corte a un valor de 0.55, separó las colectas en cuatro grupos, la definición de los grupos la influenció el peso volumétrico, peso de mil semillas, longitud de plántula, germinación con y sin envejecimiento acelerado (Figura 2). El grupo I incluyó las colectas de Ejutla de Crespo, Santiago Apóstol, San Juan Guelavia I y II, San Sebastián Teitipac, San Jerónimo, Tlacochahuaya, Villa Sola de Vega y Zimatlán de Álvarez y se caracterizó por presentar menor porcentaje de germinación con y sin envejecimiento acelerado, 74 y 36%, respectivamente, así como una menor calidad física de la semilla, reflejada en un menor peso volumétrico y peso de mil semillas (Cuadro 5).
Figura 1. Representación de la diversidad de 14 colectas de chile de agua (Capsicum annuum L.) en función de los dos primeros componentes principales.
CP2...
0.0
0.6
0.4
0.2
0.0
-0.2
-0.4
-0.6
-0.8
-1.0
-1 -2 0 2 4 6 CP1
TCH
EJU SAP
TLAI
TLAII
SSA
SAN
CP1 (94.5 %) PMS, PV y LPL
SFL
SPH CP2 (3.86 %) GE, EA
SV SUGII ZIM SST
SJGI
DISTANCIA
PROMEDIO
ENTRE
GRUPOS
1.50
1.25
1.00
0.75
0.50
0.25
0.00 EJU SAP SJGI SST TCH SJGII ZIM SV TLI TLII SSA SAH SF SPH
GI GII GIII GIV
Figura 2. Agrupamiento por el método UPGMA en 14 colectas de chile de agua (Capsicum annum L.).
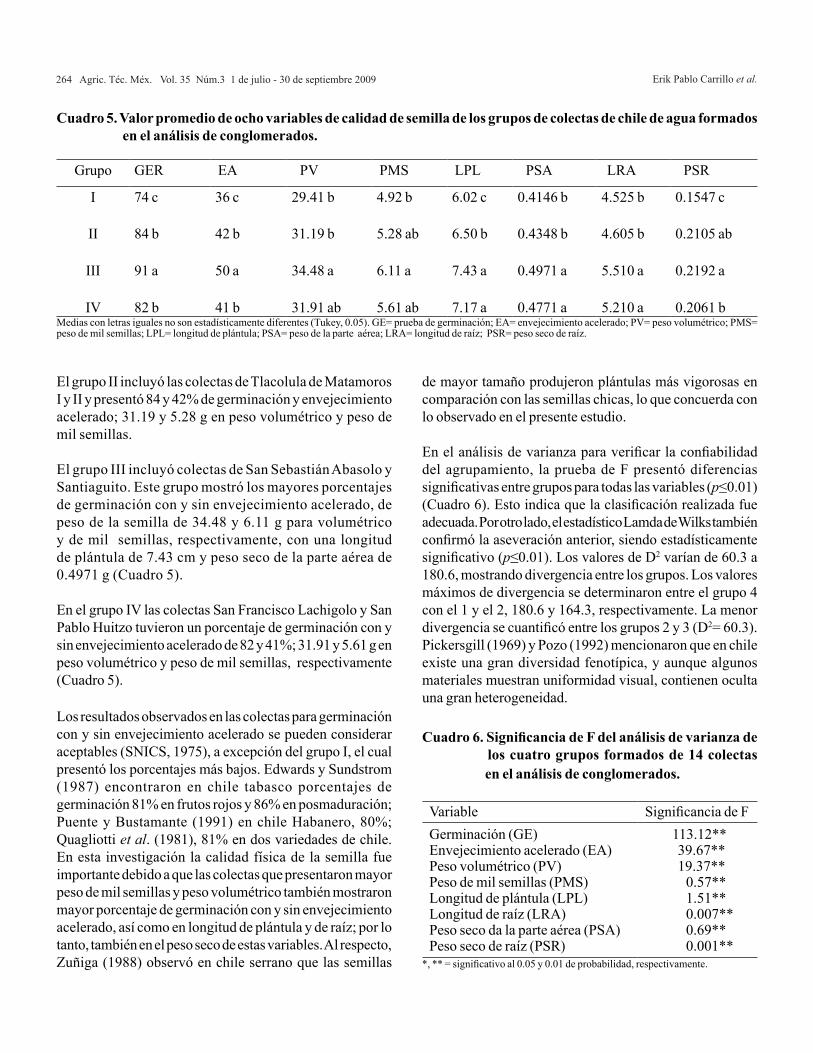
Erik Pablo Carrillo et al.264 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
El grupo II incluyó las colectas de Tlacolula de Matamoros I y II y presentó 84 y 42% de germinación y envejecimiento acelerado; 31.19 y 5.28 g en peso volumétrico y peso de mil semillas.
El grupo III incluyó colectas de San Sebastián Abasolo y Santiaguito. Este grupo mostró los mayores porcentajes de germinación con y sin envejecimiento acelerado, de peso de la semilla de 34.48 y 6.11 g para volumétrico y de mil semillas, respectivamente, con una longitud de plántula de 7.43 cm y peso seco de la parte aérea de 0.4971 g (Cuadro 5).
En el grupo IV las colectas San Francisco Lachigolo y San Pablo Huitzo tuvieron un porcentaje de germinación con y sin envejecimiento acelerado de 82 y 41%; 31.91 y 5.61 g en peso volumétrico y peso de mil semillas, respectivamente (Cuadro 5).
Los resultados observados en las colectas para germinación con y sin envejecimiento acelerado se pueden considerar aceptables (SNICS, 1975), a excepción del grupo I, el cual presentó los porcentajes más bajos. Edwards y Sundstrom (1987) encontraron en chile tabasco porcentajes de germinación 81% en frutos rojos y 86% en posmaduración; Puente y Bustamante (1991) en chile Habanero, 80%; Quagliotti et al. (1981), 81% en dos variedades de chile. En esta investigación la calidad física de la semilla fue importante debido a que las colectas que presentaron mayor peso de mil semillas y peso volumétrico también mostraron mayor porcentaje de germinación con y sin envejecimientoacelerado, así como en longitud de plántula y de raíz; por lo tanto, también en el peso seco de estas variables. Al respecto, Zuñiga (1988) observó en chile serrano que las semillas
de mayor tamaño produjeron plántulas más vigorosas en comparación con las semillas chicas, lo que concuerda con lo observado en el presente estudio.
En el análisis de varianza para verificar la confiabilidad del agrupamiento, la prueba de F presentó diferencias significativas entre grupos para todas las variables (p≤0.01) (Cuadro 6). Esto indica que la clasificación realizada fue adecuada. Por otro lado, el estadístico Lamda de Wilks también confirmó la aseveración anterior, siendo estadísticamente significativo (p≤0.01). Los valores de D2 varían de 60.3 a 180.6, mostrando divergencia entre los grupos. Los valores máximos de divergencia se determinaron entre el grupo 4 con el 1 y el 2, 180.6 y 164.3, respectivamente. La menor divergencia se cuantificó entre los grupos 2 y 3 (D2= 60.3). Pickersgill (1969) y Pozo (1992) mencionaron que en chile existe una gran diversidad fenotípica, y aunque algunos materiales muestran uniformidad visual, contienen oculta una gran heterogeneidad.
Variable Significancia de FGerminación (GE) 113.12**Envejecimiento acelerado (EA) 39.67**Peso volumétrico (PV) 19.37**Peso de mil semillas (PMS) 0.57**Longitud de plántula (LPL) 1.51**Longitud de raíz (LRA) 0.007**Peso seco da la parte aérea (PSA) 0.69**Peso seco de raíz (PSR) 0.001**
Grupo GER EA PV PMS LPL PSA LRA PSR
I 74 c 36 c 29.41 b 4.92 b 6.02 c 0.4146 b 4.525 b 0.1547 c
II 84 b 42 b 31.19 b 5.28 ab 6.50 b 0.4348 b 4.605 b 0.2105 ab
III 91 a 50 a 34.48 a 6.11 a 7.43 a 0.4971 a 5.510 a 0.2192 a
IV 82 b 41 b 31.91 ab 5.61 ab 7.17 a 0.4771 a 5.210 a 0.2061 b
Cuadro 5. Valor promedio de ocho variables de calidad de semilla de los grupos de colectas de chile de agua formados en el análisis de conglomerados.
Medias con letras iguales no son estadísticamente diferentes (Tukey, 0.05). GE= prueba de germinación; EA= envejecimiento acelerado; PV= peso volumétrico; PMS= peso de mil semillas; LPL= longitud de plántula; PSA= peso de la parte aérea; LRA= longitud de raíz; PSR= peso seco de raíz.
Cuadro 6. Significancia de F del análisis de varianza de los cuatro grupos formados de 14 colectas en el análisis de conglomerados.
*, ** = significativo al 0.05 y 0.01 de probabilidad, respectivamente.

Calidad de semilla en colectas de chile de agua (Capsicum annuum L.) de los Valles Centrales de Oaxaca, México 265
CONCLUSIONES
Las semillas de las colectas de chile de agua de los Valles Centrales de Oaxaca mostraron diferencias en calidad física y fisiológica.
Las características de la semilla de las colectas que mejor describen la calidad física y fisiológica fueron: peso de mil semillas, peso volumétrico, longitud de plántula, porcentaje de germinación y envejecimiento acelerado.
En chile de agua existe una estrecha relación entre la calidad física de la semilla y la calidad fisiológica.
LITERATURA CITADA
Besnier, R. F. 1989. Semillas: biología y tecnología. Editorial Mundi-Prensa. Madrid, España. 637 p.
Carballo, C. A. 1992. La calidad genética y su importancia en la producción de semillas. In: Mendoza, O. L.; Favela, C. E.; Cano, R. P. y Esparza, M. J. H. 1992. Situación actual de la producción, investigación y comercio de semillas en México. Memoria tercer Simposium, Torreón, Coahuila, México. pp. 80-101.
Cochran, H. L. 1974. Effect of seed size on uniformity of pimiento transplants (Capsicum annuum L.) at harvest time. J. Amer. Soc. Hort. Sci. 99:234-235.
Copeland, O. L. and McDonald, B. M. 1995. Principles of seed science and technology. Third edition. Chapman and Hall. New York, USA 409 p .
Delouche, J. C. and Baskin, C. C. 1996. Accelerated aging techniques for predicting the relative storability of seed lots. Seed Sci.Technol. 1:427-452.
Doijode, S. D. 1991. Effect of partial vaccum on viability of sweet pepper seeds. Capsicum Newsletter 6: 62-63.
Edwards, R. L. and Sundstrom, F. J. 1987. Afterripening and harvesting effects on Tabasco pepper seed germination performance. Hortscience.
22:473-475.International Seed Testing Association (ISTA). 2004.
International rules for seed testing. Bassersdorf, CH-Switzerland.
López, L. P. S. 1989. El chile de agua (Capsicum annuum L.) en los Valles Centrales de Oaxaca. Campo Experimental Valles Centrales de Oaxaca. Instituto Nacinal de Investigaciones Forestales, Agrícolas y Pecuarias. Oaxaca, México. 22 p. (Publicación Especial Núm. 2).
Mardia, K. V.; Kent, J. T. and Bibby, J. M. 1979. Multivariate analysis. Academic Press. London. 521 p.
Montes, C. F. y Martínez, C. J. 1992. Prácticas culturales relacionadas con la producción de semilla de chile. In: Mendoza, O. L.; Favela, C. E.; Cano, R. P. y Esparza, M. J. H. 1992. Situación actual de la producción, investigación y comercio
de semil las en México. Memoria tercer Simposium, Torreón, Coahuila , México. p. 80-101.Moreno, M. E.; Vásquez, E. M.; Rivera, A.; Navarrete, R.
and Esquivel, F. 1988. Effect of seed shape and size on germination of corn (Zea mays L.) stored under adverse conditions. Seed Sci. Technol. 26: 439-448.
Moreno, M. E. 1996. Análisis físico y biológico de semillas agrícolas. Tercera Ed. Instituto de Biología. Universidad Nacional Autónoma de México (UNAM). México. 393 p.
Pla, E. 1986. Análisis multivariado: método de componentes principales. Secretaría general de la Organización de los Estados Americanos. Washington, D. C. 94 p.
Perry, D. A. 1972. Seed vigour and field establishment. Horticultural Abstracts. 42:334-342.
Pickersgill, B. 1969. The domestication of chilli peppers. In: Ucko, P. J. P. and Dimbleby, G. W. The domestication and exploitation of plants and animals. Duck Worthy p. 443-450.
Pozo, C. O. 1992. Aspectos relevantes de la producción de semilla de chile en México. In: Mendoza, O. L.; Favela, C. E.; Cano, R. P. y Esparza, M. J. H. 1992. Situación actual de la producción, investigación y comercio de semillas en México. Memoria tercer Simposium, Torreón, Coahuila, México. p. 55-66.
Puente, P. C. y Bustamante, G. L. 1991. Efecto del estado de madurez y posmaduración del fruto de chile (Capsicum annuum L.) sobre la calidad de su semilla. Sociedad Mexicana de Ciencias Hortícolas A. C. IV Congreso Nacional. Saltillo, Coahuila, México. p. 187.
Quagliotti, L.; Antonucci, M. and Lantieri, S. L. 1981. Effects of postharvest ripening of the seeds within the berry in two varieties of pepper (Capsicum annuum L.). Riv. Ortoflororo Fruticoltura. 65(4):249-256.
Randle, W. M. and Honma, S. 1981. Dormancy in peppers. Scientia Horticulturae 14:19-25.

Erik Pablo Carrillo et al.266 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Van-Tangeres, O. F. R. 1997. Cluster analisis: data analisis. Communication Landscape Ecology. 6:174-207.
Zúñiga, S. A. 1988. Efecto de diferentes niveles de fertilización en la producción de fruto y semilla de chile serrano (Capsicum annuum L.). Tesis Profesional FAUANL. Marín, N. L., México. 88 p.
Statical Analysis System (SAS). 1998. SAS/STAT user´s guide, release 8.2 SAS Institute, Inc. N. C. USA.
Servicio Nacional de Inspección y Certificación de Semillas (SNICS). 1975. Normas para la certificación de semillas. Secretaría de Agricultura y Ganadería, Dirección General de Agricultura. México, D. F. 91 p.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 267-276
DISTRIBUCIÓN ESPACIAL DEL pH DE LOS SUELOS AGRÍCOLAS DE ZAPOPAN, JALISCO, MÉXICO*
pH SPATIAL DISTRIBUTION IN AGRICULTURAL SOILS OF ZAPOPAN, JALISCO, MEXICO
Daniel Ibarra Castillo1, José Ariel Ruiz Corral2§, Diego Raymundo González Eguiarte3 y José Germán Flores Garnica2
1Universidad de Guadalajara-CUCBA-Departamento de Ciencias Ambientales. Km 15.5 carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco.([email protected]). 2Campo Experimental Centro Altos de Jalisco, México, INIFAP. Km 8 carretera Tepatitlan-Lagos de Moreno, Tepatitlan, C. P. 47600, Jalisco. Tel. 01 333 64 1 69 69 y 64 1 20 61 3Universidad de Guadalajara-CUCBA-Departamento de Desarrollo Rural Sustentable Km 15.5 carretera Guadalajara-Nogales, Predio las Agujas, Nextipac, Zapopan, Jalisco.([email protected]). §Autor para correspondencia: [email protected].
* Recibido: Septiembre, 2007
Aceptado: Febrero, 2009
RESUMEN
El objetivo del presente estudio fue determinar la distribución espacial del pH del suelo en la región agrícola del municipio Zapopan, Jalisco, México. En 2004 se colectaron 105 muestras de suelo georeferenciadas del estrato 0-60 cm y se les determinó el pH. Los valores de pH se capturaron en hoja de cálculo electrónico para integrar una base de datos georeferenciada, a partir de la cual se probaron dos métodos de interpolación espacial: Kriging ordinario y Distancia Inversa Ponderada. Se determinó el cuadrado medio del error y la validación cruzada de los dos métodos y se seleccionó el Kriging Ordinario para generar una imagen raster del pH, a partir de la cual, mediante análisis espacial, se efectuaron diferentes interpretaciones sobre la distribución espacial del pH. Se utilizó el sistema de información geográfica Idrisi32.Se determinó que la totalidad de los suelos agrícolas de Zapopan son de condición ácida, aunque en diferentes niveles de intensidad. El 61.10% de la superficie agrícola le correspondió la condición de pH fuertemente ácido y al 38.90% moderadamente ácido.
Palabras clave: geoestadística, interpolación espacial, mapas temáticos, SIG, pH.
ABSTRACT
The object ive of th is s tudy was to determine the s p a t i a l d i s t r i b u t i o n o f s o i l p H i n a g r i c u l t u r a llands in the Zapopan county, Jalisco, Mexico. In 2004, 105 geo-referenced soil samples were takenin the county from 0-60cm depths cores and analyzed for pH. The determined pH values were organized i n an e l ec t ron i c sp r eadshee t t o i n t eg ra t e a geo-referenced data base. Two spatial interpolation techniques were tested: Ordinary Kriging andInverse Distance Weighting. By comparing the squaremean error and a cross validation, Ordinary Kriging was selected for generating a raster image of soil pH. With this image and using Idrisi32 geographical informat ion system i t was poss ible to obtain d i f f e r e n t i n t e r p r e t a t i o n s o f s o i l p H s p a t i a l distribution. All the agricultural soils of Zapopan are acid; although, at different levels of intensity. A strong acidic condition was found in 61.10% of the area and moderate acidic condition in 38.90%.
Key words: geostatistics, GIS, pH, spatial interpolation, thematic maps.

Daniel Ibarra Castillo et al.268 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
INTRODUCCIÓN
En general, el pH óptimo de los suelos agrícolas debe variar entre 6.5 y 7.0 para obtener los mejores rendimientos y la mayor productividad (Prasad and Power, 1997). El pH de un suelo ácido se puede mejorar gradualmente mediante el manejo apropiado y con aplicacion de cal (Prasad and Power, 1997; Havlin et al., 2005). El pH del suelo influye en la disponibilidad de los nutrimentos para las plantas, es decir, este factor puede ser la causa de que se presente deficiencia, toxicidad o que los elementos no se encuentren en niveles adecuados (Benton, 2003). Por otra parte, valores extremos del pH pueden afectar la estructura del suelo (Edward, 2000).
La práctica más utilizada para corregir el pH ácido del suelo es el encalado, con el cual se logra establecer un ambiente propicio en la raíz para el desarrollo normal de los cultivos (Benton, 2003). Tomando en cuenta el efecto del pH sobre la productividad de los cultivos, es importante no solo su determinación en el suelo, sino también conocer y cuantificar las superficies afectadas por valores extremos de este factor (Brejda et al., 2000).
En el municipio de Zapopan, Jalisco los suelos agrícolas son predominantes son Regosoles (SEMADES, 2006) y se caracterizan por ser de textura arenosa, bajo contenido de materia orgánica (Ibarra et al., 2007) y pH ácido (Ruiz et al., 1997); para contrarrestar el bajo nivele de pH, es práctica común la aplicación de cal al suelo, lo que coadyuva a que la zona sea considerada como una de las más productivas del país bajo condiciones de temporal (Ruiz et al., 1997); Lo anterior, debido a la profundidad y alta capacidad de retención de humedad de los suelos y al régimen de lluvias favorable, tanto en cantidad como en distribución a lo largo del ciclo de cultivo (Ruiz et al., 2003a).
La evaluación del estado actual del recurso edáfico en una región, implica la distribución espacial de una o varias características, lo cual se realiza con base en el muestreo de campo y procedimientos de interpolación geográfica para generar imágenes y/o mapas temáticos (Flores et al., 2002; Anguiano et al., 2003). Dos de los procedimientos de interpolación geográfica de más uso son: distancia inversa ponderada (DIP) (Hartkamp et al., 1999) y el método kriging ordinario (KO) (Bruz et al. 1996). DIP es el método de interpolación más comúnmente utilizado, debido a la sencillez de los cálculos. Sin embargo, la técnica de KO ha probado su utilidad y ventaja sobre la mayoría de los métodos
de interpolación para la descripción de la distribución espacial de variables geológicas y edáficas, tales como fósforo disponible, potasio intercambiable y pH (Webster y McBratney, 1987). Bruz et al. (1996), compararon la eficiencia de seis métodos de interpolación geográfica para la estimación de propiedades de suelos en puntos no muestreados y reportaron que KO fue el más consistente y confiable. Dicha consistencia se manifestó tanto en la condición de muestreo libre como estratificado por tipo de suelo. Las ventajas de KO derivan de la capacidad de este método para proveer el mejor estimador insesgado (Olea, 1991).
El método KO se basa en la teoría de variables regionalizadas desarrolladas en los años 60’s por Matheron (Hemyari y Nofziger, 1987) y utiliza semivariogramas, los cuales son una medida de la correlación espacial entre observaciones hechas en varios puntos y describe la función de varianza sobre una distancia en incremento; para lo cual, se requiere formar pares de observaciones. Cuando se utiliza el semivariograma, la atención se enfoca sobre las diferencias entre las observaciones, por lo que el peso de las observaciones cambia de acuerdo con el arreglo espacial de las muestras. El método KO utiliza una combinación lineal de pesos en puntos conocidos ΣλiYi para estimar el valor en un punto desconocido, donde Yi son las variables evaluadas en los sitios de observación, λi son los pesos Kriging (De Beurs, 1988).
En comparación, el método DIP calcula valores para los sitios no muestreados con base en el promedio ponderado de los valores de sitios cercanos sí muestreados. La ponderación está dada por una función inversamente proporcional a la distancia entre el punto a ser estimado y el punto muestreado (De la Mora et al., 2004).
Para orientar debidamente los programas de rehabilitación de suelos en una región, así como para facilitar la generación y la transferencia de tecnología, se requiere de un diagnóstico cuantitativo y ágil de la situación del pH en el suelo. El objetivo del presente estudio fue determinar la distribución espacial del pH del suelo en la región agrícola de Zapopan, Jalisco.
MATERIALES Y MÉTODOS
Área de estudio
El estudio se llevó a cabo en la zona agrícola del municipio de Zapopan, Jalisco, ubicado en el Valle de Zapopan, entre las coordenadas extremas 20° 50´ 54.4” y 20° 41´ 24.03” de

Distribución espacial del pH de los suelos agrícolas de Zapopan, Jalisco, México 269
latitud norte y 103° 24´ 19.83” y 103° 33´ 23.17” de longitud oeste, con altitud promedio de 1 600 m (INEGI, 2001). La precipitación pluvial media anual es 979 mm, distribuidos entre los meses de mayo y octubre; el mes más lluvioso es julio con 251 mm. La temperatura media anual es de 19.7 ºC; mayo, junio y julio, el trimestre más cálido (Ruiz et al., 2003a). Predominan los suelos del tipo regosol, que cubren más de 90% de la superficie agrícola (SEMADES, 2006). Los regosoles son suelos formados a partir de materiales no consolidados, de textura arenosa, se pueden encontrar en distintos climas y con diversos tipos de vegetación. Se caracterizan por no presentar estratos distintos. En general son claros, parecidos a la roca que los subyace cuando no son profundos. Muchas veces están acompañados de afloramiento de rocas o tepetate y son susceptibles a la erosión (FAO, 1985).
Geológicamente, existen rocas predominantes ígneas extrusivas ácidas: riolita porfírica, toba liparítica, pómez y obsidiana, asociadas con algunos afloramientos de basalto, que son los materiales que dan origen a los suelos; de éstos, los más recientes son los de la Sierra de La Primavera (Ibarra et al., 2007; Secretaría de Gobierno de Jalisco, 1988), la cual manifiesta un relieve, producto de una caldera de tipo explosivo de aproximadamente 11 km de diámetro, modificada por diversos domos riolíticos en la zona de fractura anular, cuyos productos piroclásticos se depositaron en la región central del municipio, dando origen a los suelos del Valle de Zapopan (Curiel et al., 1991).
Inicialmente, el área agrícola del municipio de Zapopan se delimitó tomando como fuente las imágenes vectoriales de uso del suelo correspondientes al proyecto de ordenamiento ecológico territorial del estado de Jalisco (SEMADES, 2006). Con esta fuente y mediante el uso del Sistema de Información Geográfica (SIG) Idrisis32 (Eastman, 1999), se estimó una superficie de 21 037.71 ha (de 1980 a 1990).
Sin embargo, durante el trabajo de muestreo en campo, se observó que una proporción de esta superficie ya no es de uso agrícola, por lo que se procedió a realizar un levantamiento de la superficie agrícola real mediante geoposicionamiento, utilizando un GPS Garmin XL-12. Como producto de este trabajo se determinó que la superficie agrícola del municipio de Zapopan en 2004 fue de 11 871.41 ha; la diferencia entre las dos superficies obedece al desarrollo urbano en las últimas dos décadas.
Trabajo de campo y laboratorio
Para el muestreo de suelos, se partió del mapa del área agrícola del municipio de Zapopan y por medio del SIG Arc View 3.2, se estableció un diseño sistemático con base a una plataforma de proyección de cuadrícula UTM (universal transverso de mercator), con distancia cada 900 m entre sitios de muestreo georeferenciado, lo que produjo un total de 242 sitios a muestrear (Figura 1) y de los cuales sólo se colectaron 105 muestras en 2004 debido a que al revisar información cartográfica actualizada se observó que el área representada por las muestras restantes ya estaba urbanizada. De esta forma, la superficie total estudiada fue 11 871.41ha de uso agrícola. La distancia entre sitios de muestreo se determinó con los siguientes criterios: a) los suelos en la región son de pH ácido INEGI (2001), b) el material madre da origen a suelos ácidos y que estos han estado expuestos a degradación por el manejo intensivo, que involucra el uso excesivo de insumos agrícolas (Ruiz et al., 2003b), y c) un muestreo previo del pH (en 25 ha) a intervalos de 50, 100, 250 y 500 m, realizado en 2002, dentro del marco de esta misma investigación y que reflejó muy poca variabilidad del pH, aún a distancias de 500 m. El muestreo se realizó en una sola toma en el estrato de 0-60 cm, los sitios de muestreo se localizaron con el GPS ya descrito. Las muestras colectadas se transportaron al laboratorio de Física y Química de Suelos de la Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA), para la determinación del pH, que se hizo con el método potenciométrico (NOM 021, 2000). Los resultados del análisis de laboratorio se capturaron en hoja de cálculo electrónico de Windows (Microsoft Excel), para integrar la base de datos georeferenciada.
Métodos de interpolación espacial
Se utilizó la matriz de datos georreferenciados de los valores de pH para desarrollar un proceso de interpolación espacial, considerando dos métodos: Kriging ordinario (KO) y Distancia inversa ponderada (DIP). Estos dos métodos de interpolación se ejecutaron mediante el sistema geoestadístico GS+ ver. 5.1 (Gamma, 2001). La interpolación con el método KO permite convertir los datos de la variable de estudio (en este caso pH) en superficies continuas (Burrough y McDonnell, 1998); se puede calcular mediante la ecuación siguiente (De la Mora et al., 2004):

Daniel Ibarra Castillo et al.270 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
(1)
donde, muestreado x0; λi = ponderación para el punto de muestreo i en el sitio xi; Z (xi)= valor de la variable Z (pH), para un punto de muestreo i, en el sitio xi; n= numero de puntos de muestreo. La varianza de KO se calcula con la expresión:
(2)
donde, ado x0; C(x0,x0)= covarianza del punto a estimar en el sitio x0 consigo mismo; C (xi,x0)= covarianza del punto de muestro en el sitio xi y el punto a estimar en el sitio x0 . Los valores de pH se interpolaron con base en KO, con la finalidad de obtener superficies continuas de valores. Para esto se obtuvo el variograma de los datos que es la herramienta básica del método KO y se utiliza para modelar la correlación espacial entre observaciones (Czaplewski et al., 1994). La variación espacial definida por el variograma, se modeló para estimar los valores de pH en los sitios que no se muestrearon.
El método DIP se basa en el cálculo de un valor para sitios no muestreados, para lo cual utiliza un promedio ponderado de los valores de sitios cercanos que sí se muestrearon. La
ponderación es producto de una función inversamente proporcional a la distancia entre el punto a estimar y el punto muestreado, como se expresa en la siguiente función lineal (De la Mora et al., 2004):
(3)
donde, = valor estimado en el sitio no muestreado x0; desde cada sitio muestreado hacia el sitio no muestreado a estimar; p= exponente de distancia (el cual es definido iterativamente hasta lograr el mínimo error medio); n= número de sitios muestreados. En este estudio se probaron tres niveles de ponderación de DIP: a) DIP simple (exponente de distancia igual a 1); DIP cuadrático (exponente de distancia igual a 2) y DIP cúbico (exponente de distancia igual a 3).
El ajuste de los métodos de interpolación se probó mediante un procedimiento de validación cruzada, el cual se ejecutó desde el sistema GS+ ver. 5.1, y consiste en la generación de residuales, que representan la diferencia entre valores observados y estimados. Estos úl t imos se generan removiendo el valor
Figura 1. Sitios de muestreos georeferenciados (242) en campo, del área agrícola del municipio de Zapopan, Jalisco.
Escala 1:300,0002 0 2 4 6 8 10
Kilometros
N
ZAPOPAN
Proyección Cuadrícula Universal Transversa de Mercator(UTM) (Zona 13, Datum: NAD27)
SITIOS DE MUESTREOS
GEORREFERENCIADOS
1700000
1700000
1800000
1800000
1900000
1900000
2000000
2000000
2200
000 2200000
2300
000 2300000
2400
000 2400000
2500
000 2500000
2600
000 2600000
L E Y E N D A
Sitios de MuestreosFuente: Sitios Localizados mediante el Diseño Sistemático, basado en una Plataforma de Proyección de Cuadrícula UTM (Universal Transversa de Mercato), con distancias cada 900 m, entre Sitios de muestreos), 2004.
SIMBOLOGÍA
Límite Municipal
Mancha Urbana
MAPA: Figura 3.Elaboro: M.C. Daniel Ibarra Castillo.
Fecha: 28 de J unio de 2009.
1830000
1830000
1840000
1840000
1850000
1850000
1860000
1860000
2340
000 2340000
2350
000 2350000
2360
000 2360000
2370
000 2370000
2380
000 2380000
Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ
Ñ ÑÑ
Ñ Ñ
Sitios de Muestreos Georreferenciados (242) en Campo, del Área de Estudio de la Zona Agrícola del
Municipio de Zapopan, Jalisco.
Fuente:Información Básica y Topográfica: INEGI; 1974-1984.lLímites Políticos: Instituto de Geografía. UdeG, 1990.Vías de Comunicación y Poblados : SIGSA, INEGI, 1997.Ordenamiento Ecológico Territorial del Estado de J alisco: Gob. del Edo. Jal., INE, UdeG.COPLADE: Vegetación,1980
)( 0~ xZ K O
)( 0~ xZ K O )(
1xi
n
ii Z⋅∑
=λ=
= valor estimado de KO en el sitio no
µλσ +⋅−= ∑=
)()()( ,, 01
0002 xxxx i
n
iiKO CCx
)( 02 xKOσ = varianza de KO en el sitio no muestre-
∑
∑
=
== n
ip
i
i
n
ip
i
d
xdx
1
10 1
)(1
)(*ˆβ
β
)(*ˆ0xβ
)( ixβ = valor observado en el sitio xi; = distanciaspid
^
^
Sitios de muestreos georeferenciados (242) en campo, del área de estudios de la zona agrícola del municipio de Zapopan, Jalisco.
SITIOS DE MUESTREOS
GEOREFERENCIADOS
MAPA: Figura 3.Elaboro: M.C. Daniel Ibarra Castillo. Fuente:Información Básica y topográfica:INEGI; 1974-1984. ILímites Políticos: Instituto de geografía. UdeG.,1990.Vías de Comunicación y poblados:SIGSA, INEGI, 1997.Ordenamiento ecológico territorial del Estado de Jalisco: Gob. del edo. Jal.,INE,UdeG.COPLADE: Vegetación, 1980Fecha: 28 de Junio de 2009.
Escala 1:300,0002 0 2 4 6 8 10
Kilometros
N
ZAPOPAN
Proyección Cuadrícula Universal Transversa de Mercator(UTM) (Zona 13, Datum: NAD27)
SITIOS DE MUESTREOS
GEORREFERENCIADOS
1700000
1700000
1800000
1800000
1900000
1900000
2000000
2000000
2200
000 2200000
2300
000 2300000
2400
000 2400000
2500
000 2500000
2600
000 2600000
L E Y E N D A
Sitios de MuestreosFuente: Sitios Localizados mediante el Diseño Sistemático, basado en una Plataforma de Proyección de Cuadrícula UTM (Universal Transversa de Mercato), con distancias cada 900 m, entre Sitios de muestreos), 2004.
SIMBOLOGÍA
Límite Municipal
Mancha Urbana
MAPA: Figura 3.Elaboro: M.C. Daniel Ibarra Castillo.
Fecha: 28 de J unio de 2009.
1830000
1830000
1840000
1840000
1850000
1850000
1860000
1860000
2340
000 2340000
2350
000 2350000
2360
000 2360000
2370
000 2370000
2380
000 2380000
Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ
Ñ ÑÑ
Ñ Ñ
Sitios de Muestreos Georreferenciados (242) en Campo, del Área de Estudio de la Zona Agrícola del
Municipio de Zapopan, Jalisco.
Fuente:Información Básica y Topográfica: INEGI; 1974-1984.lLímites Políticos: Instituto de Geografía. UdeG, 1990.Vías de Comunicación y Poblados : SIGSA, INEGI, 1997.Ordenamiento Ecológico Territorial del Estado de J alisco: Gob. del Edo. Jal., INE, UdeG.COPLADE: Vegetación,1980
LEYENDA Sitios de muestreos
Fuente: Sitos localizados mediante el diseño sistemático, basado en una plataforma de proyec-ción de cuadrícula (UTM) universal transversa de mercado, con distancias cada 900 m, entre sitiosde muestreos), 2004.
SIMBOLOGÍA
Límite municipal
Mancha urbana
Escala 1:300,0002 0 2 4 6 8 10
Kilometros
N
ZAPOPAN
Proyección Cuadrícula Universal Transversa de Mercator(UTM) (Zona 13, Datum: NAD27)
SITIOS DE MUESTREOS
GEORREFERENCIADOS
1700000
1700000
1800000
1800000
1900000
1900000
2000000
2000000
2200
000 2200000
2300
000 2300000
2400
000 2400000
2500
000 2500000
2600
000 2600000
L E Y E N D A
Sitios de MuestreosFuente: Sitios Localizados mediante el Diseño Sistemático, basado en una Plataforma de Proyección de Cuadrícula UTM (Universal Transversa de Mercato), con distancias cada 900 m, entre Sitios de muestreos), 2004.
SIMBOLOGÍA
Límite Municipal
Mancha Urbana
MAPA: Figura 3.Elaboro: M.C. Daniel Ibarra Castillo.
Fecha: 28 de J unio de 2009.
1830000
1830000
1840000
1840000
1850000
1850000
1860000
1860000
2340
000 2340000
2350
000 2350000
2360
000 2360000
2370
000 2370000
2380
000 2380000
Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ Ñ Ñ Ñ Ñ Ñ Ñ
Ñ Ñ Ñ Ñ Ñ Ñ ÑÑ Ñ Ñ
Ñ ÑÑ
Ñ Ñ
Sitios de Muestreos Georreferenciados (242) en Campo, del Área de Estudio de la Zona Agrícola del
Municipio de Zapopan, Jalisco.
Fuente:Información Básica y Topográfica: INEGI; 1974-1984.lLímites Políticos: Instituto de Geografía. UdeG, 1990.Vías de Comunicación y Poblados : SIGSA, INEGI, 1997.Ordenamiento Ecológico Territorial del Estado de J alisco: Gob. del Edo. Jal., INE, UdeG.COPLADE: Vegetación,1980

Distribución espacial del pH de los suelos agrícolas de Zapopan, Jalisco, México 271
RESULTADOS Y DISCUSIÓN
pH del suelo
De acuerdo con los valores de pH del suelo obtenidos en los muestreos de campo, se apreció que en promedio el pH en el área de estudio es de 5.4, con coeficiente de variación de 0.134. Los valores más bajo y más alto fueron 4.1 y 7.1, respectivamente. Estos resultados coinciden de manera aproximada con los reportados por la Fundación Produce Jalisco en 2000 (FPJ, 2000), en los que, en promedio, se obtuvo un valor de 5.3 para 25 muestreos dentro del área de estudio, con valor mínimo de 4.4 y un valor máximo de 6.9.
Aproximadamente dos tercios de las muestras correspondieron a la condición fuertemente ácido, con valores de pH de 4.2 a 5.5, mientras que el tercio restante, se clasificó como moderadamente ácido, con valores entre 5.6 y 6.5, Cuadro 1. De las 105 muestras sólo en dos de ellas se presentó la condición extremadamente ácida, con valores menores a 4.2; en 59 muestras la condición resultante fue de fuertemente ácido, con valores entre 4.2 y 5.5; en 36 muestras se observó la condición moderadamente ácido, con valores entre 5.6 y 6.5; en seis muestras mostraron valores de 6.6 a 7.3 la condición fue neutro y a dos de las muestras correspondió la condición moderadamente alcalino, con valores entre 7.4 y 8.3. De acuerdo con estos resultados, en términos generales, los suelos de la región agrícola de Zapopan son predominantemente ácidos, lo que coincide con otros estudios efectuados en la región (INEGI, 2004).
El pH “fuertemente ácido” en los suelos de la región se explica en función de varios factores, entre ellos la roca madre que los subyace (Curiel et al 1991; Aguirre, 2001), la actividad agrícola intensiva (Ruiz et al., 1997) y la aplicación de insumos que provocan acidez (Guerrero, 1996; Edward, 1997).
Variación espacial del pH
En el proceso de análisis de la distribución espacial, al interpolar los valores de pH con los métodos KO y DIP y al validar las estimaciones generadas mediante los procedimientos de validación cruzada y comparación del valor del cuadrado medio del error (Cuadro 2), se determinó que los métodos de interpolación espacial se comportaron de manera similar. Sin embargo, se aprecia que el método KO fue ligeramente superior.
muestreado de un sitio en particular, después de lo cual su valor es estimado con base a los restantes sitios (Isaaks y Srivastava, 1989).
Se calculó el cuadrado medio del error (CME) de los residuales para identificar cuál de los métodos de interpolación se ajustó mejor a los datos de pH determinados en campo. En este sentido se selecciona el que presenta un menor valor de CME (De la Mora et al., 2004). El cuadrado medio del error resume estadísticos que incorporan tanto el sesgo como la dispersión de la distribución del error (CME= varianza + sesgo2), el cual se calcula de la forma siguiente (Isaaks y Srivastava, 1989):
(4)
donde, CME= cuadrado medio del error; n= número de sitios muestreados y r= residuales (diferencia entre los valores estimados y los valores observados).
Generación de la imagen raster de valores de pH
Los parámetros del modelo de interpolación de mejor ajuste se utilizaron para generar una imagen raster del pH en el módulo de análisis de superficie del SIG Idrisi32 (Eastman, 1999). A partir de la imagen raster y por medio del mismo SIG, se realizó el análisis de la variación espacial del pH en el área agrícola de Zapopan, para determinar la acidez o alcalinidad de los suelos del área de estudio con la clasificación propuesta por Moreno-Dahme (González et al., 1996): a) extremadamente ácido. Representa un valor de pH del suelo inferior a 4.2, condición que inhibe el desarrollo radical, por lo que la presencia de cultivos es prácticamente imposible, b) fuertemente ácido. Valor de pH del suelo entre 4.2 y 5.5, el cual permite el desarrollo radical de una gama estrecha de especies de cultivo que se desarrollan en suelos con tendencia ácida, c) moderadamente ácido. pH de 5.6 a 6.5 que representa una condición de suelo que permite el uso agrícola, considerando la implementación de una gama más o menos amplia de cultivos con preferencia de suelos de tendencia ácida, d) neutro. pH con valor de 6.6 a 7.3 que equivale a una condición del suelo que permite el desarrollo de una amplia gama de cultivos, e) moderadamente alcalino. Valores de pH que van de 7.4 a 8.3, que se traduce en suelos que permiten el desarrollo de cultivos con tendencia alcalina, y f) fuertemente alcalino. pH mayor que 8.3, lo cual restringe al cultivo de un estrecho grupo especies con preferencia evidente hacia los suelos alcalinos.
∑=−
=n
ir
nCME
1
2
11

Daniel Ibarra Castillo et al.272 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
En el variograma isotrópico del método KO se observó tendencia a la variabilidad espacial del pH (Sahin y Hassan, 1998). La tendencia de los datos se ajustó a un modelo exponencial, con una continuidad espacial definida (Journel y Huijbregts, 1978), Figura 2. El rango (Ao) correspondiente en este modelo, es decir, donde los valores del semivariograma tienden a estabilizarse, se ubica en un valor de 31 100 m. Esta distancia es la máxima entre dos pares de sitios en la que existe correlación espacial entre los mismos; más allá de esta distancia
no existe similitud suficiente entre los valores de los sitios para que pueda ser modelada (De la Mora et al., 2004). El parámetro “sill” (Co+C), que es el valor del semivariograma para distancias muy grandes (Issaks y Srivastava, 1989) se ubicó en 0.873. El efecto “nugget” (Co), que define una discontinuidad en el origen (Samra et al., 1989), tiene un valor de 0.436, esto es aproximadamente la mitad del valor del “sill”. Este valor de Co no manifiesta ausencia de estructura en el semivariograma (Journel y Huijbregts, 1978).
Intervalode pH
Condición de acidez o alcalinidad Superficie(ha)
Superficie agrícola (%) Núm. de muestras
< 4.2 Extremadamente ácido - - 24.2 - 5.5 Fuertemente ácido 7 259.28 61.10 595.6 - 6.5 Moderadamente ácido 4 612.13 38.90 366.6 - 7.3 Neutro - - 67.4 - 8.3 Moderadamente alcalino - - 2
> 8.3 Fuertemente alcalino - - 0Total 11 871.41 100 105
Método Cuadrado medio del errorKriging ordinario (KO) 0.63248Distancia inversa ponderada (exponente 1) 0.63895Distancia inversa ponderada (exponente 2) 0.64884Distancia inversa ponderada (exponente 3) 0.66793
Figura 2. Variograma isotrópico del pH del suelo agrícola en el municipio de Zapopan, Jalisco, México.
Cuadro 1. Valores de pH y condiciones de acidez del suelo agrícola de Zapopan, Jalisco.
Cuadro 2. Valores del cuadrado medio del error en los métodos de interpolación para estimar el pH del suelo de uso agrícola en Zapopan, Jalisco.
0.600
0.450
0.300
0.150
0.000
Sem
ivar
ianz
a
0 3592.05 7184.1 10776.15 14368.2
Distancia de separación (m)
Co= 0.436; Co+C= 0.873; Ao= 31100.00

Distribución espacial del pH de los suelos agrícolas de Zapopan, Jalisco, México 273
Con base en el análisis anterior y debido a que el método KO es considerado como el mejor estimador lineal insesgado (Olea, 1991) se decidió utilizar este método para generar la imagen raster de los valores de pH. De esta forma, los parámetros del modelo seleccionado se implementaron en el sistema Idrisi32 (Eastman, 1999) y se generó la imagen raster de pH del área de estudio (Figura 3a). En ella se aprecia la distribución espacial de los puntos de muestreo dentro de la zona de estudio, con valores de <4.5 a 6.2. Estos valores
La distribución espacial de la desviación estándar del pH en el área interpolada se representa en fajas concéntricas de dispersión, Figura 3b. Con base en la información obtenida las estimaciones de pH están en el rango de desviación estándar de 0.69 a 0.77. Esto implica que las estimaciones tienen una precisión alta. Se observó más alta variación en la faja concéntrica central que en las sucesivas fajas hacia el exterior de la Figura 3b, por lo que habría que recomendar que muestreos futuros se realicen en las últimas dos fajas exteriores del área establecida y sobre todo, en las áreas noreste y suroeste que corresponden a la superficie agrícola de Zapopan (Figura 3a).
En la Figura 4 se muestra la reclasificación de los valores de pH interpolados, se puede observar que los suelos “fuertemente ácidos” se distribuyen en las porciones
Figura 3a) Distribución del pH en el suelo agrícola del municipio de Zapopan, Jalisco, y 3b) valores de desviación estándar de pH en el área interpolada.
a) b)
extremos difieren ligeramente con los valores obtenidos de las muestras, debido a que ya son valores generados a partir de un modelo. Sin embargo, la información de esta imagen ratifica el diagnóstico preliminar (Cuadro 1), en el sentido de que, en términos generales, la condición de acidez en los suelos de Zapopan se ubica entre fuertemente ácido y moderadamente ácido, por lo que se concluye que la interpolación espacial con el método KO representa el nivel de acidez determinado con los valores muestrales.
noreste, noroeste, parte del centro, suroeste y sureste, en una superficie de 7 259.28 ha que equivale al 61.10% del área agrícola. Los suelos “moderadamente ácidos” se concentran en la parte norte, noroeste, oeste,cruzando por el centro hacia una porción de la zona suroeste en 4 612.13 ha, que equivalen a 38.90% del área agrícola.
Los mapas de las Figura 3a y 4 son fuente de información clave para planteamiento de programas de encalado de suelos Los niveles de acidez determinados en la presente investigación podrá variar en los años siguientes debido al manejo que reciban los suelos. Sin embargo, la delimitación obtenida de las dos condiciones resultantes (fuertemente y moderadamente ácido) de la acidez del suelo, difícilmente podrá cambiar en el futuro cercano.
<4.54.64.84.95.05.15.25.35.45.55.65.75.85.96.06.16.2
Desviación estándar del pH
0.77
0.75
0.730.710.60
2306216
2301631
2297238
2292644
2288456
650275 654206 658137 662068 666000Longitud (UTM)
Lat
itud
(UT
M)

Daniel Ibarra Castillo et al.274 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
CONCLUSIONES
Los métodos de interpolación utilizados permitieron determinar con precisión razonable la distribución espacial del pH en los suelos agrícolas de Zapopan, Jalisco.
El método Krigin Ordinario mostró ser ligeramente superior a los otros métodos en la interpolación espacial del pH del suelo. En el municipio de Zapopan, Jalisco, 61.10% de los suelos agrícolas son fuertemente ácidos y el resto moderadamente ácidos.
AGRADECIMIENTOS
A las siguientes instituciones: Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP), Fundación Produce-Jalisco y Universidad de Guadalajara (U de G).
LITERATURA CITADA
Aguirre, G. A. 2001. Química de suelos ácidos templados y tropicales. 1er. edición. Editorial UNAM. Universidad Nacional Autónoma de México, Cd. Universitaria. México, D. F. 272 p.
Anguiano, C. J.; Alcántar, R. J. J.; Ruiz, C. J. A.; González, A. I. J.; Vizcaíno, V. I.; Regalado, R. J. R. y De la Mora, O. C. 2003. Recursos edafo-climáticos para la planeación del sector productivo en el estado de Michoacán. INIFAP-CIRPAC-Campo Experimental Uruapan. 173 p. (Libro Técnico Núm. 1).
Bruz, D. J.; De Gruijter, J. J.; Barsman, B. A.; Visschers, R.; Bregt, A. K.; Brewuwsma, A. and Bouma, J. 1996. The performance of spatial interpolation methods and cloropleth maps to estimate properties at points: a soil survey case study. Environmetrics. 7(1):1-16.
Benton, J. J. Jr. 2003. Agronomic handbook. Management of crops, soils, and their fertility. CRC PRESS. Boca Raton. London. New York. Washington, D.C. USA. 450 p.
Brejda, J. J.; Moorman, T. B.; Smith, J. L.; Karlen, D. L.; Allan, D. L. and Dao, T. H. 2000. Distribution and variability of surface soil properties at a regional scale. Soil Sci. Soc. Am. J. 64: 974-982.
Burrough, P. A. and McDonnell, R. 1998. Principles of geographical information systems. Claredon Press Oxford. New York. 333 p.
Curiel, B. A.; Rangel, R. A.; Reyna, B. O.; Ponce, R. J.; Ramírez, Q. A. e Ibarra, C. D. 1991. Ordenamiento ecológico de Zapopan. Laboratorio bosque la primavera. Universidad de Guadalajara-H. Ayuntamiento de Zapopan. Zapopan, Jalisco. 145 p.
Figura 4. Distribución espacial del pH en los suelos agrícolas en el municipio de Zapopan, Jalisco.
650000
650000
655000
655000
660000
660000
665000
665000
670000
670000
2295
000 2295000
2300
000 2300000
2305
000 2305000
# #
#
#
Nextipac
San Francisco Tesistan
Santa Lucia
La Magdalena
Moderadamente ácido
Fuertemente ácido 7,259.28
LEYENDACONDICIONES DE pH SUPERFICIE AGRÍCOLA
ha. %61.10
38.904,612.1311,871.41 100.00
Escala: 1:100,0001 0 1 2 3
KilometrosProyección Cuadrícula Transversa de Mercator (UTM)
(Zona: 13; Datum: NAD27Elaboro: Daniel Ibarra Castillo.
Universidad de Guadalajara
N
Municipio de
Zapopan

Distribución espacial del pH de los suelos agrícolas de Zapopan, Jalisco, México 275
Czaplewski, R. L.; Reich, R. M. and Bechtold, W. A. 1994. Spatial autocorrelation in growth of undisturbed natural pine stand across Georgia. Fort. Sci. 40:314-328.
De Beurs, K. 1988. Evaluation of spatial interpolation techniques for climate variables (case study of Jalisco, Mexico). (NRG/GIS modeling lab). Department of Mathematics, Wageningen Agricultural University. Supervisor from CIMMYT, México: 68 p.
De la Mora, O. C.; Flores, G. J. G.; Ruiz, C. J. A. y García, V. J. 2004. Modelaje estocástico de la variabilidad espacial de la calidad de agua en un ecosistema lacustre. Rev. Int. Contam. Ambient. 20(3):99-108.
Edward, J. P. 1997. Soil science management. 3rd. Edition. Delmar publishers. Albany, New York. 393 p.
Edward, J. P. 2000. La ciencia del suelo y su manejo. Editorial Paraninfo. Madrid, España. 405 p.
Eastman, J. R. 1999. Idrisi32. Guide to GIS and image processing Volume I. Clark Labs, Clark University. Worcester, MA, USA. 193 p.
Organización de las Naciones Unidas para la Agricultura y la Alimentación (FAO). 1985. Mapa mundial de los suelos (1:5´000,000) FAO-UNESCO. Leyenda revisada. Roma. pp. 140.
Flores, L. H. E.; Byerly, M. K. F.; Aceves, R. J. J. y Ruiz, C. J. A. 2002. Diagnóstico del sistema de producción de agave con énfasis en problemas fitosanitarios. In: análisis agroecológico del agave tequilana Weber var. Azul con énfasis en problemas fitosanitarios en Jalisco. INIFAP-CIRPAC-C.E. Altos de Jalisco. Tepatitlán, Jalisco., México. pp. 63-95. (Publicación Especial Núm. 1).
Fundación Produce Jalisco (FPJ). 2000. Base de datos de muestreos de suelo en campo en las áreas agrícolas del estado de Jalisco. Archivos electrónicos. Guadalajara, Jalisco, México.
Gamma. 2001. Geostatistics for the Environmental Sciences (GS+) versión 5.1.1. Disco compacto. Gamma Design Software. Plainwell, MI, USA.
González, A. I. J.; Ruiz, C. J. A.; López, A. J. G.; Mena, H. L.; Martínez, P. R. A.; Camarena, B. J. A.; Pérez, B. M. H.; Vázquez, V. V. y Villa, H., R. 1996. Diagnóstico del potencial agroecológico para caña de azúcar en áreas de ampliación del Ingenio de Puga, en el estado de Nayarit. INIFAP-CIRPAC. Tepic, Nayarit, México. 86 p. (Publicación Especial Núm. 1).
Guerrero, G. A. 1996. El suelo, los abonos y la fertilización de los cultivos. Ediciones Mundi-Prensa Grafo, S. A. Bilbao. España. 206 p.
Hartkamp, A. D.; De Beurs, K.; Stein, A. and White, J. W. 1999. Interpolation techniques for climate variables. NRG-GIS Series 99-01. CIMMYT. México, D. F. 26 p.
Hemyari, P. and Nofziger, D. L. 1987. Analytical solution for punctual kriging in one dimension. Soil Sci. Soc. Am. J. 51(1):268-269.
Havlin, J. L.; Beaton, J. D.; Tisdale, S. L. and Nelson, W. L. 2005. Soil fertility and fertilizers. An introduction to nutrient management. Pearson Prentice Hall. 7th Edition. Upper Saddle River, NJ, USA. 515 p.
Ibarra, C. D.; Ruíz, C. J. A.; Flores, G. J. G. y González, E. D. R. 2007. Distribución espacial del contenido de materia orgánica de los suelos agrícolas de Zapopan, Jalisco. Revista Terra Latinoamericana 25(2). En Prensa.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2001. Modelo digital de elevación escala 1:50,000. In: modelos digitales de elevación. Disco compacto. Aguascalientes, Aguascalientes, México.
Instituto Nacional de Estadística, Geografía e Informática (INEGI). 2004. Edafología, guía para la interpretación de cartografía. Aguascalientes, Aguascalientes, México. 28p.
Isaaks, E. H. and Srivastava, R. M. 1989. An introduction to applied geostatistics. Oxford University Press. New York. 561 p.
Journel, A. G. and Huijbregts, A. V. 1978. Mining geostatistics, Academic Press, New York. 300p.
Norma Oficial Mexicana 021 (NOM 021) 2000. SEMARNAT año 2000, que establece las especificaciones de fertilidad, salinidad y clasificación de suelos. Estudios, muestreo y análisis. Estados Unidos Mexicanos. Secretaría de Medio Ambiente, Recursos Naturales y Pesca (SEMARNAT). 85 p.
Olea, R. A. 1991. Geostatistical glossary and multilingual dictionary. Oxford University Press, New York. 256p.
Prasad, R. and Power, J. F. 1997. Soil fertility management for sustainable agriculture. Lewis Publishers. Boca Raton. New York. 356p.

Daniel Ibarra Castillo et al.276 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Ruiz, C. J. A.; Flores, L. H. E.; Martínez, P. R. A.; González, E. D. R. y Nava, V. L. 1997. Determinación del potencial productivo de especies vegetales para el Distrito de Desarrollo Rural de Zapopan, Jalisco. INIFAP-CIRPAC-C. E. Centro de Jalisco. Tlajomulco de Zúñiga., Jalisco, México. 60 p. (Folleto Técnico Núm. 5).
Ruiz, C. J. A.; González, A. I. J.; Anguiano, C. J.; Vizcaíno, V. I.; Ibarra, C. D.; Alcalá, G. J.; Espinosa, V. S. y Flores, L. H. E. 2003a. Estadísticas climatológicas básicas para el estado de Jalisco (período 1961-2000). INIFAP-CIRPAC-Campo Experimental Centro de Jalisco. 1ª. Ed. Conexión Gráfica. Guadalajara, Jalisco, México. 281 p. (Libro Técnico Núm. 1).
Ruiz, C. J. A.; González, A. I. J.; Regalado, R. J. R.; Anguiano, C. J.; Vizcaíno, V. I. y González, E. D. R. 2003b. Recursos edafo-climáticos para la planeación del sector productivo en el estado de Jalisco. INIFAP-CIRPAC. Guadalajara, Jalisco, México. 172 p. (Libro Técnico Núm. 2).
Secretaría de Gobierno del Estado de Jalisco (SGJ). 1988. Los Municipios de Jalisco. Secretaría de Gobierno del estado de Jalisco, Centro estatal de estudios municipales de Jalisco. Colección: Enciclopedia de los municipios de México. México, D. F. 189 p.
Sahin, A. and Hassan, H. M. 1998. Enhancement of permeability variograms using outcrop data. The Arabian J. Sci. and Eng. 23(1):137-144.
Samra, J. S.; Gill, H. S. and Bhatia, V. K. 1989. Spatial stochastic modeling of growth and forest resource evaluation. For. Sci. 35:663-676.
Secretaría de Medio Ambiente para el Desarrollo Sustentable (SEMADES). 2006. Ordenamiento ecológico territorial del estado de Jalisco. Última modificación, 27 de julio de 2006. Disponible en: http://www.semades.jalisco.gob.mx/06/ordenamiento.htm. (consultada el 28 de noviembre de 2006).
Webster, R. and McBratney, A.B. 1987. Mapping soil fertility at broom’s barn by simple Kriging. J. Sci. Food Afric. 38:97-115.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 277-283
BIOLOGICAL NITROGEN FIXATION AND TUBER YIELD OF YAM BEAN IN CENTRAL MEXICO*
FIJACIÓN BIOLÓGICA DE NITRÓGENO Y RENDIMIENTO DE LA JÍCAMA EN EL CENTRO DE MÉXICO
Javier Zaragoza Castellanos-Ramos1§, Jorge Alberto Acosta-Gallegos1, Norberto Rodríguez Orozco1 y José Jesús Muñoz-Ramos2 1Plant Nutrition Program, Bajio Experimental Station, INIFAP, A. P. 112, Celaya, Guanajuato, Mexico. C. P. 38110. ([email protected]). Tel. 01 461 6 11 53 23 Ext. 200. 2Universidad Politécnica de Durango. Durango, Durango. Carretera Durango-México, km 9.5 C. P. 34000. Tel. 01 681 15 0 13 00. §Autor para correspondencia: [email protected].
The Mexican tuber bearing edible Pachyrhizus erosus is widely grown and consumed in central Mexico. The aim of this research was to study the relationships between the dynamics of N2 fixation and biomass accumulation and final tuber yield of two Phachyrrizus species, P. erosus bred cultivar San Juan EC550 and P. ahipa accession 102 introduced from Bolivia. The experiment was established on April 2nd, 2005 at the Bajio Experimental Station located near Celaya, Guanajuato, Mexico. Plant samples from one m2 were periodically taken from 67 days after planting (DAP) onwards, on those, nodule number and mass, as well as plant biomass and leaf area index (LAI), were recorded. For tuber yield an area of 3.8 m2 was harvested per plot. P. erosus reached the highest number of nodules at 170 DAP, while P. ahipa at 123 DAP. P. ahipa had a higher number of nodules than P. erosus, but of smaller size, thus, nodule dry mass was significantly higher (p<0.05) in P. erosus in most sampling dates. Fresh tuber yield of P. erosus was significantly (p<0.01) higher than that of P. ahipa and P. ahipa displayed a larger dry tuber weight that was due to a higher concentration of solids and fiber in the tuber. Higher tuber yield in P. erosus was related to a longer growth cycle, higher LAI and biomass accumulation.
Key words: Phachirizus erosus and P. ahipa, nitrogen harvest index, plant biomass.
RESUMEN
El tubérculo comestible mexicano Phachirizus erosus se produce y consume en la región central de México. El objetivo de esta investigación fue estudiar la asociación entre la dinámica de la fijación de N2, la acumulación de biomasa y el rendimiento de dos especies de Phachyrrizus, P. erosus cv. San Juan EC550 y P. ahipa accesión 102 introducida de Bolivia. El experimento se estableció el 2 de abril 2005 en el Campo Experimental Bajío, de Celaya, Guanajuato. A partir de los 67 días después de la siembra (DDS) se tomaron muestras periódicas de plantas de 1 m2; en las muestras se determinaron el número y peso de los nódulos, la biomasa y el índice de área foliar (IAF). El rendimiento de tubérculos se determinó en todas las parcelas en una área de 3.8 m2. P. erosus alcanzó el mayor número de nódulos a los 170 DDS, mientras que P. ahipa lo hizo a los 123 DDS. P. ahipa desarrolló un mayor número de nódulos que P. erosus, pero de menor tamaño, por lo tanto, el peso de los nódulos fue significativamente mayor (p<0.05) en P. erosus en la mayoría
* Recibido: Octubre, 2007
Aceptado: Enero, 2009
ABSTRACT

Javier Zaragoza Castellanos-Ramos et al.278 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
de los muestreos. El rendimiento de tubérculos frescos fue significativamente mayor en P. erosus (p<0.01) y P. ahipa mostró un mayor peso seco, lo que fue debido a una mayor concentración de sólidos solubles y fibra. El mayor rendimiento de P. erosus se relacionó a un ciclo de cultivo más largo, mayor IAF y biomasa.
Palabra clave: Phachirizus erosus y P. ahipa, biomasa de la planta, índice de cosecha de nitrógeno.
INTRODUCTION
The Mesoamerican yam bean crop Phachyrizus erosus is native to Mexico, Guatemala and Northern Nicaragua, where it is known as “jicama”. This crop is widely grown in the Bajio region of central Mexico under irrigated plus rainfall conditions. Edible tuber production is best suited to sandy, light textured soils with moderate to favorable fertility and high soil organic matter content (Lind and Purcino, 1987). The yam bean is a crop of low nitrogen requirements (Tamez, 1987), since it has a high capacity to fix atmospheric N2 in symbiosis with nitrogen-fixing Rhizobium and Bradyrhizobium bacteria (Hafini et al., 1994; Badillo and Castellanos, 1996) and for biomass and tuber production (Sorensen, 1996).
In contrast with many of the grain legumes, a substantial amount of the fixed nitrogen is returned to the soil if the vegetative above ground parts are left in the field (Badillo and Castellanos, 1996; Sorensen, 1996). Therefore, the crop can be an important component in any agronomic rotation and of a sustainable land use, from both an ecological and socioeconomic standpoint (Hafini et al., 1994). This crop is particularly interesting to be used in crop rotations under organic farming. Adequate available soil P favorably influences Yam bean tuber growth and atmospheric N2 fixation, being P a first limiting factor for high tuber yields. Nodulation occurs in clusters of sparse fibrous roots closely proximate to the developing tuber. Nitrogen content of the tuber is from three to five times greater than potatoes, cassava, taro and sweet potato (Lind and Purcino, 1987).
Castellanos et al. (1997) conducted a field test in the central region of Mexico to assess the nitrogen fixation of two P. ahipa accessions (58 to 80 kg N ha-1) and three P. erosus bred cultivars (162 to 215 kg N ha-1). The amount of nitrogen recorded in the residues of P. erosus was
from 120 to 150 kg ha-1 twice the amount recorded in P. ahipa residue and is higher than the quantity recorded in practically all grain legumes (Sorensen, 1996). Following that line of research, an experiment was conducted to determine if there is an association between the dynamics of N2 fixation and biomass accumulation and tuber yield in P. erosus bred cultivar San Juan EC550 and P. ahipa accession 102, introduced from Bolivia.
MATERIALS AND METHODS
Plant culture
San Juan EC550 and AC-102 seeds, along with the reference crop maize hybrid H 358, were planted on April 2nd, 2005 in the field at the Bajio Experimental Station of National Research Institute for Forestry, Agriculture and Livestock (INIFAP) in Celaya, Guanajuato, Mexico (20o 34’ N, 100o 46’ W and 1750 masl). The soil is a Typic Pellusterts (pH 7.6, 1.8% organic matter and clay in texture) that during the previous season was planted with oat (Avena sativa) without the addition of chemical fertilizer. Plots 3.68 X 10 m were used in the experiment. Each plot contained four rows spaced 0.92 m apart. Plant stand for the yam bean species was of 170 000 plants ha-1 and 110 000 for the maize hybrid. Eighty kg ha-1 of P2O5 was applied to all treatments. Since there is abundant natural Rhyzobium and Bradyrhyzobium N2fixing bacteria in the experimental soil (Badillo and Castel lanos, 1997) , none inoculants were applied to the seed.
The weather was typical for the region, hot in spring and rainy during the summer (Figure 1). All plots were furrow-irrigated as needed with about 5 cm of water every month. In addition, during the growing season a total rainfall of 584 mm was registered near the experimental plots from June onwards.
After the onset of flowering, at 70 and 80 days after planting (DAP) in P. ahipa and P. erosus, respectively, flowers and young pods were removed from all plants in each plot every week to avoid the formation of pods and seeds. This is a common practice for yam bean production in the region since it allows for high tuber yields (Heredia, 1996). The removal of reproductive structures was suspended at 199 and 226 DAP for each species, respectively.

Biological nitrogen fixation and tuber yield of yam bean in central Mexico 279
Plant samples and harvest
In addition to the final harvest, during the growing cycle seven plant samples were taken in P. erosus and six in P. ahipa and in the maize crop at monthly intervals, starting at 67 DAP and finishing near physiological maturity. All samples were taken from 1.0 m2 per plot in eight replicates for the two yam beam species and in four replicates for the H 358 maize hybrid.
Plant samples were dissected to record leaf area, biomass components and number and weight of nodules. Samples were air dried first in a greenhouse and subsequently oven dried at 70 oC until constant weight was reached. After recording the weight in all plant samples, they were grounded and N content determined by the microkjeldhal method
(Bremmer and Mulvaney, 1982). The amount of nitrogen fixed by the two yam species was calculated by the difference method (Weaber, 1986) utilizing maize as the reference crop. With the tuber and total biomass and nitrogen content data, the harvest index (HI) and nitrogen harvest index (NHI) were calculated as follows: HI= tuber yield/total biomass (100) and NHI= tuber nitrogen content/biomass nitrogen content (100).
Tuber harvest of P. ahipa and H 358 was on October 18th and November 15th for P. erosus. For tuber yield determination, an area of 3.8 m2 was harvested per plot. Tuber production was recorded as fresh and dry weight.
RESULTS AND DISCUSION
Plant nodulation
In both species fixing nodules were present since the first sampling date (67 DAP) and its number and mass increased in subsequent samplings (Table 1). During the growing cycle, P. erosus reached the highest number of nodules at 170 DAP and its number was similar to the observed in previous reports (Tamez, 1987; Castellanos et al., 1997), while in P. ahipa it was observed at 123 DAP. P. ahipa had a higher number of nodules than P. erosus, but of smaller size, therefore, the dry weight of nodule mass was significantly higher (p>0.01) in P. erosus from 123 DAP onwards.
previous sampling dates, although there was some variation in this response across sampling dates. Since the reproductive structures were weekly removed, thus senescence was
Figure 1. Average maximum and minimum temperatures during the growing season in the Bajio Experimental Station, Celaya, Guanajuato.
Days after planting Nodules m2 Dry weigh g m2
P. ahipa P. erosus P. ahipa P. erosus67 20 20 0.28 0.2897 89 39 0.98 0.84123 326 230 1.99 7.42143 293 85 0.81 3.86170 (183)1 515 128 1.61 6.31199 (210) 144 146 2.08 8.24(226) - 86 - 5.82
35
30
25
20
15
10
5
0 April May June July August Sept. Oct. Nov.
Min Tem
Max Tem
Months of cultive
Tem
pera
ture
°C
Table 1. Number and dry weight of nodules per m2 of two yam bean species grown under irrigation plus rainfall conditions in Celaya, Guanajuato, Mexico.
1 (#)= number of DAP for P. erosus.
From the third and fifth sampling onwards, 123 and 170 DAP in P. erosus and P. ahipa, respectively, nodules were senescent, decreased in number and weighted less than in
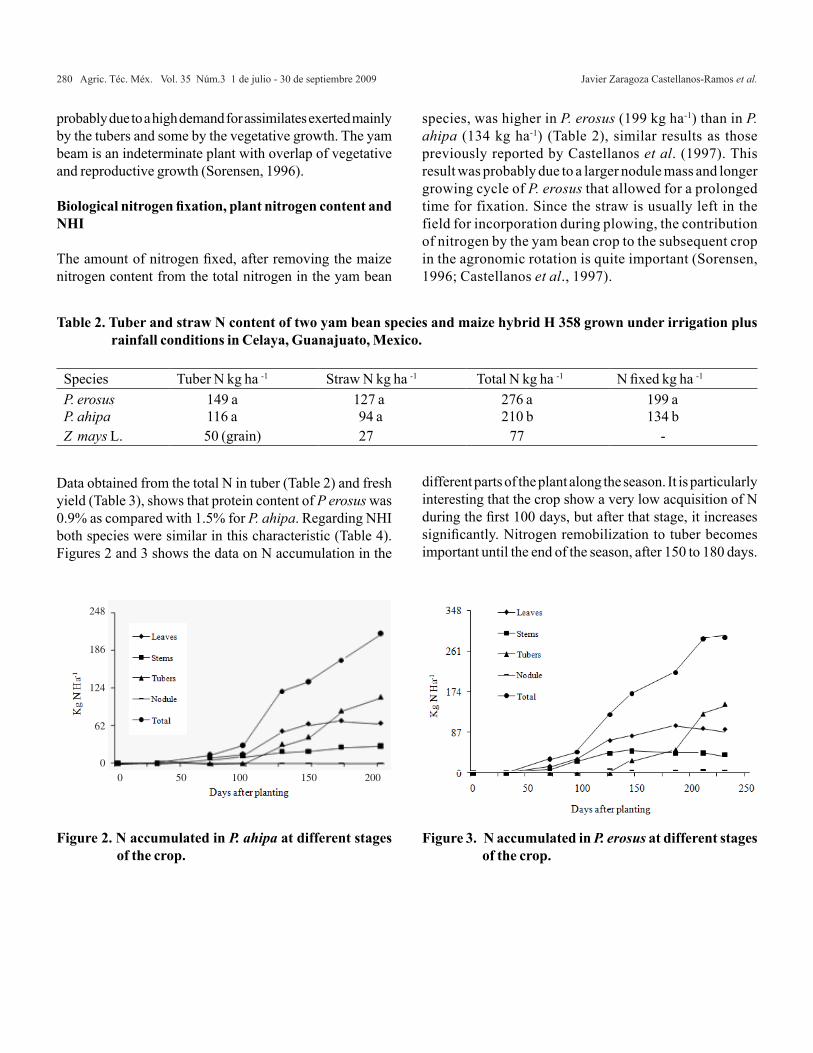
Javier Zaragoza Castellanos-Ramos et al.280 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
probably due to a high demand for assimilates exerted mainly by the tubers and some by the vegetative growth. The yam beam is an indeterminate plant with overlap of vegetative and reproductive growth (Sorensen, 1996).
Biological nitrogen fixation, plant nitrogen content and NHI
The amount of nitrogen fixed, after removing the maize nitrogen content from the total nitrogen in the yam bean
different parts of the plant along the season. It is particularly interesting that the crop show a very low acquisition of N during the first 100 days, but after that stage, it increases significantly. Nitrogen remobilization to tuber becomes important until the end of the season, after 150 to 180 days.
Figure 2. N accumulated in P. ahipa at different stages of the crop.
Figure 3. N accumulated in P. erosus at different stages of the crop.
Species Tuber N kg ha -1 Straw N kg ha -1 Total N kg ha -1 N fixed kg ha -1
P. erosus 149 a 127 a 276 a 199 aP. ahipa 116 a 94 a 210 b 134 bZ mays L. 50 (grain) 27 77 -
Table 2. Tuber and straw N content of two yam bean species and maize hybrid H 358 grown under irrigation plus rainfall conditions in Celaya, Guanajuato, Mexico.
species, was higher in P. erosus (199 kg ha-1) than in P. ahipa (134 kg ha-1) (Table 2), similar results as those previously reported by Castellanos et al. (1997). This result was probably due to a larger nodule mass and longer growing cycle of P. erosus that allowed for a prolonged time for fixation. Since the straw is usually left in the field for incorporation during plowing, the contribution of nitrogen by the yam bean crop to the subsequent crop in the agronomic rotation is quite important (Sorensen, 1996; Castellanos et al., 1997).
Data obtained from the total N in tuber (Table 2) and fresh yield (Table 3), shows that protein content of P erosus was 0.9% as compared with 1.5% for P. ahipa. Regarding NHI both species were similar in this characteristic (Table 4). Figures 2 and 3 shows the data on N accumulation in the
248
186
124
62
00 50 100 150 200

Biological nitrogen fixation and tuber yield of yam bean in central Mexico 281
was in part due to a larger growing cycle in P. erosus, which initiated the reproductive period ten days after P. ahipa (Figure 5). This difference in the duration of the vegetative period between the two species might be due to differences in photoperiod sensitivity, being P. ahipa less sensitive.
Species Yield Dry weight Above ground biomass Above ground/tuber biomass2
t ha-1
P. ahipa 48.3 b 9.00 a 2.75 b 0.305
P. erosus 105.3 a 7.96 a 4.33 a 0.544
Z mays L. 5.0 (grain) 4.33 10.44 -
Figure 4. Leaf area index of two yam bean genotypes during the growing cycle at Celaya, Guanajuato, Mexico.
a) b)
Table 3. Tuber yield1 and dry weight of two yam bean species and grain yield of maize grown under irrigation plus rainfall conditions in Celaya, Guanajuato, Mexico.
1 fresh weight; 2 on a dry weight basis.
Biomass and LAI
Early during the cycle, in a similar way as for biomass, the LAI was greater in P. ahipa and from 65 DAP onwards P. erosus displayed a significantly larger LAI (Figure 4). This
Figure 5. Biomass of plant parts along of the growing cycle of P. erosus a) y P. ahipa b) grown at Celaya, Guanajuato, Mexico.
a) b)a) b)a) b)
a) b)
5
4
3
2
1
00 50 100 150 200 250
Days after planting
LA
I
P. erosus
P. ahipa

Javier Zaragoza Castellanos-Ramos et al.282 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Tuber yield and HI
Tuber yield of P. erosus was significantly higher than that of P. ahipa (Table 4) and the dry matter was similar; thus the water content of P. erosus was higher and that may be the reason for its vernacular name ‘jicama de agua’ in Mexico, in contrast to other yam bean cultivars with less water called ‘jicama de leche’, P. ahipa belongs to the later vernacular classification. The higher water content of P. erosus makes it suitable for fresh consumption rather than processed (Sorensen, 1996). P. ahipa displayed a larger dry tuber weight and this was due to a higher concentration of solids in the tuber.
In contrast to the NHI that was similar between species, the tuber HI was significantly higher in P. erosus as compared to P. ahipa. In spite of its lower yield potential, the introduction of P. ahipa into central Mexico might have been due to other desirable traits, such as longer shelf life.
CONCLUSIONS
The dynamics of nitrogen fixation of the species P. ahipa and P. erosus was similar up to 170 DAP, from that date onwards the amount fixed was greater in P. erosus due to a longer growing cycle.
The number of nodules was larger in P. ahipa than in P. erosus, but the nodular mass was greater in the last species due to a larger nodule size.
Early during the cycle the LAI was greater in P. ahipa and from 65 DAP P. erosus displayed a significantly larger LAI. This was in part due to a longer growing cycle in P. erosus.
Fresh tuber yield in P. erosus was superior to P. ahipa; however, this last species displayed a larger dry tuber weight and this was due to a higher concentration of solids in the tuber.
The straw of the yam bean crop left in the field contained 127 kg of N ha-1 in P. erosus and 97 kg of N in P. ahipa. Therefore, this crop can be an excellent component in agronomic rotations.
ACKNOWLEDGEMENT
We appreciate the collaboration of Juan Pablo Tehuácatl for his helpful assistance during the preparation of the manuscript.
LITERATURE CITED
Badillo, V. y Castellanos, R. J. Z. 1996. Fijación simbiótica de nitrógeno bajo condiciones de campo en jícama (Pachyrhizus sp.). En resúmenes del II Simposio Internacional sobre Leguminosas Tuberosas. Celaya, Guanajuato, 5-8 de agosto. European Commission, ICTA, INIFAP, KVL y SAGAR. Guanajuato, México.
Bremmer, J. M. and Mulvaney, C. S. 1982. Nitrogen-total. In: Page, A. L.; Miller, R. H. and Keeney, D. R. (Eds.), 1982. Methods of soils analysis. Agronomy Núm. 9, Part 2, 2nd Edition.
Castellanos, R. J. Z.; Zapata, F.; Badillo, V.; Peña-Cabriales, J. J.; Jensen, E. S. and Heredia-García, E. 1997. Symbiotic nitrogen fixation and yield of Pachyrhizus erosus (L) Urban cultivars and Pachyrhizus ahipa (Wedd) Parodi landraces as affected by flower pruning. Soil Biol. and Biochem. 29(5-6):973-981.
Hafini, M. M. Grum, Stolen, O. and Sorensen, M. 1994. Biological nitrogen fixation in Pachyrhizus Rich. Ex DC. pp. 215-226 in proceedings of the first international symposium on tuberous legumes (Sorensen, M. ed.), Guadeloupe, FWI, April 21-24, 1992. Jordbrugsforlaget, Copenhagen, Denmark.
Heredia, Z. A. 1996. El cultivo de la jícama (Pachyrhizus erosus (L.) Urban) en la región Bajío en el Estado de Guanajuato, México. En resúmenes del II Simposio Internacional sobre Leguminosas Tuberosas Celaya, Guanajuato, 5-8 de agosto. European Commission, ICTA, INIFAP, KVL y SAGAR. Guanajuato, México.
Species Harvest index Nitrogen Harvest indexP. erosus 0.65 b 0.54 aP. ahipa 0.76 a 0.55 aZ. mays 0.48 0.65
Table 4. Tuber harvest index and nitrogen harvest index of two yam bean species and maize hybrid H 358 grown under irrigated plus rainfall conditions in Celaya, Guanajuato, Mexico.

Biological nitrogen fixation and tuber yield of yam bean in central Mexico 283
Lynd, J. Q. and Purcino, A. A. C. 1987. Effects of soil fertility on growth, tuber yield, nodulation and nitrogen fixation of yam bean (Pachyrhizus erosus (L.) Urban) grown on a Typic eutrustox. J. Plant Nutr. 10:485-500.
Sorensen, M. 1996. Yam bean (Pachyrhizus D C.). Promoting the conservation and use of underutilized and neglected crops. 2. Institute of plant genetics and crop plant research, Gatersleben/International Plant genetic Resources Institute, Rome.
Weaber, R.W. 1986. Measurement of biological dinitrogen fixation in the field. pp. 1-10, In: Hauck, R. D. and Weaber, R. D. (eds.), field measurement of dinitrogen fixation and denitrification. SSSA (Special Publication Number 18).
Tamez, G. P. 1987. Estudio sobre la simbiosis Rhizogium-Jicama (Pachyrhizus erosus (L.) (Urban). Tesis de Maestría en Ciencias. Centro de Investigaciones y Estudios Avanzados del Instituto Politécnico Nacional Unidad Irapuato, México.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 284-293
APTITUD COMBINATORIA Y EFECTOS RECÍPROCOS EN LÍNEAS ENDOGÁMICAS DE MAÍZ DE VALLES ALTOS DEL CENTRO DE MÉXICO*
COMBINIG ABILITY AND RECIPROCAL EFFECTS IN MAIZE INBRED LINES FROM THE HIGHLANDS OF CENTRAL MEXICO
Miguel Angel Avila Perches1§, Sergio Alfredo Rodríguez Herrera2, Mario Ernesto Vázquez Badillo2, Fernando Borrego Escalante2, Alejandro Javier Lozano del Río2 y Alfonso López Benítez2
1Campo Experimental Valle de Toluca, INIFAP. Km. 4.5 carretera Toluca-Zitácuaro, vialidad Adolfo López Mateos. 51350. Zinacantepec, México. Tel. y Fax 01 722 27 8 00 31. 2Departamento de Fitomejoramiento. Universidad Autónoma Agraria Antonio Narro. Buenavista, Saltillo, Coahuila. ([email protected]), ([email protected]), ([email protected]), ([email protected]), ([email protected]). §Autor para correspondencia: [email protected].
RESUMEN
El objetivo de este estudio fue determinar el tipo de acción génica involucrada y la influencia de los efectos recíprocos en el rendimiento y características agronómicas de cruzas dialélicas, directas y reciprocas, de ocho líneas endogámicas de maíz de Valles Altos del centro de México. Se evaluaron ocho progenitores y 56 híbridos simples en dos fechas de siembra, para determinar los efectos de aptitud combinatoria general, específica y efectos recíprocos, se utilizó el método I de Griffing. El estudio se llevó a cabo en Metepec, Estado de México, México en 2005. Los resultados mostraron diferencias entre genotipos y en los efectos de ACG y ACE para la mayoría de los caracteres evaluados. Los efectos genéticos no aditivos fueron de mayor importancia en rendimiento de grano, días a floración masculina y longitud de mazorca, en tanto que los efectos aditivos fueron de mayor relevancia en altura de planta. Los progenitores L1, L2 y L4 presentaron efectos positivos de ACG en rendimiento. El progenitor L4 junto con L3, L6 y L7 mostraron altos promedios en las características agronómicas estudiadas. Entre los híbridos de alto rendimiento, las cruzas 8 X 4, 4 X 1 y 2 X 4 mostraron efectos positivos de ACE; además, el último genotipo mostró la menor altura de planta. Los
* Recibido: Junio, 2008
Aceptado: Julio, 2009
efectos recíprocos fueron significativos en altura de planta y longitud de mazorca; no obstante, mostraron menores efectos que los de la ACG y ACE.
Palabras clave: Zea mays L., cruzas dialélicas, efectos maternos y no maternos, híbridos simples.
ABSTRACT
The aim of this study was to determine the type of gene action involved and the reciprocal effects upon seed yield and agronomic traits in eight inbred maize lines from the high valleys of central Mexico. Diallel crosses, direct and reciprocals were obtained with the eight lines. Eight parental lines and 56 simple crosses were evaluated in two planting dates in order to determine the general combining ability, specific combining ability and the reciprocal effects using Griffing’s method I. The research was carried out during 2005 at Metepec, Estado de Mexico, Mexico. Results show differences among genotypes and in GCA and SCA for most of the evaluated traits. Non-additive genetic effects were

Aptitud combinatoria y efectos recíprocos en líneas endogámicas de maíz de Valles Altos del centro de México 285
important for grain yield, days to anthesis and ear length, whereas the additive effects were significant for plant height. Parents L1, L2 and L4 showed positive effects in GCA for grain yield. Besides L4, parents L3, L6 and L7 showed high values for the agronomic traits considered. Crosses 8 X 4, 4 X 1 and 2 X 4 were among the high yielding hybrids that also showed high positive values of SCA; moreover, the last genotype registered the lowest plant height. The reciprocal effects were significant for plant height and ear length, but of small magnitude than the GCA and SCA parameters.
Key words: Zea mays L. diallel crosses, maternal and non-maternal effects, simple hybrids.
INTRODUCCIÓN
En los Valles Altos del centro de México, situados a más de 2 200 m se siembran 1.5 millones de ha con maíz, lo cual representa 20% de la superficie nacional. De esta superficie, aproximadamente 50% (750 000 ha) se cultivan bajo condiciones de punta de riego y buen temporal en las que se obtienen altos rendimientos; no obstante, el rendimiento promedio regional es menor a cuatro t ha-1; por lo anterior, el uso generalizado de semilla de variedades mejoradas es una alternativa viable para incrementar la rentabilidad del cultivo a corto plazo, por lo que es necesaria la formación de nuevos genotipos superiores a los actuales.
El método de cruzas dialélicas permite estimar la ACG y ACE y se considera eficaz para detectar fuentes de germoplasma útiles para el mejoramiento genético del maíz. El apareamiento de cruzamientos dialélicos es útil para la evaluación de componentes genéticos en la variación del rendimiento de los progenitores y para calcular la capacidad productiva de sus cruzas (Baker, 1978).
La ACG se determina el desempeño promedio de una línea en sus combinaciones híbridas, mientras que, la ACE separa las combinaciones híbridas específicas que resulten mejor o peor de lo que se esperaría en relación con la media de la ACG de las dos líneas progenitoras (Sprague y Tatum, 1942). Con relación al tipo de acción génica que determina la aptitud combinatoria de las líneas, se considera que la ACG determina la porción aditiva de los efectos genéticos, en tanto que la ACE, los efectos no aditivos, esto es, la acción génica de dominancia y epistasis (Poehlman y Allen, 2003). Griffing (1956) propuso cuatro métodos para el análisis dialélico los cuales son de uso más frecuente para estimar
los efectos de ACG y ACE, así como para el estudio genético de poblaciones biológicas y la comprensión de la acción génica en caracteres cuantitativos de importancia agrícola; asimismo, suministra al fitomejorador, las herramientas necesarias para la adecuada aplicación de los planes de mejoramiento a emplear (Hallauer y Miranda, 1981). En México, varios fitomejoradores han utilizado algunos de los diseños dialélicos de Griffing, como son los casos de Antuna et al. (2003), De la Cruz et al. (2003) y Reyes et al. (2004) y específicamente en los Valles Altos del centro de México, Pérez et al. (1991), Vasal et al. (1995) y Balderrama et al. (1997) quienes detectaron mayor efecto de la ACG en la manifestación de los caracteres estudiados.
La genética de características ha sido utilizada para el mejoramiento de numerosas especies vegetales, en la mayoría de los estudios se ha resaltado la importancia de los efectos de ACG y ACE e ignoran los factores maternales y recíprocos; es decir, se acepta que existe diferencia en el desempeño de cruzas directas y recíprocas (Mosjidis et al., 1989); sin embargo, en algunas investigaciones se han mostrado valores inconsistentes (en diferentes ambientes) y más pequeños que los obtenidos para ACG o ACE (Velázquez et al., 1992). En algunos casos, el efecto recíproco (ER) ha mostrado ser un factor relevante y de cierta consistencia, por lo que puede tener implicaciones en el mejoramiento genético del maíz (Hansen y Bagget, 1977). La diversidad genética de los progenitores es un aspecto importante que influye en la expresión de los ER en varios caracteres del maíz (Kalsy y Sharma, 1972; Khehra y Bhalla, 1976). Zhang y Kang (1997) y Kang et al. (1999) reportaron que la división del ER en efectos maternos (EMAT) y no maternos (ENMAT) en el análisis dialélico, es útil para determinar si factores maternales o extranucleares están involucrados en la expresión de un carácter. El objetivo de esta investigación fue determinar el tipo de acción génica involucrada, así como la influencia de los efectos recíprocos, en la expresión del rendimiento y algunas características agronómicas de un grupo de líneas endogámicas y sus combinaciones híbridas.
MATERIALES Y MÉTODOS
La investigación se realizó en Metepec, Estado de México, en el Campo Experimental Valle de Toluca del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) ubicado a 2 650 m, con clima templado subhúmedo con lluvias en verano, precipitación media anual de 805.4 mm y temperatura media anual de 14.4 ºC.

Miguel Angel Avila Perches et al.286 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
el período de abril a octubre de 2005, la precipitación fue de 820 m, la temperatura media de 14.8 ºC y las primeras heladas (-4 °C) ocurrieron el 21 de octubre.
Se utilizó el diseño experimental látice triple 8 X 8, la parcela experimental y útil fue de dos surcos 5 m de largo y 0.80 m de separación. La densidad de población fue de 65 000 plantas por ha-1, el manejo agronómico se realizó con base en las recomendaciones generales del INIFAP para el cultivo de maíz en el valle de Toluca. Se determinaron las siguientes características: días a floración masculina, altura de planta, longitud de mazorca y rendimiento de grano en t ha-1 al 14% de humedad.
En el análisis de la eficiencia del diseño experimental utilizado se observó que fue de 5 a 10% para cada ambiente, por lo que las características agronómicas fueron analizadas como bloques al azar en forma independiente para cada fecha de siembra y se efectuó el análisis combinado. Para la estimación de la ACG, ACE y ER se empleó el análisis dialélico de Griffing (1956) mediante el método I y modelo I, en el que se examinan las líneas parentales, las cruzas F1 directas y recíprocas, para lo cual se utilizó el programa DIALLEL-SAS05 propuesto por Zhang et al. (2005), que permite la división de los ER en maternos (MAT) y no maternos (NMAT).
RESULTADOS Y DISCUSIÓN
El análisis de varianza combinado de los dos experimentos mostró diferencias altamente significativas para los factores ambiente (AMB), genotipo (GEN) y para la interacción GEN X AMB (Cuadro 2). Lo anterior significa que el ambiente (determinado por la fecha de siembra) tuvo efecto sobre los genotipos y las características estudiadas. Lo cual coincide con de la Cruz et al. (2003) en el sentido de que el ambiente modifica las características agronómicas, por lo que es conveniente establecer los ensayos en diferentes ambientales para estimar el valor de los componentes genéticos y separar el efecto del GEN y del AMB.
Se incluyeron ocho líneas endogámicas, siete de estas fueron desarrolladas por el INIFAP, con entre tres y cinco generaciones de autofecundación; además, se utilizó una línea del Centro Internacional de Mejoramiento de Maíz y Trigo (CIMMYT) con nivel de endogamia S7. La composición de las poblaciones 85 y 87 (Pob 85 y Pob 87) contienen 60% de germoplasma de Valles Altos, 20% de clima templado y 20% de subtropical-tropical (Eagles y Lothrop, 1994), mientras que el sintético intermedio “B” (SIB) de INIFAP se formó con líneas selectas de Valles Altos (Cuadro 1).
Durante el ciclo primavera-verano de 2004, se realizaron las cruzas dialélicas, directas y recíprocas de las ocho líneas, de las que se generaron 56 cruzas F1; además, se obtuvo semilla de los progenitores por medio de cruzas fraternales. Los 64 genotipos se evaluaron en dos experimentos sembrados el 2 de abril y 6 de mayo de 2005; el primero, se estableció con la aplicación de un riego de presiembra que aportó la humedad necesaria para la germinación y emergencia, el resto del ciclo se mantuvo bajo condiciones de temporal; el segundo, se sembró bajo condiciones de temporal. Cada fecha de siembra (FS) se consideró como un ambiente diferente (AMB) de evaluación. Durante
Línea GenealogíaL1 Mich21-181-14-1-16-5*L2 Mich21CI-7-2-14-5-2*L3 VS102-81-4-4-3-2*L4 Pob87SIB/1-4-3-3*L5 Pob85SIB/1-8-3-2*L6 Tlax151SFC2-77-2-1-2*L7 Tlax151SFC2-11-2-2-3*L8 B8785MH10-1-1-2TL-1-3TL-3-1T
(CML-242)**
Fuente de variación Rendimiento de grano Días a floración masculina Altura de planta Longitud de mazorcaAmbiente (AMB) 152701682.00** 71.76** 75488.16** 60.16**Genotipo (GEN) 24893432.00** 68.21** 4343.76** 11.76**GEN X AMB 2050367.00** 30.18** 372.01** 3.12**
Cuadro 1. Líneas endogámicas de maíz de Valles Altos del centro de México utilizadas como progenitores.
*línea de INIFAP; **línea de CIMMYT.
Cuadro 2. Cuadrados medios y significancia del análisis de varianza combinado de las características agronómicas evaluadas.
** altamente significativo al 0.01.
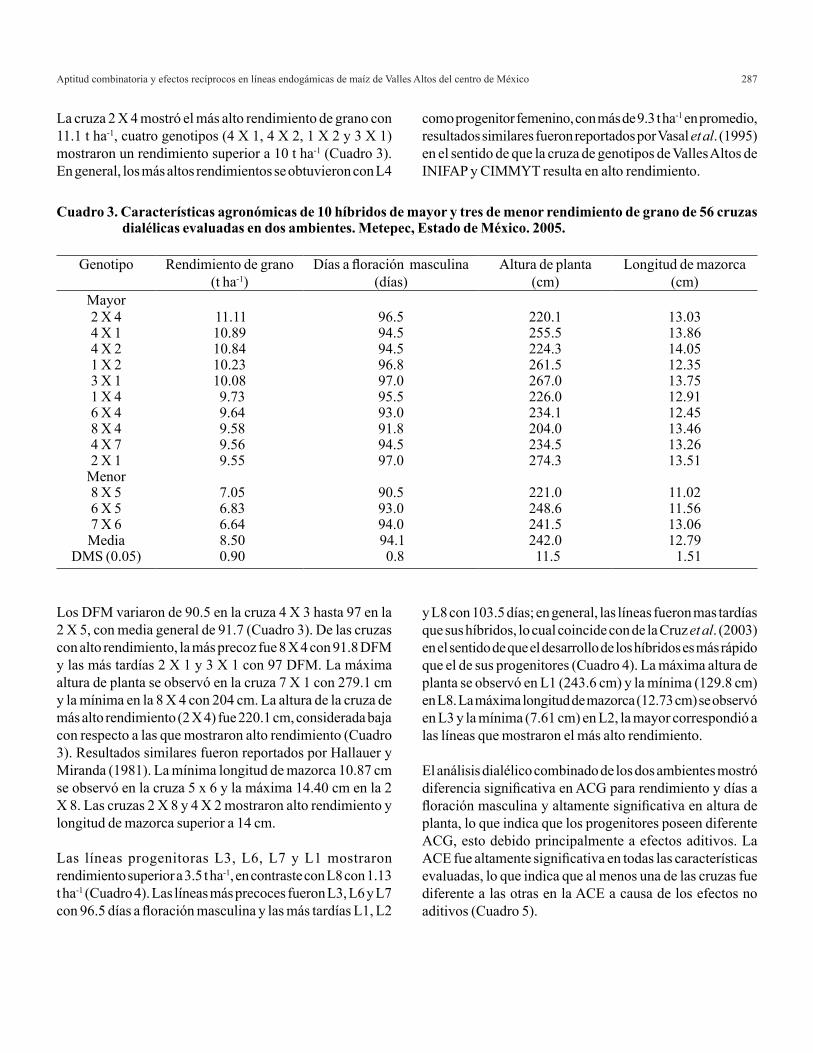
Aptitud combinatoria y efectos recíprocos en líneas endogámicas de maíz de Valles Altos del centro de México 287
como progenitor femenino, con más de 9.3 t ha-1 en promedio, resultados similares fueron reportados por Vasal et al. (1995) en el sentido de que la cruza de genotipos de Valles Altos de INIFAP y CIMMYT resulta en alto rendimiento.
y L8 con 103.5 días; en general, las líneas fueron mas tardías que sus híbridos, lo cual coincide con de la Cruz et al. (2003) en el sentido de que el desarrollo de los híbridos es más rápido que el de sus progenitores (Cuadro 4). La máxima altura de planta se observó en L1 (243.6 cm) y la mínima (129.8 cm) en L8. La máxima longitud de mazorca (12.73 cm) se observó en L3 y la mínima (7.61 cm) en L2, la mayor correspondió a las líneas que mostraron el más alto rendimiento.
El análisis dialélico combinado de los dos ambientes mostró diferencia significativa en ACG para rendimiento y días a floración masculina y altamente significativa en altura de planta, lo que indica que los progenitores poseen diferente ACG, esto debido principalmente a efectos aditivos. La ACE fue altamente significativa en todas las características evaluadas, lo que indica que al menos una de las cruzas fue diferente a las otras en la ACE a causa de los efectos no aditivos (Cuadro 5).
La cruza 2 X 4 mostró el más alto rendimiento de grano con 11.1 t ha-1, cuatro genotipos (4 X 1, 4 X 2, 1 X 2 y 3 X 1) mostraron un rendimiento superior a 10 t ha-1 (Cuadro 3). En general, los más altos rendimientos se obtuvieron con L4
Genotipo Rendimiento de grano(t ha-1)
Días a floración masculina(días)
Altura de planta(cm)
Longitud de mazorca(cm)
Mayor2 X 4 11.11 96.5 220.1 13.034 X 1 10.89 94.5 255.5 13.864 X 2 10.84 94.5 224.3 14.051 X 2 10.23 96.8 261.5 12.353 X 1 10.08 97.0 267.0 13.751 X 4 9.73 95.5 226.0 12.916 X 4 9.64 93.0 234.1 12.458 X 4 9.58 91.8 204.0 13.464 X 7 9.56 94.5 234.5 13.262 X 1 9.55 97.0 274.3 13.51
Menor8 X 5 7.05 90.5 221.0 11.026 X 5 6.83 93.0 248.6 11.567 X 6 6.64 94.0 241.5 13.06Media 8.50 94.1 242.0 12.79
DMS (0.05) 0.90 0.8 11.5 1.51
Cuadro 3. Características agronómicas de 10 híbridos de mayor y tres de menor rendimiento de grano de 56 cruzas dialélicas evaluadas en dos ambientes. Metepec, Estado de México. 2005.
Los DFM variaron de 90.5 en la cruza 4 X 3 hasta 97 en la 2 X 5, con media general de 91.7 (Cuadro 3). De las cruzas con alto rendimiento, la más precoz fue 8 X 4 con 91.8 DFM y las más tardías 2 X 1 y 3 X 1 con 97 DFM. La máxima altura de planta se observó en la cruza 7 X 1 con 279.1 cm y la mínima en la 8 X 4 con 204 cm. La altura de la cruza de más alto rendimiento (2 X 4) fue 220.1 cm, considerada baja con respecto a las que mostraron alto rendimiento (Cuadro 3). Resultados similares fueron reportados por Hallauer y Miranda (1981). La mínima longitud de mazorca 10.87 cm se observó en la cruza 5 x 6 y la máxima 14.40 cm en la 2 X 8. Las cruzas 2 X 8 y 4 X 2 mostraron alto rendimiento y longitud de mazorca superior a 14 cm.
Las líneas progenitoras L3, L6, L7 y L1 mostraron rendimiento superior a 3.5 t ha-1, en contraste con L8 con 1.13 t ha-1 (Cuadro 4). Las líneas más precoces fueron L3, L6 y L7 con 96.5 días a floración masculina y las más tardías L1, L2

Miguel Angel Avila Perches et al.288 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
(1999) señalan que el componente no materno (recíproco específico) es la interacción entre factores extranucleares y nucleares en las cruzas.
La interacción ACG X AMB fue significativa para altura de planta y altamente significativa para rendimiento y longitud de mazorca. La interacción ACE X AMB fue significativa para días a floración masculina y altamente significativa para rendimiento y altura de planta; lo anterior, resalta la importancia de repetir los experimentos bajo diferentes ambientes para obtener estimaciones consistentes, Vasal et al. (1995). Asimismo, el factor ER X AMB fue significativo en días a floración masculina y altamente significativo en rendimiento. Mientras que EMAT X AMB fue sólo significativo en días a floración masculina y altura de planta;
Línea Rendimiento de grano(t ha-1)
Días a floración masculina (días)
Altura de planta(cm)
Longitud de mazorca(cm)
L3 4.50 96.5 236.8 12.73L6 4.40 96.5 232.0 12.43L7 4.22 96.5 229.6 12.21L1 3.69 103.5 243.6 11.88L4 2.95 100.0 156.3 9.61L5 2.41 100.0 185.3 7.70L2 2.39 103.5 164.1 7.61L8 1.13 103.5 129.8 8.43
Media 3.21 100.0 197.2 10.32DMS (0.05) 0.90 0.8 11.5 1.51
Fuente de variación Rendimiento de grano Días a floración masculina Altura de planta Longitud de mazorca
GEN 1568286205.00** 4297.23** 273656.90** 741.11**ACG 166650969.00* 974.22* 138411.56** 209.35ACE 1365005524.70** 2498.84** 120973.59** 438.58**ER 36629711.10 824.16 14271.75* 93.17*EMAT 11992193.70* 315.62 4150.87 17.37ENMAT 24637517.30 508.54 10120.87* 75.79**GEN X AMB 129173100.00** 1901.90** 23437.16** 197.07**ACG X AMB 38605811.8** 252.27 3385.20* 101.15**ACE X AMB 48001075.20** 819.46* 12220.87** 57.13ER X AMB 42566212.60** 830.16* 7831.08 38.78EMAT X AMB 2882371.50 311.75* 3265.41* 12.37ENMAT X AMB 39683841.10** 518.41 4565.66 26.41
Cuadro 4. Características agronómicas de ocho líneas endogámicas progenitoras de cruzas dialélicas evaluadas en dos ambientes. Metepec, Estado de México. 2005.
Cuadro 5. Suma de cuadrados y significancia del análisis dialélico combinado para características agronómicas.
*significativo al 0.05; **altamente significativo al 0.01; GEN= genotipo; ACG= aptitud combinatoria general; ACE= aptitud combinatoria específica; ER= efecto recíproco; EMAT= efecto materno; ENMAT= efecto no materno; AMB= ambiente.
Los ER fueron significativos en altura de planta y longitud de mazorca, lo que permite inferir que para estas características el valor que se obtiene de la cruza (i, j) es diferente al de la cruza inversa (j, i); Hansen y Bagget (1977), reportaron diferencias recíprocas en altura de planta y longitud de mazorca. Los efectos maternos fueron significativos sólo en rendimiento, en este sentido, Velázquez et al. (1992) observaron EMAT para rendimiento sólo en una de tres localidades (Valles Altos del centro de México). Los efectos maternos fueron significativos y altamente significativos en altura de planta y longitud de mazorca, respectivamente (Cuadro 5). Es importante mencionar, que de acuerdo con Zhang y Kang (1997) el factor materno (recíproco general) se refiere al efecto del genotipo o tejido materno sobre alguna característica de su descendencia, mientras que, Kang et al.

Aptitud combinatoria y efectos recíprocos en líneas endogámicas de maíz de Valles Altos del centro de México 289
además, ENMAT X AMB fue altamente significativo en rendimiento (Cuadro 5); lo anterior, permite inferir que para estas características, la manifestación del efecto recíproco, materno y no materno dependen del ambiente, lo que coincide con lo reportado por Kang et al. (1999).
Al desglosar las cruzas en ACG y ACE y no obstante de haber mostrado las diferencias significativas mencionadas (Cuadro 5), se determinó que la ACE contribuyó con más de 80% a la suma de cuadrados de genotipos en rendimiento y con más de 58% en días a floración masculina y longitud de mazorca. La importancia de los efectos no aditivos en estas características coincide con de la Cruz et al. (2003); sin embargo, es contrario a lo reportado por Pérez et al. (1991) y podría deberse a que estos últimos autores utilizaron variedades criollas de Valles Aaltos. La ACG contribuyó con 51% en altura de planta, la relevancia de los efectos aditivos en altura de planta también fue reportada por Antuna et al. (2003).
Los ER fueron significativos en altura de planta y longitud de mazorca; en ambos casos, se debió a los efectos no maternos ya que estos fueron significativos en altura de planta y altamente significativos en longitud de mazorca. En general, los ER mostraron valores bajos y aún más bajos que
los observados para ACG y ACE los que sólo contribuyeron con 10% en promedio en el desglose de la suma de cuadrados de genotipos del análisis dialélico en las variables evaluadas (Cuadro5), lo anterior coincide con lo observado por Hansen y Bagget (1977).
La estimación de los efectos de ACG de las líneas endogámicas mostraron que en rendimiento, los valores positivos más altos fueron para la las líneas L4, L1 y L2 con 0.766, 0.443 y 0.378 respectivamente, esto sugiere que estos materiales contienen genes de efectos aditivos que se expresan favorablemente en el rendimiento de grano (Cuadro 6). En contraste, L5, L6, L7 y L8 mostraron valores negativos, lo anterior coincide con Velázquez et al. (1992) que también estimó ACG positivas en líneas derivadas de Mich. 21 y Pérez et al. (1991) quienes reportaron valores altos de ACG en la población Mich. 21 (Cuadro 6). En días a floración masculina, L5 obtuvo el efecto negativo más alto (-1.244 días) y L2 el más alto efecto positivo con 2.244 días. La línea L1 mostró el efecto positivo más alto en altura de planta con 23.567 cm y L4 el más negativo con -16.734 cm. El análisis dialélico combinado no se observó diferencia para ACG en longitud de mazorca (Cuadro 5), por lo que no se presentan estos efectos en el Cuadro 6.
El efecto negativo más alto de ACE para días a floración masculina se observó en la cruza 8 X 4 (-10.30), lo que resulta favorable ya que podría aportar precocidad y rendimiento de grano (9.58 t ha-1); no obstante, también mostró efecto positivo alto en altura de planta (47.35 cm ), lo cual demerita esta característica. Otras cruzas que mostraron efectos positivos en altura de planta fueron 1 X 2 y 4 X 7, (Cuadro 7). Las cruzas 2 X 8 y 8 X 4 mostraron los e f e c t o s p o s i t i v o s d e A C E m á s a l t o s p a r a longi tud de mazorca con 5 .383 y 3 .695 cm,
Línea Rendimiento de grano Días a floración masculina Altura de planta
L1 0.443** 0.984* 23.567**L2 0.378** 2.244** -5.182*L3 0.058 -1.151** 10.109**L4 0.766** -0.901* -17.630**L5 -0.708** -1.244** -8.151**L6 -0.295** -0.307 7.932**L7 -0.260** -0.192 6.088**L8 -0.383** 0.567 -16.734**
Cuadro 6. Estimación de los efectos de ACG de las ocho líneas endogámicas progenitoras de las cruzas dialélicas.
*, ** diferentes de cero al 0.05 y 0.01, respectivamente.
La estimación de los efectos de ACE de las cruzas dialélicas en rendimiento mostraron el más alto efecto positivo en 2 X 8 y 8 X 4 con 7.52 y 7.22 t ha-1, respectivamente (Cuadro 7). Se observó que en siete de las cruzas de más alto rendimiento mostraron efectos positivos superiores a 1.0 t ha-1, lo que coincide parcialmente con lo reportado por Reyes et al. (2004) en el sentido de que una cruza simple será de alto rendimiento si sus dos líneas progenitoras son de alta ACG o si el efecto de ACE es alto y al menos uno de sus progenitores es de alta ACG.

Miguel Angel Avila Perches et al.290 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
ER negativos de -14.7 y -22.4 cm, respectivamente (Cuadro 8); asimismo, los mismos genotipos mostraron los mayores ENMAT negativos con -10.5 y -18.0 cm, respectivamente, por lo que utilizados en ese sentido contribuyen al menor porte de planta; por el contrario, si se utilizan las cruzas recíprocas (4 X 1 y 8 X 7), los ER serían los mismos pero positivos y en este caso incrementarían la altura de planta.
negativos en estos dos factores con -1.03 y -0.68 cm, respectivamente; es decir, que al utilizar el híbrido 2 X 8 se incrementa la longitud de mazorca; en la cruza recíproca (8 X 2) se obtendría mismo efecto de ER y ENMAT pero negativo y aportaría una reducción en longitud de mazorca (Cuadro 9).
respectivamente. Entre los híbridos con alto rendimiento, 2 X 4 mostró efecto positivo de ACE en longitud demazorca de 1.189 cm.
En la característica altura de planta, el análisis dialélico mostró diferencia significativa en ER, lo cual se debió principalmente a los ENMAT, los híbridos 1 X 4 y 7 X 8 mostraron
Línea LíneaL1 L2 L3 L4 L5 L6 L7 L8
L1 -6.4 5.5 -14.7* 0.4 -7.1 -3.3 -1.5L2 -2.0 -1.9 -2.0 1.3 3.8 -1.5 1.3L3 6.3 -5.3 -3.9 -6.5 -6.9 -2.3 2.7L4 -10.5* -2.1 -0.5 -9.3 -3.2 -2.3 0.5L5 6.1 2.7 -1.5 -7.7 -1.5 6.5 -0.1L6 -0.9 5.7 -1.5 -1.2 -1.1 1.7 5.7L7 -2.5 -5.1 -2.4 -5.7 1.5 -3.7 -22.4**L8 3.5 2.1 7.0 1.5 -0.8 4.6 -18.0**
Cruza Rendimiento de grano(t ha-1)
Días a floración masculina(días)
Altura de planta (cm)
Longitud de mazorca(cm)
Mayor2 X 4 1.98** -0.57 9.13 1.189**4 X 1 1.26** 0.18 -1.11 0.4184 X 2 1.98** -0.57 9.13 1.189**1 X 2 1.23** -1.04 13.59* -0.0603 X 1 1.07** 2.18* 2.89 0.5571 X 4 1.26** 0.18 -1.11 0.4186 X 4 1.16** 0.22 4.68 -0.3798 X 4 7.22** -10.30** 47.35** 3.695**4 X 7 0.91** 0.10 12.44* 0.3192 X 1 1.23** -1.04 13.59* -0.060
Menor8 X 5 4.50** 9.31** 44.08** 2.601**6 X 5 0.18 0.56 11.36* -0.1687 X 6 -0.24 0.09 -6.70 -0.361
Cuadro 7. Estimación del efecto de ACE de los 10 híbridos de mayor y los tres de menor rendimiento de grano evaluadas en dos ambientes.
*, ** diferente de cero al 0.05 y 0.01 de probabilidad, respectivamente
*, ** diferente de cero al 0.05 y 0.01 de probabilidad, respectivamente; efectos recíprocos sobre la diagonal; efectos no maternos bajo la diagonal.
Cuadro 8. Estimación de los efectos recíprocos y no maternos para altura de planta.
En longitud de mazorca el análisis dialélico detectó diferencias en ER, lo cual se debió a la alta significancia de los ENMAT, se observó que los mas altos ER y ENMAT positivos se presentaron en el genotipo 2 X 8 con 1.38 y 0.97 cm, respectivamente; además, la cruza 3 X 5 mostró efectos

Aptitud combinatoria y efectos recíprocos en líneas endogámicas de maíz de Valles Altos del centro de México 291
LITERATURA CITADA
Antuna, G. O.; Rincón, S. F.; Gutiérrez del R. E.; Ruiz, T. N. A. y Bustamante, G. L. 2003. Componentes genéticos de caracteres agronómicos y de calidad fisiológica de semillas en líneas de maíz. Rev. Fitotec. Mex. 26:11-17.
Baker, R. J. 1978. Issues in diallel analysis. Crop Sci. 18:533-536.
Balderrama, C. S.; Mejía, C. A.; Castillo, G. F. y Carballo, C. A. 1997. Efectos de aptitud combinatoria en poblaciones de maíz nativas de Valles Altos de México. Rev. Fitotec. Mex. 20:137-147.
de la Cruz, L. E., Gutiérrez del R., E.; Palomo, G. A. y Rodríguez, H. S. 2003. Aptitud combinatoria y heterosis de líneas de maíz en la Comarca Lagunera. Rev. Fitotec. Mex. 26:279-284.
Eagles, H. A. and Lothrop, J. E. 1994. Highland maize from Central Mexico. Its origin, characteristics and use in breeding programs. Crop Sci. 34:11-19.
Griffing, B. 1956. Concept of general and specific combining ability in relation to diallel crossing systems. Aust. J. Biol. Sci. 9:463-493.
Hallauer, A. R. and Miranda, F. J. B. 1981. Quantitative genetics in maize breeding. Iowa State University Press. Ames, Iowa. USA.
Hansen, L. A. and Bagget, J. R. 1977. Reciprocal differences for plant and ear characteristics in sweet corn. Hort. Sci. 12:60-62.
Kalsy, H. S. and Sharma, D. 1972. Study of cytoplasmic effects in reciprocal crosses of divergent varieties of maize (Zea mays L.). Euphytica 21:527-533.
Kang, M. S.; Kushairi, D. A.; Zhang, Y. and Magari, R. 1999. Combining ability for rind puncture resistance in maize. Crop Sci. 39:368-371.
Línea LíneaL1 L2 L3 L4 L5 L6 L7 L8
L1 -0.65 0.12 -0.48 -0.23 -0.09 0.53 0.36L2 -0.41 -0.66 -0.50 -0.01 0.59 0.03 1.38**L3 0.10 -0.92** 0.38 -1.03** 0.23 -1.02 0.33L4 -0.41 -0.67* 0.46 -0.24 -0.22 0.11 -0.15L5 0.10 0.08 - 0.68* 0.02 -0.34 0.56 0.49L6 -0.12 0.31 0.21 -0.32 -0.71* -0.40 -0.15L7 0.55 -0.18 -0.98** 0.06 0.25 -0.34 -0.45L8 0.19 0.97** 0.17 -0.39 -0.01 -0.28 -0.64
Cuadro 9. Estimación de los efectos recíprocos y no maternos para longitud de mazorca.
*, ** diferente de cero al 0.05 y 0.01 de probabilidad, respectivamente; efectos recíprocos sobre la diagonal; efectos no maternos bajo la diagonal.
En los híbridos que mostraron valores positivos y negativos significativos de los ER y de los ENMAT en altura de planta y longitud de mazorca y en la mayoría de las cruzas (1 X 4, 7 X 8, 2 X 8, 3 X 5, 4 X 8, 5 X 8 y 1 X 8), intervinieron como progenitores femeninos o masculinos las líneas 4, 5 y 8 las cuales fueron formadas con la integración de germoplasma de Valles Altos, subtrópico y trópico; además, como contrapartes participaron las líneas L1, L2, L3 y L7 de Valles Altos, lo que coincide parcialmente con Kalsy y Sharma (1972) y con Khehra y Bhalla (1976) en el sentido de que la diversidad genética y el diferente origen geográfico de los progenitores influyen en la manifestación de los efectos recíprocos y/o citoplásmicos.
CONCLUSIONES
En las líneas endogámicas de Valles Altos estudiadas, los efectos genéticos no aditivos son relevantes en las características de rendimiento de grano, días a floración masculina y longitud de mazorca en tanto que los efectos aditivos son para altura de planta.
Los progenitores L1, L2 y L4 mostraron los más altos efectos positivos de ACG en rendimiento de grano y sobresalen las cruzas de 8 X 4, 4 X 1 y 2 X 4 con valores altos positivos de ACE
El híbrido 2 X 4 destaca por las características de rendimiento, altura de planta y longitud de mazorca; asimismo, las líneas endogámicas L3, L6, L7 y L1 mostraron los valores más altos en las características evaluadas.
Los ER observados fueron significativos en altura de planta y longitud de mazorca; sin embargo, presentaron valores menores a los observados en ACG y ACE.

Miguel Angel Avila Perches et al.292 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Khehra, A. S. and Bhalla, S. K. 1976. Cytoplasmic effects on quantitative characters in maize (Zea mays L.). Theor. Appl. Genet. 47:271-274.
Mosjidis, J. A.; Waines, J. G.; Yermanos, D. M. and Rosielle, A. A. 1989. Methods for the study of cytoplasmic effects on quantitative traits. Theor. Appl. Genet. 77:195-199.
Pérez, T. R. A.; Carballo, C. A.; Castillo, G. F. y Covarrubias, P. J. 1991. Identificación de patrones heteróticos en un grupo de variedades precoces de maíz. Agrociencia serie Fitociencia 2:69-79.
Poehlman, J. M. y Allen, D. 2003. Mejoramiento genético de las cosechas. Guzmán, O. M.; Hernández, C. M. A. y Serrano, C. L. M. (Trad.). Editorial Limusa. México.
Reyes, L. D.; Molina, G. J. D.; Oropeza, R. M. A. y Moreno, P. E. C. 2004. Cruzas dialélicas entre líneas autofecundadas de maíz derivadas de la raza tuxpeño. Rev. Fitotec. Mex. 27:49-56.
Sprague, G. F. and Tatum, L. A. 1942. General vs specific combining ability in single crosses of corn. J. Am. Soc. Agron. 34:923-932.
Vasal, S. K.; Srinivasan, G.; Vergara, A. N. y González, C. F. 1995. Heterosis y aptitud combinatoria en germoplasma de maíz de Valles Altos. Rev. Fitotec. Mex. 18:123-139.
Velázquez, C. G. A.; Castillo, G. F.; Molina, G. J. D. y Arellano, V. J. L. 1992. Aptitud combinatoria y efectos recíprocos en líneas de maíz (Zea mays L.) con diferente nivel de endogamia. Agrociencia serie Fitociencia 3:111-123.
Zhang, Y. and Kang, M. S. 1997. DIALLEL-SAS: A SAS program for Griffing’s diallel analyses. Agron. J. 89:176-182.
Zhang, Y.; Kang, M. S. and Lamkey, K. R. 2005. DIALLEL-SAS05: A comprehensive program for Griffing’s and Gardner-Eberhart analyses. Agron. J. 97:1097-1106.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 293-302
RENDIMIENTO DE GRANO Y FORRAJE EN HÍBRIDOS DE MAÍZ Y SU GENERACIÓN AVANZADA F2
*
GRAIN AND FORAGE YIELD IN MAIZE HYBRIDS AND THEIR ADVANCED F2 GENERATION
Rodolfo Gaytán-Bautista1, Ma. Isabel Martínez-Gómez1 y Netzahualcóyotl Mayek-Pérez2§
1Campo Experimental Pabellón, INIFAP. Carretera Aguascalientes-Zacatecas km 32.5, Pabellón de Arteaga, C. P. 20671, Aguascalientes. México, ([email protected]).2Centro de Biotecnología Genómica- Instituto Politécnico Nacional. Blvd. del Maestro esq. Elías Piña s/n, colonia Narciso Mendoza, C. P. 88710, Reynosa, Tamaulipas, México. §Autor para correspondencia: [email protected].
RESUMEN
Ante el alto costo de la semilla de híbridos de maíz, los productores a pequeña escala usan semilla en generaciones F2 y F3. El objetivo de este trabajo fue determinar el efecto de utilizar semilla autofecundada de híbridos sobre el rendimiento de grano y forraje seco de híbridos de maíz. El estudio se estableció bajo condiciones de riego en Tepezalá y Pabellón de Arteaga, Aguascalientes y Calera de Víctor Rosales, Zacatecas, México en el ciclo 2001. Se sembraron 22 híbridos comerciales y su respectiva generación avanzada F2. Se determinó rendimiento de grano y forraje, altura de planta, días a floración masculina y femenina y altura de mazorca. Se utilizó un arreglo en parcelas subdivididas con diseño en bloques completamente al azar y tres repeticiones. Se efectuó el análisis conjunto de la relación beneficio/costo. En general, usar semilla F2 redujo 22% el rendimiento de grano y 8% de forraje seco. La mayoría de los híbridos y sus generaciones avanzadas mostraron estabilidad en el rendimiento de grano y forraje seco en las tres localidades. En promedio, el avance generacional redujo 9% la relación beneficio/costo en la producción de grano; sin embargo, la incrementó 11% en la producción de forraje seco. Los híbridos SB-302 y AS-948 mostraron alto rendimiento de grano y forraje, menor reducción con el avance de generación y alta relación beneficio/costo.
Palabras clave: Zea mays L., beneficio/costo, generaciones F1 y F2 de híbridos.
ABSTRACT
To cope with the high seed cost of maize hybrids, small-scale farmers use F2 and F3 seed. The objective of this study was to determine the effect of using seed from self-pollinated hybrids on grain and forage yields of maize hybrids. The study was conducted under irrigated conditions at Tepezalá and Pabellón de Arteaga, Aguascalientes, and Calera de Victor Rosales, Zacatecas, Mexico during 2001. Twenty-two commercial hybrids and their respective advanced F2 generation were sown under a split split plot arrangement in a complete random block design with three replications. Grain and forage yield, plant height, days to masculine and feminine flowering and husk height were determined. A profit/cost analysis was performed. On average, the use of F2 seed diminished 22% grain and 8% forage yields. Most of the hybrids and their advanced generations did show grain and dry forage yield stability across locations. Due to generational advance, the profit/cost ratio diminished
* Recibido: Junio, 2008
Aceptado: Julio, 2009

Rodolfo Gaytán-Bautista et al.294 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
9% for grain production and increased 11% for dry forage production. Hybrids SB-302 and AS-948 showed high grain and dry forage yields, less reduction with the advance of generation and high profit/cost ratios.
Key words: Zea mays L., F1 and F2 hybrid generations, profitability.
INTRODUCCIÓN
En los estados de Aguascalientes y Zacatecas se siembran 34 mil ha con maíz bajo riego para la producción de grano y 15 500 para forraje, con rendimientos promedio de 5.2 y 50.0 t ha-1, respectivamente (SIAP, 2007). A partir de 1990 se incrementó el uso de híbridos de maíz en Aguascalientes, debido a los subsidios que se aplicaron para la adquisición de semilla certificada, y promoción de las empresas comercializadoras de semilla (Reyes y Robles, 1997). A nivel nacional, sólo 25% de la superficie sembrada con maíz se cultiva con híbridos o variedades registradas de polinización libre (Nadal, 2000). Ante el alto costo que representa la adquisición de semilla de híbridos F1, los productores usan semilla de generaciones avanzadas F2 y F3 de híbridos comerciales, a la espera de que el rendimiento y calidad del grano no se vea afectado en forma significativa. La semilla la obtienen mediante la conservación de grano de la cosecha anterior y con ella siembran el siguiente ciclo de cultivo (Valdivia y Vidal, 1995; De León et al., 1998). Por ello, el desarrollo de híbridos de maíz con alto rendimiento de grano en F1 y mínima depresión endogámica en F2 podría favorecer a los productores a pequeña escala de Aguascalientes y Zacatecas. Lo anterior, es posible en el caso de que el vigor híbrido este determinado por la acción equilibrada de genes co-dominantes y semi-dominantes (De León et al., 1998).
La pérdida de vigor híbrido después de la autofecundación de individuos heterocigóticos es un fenómeno de importancia práctica para los fitomejoradores. Los estudios sobre el efecto del avance generacional por autofecundación en maíz difieren en los resultados; por ejemplo, Ramírez et al. (1986) observaron reducciones de 12 a 18% en el rendimiento de grano en las F2 respecto a la F1 de híbridos tropicales, Valdivia y Vidal (1995) observaron reducciones 45% en el estado de Nayarit y Coutiño et al. (2004) de 22.6% en Chiapas (con 17% de reducción en la relación beneficio/costo). Espinosa-Calderón et al. (1990; 1993) citados por Morris et al. (1999) observaron 25 y 51%
menor rendimiento de grano en F2 en comparación con los híbridos F1 de cruza simple (H-34) y de cruza doble (H-30, H-33, H-36), respectivamente.
Ortiz y Espinosa (1990) reportaron reducción de 35% en rendimiento de grano del híbrido H-149E con semilla F2. Por su parte, Patto-Pacheco et al. (2002) observaron 49% de reducción en variedades de maíz de Brasil. Otros efectos de la autofecundación en maíz son: ampliación del ciclo biológico, incremento en acame, incremento en el porcentaje de pudrición de mazorca y reducción de altura de planta y de la prolificidad (Valdivia y Vidal, 1995; Coutiño et al., 2004). Pixley y Bänziger (2001) reportaron que el avance generacional redujo 32% el rendimiento de grano de híbridos de maíz y 5% de variedades de polinización libre.
Los híbridos de maíz que se utilizan en México se han desarrollado para la producción de grano y los progenitores muestran diferencias en altura de planta y ciclo biológico, entre otras características. Los híbridos de maíz forrajero que se cultivan en México tienen bajo valor energético (en comparación con los de EE.UU) debido a que sólo se dio énfasis al mejoramiento del rendimiento de grano, sin considerar la calidad del forraje (Núñez et al., 2003); el cual es de bajo valor nutritivo por el alto contenidos de fibra (digestibilidad) y baja energía metabolizable (Núñez et al., 2005). En general, los híbridos de maíz de alto rendimiento de grano muestran alto contenido de energía metabolizable (Núñez et al., 2003) y alta digestibilidad (Núñez et al., 2001). En la región de La Laguna, bajo condiciones de riego, el rendimiento de forraje seco alcanzó 19 t ha-1 (Núñez et al., 2003), mientras que en ambientes favorables, Peña et al. (2006) reportaron rendimientos de forraje seco de 17.6 a 23 t ha-1 con el híbrido H-376 y de 20 a 23.9 t ha-1 con H-157. El avance generacional en cuanto a producción de forraje muestra resultados contrastantes. Peña et al. (2003) observaron que las F2 de híbridos forrajeros adaptados a las condiciones agroecológicas de Aguascalientes no difirieron de los respectivos F1 en calidad forrajera. En otro estudio, Peña et al. (2002) observaron que los híbridos F1 mostraron mayor eficiencia productiva y rendimiento de forraje seco en comparación con sus F2. El desarrollo de híbridos de doble propósito es la alternativa en el corto y mediano plazos para incrementar la rentabilidad del maíz en el centro de México.
Existen híbridos con reducido efecto negativo en el rendimiento de grano y forraje al ser utilizadas en F2 por lo que es posible utilizar las generaciones avanzadas

Rendimiento de grano y forraje en híbridos de maíz y su generación avanzada F2 295
de 12 a 18 °C (García, 1988); los suelos son del tipo Xerosol lúvico con textura media y profundidad de 50 a 100 cm, con pendientes menores a 8%.
Se utilizaron 22 híbridos comerciales: 20 de siete empresas productoras de semilla y dos (H-361 y H-311) desarrollados y liberados por el programa de maíz del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) (Cuadro 1). Se obtuvo semilla F2 de cada híbrido por medio de cruzas fraternales en Pabellón de Arteaga, Aguascalientes en 2000, se polinizaron 30 plantas de cada híbrido con una mezcla de polen de las mismas. En Pabellón de Arteaga se sembró el 18 de mayo, en Tepezalá el 14 de junio y en Calera de Víctor Rosales el 25 de abril de 2001. Durante el ciclo. la media de las temperaturas máxima y mínima y la precipitación pluvial fueron: 27.3 y 11.1 °C y 404 mm, respectivamente en Pabellón de Arteaga; 27.8 y 12.6 °C y 311 mm en Tepezalá y 25.2 y 11.0 °C y 291 mm en Calera de Víctor Rosales.
surcos de 10 m de largo, separados a 0.76 m; en dos surcos se evaluó el rendimiento de grano y en los otros dos el de forraje. La densidad de siembra fue a 0.2 m entre plantas para la determinación de rendimiento de grano y a 0.16 m para
para reducir el costo de producción del cultivo (Luna, 2003). El objetivo de esta investigación fue determinar el efecto del avance generacional sobre el rendimiento de grano y forraje seco de 22 híbridos comerciales de maíz y su respectiva generación avanzada F2, adaptados a las regiones productoras de Aguascalientes y Zacatecas.
MATERIALES Y MÉTODOS
La investigación se realizó durante 2001 bajo condiciones de riego en las localidades de Pabellón de Arteaga (22° 09' latitud norte 102° 17' longitud oeste, 1 912 m) y Tepezalá, Aguascalientes (22° 13’ latitud norte, 102° 10’longitud oeste, 2 090 m) y Calera de Víctor Rosales, Zacatecas (22° 33’ latitud norte, 102° 51’ longitud oeste, 2 197 m). Las tres localidades presentan clima seco-árido con lluvias en verano y temperatura media anual
Híbrido Origen Ciclo vegetativo* Textura de grano Color de granoSB-302 Semillas Berentsen Intermedio Dentado Blanco cremosoSB-304 “ “ “ “AS-31 Aspros “ Semi-dentado “AS-910 “ “ Dentado “AS-948 “ “ Semi-dentado “AS-820 “ Precoz “ “Gilsa-120 Gilsa Intermedio Semi-cristalino “Z-21 Heertz Seed “ “ “Z-60 “ Precoz Semi-dentado “C-922 “ Intermedio Semi-cristalino “C-220 “ “ Semi-dentado “C-526 “ “ Semi-cristalino “30G40 Pioneer “ “ “A7597 Asgrow “ “ “Halcón " Precoz Semi-cristalino BlancoPantera “ Intermedio “ Blanco cremosoTromba Ceres “ Semi-dentado “Tornado “ “ “ “Trueno “ “ “ “Fuego “ “ “ “H-311 INIFAP “ “ “H-361 “ “ “ “
Cuadro 1. Origen, ciclo vegetativo y características del grano de 22 híbridos de maíz.
* precoz= 68 días a floración masculina media; intermedio= 85 días a floración masculina media.
Se utilizó un arreglo de parcelas subdivididas con diseño de bloques completos al azar con tres repeticiones en cada localidad. Los factores estudiados fueron: localidad, híbrido y generación. La unidad experimental constó de cuatro
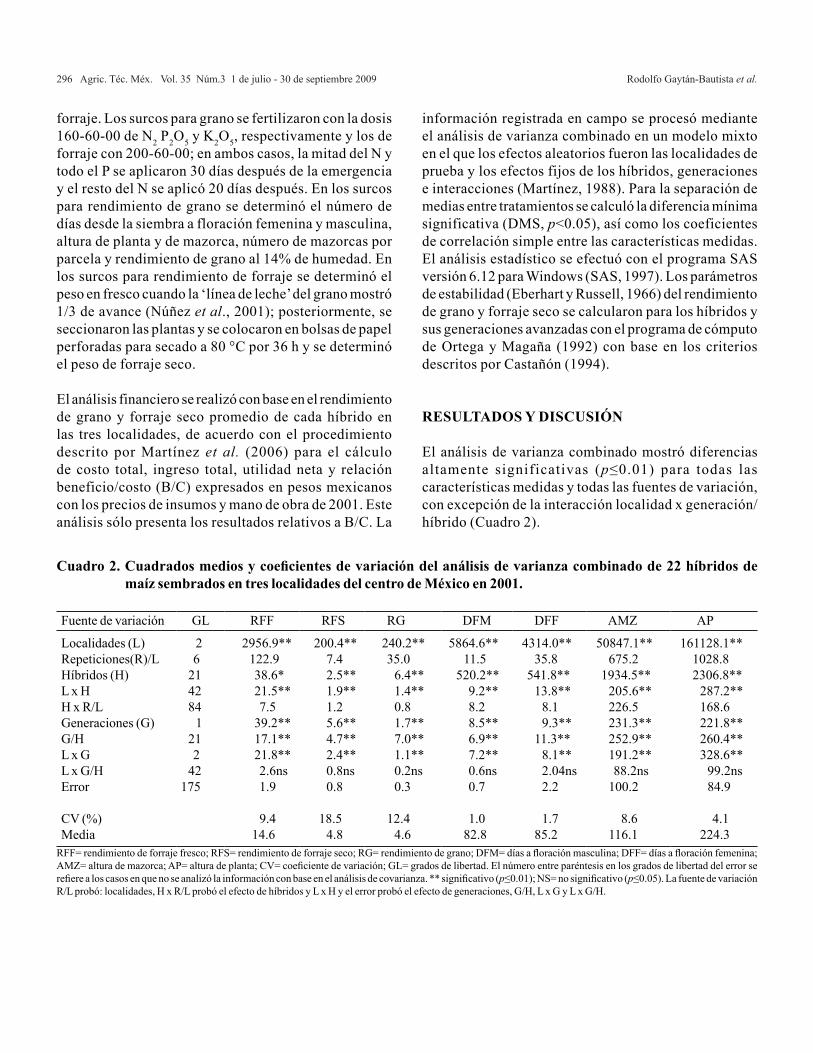
Rodolfo Gaytán-Bautista et al.296 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
forraje. Los surcos para grano se fertilizaron con la dosis 160-60-00 de N2 P2O5 y K2O5, respectivamente y los de forraje con 200-60-00; en ambos casos, la mitad del N y todo el P se aplicaron 30 días después de la emergencia y el resto del N se aplicó 20 días después. En los surcos para rendimiento de grano se determinó el número de días desde la siembra a floración femenina y masculina, altura de planta y de mazorca, número de mazorcas por parcela y rendimiento de grano al 14% de humedad. En los surcos para rendimiento de forraje se determinó el peso en fresco cuando la ‘línea de leche’ del grano mostró 1/3 de avance (Núñez et al., 2001); posteriormente, se seccionaron las plantas y se colocaron en bolsas de papel perforadas para secado a 80 °C por 36 h y se determinó el peso de forraje seco.
El análisis financiero se realizó con base en el rendimiento de grano y forraje seco promedio de cada híbrido en las tres localidades, de acuerdo con el procedimiento descrito por Martínez et al. (2006) para el cálculo de costo total, ingreso total, utilidad neta y relación beneficio/costo (B/C) expresados en pesos mexicanos con los precios de insumos y mano de obra de 2001. Este análisis sólo presenta los resultados relativos a B/C. La
información registrada en campo se procesó mediante el análisis de varianza combinado en un modelo mixto en el que los efectos aleatorios fueron las localidades de prueba y los efectos fijos de los híbridos, generaciones e interacciones (Martínez, 1988). Para la separación de medias entre tratamientos se calculó la diferencia mínima significativa (DMS, p<0.05), así como los coeficientes de correlación simple entre las características medidas. El análisis estadístico se efectuó con el programa SAS versión 6.12 para Windows (SAS, 1997). Los parámetros de estabilidad (Eberhart y Russell, 1966) del rendimiento de grano y forraje seco se calcularon para los híbridos y sus generaciones avanzadas con el programa de cómputo de Ortega y Magaña (1992) con base en los criterios descritos por Castañón (1994).
RESULTADOS Y DISCUSIÓN
El análisis de varianza combinado mostró diferencias altamente significativas (p≤0.01) para todas las características medidas y todas las fuentes de variación, con excepción de la interacción localidad x generación/híbrido (Cuadro 2).
Fuente de variación GL RFF RFS RG DFM DFF AMZ AP
Localidades (L) 2 2956.9** 200.4** 240.2** 5864.6** 4314.0** 50847.1** 161128.1**Repeticiones(R)/L 6 122.9 7.4 35.0 11.5 35.8 675.2 1028.8Híbridos (H) 21 38.6* 2.5** 6.4** 520.2** 541.8** 1934.5** 2306.8**L x H 42 21.5** 1.9** 1.4** 9.2** 13.8** 205.6** 287.2**H x R/L 84 7.5 1.2 0.8 8.2 8.1 226.5 168.6Generaciones (G) 1 39.2** 5.6** 1.7** 8.5** 9.3** 231.3** 221.8**G/H 21 17.1** 4.7** 7.0** 6.9** 11.3** 252.9** 260.4**L x G 2 21.8** 2.4** 1.1** 7.2** 8.1** 191.2** 328.6**L x G/H 42 2.6ns 0.8ns 0.2ns 0.6ns 2.04ns 88.2ns 99.2nsError 175 1.9 0.8 0.3 0.7 2.2 100.2 84.9
CV (%) 9.4 18.5 12.4 1.0 1.7 8.6 4.1Media 14.6 4.8 4.6 82.8 85.2 116.1 224.3
Cuadro 2. Cuadrados medios y coeficientes de variación del análisis de varianza combinado de 22 híbridos de maíz sembrados en tres localidades del centro de México en 2001.
RFF= rendimiento de forraje fresco; RFS= rendimiento de forraje seco; RG= rendimiento de grano; DFM= días a floración masculina; DFF= días a floración femenina; AMZ= altura de mazorca; AP= altura de planta; CV= coeficiente de variación; GL= grados de libertad. El número entre paréntesis en los grados de libertad del error se refiere a los casos en que no se analizó la información con base en el análisis de covarianza. ** significativo (p≤0.01); NS= no significativo (p≤0.05). La fuente de variación R/L probó: localidades, H x R/L probó el efecto de híbridos y L x H y el error probó el efecto de generaciones, G/H, L x G y L x G/H.

Rendimiento de grano y forraje en híbridos de maíz y su generación avanzada F2 297
Resultados similares se observaron en variedades de maíz con origen y estatus genético diferente, tanto en México (Arellano et al., 2003; Núñez et al., 2003; Coutiño et al., 2004; Peña et al., 2006) como en otros países (Meeske et al., 2000; Guimaraes et al., 2002). Los más altos rendimientos de grano y forraje seco se observaron en Tepezalá; la máxima altura de planta y días a floración femenina y masculina en Calera de Víctor Rosales y la máxima altura de mazorca en Tepezalá y Calera de Víctor Rosales (Cuadro 3). Lo anterior indicó que las condiciones de cultivo en la localidad de Pabellón de Arteaga fueron las menos favorables, posiblemente debido a la menor fertilidad y profundidad del suelo en comparación con las otras. Guimaraes et al. (2002) observaron en Brasil
diferencias significativas entre híbridos de maíz debido a la diversidad genética del germoplasma y a la variación entre ambientes de prueba.
Los más altos rendimientos de grano en F1 se observaron en los híbridos Pantera, 30G40, Z-21, SB-302 y SB-304 y en F2 en SB-302 y Pantera. Los híbridos con alto rendimiento de forraje en F1 se observaron en Halcón, Z-60, AS-31, Fuego y H-311 y en F2 en AS-948, SB-302, AS-31 y Pantera. El avance generacional redujo 22% rendimiento de grano y 8% de forraje seco. Todos los híbridos mostraron reducción en rendimiento de grano por efecto del avance generacional, los menos afectados fueron SB-302, Gilsa-120, AS-948 y Halcón; en contraste, AS-948 produjo más forraje seco en F2 que en F1.
Patto-Pacheco et al. (2002) (49%). Peña y Arellano (1990); Ramírez et al. (1986) y De León et al. (1998) observaron reducción de 9.5, 12 a 18 y 15%, respectivamente por efecto del avance generacional en maíz; mientras que Coutiño et al. (2004) reportó 22% de reducción en rendimiento de grano.
Se observó una ligera disminución en la producción de forraje seco, a diferencia de Peña et al. (2002) quienes reportaron reducción de 25% con el avance generacional de híbridos de maíz. Núñez et al. (2003) observaron baja asociación entre productividad y calidad del forraje de maíz. Aunado a lo anterior, los rendimientos de grano y de forraje seco fueron menores que los reportados por otros autores en experimentos conducidos en regiones tropicales y subtropicales de México (Ramírez et al., 1986; Valdivia y Vidal, 1995; De León et al., 1998; Núñez et al., 2003; Coutiño et al., 2004; Peña et al., 2006) lo que indica que hubo efecto ambiental restrictivo en la productividad del germoplasma de maíz
Localidad Rendimiento(t ha-1) Días a floración Altura (m)Grano Forraje seco Forraje fresco Masculina Femenina Planta Mazorca
Calera de Víctor Rosales 4.5 4.4 18.3 90 91 2.6 1.3Pabellón de Arteaga 3.7 3.5 10.3 77 80 1.9 0.9Tepezalá, Aguascalientes 6.1 5.7 15.3 81 84 2.2 1.3DMS (p<0.05) 0.3 0.2 0.6 1 2 0.1 0.1
Cuadro 3. Promedios de características agronómicas de 22 híbridos de maíz y sus generaciones avanzadas F2 en tres localidades del centro de México. 2001.
Los híbridos que mostraron alta disminución promedio en rendimiento de grano y forraje seco fueron: Fuego, AS-820, Halcón y Z-60 y el de menor efecto negativo, SB-302 (Cuadro 4). El avance generacional no afectó significativamente los días a floración femenina y masculina ni redujo la altura de planta y mazorca y los valores del error estándar de todas las variables se redujeron en F2 (Cuadro 5).
Las diferencias en rendimiento de grano y forraje observadas entre híbridos como consecuencia del avance de generación se deben principalmente al origen del germoplasma progenitor (patrones heteróticos, aptitud combinatoria, nivel de endogamia) más que al número de líneas que participan en la formación de cada híbrido dado que la mayoría de ellos son cruzas trilineales (Morris et al., 1999; Gaytán et al., 2005). El efecto negativo en rendimiento de grano de F1 a F2 fue menor que el reportado por Valdivia y Vidal (1995) y Espinosa et al. (1990; 1993) citados por Morris et al. (1999) quienes observaron reducción de 45, 42 y 51%, respectivamente; así como Ortíz y Espinosa (1990) (35%) y
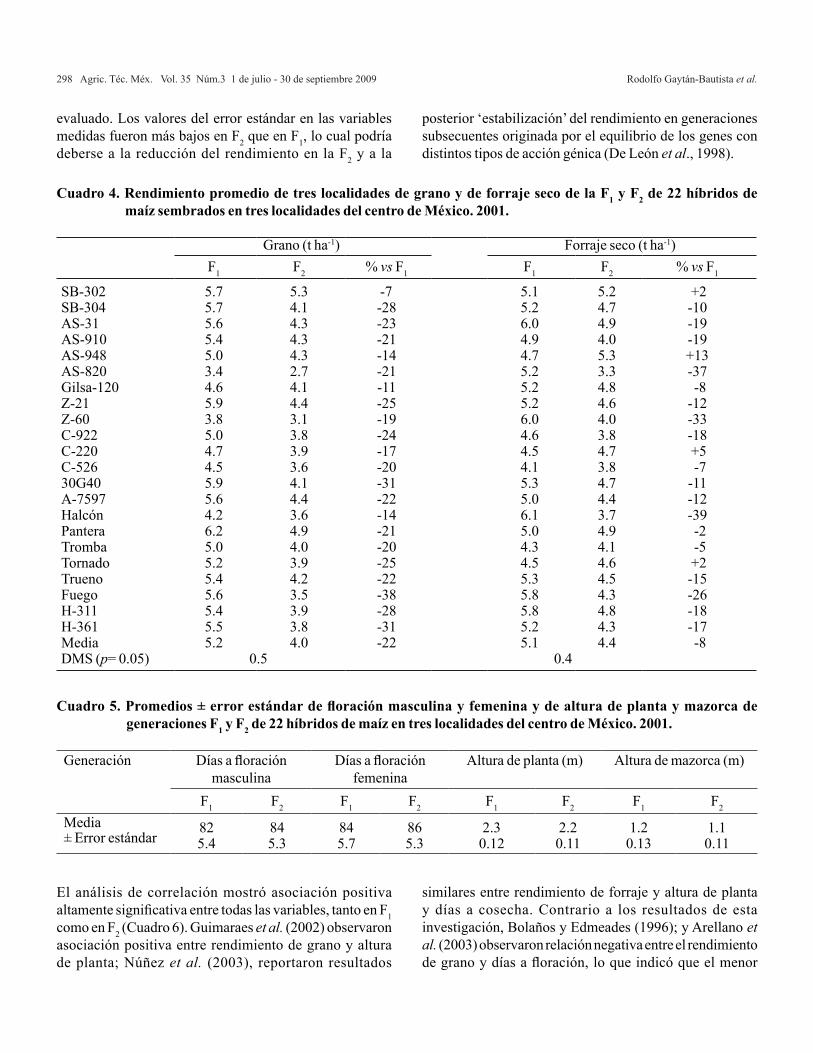
Rodolfo Gaytán-Bautista et al.298 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
evaluado. Los valores del error estándar en las variables medidas fueron más bajos en F2 que en F1, lo cual podría deberse a la reducción del rendimiento en la F2 y a la
similares entre rendimiento de forraje y altura de planta y días a cosecha. Contrario a los resultados de esta investigación, Bolaños y Edmeades (1996); y Arellano et al. (2003) observaron relación negativa entre el rendimiento de grano y días a floración, lo que indicó que el menor
Generación Días a floración masculina
Días a floración femenina
Altura de planta (m) Altura de mazorca (m)
F1 F2 F1 F2 F1 F2 F1 F2
Media 82 84 84 86 2.3 2.2 1.2 1.1± Error estándar 5.4 5.3 5.7 5.3 0.12 0.11 0.13 0.11
Grano (t ha-1) Forraje seco (t ha-1)F1 F2 % vs F1 F1 F2 % vs F1
SB-302 5.7 5.3 -7 5.1 5.2 +2SB-304 5.7 4.1 -28 5.2 4.7 -10AS-31 5.6 4.3 -23 6.0 4.9 -19AS-910 5.4 4.3 -21 4.9 4.0 -19AS-948 5.0 4.3 -14 4.7 5.3 +13AS-820 3.4 2.7 -21 5.2 3.3 -37Gilsa-120 4.6 4.1 -11 5.2 4.8 -8Z-21 5.9 4.4 -25 5.2 4.6 -12Z-60 3.8 3.1 -19 6.0 4.0 -33C-922 5.0 3.8 -24 4.6 3.8 -18C-220 4.7 3.9 -17 4.5 4.7 +5C-526 4.5 3.6 -20 4.1 3.8 -730G40 5.9 4.1 -31 5.3 4.7 -11A-7597 5.6 4.4 -22 5.0 4.4 -12Halcón 4.2 3.6 -14 6.1 3.7 -39Pantera 6.2 4.9 -21 5.0 4.9 -2Tromba 5.0 4.0 -20 4.3 4.1 -5Tornado 5.2 3.9 -25 4.5 4.6 +2Trueno 5.4 4.2 -22 5.3 4.5 -15Fuego 5.6 3.5 -38 5.8 4.3 -26H-311 5.4 3.9 -28 5.8 4.8 -18H-361 5.5 3.8 -31 5.2 4.3 -17Media 5.2 4.0 -22 5.1 4.4 -8DMS (p= 0.05) 0.5 0.4
posterior ‘estabilización’ del rendimiento en generaciones subsecuentes originada por el equilibrio de los genes con distintos tipos de acción génica (De León et al., 1998).
Cuadro 4. Rendimiento promedio de tres localidades de grano y de forraje seco de la F1 y F2 de 22 híbridos de maíz sembrados en tres localidades del centro de México. 2001.
Cuadro 5. Promedios ± error estándar de floración masculina y femenina y de altura de planta y mazorca de generaciones F1 y F2 de 22 híbridos de maíz en tres localidades del centro de México. 2001.
El análisis de correlación mostró asociación positiva altamente significativa entre todas las variables, tanto en F1 como en F2 (Cuadro 6). Guimaraes et al. (2002) observaron asociación positiva entre rendimiento de grano y altura de planta; Núñez et al. (2003), reportaron resultados
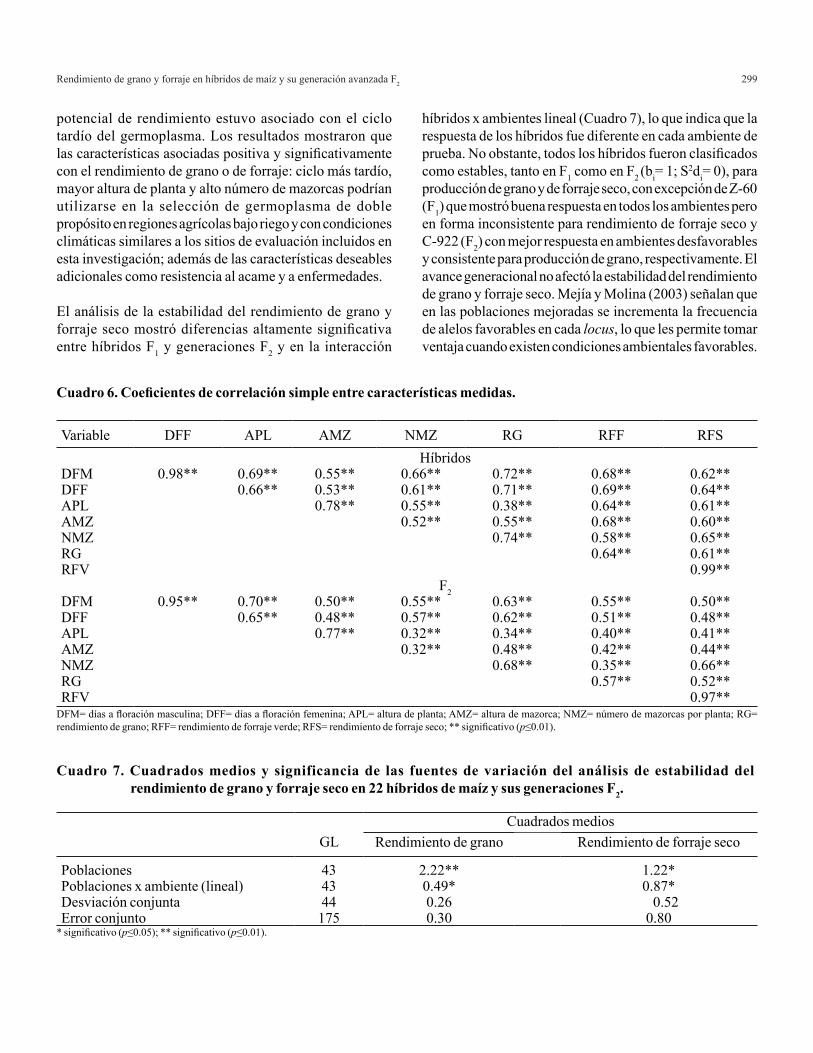
Rendimiento de grano y forraje en híbridos de maíz y su generación avanzada F2 299
potencial de rendimiento estuvo asociado con el ciclo tardío del germoplasma. Los resultados mostraron que las características asociadas positiva y significativamente con el rendimiento de grano o de forraje: ciclo más tardío, mayor altura de planta y alto número de mazorcas podrían utilizarse en la selección de germoplasma de doble propósito en regiones agrícolas bajo riego y con condiciones climáticas similares a los sitios de evaluación incluidos en esta investigación; además de las características deseables adicionales como resistencia al acame y a enfermedades.
El análisis de la estabilidad del rendimiento de grano y forraje seco mostró diferencias altamente significativa entre híbridos F1 y generaciones F2 y en la interacción
híbridos x ambientes lineal (Cuadro 7), lo que indica que la respuesta de los híbridos fue diferente en cada ambiente de prueba. No obstante, todos los híbridos fueron clasificados como estables, tanto en F1 como en F2 (bi= 1; S2di= 0), para producción de grano y de forraje seco, con excepción de Z-60 (F1) que mostró buena respuesta en todos los ambientes pero en forma inconsistente para rendimiento de forraje seco y C-922 (F2) con mejor respuesta en ambientes desfavorables y consistente para producción de grano, respectivamente. El avance generacional no afectó la estabilidad del rendimiento de grano y forraje seco. Mejía y Molina (2003) señalan que en las poblaciones mejoradas se incrementa la frecuencia de alelos favorables en cada locus, lo que les permite tomar ventaja cuando existen condiciones ambientales favorables.
GLCuadrados medios
Rendimiento de grano Rendimiento de forraje seco
Poblaciones 43 2.22** 1.22*Poblaciones x ambiente (lineal) 43 0.49* 0.87*Desviación conjunta 44 0.26 0.52Error conjunto 175 0.30 0.80
Variable DFF APL AMZ NMZ RG RFF RFSHíbridos
DFM 0.98** 0.69** 0.55** 0.66** 0.72** 0.68** 0.62**DFF 0.66** 0.53** 0.61** 0.71** 0.69** 0.64**APL 0.78** 0.55** 0.38** 0.64** 0.61**AMZ 0.52** 0.55** 0.68** 0.60**NMZ 0.74** 0.58** 0.65**RG 0.64** 0.61**RFV 0.99**
F2DFM 0.95** 0.70** 0.50** 0.55** 0.63** 0.55** 0.50**DFF 0.65** 0.48** 0.57** 0.62** 0.51** 0.48**APL 0.77** 0.32** 0.34** 0.40** 0.41**AMZ 0.32** 0.48** 0.42** 0.44**NMZ 0.68** 0.35** 0.66**RG 0.57** 0.52**RFV 0.97**
Cuadro 6. Coeficientes de correlación simple entre características medidas.
DFM= días a floración masculina; DFF= días a floración femenina; APL= altura de planta; AMZ= altura de mazorca; NMZ= número de mazorcas por planta; RG= rendimiento de grano; RFF= rendimiento de forraje verde; RFS= rendimiento de forraje seco; ** significativo (p≤0.01).
Cuadro 7. Cuadrados medios y significancia de las fuentes de variación del análisis de estabilidad del rendimiento de grano y forraje seco en 22 híbridos de maíz y sus generaciones F2.
* significativo (p≤0.05); ** significativo (p≤0.01).
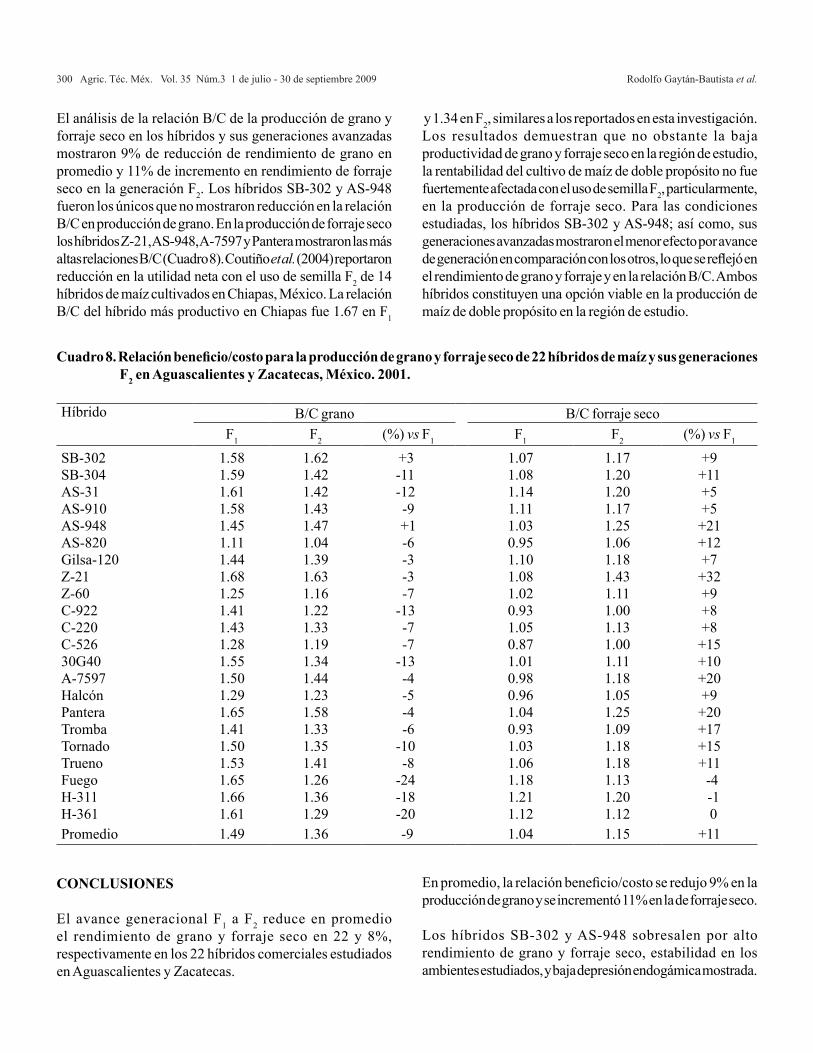
Rodolfo Gaytán-Bautista et al.300 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
El análisis de la relación B/C de la producción de grano y forraje seco en los híbridos y sus generaciones avanzadas mostraron 9% de reducción de rendimiento de grano en promedio y 11% de incremento en rendimiento de forraje seco en la generación F2. Los híbridos SB-302 y AS-948 fueron los únicos que no mostraron reducción en la relación B/C en producción de grano. En la producción de forraje seco los híbridos Z-21, AS-948, A-7597 y Pantera mostraron las más altas relaciones B/C (Cuadro 8). Coutiño et al. (2004) reportaron reducción en la utilidad neta con el uso de semilla F2 de 14 híbridos de maíz cultivados en Chiapas, México. La relación B/C del híbrido más productivo en Chiapas fue 1.67 en F1
y 1.34 en F2, similares a los reportados en esta investigación. Los resultados demuestran que no obstante la baja productividad de grano y forraje seco en la región de estudio, la rentabilidad del cultivo de maíz de doble propósito no fue fuertemente afectada con el uso de semilla F2, particularmente, en la producción de forraje seco. Para las condiciones estudiadas, los híbridos SB-302 y AS-948; así como, sus generaciones avanzadas mostraron el menor efecto por avance de generación en comparación con los otros, lo que se reflejó en el rendimiento de grano y forraje y en la relación B/C. Ambos híbridos constituyen una opción viable en la producción de maíz de doble propósito en la región de estudio.
En promedio, la relación beneficio/costo se redujo 9% en la producción de grano y se incrementó 11% en la de forraje seco.
Los híbridos SB-302 y AS-948 sobresalen por alto rendimiento de grano y forraje seco, estabilidad en los ambientes estudiados, y baja depresión endogámica mostrada.
Híbrido B/C grano B/C forraje secoF1 F2 (%) vs F1 F1 F2 (%) vs F1
SB-302 1.58 1.62 +3 1.07 1.17 +9SB-304 1.59 1.42 -11 1.08 1.20 +11AS-31 1.61 1.42 -12 1.14 1.20 +5AS-910 1.58 1.43 -9 1.11 1.17 +5AS-948 1.45 1.47 +1 1.03 1.25 +21AS-820 1.11 1.04 -6 0.95 1.06 +12Gilsa-120 1.44 1.39 -3 1.10 1.18 +7Z-21 1.68 1.63 -3 1.08 1.43 +32Z-60 1.25 1.16 -7 1.02 1.11 +9C-922 1.41 1.22 -13 0.93 1.00 +8C-220 1.43 1.33 -7 1.05 1.13 +8C-526 1.28 1.19 -7 0.87 1.00 +1530G40 1.55 1.34 -13 1.01 1.11 +10A-7597 1.50 1.44 -4 0.98 1.18 +20Halcón 1.29 1.23 -5 0.96 1.05 +9Pantera 1.65 1.58 -4 1.04 1.25 +20Tromba 1.41 1.33 -6 0.93 1.09 +17Tornado 1.50 1.35 -10 1.03 1.18 +15Trueno 1.53 1.41 -8 1.06 1.18 +11Fuego 1.65 1.26 -24 1.18 1.13 -4H-311 1.66 1.36 -18 1.21 1.20 -1H-361 1.61 1.29 -20 1.12 1.12 0Promedio 1.49 1.36 -9 1.04 1.15 +11
Cuadro 8. Relación beneficio/costo para la producción de grano y forraje seco de 22 híbridos de maíz y sus generaciones F2 en Aguascalientes y Zacatecas, México. 2001.
CONCLUSIONES
El avance generacional F1 a F2 reduce en promedio el rendimiento de grano y forraje seco en 22 y 8%, respectivamente en los 22 híbridos comerciales estudiados en Aguascalientes y Zacatecas.

Rendimiento de grano y forraje en híbridos de maíz y su generación avanzada F2 301
AGRADECIMIENTOS
Los autores agradecen el financiamiento de este trabajo al SIHGO-CONACYT (Proyecto 19990201021). Así como al Dr. Luis Reyes-Muro (INIFAP-Pabellón de Arteaga, México) en el análisis financiero incluido en este trabajo. Netzahualcóyotl Mayek-Pérez fue becario del Sistema Nacional de Investigadores (SIN) y de los programas EDI y COFAA del Instituto Politécnico Nacional.
LITERATURA CITADA
Arellano, V. J. L.; Tut, C. C.; María, M. R.; Salinas, M. Y. y Taboada, G. O. R. 2003. Maíz azul de los Valles Altos de México. I. Rendimiento de grano y caracteres agronómicos. Rev. Fitotec. Mex. 26:101-107.
Bolaños, J. and Edmeades, G. O. 1996. The importance of the anthesis silking interval in breeding for drought tolerance in tropical maize. Field Crops Res. 48:65-80.
Castañón, G. 1994. Estudio de la estabilidad en líneas avanzadas y variedades de arroz (Oryza sativa L.) usando dos metodologías. Agron. Mesoamer. 5:118-125.
Coutiño, E. B.; Sánchez, G. G. y Vidal, M. V. A. 2004. El uso de semilla F2 de híbridos de maíz en la Frailesca, Chiapas, reduce el rendimiento y las ganancias netas. Rev. Fitotec. Mex. 27:261-266.
De León, H.; Jaramillo, A.; Martínez, G. y Rodríguez, S. 1998. Híbridos dobles de maíz de baja depresión endogámica en F2. Agron. Mesoamer. 9:38-41.
Eberhart, S. A. and Russell, W. A. 1966. Stability parameters for comparing varieties. Crop Sci. 6:22-35.
García, E. 1988. Modificaciones al sistema de clasificación climática de Köppen (para adaptarlo a las condiciones de la república mexicana). Cuarta edición. UNAM. México. 217 p.
Gaytán, B. R.; Reyes, M. L.; Martínez, G. M. I.; Mayek, P. N.; Padilla, R. S. y Luna, F. M. 2005. Depresión del rendimiento de grano y forraje de híbridos de maíz en generaciones avanzadas. Agric. Téc. Méx. 31:165-175.
Guimaraes, P.; Juliatti, F. C.; Lopes, A. e Toshyiuki, O. 2002. Avaliacao do desempenho agronomico de hibridos de milho em Uberlandia, MG. Pesq. Agropec. Bras. 37:597-602.
Luna, F. M. 2003. ¿Porqué no se deja de producir maíz en México? In: El campo no aguanta más. Schwentesius, R.; Gómez, M. A. y Calva, J. L. (coords.). Universidad Autónoma Chapingo. Chapingo, Texcoco, Estado de México, México. pp. 115-132.
Martínez, G. A. 1988. Diseños experimentales. Métodos y elementos de teoría. Trillas. México. 756 p.
Martínez, G. M. I.; Gaytán, B. R.; Reyes, M. L.; Mayek, P. N.; Padilla, R. J. S. y Luna, F. M. 2006. Rentabilidad de las generaciones F1, F2 y F3 de híbridos de maíz. Agrociencia 40:677-685.
Mejía, C. A. y Molina, G. J. D. 2003. Cambios de estabilidad en el rendimiento de variedades tropicales de maíz. Rev. Fitotec. Mex. 26:89-94.
Meeske, R.; Basson, H. M.; Peinar, J. P. and Cruywagnen, C. W. 2000. A comparison of the yield, nutritional value and predicted production potential of different maize hybrids for silage production. South African J. Anim. Sci. 30:18-21.
Morris, M. L.; Risopoulos, J. and Beck, D. 1999. Genetic change in farmer-recycled maize seed: a review of the evidence. CIMMYT Economics Working Paper No. 99-07. México, D. F. 69 p.
Nadal, A. 2000. El caso del maíz mexicano en el NAFTA: Variabilidad genética y liberalización comercial. Biodiversidad 24:3-12.
Núñez, H. G.; Faz, C. R.; Tovar, G. M. R. y Zavala, G. A. 2001. Híbridos de maíz para la producción de forraje con alta digestibilidad en el norte de México. Téc. Pecu. Méx. 39:77-88.
Núñez, H. G.; Contreras, G. E. F. y Faz, C. R. 2003. Características agronómicas y químicas importantes en híbridos de maíz para forraje con alto valor energético. Téc. Pecu. Méx. 41:37-48.
Núñez, H. G.; Faz, C. R.; González, C. F. y Peña, R. A. 2005. Madurez de híbridos de maíz a la cosecha para mejorar la producción y calidad del forraje. Téc. Pecu. Méx. 43:69-78.
Ortega-Alcalá, J. y Magaña-Torres, O. S. 1992. Parámetros de estabilidad en base al modelo propuesto por Eberhart y Russell (Computer File). Campo Experimental Valle de México-INIFAP. Chapingo, México.
Ortiz, T. E. y Espinosa, C. A. 1990. Rendimiento de híbridos de maíz (Zea mays L.) en la zona de transición El Bajío-Valles Altos por efecto de la utilización de semilla de generación F1 y F2. Rev. Chapingo 15:49-52.

Rodolfo Gaytán-Bautista et al.302 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Patto-Pacheco, C. A.; Dos Santos, M. X.; Damiao-Cruz, C.; Netto-Parentoni, S.; De Oliveira-Guimaraes, P. E.; Gomes E Gama, E. E.; Da Silva, A. E.; Lemos de Carvalho, H. W. and Vieira-Junior, P. A. 2002. Inbreeding depression of 28 maize elite open pollinated varieties. Genet. Mol. Biol. 25:441-448.
Pixley, G. and Bänziger, M. 2001. Open-pollinated maize varieties: a backward step or valuable option for farmers? In: integrated approaches to higher maize productivity in the new millennium. Proceedings of the seventh eastern and southern Africa regional maize conference. Friesen, D. K. and Palmer, A. F. E. (eds.). Nairobi, Kenya. pp. 22-28.
Peña, R. A. y Arellano, V. J. L. 1990. Efecto de la depresión endogámica sobre el rendimiento de grano y sus componentes en el maíz híbrido H-131. Rev. Chapingo 15:12-16.
Peña, R. A.; Núñez, H. G. y González, C. F. 2002. Potencial forrajero de poblaciones de maíz y relación entre atributos agronómicos con la calidad. Téc. Pecu. Méx. 40:215-228.
Peña, R. A.; Núñez, H. G. y González, C. F. 2003. Importancia de la planta y el elote en poblaciones de maíz para el mejoramiento genético de la calidad forrajera. Tec. Pecu. Méx. 41:63-74.
Peña, R. A.; González, C. F.; Núñez, H. G.; Tovar, G. M. R.; Preciado, O. E.; Terrón, I. A.; Gómez, M. N. y Ortega, C. A. 2006. Estabilidad del rendimiento y calidad forrajera de híbridos de maíz. Rev. Fitotec. Mex. 29:109-114. (Número Especial 2).
Ramírez, V. P.; Balderas, M. M. y Gerón, X. F. 1986. Potencial productivo de las generaciones avanzadas de los híbridos tropicales de maíz H-503, H-507 y H-510. Rev. Fitotec. Mex. 8:20-34.
Reyes, M. L. y Robles, E. F. J. 1997. Impacto de la validación de tecnología en el programa kilo por kilo de maíz en Aguascalientes. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias, Campo Experimental Pabellón. Pabellón de Arteaga, México. 20 p. (Folleto Informativo Núm. 7).
Servicio de Información Agroalimentaria y Pesquera (SIAP). 2007. Anuario estadístico 2005. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA) (En línea). Disponible en http://www.siap.gob.mx. (Consultado el 25 de junio de 2007).
Statistical Analysis System (SAS Institute). 1997. Version 6.12. Cary, NC, USA.
Valdivia, B. R. y Vidal, M. V. A. 1995. Efecto de generaciones avanzadas en la producción de diferentes tipos de híbridos de maíz. Rev. Fitotec. Mex. 18:69-76.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 303-311
ASPECTOS TÉCNICOS Y CARACTERIZACIÓN DEL PRODUCTOR DE DURAZNO EN EL ESTADO DE MÉXICO, MÉXICO*
TECHNICAL ISSUES AND PEACH FARMER TYPOLOGY IN THE STATE OF
MEXICO, MEXICO
Bertha Sofía Larqué Saavedra1, Dora Ma. Sangerman-Jarquín1§, Benito Ramírez Valverde2, Agustín Navarro Bravo3 y María Elena Serrano Flores4
1Programa de Economía, Campo Experimental Valle de México, INIFAP, km 18.5 carretera Los Reyes- Lechería, A. P. 10, C. P. 56230 Chapingo, Texcoco, Estado de México. Tel. y Fax. 01 595 95 4 29 64 y 01 595 95 42877 y 43536 Ext. 162, ([email protected]), 2Posgrado en Estrategias para el Desarrollo Agrícola Regional, Campus- Puebla, Colegio de Postgraduados en Ciencias Agrícolas, km 125.5 carretera Federal México- Puebla C. P. 72130, Puebla, Puebla, A. P. 2-12, colonia Libertad, Tel: 22 85 14 42, ([email protected]), Recursos Naturales-Postgrado en Edafología, Colegio de Postgraduados en Ciencias Agrícolas, km 35.5 carretera México-Texcoco, C. P. 56230, Montecillos, Texcoco Estado de México, México. 4Centro Interdisciplinario de Investigaciones y Estudios sobre Medio Ambiente y Desarrollo-CIIEMAD- Unidad Académica, Instituto Politécnico Nacional, calle 30 de junio de 1520 s/n, colonia Barrio Laguna Ticomán, C. P. 07330, ([email protected]), 01 55 729 6000 ext. 52707 y 52708. §Autora para correspondencia: [email protected].
* Recibido: Julio, 2008 Aceptado: Septiembre, 2009
RESUMEN
El cultivo de durazno en México cuenta con una superficie sembrada de 45 584 ha, y rendimiento promedio de 4.6 t ha-1, genera un valor de producción de $ 1 185 558. En el centro del país, el Estado de México ocupa el tercer lugar en producción de esta fruta. En el estado, el municipio de Coatepec de Harinas tiene una superficie sembrada de 1 988 ha, un rendimiento promedio de 14.8 t ha-1 y un volumen de producción de 29 350 t. El estudio se realizó en 2007 para analizar la caracterización técnica y tipología del productor de durazno. El método de muestreo utilizado fue una muestra simple aleatoria, la cual consistió en aplicar un cuestionario con preguntas cerradas y abiertas a 50 productores ejidales y pequeños propietarios. Los resultados indicaron que los principales problemas técnicos fueron: manejo agronómico, calidad del fruto, tamaño de la huerta y los mercados en los que venden su producto. Con relación al productor, se identificó la necesidad de promover la organización de los mismos para la búsqueda de apoyos financieros suficientes y oportunos para la producción y comercialización de durazno, para facilitar la venta directa al consumidor o al mayorista y para dar valor agregado al producto.
Palabras clave: Prunus persica L., Batsch., caracterización técnica, productor, tipología.
ABSTRACT
The peach crop in Mexico has a planted area of 45 584 ha and average yield of 4.6 t ha-1, generating a production value of $ 1 185 558. In the center of the country, the State of Mexico is third in the production of this fruit. In the state, the county of Coatepec de Harinas, has a planted area of, 1 988 ha, an average yield of 14.8 t ha-1 and a production volume of 29 350 t. The study was conducted in 2007 to analyze the technical aspects of production and type of peach farmer. The sampling method used was a simple random sample with proportional allocation to the size of the stratum, which consisted of applying a questionnaire with closed and open questions to 50 ejidal (communal) producers and smallholders. The results indicated that major technical problems were: agronomic management, fruit quality, size of the orchard and the markets in which they sell their

Bertha Sofía Larqué Saavedra et al.304 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
product. In relation to farmers, there is a need to promote their organization seeking to secure financial support for production and timely marketing, to facilitate the direct sale to consumers or to wholesale and to add value to their product.
Key works: Prunus persica L. Batsch., characterization technical, farmer, typology.
INTRODUCCIÓN
El cultivo del durazno (Prunas persica L. Bastch), originario de China, se introdujo a México con los españoles hace más de 450 años. En 1927, en México se establecen plantaciones de durazno con fines comerciales, con un total de 1 579 ha a nivel nacional (Claridades, 2000). El Estado de México inició su producción comercial en la década de los 70’s. La mayoría de las huertas de durazno (90%) están plantadas con material vegetativo proveniente de San Gabriel, conocido también como Irene o Lucero, el cual concentra la producción en un período, por lo que es importante adoptar variedades y selecciones criollas con características de floración tardía, alta productividad, sin alternancia en la producción y que brinde producción escalonada (SAGARPA, 2006 a-c). De acuerdo con Gutiérrez (2006), el durazno tipo San Gabriel fue el material genético más importante en el Estado de México durante las décadas de los 80 y 90.
En el Estado de México la fruticultura se ha desarrollado como una actividad económica importante, y ha constituido en un mercado de fuerza laboral especializada en tareas propias de esta actividad. La superficie que los productores destina a la producción de durazno (Prunus Persica L. Batsch), es cultivada principalmente con las variedades Diamante, Amapre, Dorado, Oro y Criollo. El municipio de Coatepec de Harinas destaca por sus altos rendimientos y volúmenes de producción. Los productores se enfrentan a problemas técnicos los cuales se derivan principalmente de la falta de asesoría técnica que les apoye en la toma de decisiones para el cultivo hasta canales de comercialización destinados a este proceso; así como gestionar financiamientos para la producción y darle un valor agregado a su producto.
En este estudio estamos proporcionando información técnica, socioeconómica y tipología del productor, que sirva para seguir impulsando el desarrollo de este cultivo tan importante para el Estado de México. El objetivo de esta investigación fue analizar la caracterización técnica del sistema de producción; así como la caracterización del productor de durazno.
MARCO REGIONAL DE LA ZONA DE ESTUDIO
El municipio de Coatepec de Harinas se localiza al sur del Estado de México (PMDUT, 2007), colinda al norte con el municipio de Toluca, Tenango del Valle y Zinacantepec; al sur con Almoloya de Alquisiras, Zacualpan e Ixtapan de la Sal; al este con Villa Guerrero y al oeste con Temascaltepec y Texcaltitlan. Clima semicálido, semifrío y templado, precipitación anual de 1 300 y 1 700 m (García, 2004).
MATERIALES Y MÉTODOS
La investigación se realizó en 2007, de acuerdo a la sistematización de la información se analizó la tipología del productor del cual (Duch, 1998) refiere al medio conceptual, que agrupa a las unidades de producción agrícola en conjuntos con características similares, idéntica y precisa la problemática técnica, económica y social de cada tipo de productor. En este caso se analizó: el número de productores (información general de la familia), aspectos sociales y económicos, tecnología de producción, superficie plantada, tenencia de la tierra (privada y ejidal), aspectos agronómicos (plagas, enfermedades y malezas y su combate), fertilizantes, aspectos de asistencia técnica y apoyo financiero, aspecto destacado por varios autores en diferentes estudios (Ekboir, 2004; USDA, 20005; De Grammont, 2006).
Se usó un muestreo aleatorio donde el tamaño de muestra es definido por (Sukhatme y Balkrishna, 1970):
Figura 1. Localización geográfica del municipio de Coatepec de Harinas, Estado de México, México, 2007.
,
yS),(t
N11
yS
2),(t
n
2N
2
2
2
2N
22
ε∞α+
ε
∞α
=
COATEPEC HARINAS
Aspectos técnicos y caracterización del productor de durazno en el Estado de México, México 305
donde, εyN es el error permisible en por ciento de la media de la variable de interés y 1 - α el grado de confianza. La variable considerada fue el rendimiento en kg ha-1, la cual describe una de las propiedades más importantes en la caracterización del sistema de producción del duraznero. Con base a lo anterior y considerando un error permisible de ε = 0.1 y α= 0.05 se determinó un tamaño de muestra total de 50 unidades de producción, aplicando el mismo número de cuestionarios a los productoresde la comunidad de Coatepec de Harinas, Estado de México.
Como parte de la metodología se entrevistó a informantes clave: autoridades municipales, personal de la Secretaría de Desarrollo Agropecuario (SEDAGRO), Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA) y Presidentes de las Uniones de Productores.
Producción de durazno
Por superficie de riego en el Distrito de Desarrollo Rural (DDR) 078, Coatepec de Harinas, ocupó el nivel más alto de superficie sembrada de durazno con 1 988 ha, un rendimiento promedio de 14.8 t ha-1 y un volumen de 29 350 t, en comparación con otros municipios que conforman este distrito (Cuadro 1). En Coatepec de Harinas, la variedad más difundida de durazno es Diamante, seguido de otras variedades como: Amapre, Oro, Dorado y Criollo (Gutiérrez, 2006;Pérez, 2004).
En cuanto a la superficie de temporal, vuelve a destacar en el DDR 078 Coatepec de Harinas con 236 ha sembradas, un rendimiento promedio de 10 t ha-1 y volumen de producción de 2 360 t (Cuadro 2).
Cochisquilla, 1ª y 2ª de San Miguel, San José, El Potrero e Ixtlahuaca (base de datos de la campaña contra la mosca nativa de la fruta del municipio de Coatepec de Harinas. Comité de Sanidad Vegetal del Estado de México, México (Cuadro 3).
DDR Superficie sembrada (ha) Volumen de producción (t) Rendimiento (t ha-1)Atlacomulco 3 8 2.7Coatepec de Harinas 1 988 29 350 14.8Jilotepec 50 389.5 7.8Tejupilco 16 165 10.3Valle de Bravo 28 214 7.6Zumpango 5 2.50 0.5
DDR Superficie sembrada (ha) Volumen de producción (t) Rendimiento (t ha-1)
Atlacomulco 9 36.50 4.1Coatepec de Harinas 236 2360 10Texcoco 17 130.60 7.7Toluca 0.50 1 2Valle de Bravo 11 102 9.3Zumpango 8 0 0
_
Cuadro1. Superficie sembrada, rendimiento y volumen de producción en riego de durazno Diamante de Coatepec de Harinas, Estado de México (2007).
Fuente: trabajo de campo, 2007 (SIAP, 2006; PRSPD, 2007).
Cuadro 2. Superficie sembrada, rendimiento y volumen de producción en temporal de durazno Diamante, Coatepec de Harinas, Estado de México (2007).
Fuente: trabajo de campo, 2007 (SIAP, 2006; PRSPD, 2007).
Tamaño de las parcelas de los productores de durazno
Las localidades del municipio de Coatepec de Harinas dedicadas a la producción de durazno son: Las Vueltas, Cruz de Piedra, Las Mesas, Plan de San Francisco, Chiltepec,

Bertha Sofía Larqué Saavedra et al.306 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
número de árboles plantados; 42 200, con un promedio de árboles por productor de 796.23; 46% de los productores alcanzan este promedio e incluso más. Puede interpretarse que en esta localidad, existe una mayor distribución de árboles por productor.
El dato promedio más alto de árboles por localidad está en Chiltepec, seguido de Las Mesas y El Potrero en tercer lugar. Del total de estas localidades, esta última es la que registra la menor concentración de árboles por productor, mientras que Plan de San Francisco tiene mayor concentración (SAGAPA, 2006a-b).
Localidad Rango mínimo y superior de árboles por productor
Rango mínimo y superior de (ha) por productor
Chiltepec 200-7 200 0.75-12.00Cruz de Piedra 200-2 500 0.50-5.00Ixtlahuaca n d 1.00-10.00El Potrero 500-2 500 0.50-30.00Plan de San Francisco 50-1 000 0.25-1.50Las Vueltas 300-1 200 0.25-3.40Las Mesas 600-2 700 1.00-6.001ª y 2ª de San Miguel 120-1 500 0.08-4.00San José 400-3 500 1.00-15.00Cochisquilla 50-4 000 0.25-2.00
Cuadro 4. Total de productores, árboles plantados y hectáreas sembradas de durazno, por localidades del municipio de Coatepec de Harinas, Estado de México, México.
Localidad Productores Árboles ( ha- 1)
Chiltepec 47 57 130 113.50Cruz de Piedra 74 41 900 170.75Ixtlahuaca 61 n d 222.00El Potrero 53 42 200 239.75Plan de San Francisco 39 12 100 29.25Las Vueltas 68 34 200 76.45Las Mesas 14 19 500 31.001ª y 2ª de San Miguel 14 3 690 14.16San José 8 5 100 43.00Cochisquilla 51 17 940 42.95
Cuadro 3. Localidades del municipio de Coatepec de Harinas, Estado de México, México, productoras de durazno, 2007.
Fuente: trabajo de campo, 2007 (SIAP, 2006; PRSPD, 2007).
Como se puede observar en el Cuadro 3, el número de árboles por productor varían de 50 en Cochisquilla y Plan de San Francisco, a 7 200 en Chiltepec. Esta última localidad es la que registra el mayor número de árboles plantados, seguido por El Potrero. Asimismo, tiene superioridad en el número hectáreas sembradas de durazno (Ixtlahuaca no reportó datos en este renglón). La localidad con mayor número de productores es Cruz de Piedra (Cuadro 4).
Chiltepec tiene el más alto promedio de árboles por productor de 1 215.5, y solamente 14% de los productores alcanzan esta cantidad o más. El Potrero es la segunda localidad en
Fuente: trabajo de campo, 2007 (PPMNF, 2007; Comité Estatal de Sanidad Vegetal del Estado de México, México, 2007).
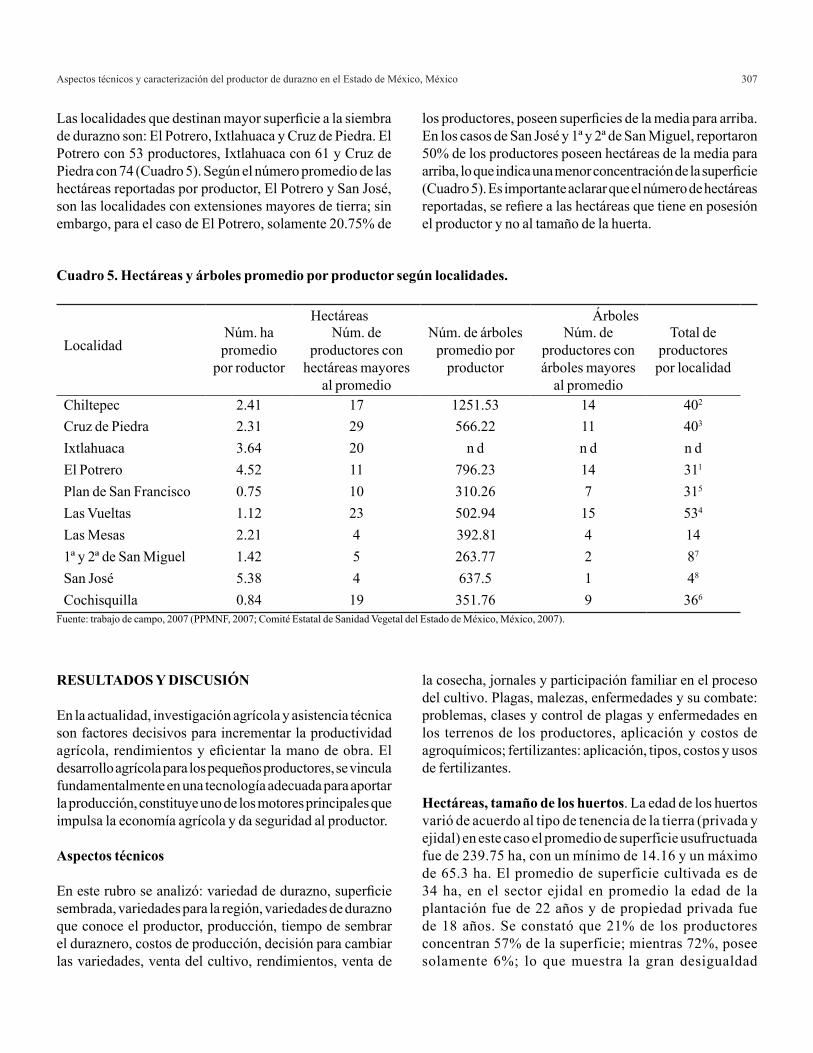
Aspectos técnicos y caracterización del productor de durazno en el Estado de México, México 307
la cosecha, jornales y participación familiar en el proceso del cultivo. Plagas, malezas, enfermedades y su combate: problemas, clases y control de plagas y enfermedades en los terrenos de los productores, aplicación y costos de agroquímicos; fertilizantes: aplicación, tipos, costos y usos de fertilizantes.
Hectáreas, tamaño de los huertos. La edad de los huertos varió de acuerdo al tipo de tenencia de la tierra (privada y ejidal) en este caso el promedio de superficie usufructuada fue de 239.75 ha, con un mínimo de 14.16 y un máximo de 65.3 ha. El promedio de superficie cultivada es de 34 ha, en el sector ejidal en promedio la edad de la plantación fue de 22 años y de propiedad privada fue de 18 años. Se constató que 21% de los productores concentran 57% de la superficie; mientras 72%, posee solamente 6%; lo que muestra la gran desigualdad
Las localidades que destinan mayor superficie a la siembra de durazno son: El Potrero, Ixtlahuaca y Cruz de Piedra. El Potrero con 53 productores, Ixtlahuaca con 61 y Cruz de Piedra con 74 (Cuadro 5). Según el número promedio de las hectáreas reportadas por productor, El Potrero y San José, son las localidades con extensiones mayores de tierra; sin embargo, para el caso de El Potrero, solamente 20.75% de
Localidad
Hectáreas ÁrbolesNúm. ha promedio
por roductor
Núm. de productores con
hectáreas mayores al promedio
Núm. de árboles promedio por
productor
Núm. de productores con árboles mayores
al promedio
Total de productores por localidad
Chiltepec 2.41 17 1251.53 14 402
Cruz de Piedra 2.31 29 566.22 11 403
Ixtlahuaca 3.64 20 n d n d n dEl Potrero 4.52 11 796.23 14 311
Plan de San Francisco 0.75 10 310.26 7 315
Las Vueltas 1.12 23 502.94 15 534
Las Mesas 2.21 4 392.81 4 141ª y 2ª de San Miguel 1.42 5 263.77 2 87
San José 5.38 4 637.5 1 48
Cochisquilla 0.84 19 351.76 9 366
los productores, poseen superficies de la media para arriba. En los casos de San José y 1ª y 2ª de San Miguel, reportaron 50% de los productores poseen hectáreas de la media para arriba, lo que indica una menor concentración de la superficie (Cuadro 5). Es importante aclarar que el número de hectáreas reportadas, se refiere a las hectáreas que tiene en posesión el productor y no al tamaño de la huerta.
Cuadro 5. Hectáreas y árboles promedio por productor según localidades.
Fuente: trabajo de campo, 2007 (PPMNF, 2007; Comité Estatal de Sanidad Vegetal del Estado de México, México, 2007).
RESULTADOS Y DISCUSIÓN
En la actualidad, investigación agrícola y asistencia técnica son factores decisivos para incrementar la productividad agrícola, rendimientos y eficientar la mano de obra. El desarrollo agrícola para los pequeños productores, se vincula fundamentalmente en una tecnología adecuada para aportar la producción, constituye uno de los motores principales que impulsa la economía agrícola y da seguridad al productor.
Aspectos técnicos
En este rubro se analizó: variedad de durazno, superficie sembrada, variedades para la región, variedades de durazno que conoce el productor, producción, tiempo de sembrar el duraznero, costos de producción, decisión para cambiar las variedades, venta del cultivo, rendimientos, venta de

Bertha Sofía Larqué Saavedra et al.308 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
en la tenencia de la tierra y obviamente en los ingresos. El rendimiento también presenta una gran variación, tomando valores entre 2.5 y 25 ha.
Manejo agronómico. En el proceso de las actividades de labor que realizan los productores, se detectó mayor diversidad en la fertilización. Se encontraron diferencias en los fertilizantes que se emplean, así como en cantidades y fechas de aplicación. Los fertilizantes más usados fueron 18-46-00, 14-00-40, triple 16, triple 18, urea, potasio, fósforo, fosfonitrato, cloruro de potasio, micro elementos como zinc y magnesio, cal agrícola y abono orgánico de res.
En la utilización del cultivar diamante, la variedad más usada, compran los injertos en viveros del mismo municipio
Coatepec de Harinas, donde trabajan con semillas mejoradas y certificada de Estados Unidos de América y los patrones son Nemaguard resistentes a nemátodos; en viveros del municipio de Temascaltepec o bien, en Uruapan Michoacán.
El sistema de conducción que se presentó con mayor frecuencia en los casos estudiados fue el de tatura. Cuentan con riego rodado, aspersión y goteo. Se observó que existen varias huertas, donde siembran durazno con aguacate, esta actividad baja el rendimientode ambos frutos.
En el Cuadro 6 se observa el diseño de la huerta y densidad por árbol, fertilización, podas y rendimientos de durazno en Coatepec de Harinas.
Diseño de huerta y densidad (árbol ha)
Fertilización Podas y riegos Rendimiento
1. Marco realD e n s i d a d . 4 0 0 á r b o l e s a u n a distancia de 3.50*4 m. Altura de árbol de 2.50 m
Al plantar 250 g de 18-40-00. 1º año fertilizan dos veces con 250 g de 18-46-00. Al 2º año dos veces, una en secas y otra en temporada de lluvia junto con la primera poda. Fertiliza con abono orgánico. Del 3º año en adelante por árbol una mezcla 18-46-00 y triple 18.
Realiza una poda de formación en el 1º y 2º año de vida en octubre o noviembre. La poda de fructificación la realiza a partir del 3º año.
20 kg por árbol a part ir del tercer año.
2. Marco realD e n s i d a d . 5 0 0 á r b o l e s a u n a distancia de 3*4. Altura de árbol 3 m.
Al plantar no fertiliza. En el 1º y 2º año de vida aplica por árbol, tres dosis de 200 g de 18-46-00 y urea. Del 3º año en adelante aplica los mismos fertilizantes, con una dosis de 1.5 kg por árbol. Cuando está en producción coloca abono de res de 15 a 20 kg por árbol
Realiza una poda de formación entre agosto, septiembre y octubre. Las podas de rejuvenecimiento son esporádicas, solamente cuando el árbol crece mucho. Si un árbol de siete años produce durazno “canica” se tiene que hacer una poda de desahije.Cuenta con sistema de riego por micro aspersión y rodado. De diciembre a marzo realiza uno cada quince días con una duración de 3 h por riego. Con el riego rodado, uno cada 25 días o al mes, y deja correr el agua por espacio de 20 min.
20 kg a los tres años del árbol. A los cinco años, de 40 a 50 kg por árbol.
3. Marco realDis tanc ia en t re árboles de 3*5 m
La fertilización la realiza sólo en la época de producción. Aplica un kg de Triple 18, combinando con 100 g de cal y lama de res.
Realiza podas de control de crecimiento y una anual de fructificación.Lleva a cabo seis riegos anuales en los meses de enero a mayo, de tres h cada uno.
21 t ha-1
Cuadro 6. Diseño, densidad, fertilización, podas y rendimiento del municipio de Coatepec de Harinas, Estado de México, México. 2007.
Fuente: trabajo de campo 2007.

Aspectos técnicos y caracterización del productor de durazno en el Estado de México, México 309
Aspectos socioeconómicos Edad del productor. La edad de los productores varió de 35 a 77 años, con un promedio de 45. Una de las variables que se consideró de importancia fue la edad, autores como Damián (2007), señala en este sentido, en un estudio realizado en el estado de Tlaxcala, sobre apropiación de tecnología agrícola, que los productores encuestados con respecto a la escolaridad la apropiación se basa en la edad, el nivel de escolaridad tienen una alta apropiación, tierra, desempeño de otras actividades complementarias, relevancia de las técnicas campesinas; así como acceden a folletos y revistas técnicas con información agrícola. Se encontró que la edad del productor son determinantes para las prácticas agronómicas y esto presenta un impacto en el rendimiento del cultivo, como ha sido observado por otros autores (Ruiz et al. 2001; INEGI, 2002; Rueda, 2003).
Crédito y comercialización. El enfoque tradicional, de programas de crédito agrícola dirigidos por la oferta, concibe al crédito como un insumo adicional a la producción, utilizado para subordinar las decisiones de los productores a las propuestas de los proyectos. La noción que los préstamos son indispensables para acelerar el cambio tecnológico en la agricultura, sin embargo, el efecto productivo del crédito es altamente discutible y difícil de medir, porque forzosamente deben ir asociados a la incorporación de tecnología adecuada al cultivo. La experiencia en este rubro muestra, con abundantes ejemplos, que la aplicación de este enfoque ha tenido como resultados: a) instituciones crediticias fracasadas (crisis financieras en la banca como el extinto Banrural, etc.), b) altos costos de transacción y baja calidad de los servicios crediticios para los prestatarios, c) los recursos crediticios no siempre han llegado a sus poblaciones objetivo, y d) el acceso a crédito ha sido suspendido una vez concluidos los programas oficiales.
Solamente el 36% de los agricultores obtuvo apoyo de los programas gubernamentales en 2007. Las instituciones o programas que otorgó el apoyo gubernamental a los agricultores son de instituciones como: PROCAMPO quien ha proporcionado el apoyo en 16%, PROGRESA y KILO x KILO en 10% respectivamente, 8% ACERCA, SEFOA-SAGARPA 4% y por último el CADER con el resto de los productores con apoyo. El promedio obtenido por los apoyos fue de $ 22 471.42, máximo 16 900, y mínimo $2 800.
Organización de productores. Una observación fundamental como resultado de las entrevistas y cuestionarios es la falta de organización de los productores, ya que se enfrentan a los intermediarios que son los que definen el precio al producto y les afecta en gran medida a los costos de producción.
Factores que limitan la comercialización de durazno. El 68.5% de los productores respondió que una de las limitantes a las que enfrenta en la comercialización de durazno es: el producto tiene daños por insectos, magulladuras, rajaduras, granizo y otros que afectan la calidad de la fruta. Con relación a la rentabilidad del duraznero, 58% respondieron que es rentable y que si tuvieran extensiones de tierra más grandes sembrarían más duraznos.
De los productores entrevistados, 32.7% contó con la asistencia técnica, y 67.3% restante no tuvo acceso a este servicio. De los agricultores que recibieron asistencia técnica; 61.5% participa en alguna organización comunal. Los productores que no recibió este servicio (asistencia técnica) el (58.3%), tiene algún cargo en la comunidad (comisariado ejidal, representante de las fiestas patronales, topil, entre otros).
Expectativas de los campesinos. Las crisis económicas y agrícolas que se ha padecido en el sector rural, desde el principio de la década de 1970, han evidenciado el agotamiento del modelo de Industrialización de Sustitución de Importaciones. La implementación del nuevo modelo de desarrollo secundario exportador por medio de las políticas de ajuste fondomonetaristas, aplicadas durante los últimos tres sexenios ha enfrentado serias dificultades mostradas por la inestabilidad y el estancamiento económico. La aplicación continua de la “políticas de ajuste” ha conducido a un cambio estructural. En este contexto se profundiza la crisis de la transferencia de tecnología que vive el sector agrícola mexicano desde principios de 1980. Lo anterior, ha propiciado una problemática que los productores observan que existe en torno al cultivo de durazno. Refieren: no existen apoyos gubernamentales, mucha burocracia en las instituciones, pocos créditos, e inoportunos, por ejemplo, llegan cuando ya están cosechando, costos de producción muy altos, y bajos rendimientos. También se presenta una queja permanente de los agricultores que no son escuchados por las instituciones y mucho menos en la toma de decisiones sobre los programas gubernamentales que afectan su vida cotidiana y su agricultura.

Bertha Sofía Larqué Saavedra et al.310 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
CONCLUSIONES
La caracterización del productor permitió conocer que existen disparidad entre las superficies cultivadas de durazno, el manejo técnico del cultivo y las formas de comercializar el producto.
Los problemas técnicos que enfrenta el sector ejidal son similares a los de la pequeña propiedad; sin embargo, el número de productores es mayor en el primer grupo. Los elementos requeridos para disminuir el factor de restrictivos que enfrenta el sistema productivo se refieren a la necesidad de realizar un programa de transferencia de tecnología y capacitación continuo, que detecte las necesidades del productor, con la finalidad de mejorar la producción y la productividad, así como minimizar los riesgos que se tienen en el sistema-producto.
Las prácticas agrícolas en la región son, en términos generales, eficientes, sin embargo, el productor requiere de paquetes tecnológicos que le permitan hacer frente a los aspectos de control de plagas malezas y enfermedades en su cultivo.
El 95% de los productores refirió que una de las limitantes es la falta de organización para la venta de su producto. Organizados tratarían de mantener el rumbo del proyecto que buscaría una vida mejor para los socios y sus familias, una mayor autonomía en la toma de decisiones, la conservación de los recursos productivos y una relación más equitativa con el mercado.
LITERATURA CITADA
Claridades Agropecuarias. 2000. El durazno mexicano. www.infoaserca.gob.mx. (fecha de consulta, diciembre de 2000).
Comité Estatal de Sanidad Vegetal del Estado de México. Temporada 2006-2007. Relación de productores inscritos en la campaña de la mosca de la fruta. (Documento inédito).
Damián, H. M. A. 2007. Apropiación de tecnología por actividades del ciclo agrícola del maíz. In: apropiación de tecnología agrícola. Damián, H. M. A (coord.) Benemérita Universidad Autónoma de Puebla- CONACYT-Siza- H. Congreso del Estado Tlaxcala, LVII Legislatura, Puebla, Puebla.
De Grammont, H. 2006. La nueva estructura ocupacional en los hogares rurales mexicanos: de la unidad económica campesina a la unidad familiar pluriactiva. Asociación Latinoaméricana de Sociología Rural (ALASRU). Quito, Ecuador. p 42.
Díaz, M. D. H. 1987. Requerimiento de frío en frutales caducifolios. INIFAP-SARH. México, D. F. 54 p. (Tema Didáctico Núm. 2).
Duch, G. J. 1998. Tipologías empíricas de productores agrícolas y tipos ideales en el estudio de la agricultura regional. Revista de Geografía Agrícola. 57:27-38.
United States Departament of Agriculture (USDA). 2005. Economic Research Service (ERS). http/: www. Fas.usda. go Vustrade (fecha de consulta, mayo de 2005).
Ekboir, J. 2004. Can impact analysis be used for research evaluation? Centro Internacional del Mejoramiento del Maíz y Trigo, El Batán, Texcoco, Estado de México, México, 29 p.
Escalante, R. 2006. Desarrollo rural regional y medio ambiente. Economía. Universidad Nacional Autónoma de México (UNAM), México, D. F. 3(8):69-94.
García, E. 2004. Modificaciones al sistema de clasificación de Köppen. 5ª. Ed. Instituto de Geografía- Universidad Nacional Autónoma de México. México, D. F. 90 p.
Gutiérrez, A. F. 2006. Características de selecciones s o b r e s a l i e n t e s d e d u r a z n o c r i o l l o e n Aguascalientes.
Gutiérrez, A. y Padilla. R. 2007 Rendimiento y calidad del fruto de durazno tipo San Gabriel de maduración temprana. Campo Experimental Pabellón. Aguascalientes-Zacatecas. http://www.inifap.gob.mx/otros_sitios/enero_junio2007.pdf. (fecha de consulta, octubre de 2007).
Coatepec de Harinas, Estado de México. Plan de Coatepec de Harinas, Estado de México. http://seduv.edomexico.gob.mx/planes_municipales/Coatepec de Harinas /Documento del plan de Coatepec de Harinas.pdf. (consultada febrero de 2007).
Instituto Nacional de Estadística Geografía e Informática. (INEGI). 2002. Censo general de población y vivienda. Información de cabeceras municipales. México, D. F.
Padrón de Productores contra Moscas Nativas de la Fruta (PPMNF). 2007. Municipio de Coatepec de Harinas (Documento inédito).

Aspectos técnicos y caracterización del productor de durazno en el Estado de México, México 311
Pérez, G. S. 2004. Manual para cultivar duraznero. Ed. Limusa. 108 p.
Plan Municipal de Desarrollo Urbano de Coatepec de Harinas Estado de México.
Plan Municipal de Desarrollo Urbano de Temascaltepec. Estado de México. Tomado de: http://www.e-mexico.gob.mx/work/méxico/mpios/15086a.htm) (fecha de consulta, febrero de 2008).
Plan Rector Sistema Producto Durazno (PRSPD). http://www.amsda.com.mx/preestatales/estatales/durazno.pdf (fecha de consulta, octubre 2007).
Rueda, B. M. C. 2003. Evaluación de variedades de durazno (Prunus persica L. Batsch) en el noreste del estado de Morelos. Tesis de Licenciatura- Fitotecnia. Universidad Autónoma Chapingo. pp. 6-22.
Ruiz, C. J. A.; Medina, G. G.; González, A. I. J.; Ortiz, T. C.; Flores, L. H. E.; Martínez, P. R. A. y Byerly, M. K. F. 2001. Requerimientos agroecológicos de cultivos. INIFAP-SAGAR. Guadalajara, Jalisco, México. 324 p. (Libro Técnico Núm. 3).
Secretaría de Agricultura Ganadería, Desarrollo Rural, Pesca y Alimentación. (SAGARPA) 2006a.Tomado de: http://w4.siap.gob.mx/sispro/SP_AG/sp_durazno.html. (fecha de consultada, agosto 2007).
Secretaría de Agricultura Ganadería, Desarrollo Rural, Pesca y Alimentación. (SAGARPA) 2006b. Anuario estadístico de la producción agropecuaria para el Estado de México. México. 250 p.
Secretaría de Agricultura Ganadería, Desarrollo Rural, Pesca y Alimentación. (SAGARPA). 2006c. Anuario estadístico de la de la producción agrícola. Servicio de información agroalimentaria y pesquera. México, D. F. (fecha de consulta, mayo 2006).
Servicio de Información Agroalimentaria y Pesquera (SIAP). 2006. Estadísticas sistema-producto-durazno. www.siap.sagarpa.gob.mx. (fecha de consulta, febrero 2007).
Sukhatme, P y Balkrishna, V. S. 1970. Sampling theory of surveys with applications. Iowa State University Press. Ames, Iowa, USA. 358 p.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 312-319
CARACTERIZACIÓN DE SELECCIONES DE GUAYABA PARA EL BAJÍO DE GUANAJUATO, MÉXICO*
CHARACTERIZATION OF GUAVA SELECTION FOR THE BAJIO REGION OF
GUANAJUATO, MEXICO
Candelario Mondragón Jacobo1§, Luis Miguel Toriz Ahumada1 y Salvador Horacio Guzmán Maldonado1
1Campo Experimental Bajío, INIFAP. Km 6.5 carretera Celaya-San Miguel de Allende. C. P. 38110. Celaya, Guanajuato, México. Tel. 01 461 611 53 23. ([email protected]), ([email protected]). §Autor para correspondencia: [email protected].
RESUMEN
En México, la producción y consumo de guayaba (Psidium guajava L.) se basa en las variedades tradicionales “Media China” y “Peruana” entre otras, seleccionadas por los productores a partir de germoplasma de la región semiárida del centro norte de México. El objetivo de este estudio fue caracterizar siete selecciones de guayaba de pulpa rosa y una de pulpa blanca desarrolladas por el programa de fruticultura del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) para la región del Bajío de Guanajuato. El estudio se efectuó en el Campo Experimental Bajío, Celaya Guanajuato, México, durante 2007 y 2008. Los árboles provinieron de semillas derivadas de la polinización abierta en una colección que contiene árboles de semilla de Brasil y México. La colección de estudio consistió de 248 árboles que representan la segunda generación de selección individual. Se determinaron las características de ocho selecciones y dos testigos comerciales. Los caracteres de planta estudiados fueron: productividad, vigor, fenología, tolerancia a la enfermedad conocida como “clavo” (Pestalotiopsis psidii) y tolerancia a baja temperatura (heladas). Los caracteres de fruto fueron: color de pulpa, peso, calibre, forma, contenido de semilla, acidez titulable, sólidos solubles totales, vitamina C y licopeno. Las selecciones de pulpa rosa y salmón fueron: 23-1, 26-8, 23-5, 25-5, 21-17, 22-11 y 25-9 y la de pulpa blanca: 21-9. Las selecciones con pulpa color rosa mostraron: frutos
grandes, de 118.4 a 207.6 g, produjeron de 6.5 a 34.7 kg árbol, contenido intermedio de sólidos solubles totales 10.1 a 13.1 ºBrix, de 83.29 a 185.75 mg 100 g de vitamina C, de 0.54 a 3.28 mg 100 g de licopeno con época de cosecha del 1 de septiembre al 15 de noviembre. Comparativamente, el testigo Media China mostró frutos amarillos de 48 g en promedio, con 11.6 ºBrix, 231.49 mg 100 g de vitamina C, 0.53 mg 100 g de licopeno y con época de cosecha de noviembre a enero. Las selecciones estudiadas pueden complementar a las variedades comerciales en cuanto a tipo de fruta y época de cosecha. Las características de tamaño, apariencia visual, contenido de semilla del fruto, ºBrix y licopeno de las selecciones compiten con las de las variedades comerciales.
Palabras clave: Psidium guajava L, calidad de fruta, mejoramiento genético de guayaba, vitamina C.
ABSTRACT
Guava (Psidium guajava L.) production and consumption in Mexico is based on traditional varieties types “Media China” and “Peruana” among others. They were selected by farmers using the mexican germplasm available in the central semiarid region of Mexico. The objective of this
* Recibido: Enero, 2009
Aceptado: Septiembre, 2009

Caracterización de selecciones de guayaba para el Bajío de Guanajuato, México 313
study was to characterize eight guava selections: seven with pink flesh and one of white flesh, developed by the horticulture program of the National Research Institute for Forestry, Agriculture and Livestock (INIFAP). The study was conducted at the Bajio research station, Celaya, Guanajuato, during 2007 and 2008. The selected trees were grown from seed obtained from an open pollinated collection of guava trees from Brazil and Mexico. The studied trees represented the second generation of individual selection. The plant characters studied were: vigour, phenology, tolerance to the disease locally known as “clavo” (Pestalotiopsis psidii) and low temperatures. The fruit characters were: flesh color, weight, shape, seed content, titratable acidity, total solid content, C vitamin and lycopene. Seven selections of pink flesh color were characterized, their IDs are: 23-1, 26-8, 23-5, 25-5, 21-17, 22-11 and 25-9 and one with white flesh; 21-9. All pink selections show: large fruits, 118.4 to 207.6 g, produced from 6.5 to 34.7 kg per tree, its sugar content varied from 10.1 to 13.1 ºBrix, C vitamin from 83.29 to 185.75 mg 100 g and lycopene from 0.54 to 3.28 mg 100 g. Its natural harvest season extended from September 1 to November 15. In comparison, “Media China” shows a mean weight of 48 g per fruit, 11.6 ºBrix, 231.49 mg 100 g of C vitamin, 0.53 mg 100 g of lycopene and a natural harvest season from November to January. The studied selections can be a complement for the commercial varieties in terms of fruit type and harvest season. Fruit characters observed in the selections compete with those of the commercial varieties. Key words: Psidium guajava L., fruit quality, guava breeding, vitamin C.
INTRODUCCIÓN
La guayaba es una de las frutas más importantes en México, se cultiva en condiciones subtropicales y tropicales en 23 000 ha. La producción comercial se concentra en dos diferentes condiciones agroclimáticas, ambas localizadas en el centro de México; la región de Calvillo, Aguascalientes y el oriente del estado de Michoacán, conformado por los municipios de Jungapeo, Juárez, Zitácuaro, Susupuato, Tuxpan y Tuzantla, en los cuales se cultiva 87% de la superficie con este cultivo en el estado, (Aguilar et al., 2003). La guayaba es también uno de los frutales domésticos más populares en México. La guayaba se consume principalmente como fruta fresca (87 al 92%) y el resto en las industrias de bebidas, mermeladas y otros productos afines (González et al., 2002).
La cadena productiva “guayaba” se fundamenta en el cultivo de la variedad “Media China” (90% de la superficie) que se caracteriza por producir frutos de forma aperada, de tamaño pequeño a mediano (<80 g), con epidermis de color amarillo brillante en frutos maduros, con pulpa de color crema a amarilla de aroma agradable y alto contenido de semilla (González et al., 2000). Los árboles son productivos y adaptados a la época normal de producción o al manejo agronómico para inducir la producción fuera de temporada, usualmente en invierno.
La producción actual se destina casi en la totalidad al mercado nacional, esfuerzos gubernamentales recientes permitieron la exportación de un pequeño volumen de fruta a los EUA, la principal limitante para la exportación es la cuarentena contra mosca de la fruta impuesta a la guayaba de México. Otra barrera al producto nacional es la preferencia del mercado internacional por fruta de mayor tamaño y pulpa de color rosado y blanco, características que no tienen las variedades comerciales de México. La industria internacional también demanda pasta y puré de color rosado (SAGARPA, 2005). Un aspecto sobresaliente de la guayaba rosada es el contenido carotenoides, en particular de licopeno, el cual se ha demostrado previene enfermedades como el cáncer de próstata (Giovannucci et al., 2002). Sin embargo, los pocos estudios realizados sobre las propiedades nutracéuticas de la guayaba, se han centrado en la vitamina C y los carotenoides (Pal et al., 2004; Sanjinez-Argadoña et al., 2005). El determinar el contenido de licopeno en materiales de guayaba que puedan ser liberados como nuevas variedades dará valor agregado al producto.
Es conocido que en el germoplasma nativo existe el color rosado de pulpa y fruta de tamaño grande por separado; sin embargo, no se han desarrollado variedades que conjunten las dos características. El objetivo de este estudio fue caracterizar siete selecciones de guayaba de pulpa rosa y una de pulpa blanca desarrolladas por el programa de fruticultura del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) para la región del Bajío, Guanajuato.
MATERIALES Y MÉTODOS
El proceso de selección fue realizado en el Campo Experimental Bajío del Instituto Nacional de Investigaciones Forestales y Agrícolas y Pecuarias (INIFAP) ubicado en Celaya, Guanajuato, México cuya posición geográfica es

Candelario Mondragón Jacobo et al.314 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
20º 34’ 00” latitud norte y 100º 50’ 00” longitud oeste a 1 765 m. El clima del sitio es subtropical semiárido con 601 mm de precipitación anual promedio, con 85% de ocurrencia en el verano (Terrones et al., 2000). La clasificación del suelo en el sitio experimental es Vertisol pélico de color obscuro. Los árboles fueron plantados en la primavera de 2002, a densidad de 5x2.5 m (800 árboles ha), fueron conducidos a tronco único y mantenidos en forma de vaso ligeramente abierto por medio de podas anuales. La duración de la fase juvenil fue de 2 a 3 años, por lo que el rendimiento se registró hasta el 4º año, la información de rendimiento por árbol presentada en este trabajo incluye los años 2007 y 2008, una vez que este parámetro se estabilizó en todos los arboles.
Los árboles fueron obtenidos de semillas derivadas de la polinización abierta en una colección (iniciada en 1990) que contiene árboles de semilla originarios de Brasil y de México. La colección de estudio consistió de 248 árboles que representan la segunda generación de selección individual. Los criterios de selección para los caracteres de planta fueron: productividad, vigor, fenología, tolerancia a la enfermedad conocida como “clavo” (Pestalotiopsis psidii), hábito de crecimiento y tolerancia a baja temperatura (heladas). Los caracteres de fruto fueron: color de pulpa, peso de fruta, calibre, forma y contenido de semilla. Para las variables anteriores se usaron 20 frutos por planta. Del análisis químico se obtuvieron acidez, contenido de azúcares, vitamina C y licopeno, las determinaciones fueron hechas por triplicado en tres frutos por selección. Durante 2007 y 2008 no se aplicaron fungicidas para determinar la resistencia a “clavo” del fruto. Las siete selecciones representan 3.2% de la colección.
El análisis químico se efectuó con el siguiente procedimiento: los sólidos solubles totales (SST) se determinaron con un refractómetro manual (Atago N-1E) en una mezcla de pulpa, el pH del jugo con un potenciómetro usando pulpa sin diluir, la acidez fue calculada de acuerdo al método de la AOAC 942.15 (AOAC, 2000). La determinación de vitamina C se efectuó por medio de HPLC (Agilent 1100 series con DAD), se utilizó una columna ZORBAX Eclipse XDB-C18 de 5 µm de acuerdo al protocolo propuesto por Scartezzini et al. (2006) con dos modificaciones: 5 000 µL y 25 mg de muestra liofilizada. El licopeno se determinó con el mismo equipo de HPLC de acuerdo con el protocolo descrito por Sadler and Dezman, (1990) y
Klaus, (2002), se ajustó el número de extracciones de uno a tres en función de la intensidad del color en la muestra, el tamaño de muestra liofilizada en este caso fue de 500 mg, la temperatura del detector fue 25 °C a una longitud de onda de 502 nm. La muestra usada fue de 20 μl, la velocidad de flujo 1.5 ml min-1 y el tiempo 20 min.
RESULTADOS Y DISCUSIÓN
Caracterización de los árboles
Todas las selecciones mostraron árboles de vigor alto, expresado en crecimiento significativo de tronco, ramas principales gruesas y hojas grandes, típicas del germoplasma de guayaba de origen tropical, significativamente mayores que los observados en las variedades Media China y Peruana. El rendimiento de fruta por árbol se incrementó con la edad, desde el doble en las selecciones 25-5, 23-5 y 26-8 hasta casi cuatro veces en la selección 21-9. El rendimiento promedio más alto fue registrado en la selección de pulpa blanca 21-9 (54.87 kg árbol), mientras que la mejor selección de pulpa rosa fue la 23-5, todas ellas fueron superiores a los testigos comerciales Peruana y Media China. La guayaba es un árbol productivo en México, al grado de que el peso de la fruta puede dañar las ramas lo que reduce la calidad de la fruta y la vida productiva del árbol. Los factores asociados a la falla de la estructura del árbol son: alta productividad natural, falta de poda y raleo de fruta, esta última resulta impráctica en los sistemas de producción nacionales. Las siete selecciones mostraron estructura de copa con resistencia de moderada a fuerte al colapso de ramas (Cuadro 1).
Aunque la región Bajío está clasificada como subtropical, en los últimos cinco años del estudio se registraron temperaturas bajas de moderadas a severas, la guayaba no es tolerante a heladas (Manica et al., 2000) y los genotipos obtenidos de zonas tropicales son más susceptibles, lo cual se evidenció en este estudio, ya que se registraron daños hasta 100% en el follaje debido a baja temperatura. El menor daño se observó en la selección 22-5 con 85% de follaje afectado.
La tolerancia a “clavo” del fruto es muy importante en México, dado que es una enfermedad endémica que deteriora la apariencia del fruto y determina la aceptación en el mercado. Por lo anterior, el control químico es rutinario en

Caracterización de selecciones de guayaba para el Bajío de Guanajuato, México 315
las zonas productoras de guayaba. Las selecciones 21-9, 21-17, 23-1 fueron clasificadas como tolerantes debido a que presentaron daño mínimo, similar al observado en Media China, bajo las condiciones del estudio.
El hábito de fructificación de flor y fruto individual fue seleccionado como el mejor ya que el de racimo induce floración y fructificación extendida; además dificulta la cosecha de los primeros frutos.
fruto, respectivamente. En general, el número de semillas por fruto o la proporción de semilla respecto al peso del fruto fue bajo en las selecciones, el contenido de semilla observado en las selecciones no representa una limitante para la aceptación por el consumidor (Cuadro 2).
El color de la pulpa es una característica importante. El programa de mejoramiento genético de la guayaba del Campo Experimental Bajío, busca genotipos con color de pulpa diferente a crema-amarillo de las variedades tradicionales, de preferencia tonos de color rosado observados en las selecciones 23-1, 23-5, 25-5, 25-9 y 26-8, salmón (22-11) y combinaciones de amarillo/rosa como la 21-17, que podrían añadir diversidad al mercado. (Cuadro 2) y (Figura 1).
Selección o variedad
Rend. 2007 (kg árbol)
Rend. 2008 (kg árbol)
Daño de helada (%)
Tolerancia a clavo
Vigor Hábito de fructificación
Resistencia carga de fruta
Época de cosecha
21-9 13.249 55. 361 100 A In Individual Alta Oct.
21-17 9.843
48.108 100 A H
Individual Alta Oct.-Nov. 15
22-11 12.606 41.594 90 I In Individual Moderada Sep.-Oct.
23-1 6.574
26.594 100 A In
Individual Alta Sep. 15-Nov. 15
23-5 34.699 75.057 100 A H Racimo Moderada Oct.-Nov. 15
25-5 9.503
22.549 85 I In
Individual Alta Sep. 15-Nov.
25-9 11.056
45.616 100 I In
Individual Alta Sep. 15-Nov. 15
26-8 11.004
25.786 100 I In
Individual Moderada Sep. 15-Nov. 15
Peruana(t) NA 33.850 100 B In Individual AltaOct.-Nov.
Media China(t) NA 24.167 100 A In Racimo Alta Nov.-Ene.
Cuadro 1. Caracteres agronómicos de selecciones de guayaba. Celaya, Guanajuato.
Rend.= rendimiento por árbol; A= alto; I= intermedio; B= bajo; H= alto vigor; In= vigor intermedio; t= testigo.
Caracteres de fruto
Todas las selecciones produjeron frutos más grandes que los testigos, los frutos más pequeños observados en la selección 23-5 pesaron el doble que Peruana, lo cual puede representar un ventaja porque en el mercado nacional no se han ofertado frutos de tamaño grande; en contraste, en otras frutas como la tuna, el tamaño es el principal criterio de calidad (Mondragón, 2001). Una posible desventaja del tamaño grande es la necesidad de empaque especial debido a que el calibre de la fruta de estas selecciones podría requerir empaque individual. Dos de las selecciones: 21-9 y 21-17,produjeron frutos de forma aperada; el resto, redondos o casi redondos. La selección 25-5 mostró menor cantidad de semilla que Media China 1.2 y 1.5 g
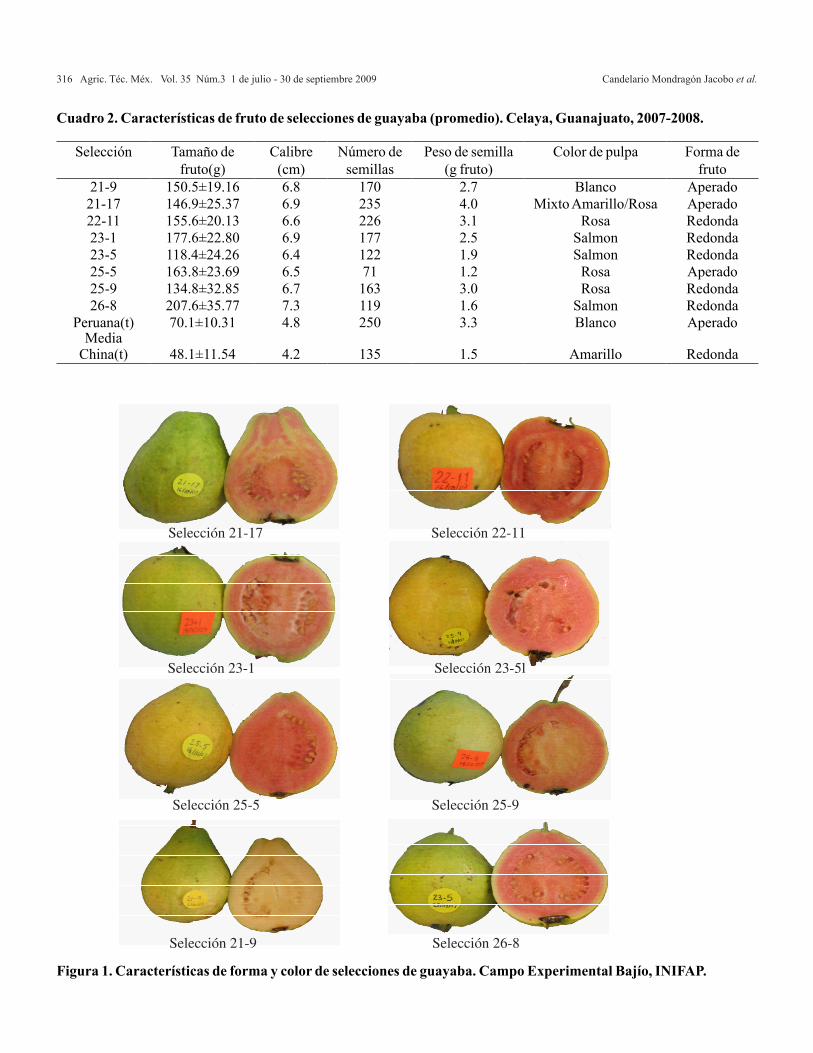
Candelario Mondragón Jacobo et al.316 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Selección Tamaño de fruto(g)
Calibre (cm)
Número de semillas
Peso de semilla (g fruto)
Color de pulpa Forma de fruto
21-9 150.5±19.16 6.8 170 2.7 Blanco Aperado 21-17 146.9±25.37 6.9 235 4.0 Mixto Amarillo/Rosa Aperado 22-11 155.6±20.13 6.6 226 3.1 Rosa Redonda 23-1 177.6±22.80 6.9 177 2.5 Salmon Redonda 23-5 118.4±24.26 6.4 122 1.9 Salmon Redonda 25-5 163.8±23.69 6.5 71 1.2 Rosa Aperado 25-9 134.8±32.85 6.7 163 3.0 Rosa Redonda 26-8 207.6±35.77 7.3 119 1.6 Salmon Redonda
Peruana(t) 70.1±10.31 4.8 250 3.3 Blanco Aperado Media
China(t) 48.1±11.54 4.2 135 1.5 Amarillo Redonda
Cuadro 2. Características de fruto de selecciones de guayaba (promedio). Celaya, Guanajuato, 2007-2008.
Figura 1. Características de forma y color de selecciones de guayaba. Campo Experimental Bajío, INIFAP.
Selección 23-1 Selección 23-5l
Selección 21-17 Selección 22-11
Selección 25-5 Selección 25-9
Selección 21-9 Selección 26-8

Caracterización de selecciones de guayaba para el Bajío de Guanajuato, México 317
Análisis químico
Los valores de pH observados fueron similares a los reportados por Marquina et al. (2008). El contenido de azúcares fue similar al de otras variedades de guayaba y otras frutas silvestres poco conocidas como el garambullo
(Mirtillocactus geometrizans) (Hernández-López et al., 2008) aunque más bajo que el de otras frutas tradicionales en México como la tuna (Mondragón y Gallegos, 2007). En promedio, las nuevas selecciones mostraron menor contenido de sólidos solubles totales (SST) que Media China y Peruana, pero dentro del rango comercialmente aceptable (Cuadro 3).
asumiendo una bio-disponibilidad de 30% por diversos factores, una ración de 100 g de cualquiera de las selecciones cubrirán entre 20 y 35% de este requerimiento.
Asumiendo que una persona consuma un fruto de 150 g (peso promedio del fruto de las selecciones) cubriría entre 30 y 50% del requerimiento diario de vitamina C. El contenido de licopeno observado en todas las selecciones de guayaba fue menor al reportado para la variedad 'Horana Red' (45 mg 100 g), (Chandrika et al., 2008) (Cuadro 3). Es importante mencionar que estos autores reportaron el contenido total de carotenoides que incluyen además del licopeno, el beta-caroteno y beta-criptoxantina las cuales no fueron determinadas en este estudio. Por esta razón, se puede inferir que el contenido de licopeno en la variedad Horana Red fue sobre estimado. El contenido de licopeno observado en las selecciones fue similar al reportado en tomate de color amarillo a anaranjado. El tomate de color rojo intenso contiene hasta 9 mg 100 g (Arias et al., 2000) es decir, hasta tres veces el observado en la variedad Peruana de guayaba. Sin embargo, no se ha determinado la dosis de licopeno requerida para que ejerza efecto preventivo sobre el cáncer de próstata, se recomienda que el consumo sea lo más alto posible.
Selección pH SST ºBrix AT (%) AA (mg 100 g) Licopeno (mg 100 g)21-9 4.0±0.04 11.4±0.67 0.53±0.12 139.59±5.59 0.54±0.1421-17 4.2±0.20 13.1±0.19 0.67±0.27 149.94±0.85 1.99±0.0622-11 4.0±0.11 11.6±0.69 0.58±0.05 185.75±7.81 2.20±0.0223-1 3.9±0.03 10.1±0.70 0.52±0.09 121.11±2.31 1.06±0.01423-5 4.0±0.02 11.9±1.97 0.53±0.02 116.51±4.95 1.64±0.0025-5 3.9±0.58 12.9±2.06 0.38±0.06 121.31±7.81 2.77±0.0525-9 3.7±0.31 10.5±2.48 0.64±0.14 124.59±4.48 2.09±0.0626-8 4.1±0.19 12.1±0.12 0.45±0.05 83.29±2.23 3.28±0.13
Peruana(t) 4.0±0.11 13.7±0.92 0.90±0.19 129.74±4.66 NDMedia China(t) 4.0±0.06 11.6±0.97 0.77±0.00 231.49±11.31 0.53±0.01
Cuadro 3. Caracterización química de selecciones de guayaba. Celaya, Guanajuato, 2007-2008.
SST= sólidos solubles totales; AT= acidez titulable; AA= ácido ascórbico (vitamina C).
Con excepción de la variedad Peruana, la acidez titulable (AT) de las selecciones estudiadas fue menor comparada con los valores reportados por Marquina et al. (2008). La AT es un parámetro que estima el contenido de ácidos orgánicos en frutas (Hernández-Pérez et al., 2005). El bajo contenido de ácidos orgánicos observado puede disminuir el contenido de compuestos volátiles los cuales imparten el aroma característico a la guayaba. Esta puede ser una característica positiva dado que algunos mercados internacionales rechazan el fuerte aroma de esta fruta.
Las diferencias en el contenido de SST y la AT observadas entre las selecciones y los testigos podrían deberse a sutiles desigualdades en el proceso de madurez. El testigo “Media China” y la selecciones 21-9, 21-17 y 22-11 mostraron alto contenido de vitamina C comparados con los valores observados por Pal et al. (2004); quienes reportaron 140 mg 100 g de fruta en guayaba (Cuadro 3). Se ha reportado diferente contenido de vitamina C en variedades y selecciones de guayaba como resultado de diferencias en la maduración, dado que el contenido de esta vitamina decrece durante el proceso (Carvalho et al., 2008). El requerimiento mínimo de vitamina C para un adulto es de 60 mg diarios,

Candelario Mondragón Jacobo et al.318 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
CONCLUSIONES
Las selecciones estudiadas pueden complementar a las variedades comerciales en cuanto a tipo de fruta y época de cosecha, ya que el inicio de cosecha ocurre hasta seis semanas antes que la variedad Media China y dos semanas antes que la variedad Peruana.
Las características de tamaño, apariencia visual, contenido de semilla del fruto, sólidos solubles totales y licopeno observadas en las selecciones compiten con las de las variedades comerciales.Los frutos de guayaba con colores de pulpa rosa, salmón o blanca podrían ocupar el nicho de mercado que actualmente no se aprovecha.
LITERATURA CITADA
Aguilar, A. A.; Mendoza, L. R. M. y Mena, Ch. J. 2003. Comportamiento del guayabo en la zona oriente de Michoacán. Memoria del 1er. Simposio Internacional de la Guayaba-Guava-Goiabeira. Aguascalientes, Aguascalientes. México. p. 124-146.
Arias, R. L.; Tung-Ching, L.; Logan, L. and Harry, J. 2000. Correlation of lycopene measured by HPLC with the L*, a*, b* color readings of a hydroponic tomato and the relationship of maturity with color and lycopene content. J. Agric. Food Chem. 48:1697-1702.
Official Methods of Analysis (AOAC). 2000. Method 142.15. Acidity (Titratable) of fruit products.
Carvalho, A. B.; de Assis, S. A.; Leite, K. M.; Bach, E. E. and Oliveira, O. M. 2008. Pectin methylesterase activity and ascorbic acid content from guava fruit, cv. Predilecta, in different phases of development Int. J. Food Sci. Nutr. 12:1-11.
Chandrika, U. G.; Fernando, K. S. and Ranaweera, K. K. 2008. Carotenoid content and in vitro bioaccessibility of lycopene from guava (Psidium guajava) and watermelon (Citrullus lanatus) by high-performance liquid chromatography diode array detection. Int. J. Food Sci. Human Nutr. 18:1-9.
Giovannucci, E.; Rim, E. B.; Liu, Y.; Stampfer, M. J. and Willett, W. C. 2002. Prospective study of tomato products, lycopene, and prostate cancer risk. J. Cancer Nat. Inst. 94:391-398.
González, G. E.; Padilla, R. J. S.; Esquivel, V. F.; Robles, E. F. J. y Perales, C. M. A. 2000. Tecnología para producir guayaba en Calvillo, Aguascalientes. CIRNE- Campo Experimental Pabellón. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Pabellón. (Folleto para productores Núm. 28).
González, G. E.; Padilla, R. J. S.; Reyes, M. L.; Perales de la C., A. y Esquivel, V. F. 2002. Guayaba su cultivo en México. CIRNE- Campo Experimental Pabellón. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Pabellón, Aguascalientes. (Libro Técnico Núm. 1).
Hernández-Pérez,T.; Carrillo-López, A.; Guevara-Lara, A.; Cruz-Hernández, A. and Paredes-López, O. 2005. Biochemical and nutritional characterization of three prickly pear species with different ripening behavior. Plant Foods Human Nutrition. 60:195-200.
Hernández-López, D.; Vaillant, F.; Reynoso-Camacho, R.; and Guzman-Maldonado, S. H. 2008. Myrtillocactus geometrizans (Cactaceae): a botanical, agronomic, physicochemical and chemical characteristics of fruits. Fruits 63(1):25-29.
Klaus, A. 2002. On-line LC-NMR and related techniques: 132.
Manica, I.; Icuna, L. M.; Junqueira N., T. V.; Salvador, J. O.; Moreira, A. y Maravolta, E. 2000. Fruticultura Tropical 6. Goiaba. Cinco continentes Editora. Porto Alegre RS. Brasil. p. 37.
Marquina, V.; Araujo, L.; Ruíz, J.; Rodríguez M., A. y Vit, P. 2008. Composición química y capacidad antioxidante en fruta, pulpa y mermelada de guayaba (Psidium guajava L.).
Mondragón, J. C. 2001. Cactus pear domestication and breeding. Plant Breed. Rev. 20:135-166.
Mondragón, J. C y Gallegos, V. C. 2007. Cactus pear genetic resources the cornerstone of the mexican cactus pear industry. Acta Hort. 811:39-46.
Pal, R. K.; Ahmad, M. S.; Roy, S. K. and M. Singh. 2004. Influence of Storage Environment, surface oating, and individual shrink wrapping on quality assurance of guava (Psidium guajava L.) Fruits. Plant Foods for Hum. Nutr. 59:67-72.
Sadler, G. and Dezman, D. 1990. Rapid extraction of lycopene and β-carotene from reconstituted tomato paste and pink grapefruit homogenates. J. Food Sci. 55:1460-1461.

Caracterización de selecciones de guayaba para el Bajío de Guanajuato, México 319
Sanjinez-Argadoña, E. J.; Cunha, R. L.; Mengalli, F. C. and Hubinger, M. D. 2005. Evaluation of total carotenoids and ascorbic acid in osmotic pretreated guavas during convective drying. Ital. J. Food Sci. 17:305-314.
Scar tezz in i , P. ; Antognoni , F. ; Raggi , M. A. ; Poli, F. and Sabbiani, C. 2006. Vitamin C
content and antioxidant activity of the fruit and of the Ayurvedic preparation of Emblica officinalis Gaertn. J. Ethnopharmacology. 104:113-118.
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA), Instituto Tecnológico de Estudios Superiores Monterrey (ITESM), Instituto Nacional de Capacitación Agrícola (INCA). 2005. Plan rector del sistema nacional de guayaba. Segunda fase: diagnóstico inicial base de referencia estructura estratégica.
Terrones, T. del R.; Mejía, A. C. y García, N. H. 2000. Índices agroclimáticos de Guanajuato. SAGAR, INIFAP-CIRCE, CEBAJ. Celaya, Guanajuato. (Publicación Técnica Núm. 2).

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 320-328
GENETIC DIFFERENCES BETWEEN Anastrepha ludens (Loew) POPULATIONS STEMMING FROM A NATIVE AND AN EXOTIC HOST IN NE MEXICO*
DIFERENCIAS GENÉTICAS ENTRE POBLACIONES DE Anastrepha ludens (Loew) DE HOSPEDEROS
NATIVOS Y EXÓTICOS EN EL NE MÉXICO
Víctor Pecina Quintero1§, José Isabel López Arroyo1, Jesús Loera Gallardo1, Juan Rull2, Enrique Rosales Robles1, Edgardo Cortez Mondaca1, Sanjuana Hernández Delgado3, Netzahualcóyotl Mayek Perez3 and Martín Aluja Schuneman2
1Campo Experimental Río Bravo, INIFAP. A. P. 172, C. P. 88900, Río Bravo, Tamaulipas, México. Tel. 01 899 944 10 46. ([email protected]), ([email protected]), ([email protected]), ([email protected]). 2Instituto de Ecología, A. C., A. P. 63 C. P. 91000 Xalapa, Veracruz, México. Tel. 01 228 842 18 41. ([email protected]), ([email protected]). 3Centro de Biotecnología Genómica, Instituto Politécnico Nacional. A. P. 152, C. P. 88730. Ciudad Reynosa, Tamaulipas, México. ([email protected]), ([email protected]). §Autor para correspondencia: [email protected].
ABSTRACT
The objective of this study was to determine the genetic structure of populations of the mexican fruit fly Anastrepha ludens (Loew) in NE citrus growing regions of Mexico. The work was conducted during 2005 at the Center of Genomic Biotechnology in Reynosa, Tamaulipas, Mexico. AFLP markers using four different pairs of initiators were used on specimens collected from the native, ancestral host, yellow chapote (Casimiroa greggii S. Wats) and the exotic host, citrus (Citrus sinensis (L.) Osbeck cv Valencia). The four combinations of initiators amplified an average of 95 AFLP fragments. A total of 382 products were obtained and 259 (67%) of them were polymorphous. The overall index of genetic diversity was 28%, with 3% difference between the genotypes collected from yellow chapote as compared to the genotypes collected from sweet orange. Cluster analysis and principal components showed a close genetic relation among A. ludens specimens regardless of the origin. Although, the dendrogram showed two main groups, the genetic distance varied from 0.0 to 10%, suggesting that despite the reproductive isolation, host groups are not evolving. These results are discussed in the context of evolution and conclude on practical implications of the ongoing eradication program through sterile insect technique and the geographical origin of this species.
Key words: AFLP markers, genetic diversity, host fidelity.
RESUMEN
El objetivo de este trabajo fue determinar la estructura genética de poblaciones de mosca mexicana de la fruta Anastrepha ludens (Loew) en las regiones citrícolas del noreste de México. La caracterización molecular se realizó durante 2005 en Centro de Biotecnología Genómica del Instituto Politécnico Nacional en Reynosa, Tamaulipas. Se utilizaron marcadores AFLP con cuatro diferentes pares de iniciadores para analizar individuos de A. ludens colectados del hospedero nativo chapote amarillo (Casimiroa greggii S. Wats) y del hospedero exótico naranja dulce (Citrus sinensis L. Osbeck cv Valencia). Las cuatro combinaciones de iniciadores amplificaron en promedio 95 fragmentos de AFLP. Se obtuvieron 382 productos de los cuales, 259 (67%) fueron polimórficos. El índice general de diversidad genética fue de 28% con 3% de diferencia entre los genotipos colectados de chapote amarillo y los de naranja dulce. El análisis de conglomerados y de componentes principales mostró estrecha relación genética entre individuos de A.
* Recibido: Febrero, 2009
Aceptado: Septiembre, 2009

Genetic differences between Anastrepha ludens (Loew) populations stemming from a native and an exotic host in NE Mexico 321
ludens independientemente de su origen. No obstante de que el dendrograma divide las poblaciones de mosca mexicana de la fruta en dos grandes grupos, la distancia genética varió de 0.0 a 10%, lo que sugiere que no obstante el aislamiento reproductivo entre los grupos, estos no están en evolución. Estos resultados se discuten en el contexto de evolución (historia de vida) y se concluye sobre las implicaciones que tienen sobre la campaña en curso para la erradicación mediante la técnica de insectos estériles y para determinar el origen geográfico de estas especies.
Palabras clave: diversidad genética, fidelidad al hospedero, marcadores AFLP.
INTRODUCTION
The genetic structure of A. ludens populations can be an important component for a successful region-wide pest management (Via, 1990; Krafsur, 2005) and it is fundamental to establish the origin and identity of the invasive species into agricultural regions (Roderick, 1996; Armstrong and Ball, 2005). The mexican fruit fly complex is different enough from others that renaming it has been long overdue (Baker et al., 1944; Aluja et al., 2003; Hernández-Ortiz et al., 2004). In the A. fraterculus sibling complex, some sibling species appear to have different geographic range, but others do not. Sibling species exhibit differences in biology, host diversity (Aluja et al., 2000a, 2003) and consistent morphological characteristics (Hernandez-Ortíz et al., 2004). Mating compatibility tests among different populations of South America A. fraterculus have provided evidence of prezygotic reproductive isolation due in part to asynchronies in the daily patterns of sexual activity (Vera et al., 2006). The latter finding has important management implications, particularly on the sterile insect technique (SIT). A. ludens is the most devastating pest of citrus in Mexico and Central America (Aluja, 1993; Thomas, 2003); however, its genetic structure is almost unknown. Up to date the only published report indicate that it is difficult to genetically differentiate A. ludens populations, because it’s high intra-population variability (Mangan and Moreno, 2003). Currently, this species are subject to an area-wide management program based on the sterile insect technique that has resulted in the partial/temporary declaration of several fruit fly free areas in NW Mexico (Reyes et al., 2000). Introduced A. ludens populations have been eradicated through SIT from the San Diego area in the US. (Dowell et al., 2000) and have been a recurrent problem in the Rio Grande valley of Texas (Thomas, 2003).
In NE Mexico, the widespread abundance of naturally occurring ancestral hosts of A. ludens such as the yellow chapote (Casimiroa greggi S. Wats), hinders the official declaration of low pest prevalence area; therefore, a stiff regulation on citrus fruit transport has been impose. It is assumed that there is a permanent massive flow of mexican fruit flies from their native hosts (yellow chapote and white zapote [Casimiroa edulis Llave et Lex]) into citrus groves (Enkerlin, 1987), that complicates an effective pest control. However, there is no solid evidence based on the genetic structure of the populations that demonstrates that there is a flow of flies from the native habitats to crop areas. Studies on mobility and longevity of sterile mexican fruit flies revealed that adults can fly up to 9 km; however, 98% were recaptured within 100 m from the release site (Enkerlin 1987, Thomas & Loera-Gallardo, 1998). The latter study determined a mean mobility distance of 240 m (using a regression model), which suggest that mexican fruit flies in citrus, reproduce within the orchard or at nearby neglected plantations. Nevertheless, studies on the reproductive phenology of A. ludens in NE Mexico (Enkerlin, 1987; Thomas, 2003) suggest that the fly is essentially bivoltine in the area and may exploit yellow chapote fruits during spring and citrus at fall. A recent study compared the reaction of wild and lab-reared A. ludens strains to host fruits, the results showed that wild flies were incapable of recognize introduced grapefruit as host, while laboratory flies did. Such reaction is the effect of selective pressures to which the mexican fruit flies have been submitted at the laboratory (Robacker and Fraser, 2004) and a manifestation of the inherent plasticity of this species. Also, when wild and lab-reared flies had previous contact with yellow chapote, the attraction towards this host increased.
The objective of this study was to determine the genetic structure of A. ludens populations from two of the most important citrus producing states of Mexico: Tamaulipas and Nuevo Leon. Molecular characterization of A. ludens populations is important for the successful application of the SIT in pest management programs. If A. ludens, as is the case with A. fraterculus, is found to enclose a complex of cryptic species, then the strain currently being release on SIT programs in Mexico would be useless at certain areas due to reproductive isolation. Furthermore, molecular characterization of A. ludens populations will be an aid to confirm and identify the origin of recurrent putative introductions into citrus growing areas of California, Texas and Florida.

Víctor Pecina Quintero et al.322 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
MATERIALS AND METHODS
DNA collection and extraction
A total of 25 specimens (adult and larvae) of the mexican fruit fly, A. ludens were directly collected from infested C. greggii (yellow chapote) and Citrus sinensis cv. Valencia (citrus), in different geographical regions within NE Mexico
during 2005. (Table 1). The specimens were preserved in ethanol (90%), then washed with sterilized water (to remove alcohol) and stored at- 80 ºC. The specimens were ground to a fine powder with a mortar and pestle. The protocol for DNA extraction was according to Corsini et al., 1999, with extraction solution (50 mM Tris HCl, pH 8.0, 50 mM, EDTA, 3% w/v SDS and 0.1 M 2-mercaptoetanol) and 25 µl proteinase K (10 mg ml).
(Applied Biosystems®). The following PCR program was used: 94 ºC for 30 s, 56 ºC for 60 s and 72 ºC for 60 s for 20 cycles. Subsequently, 2.5 µL of pre-amplified DNA was added with 0.5 µL of the oligonucleotide EcoRI + 3 (EcoRI + AAA), 0.6 µL of oligonucleotide MseI + 3 (AAA, AAC, ATG and AGG), both with a concentration of 50 ng µL-1, and 0.4 µL of 10 mM dNTPs mix, for a total volume of 5 µL, plus 0.2 µL of Taq DNA polymerase 5 U µL-1 and 1.4 µL of 10 X PCR buffer, for a total volume of 11 µL.
The reaction mixture was subjected to the following PCR program: 94 ºC for 30 s, 65 ºC for 30 s and 72 ºC for 60 s; this for 11 cycles where the temperature was reduced by alignment of 0.7 ºC per cycle and then continued with the following PCR program: 94 ºC for 30 s, 56 ºC for 30 s and 72 ºC for 60 s for 23 cycles. Amplified products were separated in acrylamide gel at 6%. Electrophoresis was performed at 2 000 V for 3 h. For detection of amplified products using the Silver Sequence Staining Reagents kit Promega®. The AFLP analysis was conducted at the laboratory of plant biotechnology of the Genomic Biotechnology Center of the National Polytechnic Institute in Reynosa, Tamaulipas, Mexico.
Num. of collection Host Location /CoordinatesCHATAM1, 2, 3, 4 and 5 Casimiroa greggii Tamaulipas, Mexico
N 23º 45' W 99º 03'CHATAM6, 7, 8, 9 and 10 Casimiroa greggii Tamaulipas, Mexico
N 23º 59' W 99º 06' CHANL1, 2, 3 and 4 Casimiroa greggii Nuevo Leon, Mexico
N 24º 52' W 99º 43'CITAM 1, 2, 3, 4, 5, 6, 7, 8, 9 and 10 Citrus sinensis Tamaulipas, Mexico
N 23º 45' W 98º 59'CITSLP (C) Citrus sinensis San Luis Potosi, Mexico
N 21º 55' W 99º 59'
Table 1. Host of origin and location where Anastrepha ludens specimens were collected.
Numbers refer to individuals labeled in Figures 1 and 2.
AFLP analysis
The AFLP protocol utilized was that of Vos et al. (1995). It consists in the genomic DNA digestion and ligation of specific adaptors, followed by two PCR selective amplifications. For DNA digestion a reaction mixture of 30 µL containing 3 µL of DNA (150 ng µL-1), 1.5 µL of EcoRI (Roche®) of 10 U µL-1, 1.5 µL of Tru9I (Roche®) of 10 U µL-1 and 3 µL of 10X RL was used. The samples were incubated for 4 h at 37°C, followed by heating the reaction mixture to 70°C for 15 min to inactivate the restriction enzymes. The specific adapters were united to the sites of restriction. The mixture consisted of 1 µL of EcoRI adapter (5 pmol), 1 µL of MseI adapter (50 pmol), 1.2 µL of 10 mM ATP, 1 µL of 10X RL buffer and 1 µL of T4 DNA ligase (Roche®) of 1 U µL-1 in a total volume of 10 µL. The preamp included 5 µL of the ligation mixture plus the addition of a mixture of oligonucleotides specific AFLP (50 ng µL-1), 2 µL of 10 mM dNTPs in a total volume of 25 µL. The reaction mixture consisted of 0.2 µL addition of Taq DNA polymerase (Roche®) (5 U µL-1) and 5 µL of 10X PCR buffer in a final volume of 20 µL. The total volume of the reaction mixture was 50 µL. The mixture was subjected to PCR amplification in a GeneAmp 9 700 thermocycler
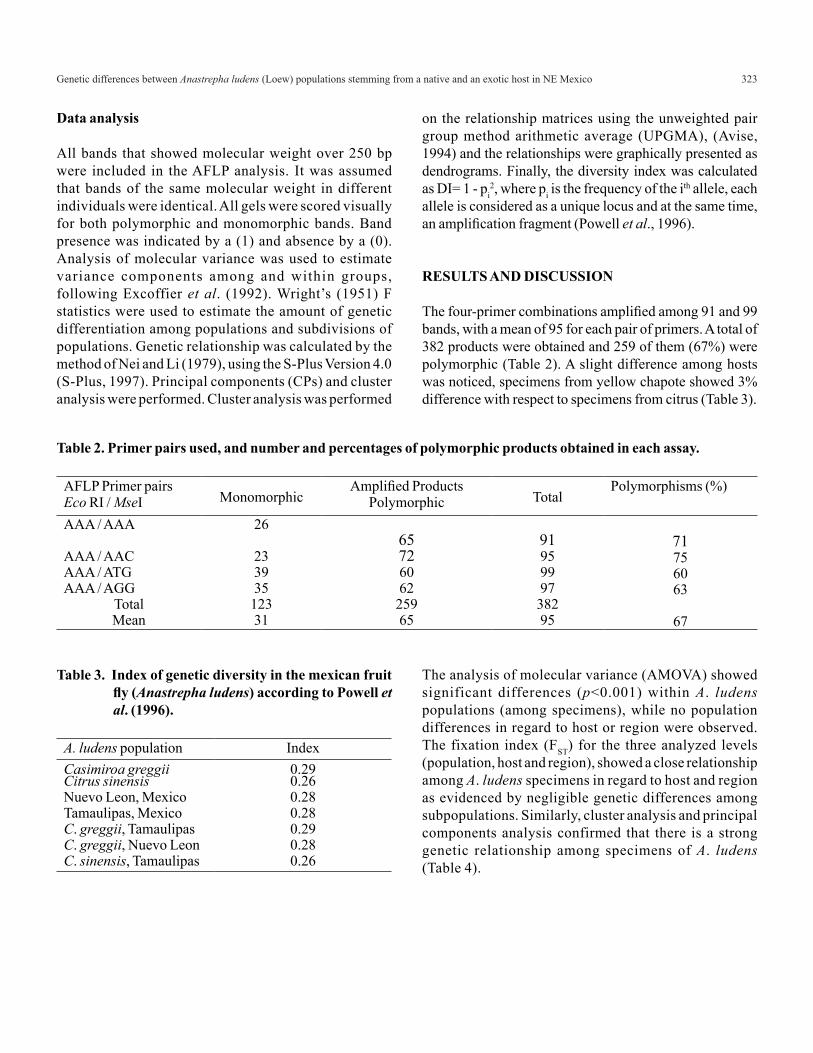
Genetic differences between Anastrepha ludens (Loew) populations stemming from a native and an exotic host in NE Mexico 323
Data analysis
All bands that showed molecular weight over 250 bp were included in the AFLP analysis. It was assumed that bands of the same molecular weight in different individuals were identical. All gels were scored visually for both polymorphic and monomorphic bands. Band presence was indicated by a (1) and absence by a (0). Analysis of molecular variance was used to estimate variance components among and within groups, following Excoffier et al. (1992). Wright’s (1951) F statistics were used to estimate the amount of genetic differentiation among populations and subdivisions of populations. Genetic relationship was calculated by the method of Nei and Li (1979), using the S-Plus Version 4.0 (S-Plus, 1997). Principal components (CPs) and cluster analysis were performed. Cluster analysis was performed
on the relationship matrices using the unweighted pair group method arithmetic average (UPGMA), (Avise, 1994) and the relationships were graphically presented as dendrograms. Finally, the diversity index was calculated as DI= 1 - pi
2, where pi is the frequency of the ith allele, each allele is considered as a unique locus and at the same time, an amplification fragment (Powell et al., 1996).
RESULTS AND DISCUSSION
The four-primer combinations amplified among 91 and 99 bands, with a mean of 95 for each pair of primers. A total of 382 products were obtained and 259 of them (67%) were polymorphic (Table 2). A slight difference among hosts was noticed, specimens from yellow chapote showed 3% difference with respect to specimens from citrus (Table 3).
The analysis of molecular variance (AMOVA) showed significant differences (p<0.001) within A. ludens populations (among specimens), while no population differences in regard to host or region were observed. The fixation index (FST) for the three analyzed levels (population, host and region), showed a close relationship among A. ludens specimens in regard to host and region as evidenced by negligible genetic differences among subpopulations. Similarly, cluster analysis and principal components analysis confirmed that there is a strong genetic relationship among specimens of A. ludens (Table 4).
AFLP Primer pairs Amplified Products Polymorphisms (%)Eco RI / MseI Monomorphic Polymorphic Total
AAA / AAA 2665 91 71
AAA / AAC 23 72 95 75AAA / ATG 39 60 99 60AAA / AGG 35 62 97 63
Total 123 259 382Mean 31 65 95 67
A. ludens population IndexCasimiroa greggii 0.29Citrus sinensis 0.26Nuevo Leon, Mexico 0.28Tamaulipas, Mexico 0.28C. greggii, Tamaulipas 0.29C. greggii, Nuevo Leon 0.28C. sinensis, Tamaulipas 0.26
Table 2. Primer pairs used, and number and percentages of polymorphic products obtained in each assay.
Table 3. Index of genetic diversity in the mexican fruit fly (Anastrepha ludens) according to Powell et al. (1996).
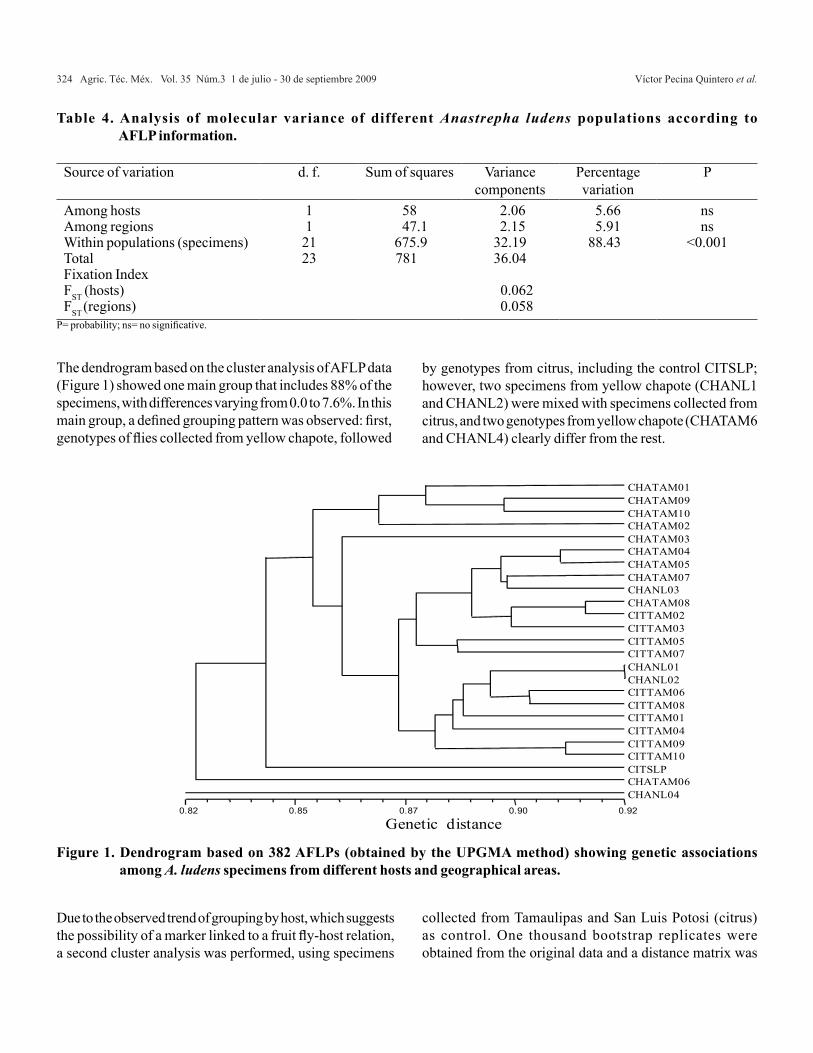
Víctor Pecina Quintero et al.324 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Source of variation d. f. Sum of squares Variance components
Percentage variation
P
Among hosts 1 58 2.06 5.66 nsAmong regions 1 47.1 2.15 5.91 nsWithin populations (specimens) 21 675.9 32.19 88.43 <0.001Total 23 781 36.04Fixation IndexFST (hosts) 0.062FST (regions) 0.058
Geneti cdistance0.82 0.85 0.87 0.90 0.92
CHATAM10MW
CHATAM01 CHATAM09 CHATAM10 CHATAM02 CHATAM03 CHATAM04 CHATAM05 CHATAM07 CHANL03 CHATAM08 CITTAM02 CITTAM03 CITTAM05 CITTAM07 CHANL01 CHANL02 CITTAM06 CITTAM08 CITTAM01 CITTAM04 CITTAM09 CITTAM10 CITSLP CHATAM06 CHANL04
Genetic distance0.82 0.85 0.87 0.90 0.92
CHATAM10MW
CHATAM01 CHATAM09 CHATAM10 CHATAM02 CHATAM03 CHATAM04 CHATAM05 CHATAM07 CHANL03 CHATAM08 CITTAM02 CITTAM03 CITTAM05 CITTAM07 CHANL01 CHANL02 CITTAM06 CITTAM08 CITTAM01 CITTAM04 CITTAM09 CITTAM10 CITSLP CHATAM06 CHANL04
Figure 1. Dendrogram based on 382 AFLPs (obtained by the UPGMA method) showing genetic associations among A. ludens specimens from different hosts and geographical areas.
by genotypes from citrus, including the control CITSLP; however, two specimens from yellow chapote (CHANL1 and CHANL2) were mixed with specimens collected from citrus, and two genotypes from yellow chapote (CHATAM6 and CHANL4) clearly differ from the rest.
collected from Tamaulipas and San Luis Potosi (citrus) as control. One thousand bootstrap replicates were obtained from the original data and a distance matrix was
Table 4. Analysis of molecular variance of different Anastrepha ludens populations according to AFLP information.
P= probability; ns= no significative.
The dendrogram based on the cluster analysis of AFLP data (Figure 1) showed one main group that includes 88% of the specimens, with differences varying from 0.0 to 7.6%. In this main group, a defined grouping pattern was observed: first, genotypes of flies collected from yellow chapote, followed
Due to the observed trend of grouping by host, which suggests the possibility of a marker linked to a fruit fly-host relation, a second cluster analysis was performed, using specimens

Genetic differences between Anastrepha ludens (Loew) populations stemming from a native and an exotic host in NE Mexico 325
calculated for each replicate. In this new analysis, two main genotype groups were observed; the first one, included yellow chapote and citrus specimens from Tamaulipas and the other the citrus specimen from San Luis Potosi (control). The dendrogram showed that all specimens from yellow chapote are closely related, followed by citrus specimens collectedin Tamaulipas (Figure 2).
This suggests the existence of a DNA marker linked to the fruit fly-host relation. Moreover, in the matrices of AFLP, it was observed that the Eco/Mse AAA/AAA pair of primers amplify a DNA band which was present in the whole set of specimens collected in citrus; meanwhile, in the yellow chapote specimens this DNA band was absent, except for one individual.
among specimens from the same host with genotypes collected from yellow chapote separated from those collected from citrus.
Phenotypic plasticity has been defined as the ability of a single genotype to produce more than one alternative form in response to environmental conditions (West-Eberhard, 1989). It has been recognized that plasticity itself is a trait subject to natural selection and evolutionary change (Williams, 1966). The results of this research indicate that A. ludens exploiting different hosts have a slightly different genetic composition, probably owing to the fact that citrus colonization by yellow chapote flies implies going through
0.1
CITSLP
CHATAM03
CHATAM01
CHATAM0278
55
CHATAM04
CHATAM0531
27
CHATAM06
CHATAM0729
CHATAM08
CHATAM0931
10
18
CHATAM10
CITAM1032
CITAM02
CITAM0333
10
15
CITAM04
CITAM0533
CITAM06
CITAM0729
10
CITAM10
CITAM08
CITAM0929
16
5
42
100
Genetic distance
Figure 2. Dendrogram based on 382 AFLPs (obtained by the UPGMA method) showing genetic associations among A. ludens specimens from different hosts in Tamaulipas, Mexico. Numbers above the branches represent bootstrap support values for 1 000 replicates.
The AFLP analysis showed a high level of polymorphism (67%), with a mean index of diversity of 28%. The technique was sensitive enough to detect differences among individuals. A slight trend toward greater genetic diversity of A. ludens populations stemming from native yellow chapote was observed, which is possibly due to the fact that these populations have been less perturbed (Robacker and Fraser, 2004) and subjected to a lesser selection than citrus populations subjected to frecuent insecticide sprays.
A weak grouping trend related to host origin was observed; particularly in the specimens from Tamaulipas (Figure 2). The analyses showed a close relation

Víctor Pecina Quintero et al.326 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
a series of bottlenecks as described by Alberti et al. (1999) for A. fraterculus in Argentina. Nevertheless, the detected differences were not significant, suggesting that reproductive isolation is not evolving among the studied populations. It is important to note that A. ludens is polyphagous and multivoltine (Aluja, 1994; Thomas, 2003). Multivoltine fruit flies have to cope with long periods when host fruit is not available without undergoing diapuase. There are a few possible life strategies to overcome this hurdle. The first one would be through evolution of adult longevity, which has occurred in some Anastrepha species (Aluja et al., 2000a) but not in the case of A. ludens; the second would be by exploiting hosts with long fruiting periods, as it is the papaya fruit fly, Toxotrypana curvicauda Gerstaecker. Multivoltine fruit flies can bridge the fruiting gaps of their main host by exploiting alternate hosts (Aluja and Mangan, 2008). This survival strategy might select for genetic plasticity but also restricts genetic differentiation of host plant races. A likely scenario would explain the observed results, where an A. ludens population switching from chapote to citrus undergoes a genetic bottleneck that can change allele frequency, followed by a return to the ancestral host that may result in homogenizing the gene pool. This life strategy and the phenotypic plasticity observed may help to explain the findings of Robacker and Fraser (2004) showing that mexican fruit flies with previous contact with yellow chapote will prefer this host over other alternatives regardless of their own origin. It also provides support to the idea that chapote populations move back and forth from chapote to citrus as documented by Enkerlin (1987) and more recently by Thomas (2003).
Practical implications of these findings suggest that at a regional scale, the sterile insect technique can be applied for A. ludens without the need to colonize different strains stemming from different hosts. AFLP markers proved to be a useful tool to differentiate among A. ludens specimens allowing to detect a high level of polymorphism, measure genetic diversity and to detect genetic differences in host relation. It is important to mention that this is the first step of several more to follow attempting to characterize A. ludens populations within their geographical range. Molecular scrutiny of these populations at a wider scale is essential for a thorough understanding of the evolution of A. ludens life strategies. Wide range sampling is also critical to rule out geographical effects (Atlantic-Pacific, altiplano-coastal
plains) on A. ludens population’s genetic composition and to get evidence that these factors may be leading to the evolution of inter population reproductive isolation. That attempt will also contribute to determine the origin of recurrent introductions of A. ludens into citrus growing regions of the U. S., whether occur by natural dispersal (the case of Texas and Nuevo Leon), or human transport. It is also necessary to widen the scope of sampling of host-associated populations considering that both chapote and citrus belong to Rutaceae and A. ludens has been reported infesting plants in very different families: Solanaceae (Thomas, 2004), Anacardaceae, and Rosaceae (Aluja et al., 2000b; Norrbom, 2003). Based on the above, the next step to genetically characterize the A. ludens populations in Mexico, will be to extend the sampling to other regions and other host plants and to incorporate other molecular tools as microsatellites.
CONCLUSIONS
Despite there is a high level of polymorphism (67%) in A. ludens in NE Mexico, the genetic diversity of this species is low (ID 28%).
The AFLP technique was sensitive enough to detect differences among individuals, a silght 3% difference was observed among specimens collected from yellow chapote as compared to those collected from sweet orange.
Cluster analysis and principal components showed a tight genetic relation among A. ludens individual regardless of origin.
Although the dendrogram divided divide the mexican fruit fly populations into two main groups, the genetic distance (0.0 to 10%) suggest that despite the reproductive isolation, host groups are not evolving.
ACKNOWLEDGEMENTS
We thank Victor Maya Hernandez for the collection of flies in Tamaulipas, Mexico and Alberto Anzures for the review of this manuscript. This study was sponsored by INIFAP, Fondo Mixto CONACyT-Gobierno de Nuevo Leon and Fundacion Produce Tamaulipas A. C. MA and JR acknowledge the support from the Campaña Nacional Contra Moscas de la Fruta.

Genetic differences between Anastrepha ludens (Loew) populations stemming from a native and an exotic host in NE Mexico 327
LITERATURE CITED
Alberti, A. C.; Calcagno, G.; Saidman, B. O and Vilardi, J. C. 1999. Analysis of the genetic structure of a natural population of Anastrepha fraterculus (Diptera: Tephrititdae). Ann. Entomol. Soc. Am. 92:731-736.
Aluja, M. 1993. Manejo integrado de las moscas de la fruta. Ed. Trillas, Mexico D. F. México.
Aluja, M. 1994. Bionomics and management of Anastrepha. Ann. Rev. Entomol. 39:155-178.
Aluja, M. and Mangan, R. 2008. Fruit fly (Diptera: Tephritidae) host status determination: critical conceptual, methodological and regulatory considerations. Annu. Rev. Entomol. 53:
In press.Aluja, M.; Piñero, J.; Jácome, I.; Díaz-Fleischer, F. and
Sivinski, J. 2000a. Behavior of flies in the genus Anastrepha (Trypetinae: Toxotrypanini), p. 375-408. In: Aluja, M. and Norrbom, A. L. (eds.), Fruit flies (Tephritidae): phylogeny and evolution of behaviour. CRC Press, Boca Raton, FL.
Aluja, M.; Piñero, J.; López, M.; Ruíz, C.; Zúñiga, A.; Piedra, E.; Díaz-Fleischer, F. and Sivinski. F. 2000b. New host plant and distribution records in Mexico for Anastrepha spp., Toxotrypana curvicauda Gerstacker, Rhagoletis zoqui Bush, Rhagoletis sp., and Hexachaeta sp. (Diptera: Tephritidae). Proc. Entomol. Soc. Wash. 102:802-815.
Aluja, M.; Pérez-Staples, D.; Macías-Ordóñez, R.; Piñero, J.; McPheron, B. and Hernández-Ortiz, V. 2003. Nonhost status of citrus sinensis cultivar Valencia and C. paradisi cultivar ruby red to mexican Anastrepha fraterculus (Diptera: Tephritidae). J. Econ. Entomol. 96:1693-1703.
Armstrong, K. F. and Ball, S. L. 2005. DNA barcodes for biosecurity: invasive species identification. Philos. Trans. R. Soc. Lond. Biol. Sci. 360:1813-1823.
Avise, J. C. 1994. Molecular markers, natural history and evolution. Chapman & Hall, New York, NY.
Baker, A. C.; Stone, W. E.; Plummer, C. C. and McPhail, M. 1944. A review of studies on the mexican fruit fly and related mexican species. USDA, Miscellaneous publication No. 531.
Corsini, G.; Manubens, A.; Lladser, A.; Lobos, S.; Seelenfreund, D. and Lobos, C. 1999. AFLP analisis of fruit fly Ceratitis capitata. Focus 21(3):72-73.
Dowell, R. V.; Sidiqui, I. A.; Meyer, F. and Spaugy, L. 2000. Mediterranean fruit fly preventive release program in Southern California, p. 369-375. In: Tan, K. H. (ed), Area-wide control of fruit flies and other insect pests. Universiti Sains Malaysia, Penang, Malaysia.
Enkerlin, W. 1987. Orientación y dispersión de poblaciones de la mosca mexicana de la fruta (Anastrepha ludens, Loew), estériles y silvestres, en el municipio de Allende, N. L., en el período de septiembre de 1985 a agosto de 1986. M. S. Thesis, Instituto Tecnológico y de Estudios Superiores de Monterrey (ITESM), Campus Monterrey, Monterrey, Nuevo León, México.
Excoffier, L.; Simouse, P. and Quattro, J. 1992. Analysis of molecular variance inferred for metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics 131:479-491.
Hernandez-Ortiz, V.; Gomez-Amaya, J. A.; Sanchez, A.; McPheron, B. A. and Aluja, M. 2004. Morphometric analysis of mexican and South American populations of the Anastrepha fraterculus complex (Diptera: tephritidae) and recognition of a distinct mexican morphotype. Bull. Entomol. Res. 94:487-499.
Krafsur, E. S. 2005. Role of population genetics in the sterile insect technique, pp. 389- 406. In: Dyck, V. A., Hendrichs, J. and Robinson, A. S. (eds). Sterile insect technique: principles and practice in area-wide integrated pest management. Springer, Dordrech, The Netherlands.
Mangan, R. L. and Moreno, D. S. 2003. Molecular analysis of Anastrepha ludens genetic variation. http://www.tard.state.tx.us/listing.php?id=3d6d01c202fe8.(05-08-2003).
Nei, M. and Li, W. H. 1979. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc. Natl. Acad. Sci. USA. 76:5269-5273.
Norrbom, A. L. 2003. Host plant data base for Anastrepha and Toxotrypana (Diptera: Tephritidae: Toxotrypanini). Diptera data dissemination disk. CD-ROM, North Amer. Dipterist’s Soc. Washington, D. C.
Powell, W.; Morgante, M.; Andre, C.; Hanafey, M.; Vogel, J.; Tingey, S. and Rafalski, A. 1996. The comparasion of RFLP, RAPD, AFLP and SSR (microsatellite) markers for germplasm analysis. Mol. Breeding 2:225-238.

Víctor Pecina Quintero et al.328 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Reyes, J.; Santiago, G. and Hernández, P. 2000. The mexican fruit fly eradication programme, p. 377-380. In: Tan, K. H. (ed.). Area-wide control of fruit flies and other insect pests. University Sains Malaysia, Penang, Malaysia.
Robacker, D. C. and Fraser, I. 2004. Do mexican fruit flies (Diptera: Tephritidae) prefer grapefruit to yellow chapote, a native host? Fla. Entomol. 85:481-487.
Roderick, G. K. 1996. Geographic structure of insect populations: gene flow, phylogeography, and their uses. Ann. Rev. Entomol. 41:325-352.
S-Plus (S-Plus) User`s Guide Version 4.0. 1997. Seattle: Math Soft. 525 p.
Thomas, D. B. and Loera-Gallardo, J. 1998. Dispersal and longevity of mass-released mexican fruit files (Diptera: Tephritidae). Environ. Entomol. 27:1045-1052.
Thomas, D. B. 2003. Reproductive phenology of the mexican fruit fly Anastrepha ludens (Loew) (Diptera: Tephritidae) in the Sierra Madre Oriental, Northern Mexico. Neotrop. Entomol. 32:385-397.
Thomas, D. B. 2004. Hot peppers as a host for the mexican fruit fly Anastrepha ludens (Diptera: Tephritidae). Fla. Entomol. 87:603-608.
Vera, M. T.; Cáceres, C.; Wornoayporn, V.; Islam, A.; Robinson, A. S.; De La Vega, M. H.;
Hendrichs, J. and Cayol, J. P. 2006. Mating in compatibility among populations of the South American fruit fly Anastrepha fraterculus (Diptera: Tephritidae). Ann. Entomol. Soc. Am. 99:387-397.Via, S. 1990. Ecological genetics and host adaptation in
herbivorous insects: The experimental study of evolution in natural and agricultural systems. Ann. Rev. Entomol. 35:421- 446.
Vos, P.; Hogers, R.; Bleeker, M.; Reijans, M.; Van de Lee, T.; Hornes, M.; Frijters, A.; Pot, J.; Peleman, J.; Kuiper, M. & Zabeau, M. 1995. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res. 23:4407-4414.
West-Eberhard, M. J. 1989. Phenotypic plasticity and the origins of diversity. Annu. Rev. Ecol. Syst. 20:249-278.
Williams, G. C. 1966. Adaptation and natural selection. Princeton University Press, Princeton, USA.
Wright, S. 1951. The genetical structure of populations. Ann. Eugen. 15:323-354.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 329-334
ÁREAS PRIORITARIAS PARA COLECTAR GERMOPLASMA DE (Crataegus L.) EN MÉXICO CON BASE EN LA DIVERSIDAD Y RIQUEZA DE ESPECIES*
PRIORITY AREAS TO COLLECT (Crataegus L.) GERMPLASM IN MEXICO ON THE BASIS OF
DIVERSITY AND SPECIES RICHNESS
Carlos Alberto Núñez-Colín§
Campo Experimental Bajío, INIFAP. Km. 6.5 carretera Celaya-San Miguel de Allende. A. P. 112. C. P. 38110 Celaya, Guanajuato, México. §Autor para correspondencia: [email protected].
* Recibido: Enero, 2009
Aceptado: Septiembre, 2009
RESUMEN
El tejocote (Crataegus spp.) es un árbol frutal distribuido en la mayor parte del territorio mexicano, se reportan 13 especies para las regiones del norte y centro y dos o más especies pueden existir en el sur. Se desconoce la ubicación de las regiones con mayor biodiversidad de este género en el país. Por lo tanto, el presente estudio biogeográfico tuvo como objetivos determinar las áreas prioritarias para la colecta de germoplasma con base en índices de diversidad y riqueza específica del género Crataegus en México, en 2007. Para obtener la riqueza de especies y diversidad se utilizó el programa DIVA GIS versión 2 en el cual se ingresó en cada celda el número de especies identificadas en una cuadrícula de un grado de latitud por uno de longitud. Tres diferentes regiones mostraron alta diversidad: la primera en el noreste, específicamente en el suroeste del estado de Nuevo León en la colindancia con el sureste de Coahuila; la segunda en el centro de México, incluyendo el este del Estado de México, todo el estado de Tlaxcala y el área central del estado de Puebla. La tercera, en el sur, en la región de los altos en el centro del estado de Chiapas. Estas regiones pueden considerarse prioritarias para colectar germoplasma representativo del género Crataegus.
Palabras clave: índice de diversidad, índice de diversidad beta, tejocote Rosaceae subfam. Maloideae.
ABSTRACT
The tejocote (Crataegus spp.) is a fruit tree widely distributed through the mexican territory, 13 species had been reported in the northern and central regions and two or more species can exist in southern Mexico. The location of the highest biodiversity of this genus in Mexico is unknown. Therefore, the present biogeographic study aims to determine the priority areas for germplasm collection on the basis of diversity and species richness of the genus Crataegus in Mexico, in 2007. The software DIVA-GIS version 2 was utilized to obtain the species richness and diversity considering the number of species per cell in a grid of one degree of latitude per one degree of longitude. High diversity was observed in three different regions of Mexico. The first region is located at northeastern Mexico, specifically, at the southwest of the state of Nuevo Leon bordering the southeast of Coahuila.

330 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Carlos Alberto Núñez-Colín
The second at central Mexico, at the east of the Estado de Mexico, the state of Tlaxcala and the central area of Puebla. The third region is at the central highlands in the state of Chiapas. These regions could be considered the priority areas for collecting representative Crataegus germplasm.
Key words: beta diversity index, diversity index, tejocote, Rosaceae subfam. Maloideae.
INTRODUCCIÓN
El género Crataegus es el más amplio y taxonómicamente complejo de los géneros de plantas superiores en Estados Unidos de América (Phipps et al., 2003), se han reportado 13 especies en el norte y centro de México y dos o más especies pueden existir en el sur de México (Eggleston, 1909; Phipps et al., 1990; Phipps, 1997; Núñez-Colín et al., 2004). Phipps et al. (2003) reportaron que las especies mexicanas pertenecen a seis diferentes series taxonómicas (Baroussaneae, Greggianae, Parvifoliae, Madrenses, Crus-galli y Mexicanae), pero se desconoce la ubicación de la más amplia biodiversidad de Crataegus en México. Adicionalmente, la identificación de ejemplares de este género es problemática, se cometen errores al identificar ejemplares de tejocote como C. pubescens (Kunth) Steud. que es un nombre incorrecto desde 1997, siendo correctamente referido como C. gracilior Phipps para ciertos especímenes (Phipps, 1997); además, que a partir del trabajo de Eggleston (1909) es conocido que C. mexicana DC y C. pubescens (C. gracilior) son dos especies diferentes, lo que fue desapercibido por los taxónomos mexicanos hasta la reclasificación de Phipps (1997), esto fue ampliamente discutido por Núñez-Colín (2008) quien cuestionó la validez de nombres específicos como C. stipulosa (Kunth) Steud para especímenes identificados en el sur de México.
El nombre común que se da en México para especies del género Crataegus es “tejocote”, este nombre es derivado de la palabra náhuatl “texocotl” que literalmente significa fruta dura (Cabrera, 1992). Desde la época prehispánica, los frutos de tejocote fueron colectados para consumo y sembrados en huertas de los pueblos indígenas, posteriormente fueron cultivados con fines comerciales (Nieto-Ángel y Borys, 1993). Los frutos de las especies de Crataegus son utilizadas en las celebraciones tradicionales de todos santos y navidad, principalmente en la elaboración de bebidas (ponche) y
consumo en fresco (dentro de las piñatas). El tejocote, por lo tanto, es una parte importante de la cultura tradicional mexicana (Borys y Lesczcyñska-Borys, 1994).
Núñez-Colín et al. (2008b) observaron tres diferentes condiciones eco-climáticas en las que el género Crataegus se desarrolla; la primera región fue en la parte noreste de México (incluyendo algunas áreas en el noroeste) con clima semidesértico; la segunda región fue en el eje neo-volcánico con clima templado, en esta región se presentaron especímenes hortícolas y la tercera región, con clima subtropical, en el sur de México en la frontera con Guatemala. Por lo tanto, el género Crataegus está distribuido en la mayoría de las regiones montañosas en México.
Cuando es necesario colectar germoplasma, el investigador debe hacerlo en regiones con la mayor diversidad para incrementar las posibilidades de obtener la máxima diversidad genética (Zagaja, 1988); sin embargo, esta actividad resulta difícil cuando no se cuenta con estudios biogeográficos.
Por esta razón, el presente estudio tuvo como objetivo determinar las áreas prioritarias para la colecta de germoplasma de Crataegus por medio de índices de riqueza y diversidad biogeográfica; así como determinar la diversidad y riqueza específica del germoplasma de Crataegus en México.
MATERIALES Y MÉTODOS
Se utilizaron 363 datos de pasaporte de especímenes de herbario (MO, LL, TEX, ARIZ, MEXU, IEB, XAL), así como los datos de pasaporte de accesiones del banco de germoplasma de tejocote (Crataegus spp.) de la Universidad Autónoma Chapingo (BGT-UACh) que fueron clasificados, así como correctamente identificados, los datos de herbario fueron obtenidos en su mayoría de la red mesoamericana de información sobre la biodiversidad (REMIB). Los ejemplares de los herbarios mexicanos (MEXU, IEB y XAL) y del BGT-UACh fueron revisados minuciosamente para descartar identificaciones, erróneas; para los ejemplares del norte y centro de México se utilizaron las claves de Phipps (1997) y para los del sur las claves de Eggleston (1909). Los datos de pasaporte con el nombre C. pubescens

Áreas prioritarias para colectar germoplasma de (Crataegus L.) en México con base en la diversidad y riqueza de especies 331
(Kunth) Steud que no pudieron ser identificados con el nombre correcto no fueron tomados en cuenta; asimismo, se tuvo cuidado de que los ejemplares que no se revisaron personalmente hubiesen sido colectados e identificados por expertos como el Dr. James Phipps de la Universidad de Western Ontario.
Se utilizó el programa DIVA-GIS versión 2 (Hijmans et al., 2002) para determinar la riqueza específica con base en el número de especies en cada celda de una cuadrícula delimitada por un grado de latitud por uno de longitud. Aunado a lo anterior, se calculó el estimador de riqueza de Chao, esta fórmula presupone que todos las celdas de la cuadrícula tuvieran el mismo número de sitios de colecta. También se obtuvo el índice de diversidad de Brillouin para determinar la posible biodiversidad con base en el número de especímenes y de especies diferentes ubicados en cada celda. Adicionalmente, se calculó el índice de beta diversidad (o diversidad beta) en la modalidad de “queen case” (tomando en cuenta las ocho celdas
alrededor) para determinar el intercambio de especies en las celdas, cuando en la cuadrícula se presentan diferentes especies en todos las celdas, existe alta diversidad beta en ellas (Hijmans et al., 2002).
RESULTADOS
Riqueza de especies
Se observó alta riqueza de especies en cuatro celdas en dos diferentes regiones en el noreste de México. Uno de las celdas se ubica en el área norte del estado de Coahuila en frontera con el estado de Texas, Estados Unidos de América; tres celdas vecinas, en el sur del estado de Nuevo León, una de ellas colindante con la región oeste del estado de Tamaulipas y otra con el sureste del estado de Coahuila y norte de San Luis Potosí (Figura 1). Las accesiones reportadas para el centro y sur de México presentaron baja riqueza de especies.
presentaron baja riqueza estimada (Figura 2). Adicionalmente, se localizaron áreas secundarias de riqueza estimada en las otras áreas de alta riqueza específica observada y en otras regiones en el este del estado Hidalgo en colindancia con el noroeste del estado de Puebla y noreste de Veracruz (Figura 2).
Rishness 1-2 2-3 3-4 4-5 5-6 Nodata
Figura 1. Riqueza específica observada del género Crataegus en México obtenida con 363 accesiones geo-referenciadas y el programa DIVA-GIS versión 2 (Hijmans et al., 2002).
El estimador de riqueza de Chao presentó algunas diferencias con la riqueza específica observada porque sólo determinó una celda con alta riqueza estimada en el norte del estado de Coahuila en frontera con Texas, que fue similar al de la riqueza observada, pero todas las demás áreas de México
332 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Carlos Alberto Núñez-Colín
Índice de biodiversidad
La más alta diversidad se observó en tres diferentes regiones. La primera en el noreste, específicamente en el suroeste del estado de Nuevo León en colindancia con el sureste de Coahuila. La segunda, que incluye dos celdas juntas en el
centro de México, específicamente en el oriente del Estado de México, todo Tlaxcala y el área central de Puebla, que fueron consideradas como un sólo acervo genético (Núñez-Colín et al., 2008a). La tercera región se ubicó en el sur de México, específicamente en el área central del estado de Chiapas (Figura 3).
1-3 3-5 5-7 7-9 9-11 11-13 Nodata
Figura 2. Estimador de riqueza específica de Chao del género Crataegus en México obtenido con 363 accesiones geo- referenciadas y el programa DIVA-GIS versión 2 (Hijmans et al., 2002).
0-0.5 0.5-1 1-1.5 1.5-2 2-2.5 2.5-3 Nodata
Figura 3. Índice de diversidad de Brillouin del género Crataegus en México obtenido de 363 accesiones geo-referenciadas y el programa DIVA-GIS versión 2 (Hijmans et al., 2002).

Áreas prioritarias para colectar germoplasma de (Crataegus L.) en México con base en la diversidad y riqueza de especies 333
Las áreas secundarias de diversidad se ubicaron en el área central del estado de Michoacán, sureste del estado de Puebla, centro del estado de Veracruz, este del estado de Hidalgo, norte de Coahuila y la parte media del oeste de Tamaulipas en colindancia con el sureste del estado de Nuevo León. Todas las otras regiones de México donde
DISCUSIÓN
Las regiones de alta riqueza determinadas en esta investigación coinciden con lo reportado por Phipps (1997), quien identificó once especies diferentes en cinco series en las mismas áreas en comparación con tres especies diferentes en dos series reportadas por Eggleston (1909); Phipps et al. (1990) y Phipps (1997) para el centro y sur del país.
Es posible que la riqueza específica del género Carataegus en el sur de México esté subestimada, debido a que no existe una taxonomía actualizada para esa región posterior al trabajo de Eggleston (1909). Por lo tanto, este parámetro no debe ser utilizado para estimar zonas prioritarias de colecta de germoplasma.
La diversidad beta debe explicarse con las mismas razones que los índices de riqueza; por lo tanto, las regiones del noreste de México y la mayoría de las regiones identificadas en el trabajo de Phipps (1997), son las regiones más importantes para la colecta taxonómica porque en ellas se
se reportaron accesiones de Crataegus mostraron baja diversidad (Figura 3). La mayor diversidad beta se ubicó en el área noreste de México en los estados de Coahuila, Nuevo León, Tamaulipas y San Luis Potosí. En todas las otras regiones de México, accesiones de Crataegus fueron reportadas con baja diversidad beta (Figura 4).
encuentra el mayor número de especies de Crataegus en México. El centro y sur de México mostraron una importante variación intra serie en las series Mexicanae y Crus-galli pero baja diversidad beta porque sólo presentó dos series diferentes. Debido a lo anterior, este parámetro no debe ser utilizado en la determinación de las áreas prioritarias para la colecta de germoplasma. Sin embargo, las regiones del noreste de México son importantes para obtener material de la mayoría de las especies reportadas en México.
El índice de diversidad de Brillouin puede ser utilizado para priorizar regiones para la colecta de germoplasma. Las tres regiones con la más alta diversidad determinada con este índice podrían representar toda la diversidad genética del género Crataegus. Sin embargo, es necesario considerar las regiones secundarias de diversidad.
Las regiones con alta diversidad corresponden a las tres regiones eco-climáticas reportadas por Núñez-Colín et al. (2008b). En ellas, es posible encontrar más genes útiles para programas de mejoramiento genético que en zonas con baja
Figura 4. Índice de diversidad beta de Whitaker en la modalidad queen case del género Crataegus en México obtenido con 363 accesiones geo-referenciadas y el programa DIVA-GIS versión 2 (Hijmans et al., 2002).
-1-0 0-1 1-2 2-3 3-4 Nodata

334 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Carlos Alberto Núñez-Colín
diversidad (Guarino et al., 2002). Por estas razones, estas tres regiones son consideradas como prioritarias para la colecta de germoplasma del género Crataegus.
CONCLUSIONES
Las regiones con mayor diversidad beta y riqueza de especies se ubican en el noreste y las de más alta diversidad en el noreste, centro y sur de México.
De acuerdo con el índice de diversidad de Brillouin, las zonas prioritarias para la colecta de germoplasma del género Crataegus se localizan en los estados de Tamaulipas, Nuevo León, San Luis Potosí, Coahuila, Estado de México, Tlaxcala, Chiapas y Puebla .
AGRADECIMIENTOS
Esta investigación fue financiada en parte con la beca número 169581 del Consejo Nacional de Ciencia y Tecnología de México (CONACYT).
LITERATURA CITADA
Borys, M. W. y Leszcyñska-Borys, H. 1994. Tejocote (Crataegus spp.)-planta para solares, macetas e interiores. Revista Chapingo Serie Horticultura (México) 1:95-107.
Cabrera, L. G. 1992. Diccionario de Aztequismos. Ediciones Colofón, México, D. F., México. 166 p.
Eggleston, W. W. 1909. The Crataegi of Mexico and Central America. Bull. Torrey Bot. Club (USA)
36:501-514.Guarino, L.; Jarvis, A.; Hijmans, R. J. and Maxted, N. 2002.
(36) Geographic information systems (GIS) and the conservation and use of plant genetic resources. p. 387-404. In: Engels, J. M. M.; Ramanatha Rao, V.; Brown, A. H. D. and Jackson, M. T. (eds.). Managing plant genetic diversity. International Plant Genetic Res. Institute (IPGRI), Rome, Italy.
Hijmans, R. J.; Guarino, L.; Bussink, C.; Barrantes, I. y Rojas, E. 2002. DIVA-GIS Versión 2. Sistema de información geográfica para el análisis de datos de biodiversidad. Internacional Potato Center, Lima, Perú. 64 p.
Nieto-Ángel, R. y Borys, M. W. 1993. El tejocote (Crataegus spp.), un potencial frutícola de zonas templadas. Revista Fruticultura Profesional (España)
54:64-71.Núñez-Colín, C. A.; Pérez-Ortega, S. A.; Segura, S.;
Nieto-Ángel, R. y Barrientos-Priego, A. F. 2004. Variabilidad morfológica de tejocote (Crataegus spp.) en México. Proc. of the Interamer. Soc. Tropical Hort. (USA) 48:144-148.
Núñez-Colín, C. A. 2008. Variabilidad inter e intra específica del germoplasma de tejocote (Crataegus spp.) del centro y sur de México. Tesis doctoral. Instituto de Horticultura, Universidad Autónoma Chapingo, Chapingo, México. 120 p.
Núñez-Colín, C. A.; Nieto-Ángel, R.; Barrientos-Priego, A. F.; Sahagún-Castellanos, J.; Segura, S. and González-Andrés, F. 2008a. Variability of three regional sources of germplasm of tejocote (Crataegus spp.) from central and southern Mexico. Genet. Res. & Crop Evol. (Holland) 55:1159-1165.
Núñez-Colín, C. A.; Nieto-Ángel, R.; Barrientos-Priego, A. F.; Segura, S.; Sahagún-Castellanos, J. y González-Andrés, F. 2008b. Distribución y caracterización eco-climática del género Crataegus (Rosaceae subfam. Maloideae) en México. Revista Chapingo serie Horticultura (México) 14:177-184.
Phipps, J. B.; Robertson, K. R.; Smith, P. G. and Rohrer, J. R. 1990. A checklist of the subfamily Maloideae (Rosaceae). Can. J. Bot. 68:2209-2269.
Phipps, J. B. 1997. Monograph of northern mexican Crataegus (Rosaceae, subfam. Maloideae). SIDA Botanical Miscellany (USA) 15:1-94.
Phipps, J. B.; O’Kennon, R. J. and Lance, R. W. 2003. Hawthorns and medlars. Timber Press, Portland, USA. 139 p.
Zagaja, S. W. 1988. Exploración de recursos genéticos. In: Moore, J. N.; Janick, J. (eds.). Métodos genotécnicos en frutales. AGT Editor, México, D. F. México. p. 3-12.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 335-341
AMARRE, RENDIMIENTO Y CALIDAD DEL FRUTO EN NARANJA CON APLICACIÓN DE UN COMPLEJO HORMONAL*
FRUIT SET, YIELD AND QUALITY IN NAVEL ORANGE WITH THE APLICATION OF A HORMONAL COMPLEX
Juan José Galván-Luna1§, Florencio Briones-Encinia1, Patricio Rivera-Ortiz1, Luis Alonso Valdes-Aguilar2, Marcos Soto-Hernández3, Jorge Rodríguez-Alcázar3 y Ovidio Salazar-Salazar1
1Unidad Académica Multidisciplinaria Agronomía y Ciencias (UAMAC), Universidad Autónoma de Tamaulipas (UAT) Centro Universitario Adolfo López Mateos, C. P. 87149, Ciudad Victoria, Tamaulipas. Tel. y Fax 01 834 31 8 17 21, Ext. 2120. ([email protected]). 2Centro de Investigación en Química Aplicada (CIQA). Saltillo, Coahuila. ³Colegio de Postgraduados. Km 36.5 carretera México-Texcoco, Montecillo, Estado de México, C. P. 56230. Montecillo, México. §Autor para correspondencia: [email protected].
* Recibido: Noviembre, 2006
Aceptado: Abril, 2009
RESUMEN
En cítricos el uso de reguladores de crecimiento incrementa el amarre de frutos y en consecuencia la productividad. El objetivo del presente trabajo fue determinar el efecto de un complejo hormonal de origen natural (auxinas, giberelinas y citocininas) en el amarre, rendimiento y calidad del fruto en naranja (Citrus sinensis (L.) Osb) del grupo navel cvs. “Washington” y “Thomson”, en árboles de 15 años de edad. El estudio se realizó en el municipio de Ciudad Victoria, Tamaulipas, México durante 2004-2005. Se utilizó un arreglo en parcelas divididas en un diseño de bloques completos al azar con cinco tratamientos y cuatro repeticiones.Los resultados mostraron diferencias significativasentre tratamientos e interacción entre cultivares y t ratamientos en el número de frutos retenidos hasta los 129 días después de floración y porcentaje de amarre final. La dosis del complejo hormonal que incluye 32.2 ppm de auxinas, 32.2 ppm de giberelinas y83 .2 ppm de c i toc in inas más 19 .34 g L -1 de mic roe lementos mos t ró e l mayor amar re de lfruto con 1.2%.
Palabras clave: auxinas, citocininas, giberelinas, naranja navel “Washington” y “Thomson”.
ABSTRACT
The use of growth regulators improves fruit set in citrus; consequently, they increase crop productivity. The aim of the present research was to determine the effects of a hormonal complex of natural origin (auxins, gibberellins and cytokinins) on fruit set, yield and fruit quality of orange navel (Citrus sinensis (L.) Osb) cvs. “Washington” and “Thomson” of 15 years old trees. The study was conducted at Ciudad Victoria, Tamaulipas, Mexico, during 2004-2005. A split plot arrangement under a complete random block design with five treatments and four replications was used. Results show significant differences among treatments and for the interaction treatment x cultivar in the number of fruits retained until 129 days after flowering and in the percentage of fruit set. The hormonal complex at a dose that includes 32.2ppm of auxins, 32.2 ppm of gibberellins and 83.2 ppm of cytokinins plus 48.3 g L-1 show the greater effect at increasing fruit set, up to 1.2%. Key words: auxins, gibberellins cytokinins, orange navel “Washington” and “Thomsom”.
La importancia del cultivo de la naranja es incuestionable si se considera que México ocupa el tercer lugar a nivel mundial en producción, superado por Brasil y Estados Unidos de América

Juan José Galván-Luna et al.336 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
y que de acuerdo a la clasificación de productos alimenticios básicos, la naranja se encuentra en el 5º lugar (FAO, 2004). El principal destino de la producción de naranja es el mercado nacional en fresco, al cual se destina entre 80 y 85%, mientras que la agroindustria procesa entre 15 y 20% (Gómez y Schwentesius, 1997).
La mayoría de las variedades de naranja navel son de maduración temprana (se cosechan de noviembre a marzo) y sus frutos son partenocárpicos (sin semillas) por lo que son más sensibles a los cambios extremos de condiciones climáticas que las naranjas comunes (con semillas), por lo que la producción es variable en algunas regiones. También depende del clima (temperaturas cálidas en el día y templadas en la noche) la coloración de la cáscara, en condiciones de clima mediterráneo es en el que manifiestan con más intensidad los pigmentos que le dan color a la corteza y al jugo del fruto (Loussert, 1992). La corteza es ligeramente rugosa de grueso normal (5 mm) y fácil de pelar (Saunt, 1992), por lo general son frutos de bajo pH y alto contenido de jugo (Davies, 1986).
La variedad Washington navel se caracteriza por ser un árbol vigoroso y productivo, con hojas grandes y acusadas, el fruto es de tamaño medio a grande (65 a 75 mm de diámetro y de 200 a 250 g de peso), de forma redonda a ligeramente ovalado; corteza de color anaranjado intenso (cuando se presentan las condiciones climáticas adecuadas) ligeramente rugosa y gruesa, relativamente fácil de pelar. Los gajos se separan fácilmente y contiene una pulpa firme con mucho jugo, de sabor agradable (Agustí, 1999).
La naranja Thomson pertenece al grupo de naranjas navel de reciente aparición y tienen poca o nula comercialización, por lo tanto la información y problemática para su implantación, es escasa. En la zona centro de Tamaulipas se comercializa junto con el cultivar Washington navel, siendo los dos cultivares mas importantes de este grupo en Tamaulipas (García, 2004).
En general, la formación de flores en los cítricos se da en un proceso de desarrollo unitario, en dos fases: diferenciación de meristemo y desarrollo de la flor. Éste es un proceso continuo y termina con la apertura floral o antesis (Bernier, 1988). Sin embargo, en naranjas dulces el desarrollo del fruto sigue una curva sigmoidal, caracterizada por tres etapas bien diferenciadas (Agustí, 1999).
El amarre del fruto requiere de la conjunción de dos factores: un estímulo inicial que provoque el crecimiento del ovario y su capacidad de acumular metabolitos y una
disponibilidad suficiente de éstos. Esta disponibilidad es crítica durante la fase de abscisión y determina el cuajado final del fruto. Es el cuajado, o amarre de frutos y no la floración el factor que determina la cosecha en los cítricos (Guardiola, 2004).
En la región citrícola del centro del estado de Tamaulipas, la superficie establecida de naranja navel es de 1 106 ha (SAGARPA, 2001). Las temperaturas cálidas que se presentan en los períodos de floración, cuajado y crecimiento del fruto se consideran la causa principal de la problemática asociada a dichos procesos y por consecuencia de la baja producción de este grupo de naranjas cultivadas en la región (García, 2004).
El tamaño final del fruto está regulado por un conjunto de factores de índole e incidencia variable. La imposibilidad de su control y su interrelación complican su estudio y solo permiten tener un conocimiento parcial de algunos de ellos, lo que unido a diferencias varietales, edáficas, etc., obliga al estudio fragmentado de los mismos (Agustí, 2001).
En naranja cv “Valencia” se determinó que la aplicación de un complejo hormonal a base de auxinas, giberelinas y citocininas incrementó el rendimiento de fruto y jugo concentrado con la aplicación de 3 L ha-1 (Ruiz, 1999).
La naranja navel es un cultivo comercial en el cual sólo 1% de las flores alcanzan a llegar a fruto maduro (porcentaje de amarre). En consecuencia se han ensayado diversas técnicas para incrementar el amarre y rendimiento de frutos; entre ellas, la polinización cruzada, el anillado de troncos, la aplicación de fitorreguladores y la práctica del riego por encima del dosel de los árboles con el objeto de reducir las altas temperaturas (García, 2004).
En el estado de Tamaulipas, el enorme potencial que representa el uso de los reguladores no ha sido debidamente explotado y los resultados experimentales disponibles corresponden a ambientes diferentes. Si bien las aplicaciones hormonales no constituyen una panacea universal, existen determinados aspectos del desarrollo que son en mayor o menor medida susceptibles de modificación (Talón, 2001).
El uso de reguladores de crecimiento es una alternativa para incrementar el porciento de amarre de frutos, lo cual se traduce en incrementos de la producción. En la época de

Amarre, rendimieto y calidad del fruto en naranja con aplicación de un complejo hormonal 337
floración, el uso de reguladores hormonales complejos, han mostrado ser más efectivos para mejorar la producción que los productos que contienen una sola hormona (Ruiz, 1999).
Ante la falta de investigación sobre el uso de reguladores del crecimiento en la región y el bajo porcentaje de amarre observado en las naranjas navel, el presente trabajo de investigación tuvo como objetivo evaluar el efecto de un complejo hormonal de origen natural (auxinas, giberelinas y citocininas) en el amarre, rendimiento y calidad del fruto en naranja en la zona centro del estado de Tamaulipas.
Ubicación y características de la unidad experimental. El experimento se realizó en el período de febrero de 2004 a febrero de 2005, en árboles de los cultivares de naranja navel “Washington” y “Thomson” de 15 años de edad, ubicados en el municipio de Ciudad Victoria, Tamaulipas, México, plantados en suelo calcáreo (34.9% de carbonatos), arcilloso, con un pH de 8.2, con nivel deficiente de zinc y bajo contenido de nitrógeno, fósforo y magnesio, con riego por micro aspersión. Se aplicó un complejo hormonal de origen natural que contiene auxinas (ácido indolacético 32.2 ppm), giberelinas (32.2 ppm), citocininas (zeatina 83.2 ppm) y microelementos (19.34 g L-1) en los porcentajes siguientes: (Mn 0.12%, Zn 0.37%, Fe 0.49%, Mg 0.14%, B 0.30% y S 0.44%) utilizando como fuente el producto Biozyme * TF, que en términos porcentuales contiene 1.86% de microelementos, 78.87 de fitohormonas y 19.27 de diluyentes y acondicionadores.
Variables respuesta. Las variables fueron: número de flores por árbol (calculadas tomando como base las ramas terminales de los cuatro puntos cardinales contabilizando el número de flores y multiplicado por el número de ramas en el árbol), el número de frutos por árbol para determinar el porcentaje de amarre final (PAF) y el número de frutos retenidos (NFR) hasta los 129 días después de la floración
(DDF) (etapa del crecimiento inicial del fruto en la que ya no se observó caída de fruto). Se determinó el diámetro polar (DP) y ecuatorial (DE) utilizando el vernier Scienceware de 150 mm. El peso del fruto (PF) se cuantificó con una balanza electrónica de precisión ohaus scout, el grosor de la cáscara (GC) con el mismo vernier. Se determinaron los grados brix (GB) con el refractómetro manual Atago ATC-1E. El pH del fruto se determinó con el potenciómetro manual Corning y el espacio de color L* a* b* con el colorímetro Minolta CR-300. Con base en el diámetro polar y ecuatorial, tomados durante la fase de fructificación, a los 100, 150, 200, 250 y 300 días después de plena floración se estimaron las curvas de crecimiento de acuerdo con las proporciones del complejo hormonal utilizado en la presente investigación.
Tratamientos y diseño experimental. Se utilizó un diseño en bloques completos al azar con cuatro repeticiones, con arreglo en parcelas divididas, donde la parcela grande correspondió a los cultivares y la parcela chica a los cinco tratamientos descritos en el Cuadro 1. Se realizaron dos aplicaciones una en plena floración y la otra a la caída de pétalos (15 y 31 de marzo en el cultivar de naranja navel “Washington” y 19 y 31 de marzo de 2004 en “Thomson”). Cada aplicación se efectuó de 7:00 a 10:00 con una mochila manual utilizando 3 L por árbol y las cantidades del producto de acuerdo a las dosis descritas, mismas que para efectos de identificación se clasificaron en dosis baja, media, alta y muy alta, teniendo un testigo al cuál sólo se le aplicó agua. En cada tratamiento se adicionó una mezcla de ácidos con el producto “Bionex” a razón de 2 cc por L de agua para disminuir el pH.
Análisis estadístico. Se realizó el análisis de varianza y la prueba de comparación de medias de Tukey con α= 0.05 con el paquete estadístico de SAS (2000). Para el análisis del número de frutos se aplicó la transformación raíz cuadrada y para el porcentaje de amarre la transformación seno inverso, éstas con la finalidad de darle una distribución normal.
Tratamiento (dosis) Auxinas (ppm) Giberelinas n (ppm) Citocininas (ppm) Microelementos (gL-1)TestigoBajaMediaAltaMuy alta
0016.132.248.364.4
0016.132.248.364.4
0041.683.2124.8166.4
009.1718.3427.5136.68
Cuadro 1. Tratamientos aplicados en ppm de complejo hormonal que incluye auxinas, giberelinas, citocininas y g L-1 de microelementos en los cultivares de naranja navel “Washington” y “Thomson”, en la zona centro de Tamaulipas. 2005.

Juan José Galván-Luna et al.338 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Amarre del fruto. El mayor porcentaje de amarre final se observó con la dosis media (1.22%), resultando significativamente (p<0.05) superior al testigo (0.40%), no hubo diferencia significativa en el número de
frutos retenidos entre las dosis media, alta y muy alta; sin embargo, el número de frutos retenidos en estas dosis fue aproximadamente 100% mayor que el tratamiento testigo, Cuadro 2.
mayor número de frutos retenidos fue 48.3 de auxinas, 48.3 de giberelinas y 124.8 ppm de citocininas más 29.01 g L-1
de microlementos, con una media de 34.5 frutos superior al testigo (12.38) respectivamente, con mayor número de frutos en promedio observados en el cultivar Thomson para las proporciones mas bajas (Figura. 1).
Dosis Amarre Final (%) Numero de frutos retenidos (Núm)Testigo
Baja
Media
Alta
Muy alta
0.40 ± 0.325 b
0.72 ± 0.735 b
1.22 ± 0.675 a
1.15 ± 0.770 a
0.76 ± 0.420 b
12.38 ± 9.80 b
18.63 ± 7.73 b
31.75 ± 12.19 a
34.50 ± 18.08 a
29.50 ± 14.73 a
05
1015202530354045
Núm
ero de frutos
Testigo Dosisbaja
Dosismedia
Dosisalta
Dosismuy alta
Tratamientos hormonales aplicados.
WnTh
Figura 1. Número de frutos retenidos a los 129 días después de floración en naranja navel Washington (Wn) y Thomson (Th) con las dosis del complejo hormonal que contiene auxinas, giberelinas y citocininas. Zona centro de Tamaulipas. 2005.
Cuadro 2. Porcentaje de amarre final y número de frutos retenidos en naranja navel “Washington” y “Thomson” de acuerdo con las dosis aplicadas de complejo hormonal. Valores zona centro de Tamaulipas. 2005.
Medias con la misma letra no son estadísticamente iguales (Tukey, 0.05). CV. 13.05. X ± S.
El número de frutos retenidos hasta los 129 días después de floración y porcentaje de amarre final, mostraron diferencia significativa (p<0.05) entre los cultivares estudiados, aún cuando estadísticamente no se observaron diferencias con los tratamientos aplicados en las dosis media, alta y muy alta, la proporción del complejo hormonal que resultó con
Tratamientos hormonales aplicados
Núm
ero
de fr
utos
Testigo Dosis Dosis Dosis Dosis baja media alta muy alta

Amarre, rendimieto y calidad del fruto en naranja con aplicación de un complejo hormonal 339
La dosis del complejo hormonal que significativamente mostró mayor porcentaje de amarre final fueron las que incluyeron 32.2 de auxinas, 32.2 de giberelinas y 83.2 ppm de citocininas más 19.34 g L-1 de microelementos y 48.3 de auxinas, 48.3 de giberelinas y 124.8 ppm de citocininas más 29.01 g L-1 de microelementos (1.22 y 1.15%, respectivamente), diferentes (p<0.05) al testigo (0.40%),
giberelinas incrementan su concentración en los ovarios en desarrollo durante el período de antesis como si formaran parte del estímulo hormonal que reactiva la división celular y estimula el crecimiento del fruto posibilitando su amarre. De acuerdo con Talón (2001) las aplicaciones exógenas de auxinas para mejorar el amarre del fruto no son eficaces, por lo que se desconoce su función en el amarre de frutos cítricos. Sin embargo, al aplicarlas en un complejo hormonal, como en este caso, aún cuando se desconoce su función en aplicaciones individuales, en conjunto los tres grupos de hormonas parecen potenciar los efectos individuales al incrementar los porcentajes de amarre del fruto.
Los resultados de los porcentajes de amarre obtenidos están de acuerdo con Agustí y Almela (1991) que señalan 1% para naranja valencia y 0.2% para cítricos sin semilla, por lo que al incrementar a más de 1% con la aplicación del complejo hormonal, resulta una práctica favorable.
Peso del fruto. El peso del fruto no mostró diferencia significativa entre las proporciones hormonales, los resultados en promedio fueron de 325.78 g en “Washington”
0
0.2
0.4
0.6
0.8
1
1.2
1.4
Amarre (%
)
Testigo Dosisbaja
Dosismedia
Dosisalta
Dosismuy alta
Dosis de fitohormonas
WnTh
Figura 2. Porcentaje de amarre de frutos en naranja navel “Washington” (Wn) y “Thomson” (Th) con la aplicación de complejo hormonal que incluye auxinas, giberelinas y citocininas. Zona centro de Tamaulipas. 2005.
Figura 2. Resultados similares fueron reportados por Ruiz (1999), en el cultivar “Valencia”, con el rendimiento y producción de jugo, lo que confirma que las aplicaciones hormonales estimulan el amarre del fruto y la producción del mismo (Talón, 2001) y que la imposibilidad del control de los factores que influyen en el tamaño final del fruto determina el estudio fragmentado de los mismos (Agustí, 2001).
Dosis de fitohormonas
Am
arre
(%)
El amarre del fruto resulta de la conjunción de dos factores; la concentración de hormonas apropiadas, que estimulen el crecimiento del fruto e impiden su abscisión y el suministro de metabolitos suficiente para atender las necesidades nutricionales (Guardiola, 2004), en este trabajo se detectaron diferencias al aplicar las diferentes dosis del complejo hormonal, dado que el manejo del huerto fue similar para todos los tratamientos.
Las giberelinas en la fase de fructificación, reactivan el crecimiento del fruto, atraen nutrientes hacia los frutos y parecen sostener el crecimiento hasta la caída de junio, en este sentido las giberelinas son factores que limitan y condicionan el amarre del fruto (Talón, 2001), de ahí que al aplicarlas exógenamente incrementan el porcentaje de amarre. Si bien su contenido no es limitante en la mayor parte de los casos, también auxinas y citocininas están implicadas en el amarre del fruto. Esta participación se demuestra utilizando inhibidores de su síntesis o su acción, que interfieren con el cuajado; además, en unas pocas ocasiones se ha descrito que el amarre puede aumentarse mediante la aplicación de estas hormonas (Guardiola, 2004). Las citocininas al igual que las
Testigo Dosis Dosis Dosis Dosis baja media alta muy alta
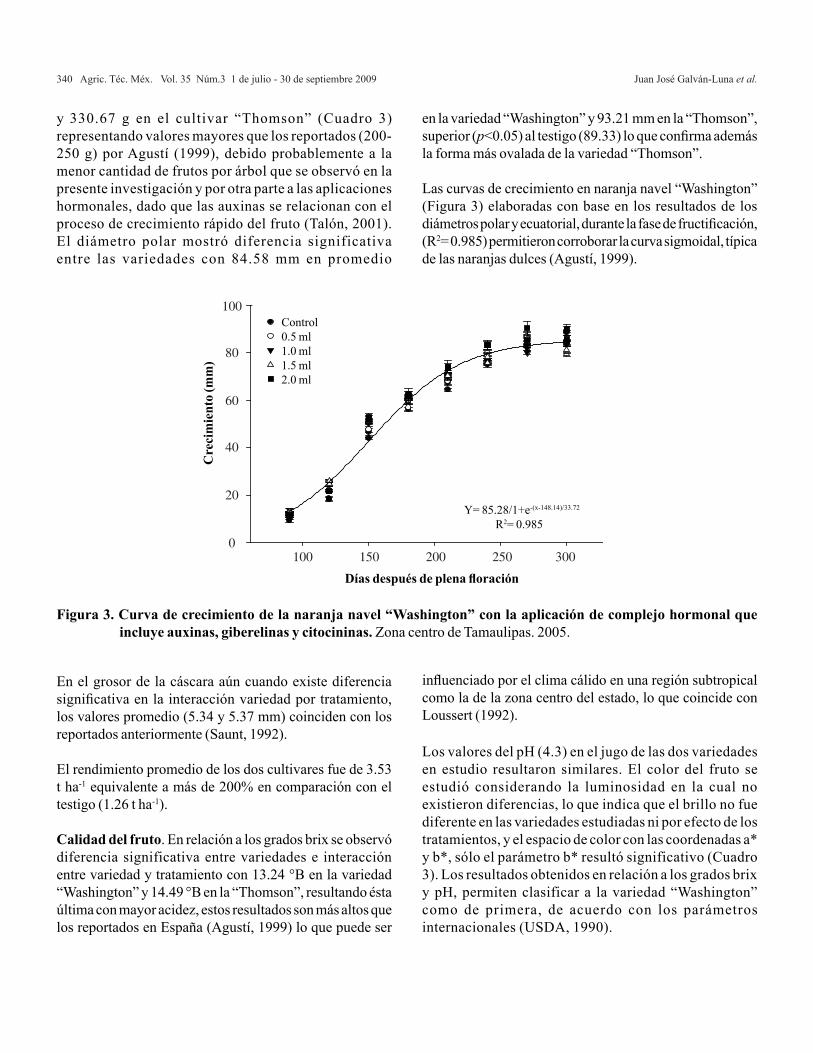
Juan José Galván-Luna et al.340 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
y 330.67 g en el cultivar “Thomson” (Cuadro 3) representando valores mayores que los reportados (200-250 g) por Agustí (1999), debido probablemente a la menor cantidad de frutos por árbol que se observó en la presente investigación y por otra parte a las aplicaciones hormonales, dado que las auxinas se relacionan con el proceso de crecimiento rápido del fruto (Talón, 2001). El diámetro polar mostró diferencia significativa entre las variedades con 84.58 mm en promedio
influenciado por el clima cálido en una región subtropical como la de la zona centro del estado, lo que coincide con Loussert (1992).
Los valores del pH (4.3) en el jugo de las dos variedades en estudio resultaron similares. El color del fruto se estudió considerando la luminosidad en la cual no existieron diferencias, lo que indica que el brillo no fue diferente en las variedades estudiadas ni por efecto de los tratamientos, y el espacio de color con las coordenadas a* y b*, sólo el parámetro b* resultó significativo (Cuadro 3). Los resultados obtenidos en relación a los grados brix y pH, permiten clasificar a la variedad “Washington” como de primera, de acuerdo con los parámetros internacionales (USDA, 1990).
Y= 85.28/1+e-(x-148.14)/33.72
R2=0.985
Días después de plena floración
100 150 200 250 300
Crecimiento
(mm
)
0
20
40
60
80
100Control0.5 ml1.0 ml1.5 ml2.0 ml
Figura 3. Curva de crecimiento de la naranja navel “Washington” con la aplicación de complejo hormonal que incluye auxinas, giberelinas y citocininas. Zona centro de Tamaulipas. 2005.
en la variedad “Washington” y 93.21 mm en la “Thomson”, superior (p<0.05) al testigo (89.33) lo que confirma además la forma más ovalada de la variedad “Thomson”.
Las curvas de crecimiento en naranja navel “Washington” (Figura 3) elaboradas con base en los resultados de los diámetros polar y ecuatorial, durante la fase de fructificación, (R2= 0.985) permitieron corroborar la curva sigmoidal, típica de las naranjas dulces (Agustí, 1999).
Días después de plena floración100 150 200 250 300
100
80
60
40
20
0
Cre
cim
ient
o (m
m)
Control0.5 ml1.0 ml1.5 ml2.0 ml
Y= 85.28/1+e-(x-148.14)/33.72
R2= 0.985
En el grosor de la cáscara aún cuando existe diferencia significativa en la interacción variedad por tratamiento, los valores promedio (5.34 y 5.37 mm) coinciden con los reportados anteriormente (Saunt, 1992).
El rendimiento promedio de los dos cultivares fue de 3.53 t ha-1 equivalente a más de 200% en comparación con el testigo (1.26 t ha-1).
Calidad del fruto. En relación a los grados brix se observó diferencia significativa entre variedades e interacción entre variedad y tratamiento con 13.24 °B en la variedad “Washington” y 14.49 °B en la “Thomson”, resultando ésta última con mayor acidez, estos resultados son más altos que los reportados en España (Agustí, 1999) lo que puede ser
Amarre, rendimieto y calidad del fruto en naranja con aplicación de un complejo hormonal 341
Gómez, C. M. A. y Schwentesius, R. 1997. La agroindustria de naranja en México. Edit. Centro de Investigaciones Económicas, Sociales y Tecnológicas de la Agroindustr ia y la Agricultura Mundial (CIESTAAM), Chapingo, Estado de México.
Guardiola, B. J. L. 2004. Cuajado del fruto, aspectos hormonales y nutricionales. Universidad Politécnica de Valencia, España. Mayo 2004.
Food and Agriculture Organization of the United Nations (FAO). 2004. FAOSTAT. Agriculture. www.fao.org.
Loussert, R. 1992. Los agrios. Ed. Mundi prensa, Madrid, España. 303 p.Ruiz, B. O. 1999. Fisiología de la floración y reguladores
de crecimiento. In: IV Curso Internacional de Citricultura. Unidad Académica Multidisciplinaria Agronomía y Ciencias, UAT. Cd. Victoria, Tamaulipas. 21-25 de septiembre de 1999.
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA), 2001. Inventario de productores citrícolas 2001. Sistemas de Información Agropecuaria. Alianza para el Campo, SAGARPA. Gobierno de estado de Tamaulipas.
Saunt, J. 1992. Variedades de cítricos del mundo. Ed. Edipublic, Valencia, España. 126 p.
Statistical Analysis System Institute (SAS) 2000. Statistical Analysis System. Version 8.01. Cary, NC., USA.
Talón, M. 2001. Revista Comunitat Valencia Agraria No. 15.United States Department of Agriculture (USDA). 1990.
Quality control manual for citrus processing plants. Edit. Intercit, Florida, USA.
Cultivar PF (g) DP (mm) GC (mm) °B pH b*
Washington 325.78 a 84.58 a 5.34 a 13.24 a 4.3 a 59.73 aThomson 330.67 a 93.21 b 5.37 a 14.49 b 4.3 a 58.98 b
Cuadro 3. Características del fruto con la aplicación de un complejo hormonal que incluye auxinas, giberelinas y citocininas. Zona centro de Tamaulipas. 2005.
PF= peso del fruto; DP= diámetro polar; GC= grosor de la cáscara; oB= grados Brix; b*= parámetro b del color. Medias con la misma letra no son estadísticamente iguales Tukey (0.05).
El amarre del fruto se incrementa hasta 300% con la aplicación de la dosis media del complejo hormonal que incluye auxinas (32.2 ppm), giberelinas (32.2 ppm) y citocininas (83.2 ppm).
El peso del fruto no se modifica con la aplicación de auxinas, giberelinas y citocininas.
Los cultivares “Washington” y “Thomson” muestran respuesta diferencial a la aplicación de complejos hormonales en el porciento de amarre del fruto, número de frutos retenidos, grosor de la cáscara, grados brix y pH.
LITERATURA CITADA
Agustí, M. y Almela, V. 1991. Aplicación de fitorreguladores en citricultura. Editorial Aedos. Barcelona, España. 261 p.
Agustí, M. 1999. Citricultura. Ediciones Mundi-prensa. España. 418 p.
Bernier, G. 1988. The control of floral evocation and morphogenesis. Ann. Rev. Plant Physiol. 93:175-219.
Davies, F. S. 1986. The navel orange. Hort. Rev. 8:129-175.
García, D. M. A. 2004. Efecto de la nebulización en la temperatura y humedad del aire y su relación con el cuajado y rendimiento del fruto en naranjo navel. Agrociencia 38:643-651.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 342-348
CARACTERÍSTICAS DE CALIDAD DE FRUTOS DE PAPAYA MARADOL EN LA MADUREZ DE CONSUMO*
QUALITY CHARACTERISTICS IN MARADOL PAPAYA FRUITS AT THE COMSUMPTION RIPENESS STAGE
Felipe Santamaría Basulto1§, Raúl Díaz Plaza1, Enrique Sauri Duch2, Francisco Espadas y Gil3, Jorge Manuel Santamaría Fernández3 y Alfonso Larqué Saavedra3
1Campo Experimental Mocochá, INIFAP, km. 24 carretera antigua Mérida-Motul, Mocochá, Yucatán. ([email protected]). Tel. 01 991 9 16 22 15. 2 Instituto Tecnológico de Mérida. Km 5 carretera Mérida-Progreso, A. P. 9-11 Mérida, Yucatán, México, C. P. 97118. ([email protected]). Tel. y Fax (99) 44 84 79. 3 Centro de Investigación Científica de Yucatán. Calle 43 Núm. 130 Colonia Chuburná de Hidalgo. C. P. 97200 Mérida, Yucatán, México. ([email protected]), ([email protected]). Tel. 01 999 9 42 83 30 Ext. 215 y 260. §Autor para correspondencia: [email protected].
* Recibido: Enero, 2009
Aceptado: Septiembre, 2009
RESUMEN El color de la cáscara es la característica más utilizada para evaluar la maduración de los frutos de papaya; las recomendaciones para la cosecha, comercialización y consumo se basan en la aparición de porcentajes de color verde, amarillo y naranja. Esas escalas subjetivas son ambiguas y crean dificultades en la interpretación del observador. Por lo anterior, el objetivo fue evaluar los principales cambios de apariencia, firmeza y sólidos solubles totales de papaya ‘maradol’ durante su maduración poscosecha para proponer índices objetivos que describan el estado de madurez de consumo. En noviembre de 2005 se definieron seis estados de maduración en frutos obtenidos en Yucatán y se evaluó el color del fruto, contenido de clorofilas y carotenoides, firmeza de la pulpa, azúcares y contenido de sólidos solubles totales. En marzo de 2006 se obtuvieron las características de los frutos en la madurez de consumo en dos localidades y se compararon con los resultados anteriores. La madurez de consumo de papaya maradol se alcanza entre los 13 y 15 días después de la cosecha en condiciones de almacenamiento de 23 ± 1 °C y 75% de humedad relativa. El ángulo del tono de la cáscara entre 70 y 80°, el contenido de sólidos solubles totales entre 10 y 11.5 °Brix, y la firmeza de la pulpa de 4.7 a 6.9 newtons permitieron diferenciar dos estados de madurez de consumo.
Palabras clave: Carica papaya L., color CIELAB, índices de madurez, papaya maradol.
ABSTRACT
Skin fruit color is the main characteristic to evaluate papaya fruit ripening; recommendations for harvest, commercialization and consumption, are based in green, yellow and orange color percentages. These subjective scales are difficult for interpretation. The purpose of the present work was to evaluate the main changes in appearance, firmness and total soluble solids occurring during postharvest ripening of maradol papaya fruits in order to define fruit characteristics that can be suggested as quality standards for consumption maturity. In November 2005, six maturity stages were identified and parameters such as fruit color, skin chlorophyll and carotenoids content, pulp firmness, sugars and total soluble solids were measured along them. In March 2006, fruit characteristics in consumption maturity of fruits harvested from two locations were obtained and compared with previous results. It was found that the consumption maturity stage is reached between 13 and 15 days after harvest when fruits were stored at 23±1°C and

Características de calidad de frutos de papaya maradol en la madurez de consumo 343
75% of relative humidity. Skin°HUE values between 70 and 80, total soluble solids content between 10 and 11.5 °Brix and pulp firmness between 4.7 to 6.9 newtons allowed the differentiation of two consumption maturity stages.
Key words: Carica papaya L., CIELAB color, maradol papaya, maturity index.
El color de la cáscara es la característica más utilizada para evaluar el estado de maduración de los frutos de papaya. Las recomendaciones para la cosecha se basan en el cambio del color verde oscuro a verde claro y la aparición de tonos amarillos en el extremo distal (Kader, 2004). La comercialización se realiza desde el estado de rompimiento del color verde a ¼, ½ y ¾ de madurez, mientras que el consumo se recomienda cuando la cáscara de los frutos presenta 75% o más de color amarillo (Zhou et al., 2004).
Las escalas subjetivas se continúan usando en papaya como en los cultivares Tainung (Rocha et al., 2005), Caliman (De Morais et al., 2007) y en la descripción de nuevos híbridos derivados de maradol (Alonso et al., 2008). Se reportan también mediciones objetivas del color del fruto de papaya Golden en valores L*, a*, b* (De Oliveira et al., 2002) y el ángulo del tono de la cáscara (Bron y Jacomino, 2006). Con las mediciones objetivas de color se evitan subjetividades, ya que el número de franjas amarillas y los porcentajes de coloración en la cáscara puede crear dificultades en la interpretación del observador. Por otra parte, el patrón de coloración en papaya no esta restringido a las franjas amarillas y pueden aparecer sitios de color amarillo en casi cualquier parte de la cáscara del fruto (Peleg y Gómez-Brito, 1975).
En México, la principal variedad de papaya cultivada es maradol. La norma mexicana NMX-FF-041-SCFI-2007, describe el color externo de esta variedad con base en la designación de rayas y porcentajes de color verde, amarillo o naranja, de acuerdo con siete grados de maduración que incluyen a los frutos en madurez fisiológica y en madurez de consumo. Los trabajos que describen las características del fruto con variables objetivas están enfocados al efecto de la antracnosis (Acosta et al., 2001), o describen los cambios de color en las etapas iniciales o finales de la maduración (Pérez-Carrillo y Yahia, 2004; Hernández et al., 2007). Dado lo anterior, el objetivo del presente trabajo fue evaluar los principales cambios de apariencia
y sólidos solubles totales de papaya maradol durante su maduración poscosecha para proponer índices objetivos que describan el estado de madurez de consumo.
En noviembre de 2005 se obtuvieron frutos en una parcela comercial ubicada en la localidad de Techoh, municipio de Tixkokob, Yucatán, los frutos se cosecharon cuando presentaron el cambio de color verde intenso a verde claro con una franja amarilla (Gobierno del estado de Yucatán, 1997) y frutos inmaduros que presentaron la cáscara de color verde intenso sin ninguna coloración amarilla. Al día siguiente de la cosecha los frutos se lavaron y se trataron con inmersión en la solución del fungicida azoxystrobin (1 mL L-1 agua) por 1.5 min. Los frutos se almacenaron a 23 ± 1 °C y 75% de humedad relativa por 15 días, tiempo en el cual alcanzaron la madurez de consumo. Durante este tiempo se definieron seis estados de maduración tomando muestras de 3 frutos después de 1, 4, 7, 11, 13 y 15 días de realizada la cosecha. Se evaluó el contenido de clorofilas y carotenoides totales en la cáscara del fruto, la firmeza de la pulpa, contenido de sólidos solubles totales, azúcares totales, azúcares reductores y color de cáscara y pulpa.
En el segundo experimento, los frutos se cosecharon en dos plantaciones comerciales, en marzo de 2006 en la parcela Casa Blanca ubicada en el municipio de Ucú, y en abril de 2006 en la plantación San Pedro ubicada en el municipio de Sucilá. Los valores de color de cáscara se presentan en el Cuadro 1. Los frutos fueron tratados y almacenados en las mismas condiciones descritas en el experimento anterior.
Se definieron dos estados de madurez de consumo excluyendo los frutos que presentaron resina al ser cortados y los frutos con alteraciones del aspecto, aroma o pudriciones. Los frutos del primer estado de madurez de consumo (MC1) presentaron toda la cáscara y pulpa de color anaranjado característico de la variedad maradol, y la pulpa presenta una consistencia adecuada para su consumo. En el segundo
Parámetro Ucú SuciláL* (luminosidad) 43.12 ± 0.68 46.26 ± 0.89a* -18.40 ± 0.39 -19.30 ± 0.10b* 30.00 ± 0.74 31.87 ± 0.78Ángulo de tono 121.93 ± 1.11 122.11 ± 0.85
Cuadro 1. Color de la cáscara al momento de la cosecha de los frutos de dos localidades

Felipe Santamaría Basulto et al.344 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
estado de madurez de consumo (MC2) los frutos son de condiciones similares al estado MC1, pero con mayor intensidad en el color de la cáscara y pulpa, aunque la pulpa es más suave todavía es adecuada para su consumo.
El contenido de clorofila y carotenoides se determinó de acuerdo con Wellburn (1994), para ello se extrajo una muestra de aproximadamente 1 g de la cáscara y se maceró en un mortero con 1 mL de acetona al 80%. El extracto se transfirió a un tubo Eppendorf de 1.5 mL y se centrifugó a 13 400 x g por 10 min. Posteriormente el extracto se aforó a 5 mL. Se midió la absorbancia de esta dilución en un espectrofotómetro Beckman Coulter modelo DU 650 a tres longitudes de onda: 470, 633 y 645 nm.
El color se midió con un colorímetro Minolta modelo CR-200, los datos se reportan en valores L*, a*, b* de la escala CIELAB (McGuire, 1992). En cada fruto se hicieron 6 lecturas en la cáscara que corresponden al área cercana al pedúnculo, al centro y al área cercana al ápice en los dos lados opuestos del fruto. Cada fruto se cortó en dos y se midió el color de la pulpa en la parte media entre la cáscara y la cavidad de las semillas, los valores a* y b* se usaron para calcular el ángulo de tono (H= arctan (b*/a*)).
Para la determinación de la firmeza, se tomaron seis muestras de pulpa en los mismo lugares donde se midió el color, se eliminó la cáscara y las semillas y se sacó una muestra cuadrangular de aproximadamente 4.5 cm por lado. La firmeza de la pulpa se evaluó con una prensa Instron 4442 con un punzón de acero inoxidable de 8 mm de diámetro y punta plana, la cual penetró 15 mm a una velocidad de 25 mm s-1 . Los resultados se reportan en newtons (N).
La de te rminac ión de azúcares se rea l izó por espectrofotometría visible utilizando un espectrofotómetro Beckman Coulter modelo DU 650, los azúcares totales se determinaron por el método fenol-sulfúrico midiendo la absorbancia a 490 nm (Dubois et al., 1956), los azúcares reductores se determinaron por el método DNS midiendo la absorbancia a 550 nm (Miller, 1959). Los sólidos solubles totales (SST) se midieron con un refractómetro digital Palette PR-101α. Los resultados se graficaron calculando la media y la desviación estándar. A los resultados se les realizó un análisis de varianza (diseño completamente al azar) y las medias se compararon con la prueba de Tukey (p≤0.05)
usando el programa Statgraphics Plus 4.1 (Statistical Graphics Corp.), en cada estado de maduración se tomaron tres frutos y cada fruto se consideró una repetición.
Los resultados del primer experimento mostraron que la concentración de carotenoides totales presentó un incremento constante, al día 1 después de la cosecha los frutos tuvieron un contenido de 13.1 µg g-1 de cáscara y se fue incrementando paulatinamente hasta 67 µg g-1 de cáscara a los 13 y 15 días (Figura 1). Para el caso de la clorofila, los frutos presentaron un decremento constante de 290 µg g-1 al inicio de la maduración hasta 6.5 µg g-1 de cáscara en los estados de madurez de consumo.
En cuanto al color, la luminosidad de la cáscara se incrementó durante la maduración, los frutos verdes tuvieron un valor promedio de L* de 42.5 y se incrementó a 57 y 58 en la madurez de consumo (Figura 2A). En contraste, los valores de L* en la pulpa decrecieron desde un promedio de 70 en los frutos verdes a valores cercanos a 51 al final de la maduración. De todos los componentes del color, el componente a* mostró los cambios más notables, en la cáscara presentó valores negativos en los frutos verdes y hasta 11 días después de la cosecha, después alcanzó valores positivos a los 13 y 15 días después de la cosecha (Figura 2B). En la pulpa, los valores fueron positivos desde el primer día, después los valores se incrementaron hasta alcanzar promedios mayores de +23 en la madurez de consumo.
0
50
100
150
200
250
300
350
V 1 4 7 11 13 15
Días después de la cosecha
clorofilas mg g
-1de peso fresco
0
10
20
30
40
50
60
70
80
90
100
carotenoides mg g
-1de peso fresco
clorof ilascarotenoidesa a
ab
bbc
c c
a a
ab ab
c
dd
Figura 1. Contenido de clorofilas y carotenoides en la cáscara de frutos de papaya maradol durante su maduración poscosecha. Cada punto es el promedio de tres frutos ± desviación estándar. Para cada variable las medias con la misma letra son estadísticamente iguales (Tukey, 0.05).
Car
oten
oide
s mg
g-1 d
e pes
o fr
esco
Días después dela cosecha
Clo
rofil
as m
g g-1
de p
eso
fres
co

Características de calidad de frutos de papaya maradol en la madurez de consumo 345
En la mayoría de los frutos, el proceso de maduración involucra la pérdida de clorofila y la síntesis de nuevos pigmentos como carotenoides y el desenmascaramiento de otros pigmentos que se han formado durante el desarrollo del fruto (Aked, 2000; Ferrer et al., 2005). Los datos sugieren que en papaya maradol los primeros cambios del color se dan como resultado de un incremento del color amarillo y no por la pérdida de color verde,ya que el valor negativo de a* y el contenido de clorofilas de los frutos verdes es similar a los frutos al día siguiente de la cosecha. Los frutos de papaya maradol inicialmente son verde amarillento y cambian alanaranjado-amarillo en los estados de madurez de consumo.
Los frutos verdes presentaron una firmeza de pulpa de 150 N, la cual fue disminuyendo paulatinamente a 117 N a los siete días después de la cosecha, del día 7 al día 11 se observa una disminución drástica al bajar a 9.8 N, posteriormente la firmeza alcanza a 6.7 N en la madurez de consumo (Figura 3A).
0
10
20
30
40
50
60
70
80
V 1 4 7 11 13 15
Valor
L*
aa
a
bbc
ab
bcc c c
b bc c bc
-20
-15-10
-5
05
10
15
2025
30
0 1 4 7 11 13 15
Valor
a*
a
b
cc
c cc
aa a
b
ab
c
c
0
20
40
60
80
100
120
140
0 1 4 7 11 13 15
Ángu
lo de
tono
( o
)c
a
a abbc
c
de
bbcc
e
c c
10
15
20
25
30
35
40
45
50
55
0 1 4 7 11 13 15
Valor
b*
CÁSCARAPULPA
d
a
a
b
de
cd
d
cc
e
bbc
cd d
Figura 2. Componentes del color de cáscara y pulpa durante la maduración de frutos de papaya maradol. Valores L* (A), a* (B), b* (C) y ángulo de tono (D). Cada punto es el promedio de tres frutos ± desviación estándar. Para cada variable las medias con la misma letra son estadísticamente iguales (Tukey, 0.05).
C D
A B
Los valores de b* fueron similares en la cáscara y en la pulpa, los frutos verdes presentaron promedios de +20.7 y +17 en la cáscara y pulpa respectivamente y en los frutos evaluados a los siete días después de la cosecha el promedio se incrementó a +35, luego los valores de la cáscara se incrementan hasta cerca de + 46 en la madurez de consumo, en la pulpa los valores fueron ligeramente menores alcanzando promedios cercanos a +36 (Figura 2C).
El tono del color de la cáscara y pulpa mostró que los frutos fueron inicialmente verde amarillento y cambian hacia el anaranjado-amarillo. El color amarillo se presentó en la cáscara a los 11 días después de la cosecha, cuando los frutos alcanzaron valores cercanos a 90° (Figura 2D), en cambio la pulpa fue amarilla desde el primer día y desarrolló un color amarillo anaranjado desde los cuatro días después de la cosecha al alcanzar valores cercanos a 60°.
Los datos de L* y a* son similares a los reportados por Pérez-Carrillo y Yahia (2004) en la cáscara y por Hernández et al. (2007) en la pulpa de frutos maduros de papaya maradol.
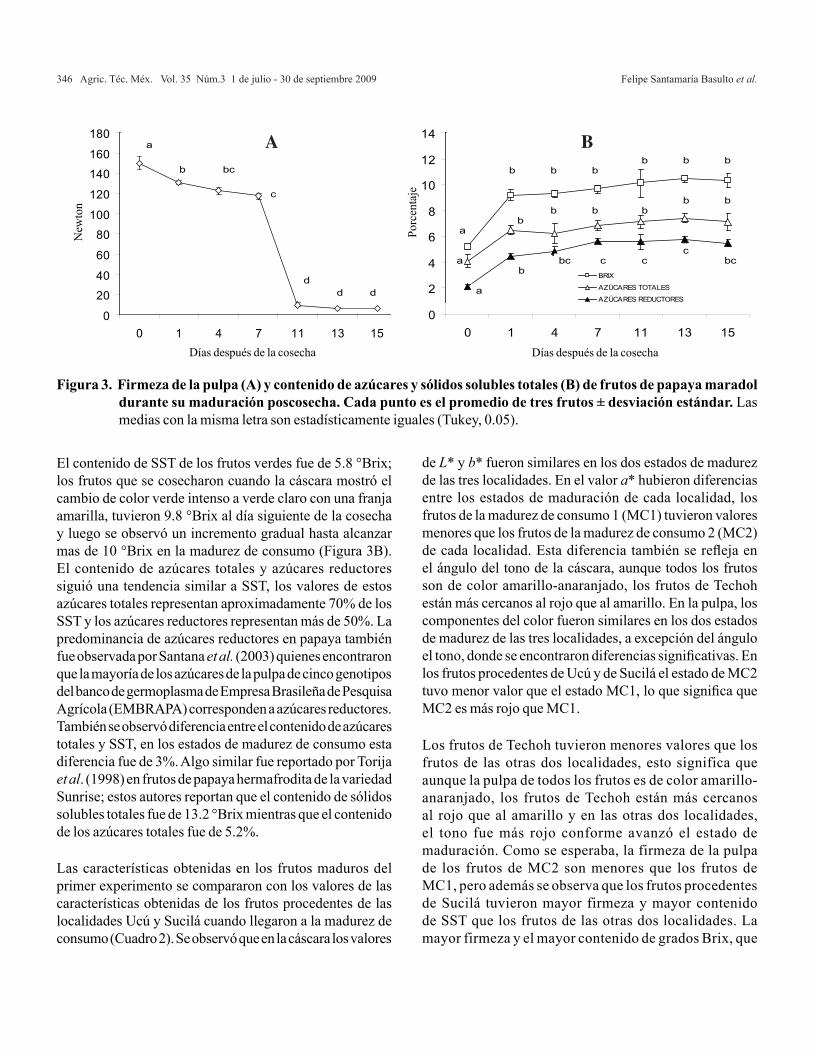
Felipe Santamaría Basulto et al.346 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
de L* y b* fueron similares en los dos estados de madurez de las tres localidades. En el valor a* hubieron diferencias entre los estados de maduración de cada localidad, los frutos de la madurez de consumo 1 (MC1) tuvieron valores menores que los frutos de la madurez de consumo 2 (MC2) de cada localidad. Esta diferencia también se refleja en el ángulo del tono de la cáscara, aunque todos los frutos son de color amarillo-anaranjado, los frutos de Techoh están más cercanos al rojo que al amarillo. En la pulpa, los componentes del color fueron similares en los dos estados de madurez de las tres localidades, a excepción del ángulo el tono, donde se encontraron diferencias significativas. En los frutos procedentes de Ucú y de Sucilá el estado de MC2 tuvo menor valor que el estado MC1, lo que significa que MC2 es más rojo que MC1.
Los frutos de Techoh tuvieron menores valores que los frutos de las otras dos localidades, esto significa que aunque la pulpa de todos los frutos es de color amarillo-anaranjado, los frutos de Techoh están más cercanos al rojo que al amarillo y en las otras dos localidades, el tono fue más rojo conforme avanzó el estado de maduración. Como se esperaba, la firmeza de la pulpa de los frutos de MC2 son menores que los frutos de MC1, pero además se observa que los frutos procedentes de Sucilá tuvieron mayor firmeza y mayor contenido de SST que los frutos de las otras dos localidades. La mayor firmeza y el mayor contenido de grados Brix, que
0
20
40
60
80
100
120
140
160
180
0 1 4 7 11 13 15Días después de la cosecha
New
tons
a
b bc
c
dd d
0
2
4
6
8
10
12
14
0 1 4 7 11 13 15Días después de la cosecha
Porc
enta
je
BRIX
AZÚCARES TOTALES
AZÚCARES REDUCTORES
a
b bb b b
b
a
bb b
b bb
a
bbc c
cbcc
A B
Figura 3. Firmeza de la pulpa (A) y contenido de azúcares y sólidos solubles totales (B) de frutos de papaya maradol durante su maduración poscosecha. Cada punto es el promedio de tres frutos ± desviación estándar. Las medias con la misma letra son estadísticamente iguales (Tukey, 0.05).
Días después de la cosecha
New
ton
Días después de la cosecha
Porc
enta
je
A B
El contenido de SST de los frutos verdes fue de 5.8 °Brix; los frutos que se cosecharon cuando la cáscara mostró el cambio de color verde intenso a verde claro con una franja amarilla, tuvieron 9.8 °Brix al día siguiente de la cosecha y luego se observó un incremento gradual hasta alcanzar mas de 10 °Brix en la madurez de consumo (Figura 3B). El contenido de azúcares totales y azúcares reductores siguió una tendencia similar a SST, los valores de estos azúcares totales representan aproximadamente 70% de los SST y los azúcares reductores representan más de 50%. La predominancia de azúcares reductores en papaya también fue observada por Santana et al. (2003) quienes encontraron que la mayoría de los azúcares de la pulpa de cinco genotipos del banco de germoplasma de Empresa Brasileña de Pesquisa Agrícola (EMBRAPA) corresponden a azúcares reductores. También se observó diferencia entre el contenido de azúcares totales y SST, en los estados de madurez de consumo esta diferencia fue de 3%. Algo similar fue reportado por Torija et al. (1998) en frutos de papaya hermafrodita de la variedad Sunrise; estos autores reportan que el contenido de sólidos solubles totales fue de 13.2 °Brix mientras que el contenido de los azúcares totales fue de 5.2%.
Las características obtenidas en los frutos maduros del primer experimento se compararon con los valores de las características obtenidas de los frutos procedentes de las localidades Ucú y Sucilá cuando llegaron a la madurez de consumo (Cuadro 2). Se observó que en la cáscara los valores

Características de calidad de frutos de papaya maradol en la madurez de consumo 347
se observó en los frutos de Sucilá probablemente sea el reflejo de las mejores condiciones de manejo agronómico, ya que la parcela de Sucilá corresponde a una muestra de
Aked, J. 2000. Fruits and vegetables. In: Kilcast, D. y Subramaniam, P. (eds.) The stability and shelf-life of food. CRC Press. Woudhead Publising Limited. p. 249-278.
Alonso, E. M.; Tornet, Q. Y.; Ramos, R. R.; Farrés, A. E.; Aranguren, G. M. y Rodríguez, M. D. 2008. Caracterización y evaluación de dos híbridos de papaya en Cuba. Agric. Téc. Méx. 34:333-339.
Bron, I. U. y Jacomino, A. P. 2006. Ripening and quality of ‘Golden’ papaya fruit harvested at different maturity stages. Braz. J. Plant Physiol. 18:389-396.
De Morais, P. L. D.; Da Silva, G. G.; Menezes, J. B.; Nogueira, M. F. E.; Dantas, D. J. y Sales, R. 2007. Pós-colheita de mamão híbrido UENF/CALIMAN 01 cultivado no rio grande do norte. Rev. Bras. Frutic., Jaboticabal 29:666-670.
De Oliveira, M. A. B.; Vianni, R.; De Souza, G. y Araújo, T. M. R. 2002. Caracterização do estádio de maturação do papaia ‘golden’ em função da cor. Rev. Bras. Frutic., Jaboticabal 24:559-561.
Techoh Ucú SuciláM C 1 M C 2 M C 1 M C 2 M C 1 M C 2
Color de cáscara
L* NS 57.21 58.02 58.47 56.09 56.06 55.39a* 10.25 ab 17.50 a 8.78 b 13.57 ab 6.52 b 14.52 abb* NS 45.77 47.76 51.93 49.16 49.48 49.87Tono (°) 77.38 ab 70.04 b 80.20 a 74.35 ab 82.48 a 73.43 ab
Color de pulpa
L* NS 50.20 51.60 52.65 49.27 49.50 47.95
a* NS 23.91 23.73 20.05 21.35 19.50 22.01
b* 35.60 37.65 40.39 37.28 39.30 38.83
Tono (°) 56.09 b 57.78 b 63.68 a 60.22 ab 63.57 a 60.45 ab
SST °Brix 10.47 b 10.35 b 10.65 b 11.0 ab 12.63 a 12.45 a
FIRMEZA N 6.74 ab 5.91 ab 6.94 ab 4.69 b 7.75 a 5.79 ab
la producción destinada para el mercado de exportación mientras que las parcelas de Techoh y Ucú son unidades de producción que destinan su producto al mercado local.
Cuadro 2. Características de color, firmeza y contenido de sólidos solubles totales de frutos en dos estados de madurez de consumo (MC1 y MC2) procedentes de tres localidades.
En cada variable, las medias con la misma letra son estadísticamente iguales (Tukey, 0.05).
Los resultados de esta investigacion permiten concluir que la madurez de consumo de papaya maradol se alcanza entre los 13 y 15 días después de la cosecha en condiciones de almacenamiento de 23 ± 1 °C y 75 % de humedad relativa. El ángulo del tono de la cáscara entre 70 y 80°, el contenido de SST entre 10 y 11.5 °Brix, y la firmeza de la pulpa de 4.7 a 6.9 N permitieron diferenciar dos estados de madurez de consumo. Estas características pueden ser propuestas como índices objetivos para describir la madurez de consumo de papaya maradol.
LITERATURA CITADA
Acosta, R. M.; Nieto-Ángel, D.; Domínguez-Álvarez, J. L. y Delgadillo-Sánchez, F. 2001. Calidad y tolerancia en frutos de papaya (Carica papaya L.) a la inoculación del hongo Colletotrichum gloeosporioides Penz., en postcosecha. Revista Chapingo Serie Horticultura.7:119-130.

Felipe Santamaría Basulto et al.348 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Dubois, M.; Gilles, K. A.; Hamilton, J. K.; Rebers, P. A. and Smith, F. 1956. Colorimetric method for determination of sugars and related substances. Anal. Chem. 28:350-356.
Ferrer, A.; Remón, S.; Negueruela, A. and Oria, R. 2005. Changes during ripening of the very late season spanish pech cultivar calanda feasibility of using CIELAB coordinates as maturity indices. Scientia Horticulturae 105:435-446.
Gobierno del estado de Yucatán. 1997. Experiencias obtenidas en el cultivo de papaya maradol en el estado. Secretaría de Desarrollo Rural. Editorial DAHEMONT, Mérida, Yucatán, 115 p.
Hernández, Y.; Lobo, M. G. y González, M. 2007. Optimización del tipo de troceado de papaya mínimamente procesada y su efecto en la translucidez. In: grupo de postrecolección y refrigeración. UPCT (ed.) V Congreso Iberoamericano de Tecnología Poscosecha y Agroexportaciones. p. 751-759.
Kader, A. A. 2004. Recommendations for maintaining postharvest quality. Papaya produce facts. postharvest technology research information center, department of pomology University of California. Davis. http://rics.ucdavis.edu/postharvest2/Produce/ProduceFacts/Fruit/papaya.sht.
McGuire, R. G. 1992. Reporting of objective color measurements. Hortscience. 27:254:1255.
Miller, G. L. 1959. Use of dinitrosalicylic acid reagent for determination of reducing sugars. Anal. Chem. 31:426-428.
NMX-FF-041-SCFI-2007. 2007. Productos alimenticios no industrializados para consumo humano fruta fresca - papaya (Carica papaya L.) - Especificaciones (Cancela a la NMX-FF-041-SCFI-2003).
Peleg, M. y Gómez-Brito, L. 1975. The red component of the external color as a maturity index of papaya fruits. Journal of Food Science 40:1105-1106.
Pérez-Carrillo, E. y Yahia, E. M. 2004. Effect of postharvest hot air and fungicide treatments on the quality of ‘maradol’ papaya (Carica papaya L.) J. Food Qual. 27:127-139.
Rocha, R. H. C.; De Carvalho, N. S. R.; Menezes, J. B.; De Souza, N. G. H. y De Oliveira, S. E. 2005. Qualidade pós-colheita do mamão formosa armazenado sob refrigeração. Rev. Bras. Frutic., Jaboticabal 27:386-389.
Santana, R. R. L.; Matsuura, F. C. A. y Cardoso, U. R. L. 2003. Genótipos melhorados de mamao (Carica papaya L.): avaliacao tecnológica dos frutos na forma de sorvete. Cien. Tecnol. Aliment., Campinas. 23:151-155.
Torija, E.; Díez, C.; Matallana, C.; Camara, M.; Camacho, E. and Mazarío, P. 1998. Influence of freezing process on free sugars content of papaya and banana fruits. J. Sci. Food Agric. 7:315-319.
Wellburn, A. R. 1994. The spectra determination of chlorophyll and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolutions. J. Plant Physiol. 144:302-313.
Zhou, L.; Paull, R. E. and Chen, N. J. 2004. Papaya. In: Gross. K. C.; Wang, Ch. Y. and Saltveit, M. The commercial storage of fruits, vegetables, and florist and nursery stocks agricultural. Agriculture Handbook. Number 66. USDA-ARS.

Agricultura Técnica en México Vol. 35 Núm.3 1 de julio-30 de septiembre 2009 p. 349-352
NEGRO GRIJALVA, NUEVO CULTIVAR DE FRIJOL PARA EL TRÓPICO HÚMEDO DE MÉXICO*
NEGRO GRIJALVA, A NEW DRY BEAN CULTIVAR FOR THE TROPICAL AREAS OF MEXICOBernardo Villar Sánchez1§, Ernesto López Salinas2 y Oscar Hugo Tosquy Valle2
1Programa de Frijol, Campo Experimental Centro de Chiapas. INIFAP. Km 3.0 carretera Ocozocuautla-Cintalapa. C. P. 29140, Ocozocuautla, Chiapas, México. Tel. 01 968 6 88 29 11. 2Campo Experimental Cotaxtla. INIFAP. ([email protected]), ([email protected]). Tel. 01 229 9 34 83 54. §Autor para correspondencia: [email protected].
ABSTRACT
In the tropical areas of southeastern Mexico the small-opaque black bean class is the most demanded by growers and consumers. In 2003 the line DOR 445 was released as a new cultivar named ‘Negro Grijalva’. This cultivar is of early maturity, resistant to BGYMV and adapted to acid soils. From 1997-2000 Negro Grijalva was compared to cv. Negro INIFAP across fifteen year-site combinations in the state of Chiapas under residual moisture conditions; the yield of Negro Grijalva was 1 219 kg ha-1, 35.9% higher than that of Negro INIFAP. In a soil of pH 4.3 in central Chiapas, the yield of Negro Grijalva was similar to Negro INIFAP and with the addition of lime at a rate of 1.5 t ha-1, its yield increased by 20.8% vs 6.0% in Negro INIFAP. Therefore, Negro Grijalva can be used in acid soils with or without the addition of lime by low income farmers. Negro Grijalva has a seed weight of 23 g per 100 seeds.
En el trópico húmedo del sureste de México, el frijol de grano negro, opaco y pequeño es la clase comercial de mayor demanda (SIAP, 2006). En esta región durante 2007 se cosecharon 175 592 ha de frijol negro, de las que se obtuvieron 114 114 t de grano, que correspondió al 11.8% de la producción nacional. El estado de Chiapas contribuyó con 56% de la producción en esa región; sin embargo, el rendimiento promedio apenas superó los 600 kg ha-1 en una superficie de 130 038 ha (SAGARPA, 2008). El bajo rendimiento obtenido se debe a varios factores que limitan
la productividad del frijol, principalmente la incidencia y severidad de enfermedades y la baja fertilidad del suelo (Villar et al., 2003). Entre las enfermedades, destaca el virus del mosaico amarillo dorado del frijol (VMADF), que ha llegado a causar la pérdida total de la cosecha y ha reducido la superficie de siembra (Villar y López, 2003) y la acidez de los suelos que causa fitotoxicidad por el alto contenido de aluminio intercambiable en las regiones de la Frailesca y la costa de Chiapas (Buerkert et al., 1990; Villar et al., 2002).
Los resultados de investigación y validación realizados por el programa de frijol del Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias (INIFAP) en varios años y localidades en el estado de Chiapas, indican que es factible obtener rendimientos mayores a 1 t ha-1, con la aplicación de tecnología moderna de bajos insumos, como la siembra de variedades mejoradas que cuentan con resistencia a los principales factores limitantes (Villar y López, 2003).
En 2003, se liberó la línea DOR 445, como nueva variedad con el nombre de ‘Negro Grijalva’ para las áreas tropicales de Chiapas y regiones similares. Esta variedad cuenta con características de precocidad, resistencia al virus VMADF y tolerancia a la acidez del suelo, características que pueden contribuir a mejorar la productividad del frijol en la entidad.
* Recibido: Febrero, 2009
Aceptado: Septiembre, 2009

Bernardo Villar Sánchez et al.350 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
Negro Grijalva se originó de la cruza doble (DOR-364 x G 18521) // (DOR-365 x IN 100), realizada en el Centro Internacional de Agricultura Tropical (CIAT), Calí, Colombia. Los progenitores DOR se utilizaron como fuente de resistencia al VMADF, ambos son de amplia adaptación en las áreas tropicales de Centroamérica, mientras que las líneas G 18521 e IN 100 se incluyeron por su resistencia a roya (López et al., 2007). El proceso de desarrollo de Negro Grijalva incluyó selección individual en F2, compuesto masal, selección individual en F4 y selección masal en F5 y en F6 realizadas en el Instituto de Ciencia y Tecnología Agrícola (ICTA) de Guatemala, dentro del programa cooperativo regional de frijol para Centroamérica, México y el Caribe (PROFRIJOL). En 1993 se introdujo a México la línea DOR 445 a través del proyecto de mejoramiento de frijol negro mesoamericano, para evaluarla en el estado de Chiapas en ensayos regionales y uniformes de rendimiento de 1997 a 2000, en los que mostró alto rendimiento y tolerancia al virus del mosaico amarillo dorado del frijol y adaptación a los suelos ácidos con alta saturación de aluminio. La validación se realizó de 2001 a 2003 en varias localidades del estado de Chiapas, bajo condiciones de temporal, humedad residual y riego.
Las principales características agronómicas de Negro Grijalva son: hábito de crecimiento indeterminado y erecto, planta arbustiva tipo II, con guías cortas en siembras de temporal y de riego y largas cuando se siembra en el ciclo otoño-invierno bajo condiciones de humedad residual. La altura de planta promedio es de 42 cm, es precoz, ya que inicia floración a los 30 días después de la siembra y madura a los 70 días. La flor es morada y vaina es de color amarillo en la etapa de madurez fisiológica y blanca en la cosecha, con granos negros, opacos y pequeños (23g/100 semillas).
Durante 1997-2000, Negro Grijalva se comparó con el testigo comercial Negro INIFAP en ensayos de rendimiento establecidos en quince localidades en Chiapas, México, bajo condiciones de humedad residual, temporal y riego. En la primera condición, el rendimiento promedio de Negro Grijalva fue de 1 219 kg ha-1, superior en 35.9% al promedio de rendimiento obtenido por Negro INIFAP. Bajo condiciones de temporal, el rendimiento de la nueva variedad fue de 1 284 kg ha-1, superior en 17.8% al obtenido por el testigo regional, mientras que en riego, el rendimiento de Negro Grijalva (2 694 kg ha-1) fue 9.7% mayor que el Negro INIFAP. El promedio general de rendimiento de la nueva variedad fue superior en 20% al testigo.
En 2000, en la localidad de Ocozocoautla, Chiapas, bajo condiciones de temporal, se evaluó la reacción de la variedad a la incidencia natural del VMADF. Negro Grijalva mostró menor incidencia de la enfermedad (2.5%), se calificó como resistente y obtuvo el mayor rendimiento de grano (1 330 kg ha-1), mientras que el testigo Negro INIFAP presentó el mayor daño (18%) y un rendimiento de 1 120 kg ha-1 (Villar et al., 2003). En la misma localidad, en una parcela de validación establecida durante el ciclo otoño-invierno 2001-2002, se corroboró la tolerancia de Negro Grijalva al VMADF, mientras que el criollo regional Sesentano fue susceptible (López et al., 2007).
La evaluación de la tolerancia a suelos ácidos se realizó en los ciclos de temporal de 1999 y 2000, en el municipio de Villaflores, ubicado en el centro del estado de Chiapas. La nueva variedad mostró rendimiento de grano similar al de Negro INIFAP, bajo condiciones naturales de acidez (pH de 4.3) y con más de 10% de saturación de aluminio. La tolerancia a la acidez del suelo de ambos genotipos, puede estar ligada a mecanismos de la planta para tolerar altas concentraciones de aluminio en el suelo (Villar, 2000), los cuales se dividen en externos, en donde este elemento es excluido del simplasto de los meristemos radicales, y en internos, en los que el aluminio que ha penetrado al plasmalema se acompleja internamente (Caver y Ownby, 1995). Negro Grijalva mostró mayor incremento en rendimiento (20.8%) que el testigo comercial (6.2%) al aplicar cal al suelo; lo anterior, debido a que se neutraliza el aluminio (disminuye el porcentaje de saturación de este elemento) y se incrementa el pH del suelo, lo que permite mayor disponibilidad de nutrimentos. Estos resultados indican que la variedad Negro Grijalva puede ser utilizada por agricultores de bajos recursos y productores empresariales que aplican la práctica de encalado (Villar et al., 2003).
De 2001 a 2003 la variedad Negro Grijalva se validó en 10 parcelas comerciales establecidas en terrenos de agricultores en el estado de Chiapas y fue comparada con la variedad Negro INIFAP. En las tres condiciones de humedad (temporal, humedad residual y riego), la nueva variedad obtuvo mayor rendimiento que el testigo comercial. El rendimiento promedio general de Negro Grijalva fue de 1 273 kg ha-1, superior en 17.1% al promedio obtenido por la variedad testigo (Cuadro 1). Los resultados de experimentación y validación, confirman el alto potencial de rendimiento que tiene esta variedad bajo los diferentes sistemas en que se produce frijol en el estado de Chiapas.
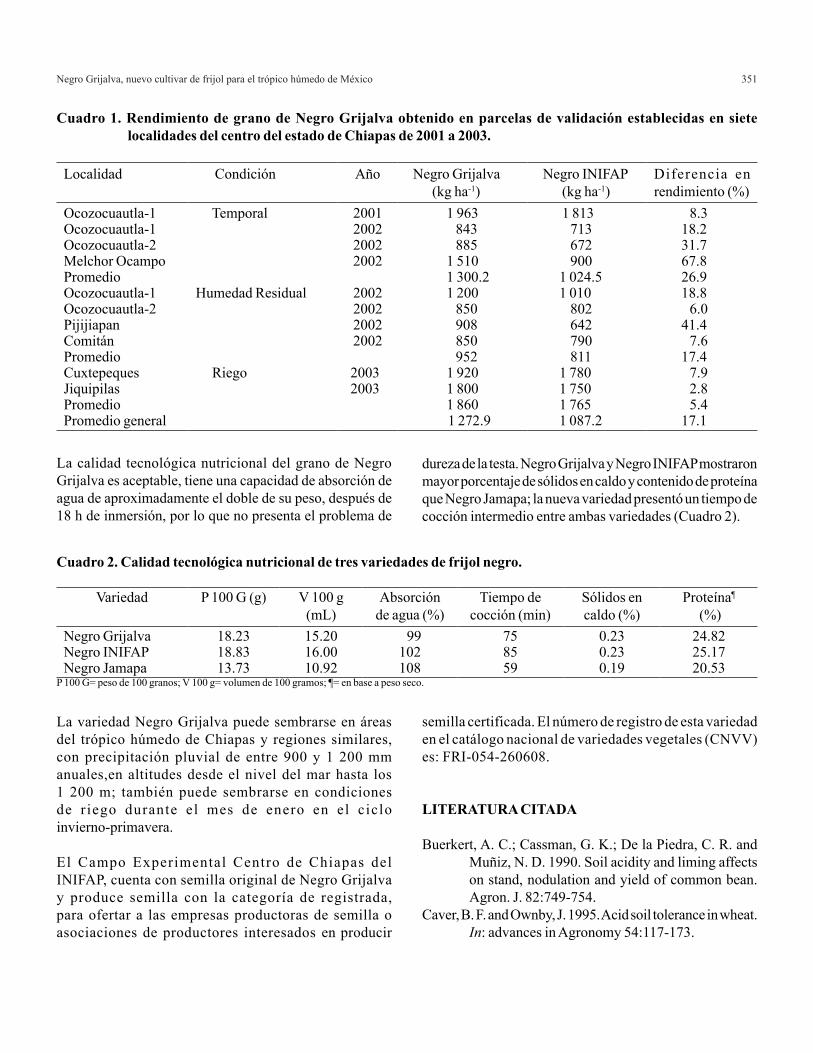
Negro Grijalva, nuevo cultivar de frijol para el trópico húmedo de México 351
dureza de la testa. Negro Grijalva y Negro INIFAP mostraron mayor porcentaje de sólidos en caldo y contenido de proteína que Negro Jamapa; la nueva variedad presentó un tiempo de cocción intermedio entre ambas variedades (Cuadro 2).
semilla certificada. El número de registro de esta variedad en el catálogo nacional de variedades vegetales (CNVV) es: FRI-054-260608.
LITERATURA CITADA
Buerkert, A. C.; Cassman, G. K.; De la Piedra, C. R. and Muñiz, N. D. 1990. Soil acidity and liming affects on stand, nodulation and yield of common bean. Agron. J. 82:749-754.
Caver, B. F. and Ownby, J. 1995. Acid soil tolerance in wheat. In: advances in Agronomy 54:117-173.
Localidad Condición Año Negro Grijalva (kg ha-1)
Negro INIFAP (kg ha-1)
Diferencia en rendimiento (%)
Ocozocuautla-1 Temporal 2001 1 963 1 813 8.3Ocozocuautla-1 2002 843 713 18.2Ocozocuautla-2 2002 885 672 31.7Melchor Ocampo 2002 1 510 900 67.8Promedio 1 300.2 1 024.5 26.9Ocozocuautla-1 Humedad Residual 2002 1 200 1 010 18.8Ocozocuautla-2 2002 850 802 6.0Pijijiapan 2002 908 642 41.4Comitán 2002 850 790 7.6Promedio 952 811 17.4Cuxtepeques Riego 2003 1 920 1 780 7.9Jiquipilas 2003 1 800 1 750 2.8Promedio 1 860 1 765 5.4Promedio general 1 272.9 1 087.2 17.1
Variedad P 100 G (g) V 100 g(mL)
Absorciónde agua (%)
Tiempo de cocción (min)
Sólidos en caldo (%)
Proteína¶
(%)Negro Grijalva 18.23 15.20 99 75 0.23 24.82Negro INIFAP 18.83 16.00 102 85 0.23 25.17Negro Jamapa 13.73 10.92 108 59 0.19 20.53
Cuadro 1. Rendimiento de grano de Negro Grijalva obtenido en parcelas de validación establecidas en siete localidades del centro del estado de Chiapas de 2001 a 2003.
La calidad tecnológica nutricional del grano de Negro Grijalva es aceptable, tiene una capacidad de absorción de agua de aproximadamente el doble de su peso, después de 18 h de inmersión, por lo que no presenta el problema de
Cuadro 2. Calidad tecnológica nutricional de tres variedades de frijol negro.
P 100 G= peso de 100 granos; V 100 g= volumen de 100 gramos; ¶= en base a peso seco.
La variedad Negro Grijalva puede sembrarse en áreas del trópico húmedo de Chiapas y regiones similares, con precipitación pluvial de entre 900 y 1 200 mm anuales,en altitudes desde el nivel del mar hasta los 1 200 m; también puede sembrarse en condiciones de r iego durante el mes de enero en el cicloinvierno-primavera.
El Campo Experimental Centro de Chiapas del INIFAP, cuenta con semilla original de Negro Grijalva y produce semilla con la categoría de registrada, para ofertar a las empresas productoras de semilla o asociaciones de productores interesados en producir

Bernardo Villar Sánchez et al.352 Agric. Téc. Méx. Vol. 35 Núm.3 1 de julio - 30 de septiembre 2009
López, S. E.; Villar, S. B.; Tosquy, V. O. H.; Ugalde, A. F. J.; Becerra, L. E. N. y Cumpeán, G. J. 2007. Negro Papaloapan, nueva variedad de frijol para las áreas tropicales de Veracruz y Chiapas. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional Golfo Centro. Campo Experimental Cotaxtla. Veracruz, México. 21 p. (Folleto Técnico Núm. 41).
López, S. E.; Tosquy, V. O. H.; Villar, S. B.; Cumpián, G. J.; Ugalde, A., F. J. y Becerra, L. E. N. 2007. Negro Papaloapan, nuevo cultivar de frijol para las áreas tropicales de México. Agric. Téc. Méx. 33(3):257-267.
Villar, S. B. 2000. Estrategia para el manejo de suelos ácidos en frijol (Phaseolus vulgaris L.) en el estado de Chiapas, México. Agron. Mesoam. 11(1):155-158.
Villar, S. B.; Garrido, R. E.; López, L. A. y Cruz, Ch. F. 2002. Manual para la producción de frijol en el estado de Chiapas. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional del Pacífico Sur. Campo Experimental Centro de Chiapas. Ocozocuautla, Chiapas, México. 154 p. (Publicación Especial Núm. 1).
Villar, S. B. y López, S. E. 2003. Negro Grijalva, nueva variedad de frijol para Chiapas y regiones similares. Instituto Nacional de Investigaciones Forestales, Agrícolas y Pecuarias. Centro de Investigación Regional del Pacífico Sur. Campo Experimental Centro de Chiapas. Ocozocuautla de Espinosa, Chiapas, México. 22 p. (Folleto Técnico Núm. 2).
Villar, S. B.; López, S. E. y Acosta, G. J. 2003. Selección de genotipos de frijol por rendimiento y resistencia al mosaico dorado y suelos ácidos. Rev. Fitotec. Méx. 26(2):109-114.
Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). 2008. Anuarios estadísticos de la producción agrícola. Servicio de Información Agroalimentaria y Pesquera (SIAP). México, D. F. http://www.siap.sagarpa.gob.mx (consultado el 7 de noviembre de 2008).
Servicio de Información y Estadística Agroalimentaria y Pesquera (SIAP). 2006. Situación actual y perspectivas de la producción de frijol en México 2000-2005. Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA). México. 34 p.





























