Embed Size (px)
Citation preview

Mechanism of platelet functional changes and effects of anti-platelet agentson in vivo hemostasis under different gravity conditions
Suping Li,1,2* Quanwei Shi,1* Guanglei Liu,1 Weilin Zhang,1 Zhicheng Wang,1 Yuedan Wang,3
and Kesheng Dai11School of Biological Science and Medical Engineering, Beihang University, Beijing; 2State Key Laboratory of SpaceMedicine Fundamentals and Application, China Astronaut Research and Training Center, Beijing; and 3Departmentof Immunology, Peking University, Beijing, China
Submitted 26 October 2009; accepted in final form 2 February 2010
Li S, Shi Q, Liu G, Zhang W, Wang Z, Wang Y, Dai K.Mechanism of platelet functional changes and effects of anti-plateletagents on in vivo hemostasis under different gravity conditions. J ApplPhysiol 108: 000–000, 2010. First published February 4, 2010;doi:10.1152/japplphysiol.01209.2009.—Serious thrombotic and hem-orrhagic problems or even fatalities evoked by either microgravity orhypergravity occur commonly in the world. We recently reported thatplatelet functions are inhibited in microgravity environments andactivated under high-G conditions, which reveals the pathogenesis forgravity change-related hemorrhagic and thrombotic diseases. How-ever, the mechanisms of platelet functional variations under differentgravity conditions remain unclear. In this study we show that theamount of filamin A coimmunoprecipitated with GPIb� was enhancedin platelets exposed to modeled microgravity and, in contrast, wasreduced in 8 G-exposed platelets. Hypergravity induced actin filamentformation and redistribution, whereas actin filaments were reduced inplatelets treated with modeled microgravity. Furthermore, intracellu-lar Ca2� levels were elevated by hypergravity. Pretreatment of plate-lets with the cell-permeable Ca2� chelator BAPTA-AM had no effecton cytoskeleton reorganization induced by hypergravity but signifi-cantly reduced platelet aggregation induced by ristocetin/hypergrav-ity. Two anti-platelet agents, aspirin and tirofiban, effectively reversedthe shortened tail bleeding time and reduced the death rate of miceexposed to hypergravity. Furthermore, the increased P-selectin sur-face expression was obviously reduced in platelets from mice treatedwith aspirin/hypergravity compared with those from mice treated withhypergravity alone. These data suggest that the actin cytoskeletonreorganization and intracellular Ca2� level play key roles in theregulation of platelet functions in different gravitational environ-ments. The results with anti-platelet agents not only further confirmthe activation of platelets in vivo but also suggest a therapeuticpotential for hypergravity-induced thrombotic diseases.
platelets; cytoskeleton; calcium; modeled microgravity; hypergravity
GRAVITATIONAL CHANGES induce pathophysiological responses invarious cell types such as vascular endothelial cells (18),neuroblastoma cells (30), and bone cells (34). Many throm-botic and hemorrhagic diseases have been documented inhuman beings exposed to microgravity or hypergravity envi-ronments (27, 32), and several possibly related organs havebeen examined to explore the mechanisms underlying thesegravity change-related diseases (28, 39). We reported recentlythat platelet functions are inhibited in microgravity environ-ments and activated under high-G conditions, which revealsthe pathogenesis of gravity change-related hemorrhagic and
thrombotic diseases (8, 11, 19). However, the mechanisms ofthe platelet functional variations under different gravity con-ditions remain unclear.
The actin cytoskeleton of cells is a highly dynamic structureand is sensitive to changes of gravity conditions (30, 38).Several studies have shown that simulated hypogravity stimu-lates endothelial cell growth and also remodels the actincytoskeleton and reduces the total amount of actin (1, 38). Incontrast, hypergravity alters the distribution of actin fiberswithout affecting the total amount of actin (38). In humanmesenchymal stem cells, F-actin stress fibers are disruptedwithin 3 h of initiation of modeled microgravity and arecompletely absent by 7 days, whereas monomeric G-actin isincreased (22). Furthermore, studies in Biorack on space shut-tle missions STS-76, STS-81, and STS-84 confirmed the col-lapse of the actin cytoskeleton in cells grown in microgravityconditions. However, flown cells under 1-G conditions main-tained normal actin cytoskeleton and fibronectin matrix (14). Inaddition, experiments on single-cell behavior in hypergravityhave shown that hypergravity stimulation results in reorgani-zation of actin fibers with an increased formation of dense actinfibers in cytosol (18). We have reported that glycoprotein (GP)Ib� surface expression and association with the cytoskeletonsignificantly decreased in platelets exposed to simulated mi-crogravity and obviously increased in hypergravity-exposedplatelets (8, 11), indicating that the reorganization of cytoskel-eton may be involved in the regulation of platelet functionalchanges under different gravity conditions.
Changes in the intracellular Ca2� concentration ([Ca2�]i) inresponse to altered gravity have been reported to occur widelyin many types of cells and to associate with cytoskeletalreorganization (4, 36). [Ca2�]i increases in response to gravitychange in hypocotyls and petioles of Arabidopsis seedlings,which depends on both Ca2� influx and Ca2� release fromintracellular Ca2� stores (35). The elevation of [Ca2�]i issufficient for promoting cytoskeletal reorganization (25); onthe other hand, dynamic remodeling of the actin cytoskeletonalso modulates intracellular Ca2� release (4, 35). The intracel-lular Ca2� concentration plays key roles in initiating andregulating platelet adhesion, aggregation, and secretion (21),thus raising the concern whether platelet functional changesunder different gravitational conditions result from variationsof [Ca2�]i.
In this study, we show that the association of GPIb� withfilamin A and the organization of actin cytoskeleton are regu-lated by different gravities. Intracellular Ca2� levels wereelevated by hypergravity, which upregulated ristocetin-inducedplatelet aggregation. Furthermore, two anti-platelet agents, aspirin
* S. Li and Q. Shi contributed equally to this work.Address for reprint requests and other correspondence: K. Dai, School of
Biological Science and Medical Engineering, Beihang Univ., 37 XueyuanRoad, Haidian district, Beijing 100083, China (e-mail: [email protected]).
J Appl Physiol 108: 000–000, 2010.First published February 4, 2010; doi:10.1152/japplphysiol.01209.2009.
8750-7587/10 $8.00 Copyright © 2010 the American Physiological Societyhttp://www.jap.org 1
AQ: 1
AQ: 2
AQ: 5
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss
DOCHEAD � ; DOCTOPIC �

and tirofiban, effectively reversed platelet activation induced byhypergravity in vivo and reduced the death rate of mice exposedto hypergravity.
MATERIALS AND METHODS
Reagents. Monoclonal antibody SZ2 against GPIb� was kindlyprovided by Dr. Changgeng Ruan (Soochow University, Suzhou,China). Monoclonal antibody against filamin A was purchased fromNovocastra Laboratories (Newcastle, UK). Polyclonal goat anti-mouse P-selectin antibody sc-6941 and FITC-labeled rabbit anti-goatIgG were purchased from Santa Cruz Biotechnology (Santa Cruz,CA). Ristocetin, ADP, EDTA, and EGTA were purchased fromSigma (St. Louis, MO). 1,2-Bis(o-aminophenoxy)ethane-N,N,N=,N=-tetraacetic acid acetoxymethyl ester (BAPTA-AM) was purchasedfrom Dojindo Molecular Technologies (Rockville, MD). The rhodam-ine-conjugated phalloidin and fluo-3 AM were obtained from Invitro-gen Molecular Probes (Eugene, OR).
Rotary cell culture system. To simulate microgravity conditions,we used a National Aeronautics and Space Administration-recom-mended and widely accepted ground-based microgravity-simulatingsystem, the Rotary Cell Culture System (RCCS; Synthecon, Houston,TX) (5, 21). Platelets were either rotated about a horizontal axisperpendicular to the gravitational vector to randomize gravitationalvectors across the surface of the platelets and generate microgravity of�10�2 G or rotated about a vertical axis parallel to the gravitationalvector to experience normal gravitational forces and serve as a control(1 G) environment (5, 12).
Hypergravity stimulation. Hypergravity was applied to platelets ormice with a custom-built slow-speed 30-cm-radius centrifuge. Plate-lets or mice were placed on the plate of the centrifuge and rotated fora certain time at 253 or 311 rpm to obtain 8 or 12 G, respectively, atroom temperature (RT). The mice were anesthetized with pentobar-bital sodium before centrifugation to avoid mechanical injury. Thesynchronous 1-G controls were placed on a plateau for the same time.
Animal model. Four- to six-wk-old adult C57 black or BALB/Cmice were anesthetized with an intraperitoneal injection of pentobar-bital sodium. To study the effects of aspirin and tirofiban treatments,anesthetized mice were intraperitoneally injected with aspirin (170mg/kg) and caudally intravenously injected with tirofiban at a dose of0.4 mg/kg, respectively, and then exposed to 12-G hypergravity for 15min. Alcohol was used as a control for aspirin; 0.15 M NaCl was usedas a control for tirofiban. The synchronous 1-G control mice that werematched for weight and sex were treated in the same way and thenplaced on a plateau for 15 min. The 1-G control mice injected withaspirin or tirofiban were used to confirm the rationality of drug doseused that would not cause spontaneous bleeding in mice. All proce-dures performed in this study were approved by the Peking UniversityInstitutional Animal Care and Use Committee.
Preparation of washed platelets and platelet-rich plasma. Forstudies involving human subjects, approval was obtained from theBeihang University institutional review board. Informed consent wasprovided according to the Declaration of Helsinki. Fresh blood fromhealthy volunteers was anticoagulated with a 1:7 volume of acid-citrate dextrose (ACD: 2.5% trisodium citrate, 2.0% D-glucose, 1.5%citric acid). After centrifugation, isolated platelets were washed twicewith CGS buffer (sodium citrate, glucose, and sodium chloride) andresuspended in modified Tyrode’s buffer (2.5 mM HEPES, 150 mMNaCl, 2.5 mM KCl, 12 mM NaHCO3, 1 mM MgCl2, 1 mM CaCl2,and 5.5 mM D-glucose, pH 7.4) to a final concentration of 3 � 108
platelets/ml and then incubated at RT for 1 h to recover to resting stateas described previously (7). In platelet-rich plasma (PRP) experi-ments, fresh blood was anticoagulated with a 1:9 volume of 3.8%trisodium citrate and centrifuged at 150 g for 12 min at RT to get PRP.PRP was then incubated at RT for 1 h to recover to resting state asdescribed previously (9).
For mouse experiments, fresh blood was immediately drawn fromthe inferior caval vein of mice after 12- or 1-G exposure andanticoagulated with a 1:7 volume of ACD. An equal volume of 4%phosphate-buffered paraformaldehyde was then added to incubate for30 min at RT. After centrifugation, isolated platelets were washedtwice with CGS and then resuspended in modified Tyrode’s buffer toa concentration of 1 � 107 platelets/ml (7).
Coimmunoprecipitation and immunoblotting. Washed human platelets(3 � 108 platelets/ml) were exposed to modeled microgravity for 30min or 1 h, 8 G for 10 or 15 min, or synchronous 1-G conditions andthen lysed with buffer containing 0.1 mM E64, 1 mM phenylmeth-ylsulfonyl fluoride (PMSF), and 1:100 aprotinin on ice for 30 min.Platelet lysates were centrifuged at 15,000 g for 4 min to removecytoplasmic actin filaments. The supernatants were immunoprecipitatedwith anti-GPIb� NH2-terminal antibody SZ2 and protein G-Sepharosebeads. Samples were subjected to SDS-PAGE and immunoblotted withSZ2 and anti-filamin A antibody, respectively. Immunoreactive bandswere visualized using enhanced chemiluminescence (ECL).
Staining of platelet F-actin. Washed human platelets (3 � 108
platelets/ml) were exposed to simulated microgravity for 1 h, 8 G for15 min, or synchronous 1-G conditions and immediately fixed with4% paraformaldehyde. F-actin was stained by rhodamine-conjugatedphalloidin according to the previously reported method (18). Toevaluate the effect of the intracellular Ca2� chelator BAPTA-AM onactin distribution under hypergravity, platelets were pretreated withBAPTA-AM at 10 �M at 37°C for 20 min, centrifuged at 8 G for 15min, and then stained. As a negative control, platelets were pretreatedwith vehicle DMSO.
Measurement of intracellular Ca2� concentration. IntracellularCa2� levels were determined with the Ca2�-sensitive fluorochromefluo-3 AM using flow cytometry. Briefly, washed human platelets (3 �108 platelets/ml) were loaded with 8 �M fluo-3 AM for 30 min at37°C in the dark. After being washed once, platelets were resuspendedat a concentration of 5 � 107 platelets/ml. The external Ca2� wasadjusted to 1 mM, and then the dyed platelets were exposed tosimulated microgravity for 1 h, 8 G for 15 min, or synchronous 1 Gand analyzed using flow cytometry. In some experiments, the loadedplatelets were preincubated with 5 mM EDTA (or EGTA) at RT for10 min and then centrifuged at 8 G for 15 min and analyzed.
Measurement of platelet aggregation. Platelet aggregation wasperformed using a turbidometric platelet aggregometer (Xinpusen,Beijing, China). Briefly, PRP was exposed to 8 or 1 G for 15 min andthen immediately added into glass aggregometer cuvettes, and aggre-gation was induced by addition of the agonist ristocetin (0.6–0.8mg/ml) or ADP at 37°C under stirring (1,000 rpm) conditions. Toevaluate the effect of BAPTA-AM on platelet aggregation, PRP waspreincubated with BAPTA-AM (10 �M) at 37°C for 20 min beforehypergravity or 1-G treatment, and then aggregation was measured.Vehicle DMSO was used as a negative control.
Tail bleeding time assay. Tail bleeding time assays were performedwith 4- to 6-wk-old adult C57 black and BALB/C mice. Afterapplication of hypergravity at 12 G for 15 min, the differently treatedand 1-G control mice were immediately subjected to tail bleeding timeanalysis. The tails were cut transversely 0.5 mm from the tip andimmersed in 0.15 M NaCl maintained at 37°C. The time to arrest ofbleeding was noted. The occurrence of rebleeding from tail woundswas assessed. Maximum bleeding times allowed were 10 min after thetail was cauterized.
Platelet surface staining. Washed mouse platelets (1 � 107 plate-lets/ml) were incubated with goat anti-mouse P-selectin antibody atRT for 30 min. Platelets were washed once and further incubated with10 �g/ml FITC-labeled rabbit anti-goat IgG for 30 min at RT in thedark. Samples were analyzed using flow cytometry, and antibodybinding was expressed as geometric mean fluorescence intensity. Asnegative controls, platelets were incubated with goat IgG and thenincubated with FITC-labeled rabbit anti-goat IgG.
2 MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
AQ: 3
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss
Statistical analysis. The Western blot was scanned, and the opticaldensity of each band from the same blot was quantitated with NIHImageJ software (image processing and analysis in Java; NationalInstitutes of Health). Data are means � SD. Statistical significancebetween means was determined by using Student’s t-test with InStatsoftware. A P value 0.05 was considered significant.
RESULTS
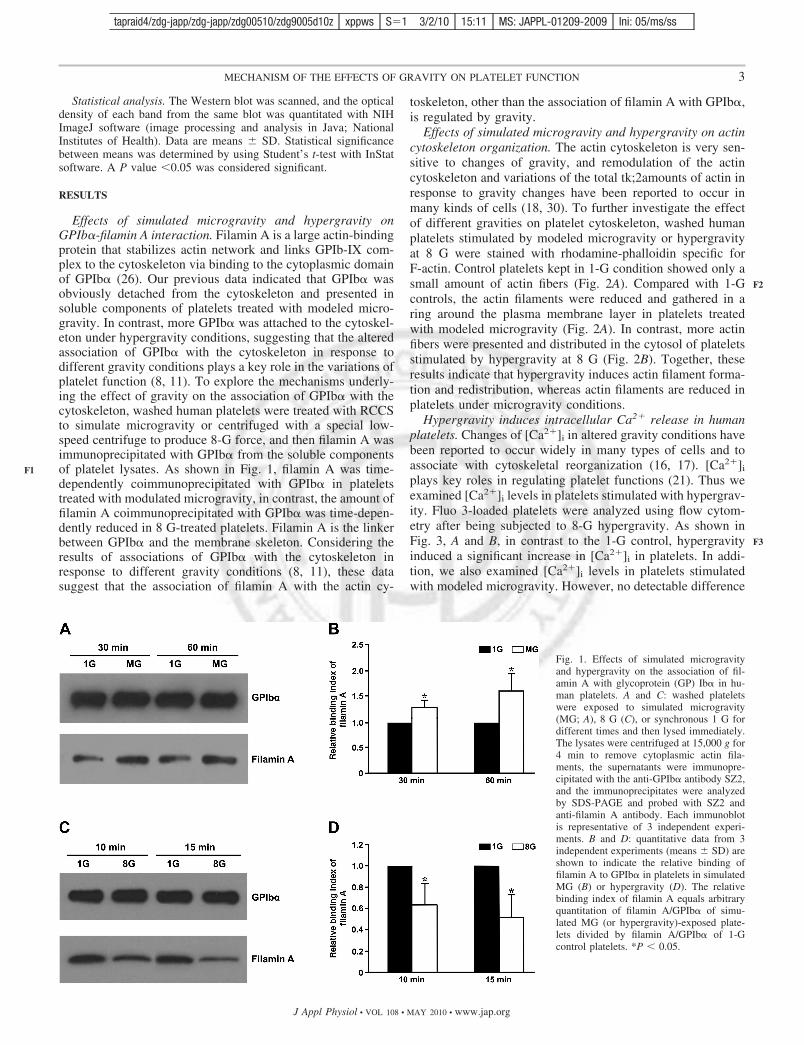
Effects of simulated microgravity and hypergravity onGPIb�-filamin A interaction. Filamin A is a large actin-bindingprotein that stabilizes actin network and links GPIb-IX com-plex to the cytoskeleton via binding to the cytoplasmic domainof GPIb� (26). Our previous data indicated that GPIb� wasobviously detached from the cytoskeleton and presented insoluble components of platelets treated with modeled micro-gravity. In contrast, more GPIb� was attached to the cytoskel-eton under hypergravity conditions, suggesting that the alteredassociation of GPIb� with the cytoskeleton in response todifferent gravity conditions plays a key role in the variations ofplatelet function (8, 11). To explore the mechanisms underly-ing the effect of gravity on the association of GPIb� with thecytoskeleton, washed human platelets were treated with RCCSto simulate microgravity or centrifuged with a special low-speed centrifuge to produce 8-G force, and then filamin A wasimmunoprecipitated with GPIb� from the soluble componentsof platelet lysates. As shown in Fig. 1, filamin A was time-dependently coimmunoprecipitated with GPIb� in plateletstreated with modulated microgravity, in contrast, the amount offilamin A coimmunoprecipitated with GPIb� was time-depen-dently reduced in 8 G-treated platelets. Filamin A is the linkerbetween GPIb� and the membrane skeleton. Considering theresults of associations of GPIb� with the cytoskeleton inresponse to different gravity conditions (8, 11), these datasuggest that the association of filamin A with the actin cy-
toskeleton, other than the association of filamin A with GPIb�,is regulated by gravity.
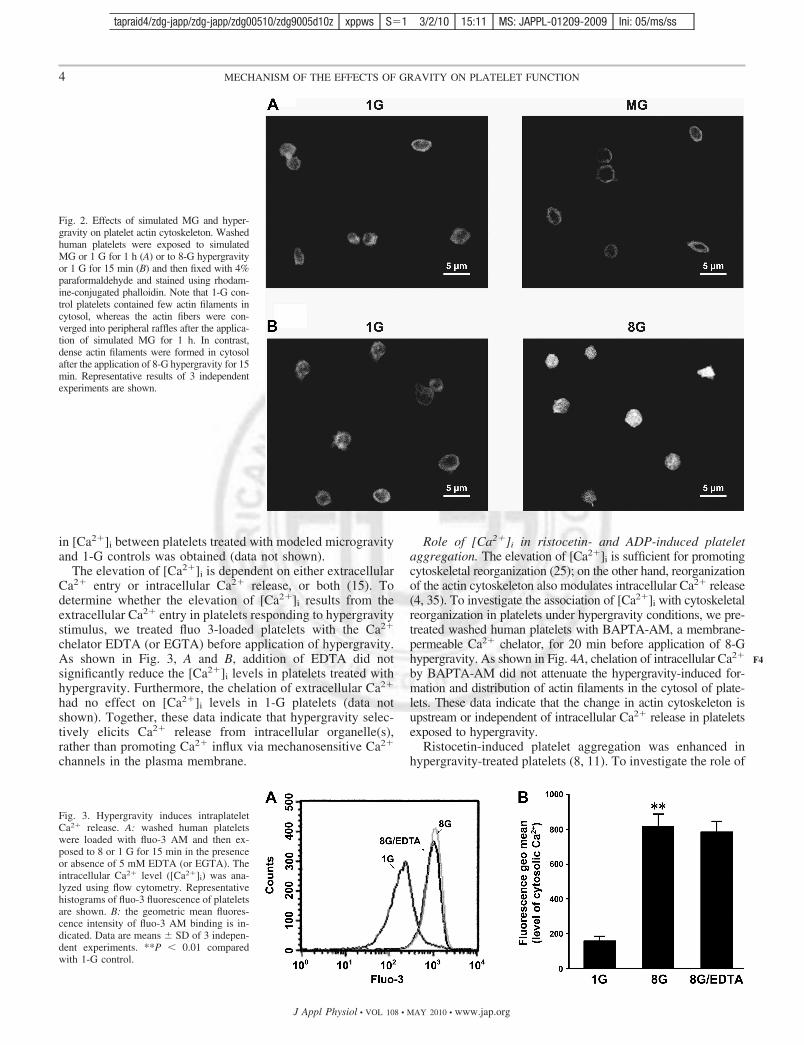
Effects of simulated microgravity and hypergravity on actincytoskeleton organization. The actin cytoskeleton is very sen-sitive to changes of gravity, and remodulation of the actincytoskeleton and variations of the total tk;2amounts of actin inresponse to gravity changes have been reported to occur inmany kinds of cells (18, 30). To further investigate the effectof different gravities on platelet cytoskeleton, washed humanplatelets stimulated by modeled microgravity or hypergravityat 8 G were stained with rhodamine-phalloidin specific forF-actin. Control platelets kept in 1-G condition showed only asmall amount of actin fibers (Fig. 2A). Compared with 1-Gcontrols, the actin filaments were reduced and gathered in aring around the plasma membrane layer in platelets treatedwith modeled microgravity (Fig. 2A). In contrast, more actinfibers were presented and distributed in the cytosol of plateletsstimulated by hypergravity at 8 G (Fig. 2B). Together, theseresults indicate that hypergravity induces actin filament forma-tion and redistribution, whereas actin filaments are reduced inplatelets under microgravity conditions.
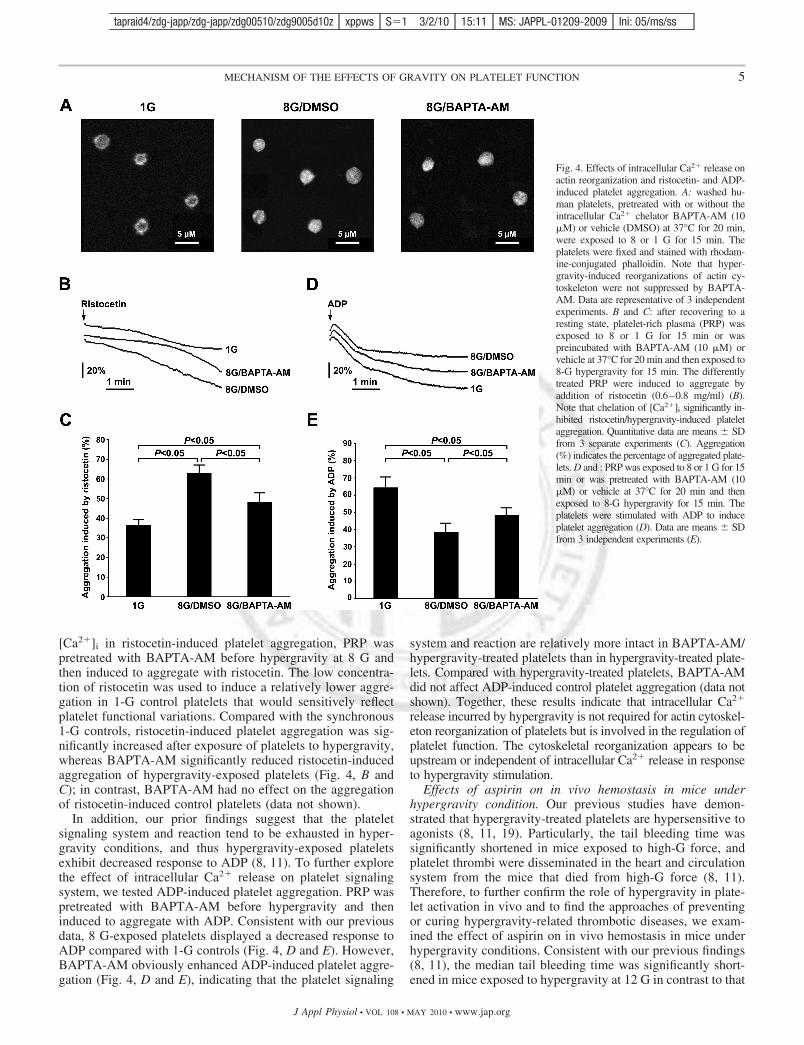
Hypergravity induces intracellular Ca2� release in humanplatelets. Changes of [Ca2�]i in altered gravity conditions havebeen reported to occur widely in many types of cells and toassociate with cytoskeletal reorganization (16, 17). [Ca2�]i
plays key roles in regulating platelet functions (21). Thus weexamined [Ca2�]i levels in platelets stimulated with hypergrav-ity. Fluo 3-loaded platelets were analyzed using flow cytom-etry after being subjected to 8-G hypergravity. As shown inFig. 3, A and B, in contrast to the 1-G control, hypergravityinduced a significant increase in [Ca2�]i in platelets. In addi-tion, we also examined [Ca2�]i levels in platelets stimulatedwith modeled microgravity. However, no detectable difference
Fig. 1. Effects of simulated microgravityand hypergravity on the association of fil-amin A with glycoprotein (GP) Ib� in hu-man platelets. A and C: washed plateletswere exposed to simulated microgravity(MG; A), 8 G (C), or synchronous 1 G fordifferent times and then lysed immediately.The lysates were centrifuged at 15,000 g for4 min to remove cytoplasmic actin fila-ments, the supernatants were immunopre-cipitated with the anti-GPIb� antibody SZ2,and the immunoprecipitates were analyzedby SDS-PAGE and probed with SZ2 andanti-filamin A antibody. Each immunoblotis representative of 3 independent experi-ments. B and D: quantitative data from 3independent experiments (means � SD) areshown to indicate the relative binding offilamin A to GPIb� in platelets in simulatedMG (B) or hypergravity (D). The relativebinding index of filamin A equals arbitraryquantitation of filamin A/GPIb� of simu-lated MG (or hypergravity)-exposed plate-lets divided by filamin A/GPIb� of 1-Gcontrol platelets. *P 0.05.
3MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
F1
F2
F3
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss
in [Ca2�]i between platelets treated with modeled microgravityand 1-G controls was obtained (data not shown).
The elevation of [Ca2�]i is dependent on either extracellularCa2� entry or intracellular Ca2� release, or both (15). Todetermine whether the elevation of [Ca2�]i results from theextracellular Ca2� entry in platelets responding to hypergravitystimulus, we treated fluo 3-loaded platelets with the Ca2�
chelator EDTA (or EGTA) before application of hypergravity.As shown in Fig. 3, A and B, addition of EDTA did notsignificantly reduce the [Ca2�]i levels in platelets treated withhypergravity. Furthermore, the chelation of extracellular Ca2�
had no effect on [Ca2�]i levels in 1-G platelets (data notshown). Together, these data indicate that hypergravity selec-tively elicits Ca2� release from intracellular organelle(s),rather than promoting Ca2� influx via mechanosensitive Ca2�
channels in the plasma membrane.
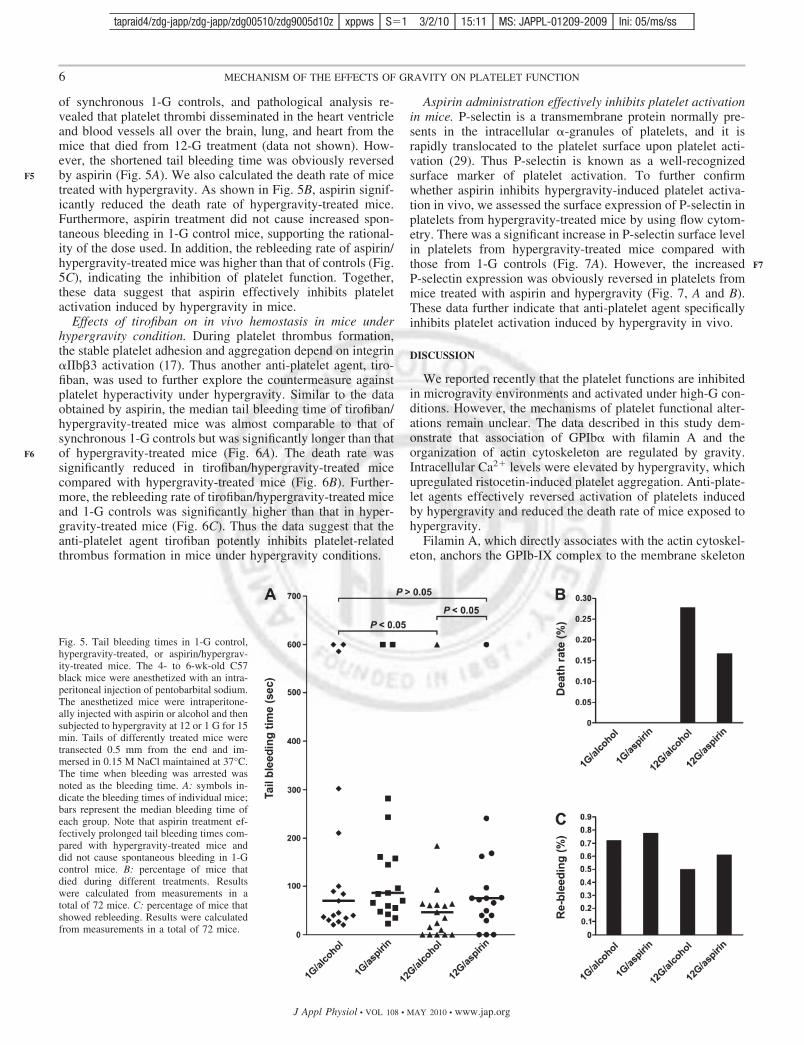
Role of [Ca2�]i in ristocetin- and ADP-induced plateletaggregation. The elevation of [Ca2�]i is sufficient for promotingcytoskeletal reorganization (25); on the other hand, reorganizationof the actin cytoskeleton also modulates intracellular Ca2� release(4, 35). To investigate the association of [Ca2�]i with cytoskeletalreorganization in platelets under hypergravity conditions, we pre-treated washed human platelets with BAPTA-AM, a membrane-permeable Ca2� chelator, for 20 min before application of 8-Ghypergravity. As shown in Fig. 4A, chelation of intracellular Ca2�
by BAPTA-AM did not attenuate the hypergravity-induced for-mation and distribution of actin filaments in the cytosol of plate-lets. These data indicate that the change in actin cytoskeleton isupstream or independent of intracellular Ca2� release in plateletsexposed to hypergravity.
Ristocetin-induced platelet aggregation was enhanced inhypergravity-treated platelets (8, 11). To investigate the role of
Fig. 2. Effects of simulated MG and hyper-gravity on platelet actin cytoskeleton. Washedhuman platelets were exposed to simulatedMG or 1 G for 1 h (A) or to 8-G hypergravityor 1 G for 15 min (B) and then fixed with 4%paraformaldehyde and stained using rhodam-ine-conjugated phalloidin. Note that 1-G con-trol platelets contained few actin filaments incytosol, whereas the actin fibers were con-verged into peripheral raffles after the applica-tion of simulated MG for 1 h. In contrast,dense actin filaments were formed in cytosolafter the application of 8-G hypergravity for 15min. Representative results of 3 independentexperiments are shown.
Fig. 3. Hypergravity induces intraplateletCa2� release. A: washed human plateletswere loaded with fluo-3 AM and then ex-posed to 8 or 1 G for 15 min in the presenceor absence of 5 mM EDTA (or EGTA). Theintracellular Ca2� level ([Ca2�]i) was ana-lyzed using flow cytometry. Representativehistograms of fluo-3 fluorescence of plateletsare shown. B: the geometric mean fluores-cence intensity of fluo-3 AM binding is in-dicated. Data are means � SD of 3 indepen-dent experiments. **P 0.01 comparedwith 1-G control.
4 MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
F4
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss
[Ca2�]i in ristocetin-induced platelet aggregation, PRP waspretreated with BAPTA-AM before hypergravity at 8 G andthen induced to aggregate with ristocetin. The low concentra-tion of ristocetin was used to induce a relatively lower aggre-gation in 1-G control platelets that would sensitively reflectplatelet functional variations. Compared with the synchronous1-G controls, ristocetin-induced platelet aggregation was sig-nificantly increased after exposure of platelets to hypergravity,whereas BAPTA-AM significantly reduced ristocetin-inducedaggregation of hypergravity-exposed platelets (Fig. 4, B andC); in contrast, BAPTA-AM had no effect on the aggregationof ristocetin-induced control platelets (data not shown).
In addition, our prior findings suggest that the plateletsignaling system and reaction tend to be exhausted in hyper-gravity conditions, and thus hypergravity-exposed plateletsexhibit decreased response to ADP (8, 11). To further explorethe effect of intracellular Ca2� release on platelet signalingsystem, we tested ADP-induced platelet aggregation. PRP waspretreated with BAPTA-AM before hypergravity and theninduced to aggregate with ADP. Consistent with our previousdata, 8 G-exposed platelets displayed a decreased response toADP compared with 1-G controls (Fig. 4, D and E). However,BAPTA-AM obviously enhanced ADP-induced platelet aggre-gation (Fig. 4, D and E), indicating that the platelet signaling
system and reaction are relatively more intact in BAPTA-AM/hypergravity-treated platelets than in hypergravity-treated plate-lets. Compared with hypergravity-treated platelets, BAPTA-AMdid not affect ADP-induced control platelet aggregation (data notshown). Together, these results indicate that intracellular Ca2�
release incurred by hypergravity is not required for actin cytoskel-eton reorganization of platelets but is involved in the regulation ofplatelet function. The cytoskeletal reorganization appears to beupstream or independent of intracellular Ca2� release in responseto hypergravity stimulation.
Effects of aspirin on in vivo hemostasis in mice underhypergravity condition. Our previous studies have demon-strated that hypergravity-treated platelets are hypersensitive toagonists (8, 11, 19). Particularly, the tail bleeding time wassignificantly shortened in mice exposed to high-G force, andplatelet thrombi were disseminated in the heart and circulationsystem from the mice that died from high-G force (8, 11).Therefore, to further confirm the role of hypergravity in plate-let activation in vivo and to find the approaches of preventingor curing hypergravity-related thrombotic diseases, we exam-ined the effect of aspirin on in vivo hemostasis in mice underhypergravity conditions. Consistent with our previous findings(8, 11), the median tail bleeding time was significantly short-ened in mice exposed to hypergravity at 12 G in contrast to that
Fig. 4. Effects of intracellular Ca2� release onactin reorganization and ristocetin- and ADP-induced platelet aggregation. A: washed hu-man platelets, pretreated with or without theintracellular Ca2� chelator BAPTA-AM (10�M) or vehicle (DMSO) at 37°C for 20 min,were exposed to 8 or 1 G for 15 min. Theplatelets were fixed and stained with rhodam-ine-conjugated phalloidin. Note that hyper-gravity-induced reorganizations of actin cy-toskeleton were not suppressed by BAPTA-AM. Data are representative of 3 independentexperiments. B and C: after recovering to aresting state, platelet-rich plasma (PRP) wasexposed to 8 or 1 G for 15 min or waspreincubated with BAPTA-AM (10 �M) orvehicle at 37°C for 20 min and then exposed to8-G hypergravity for 15 min. The differentlytreated PRP were induced to aggregate byaddition of ristocetin (0.6–0.8 mg/ml) (B).Note that chelation of [Ca2�]i significantly in-hibited ristocetin/hypergravity-induced plateletaggregation. Quantitative data are means � SDfrom 3 separate experiments (C). Aggregation(%) indicates the percentage of aggregated plate-lets. D and : PRP was exposed to 8 or 1 G for 15min or was pretreated with BAPTA-AM (10�M) or vehicle at 37°C for 20 min and thenexposed to 8-G hypergravity for 15 min. Theplatelets were stimulated with ADP to induceplatelet aggregation (D). Data are means � SDfrom 3 independent experiments (E).
5MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss
of synchronous 1-G controls, and pathological analysis re-vealed that platelet thrombi disseminated in the heart ventricleand blood vessels all over the brain, lung, and heart from themice that died from 12-G treatment (data not shown). How-ever, the shortened tail bleeding time was obviously reversedby aspirin (Fig. 5A). We also calculated the death rate of micetreated with hypergravity. As shown in Fig. 5B, aspirin signif-icantly reduced the death rate of hypergravity-treated mice.Furthermore, aspirin treatment did not cause increased spon-taneous bleeding in 1-G control mice, supporting the rational-ity of the dose used. In addition, the rebleeding rate of aspirin/hypergravity-treated mice was higher than that of controls (Fig.5C), indicating the inhibition of platelet function. Together,these data suggest that aspirin effectively inhibits plateletactivation induced by hypergravity in mice.
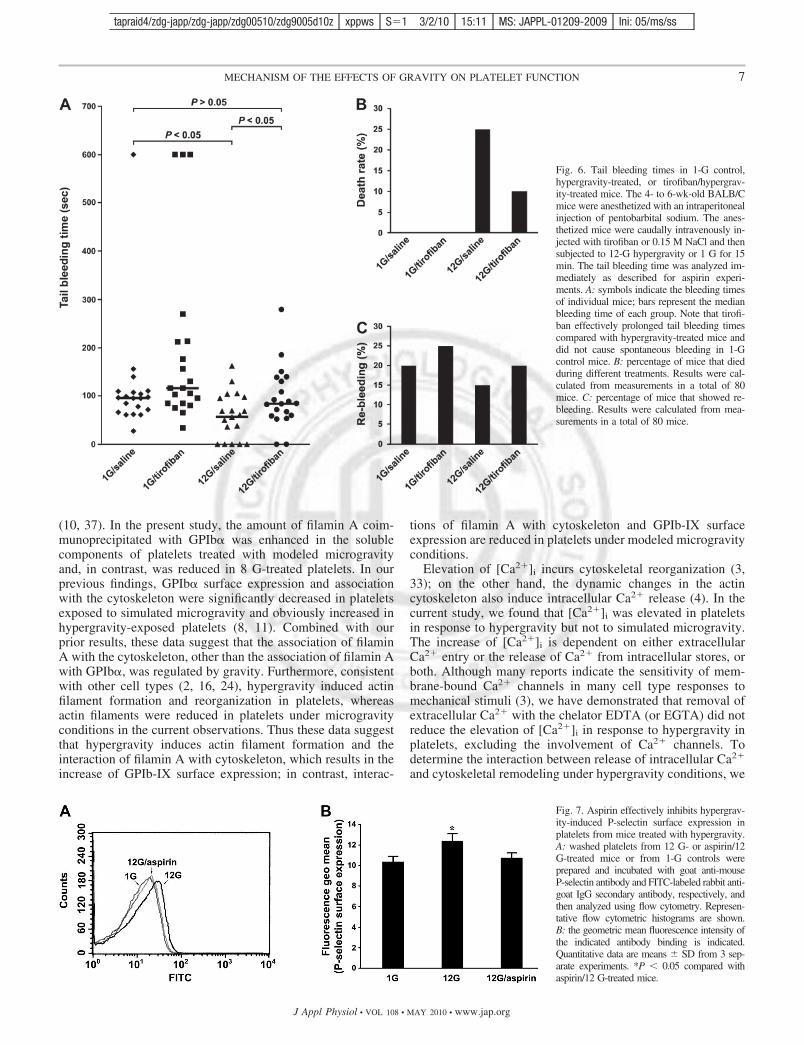
Effects of tirofiban on in vivo hemostasis in mice underhypergravity condition. During platelet thrombus formation,the stable platelet adhesion and aggregation depend on integrin�IIb3 activation (17). Thus another anti-platelet agent, tiro-fiban, was used to further explore the countermeasure againstplatelet hyperactivity under hypergravity. Similar to the dataobtained by aspirin, the median tail bleeding time of tirofiban/hypergravity-treated mice was almost comparable to that ofsynchronous 1-G controls but was significantly longer than thatof hypergravity-treated mice (Fig. 6A). The death rate wassignificantly reduced in tirofiban/hypergravity-treated micecompared with hypergravity-treated mice (Fig. 6B). Further-more, the rebleeding rate of tirofiban/hypergravity-treated miceand 1-G controls was significantly higher than that in hyper-gravity-treated mice (Fig. 6C). Thus the data suggest that theanti-platelet agent tirofiban potently inhibits platelet-relatedthrombus formation in mice under hypergravity conditions.
Aspirin administration effectively inhibits platelet activationin mice. P-selectin is a transmembrane protein normally pre-sents in the intracellular �-granules of platelets, and it israpidly translocated to the platelet surface upon platelet acti-vation (29). Thus P-selectin is known as a well-recognizedsurface marker of platelet activation. To further confirmwhether aspirin inhibits hypergravity-induced platelet activa-tion in vivo, we assessed the surface expression of P-selectin inplatelets from hypergravity-treated mice by using flow cytom-etry. There was a significant increase in P-selectin surface levelin platelets from hypergravity-treated mice compared withthose from 1-G controls (Fig. 7A). However, the increasedP-selectin expression was obviously reversed in platelets frommice treated with aspirin and hypergravity (Fig. 7, A and B).These data further indicate that anti-platelet agent specificallyinhibits platelet activation induced by hypergravity in vivo.
DISCUSSION
We reported recently that the platelet functions are inhibitedin microgravity environments and activated under high-G con-ditions. However, the mechanisms of platelet functional alter-ations remain unclear. The data described in this study dem-onstrate that association of GPIb� with filamin A and theorganization of actin cytoskeleton are regulated by gravity.Intracellular Ca2� levels were elevated by hypergravity, whichupregulated ristocetin-induced platelet aggregation. Anti-plate-let agents effectively reversed activation of platelets inducedby hypergravity and reduced the death rate of mice exposed tohypergravity.
Filamin A, which directly associates with the actin cytoskel-eton, anchors the GPIb-IX complex to the membrane skeleton
Fig. 5. Tail bleeding times in 1-G control,hypergravity-treated, or aspirin/hypergrav-ity-treated mice. The 4- to 6-wk-old C57black mice were anesthetized with an intra-peritoneal injection of pentobarbital sodium.The anesthetized mice were intraperitone-ally injected with aspirin or alcohol and thensubjected to hypergravity at 12 or 1 G for 15min. Tails of differently treated mice weretransected 0.5 mm from the end and im-mersed in 0.15 M NaCl maintained at 37°C.The time when bleeding was arrested wasnoted as the bleeding time. A: symbols in-dicate the bleeding times of individual mice;bars represent the median bleeding time ofeach group. Note that aspirin treatment ef-fectively prolonged tail bleeding times com-pared with hypergravity-treated mice anddid not cause spontaneous bleeding in 1-Gcontrol mice. B: percentage of mice thatdied during different treatments. Resultswere calculated from measurements in atotal of 72 mice. C: percentage of mice thatshowed rebleeding. Results were calculatedfrom measurements in a total of 72 mice.
6 MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
F5
F6
F7
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss
(10, 37). In the present study, the amount of filamin A coim-munoprecipitated with GPIb� was enhanced in the solublecomponents of platelets treated with modeled microgravityand, in contrast, was reduced in 8 G-treated platelets. In ourprevious findings, GPIb� surface expression and associationwith the cytoskeleton were significantly decreased in plateletsexposed to simulated microgravity and obviously increased inhypergravity-exposed platelets (8, 11). Combined with ourprior results, these data suggest that the association of filaminA with the cytoskeleton, other than the association of filamin Awith GPIb�, was regulated by gravity. Furthermore, consistentwith other cell types (2, 16, 24), hypergravity induced actinfilament formation and reorganization in platelets, whereasactin filaments were reduced in platelets under microgravityconditions in the current observations. Thus these data suggestthat hypergravity induces actin filament formation and theinteraction of filamin A with cytoskeleton, which results in theincrease of GPIb-IX surface expression; in contrast, interac-
tions of filamin A with cytoskeleton and GPIb-IX surfaceexpression are reduced in platelets under modeled microgravityconditions.
Elevation of [Ca2�]i incurs cytoskeletal reorganization (3,33); on the other hand, the dynamic changes in the actincytoskeleton also induce intracellular Ca2� release (4). In thecurrent study, we found that [Ca2�]i was elevated in plateletsin response to hypergravity but not to simulated microgravity.The increase of [Ca2�]i is dependent on either extracellularCa2� entry or the release of Ca2� from intracellular stores, orboth. Although many reports indicate the sensitivity of mem-brane-bound Ca2� channels in many cell type responses tomechanical stimuli (3), we have demonstrated that removal ofextracellular Ca2� with the chelator EDTA (or EGTA) did notreduce the elevation of [Ca2�]i in response to hypergravity inplatelets, excluding the involvement of Ca2� channels. Todetermine the interaction between release of intracellular Ca2�
and cytoskeletal remodeling under hypergravity conditions, we
Fig. 6. Tail bleeding times in 1-G control,hypergravity-treated, or tirofiban/hypergrav-ity-treated mice. The 4- to 6-wk-old BALB/Cmice were anesthetized with an intraperitonealinjection of pentobarbital sodium. The anes-thetized mice were caudally intravenously in-jected with tirofiban or 0.15 M NaCl and thensubjected to 12-G hypergravity or 1 G for 15min. The tail bleeding time was analyzed im-mediately as described for aspirin experi-ments. A: symbols indicate the bleeding timesof individual mice; bars represent the medianbleeding time of each group. Note that tirofi-ban effectively prolonged tail bleeding timescompared with hypergravity-treated mice anddid not cause spontaneous bleeding in 1-Gcontrol mice. B: percentage of mice that diedduring different treatments. Results were cal-culated from measurements in a total of 80mice. C: percentage of mice that showed re-bleeding. Results were calculated from mea-surements in a total of 80 mice.
Fig. 7. Aspirin effectively inhibits hypergrav-ity-induced P-selectin surface expression inplatelets from mice treated with hypergravity.A: washed platelets from 12 G- or aspirin/12G-treated mice or from 1-G controls wereprepared and incubated with goat anti-mouseP-selectin antibody and FITC-labeled rabbit anti-goat IgG secondary antibody, respectively, andthen analyzed using flow cytometry. Represen-tative flow cytometric histograms are shown.B: the geometric mean fluorescence intensity ofthe indicated antibody binding is indicated.Quantitative data are means � SD from 3 sep-arate experiments. *P 0.05 compared withaspirin/12 G-treated mice.
7MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss

examined the effect of the intracellular Ca2� chelatorBAPTA-AM on changes of actin cytoskeleton, and the dataindicate that addition of BAPTA-AM did not prevent thechanges in cytoskeletal reorganization, excluding the necessityof intracellular Ca2� release for actin reorganization. However,whether Ca2� release in response to hypergravity is a down-stream event or is independent of actin reorganization remainsto be further investigated.
In platelets, elevation of [Ca2�]i is involved in the regulationof almost every important function of platelets such as adhe-sion, secretion, and aggregation (13). Furthermore, elevation of[Ca2�]i leads to platelet hyperactivity (20). Thus we investi-gated the role of [Ca2�]i in platelet hyperactivity induced byhypergravity. The data indicate that [Ca2�]i increase plays akey role in platelet hyperactivity under hypergravity condi-tions. An increase of GPIb-IX surface expression by itselfcannot incur platelet activation. Thus the elevation of [Ca2�]i
might present as the main reason leading to platelet hyperac-tivity under hypergravity conditions.
Aspirin is widely used as an antithrombotic drug that effec-tively blocks the arachidonate pathway by inhibiting prosta-glandin synthesis, thus preventing platelet activation and re-ducing the rate of thrombotic formation (23). Tirofiban is anovel nonpeptide platelet antagonist and has a high degree ofselectivity for the platelet �IIb3 receptor (6). Therefore, tofurther confirm the role of hypergravity in platelet activation invivo and to preliminarily explore the countermeasure againsthypergravity-related thrombotic diseases, we examined theeffects of aspirin and tirofiban on in vivo hemostasis in miceunder hypergravity conditions. The data indicate that adminis-tration of aspirin or tirofiban effectively inhibited in vivoplatelet activation induced by hypergravity and reduced deathrate in mice under hypergravity conditions, which not onlyfurther confirms the activation of platelets in vivo but alsosuggests the possibility of preventing thrombus formation in ahypergravitational environment.
In conclusion, the association of GPIb� with filamin A andthe organization of actin cytoskeleton were regulated by dif-ferent gravity conditions. Intracellular Ca2� levels were ele-vated by hypergravity, which obviously enhanced ristocetin-induced platelet aggregation. Two anti-platelet agents, aspirinand tirofiban, effectively reversed activation of platelets in-duced by hypergravity in vivo and reduced the death rate ofmice exposed to hypergravity. Thus these data implicate themechanisms underlaying the functional variations of plateletsunder different gravity conditions and also suggest an effectivecountermeasure against hypergravity-related thrombotic dis-eases.
ACKNOWLEDGMENTS
We thank Drs. Changgeng Ruan (Soochow University, Suzhou, China) andXiaoping Du (University of Illinois, Chicago, IL) for providing reagents.
GRANTS
This work was supported by National Natural Science Foundation of ChinaGrants NSFC 30971067 and NSFC 30770795, Program for New CenturyExcellent Talents in University Grant NCET-06-0167, and Foundation for theAuthor of National Excellent Doctoral Dissertation of the People’s Republic ofChina Grant FANEDD 200560, an opening foundation of the State KeyLaboratory of Space Medicine Fundamentals and Application (China Astro-naut Research and Training Center). S. Li is a recipient of the InnovationFoundation of Beihang University of Aeronautics and Astronautics for PhDgraduates.
DISCLOSURES
No conflicts of interest are declared by the author(s).
REFERENCES
1. Buravkova LB, Romanov YA. The role of cytoskeleton in cell changesunder condition of simulated microgravity. Acta Astronaut 48: 647–650,2001.
2. Carlsson SI, Bertilaccio MT, Ballabio E, Maier JA. Endothelial stressby gravitational unloading: effects on cell growth and cytoskeletal orga-nization. Biochim Biophys Acta 1642: 173–179, 2003.
3. Chen NX, Ryder KD, Pavalko FM, Turner CH, Burr DB, Qiu J,Duncan RL. Ca2� regulates fluid shear-induced cytoskeletal reorganiza-tion and gene expression in osteoblasts. Am J Physiol Cell Physiol 278:C989–C997, 2000.
4. Chun JT, Santella L. Roles of the actin-binding proteins in intracellularCa2� signalling. Acta Physiol (Oxf) 195: 61–70, 2009.
5. Clement JQ, Lacy SM, Wilson B. Genome-wide gene expression pro-filing of microgravity effect on human liver cells. J Gravit Physiol 14:121–122, 2007.
6. Cohen M, Théroux P, Weber S, Laramée P, Huynh T, Borzak S,Diodati JG, Squire IB, Deckelbaum LI, Thornton AR, Harris KE, SaxFL, Lo M, White HD. Combination therapy with tirofiban and enoxaparinin acute coronary syndromes. Int J Cardiol 71: 273–281, 1999.
7. Dai K, Bodnar R, Berndt M, Du X. A critical role for 14-3-3� protein inregulating the VWF binding function of platelet glycoprotein Ib-IX and itstherapeutic implications. Blood 106: 1975–1981, 2005.
8. Dai K, Wang Y, Yan R, Shi Q, Wang Z, Yuan Y, Cheng H, Li S, FanY, Zhuang F. Effects of microgravity and hypergravity on plateletfunctions. Thromb Haemost 101: 902–910, 2009.
9. Dai K, Yan R, Li S, Fan Y, Zhuang F, Ruan C. Prolonged inhibition ofprotein kinase A results in metalloproteinase-dependent platelet GPIb�shedding. Thromb Res 124: 101–109, 2009.
10. Feng S, Resendiz JC, Lu X, Kroll MH. Filamin A binding to thecytoplasmic tail of glycoprotein Ib� regulates von Willebrand factor-induced platelet activation. Blood 102: 2122–2129, 2003.
11. Gerzer R. Hypergravity and microgravity influence hemostasis. ThrombHaemost 101: 799, 2009.
12. Hammond TG, Hammond JM. Optimized suspension culture: the rotat-ing-wall vessel. Am J Physiol Renal Physiol 281: F12–F25, 2001.
13. Holinstat M, Voss B, Bilodeau ML, McLaughlin JN, Cleator J, HammHE. PAR4, but not PAR1, signals human platelet aggregation via Ca2�
mobilization and synergistic P2Y12 receptor activation. J Biol Chem 281:26665–26674, 2006.
14. Hughes-Fulford M. Function of the cytoskeleton in gravisensing duringspaceflight. Adv Space Res 32: 1585–1593, 2003.
15. Hung CT, Allen FD, Pollack SR, Brighton CT. Intracellular Ca2� storesand extracellular Ca2� are required in the real-time Ca2� response of bonecells experiencing fluid flow. J Biomech 29: 1411–1417, 1996.
16. Kacena MA, Todd P, Gerstenfeld LC, Landis WJ. Experiments withosteoblasts cultured under hypergravity conditions. Microgravity SciTechnol 15: 28–34, 2004.
17. Kasirer-Friede A, Moran B, Nagrampa-Orje J, Swanson K, RuggeriZM, Schraven B, Neel BG, Koretzky G, Shattil SJ. ADAP is requiredfor normal �IIb3 activation by VWF/GP Ib-IX-V and other agonists.Blood 109: 1018–1025, 2007.
18. Koyama T, Kimura C, Hayashi M, Watanabe M, Karashima Y, OikeM. Hypergravity induces ATP release and actin reorganization via ty-rosine phosphorylation and RhoA activation in bovine endothelial cells.Pflügers Arch 457: 711–719, 2009.
19. Li S, Shi Q, Wang ZH, Yan R, Cheng H, Dai K. Hypergravity resultsin human platelet hyperactivity. J Physiol Biochem 65: 147–156, 2009.
20. Li Y, Woo V, Bose R. Platelet hyperactivity and abnormal Ca2� ho-meostasis in diabetes mellitus. Am J Physiol Heart Circ Physiol 280:H1480–H1489, 2001.
21. Mehta D, Konstantoulaki M, Ahmmed GU, Malik AB. Sphingosine1-phosphate-induced mobilization of intracellular Ca2� mediates rac ac-tivation and adherens junction assembly in endothelial cells. J Biol Chem280: 17320–17328, 2005.
22. Meyers VE, Zayzafoon M, Douglas JT, Mcdonald JM. RhoA andcytoskeletal disruption mediate reduced osteoblastogenesis and enhancedadipogenesis of human mesenchymal stem cells in modeled microgravity.J Bone Miner Res 20: 1858–1866, 2005.
8 MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
AQ: 4
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss

23. Michelson AD, Cattaneo M, Eikelboom JW, Gurbel P, Kottke-March-ant K, Kunicki TJ, Pulcinelli FM, Cerletti C, Rao AK. Aspirinresistance: position paper of the Working Group on Aspirin Resistance. JThromb Haemost 3: 1309–1311, 2005.
24. Monici M, Marziliano N, Basile V, Romano G, Conti A, Pezzatini S,Morbidelli L. Hypergravity affects morphology and function in micro-vascular endothelial cells. Microgravity Sci Technol 18: 234–238, 2006.
25. Nebe B, Bohn W, Sanftleben H, Rychly J. Induction of a physicallinkage between integrins and the cytoskeleton depends on intracellularcalcium in an epithelial cell line. Exp Cell Res 229: 100–110, 1996.
26. Ozaki Y, Asazuma N, Suzuki-Inoue K, Berndt MC. Platelet GPIb-IX-V-dependent signaling. J Thromb Haemost 3: 1745–1751, 2005.
27. Pelletier AR, Gilchrist J. Roller coaster related fatalities, United States,1994–2004. Inj Prev 11: 309–312, 2005.
28. Pringle SD, Macfarlane PW, Cobbe SM. Response of heart rate to aroller coaster ride. BMJ 299: 1575, 1989.
29. Rendu F, Brohard-Bohn B. The platelet release reaction: granulesconstituents, secretion and functions. Platelets 12: 261–273, 2001.
30. Rosner H, Wassermann T, Moller W, Hanke W. Effects of alteredgravity on the actin and microtubule cytoskeleton of human SH-SY5Yneuroblastoma cells. Protoplasma 229: 225–234, 2006.
31. Rowe WJ. Interplanetary travel and permanent injury to normal heart.Acta Astronaut 40: 719–722, 1997.
32. Rowe WJ. The Apollo 15 space syndrome. Circulation 97: 119–120,1998.
33. Schwarz U, Walter U, Eigenthaler M. Taming platelets with cyclicnucleotides. Biochem Pharmacol 62: 1153–1161, 2001.
34. Searby ND, Steele CR, Globus RK. Influence of increased mechanicalloading by hypergravity on the microtubule cytoskeleton and prostaglan-din E2 release in primary osteoblasts. Am J Physiol Cell Physiol 289:C148–C158, 2005.
35. Toyota M, Furuichi T, Tatsumi H, Sokabe M. Cytoplasmic calciumincreases in response to changes in the gravity vector in hypocotyls andpetioles of Arabidopsis seedlings. Plant Physiol 146: 505–514, 2008.
36. Toyota M, Furuichi T, Tatsumi H, Sokabe M. Hypergravity stimulationinduces changes in intracellular calcium concentration in Arabidopsisseedlings. Adv Space Res 39: 1190–1197, 2007.
37. Vanhoorelbeke K, Ulrichts H, Van de Walle G, Gerlinde V, AlexandreF, Hans D. Inhibition of platelet glycoprotein Ib and its antithromboticpotential. Curr Pharm Des 13: 2684–2697, 2007.
38. Versari S, Villa A, Bradamante S, Maier JA. Alterations of the actincytoskeleton and increased nitric oxide synthesis are common features inhuman primary endothelial cell response to changes in gravity. BiochimBiophys Acta 1773: 1645–1652, 2007.
39. Watenpaugh DE. Fluid volume control during short-term space flight andimplications for human performance. J Exp Biol 204: 3209–3215, 2001.
9MECHANISM OF THE EFFECTS OF GRAVITY ON PLATELET FUNCTION
J Appl Physiol • VOL 108 • MAY 2010 • www.jap.org
tapraid4/zdg-japp/zdg-japp/zdg00510/zdg9005d10z xppws S�1 3/2/10 15:11 MS: JAPPL-01209-2009 Ini: 05/ms/ss





























