Embed Size (px)
Citation preview

Update on Plastid Division
Mechanism of Plastid Division: From a Bacterium toan Organelle1
Shin-ya Miyagishima*
Center for Frontier Research, National Institute of Genetics, Shizuoka 411–8540, Japan; and Initiative ResearchProgram, Advanced Science Institute, RIKEN, Saitama 351–0198, Japan
Chloroplasts arose more than 1 billion years ago,when a free-living cyanobacterium became an endo-symbiont in a eukaryotic host cell. Several lines ofevidence indicate that all chloroplasts and their non-photosynthetic relatives (plastids) are directly or in-directly derived from a single endosymbiotic event(Reyes-Prieto et al., 2007; Gould et al., 2008; Archibald,2009; Fig. 1). The original endosymbiosis of the cya-nobacterium gave rise to plastids (primary plastids) inthe Archaeplastida, which are made up of the Glau-cophya, Rhodophyta (red algae), and Viridiplantae(green algae and land plants). All other plastids, suchas those found in kelps, dinoflagellates, malaria par-asites (these three belong to the Chromalveolata), eu-glenids (Excavata), and chlorarachniophytes (Rhizaria),were established by subsequent endosymbiotic eventsin which eukaryotic algae had become integrated intoother eukaryotes. Recent phylogenetic studies haveclassified eukaryotes into six supergroups: Amoebo-zoa, Opisthokonta (fungi and animals), Archaeplas-tida, Chromalveolata, Excavata, and Rhizaria. Thus,four of the six eukaryotic supergroups possess plastids(Reyes-Prieto et al., 2007; Gould et al., 2008; Archibald,2009; Fig. 1).The conversion of a cyanobacterium to plastids re-
quired several steps (Reyes-Prieto et al., 2007; Gouldet al., 2008; Archibald, 2009). Theoretically, a specificrelationship developed between an endosymbiont andhost, such as a predator-prey relationship. Subse-quently, mechanisms controlling metabolic exchangebetween the two developed. Most of the genes thatwere once in the endosymbiont genomewere either lostor transferred into the host nuclear genome. As a result,the size of the plastid genome has been reduced to lessthan one-tenth that of the free-living cyanobacterialgenome. Thus, the bulk of the plastid proteome consistsof nucleus-encoded proteins that are translated oncytoplasmic ribosomes. In order to translocate proteinsfrom the cytoplasm into chloroplasts, a protein importapparatus was developed at the envelope membrane.In addition, a mechanism that controls endosymbiotic
cell division was developed such that each daughtercell would inherit the endosymbiont during cell di-vision. Generally, algal cells contain either a singlecholoroplast or just a few chloroplasts. Thus, chloro-plast division is synchronized with the host cell cyclesuch that the chloroplast divides before cytokinesis andis thus transmitted into each daughter cell.
The requirement of division synchronization is sup-ported by several other endosymbiotic relationships. Forexample, certain species of heterotrophic dinoflagellatesengulf eukaryotic algae and use them as temporarychloroplasts (called “kleptoplasts”) for a period of daysto weeks before digesting them. In contrast, a few dino-flagellate species maintain a eukaryotic algal unicell (i.e.containing the nucleus, mitochondria, Golgi apparatuses,etc.) as a permanent endosymbiont by synchronizing theendosymbiont cell division to the host cell cycle (Wouterset al., 2009; Imanian et al., 2010). A similar situation is alsoobserved in other eukaryotic groups.Hatena arenicola hasa transient green algal endosymbiont, and this photo-synthetic endosymbiont is inherited by only one daugh-ter cell during cell division. Daughter cells that have lostthe endosymbiont again engulf the green alga (Okamotoand Inouye, 2005). H. arenicola (Katablepharidophyta)bears a close evolutionary relationship with crypto-phytes, in which permanent chloroplasts of a red algalorigin have been established by division synchronization(Fig. 1). There are also eukaryotes that possess permanentcyanobacterial endosymbionts, such as Paulinella chroma-tophora. This endosymbiont is consistently inherited byprogeny cells as a consequence of tight synchronizationof host and endosymbiotic cell cycles (Melkonian andMollenhauer, 2005).
Currently, it is largely unknown how endosymbioticcell division is regulated by host cells in the diversearray of endosymbiotic relationships. However, studiesover the last decade have rapidly provided informationon the mechanism and the regulation of primary chlo-roplast division (Kuroiwa et al., 2008; Yang et al., 2008;Maple and Møller, 2010; Miyagishima and Kabeya,2010; Pyke, 2010). In addition, several genome projectson the plastid-carrying eukaryotes have been initiated.These advances promise the eventual understanding ofthe mechanisms that coordinate the proliferation ofboth a host cell and an endosymbiont.
The mechanism of primary plastid division, espe-cially that of chloroplasts, has been summarized in
1 This work was supported by a Grant-in-Aid for ScientificResearch from the Ministry of Education, Culture, Sports, Science,and Technology of Japan (grant no. 22687020).
* E-mail [email protected]/cgi/doi/10.1104/pp.110.170688
Plant Physiology�, April 2011, Vol. 155, pp. 1533–1544, www.plantphysiol.org � 2011 American Society of Plant Biologists 1533

recent reviews (Kuroiwa et al., 2008; Yang et al., 2008;Maple and Møller, 2010; Miyagishima and Kabeya,2010; Pyke, 2010). Here, the mechanism of primarychloroplast division is briefly introduced, and then thecommon and diverse mechanisms underlying divisionin other types of plastids and photosynthetic endo-symbionts will be discussed. Note that, in this review,the term “chloroplasts” is used to indicate photosyn-thetic plastids.
PRIMARY CHLOROPLAST DIVISION BY DIVISIONCOMPLEX OF CHIMERIC ORIGIN
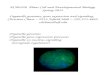
Chloroplasts usually divide by binary fission (Fig.2), but multiple fission also occurs in certain cases(Duckett and Ligrone, 1993). Earlier electron micro-scopic studies established that chloroplast division isperformed by the simultaneous constriction of theinner and outer envelopes at the division site. Since the1980s, electron-dense ring structures at the divisionsite called plastid-dividing (PD) rings have beendetected on both the inside and outside of the chloro-plast, suggesting that chloroplast division is per-formed by constriction of the ring-like divisioncomplex (Kuroiwa et al., 1998).
Recent studies, especially molecular genetic studieson leaf chloroplasts in Arabidopsis (Arabidopsis thali-ana) and chloroplasts in the unicellular red algaCyanidioschyzon merolae, have identified several com-ponents of the division complex. As observed byelectronmicroscopy, the division complex encompassesboth the inside and the outside of the two envelopes
(Fig. 2). All of the known components are encoded inthe host nuclear genome, except for a few componentsof cyanobacterial origin encoded in the chloroplastgenome in certain algal lineages (Kuroiwa et al., 2008;Yang et al., 2008; Maple and Møller, 2010; Miyagishimaand Kabeya, 2010; Pyke, 2010).
Components Descended from the
Cyanobacterial Endosymbiont
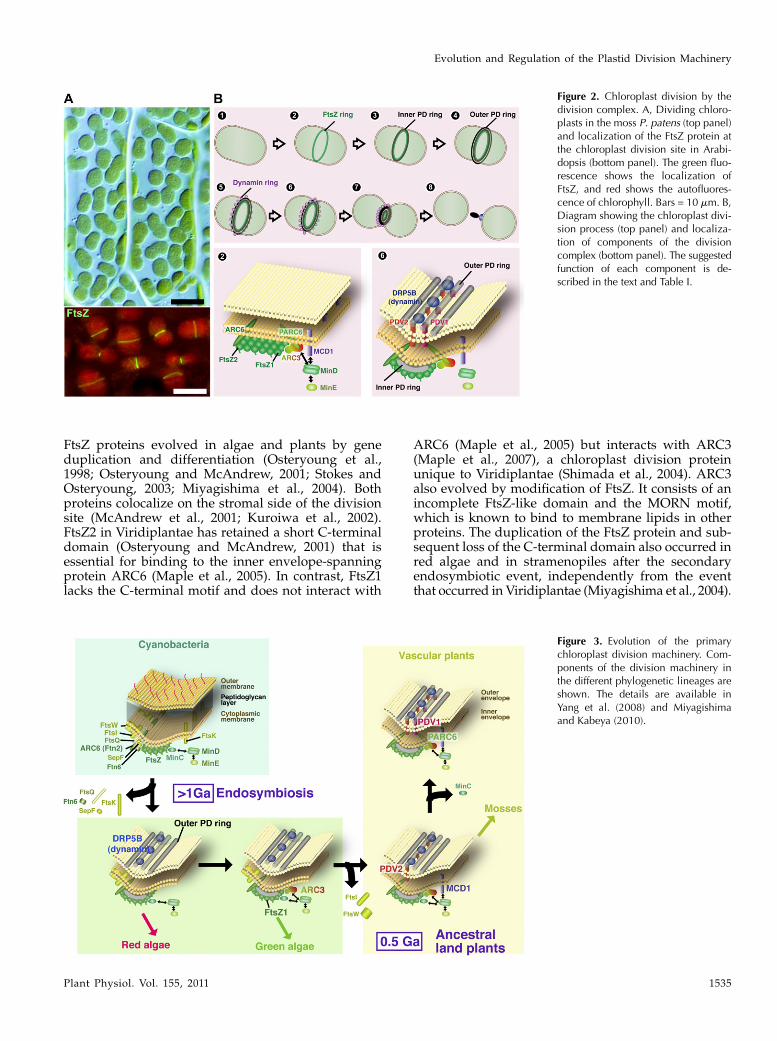
Consistent with the endosymbiotic origin of plas-tids, the stromal portion of the chloroplast divisioncomplex is descended from the cyanobacterial celldivision machinery (Figs. 2 and 3; Table I). As inbacteria, Filamenting temperature-sensitive mutant Z(FtsZ; a prokaryotic tubulin-like GTPase) self-assem-bles into a ring structure on the stromal side of thechloroplast division site and is involved in the divisionprocess (Osteryoung and Vierling, 1995; Osteryounget al., 1998; Strepp et al., 1998; Mori et al., 2001; Vithaet al., 2001). In addition to FtsZ, certain other compo-nents, such as ACCUMULATION AND REPLICA-TION OF CHLOROPLAST 6 (ARC6; an ortholog ofcyanobacterial division protein Ftn2; Vitha et al., 2003),are also descended from the cyanobacterial divisionmachinery. However, many other components werelost after the endosymbiotic event (Figs. 2 and 3).
Although plastids have retained some of the cyano-bacterial division machinery, both duplication anddifferentiation of the cyanobacteria-descended com-ponents have played significant roles in the evolutionof the plastid division machinery (Figs. 2 and 3; TableI). While cyanobacterial genomes encode a single FtsZprotein, two phylogenetically and functionally distinct
Figure 1. A schematic view of the origin andspread of plastids in the putative six eukaryoticsupergroups (modified from Archibald, 2009).The broken lines denote uncertainty of branchpositions in the tree. Red algae and groups con-taining plastids of red algal origin are shown inred. Viridiplantae (green algae and land plants)and groups containing plastids of green algalorigin are shown in green. Arrows indicate theprimary endosymbiotic event of a cyanobacte-rium (1st) and secondary endosymbiotic events(2nd). The position of secondary endosymbioticevents in the tree is still uncertain at present. Notethat some dinoflagellates possess plastids of stra-menopile, haptophyte, or green algal origin, al-though those are not indicted in the scheme. It issuggested that these dinoflagellates replaced theoriginal red algal chloroplast with chloroplasts ofother origin by another endosymbiotic event(Gould et al., 2008).
Miyagishima
1534 Plant Physiol. Vol. 155, 2011
FtsZ proteins evolved in algae and plants by geneduplication and differentiation (Osteryoung et al.,1998; Osteryoung and McAndrew, 2001; Stokes andOsteryoung, 2003; Miyagishima et al., 2004). Bothproteins colocalize on the stromal side of the divisionsite (McAndrew et al., 2001; Kuroiwa et al., 2002).FtsZ2 in Viridiplantae has retained a short C-terminaldomain (Osteryoung and McAndrew, 2001) that isessential for binding to the inner envelope-spanningprotein ARC6 (Maple et al., 2005). In contrast, FtsZ1lacks the C-terminal motif and does not interact with
ARC6 (Maple et al., 2005) but interacts with ARC3(Maple et al., 2007), a chloroplast division proteinunique to Viridiplantae (Shimada et al., 2004). ARC3also evolved by modification of FtsZ. It consists of anincomplete FtsZ-like domain and the MORN motif,which is known to bind to membrane lipids in otherproteins. The duplication of the FtsZ protein and sub-sequent loss of the C-terminal domain also occurred inred algae and in stramenopiles after the secondaryendosymbiotic event, independently from the eventthat occurred in Viridiplantae (Miyagishima et al., 2004).
Figure 2. Chloroplast division by thedivision complex. A, Dividing chloro-plasts in the moss P. patens (top panel)and localization of the FtsZ protein atthe chloroplast division site in Arabi-dopsis (bottom panel). The green fluo-rescence shows the localization ofFtsZ, and red shows the autofluores-cence of chlorophyll. Bars = 10 mm. B,Diagram showing the chloroplast divi-sion process (top panel) and localiza-tion of components of the divisioncomplex (bottom panel). The suggestedfunction of each component is de-scribed in the text and Table I.
Figure 3. Evolution of the primarychloroplast division machinery. Com-ponents of the division machinery inthe different phylogenetic lineages areshown. The details are available inYang et al. (2008) and Miyagishimaand Kabeya (2010).
Evolution and Regulation of the Plastid Division Machinery
Plant Physiol. Vol. 155, 2011 1535

Duplication and diversification of ARC6, whichoccurred in a common ancestor of vascular plants,resulted in novel functions in the division complex(Glynn et al., 2009). ARC6 promotes FtsZ assembly(Vitha et al., 2003) and recruits the PLASTID DIVI-SION 2 (PDV2) protein through direct interaction inthe intermembrane space (Glynn et al., 2008). In con-trast, PARC6 (a paralog of ARC6 unique to vascularplants) inhibits FtsZ assembly, most likely through aninteraction with ARC3, and is required for the recruit-ment of PDV1 (a paralog of PDV2 unique to vascularplants; Glynn et al., 2009).
Components Derived from the Host Cell
In addition to the modification of cyanobacteria-descended proteins, some components of eukaryotichost origin have become integrated into the chloro-plast division machinery (Figs. 2 and 3; Table II).DRP5B (ARC5) is a member of the eukaryotic dynaminfamily of GTPases and localizes on the cytosolic side ofthe chloroplast division complex (Gao et al., 2003;Miyagishima et al., 2003). Proteins of this family formhelical assemblies on the membrane, and it is suggested
that constriction and disassembly of the helix result inmembrane fission (Roux and Antonny, 2008). Phyloge-netically, DRP5B is most closely related to DRP5Aprotein. DRP5A is not involved in plastid division(Gao, 2005) but rather is involved in cytokinesis inArabidopsis and the slime mold Dictyostelium discoi-deum (Miyagishima et al., 2008). Thus, the dynamin inchloroplast division probably evolved from the cytoki-netic activity of the eukaryotic host cell. PDV1 andPDV2 are unique to land plants and are required for therecruitment of DRP5B (Miyagishima et al., 2006). Itappears that PDV proteins connect the cyanobacteria-descended stromal complex and the host-derived cyto-solic complex (Glynn et al., 2008, 2009; Figs. 2 and 3).However, it is not known whether PDV proteins di-rectly interact with DRP5B or how the two complexesare connected in algae, which do not have PDV pro-teins.
The PD ring has been detected by electron micros-copy in several different lineages of algae and landplants to usually have two structures: the outer PDring on the cytosolic side of the outer envelope, andthe inner PD ring on the stromal side of the innerenvelope (Kuroiwa et al., 1998). Occasionally, a middle
Table I. Cyanobacteria-derived components of the chloroplast division machinery
The references are indicated in the text.
Component
(Localization)Function and Notes
FtsZ1/FtsZ2 (medial ring) Nucleus-encoded FtsZ proteins are required for normal chloroplast division. In bacteria, the FtsZ ringacts as the scaffold for recruitment of other key cell division proteins. Bacterial FtsZ self-assemblesinto a ring that constricts liposomes in vitro, suggesting that the FtsZ ring generates constrictive forcein vivo. Green algae and land plants have two types of phylogenetically distinct nucleus-encodedFtsZ proteins. FtsZ1 emerged by duplication of the FtsZ2 protein after the endosymbiotic event. UnlikeFtsZ2 and cyanobacterial FtsZ, FtsZ1 does not contain a short C-terminal motif. The C-terminal motifmediates the interaction between FtsZ2 and ARC6.
ARC6/PARC6 (medialring/medial ring and poles)
Nucleus-encoded ARC6 and PARC6 (a paralog of ARC6 unique to vascular plants) are required for normalchloroplast division. Both proteins are descended from Ftn2 (ZipN), which is a component of thecyanobacterial division complex. ARC6 promotes FtsZ assembly through direct interaction with FtsZ2,while PARC6 destabilizes the FtsZ polymer, most likely via direct interaction with ARC3. ARC6recruits PDV2 through direct interaction with PDV2 in the intermembrane space, while PARC6 isrequired for the recruitment of PDV1 (it is unknown whether PARC6 and PDV1 directly interact).
SepF (medial ring?) SepF is encoded in the plastid (cyanelle) genome of the glaucophyte C. paradoxa, but the function inplastid division has not been determined. In bacteria, SepF is required for normal cell division. SepFlocalizes at the division site by interaction with FtsZ.
FtsW (medial ring?) FtsW is encoded in the plastid genome of glaucophytes and a certain green algae, but the function isunknown. In bacteria, FtsW spans the cytoplasmic membrane at the division site and recruits FtsI.
FtsI (medial ring?) FtsI is encoded in the plastid genome of a certain green algae, but the function has not been determined.FtsI spans the cytoplasmic membrane at the division site and its periplasmic domain that cross-linksthe newly synthesized peptidoglycan strand.
MinC (unknown) MinC is encoded in the nuclear genome of (some) certain algal lineages and the moss P. patens, but thefunction has not been determined. In bacteria, MinC, MinD, MinE (proteobacteria), and DivIVA(firmicutes) determine the site of FtsZ ring formation. MinC inhibits FtsZ polymerization.
MinD (medial ringand punctate structures)
MinD is a membrane-associated ATPase. The nucleus-encoded MinD protein regulates positioning of thechloroplast FtsZ ring. In bacteria, MinD localizes MinC to the cytoplasmic membrane by directinteraction with MinC.
MinE (unknown) The nucleus-encoded MinE protein regulates the positioning of the chloroplast FtsZ ring. Inproteobacteria, MinE restricts the inhibitory activity of the MinCD complex to the cell pole,thereby allowing the FtsZ ring to form at the midcell position. MinE stimulates ATP hydrolysis ofMinD, which releases the MinCD complex from the cytoplasmic membrane.
Miyagishima
1536 Plant Physiol. Vol. 155, 2011

PD ring is observed in the intermembrane space(Miyagishima et al., 2001; Sumiya et al., 2008). A veryrecent study that was performed by dissecting theplastid division complex in the red alga C. merolaeshowed that the outer PD ring is a bundle of poly-glucan filaments (Yoshida et al., 2010). These filamentsare associated with the PDR1 protein, which is mostclosely related to the eukaryotic glycogenin proteins.Because glycogenin proteins are known to polymerizeGlc, PDR1 likely synthesizes the polyglucan filamentsof the PD ring. The filaments of the outer PD ring havebeen observed in several different lineages (Kuroiwaet al., 1998), and potential orthologs of PDR1 areencoded in land plant genomes (Yoshida et al., 2010).However, the genomes of other plastid-carrying eu-karyotes, such as green algae and stramenopiles, ap-parently lack the PDR1 gene; thus, how the filament issynthesized is uncertain at present.
Division Site Selection
Chloroplasts normally divide at the midpoint, yield-ing two daughter chloroplasts of equal size (Fig. 2).The mechanism responsible for the positioning of theFtsZ ring is also descended from the cyanobacterialmachinery known as the Min system (Figs. 2 and 3;Table I). The cyanobacterial Min system involvesMinC, MinD, MinE, and probably Cdv3 (DivIVA-like) proteins (Mazouni et al., 2004; Miyagishima
et al., 2005; Marbouty et al., 2009). In bacteria, MinC isan inhibitor of FtsZ polymerization and forms a com-plex with MinD. MinE in Escherichia coli (a gram-negative proteobacterium) and DivIVA in Bacillussubtilis (a gram-positive firmicute) regulate the posi-tioning of MinCD such that the FtsZ ring forms only atthe midcell position (Harry et al., 2006). The mecha-nism of the division site selection has also been mod-ified since the endosymbiotic event (Figs. 2 and 3;Tables I and II). MinC and Cdv3 have been lost,although certain algal lineages and the moss Physco-mitrella patens still possess a MinC-like protein (Yanget al., 2008; Miyagishima and Kabeya, 2010). A recentstudy suggests that ARC3 fulfills the function of MinCin Viridiplantae (Maple et al., 2007). Overexpression ofARC3 inhibits chloroplast division, most likely thor-ough an inhibition of the FtsZ assembly, like the caseof bacterial MinC. ARC3 interacts with MinD, MinE,FtsZ1 (Maple et al., 2007), and PARC6 (Glynn et al.,2009). The arc3 mutant exhibits multiple FtsZ ringswithin a single enlarged chloroplast, similar to theArabidopsis minD mutant and bacterial minC andminD mutants (Glynn et al., 2007). MULTIPLE CHLO-ROPLAST DIVISION SITE 1 (MCD1), a protein uniqueto land plants, is also involved in the positioning of theplastid FtsZ ring. MCD1 recruits MinD to the properlocation, and the mcd1 mutant exhibits multiple FtsZrings in a single chloroplast (Nakanishi et al., 2009).The Min system is widely conserved in algae and land
Table II. Eukaryotic host-derived components of the chloroplast division machinery
The references are indicated in the text.
Component
(Localization)Function and Notes
PDV1/PDV2 (medial ring) Nucleus-encoded PDV2 and PDV1 (a paralog of PDV2 unique to vascular plants) arerequired for normal chloroplast division. Both proteins are required for therecruitment of DRP5B, but it is not known whether PDV and DRP5B directly interact.Homologs of PDV proteins are only evident in land plants.
DRP5B (ARC5) (medial ring) Nucleus-encoded DRP5B is required for normal chloroplast division. DRP5B is a memberof the dynamin superfamily of eukaryotic membrane-remodeling GTPases and isunique to plastid-containing eukaryotes. The best studied member is dynamin,which is involved in vesicle budding in endocytosis. Dynamin forms helical assembliesat the neck of membrane buds, and helix disassembly upon GTP hydrolysistriggers membrane fission.
Outer PD ring (medial ring) The outer PD ring is a bundle of polyglucan filaments in the red alga C. merolae, andsimilar filaments have been observed at the chloroplast division site in several lineagesof algae and land plants. In C. merolae, the filaments are associated with the PDR1protein, which probably synthesizes the glucan strands. Formation of the outer PDring is required for the localization of DRP5B.
Inner PD ring (medial ring) The inner PD ring has been detected as a belt-like structure of approximately 5 nm thicknessby electron microscopy in several lineages of algae and land plants. The componentsare still unknown. The components should be distinct from the known plastid divisionproteins because the inner PD ring is distinct from the FtsZ ring in C. merolae, and theC. merolae genome encodes only FtsZ among known stromal components of thedivision complex.
ARC3 (medial ring and poles) Nucleus-encoded ARC3 regulates the positioning of the chloroplast FtsZ ring. ARC3 interactswith MinD, MinE, FtsZ1, and PARC6. ARC3 probably acts as an inhibitor of FtsZ ringformation, as does bacterial MinC.
MCD1 (medial ring and punctate structures) Nucleus-encoded MCD1 regulates the positioning of the chloroplast FtsZ ring. MCD1interacts with MinD.
Evolution and Regulation of the Plastid Division Machinery
Plant Physiol. Vol. 155, 2011 1537

plants, but the genome of the red alga C. merolae doesnot encode any of the proteins described above. Cur-rently, it is not known how the FtsZ ring is positionedat the midpoint in this organism.
Formation and Constriction of the Chloroplast
Division Machinery
Studies in Arabidopsis (Miyagishima et al., 2006;Glynn et al., 2008, 2009) and C. merolae (Miyagishimaet al., 2003; Yoshida et al., 2010) suggest that the plastiddivision complex is assembled from the inside to theoutside of the chloroplast before the onset of divisionsite constriction, in the order of the FtsZ ring, inner PDring, outer PD ring, and DRP5B ring (Miyagishimaet al., 2003, 2006; Glynn et al., 2008, 2009; Yoshida et al.,2010; Fig. 2). Thus, topological information is con-veyed from the stromal complex that is descendedfrom the cyanobacterial endosymbiont to the cytosoliccomplex of host eukaryotic origin.
Currently, how the division complex constricts thetwo envelopes is essentially a matter of speculation.Bacterial FtsZ self-assembles into rings inside of lipo-somes and induces constrictions of these liposomes invitro (Osawa et al., 2008). The helical self-assembly ofdynamin tabulates liposomes, and disassembly of thehelix results in membrane fission in vitro (Roux andAntonny, 2008). Thus, FtsZ and DRP5B probably par-ticipate in the generation of constrictive force in plastiddivision. However, there is a certain time lag betweenFtsZ ring formation and the onset of plastid divisionsite constriction. Constriction starts after the othercomponents of the division complex have been assem-bled in the division complex (Miyagishima et al.,2003). A similar time lag has been observed in bacterialcell division (Aarsman et al., 2005; Gamba et al., 2009).It is proposed that the sliding of the PD ring filamentsmediated by DRP5B likely produces the constrictiveforce (Yoshida et al., 2006), but such sliding of thefilaments has not been demonstrated. DRP5B formspunctate rings (Gao et al., 2003; Miyagishima et al.,2003), unlike other components of the division com-plex, which have been observed to form continuousrings by fluorescence microscopy. In Arabidopsis, theenlarged chloroplasts in drp5B (arc5) (Pyke and Leech,1994) or pdv (Miyagishima et al., 2006) mutants can stilldivide by constriction (note that DRP5A, which is mostclosely related to DRP5B, is not involved in plastiddivision as described above). Future functional studiesin vivo and in vitro at a higher resolution will ulti-mately shed light on how the division complex con-stricts double-membraned plastids.
PLASTID DIFFERENTIATION ANDPLASTID DIVISION
In algae and mosses, chloroplasts are usually theonly type of plastid present. This is consistent with thefact that the vegetative cells of cyanobacteria remainblue-green and photosynthetic throughout their life
cycle. Thus, from an evolutionary standpoint, thechloroplast is the origin of plastids.
Vascular plants have evolved a complex plastiddifferentiation system in which all of the plastids,such as chloroplasts, chromoplasts, leucoplasts, andamyloplasts, are derived from nongreen proplastids inmeristematic cells (Lopez-Juez and Pyke, 2005). Forexample, in spinach (Spinacia oleracea), the shoot apicalmeristem contains approximately 12 proplastids, thedivision of which keeps pace with cell division so thatnewly formed cells have essentially the same numberof proplastids. During leaf development, cells expandwithout cell division (but with endoreduplication), yetthe chloroplasts still continue to divide and the num-ber of chloroplasts per cell eventually reaches approx-imately 200 (Possingham and Lawrence, 1983).
Previous microscopic studies showed that all typesof plastids are capable of division (Possingham andLawrence, 1983). However, in spite of recent remark-able advances in understanding the mechanism ofchloroplast division, few studies have addressed themechanism governing the division of the nongreenplastids (Pyke, 2010). This is largely because nongreenplastids are smaller than chloroplasts and observationrequires fluorescent staining or electronmicroscopy. Inaddition, some plastids exhibit irregular shapes withnumerous tubular connections between them calledstromules (Hanson and Sattarzadeh, 2008). In somecases, multiple FtsZ rings form in a single plastid, andmultiple fission has been observed (Momoyama et al.,2003; Yun and Kawagoe, 2009; Pyke, 2010).
Observation of nongreen plastids in chloroplastdivision mutants and analyses of the localization ofthe chloroplast division proteins have shown thatnongreen plastid division utilizes division machinerysimilar to that in chloroplasts (Table III). However,these investigations have also revealed certain differ-ences between chloroplasts and the nongreen plastids(Table III). For example, there is no detectable defectin proplastid division in the drp5B (arc5) mutant(Robertson et al., 1996; Pyke, 1999), and the DRP5Bprotein is not detected in the shoot apical meristem(Okazaki et al., 2009). Thus, land plants apparentlyhave evolved a plastid division mechanism that doesnot require DRP5B protein.
In addition, there might be a plastid division mech-anism that does not utilize the known plastid divisioncomplex. Whereas the arc6 mutation impairs FtsZassembly (Vitha et al., 2003) and results in severedefects in both chloroplast and proplastid division(Robertson et al., 1995; Pyke, 1999), the photosyntheticcells in arc6 contain irregularly shaped nongreen plas-tids along with chloroplasts. These abnormal plastidssomehow proliferate in the arc6 mutant (Chen et al.,2009). A similar type of plastid proliferation alsooccurs in a tomato (Solanum lycopersicum) chloroplastdivision mutant, where budding and fragmentation ofthe plastids are observed (Forth and Pyke, 2006). Avery recent study showed that Arabidopsis mutantsin which FtsZ proteins are completely depleted are
Miyagishima
1538 Plant Physiol. Vol. 155, 2011

viable, and the mesophyll cells still contain one or twoenlarged chloroplasts (Schmitz et al., 2009). This resultsuggests that the plastids, at least in the meristematictissues, somehow proliferate without FtsZ in the mu-tant, although the FtsZ rings are always evident in theproplastid division that occurs in the wild type(McAndrew et al., 2008; Okazaki et al., 2009). Becausethese abnormal types of plastid proliferation havebeen reported only in mutants, further studies arerequired to examine the possibility of as-yet-unknownmechanisms of plastid proliferation. In this regard,the genomes of certain lineages of bacteria and archaeado not encode FtsZ. Most of these prokaryotes donot have a peptidoglycan layer. Very recent studiesshowed that the cell division in Crenarchaeota (agroup of archaea) involves the ESCRT system, whichis involved in vesicle formation in eukaryotes. Inaddition, when the peptidoglycan layer is depletedby a mutation, B. subtilis cells are able to divide even inthe absence of FtsZ (Bernander and Ettema, 2010).
Other Proteins Related to Plastid Division
Proteins other than the ones described above havealso been implicated in plastid division in Arabidop-sis. When the GIANT CHLOROPLAST 1 (GC1; alsocalled AtSulA; Maple et al., 2004; Raynaud et al., 2004),CRUMPLED LEAF (CRL; Asano et al., 2004), Msc-likegenes ([MSL]; and MSL3; Haswell and Meyerowitz,2006), AtCDT1 (Raynaud et al., 2005), or CPN60(Suzuki et al., 2009) gene is inactivated, plastid divi-sion is impaired and giant plastids are generated, as inother bona fide plastid division genes. At present, it isnot known whether these proteins are directly in-volved in chloroplast division machinery or how theseproteins are related to plastid division.CRL (Asano et al., 2004) and GC1 (Maple et al., 2004;
Raynaud et al., 2004) are descended from the cyano-bacterial endosymbiont. Some of the bacterial ortho-logs of GC1 are annotated as SulA-like proteins. SulAfunctions as an inhibitor of FtsZ assembly in E. coli(Harry et al., 2006; de Boer, 2010). However, there is noevident similarity between GC1 and E. coli SulA at the
level of primary structure and no experimental evi-dence that GC1 is a functional counterpart of SulA.MSL proteins are homologs of bacterial mechanosen-sitive ion channels and likely control plastid size andshape (Haswell and Meyerowitz, 2006). AtCDT1 (forCdc10-dependent transcript 1) are dually targeted toboth the nucleus and plastids and likely coordinatethe cell cycle as well as plastid division (Raynaudet al., 2005).
SECONDARY PLASTID DIVISION
A diverse array of eukaryotic lineages possess plas-tids (most are chloroplasts) that are of secondaryendosymbiotic origin (Fig. 1). In addition, plastids oftertiary endosymbiotic origin have been found incertain species of dinoflagellates. The secondary en-dosymbiosis of a red algal ancestor gave rise toplastids in Chromalveolata, such as stramenopiles,haptophytes, cryptophytes, most of the photosyntheticdinoflagellates, and apicomplexan parasites. Eugle-nids and chrorarachniophytes possess chloroplasts ofa green algal origin. The question of exactly howmanyendosymbiotic events have given rise to this evidentdiversity remains elusive. However, several reportshave indicated a preference for the hypothesis that thered algal endosymbiosis and subsequent reductioninto chloroplasts occurred only once in the commonancestor of chromalveolates (Reyes-Prieto et al., 2007;Gould et al., 2008; Archibald, 2009). This scenario alsosuggests that several chromalveolate lineages, such asciliates, oomycetes, and plastid-lacking lineages ofdinoflagellates, have subsequently lost the plastids.
Little is known about how secondary plastids in thediverse eukaryotic lineages divide, but a few studies ofstramenopile chloroplast division and recent genomeprojects of Chromista (stramenopiles, haptophytes,and cryptophytes) suggest that a part of the chloro-plast division machinery in these lineages is descendedfrom a red algal endosymbiont (Fig. 4). The chloroplastsin Chromista are surrounded by four membranes. Theinner twomembranes are descended from the inner and
Table III. Involvement of chloroplast division proteins in proplastid division in the shoot or rootapical meristem
The division defect is suggested by the presence of a smaller number of larger proplastids than those inthe wild type. n/d, Not determined.
Components Localization Meristematic Proplastid Division in Mutants
FtsZ1/FtsZ2 Midplastid ringa,b Division defectc
ARC6/PARC6 Midplastid ringb Division defect (arc6)d; n/d (parc6)MinE n/d Division defecte
PDV1/PDV2 Midplastid ringb,f Division defectf
DRP5B Not detectedb Slight difference from the wild typeg
Outer PD ring Midplastid ringh n/dInner PD ring Midplastid ringh n/d
aMcAndrew et al. (2008). bOkazaki et al. (2009). cOsteryoung et al. (1998). dRobertson et al.(1995). ePyke (1999; arc12). fMiyagishima et al. (2006). gRobertson et al. (1995). hKuroiwaet al. (1998).
Evolution and Regulation of the Plastid Division Machinery
Plant Physiol. Vol. 155, 2011 1539
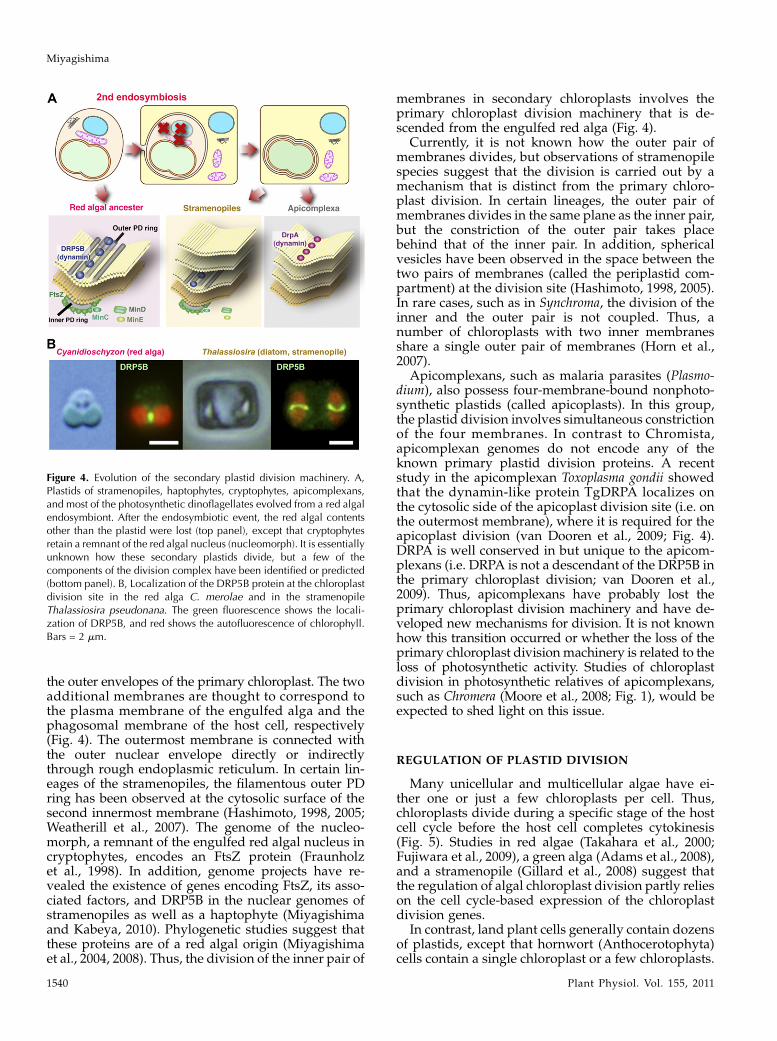
the outer envelopes of the primary chloroplast. The twoadditional membranes are thought to correspond tothe plasma membrane of the engulfed alga and thephagosomal membrane of the host cell, respectively(Fig. 4). The outermost membrane is connected withthe outer nuclear envelope directly or indirectlythrough rough endoplasmic reticulum. In certain lin-eages of the stramenopiles, the filamentous outer PDring has been observed at the cytosolic surface of thesecond innermost membrane (Hashimoto, 1998, 2005;Weatherill et al., 2007). The genome of the nucleo-morph, a remnant of the engulfed red algal nucleus incryptophytes, encodes an FtsZ protein (Fraunholzet al., 1998). In addition, genome projects have re-vealed the existence of genes encoding FtsZ, its asso-ciated factors, and DRP5B in the nuclear genomes ofstramenopiles as well as a haptophyte (Miyagishimaand Kabeya, 2010). Phylogenetic studies suggest thatthese proteins are of a red algal origin (Miyagishimaet al., 2004, 2008). Thus, the division of the inner pair of
membranes in secondary chloroplasts involves theprimary chloroplast division machinery that is de-scended from the engulfed red alga (Fig. 4).
Currently, it is not known how the outer pair ofmembranes divides, but observations of stramenopilespecies suggest that the division is carried out by amechanism that is distinct from the primary chloro-plast division. In certain lineages, the outer pair ofmembranes divides in the same plane as the inner pair,but the constriction of the outer pair takes placebehind that of the inner pair. In addition, sphericalvesicles have been observed in the space between thetwo pairs of membranes (called the periplastid com-partment) at the division site (Hashimoto, 1998, 2005).In rare cases, such as in Synchroma, the division of theinner and the outer pair is not coupled. Thus, anumber of chloroplasts with two inner membranesshare a single outer pair of membranes (Horn et al.,2007).
Apicomplexans, such as malaria parasites (Plasmo-dium), also possess four-membrane-bound nonphoto-synthetic plastids (called apicoplasts). In this group,the plastid division involves simultaneous constrictionof the four membranes. In contrast to Chromista,apicomplexan genomes do not encode any of theknown primary plastid division proteins. A recentstudy in the apicomplexan Toxoplasma gondii showedthat the dynamin-like protein TgDRPA localizes onthe cytosolic side of the apicoplast division site (i.e. onthe outermost membrane), where it is required for theapicoplast division (van Dooren et al., 2009; Fig. 4).DRPA is well conserved in but unique to the apicom-plexans (i.e. DRPA is not a descendant of the DRP5B inthe primary chloroplast division; van Dooren et al.,2009). Thus, apicomplexans have probably lost theprimary chloroplast division machinery and have de-veloped new mechanisms for division. It is not knownhow this transition occurred or whether the loss of theprimary chloroplast divisionmachinery is related to theloss of photosynthetic activity. Studies of chloroplastdivision in photosynthetic relatives of apicomplexans,such as Chromera (Moore et al., 2008; Fig. 1), would beexpected to shed light on this issue.
REGULATION OF PLASTID DIVISION
Many unicellular and multicellular algae have ei-ther one or just a few chloroplasts per cell. Thus,chloroplasts divide during a specific stage of the hostcell cycle before the host cell completes cytokinesis(Fig. 5). Studies in red algae (Takahara et al., 2000;Fujiwara et al., 2009), a green alga (Adams et al., 2008),and a stramenopile (Gillard et al., 2008) suggest thatthe regulation of algal chloroplast division partly relieson the cell cycle-based expression of the chloroplastdivision genes.
In contrast, land plant cells generally contain dozensof plastids, except that hornwort (Anthocerotophyta)cells contain a single chloroplast or a few chloroplasts.
Figure 4. Evolution of the secondary plastid division machinery. A,Plastids of stramenopiles, haptophytes, cryptophytes, apicomplexans,and most of the photosynthetic dinoflagellates evolved from a red algalendosymbiont. After the endosymbiotic event, the red algal contentsother than the plastid were lost (top panel), except that cryptophytesretain a remnant of the red algal nucleus (nucleomorph). It is essentiallyunknown how these secondary plastids divide, but a few of thecomponents of the division complex have been identified or predicted(bottom panel). B, Localization of the DRP5B protein at the chloroplastdivision site in the red alga C. merolae and in the stramenopileThalassiosira pseudonana. The green fluorescence shows the locali-zation of DRP5B, and red shows the autofluorescence of chlorophyll.Bars = 2 mm.
Miyagishima
1540 Plant Physiol. Vol. 155, 2011

In the shoot apical meristem, there likely is a correla-tion between cell cycle progression and proplastiddivision (Seguı-Simarro and Staehelin, 2009), whereasplastids generally divide nonsynchronously, even inthe same cell (e.g. chloroplasts in the moss P. patens;Fig. 2). Moreover, the plastid division rate changes inaccord with cell differentiation, resulting in changes ofplastid size and number (Fig. 5). Many earlier studieshave indicated that various environmental stimuliimpact the chloroplast division rate (Possingham andLawrence, 1983; Lopez-Juez and Pyke, 2005). Cur-rently, it is largely unknown how plastid division isregulated in land plants, but a recent study showedthat PDV proteins determine the rate of plastid divi-sion in land plants (Okazaki et al., 2009). The level ofPDV proteins in the division complex changes duringcell differentiation, where either an increase or a de-crease of the PDV1 and PDV2 levels results in acorresponding increase or decrease in the plastid divi-sion rate. For example, the PDV level is highest in theproximity of the shoot apical meristem. The level ofPDV proteins, but not other components of the divisioncomplex, decreases during leaf development, resultingin a decrease of the chloroplast division rate and anincrease in chloroplast size. It is not currently known
how PDV proteins regulate the chloroplast divisionrate, but it appears that PDV proteins regulate the rateof division site constriction (Okazaki et al., 2009).
CHLOROPLASTS IN GLAUCOPHYTES ANDINTERMEDIATES BETWEEN CYANOBACTERIAAND PLASTIDS
Unlike plastids in other organisms, the chloroplastsof glaucophyte algae (called cyanelles) have retained apeptidoglycan layer between the two envelope mem-branes that is descended from the cyanobacterial en-dosymbiont. In plastids of other lineages, the intervalbetween the inner and the outer envelopes at thedivision site is almost the same as that in other parts ofthe plastid. In contrast, the inner envelope in glauco-phytes constricts faster (accompanied by the ingrowthof the peptidoglycan layer) than the outer envelopemembrane, reminiscent of the relationship betweenthe cytoplasmic membrane and the outer membrane incyanobacterial cell division. Therefore, the intervalbetween the two envelopes at the division site be-comes much larger than that in other parts of thechloroplast in glaucophytes (Iino and Hashimoto,2003; Sato et al., 2009).
Glaucophyte chloroplast division involves FtsZ-based division machinery on the stromal side (Satoet al., 2005, 2009), and a structure similar to the innerPD ring has been detected by electron microscopy.However, the outer PD ring is not evident (Iino andHashimoto, 2003; Sato et al., 2009). In addition, all ofthe chloroplast division genes of host origin, includingdrp5B, are absent from the Cyanophora paradoxa ESTdatabase, where the ftsZ, minD, and minE genes areevident (Miyagishima and Kabeya, 2010). It appearsthat the outer envelope division in glaucophyte in-volves a mechanism that is similar to that in bacteria.However, studies on division of the peptidoglycanlayer and the outer membrane in bacteria are justbeginning (de Boer, 2010). Information from the wholeglaucophyte genome, and further studies of bacterialcell wall and outer membrane division, will ultimatelyclarify the relationship between the loss of the pepti-doglycan layer and the evolution of the plastid divi-sion machinery.
Peptidoglycans have never been detected in plastidsother than glaucophytes, but it should be noted thatsome algal and plant genomes still encode cyanobac-teria-descended proteins that are involved in or re-lated to peptidoglycan synthesis in bacteria. Disruptionof the nucleus-encoded mur or mra genes impairschloroplast division in the moss P. patens (Machidaet al., 2006; Homi et al., 2009). The chloroplast genomesof certain green algal lineages still retain ftsI and ftsWgenes. However, the functions of these gene productsin plastids are currently unknown.
Although photosynthesis in eukaryotes largelyrelies on chloroplasts, numerous eukaryotic speciescontain bacterial or eukaryotic photosynthetic endo-
Figure 5. Plastid division in algae and vascular plants. In unicellularalgae, which usually have just one or only a few chloroplasts per cell,chloroplast division occurs once per cell cycle prior to cytokinesis. Invascular plants, the rate of proplastid and cell multiplication is almostthe same in meristematic cells. After leaf cells start to differentiate,proplastids differentiate into chloroplasts. During leaf development,cell division ceases but nuclear DNA replication continues, resulting inenlarged cells with a high degree of ploidy. During leaf cell expansion,chloroplast division still continues without cell division, but thedivision rate slows down, resulting in an increase of the chloroplastsize.
Evolution and Regulation of the Plastid Division Machinery
Plant Physiol. Vol. 155, 2011 1541

symbionts other than plastids. Analyses of organismsthat have undergone more recent cyanobacterial en-dosymbiosis, such as Rhopalodia gibba (diatom, stra-menopile; Prechtl et al., 2004) and P. chromatophora(Rhizaria, Cercozoa; Melkonian and Mollenhauer,2005), should yield insights into how eukaryotes areable to acquire a permanent bacterial endosymbiont ina process reminiscent of the evolutionary origin ofplastids.
P. chromatophora contains a photosynthetic inclusionthat evolved by the endosymbiosis of a cyanobacte-rium, independently from and more recently thanplastids (Marin et al., 2005). The symbiont (called achromatophore) is permanently inherited by synchro-nization of the host and the endosymbiont cell cycle.At present, it is not known how the chromatophoredivision is controlled by the host cell. Recent studiesshowed that the chromatophore genome has alreadybeen reduced compared with free-living cyanobacte-ria, but not as much as the primary plastids (Nowacket al., 2008). The chromatophore genome still retainsmost of the known cyanobacterial cell division genes,unlike the plastid genome. The exceptions are theabsence of a cyanobacterial counterpart of GC1 andtruncation of the N-terminal portion of Ftn2 (cyano-bacterial counterpart of ARC6; Nowack et al., 2008).These or other as-yet-unknown cyanobacterial celldivision genes would be candidates for being trans-ferred to the host genome and coming to be regulatedby the host cell cycle. Recent EST analyses of the P.chromatophore nuclear genome revealed the expressionof photosynthetic genes that were probably trans-ferred from the endosymbionts (Nakayama and Ishida,2009; Nowack et al., 2011). Further characterization willelucidate how the host cell has established the control ofendosymbiont cell division in order to establish apermanent endosymbiotic relationship.
CONCLUSION AND PERSPECTIVES
Diverse eukaryotic lineages have acquired photo-synthetic activity by endosymbiotic events. Permanentinheritance of photosynthetic endosymbionts or plas-tids relies on the regulation of endosymbiont or plastiddivision by the host cell. Recent studies have yieldedsignificant progress in understanding of the mecha-nism and regulation of plastid division, but manyquestions still remain to be answered, and new ques-tions are emerging.
The most noteworthy issues concerning the mecha-nism of plastid division are how the division complexconstricts and splits chloroplasts and how the constric-tive force is generated. In terms of evolution, anotherquestion is whether there are any common trends withregard to the mechanisms that control endosymbiont ororganelle division. There likely are, because the evolu-tion of the mitochondrial division machinery is similarto that of plastids. Mitochondria evolved from ana-proteobacterial endosymbiont, and mitochondrial
division involves a dynamin-related protein. In addi-tion, mitochondrial division in some eukaryotic line-ages also involves the FtsZ protein of a-proteobacterialorigin (Kiefel et al., 2006). Further studies shouldprovide important insights not only to the under-standing of organelle division but also the mecha-nisms of membrane fission and eukaryotic evolutioneffected by endosymbiosis.
Received December 7, 2010; accepted February 2, 2011; published February
10, 2011.
LITERATURE CITED
Aarsman ME, Piette A, Fraipont C, Vinkenvleugel TM, Nguyen-Disteche
M, den Blaauwen T (2005) Maturation of the Escherichia coli divisome
occurs in two steps. Mol Microbiol 55: 1631–1645
Adams S, Maple J, Møller SG (2008) Functional conservation of the MIN
plastid division homologues of Chlamydomonas reinhardtii. Planta 227:
1199–1211
Archibald JM (2009) The puzzle of plastid evolution. Curr Biol 19: R81–R88
Asano T, Yoshioka Y, Kurei S, Sakamoto W, Machida Y, Sodmergen (2004)
A mutation of the CRUMPLED LEAF gene that encodes a protein
localized in the outer envelope membrane of plastids affects the pattern
of cell division, cell differentiation, and plastid division in Arabidopsis.
Plant J 38: 448–459
Bernander R, Ettema TJ (2010) FtsZ-less cell division in archaea and
bacteria. Curr Opin Microbiol 13: 747–752
Chen Y, Asano T, Fujiwara MT, Yoshida S, Machida Y, Yoshioka Y (2009)
Plant cells without detectable plastids are generated in the crumpled
leaf mutant of Arabidopsis thaliana. Plant Cell Physiol 50: 956–969
de Boer PA (2010) Advances in understanding E. coli cell fission. Curr Opin
Microbiol 13: 730–737
Duckett J, Ligrone R (1993) Plastid-dividing rings in fern. Ann Bot (Lond)
72: 619–627
Forth D, Pyke KA (2006) The suffulta mutation in tomato reveals a novel
method of plastid replication during fruit ripening. J Exp Bot 57:
1971–1979
Fraunholz MJ, Moerschel E, Maier UG (1998) The chloroplast division
protein FtsZ is encoded by a nucleomorph gene in cryptomonads. Mol
Gen Genet 260: 207–211
Fujiwara T, Misumi O, Tashiro K, Yoshida Y, Nishida K, Yagisawa F,
Imamura S, Yoshida M, Mori T, Tanaka K, et al (2009) Periodic gene
expression patterns during the highly synchronized cell nucleus and
organelle division cycles in the unicellular red alga Cyanidioschyzon
merolae. DNA Res 16: 59–72
Gamba P, Veening JW, Saunders NJ, Hamoen LW, Daniel RA (2009) Two-
step assembly dynamics of the Bacillus subtilis divisome. J Bacteriol 191:
4186–4194
Gao H (2005) Dynamin-related proteins involved in chloroplast divi-
sion and morphogenesis. PhD thesis. Michigan State University, East
Lansing
Gao H, Kadirjan-Kalbach D, Froehlich JE, Osteryoung KW (2003) ARC5, a
cytosolic dynamin-like protein from plants, is part of the chloroplast
division machinery. Proc Natl Acad Sci USA 100: 4328–4333
Gillard J, Devos V, Huysman MJ, De Veylder L, D’Hondt S, Martens C,
Vanormelingen P, Vannerum K, Sabbe K, Chepurnov VA, et al (2008)
Physiological and transcriptomic evidence for a close coupling between
chloroplast ontogeny and cell cycle progression in the pennate diatom
Seminavis robusta. Plant Physiol 148: 1394–1411
Glynn JM, Froehlich JE, Osteryoung KW (2008) Arabidopsis ARC6 coor-
dinates the division machineries of the inner and outer chloroplast
membranes through interaction with PDV2 in the intermembrane space.
Plant Cell 20: 2460–2470
Glynn JM, Miyagishima SY, Yoder DW, Osteryoung KW, Vitha S (2007)
Chloroplast division. Traffic 8: 451–461
Glynn JM, Yang Y, Vitha S, Schmitz AJ, Hemmes M, Miyagishima SY,
Osteryoung KW (2009) PARC6, a novel chloroplast division factor,
influences FtsZ assembly and is required for recruitment of PDV1
during chloroplast division in Arabidopsis. Plant J 59: 700–711
Miyagishima
1542 Plant Physiol. Vol. 155, 2011

Gould SB, Waller RF, McFadden GI (2008) Plastid evolution. Annu Rev
Plant Biol 59: 491–517
Hanson MR, Sattarzadeh A (2008) Dynamic morphology of plastids and
stromules in angiosperm plants. Plant Cell Environ 31: 646–657
Harry E, Monahan L, Thompson L (2006) Bacterial cell division: the
mechanism and its precison. Int Rev Cytol 253: 27–94
Hashimoto H (1998) Electron-opaque annular structure girdling the con-
stricting isthmus of the dividing chloroplasts of Heterosigma akashiwo
(Raphidophyceae, Chromophyta). Protoplasma 197: 210–216
Hashimoto H (2005) The ultrastructural features and division of secondary
plastids. J Plant Res 118: 163–172
Haswell ES, Meyerowitz EM (2006) MscS-like proteins control plastid size
and shape in Arabidopsis thaliana. Curr Biol 16: 1–11
Homi S, Takechi K, Tanidokoro K, Sato H, Takio S, Takano H (2009) The
peptidoglycan biosynthesis genes MurA and MraY are related to chlo-
roplast division in the moss Physcomitrella patens. Plant Cell Physiol 50:
2047–2056
Horn S, Ehlers K, Fritzsch G, Gil-Rodrıguez MC, Wilhelm C, Schnetter R
(2007) Synchroma grande spec. nov. (Synchromophyceae class. nov.,
Heterokontophyta): an amoeboid marine alga with unique plastid
complexes. Protist 158: 277–293
Iino M, Hashimoto H (2003) Intermediate features of cyanelle division of
Cyanophora paradoxa (Glaucocystophyta) between cyanobacterial and
plastid division. J Phycol 39: 561–569
Imanian B, Pombert JF, Keeling PJ (2010) The complete plastid genomes of
the two ‘dinotoms’ Durinskia baltica and Kryptoperidinium foliaceum.
PLoS ONE 5: e10711
Kiefel BR, Gilson PR, Beech PL (2006) Cell biology of mitochondrial
dynamics. Int Rev Cytol 254: 151–213
Kuroiwa H, Mori T, Takahara M, Miyagishima SY, Kuroiwa T (2002)
Chloroplast division machinery as revealed by immunofluorescence
and electron microscopy. Planta 215: 185–190
Kuroiwa T, Kuroiwa H, Sakai A, Takahashi H, Toda K, Itoh R (1998)
The division apparatus of plastids and mitochondria. Int Rev Cytol
181: 1–41
Kuroiwa T, Misumi O, Nishida K, Yagisawa F, Yoshida Y, Fujiwara T,
Kuroiwa H (2008) Vesicle, mitochondrial, and plastid division machin-
eries with emphasis on dynamin and electron-dense rings. Int Rev Cell
Mol Biol 271: 97–152
Lopez-Juez E, Pyke KA (2005) Plastids unleashed: their development and
their integration in plant development. Int J Dev Biol 49: 557–577
Machida M, Takechi K, Sato H, Chung SJ, Kuroiwa H, Takio S, Seki M,
Shinozaki K, Fujita T, Hasebe M, et al (2006) Genes for the peptido-
glycan synthesis pathway are essential for chloroplast division in moss.
Proc Natl Acad Sci USA 103: 6753–6758
Maple J, Aldridge C, Møller SG (2005) Plastid division is mediated by
combinatorial assembly of plastid division proteins. Plant J 43: 811–823
Maple J, Fujiwara MT, Kitahata N, Lawson T, Baker NR, Yoshida S,
Møller SG (2004) GIANT CHLOROPLAST 1 is essential for correct
plastid division in Arabidopsis. Curr Biol 14: 776–781
Maple J, Møller SG (2010) The complexity and evolution of the plastid-
division machinery. Biochem Soc Trans 38: 783–788
Maple J, Vojta L, Soll J, Møller SG (2007) ARC3 is a stromal Z-ring
accessory protein essential for plastid division. EMBO Rep 8: 293–299
Marbouty M, Saguez C, Cassier-Chauvat C, Chauvat F (2009) ZipN, an
FtsA-like orchestrator of divisome assembly in the model cyanobacte-
rium Synechocystis PCC6803. Mol Microbiol 74: 409–420
Marin B, Nowack EC, Melkonian M (2005) A plastid in the making:
evidence for a second primary endosymbiosis. Protist 156: 425–432
Mazouni K, Domain F, Cassier-Chauvat C, Chauvat F (2004) Molecular
analysis of the key cytokinetic components of cyanobacteria: FtsZ, ZipN
and MinCDE. Mol Microbiol 52: 1145–1158
McAndrew RS, Froehlich JE, Vitha S, Stokes KD, Osteryoung KW (2001)
Colocalization of plastid division proteins in the chloroplast stromal
compartment establishes a new functional relationship between FtsZ1
and FtsZ2 in higher plants. Plant Physiol 127: 1656–1666
McAndrew RS, Olson BJ, Kadirjan-Kalbach DK, Chi-Ham CL, Vitha S,
Froehlich JE, Osteryoung KW (2008) In vivo quantitative relationship
between plastid division proteins FtsZ1 and FtsZ2 and identification of
ARC6 and ARC3 in a native FtsZ complex. Biochem J 412: 367–378
Melkonian M, Mollenhauer D (2005) Robert Lauterborn (1869–1952) and
his Paulinella chromatophora. Protist 156: 253–262
Miyagishima S, Kuroiwa H, Kuroiwa T (2001) The timing and manner of
disassembly of the apparatuses for chloroplast and mitochondrial
division in the red alga Cyanidioschyzon merolae. Planta 212: 517–528
Miyagishima SY, Froehlich JE, Osteryoung KW (2006) PDV1 and PDV2
mediate recruitment of the dynamin-related protein ARC5 to the plastid
division site. Plant Cell 18: 2517–2530
Miyagishima SY, Kabeya Y (2010) Chloroplast division: squeezing the
photosynthetic captive. Curr Opin Microbiol 13: 738–746
Miyagishima SY, Kuwayama H, Urushihara H, Nakanishi H (2008)
Evolutionary linkage between eukaryotic cytokinesis and chloroplast
division by dynamin proteins. Proc Natl Acad Sci USA 105: 15202–15207
Miyagishima SY, Nishida K, Mori T, Matsuzaki M, Higashiyama T,
Kuroiwa H, Kuroiwa T (2003) A plant-specific dynamin-related protein
forms a ring at the chloroplast division site. Plant Cell 15: 655–665
Miyagishima SY, Nozaki H, Nishida K, Nishida K, Matsuzaki M,
Kuroiwa T (2004) Two types of FtsZ proteins in mitochondria and
red-lineage chloroplasts: the duplication of FtsZ is implicated in endo-
symbiosis. J Mol Evol 58: 291–303
Miyagishima SY, Wolk CP, Osteryoung KW (2005) Identification of
cyanobacterial cell division genes by comparative and mutational
analyses. Mol Microbiol 56: 126–143
Momoyama Y, Miyazawa Y, Miyagishima SY, Mori T, Misumi O, Kuroiwa
H, Tsuneyoshi K (2003) The division of pleomorphic plastids with
multiple FtsZ rings in tobacco BY-2 cells. Eur J Cell Biol 82: 323–332
Moore RB, Obornık M, Janouskovec J, Chrudimsky T, Vancova M, Green
DH, Wright SW, Davies NW, Bolch CJ, Heimann K, et al (2008) A
photosynthetic alveolate closely related to apicomplexan parasites.
Nature 451: 959–963
Mori T, Kuroiwa H, Takahara M, Miyagishima SY, Kuroiwa T (2001)
Visualization of an FtsZ ring in chloroplasts of Lilium longiflorum leaves.
Plant Cell Physiol 42: 555–559
Nakanishi H, Suzuki K, Kabeya Y, Miyagishima SY (2009) Plant-specific
protein MCD1 determines the site of chloroplast division in concert with
bacteria-derived MinD. Curr Biol 19: 151–156
Nakayama T, Ishida K (2009) Another acquisition of a primary photosyn-
thetic organelle is underway in Paulinella chromatophora. Curr Biol 19:
R284–R285
Nowack EC, Melkonian M, Glockner G (2008) Chromatophore genome
sequence of Paulinella sheds light on acquisition of photosynthesis by
eukaryotes. Curr Biol 18: 410–418
Nowack EC, Vogel H, Groth M, Grossman AR, MelkonianM, Glockner G
(2011) Endosymbiotic gene transfer and transcriptional regulation of
transferred genes in Paulinella chromatophora. Mol Biol Evol 28: 407–422
Okamoto N, Inouye I (2005) A secondary symbiosis in progress? Science
310: 287
Okazaki K, Kabeya Y, Suzuki K, Mori T, Ichikawa T, Matsui M,
Nakanishi H, Miyagishima SY (2009) The PLASTID DIVISION1 and
2 components of the chloroplast division machinery determine the rate
of chloroplast division in land plant cell differentiation. Plant Cell 21:
1769–1780
Osawa M, Anderson DE, Erickson HP (2008) Reconstitution of contractile
FtsZ rings in liposomes. Science 320: 792–794
Osteryoung KW, McAndrew RS (2001) The plastid division machine.
Annu Rev Plant Physiol Plant Mol Biol 52: 315–333
Osteryoung KW, Stokes KD, Rutherford SM, Percival AL, Lee WY (1998)
Chloroplast division in higher plants requires members of two func-
tionally divergent gene families with homology to bacterial ftsZ. Plant
Cell 10: 1991–2004
Osteryoung KW, Vierling E (1995) Conserved cell and organelle division.
Nature 376: 473–474
Possingham JV, Lawrence ME (1983) Controls to plastid division. Int Rev
Cytol 84: 1–56
Prechtl J, Kneip C, Lockhart P, Wenderoth K, Maier UG (2004) Intracel-
lular spheroid bodies of Rhopalodia gibba have nitrogen-fixing apparatus
of cyanobacterial origin. Mol Biol Evol 21: 1477–1481
Pyke KA (1999) Plastid division and development. Plant Cell 11: 549–556
Pyke KA (October 5, 2010) Plastid division. AoB PLANTS http://dx.doi.
org/10.1093/aobpla/plq016
Pyke KA, Leech RM (1994) A genetic analysis of chloroplast division and
expansion in Arabidopsis thaliana. Plant Physiol 104: 201–207
Raynaud C, Cassier-Chauvat C, Perennes C, Bergounioux C (2004) An
Arabidopsis homolog of the bacterial cell division inhibitor SulA is
involved in plastid division. Plant Cell 16: 1801–1811
Raynaud C, Perennes C, Reuzeau C, Catrice O, Brown S, Bergounioux C
Evolution and Regulation of the Plastid Division Machinery
Plant Physiol. Vol. 155, 2011 1543

(2005) Cell and plastid division are coordinated through the prerepli-
cation factor AtCDT1. Proc Natl Acad Sci USA 102: 8216–8221
Reyes-Prieto A, Weber AP, Bhattacharya D (2007) The origin and estab-
lishment of the plastid in algae and plants. Annu Rev Genet 41: 147–168
Robertson EJ, Pyke KA, Leech RM (1995) arc6, an extreme chloroplast
division mutant of Arabidopsis also alters proplastid proliferation and
morphology in shoot and root apices. J Cell Sci 108: 2937–2944
Robertson EJ, Rutherford SM, Leech RM (1996) Characterization of
chloroplast division using the Arabidopsis mutant arc5. Plant Physiol
112: 149–159
Roux A, Antonny B (2008) The long and short of membrane fission. Cell
135: 1163–1165
Sato M, Mogi Y, Nishikawa T, Miyamura S, Nagumo T, Kawano S (2009)
The dynamic surface of dividing cyanelles and ultrastructure of the
region directly below the surface in Cyanophora paradoxa. Planta 229:
781–791
Sato M, Nishikawa T, Yamazaki T, Kawano S (2005) Isolation of the plastid
FtsZ gene from Cyanophora paradoxa (Glaucocystophyceae, Glaucocys-
tophyta). Phycol Res 53: 93–96
Schmitz AJ, Glynn JM, Olson BJ, Stokes KD, Osteryoung KW (2009)
Arabidopsis FtsZ2-1 and FtsZ2-2 are functionally redundant, but FtsZ-
based plastid division is not essential for chloroplast partitioning or
plant growth and development. Mol Plant 2: 1211–1222
Seguı-Simarro JM, Staehelin LA (2009) Mitochondrial reticulation in shoot
apical meristem cells of Arabidopsis provides a mechanism for homog-
enization of mtDNA prior to gamete formation. Plant Signal Behav 4:
168–171
Shimada H, Koizumi M, Kuroki K, Mochizuki M, Fujimoto H, Ohta H,
Masuda T, Takamiya K (2004) ARC3, a chloroplast division factor, is a
chimera of prokaryotic FtsZ and part of eukaryotic phosphatidylinosi-
tol-4-phosphate 5-kinase. Plant Cell Physiol 45: 960–967
Stokes KD, Osteryoung KW (2003) Early divergence of the FtsZ1 and
FtsZ2 plastid division gene families in photosynthetic eukaryotes. Gene
320: 97–108
Strepp R, Scholz S, Kruse S, Speth V, Reski R (1998) Plant nuclear gene
knockout reveals a role in plastid division for the homolog of the
bacterial cell division protein FtsZ, an ancestral tubulin. Proc Natl Acad
Sci USA 95: 4368–4373
Sumiya N, Hirata A, Kawano S (2008) Multiple FtsZ ring formation and
reduplicated chloroplast DNA in Nannochloris bacillaris (Chlorophyta,
Trebouxiophyceae) under phosphate-enriched culture. J Phycol 44:
1476–1489
Suzuki K, Nakanishi H, Bower J, Yoder DW, Osteryoung KW, Miyagishima
SY (2009) Plastid chaperonin proteins Cpn60 alpha and Cpn60 beta
are required for plastid division in Arabidopsis thaliana. BMC Plant Biol
9: 38
Takahara M, Takahashi H, Matsunaga S, Miyagishima S, Takano H, Sakai
A, Kawano S, Kuroiwa T (2000) A putative mitochondrial ftsZ gene is
present in the unicellular primitive red alga Cyanidioschyzon merolae.
Mol Gen Genet 264: 452–460
van Dooren GG, Reiff SB, Tomova C, Meissner M, Humbel BM, Striepen
B (2009) A novel dynamin-related protein has been recruited for
apicoplast fission in Toxoplasma gondii. Curr Biol 19: 267–276
Vitha S, Froehlich JE, Koksharova O, Pyke KA, van Erp H, Osteryoung
KW (2003) ARC6 is a J-domain plastid division protein and an evolu-
tionary descendant of the cyanobacterial cell division protein Ftn2.
Plant Cell 15: 1918–1933
Vitha S, McAndrew RS, Osteryoung KW (2001) FtsZ ring formation at the
chloroplast division site in plants. J Cell Biol 153: 111–120
Weatherill K, Lambiris I, Pickett-Heaps JD, Deane JA, Beech PL (2007)
Plastid division in Mallomonas (Synurophyceae, Heterokonta). J Phycol
43: 535–541
Wouters J, Raven JA, Minnhagen S, Janson S (2009) The luggage hypoth-
esis: comparisons of two phototrophic hosts with nitrogen-fixing
cyanobacteria and implications for analogous life strategies for
kleptoplastids/secondary symbiosis in dinoflagellates. Symbiosis 49:
61–70
Yang Y, Glynn JM, Olson BJ, Schmitz AJ, Osteryoung KW (2008) Plastid
division: across time and space. Curr Opin Plant Biol 11: 577–584
Yoshida Y, Kuroiwa H, Misumi O, Nishida K, Yagisawa F, Fujiwara T,
Nanamiya H, Kawamura F, Kuroiwa T (2006) Isolated chloroplast
division machinery can actively constrict after stretching. Science 313:
1435–1438
Yoshida Y, Kuroiwa H, Misumi O, Yoshida M, Ohnuma M, Fujiwara T,
Yagisawa F, Hirooka S, Imoto Y, Matsushita K, et al (2010) Chloroplasts
divide by contraction of a bundle of nanofilaments consisting of
polyglucan. Science 329: 949–953
Yun MS, Kawagoe Y (2009) Amyloplast division progresses simulta-
neously at multiple sites in the endosperm of rice. Plant Cell Physiol
50: 1617–1626
Miyagishima
1544 Plant Physiol. Vol. 155, 2011