Embed Size (px)
Citation preview

PertanikaJ. Trop. Agric. Sci. 27(1): 1 - 10 (2004) ISSN: 1511-3701© Universiti Putra Malaysia Press
Mechanism of Paraquat Resistance in Crassocephalum crepidioides(Benth.) S. Moore During Immature Stage
ISMAIL B. S., 'CHUAH T. S. & KHATIJAH H. H.School of Environmental and Natural Resource Sciences,
Faculty of Science and Technology, Universiti Kebangsaan Malaysia,43600 UKM, Bangi, Selangor, Malaysia
faculty of Agrotechnology and Food Science,Kolej Universiti Sains dan Teknobgi Malaysia,
Mengabang Telipot, 21030 Kuala Terengganu, Terengganu, Malaysia
Keywords: Resistance, herbicide, cuticle, epicuticular wax, Crassocephalum crepidioides
ABSTRAK
Mekanisme kerintangan Crassocephalum crepidioides terhadap parakuat pada peringkat 6 helm daundikaji. Parakuat yang diekstrak daripada tisu daun biotip rintang dan rentan tidak mengalami metabolisme.Maka perbezaan metabolisme tidak memainkan peranan dalam mekanisme kerintangan. Biotip rentan menyeraplebih 44% 14C-parakuat berbanding biotip rintang. Namun, lebih daripada 98% MGparakuat yangdiserap olehbiotip rintang dan rentan berada pada daun yang telah diberikan perlakuan. Perbezaan penyerapan adalahberkolerasi dengan kuantiti lilin epikutikular dan kuantiti membran kutikel pada permukaan daun. Hasilkajian ini mencadangkan bahawa perbezaan penyerapan mungkin merupakan satu faktor yang menyebabkankerintangan spesies rumpai ini terhadap parakuat.
ABSTRACT
The mechanism of paraquat resistance in Crassocephalum crepidioides at the six-leaf stage was investigated.The extractabk paraquat was not metabolized by the leaf tissue in the resistant (R) and susceptible (S) biotypes.Therefore, differential metabolism does not appear to play a role in the mechanism of resistance. The S biotypeabsorbed 44% more HCfaraquat than the R biotype. However, more than 98% of the absorbed HC-paraquatremained on the treated leaf of both biotypes. The difference in absorption had a negative correlation with theamount of epicuticular wax as well as the cuticle of leaf surfaces in both biotypes. The results of this study suggestthat differential absorption may be a factor that accounts for resistance to paraquat at the six-leaf stage.
INTRODUCTIONParaquat is a fast acting, contact herbicide(McEwen and Stepheson 1979) with no activityin the soil due to strong adsorption to soilparticles (Kokana and Aylmore 1993). Paraquatis used for controlling a wide range of broad-leaved weeds, sedges and grasses in the rice andvegetable agro-ecosystem for weed clearancebefore embarking on a new growing season.After the first case of a paraquat-resistant weed(Conyza bonariensis (L.) Cronq.) was reported inEgypt in 1979 (Summer 1980), resistance to theherbicide paraquat has been documented for 25weed species worldwide due to extensive andconsecutive use of the herbicide (Heap 1997).However, Chun and Kim have found that
Rehmannia glutinosa (Gaertn.) Libosch. ex Fisch.& Mey., a perennial herb belonging to the familyScrophulariaceae, is tolerant to paraquatalthough this plant has never been sprayed withparaquat (Chun and Kim 1992).
Fuerst and Vaughn (1990) have suggestedseveral hypotheses on the mechanism of paraquatresistance. These include reduced cuticularpenetration of paraquat due to differentialstructure and/or properties of leaf surface,differential metabolism of paraquat, altered siteof action, paraquat sequestration from the siteof action and enzymatic detoxification ofparaquat-generated radical oxygen species. Someof these mechanisms contribute to paraquatresistance in weeds such as Hordeum glaucum Steud.

ISMAIL B. S., CHUAH T. S. & KHATIJAH H. H.
(Bishop et al 1987), Conyza bonariensis (L.) Cronq.(Norman et al. 1994) and Erigeron canadensis (L.)Cronq. (Tanaka et al 1986).
A paraquat-resistant (R) biotype of theannual broadleaf weed Crassocephalum crepidioideswas first reported in 1990 in tomato fields nearTanah Rata, Cameron Highlands, Malaysia (Itohet al. 1990). Crassocephalum crepidioides is alsoknown as redflower ragleaf, a common weed intomato and potato fields in Cameron Highlands.Paraquat had been applied twice a year at aconcentration of 1 kg a.i. ha'1 to these fields forten years. Preliminary studies had shown thatthe level of resistance in the R biotype of C.crepidioides was 100-fold higher than thesusceptible (S) biotype through leaf disc testsinvolving various concentrations of paraquat(Itoh et al 1990). It has been reported thatdifferential translocation was the factorcontributing to the mechanism of resistance inmature plants of C. crepidioides (Ismail et al.2001). The objective of this study was to comparethe mechanism of resistance between the R andS biotypes of C. crepidioides at the six-leaf stage.
MATERIALS AND METHODS
Plant Materials
Seeds of the R and S biotypes were collectedfrom the Research Station of the MalaysianAgricultural Research and Development Institute(MARDI) and from farmers' fields in Brinchang,Cameron Highlands, Pahang, Malaysia,respectively. The two populations had beencharacterized as R and S to paraquat. The seedswere used to establish the six -leaf stage seedlingsfor the experiments.
Seeds of both biotypes were germinatedseparately in 36 x 26 x 5 cm plastic trayscontaining moist sand. After two weeks, seedlingsof each biotype were transplanted into 7 cmdiameter pots containing sand and grown in thegreenhouse at 29±4°C, with a 12-h photoperiodand a light intensity of 800 mE m2 sec1. Plantsin each pot were watered twice daily and fertilizedwith 5 ml of half strength Hoagland's nutrientsolution three times weekly (Hoagland andArnon 1950).
Herbicide
The herbicide used in the study was paraquatdichloride (Gramoxone) containing 200 g activeingredient (a.i) per litre aqueous solution.Radiolabelled paraquat dichloride ([14C] methyl;
specific activity 1.74 x 103 kBq mg **) waspurchased from Sigma Chemical Co., USA.
Metabolism Study
Four plants of the R and S biotypes at the six-leaf stage were sprayed with nonlabelled paraquatdichloride at 0.50 kg a.i.ha1 using a backpacksprayer that delivered 450 litre ha"1 spray volumeat 100 kPa pressure. The plants were then treatedwith 10 |il (1.48 kBq) radiolabeled paraquat inthe central portion of the adaxial surface of thethird leaf from the plant base. Control vialscontained 14C-paraquat with no plant tissue. Thetreated plants and the control vials were placedin a growth chamber at 25°C in darkness at 65%relative humidity for 12 h. The treated leaf wasthen excised and the cut petiole end wassubmerged into a 20-ml vial containing distilledwater and then placed in the growth chambertogether with the control vial for 12 h at 30°Cand light intensity of 50 jiE m2 sec*1.
After incubation, the surface herbicideresidue was removed by rinsing with 5 ml ofdistilled water. The radioactive content of therinse was measured by liquid scintillationspectrometry (LSS) using 10 ml of the aqueous-based cocktail NBCS 104 (Amersham). The tissuewas weighed and frozen at -30 °C until extraction.The frozen tissue was then pulverized with amortar and pestle using liquid nitrogen.Radiolabelled paraquat was extracted from thetissue with 5 N H2SO4 using the fresh weight tovolume ratio of 1:4 and centrifuged at 12,000 gfor 5 min (Chun et al 1997). The resultingsupernatant was concentrated with a rotaryevaporator at 30°C and analyzed for paraquatand potential metabolites by thin-layerchromatography (TLC) in 5 M NH4C1 developingsystem. The TLC plates used were 20 x 20 cm,precoated with silica gel 60 F-254 plates (Merck).Following development, the TLC plate was air-dried at 25°C and placed on X-ray film (Kodak)at -30°C for one month (Craft and Yamaguchi1964). After the autoradiography study, the TLCplate was separated into 1.5-cm segments fromthe original to the solvent front, scraped and theradioactivity in each segment was quantified byliquid scintillation spectrometry (LSS) using 1ml of distilled water and 10 ml of the aqueous-based cocktail NBCS 104 (Amersham). Theaverage of counting efficiency was more than90%.
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004

MECHANISM OF PARAQUAT RESISTANCE IN CHASSOCEPHALUM CREPIDIOIDES
Uptake and Translocation StudiesPlants from the two biotypes were sprayed withnon-labelled paraquat dichloride at 0.50 kg a.i.ha1
and treated with 10 ml (1.48 kBq) 14C- paraquatdichloride on the third leaf as describedpreviously. The plants were then kept in a growthchamber at 25°C in darkness and 65% relativehumidity. The treated leaf was excised at differentintervals viz. 0.75, 1.5, 3, 6, 9 and 12 h afterincubation. The surface herbicide residue wasremoved with a 5 ml rinse of distilled water andthe radioactive content of the rinse was measuredby LSS as described. The excised leaf was thencut into small segments (15-30 mg) and eachpiece of the segments placed into a pre-weighed20-ml vial containing 250 (xl distilled water, 1 mlNSCII tissue solubilizer (Amersham), 30 |ll glacialacetic acid and 300 ml 30% H2O2 and incubatedfor 10 h in a water bath at 50°C (Bishop et al.1987). The vial was cooled at room temperatureand 10 ml of the cocktail NBCS 204 was added.It was then kept in the dark for three daysbefore the radioactivity was assessed using LSS.The absorbed paraquat was caculated from theleaf digest and expressed as ng mg"1 fresh weightof leaf tissue.
For the translocation study, plants from bothbiotypes were sprayed with nonlabelled paraquatdichloride and treated with 14C-labelled paraquatdichloride on the third leaf as previouslydescribed. Plants were then placed in the growthchamber at 25/30°C night/day temperature and65% relative humidity. The photoperiod was setat 12 h with a light intensity of 50 |iE m"8 sec1.After 24 h incubation, the surface herbicideresidue of the treated leaf was removed and theradioactive content of the rinse was determinedas before. The whole plant was then exposed toan X-ray film at -30°C for two weeks. After theautoradiography study, the plant was sectionedinto three parts: treated leaf, shoot above treatedleaf and shoot below treated leaf (includingroot). The leaf tissue and stem were cut intosmall segments (15-30 mg) and placed into apreweighed 20-ml vial containing 250 ml distilledwater, 1 ml NSCII tissue solubilizer (Amersham),30 ml glacial acetic acid and 300 ml 30% H^O2
and incubated for 10 h in a water bath at 50°Cas above. The radioactive content for each partof the plant was quantified using LSS as describedbefore. Translocation of the compound wasexpressed as a percentage of the total amountabsorbed.
Leaf Surface Characteristics Studies
a) Cuticle assayThe third leaves were excised from plants ofboth biotypes without paraquat application andthe total leaf area was measured using a leaf areameter. In order to remove epicuticular wax, theleaves of each biotype were individually submergedin 20 ml chloroform for 5 min. The chloroformwas then filtered through Whatman No. 1 filterpaper into a pre-weighed, acid-washed and oven-dried beaker. Following the evaporation ofchloroform in a fume hood, the wax-containingbeaker was allowed to equilibrate to roomtemperature in a desiccator before being weighedfor determining the amount of epicuticular wax(jig cm*). After the surface wax removal, thecuticle was isolated by digestion of the remainingparts of the leaf tissues in a solution consistingof zinc chloride and hydrochloric acid in a ratioof 2:3 at room temperature for 24 h (Hollowayand Baker 1968). Following digestion, the cuticlewas extracted from the remaining tissues with100 ml of methanol. The final methanol rinse(containing cuticular components) was decantedinto a pre-weighed, acid-washed and oven-driedbeaker. The beaker was dried in a fume hood atroom temperature (27QC) and placed in adesiccator prior to „ being weighed fordetermination of cuticle (|Ug cm2).
b) Type of stomata, trichome and stomata densitydeterminationThe third leaves were excised from untreatedplants of the R and S biotypes and fixedindividually in a mixture of glacial acetic acidand 70% ethanol (1:3) for 48 h. From thecentral portion of each leaf blade, a smallsegment (1.5 x 1.5 cm) was cut, submerged inbasic fuschin stain (containing 10% KOH) andplaced in an oven at 60°C for one week. The leafsegments were stained in Alcian Green anddehydrated in an ethanol series (50-100%, at 5min intervals). After dehydration, the specimenswere mounted in Euparal. Observations weremade on the type of stomata, trichome density(no. cm2) and stomatal density of the adaxialsurface (no. mm"2) using a light microscope(Leitz Diaplan) at lOOx magnification.
c) Trichome and epicuticular wax structuresThe third leaves without paraquat applicationwere excised from five plants of the R and Sbiotypes. The central portion of the leaf blades
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004

ISMAIL B. S., CHUAH T. S. & KHATIJAH H. H.
was cut into small segments (0.5 x 0.5 cm)before fixation in 2% glutaraldehyde for 48 h.Samples were dehydrated in an ethanol series(50-100%, at 5 min intervals), critical-point dried,mounted on aluminium stubs and coated withgold. The structure of trichomes and epicuticularwax of the adaxial surface was observed under ascanning electron microscope (Phillips XL 30).
Statistical Analysis
Each of the above experiments was performedusing a randomized complete block design withfour replications except for trichome andepicuticular wax structure studies that had threereplications. All data were subjected to ANOVAand the means were compared with the t-test at5% level of significance. A correlation test wasundertaken to show the relationship betweenabsorption and leaf surface characteristics inboth biotypes.
RESULTS AND DISCUSSION
Metabolism Study
The extraction of paraquat residues from leaftissues of both biotypes was limited; radioactivityrecovered in H2SO4 extract accounted forapproximately 50 % of the absorbed radioactivityby both C. crepidioides biotypes whilst the rest of14C was unextractable. It is possible that theunextractable radiolabel may have covalentlybonded to the leaf tissue (Fuerst and Vaughn1990). The average of total recovery of theradiolabel, however, was more than 90% of thatapplied.
Autoradiographs of the standard 14C-paraquat, together with the R and S plant leafextracts indicated a single spot of radioactivity(Rj. = 0.00-0.20). However, radioactivity recoveredfrom acid extracts was not only detected at Rjvalues of 0.00-0.20 (98.5-99%), but a smallamount of radioactivity also appeared at Rf valuesof 0.20-0.30 and 0.70-0.90 after the TLC platewas quantified using LSS (Table 1). This couldbe due to the lower sensitivity of the X-ray filmto radioactivity as compared to LSS. The largeamount (98.5-99%) of radioactivity recoveredwith Rf values of 0.00-0.20 in the TLC platecorresponded to authentic paraquat while therest of the radioactivity at Rf values of 0.20-0.30and 0.70-0.90 were unidentified compounds(Slade 1966). These results suggested thatextractable paraquat was not metabolized in the
leaf tissues of both biotypes and the differentialmetabolism could not account for the resistancemechanism of C crepidioides to paraquat asreported in paraquat-resistant weeds like Conyzabonariensis (L.) Cronq. (Norman et ai 1993) andcultivars like Lolium perenne L. (Harvey et aL1978).
Uptake and Translocation Studies
Differential translocation may be due to thedifference in degradation at the site of absorptionyielding some metabolites that could betranslocated and other metabolites that couldnot be translocated. However, the results fromthe metabolism study showed that 94-99% of theradioactivity recovered from treated leaf tissuecorresponded to authentic paraquat (Table 1).Hence, radioactivity observed in translocation aswell as absorption studies can be assumed asauthentic paraquat.
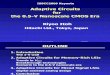
Fig. 1 shows the absorbed I4C-paraquat intreated leaves of both biotypes at the six-leafstage. There was significant difference in itsuptake by the R and S biotypes at each timeinterval. At first, the uptake rate of the S biotypewas slower compared to the R biotype. However,the uptake of labelled paraquat by the S biotypeincreased 3 h after treatment (HAT) andovertook the R biotype approximately 4 HAT.Subsequently, the total uptake by both biotypesplateaued at 6 HAT, indicating that absorptionhad reached a maximum'. There are severalbarriers to herbicide absorption into the leaftissue. The first barrier is the trichome, followedby the epicuticular wax and the cuticle. Thesusceptible biotype absorbed less paraquatcompared to the R biotype 3 HAT because it wasinhibited by higher trichome density relative tothe R biotype (Table 3). Once paraquat passedthrough the trichomes, the uptake in the Sbiotype increased and overtook the R biotypeapproximately at 4 HAT due to a lower amountof epicuticular wax as well as cuticle comparedto the R biotype (Table 3). The S biotypeabsorbed 44% more 14C-paraquat than the Rbiotype at 6 HAT, suggesting lower total uptakein the R biotype as a factor contributing toparaquat resistance. This result is in agreementwith a previous report on Erigeron philadelphicusand Ei canadensis (Tanaka et al. 1986).
However, differential translocation does notappear to play a role in the mechanism ofresistance because most of the absorbed 14C-
PERTANIKAJ. TROP, AGRIC. SCI. VOL. 27 NO. 1, 2004
MECHANISM OF PARAQUAT RESISTANCE IN CRASSOCEPHALUM CREPIDIOIDES
TABLE 1TLC analyses of radioactivity in acid extracts from leaf tissue of resistant and
susceptible biotypes treated with [14C] -paraquat dichloride for24 h at six-leaf stage
Rf value
Standard 14OparaquatRS
Distribution
0.00-0.20
94 (0.5)98.5 (0.4)99 (0.3)
(Rt) of radioactivity (*
0.20-030
6 (0.4)
Jo of recovery)
0.70-0.90
*_1.5 (0.1)1 (0.1)
SE of means are given in parentheses.* Not detected.
0 2 4 6 8Penod of uptake (hours)
Fig.l: Absorption of }4Cj)araquat by leaves of resistant (_) and .susceptible (....) biotypes at six leaf stage
TABLE 2Distribution of total 14C-paraquat recovered in plant leaf in resistant
and susceptible biotypes 24 HAT at six-leaf stage
14C-Paraquat inplant sections
RS
Distribution
Treated leaf
98.99 a98.72 a
of 14C-paraquat
Above treated
0.78 aLOO a
(% of absorbed)
leaf Below treated leaf
0.23 a0.27 a
Mean within columns with similar letters are not significant at the 5% level by T-test.
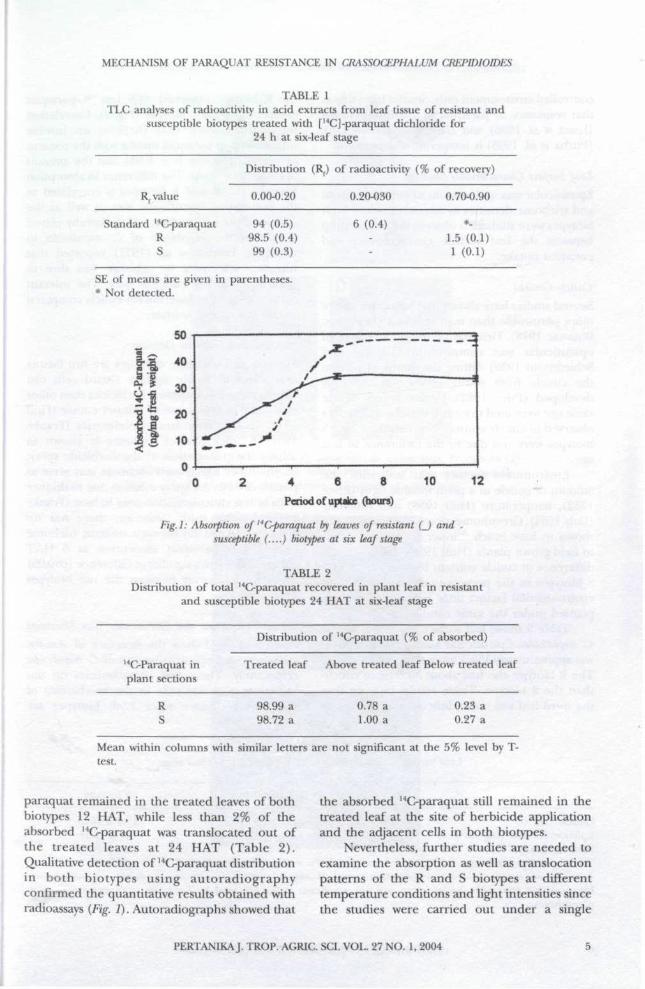
paraquat remained in the treated leaves of bothbiotypes 12 HAT, while less than 2% of theabsorbed 14C-paraquat was translocated out ofthe treated leaves at 24 HAT (Table 2).Qualitative detection of 14C-paraquat distributionin both biotypes using autoradiographyconfirmed the quantitative results obtained withradioassays (Fig. 1). Autoradiographs showed that
the absorbed 14C-paraquat still remained in thetreated leaf at the site of herbicide applicationand the adjacent cells in both biotypes.
Nevertheless, further studies are needed toexamine the absorption as well as translocationpatterns of the R and S biotypes at differenttemperature conditions and light intensities sincethe studies were carried out under a single
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004

ISMAIL B. S., CHUAH T. S. 8c KHATIJAH H. H.
controlled environment only. Studies have shownthat resistance to paraquat in Hordeum glaucum(Lasat et al 1996) and Hordeum leporinum Link(Purba et al 1995) is temperature-dependent.
Leaf Surface Characteristics StudiesEpicuticular wax and amount of cuticle, stomataand trichome densities in adaxial leaves of bothbiotypes were studied to observe the relationshipbetween the leaf surface characteristics andparaquat uptake.
Cuticle Content
Several studies have shown that young leaves aremore permeable than mature leaves (Leon andBukovac 1978). This may be due to a change inepicuticular wax composition (Loomis andSchiefertein 1959) during development or thatthe cuticle from young leaves was not fullydeveloped (Price 1982). Hence, leaves of thesame age were used to ensure that the differencesobserved in cuticle content between the R and Sbiotypes were not due to the difference in leafage.
Environment factors that influence theamount of cuticle in a plant include light (Price1982), temperature (Hull 1958) and humidity(Daly 1964). Greenhouse-grown plants have beenshown to have much thinner cuticle comparedto field-grown plants (Hull 1958). However, thedifference in cuticle content between the R andS biotypes in the present study was not due toenvironmental factors since both biotypes wereplanted under the same conditions.
Table 3 shows leaf surface characteristics ofC. crepidioides. Epicuticular wax of the R biotypewas approximately 150% more than the S biotype.The R biotype also had about 50% more cuticlethan the S biotype. These results indicate thatthe third leaf was more difficult to penetrate as
the R biotype absorbed 44% less 14C-paraquatcompared to the S biotype (Fig. 1). Correlationtests also showed that there is an inverserelationship in paraquat uptake with the contentof epicuticular wax (r = -0.95) and the amountof cuticle (r = -0.94). The difference in absorptionbetween the R and S biotypes is correlated tothe amount of epicuticular wax as well as thecuticle of the leaf surface, which thereby playeda role in the resistance of C. crepidioides toparaquat. Pereira et al. (1971) reported thatnitrofen selectivity in cabbage was due todifferences in the cuticular wax. The tolerantcultivar seemed to have thicker cuticle comparedto the susceptible cultivar.
Stomata and Trichome DensitiesStomata and trichome densities are two factorsthat affect herbicide uptake. Guard cells canabsorb more foliar-applied herbicides than otherepidermal cells because of thinner cuticle (Hull1970) and/or abundant ecotodesmata (Franke1961). A thick mat of trichome is known toinhibit the penetration of the herbicide spray;on the other hand, each trichome may serve asportals of entry for spray solution due to thinnerwalls or less cuticularization near its base (Franke1967). In this study, however, there was nocorrelation (p>0.05) between stomata, trichomedensity and paraquat absorption at 6 HATalthough there was significant difference (p>0.05)in trichome density between the two biotypes(Table 3).
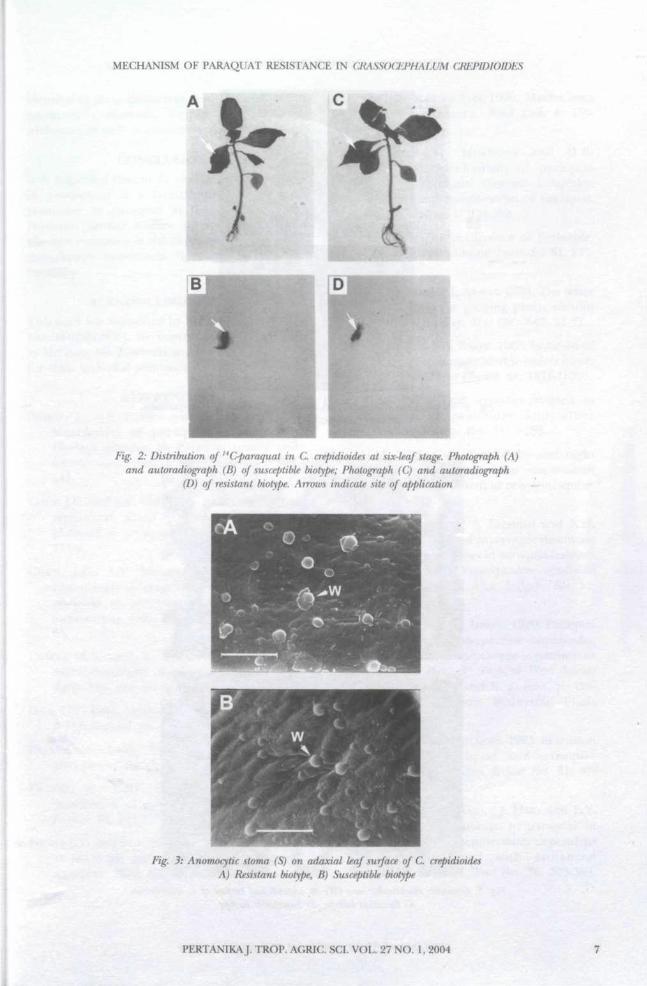
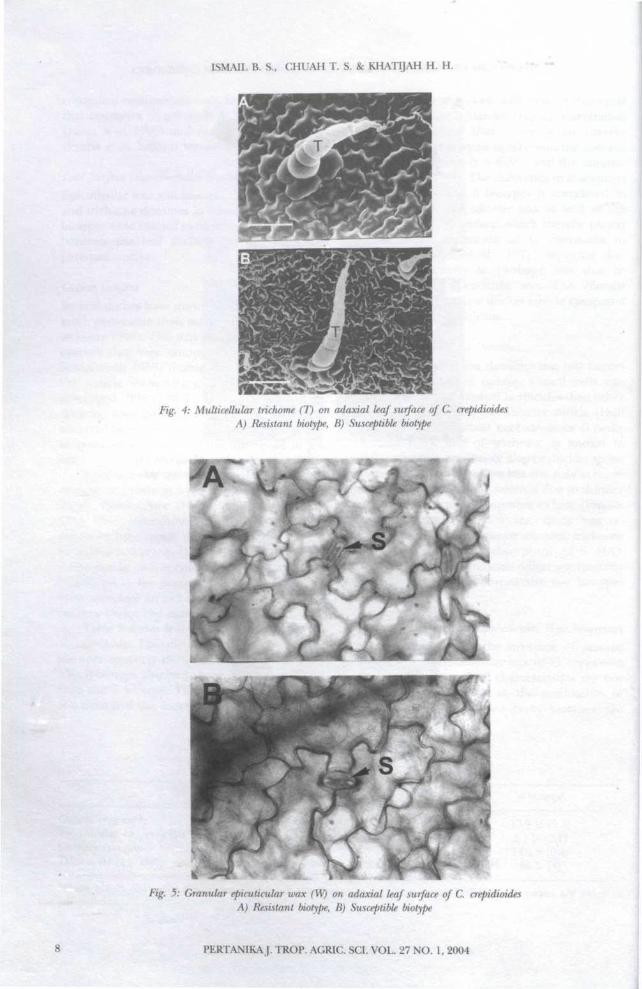
Stomata, Trichome and Epicuticular Wax StructuresFigs. 3y 4 and 5 show the structure of stomata,trichomes and epicuticular wax of C. crepidioidesrespectively. These leaf characteristics do notappear to play any role in the mechanism ofparaquat resistance since both biotypes are
TABLE 3Leaf surface characteristics of G crepidioides at six-leaf stage
R biotype S biotype
Cuticle (mg cm2)Epicuticular wax (mg cm2)Stomata (no.mnr2)Trichomes (no. cm2)
27.8 a (0.4)5.3 a (0.2)13.6 a (0.7)
22 a (5)
17.9 b (0.3)2.1 b (0.1)14.6 a (0.8)
46 h (4)
Means within rows with similar letters are not significant at the 5% level by T-test. SE of means are given inparentheses.
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004
MECHANISM OF PARAQUAT RESISTANCE IN CliASSOCMPHALUM CliKI'IDIOIDES
B
Fig. 2: Distribution of NC-paraquat in C. crepidioides at six-leaf stage. Photograph (A)and autoradiograph (B) of susceptible biotype; Photograph (C) and autoradiograph
(D) of resistant biotype. Arrows indicate site of application
Fig. 3: Anomocytic stoma (S) on adaxial leaf surface of C. crepidioidesA) Resistant biotype, B) Susceptible biotype
PERTANIKAJ. TROP. AGRIC SCI. VOL. 27 NO. 1, 2004
ISMAIL B. S., CHUAH T. S. 8c KHATIJAH H. H.
Fig. 4: Multicellular trichome (T) on adaxial leaf surface of C. crepidioidesA) Resistant biotype, B) Susceptible biotype
Fig. 5: Granular epicuticular wax (W) on adaxial leaf surface of C. crepidioidesA) Resistant biotype, B) Susceptible biotype
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004

MECHANISM OF PARAQUAT RESISTANCE IN CRASSOCblPHALUM CmPIDIOIDES
identical in these characters. Both biotypes haveanomocytic stomata, simple mutlicellulartrichomes as well as granular epicuticular wax.
CONCLUSION
It is suggested that in C crepidioides, differencein absorption is a contributing factor forresistance to paraquat at the six-leaf stage.However, further studies are required to seewhether resistance is still expressed in changingtemperature conditions, light intensities andhumidity.
ACKNOWLEDGEMENTS
This work was supported by IRPA research grantNo. 08-02-02-0004. We express our appreciationto Mr. Aziz, Ms. Normala and Mr. Cha Thye Sanfor their technical assistance.
REFERENCES
BISHOP, T., S.B. POWELS and G. CORNIC. 1987.
Mechanism of paraquat resistance inHordeum glaucum. II. Paraquat uptake andtranslocation. Aust. J. Plant Physiol 14: 539-547.
CHUN, J.C. and S.E. KIM. 1992. Resistance of themedicinal plant jiwhang {Rehmanniaglutinosa) to paraquat. Korean], Weed Sri. 12:374-9.
CHUN, J . C , S.Y. MA and H J . LEE. 1997.
Physiological responses of Rehmanniaglutinosa to paraquat and its tolerancemechanisms. Pestic. Biochem. Physiol 59: 51-63.
CRAFTS, A.S. and S. YAMAGUCHI. 1964. The
Autoradiography of plant materials. Calif,Agric. Exp. Stn. Ser. Manual 35: 143-148.
DALY, G.T. 1964. Leaf-surface wax in Poa colensoi./ Experimental Bot. 15:160-165.
FRANKE, W. 1961. Ectodesmata and foliarabsorption. Am. J. Bot. 44: 683-91.
FRANKE, W. 1967. Mechanisms of foliarpenetration of solutions. Annu. Rev. PlantPhysiol. 18: 281-300.
FUERST E.P. and M.A. NORMAN. 199L Interactionof herbicide with photosynthetic electrontransport. Weed. Sri. 39: 458-464.
FUERST, E.P. and KX. VAUGHN. 1990. Mechanismsof paraquat resistance. Weed Tech. 4: 150-156.
HARVEY, B.M.R., J. MULDOON and D.B.HARPER. 1978. Mechanism of paraquattolerance in prennial ryegrass. I. uptake,metabolism and translocation of paraquat.Plant Cell Environ. 1: 203-209.
HEAP, I.M. 1997. The occurrence of herbicide-resistant weeds worldwide Pestic. Sri. 51: 235-243.
HOAGLAND, D.R. and D.I. ARNON. 1950. The waterculture method for growing plants withoutsoil. Calif. Agric. Exp. Stn. dr. 347: 32-37.
HOLLOWAY, PJ. and E.A. BAKER. 1968. Isolation ofplant cuticles with zinc chloride-hydrochloricacid solution. Plant Physiol. 43: 1878-1879.
HULL., H.M. 1970. Leaf structure related toabsorption of pesticides and othercompounds. Residue Rev. 31: 1-155.
HULL, H.M. 1958. The effect of day and nighttemperature on growth, foliar wax contentand cuticle development of velvet mesquite.Weed 6:133-142.
ISMAIL, B.S., T.S. CHUAH, S. SALMIJAH and K.H.
KHATIJAH. 2001. Role of superoxide dismutaseand peroxidase activities in paraquat-resistantredflower ragleaf (Crassocephalum crepidioides(Benth.) S. Moore). Aust. J. Agric. Res. 52:583-586.
ITOH, K., M, AZMI and A. AHMAD. 1990. Paraquat
resistance in Crassocephalum crepidioidesy
Amaranthus lividus and Conyza sumatrensis inMalaysia. In Proc. 3rd Tropical Weed SrienceConf ed. S. A. Lee and K. F. Kon, p. 489-493. Kuala Lumpur: Malaysian PlantProtection Society.
KOKANA, R.S. and L.A.G. AYLMORE. 1993. Retensionand release of diquat and paraquatherbicides in soils. Aus. J. Soil Res. 31: 97-109.
LASAT, M.M., J.M. DITOMASO, J J. HART and L.V.
KOCHIAN. 1996. Resistance to paraquat inHordeum glaucum is temperature dependentand not associated with enhancedapoplasmic binding. Weed Res 36: 303-309.
PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004

ISMAIL B. S., CHUAH T. S. & KHATIJAH H. H.
LEON, J.M. and MJ. BUKOVAC. 1978. Cuticledevelopment and surface morphology ofolive leaves with reference to penetration offoliar-applied chemical. / Am. Soc. Hortic.ScL 103: 465-472.
LOOMIS, W.E. and R.H. SCHIEFERTEIN. 1959. Growthand differentiation of epidermal wall. Proc.9th Int. BoL Congr., p. 235-236 at Montreal.
MCEWEN, L. and G.R. STEPHENSON. 1979. The Use
and Significance of Pesticide in the Environment.Toronto: John Wiley and Sons. p. 94-95.
NORMAN, M.A., E.P. FUERST, R.J. SMEDA and K.C.
VAUGHN. 1993. Evaluation of paraquatresistance mechanisms in Conyza. Pestic.Biochem. Physiol. 46: 236-249.
NORMAN, M.A., R.J. SMEDA, K.C. VAUGHN and E.P.
FUERST. 1994. Differential movement ofparaquat in resistant and sensitive biotypesof Conyza. Pestic. Biochem. Physiol. 50: 31-42.
PEREIRA,J.F., W.E. SPLITTSTOESSER and HJ. HOPEN.
1971. Mechanism of intraspecific selectivity
of cabbage to nitrofen. Weed Sci. 19: 647-651.
PRICE, C.E. 1982. A review of factor influencingthe penetration of pesticides through plantleaves. In The Plant Cuticle ed. D.F. Cultler,K.L. Alvin and C.E. Price p. 237-252.London: Academic press.
PURBA, E., C. PRESTON and S. B. POWI.ES. 1995.
The mechanism of resistance to paraquat isstrongly temperature dependent in resistantHordeum leporinum Link and H. glaucumSteud. Planta. 196: 464-468.
SIADE, P. 1966. The fate of paraquat applied toplants. Weed Res. 6:158-167.
Si \IMER, L.A. 1980. The Bipyridinium Herbicide.London: Academic Press.
TANAKA, Y.H., H. CHISAKA and H. SAKA. 1986.
Movement of paraquat in resistant andsusceptible biotypes o{ Erigeron philadelphicusand E. canadensis. Physiol. Plant. 66: 605-608.
(Received: 27 June 2001)(Accepted: 14 September 2004)
10 PERTANIKAJ. TROP. AGRIC. SCI. VOL. 27 NO. 1, 2004












![General product leaflet [EN] - itoh-denki.com product leaflet-PPP... · or ASI module 1 3 2 4 5 6 8 7 9 10 11. ITOH DENKI CO., LTD Conception graphique : Studio MARIE-PIERRE 04 50](https://img.pdfslide.us/doc/110x75/5c81aa7c09d3f2f3348cb009/general-product-leaflet-en-itoh-denki-product-leaflet-ppp-or-asi-module.jpg)