Embed Size (px)
Citation preview

Toxicology in Vitro 27 (2013) 1320–1346
Contents lists available at SciVerse ScienceDi rect
Toxicology in Vitro
journal homepage: www.elsevier .com/locate / toxinvi t
Review
Mechanism-based testing strategy using in vitro approaches for identificationof thyroid hormone disrupting chemicals
AlberTinka J. Murk a,⇑, Eddy Rijntjes b, Bas J. Blaauboer c, Rebecca Clewell d, Kevin M. Crofton e,Milou M.L. Dingemans f, J. David Furlow g, Robert Kavlock h, Josef Köhrle b, Robert Opitz i, Theo Traas k,Theo J. Visser j, Menghang Xia l, Arno C. Gutleb m
a Wageningen University, Sub-department of Toxicology, Tuinlaan 5, 6703 HE Wageningen, The Netherlands b Institut für Experimentelle Endokrinologie, Charité-Universitätsmedizin Berlin, Berlin, Germany c Doerenkamp-Zbinden Chair, Institute for Risk Assessment Sciences, Division of Toxicology, Utrecht University, The Netherlands d The Hamner Institutes for Health Sciences, Research Triangle Park, NC, USA e National Center for Computational Toxicology, Office of Research and Development, US Environmental Protection Agency, Research Triangle Park, NC, USA f Neurotoxicology Research Group, Toxicology Division, Institute for Risk Assessment Sciences (IRAS), Faculty of Veterinary Medicine, Utrecht University, Utrecht, The Netherlands g Department of Neurobiology, Physiology, and Behavior, University of California, Davis, USA h Office of Research and Development, US Environmental Protection Agency, Washington, DC, USA i Institute of Interdisciplinary Research in Molecular Human Biology, Université Libre de Bruxelles, 1070 Brussels, Belgium j Department of Internal Medicine, Erasmus University Medical Center, Rotterdam, The Netherlands k Bureau REACH, National Institute of Public Health and the Environment, Bilthoven, The Netherlands l National Center for Advancing Translational Sciences, National Institutes of Health, Bethesda, MD 20892, USA m Department Environment and Agro-biotechnologies, Centre de Recherche Public – Gabriel Lippmann, Belvaux, Luxembourg
a r t i c l e i n f o
Article history: Received 10 November 2012 Accepted 18 February 2013 Available online 27 February 2013
Keywords:Endocrine disruption Thyroid hormone Alternatives to animal testing In vitro assaysToxicokineticsTesting strategy BatteryFunctional assay Metabolism
0887-2333/$ - see front matter Published by Elsevierhttp://dx.doi.org/10.1016/j.tiv.2013.02.012
⇑ Corresponding author. E-mail address: [email protected] (A.J. Murk).
a b s t r a c t
The thyroid hormone (TH) system is involved in several important physiological processes, including regulation of energy metabolism, growt h and differentiation , development and maintenance of brain function, thermo-regulation, osmo-regulation, and axis of regulation of other endocrine systems, sexual behaviou r and fertility and cardiovascular function. Therefore, concern about TH disru ption (THD) has resulted in strategies being developed to identify THD chemicals (THDCs). Information on potential of chemicals causing THD is typically derived from animal studies. For the majority of chemicals, how- ever, this information is either limited or unavaila ble. It is also unlikely that animal experiments will be performed for all THD relevant chemicals in the near future for ethical, financial and practical rea- sons. In add ition, typical animal experimen ts often do not provide information on the mech anism of action of THDC, making it harder to extrapolate results across species. Relevant effects may not be identified in animal studies when the effects are delayed, life stage specific, not assessed by the exper- imental paradigm (e.g., behaviour) or only occur when an organism has to adapt to environmental fac- tors by modulating TH levels. Therefore, in vitro and in silico alternatives to identify THDC and quantify their potency are needed. THD C have many potential mechanisms of action, including altered hormone produc tion, transport, metabolism, receptor activation and disruption of several feed-back mecha- nisms. In vitro assays are availabl e for many of these endpoints, and the application of modern ‘-omics’ technologies, applicable for in vivo studies can help to reveal relevant and possibl y new endpoints for inclus ion in a targeted THDC in vitro test battery. Within the framew ork of the ASAT initiative (Assur-ing Safety without Animal Testing), an international group consisting of experts in the areas of thyroid endocrinology , toxicology of endocrine disruption, neurotoxicology, high-throughput screening, com- putational biology, and regulatory affairs has reviewed the state of science for (1) known mechanisms for THD plus examples of THDC; (2) in vitro THD tests currently available or under development related to these mechanisms; and (3) in silico methods for estimating the blood levels of THDC. Based on this scientific review, the panel has recommen ded a battery of test methods to be able to classify chemicals as of less or high concern for further hazard and risk asses sment for THD. In addition, research gaps and needs are identified to be able to optimize and validate the targeted THD in vitro test battery for a mechanism-based strategy for a decision to opt out or to proceed with further testing for THD.
Published by Elsevier Ltd.
Ltd.

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1321
Contents
1. Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1321
1.1. Possible physiological consequences of thyroid hormone disruption . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1321 1.2. Environment al chemicals and THD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1322 1.3. Aim of the workshop. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13272. Thyroid hormone system and endpoints for THD . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1327
2.1. Box 1. Central regulation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13272.1.1. Endpoint: TRH receptor signalling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1327 2.1.2. In vitro assays for TRH receptor signalling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1327 2.1.3. Endpoint: TSH receptor signalling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1328 2.1.4. In vitro assays for TSH receptor signalling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1328
2.2. Box 2. TH synthesis and secretion by the thyroid gland . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1328
2.2.1. Endpoint: NIS-mediated iodide uptake . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1329 2.2.2. In vitro assays for NIS-mediated iodide uptake . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1329 2.2.3. Endpoint: TPO inhibition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1329 2.2.4. In vitro assays for TPO inhibition . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13292.3. Box 3. Transport of thyroid hormones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1330
2.3.1. Endpoint: Binding to transport proteins TTR and TBG . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1330 2.3.2. In vitro assays for binding to transport proteins TTR and TBG . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13302.4. Box 4. Metabolism and excretion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1330
2.4.1. Endpoint: Deiodination . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1331 2.4.2. Endpoint: Alanine side chain modification . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1331 2.4.3. Endpoint: Sulfation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1331 2.4.4. Endpoint: Glucuronidation . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1331 2.4.5. Inhibition of enzyme functioning . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1331 2.4.6. In vitro assays for induction and inhibition of enzymes for TH metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13322.5. Box 5. Cellular concentrations and transporters . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1332
2.5.1. Endpoint: cellular TH uptake . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13322.6. Box 6. Cellular responses . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1332
2.6.1. Endpoint: TR binding and transcriptional activity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1333 2.6.2. In vitro assays for detecting disruption of TR activity . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 13333. Chemical bio-activation and availability in vitro . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1334 4. Suggested testing strategy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1334
4.1. Tier 1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1335 4.2. Tier 2 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1335
5. Advantages of in vitro testing . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1337 6. Further outlook: using computational modelling to interpret in vitro assays for in vivo effects . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1337 7. Recommendations for assay use and development . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1337 8. Summarizing conclusions . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1338
Conflict of interest statement . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1338 Disclaimer . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1338 Acknowledgements . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1338 References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 1338
1. Introduction
Endocrine disruption (ED) by chemicals is not restricted to the sex hormone system, but also includes thyroid hormone (TH) dis- ruption (THD). THD is defined herein as a change in hormone pro- duction, transport, function or metabolism resulting in impaired homeostasi s. When the homeostasi s is not impaired it is called ahormone modulator. THD can be induced by a variety of causes including diet, disease, and exposure to environm ental chemicals.
TH are involved in several important physiolog ical processes such as regulatio n of energy metaboli sm (Cheng et al., 2010 ),growth and differentiation , development and maintenanc e of brain function and the sympathetic nervous system (Bernal, 2007; Horn and Heuer, 2010; Reinehr, 2010; Warner and Mittag, 2012 ), ther- mo-regulation (Ribeiro, 2008 ), osmo-regul ation and renal function (Vargas et al., 2006 ), regulation of onset and proper function of other endocrin e systems including the estrogen system, sexual behaviour and fertility, and cardiovascular functioning (Danziand Klein, 2012; Krassas et al., 2010; Wagner et al., 2008 ). Whether during developmen t of the organism, differentiation of cells and tis- sues, maintenanc e or alteration of physiologica l functions of adult individuals, in many cases TH effects can best be characteri zed as
‘permissive hormone action’. This indicates that the TH status of cells, tissues, and organisms provides the background and platform for other biologica l signals – hormonal, neural, immunologica l, nutritive and environmental – that are critical for maintenance of both developmen t and homeostasis of the organism as a whole (Lopez-Juarez et al., 2012; Pascual and Aranda, 2012; Sirakov et al., 2012 ).
The predominant TH in the circulation in the euthyroid situa- tion is 3,3 0,5,50-tetraiodoth yronine (thyroxine, T4), which is the precursor for the most active form of TH (3,30,5-triiodothyro nine; T3). Most of the known functions of TH are mediated by the inter- action of T3 with the nuclear T3-receptors (TRs), which act as li- gand-mo dulated transcrip tion factors. While almost none of the genes regulated by T3 are exclusively responsive to T3, virtually all molecular, cellular and metabolic events are more or less sensi- tive to TH (Grimaldi et al., 2012; König and Moura Neto, 2002; Oetting and Yen, 2007 ).
1.1. Possible physiological conseque nces of thyroid hormone disruption
Lessons learned from decades of biomedical studies of iodide deficiency, congenit al hypothyroid ism, genetic diseases related to

Table 1Physiological and related pathological process es related to the thyroid hormone system.
Thyroid-hormone physiological systems and processes
Related diseases References
Cellular metabolism Metabolic disease Brent (2012) and Song et al. (2011)Cell cycle/apoptosis Neoplasia Kim and Cheng (2012) and Piekielko-Witkowska and Nauman (2011)
Cancer Kim and Cheng (2012) and Pascual and Aranda (2012)Angiogenesis Cancer; cardiac hypertrophy; neurological
deficit (brain)Berg et al. (2009), Cheng et al. (2010), Davis et al. (2009), Judson et al. (2010),Luidens et al. (2010) and Zhang et al. (2010a)
Thyroid gland Goitrogenesis Paschke (2011a,b)Metabolism Obesity; metabolic syndrome Baxter and Webb (2009), Brent (2012), Cioffi et al. (2010), Liu and Brent (2010) and
Reinehr (2010)Nervous system Learning; memory; IQ Patel et al. (2011b)
Neurobehavioral disorders (e.g.hyperactivity; bipolar disorder)
Chakrabarti (2011) and de Cock et al. (2012)
Sensory development (incl. hearing) Crofton and Zoeller (2005), Forrest and Swaroop (2012) and Sharlin et al. (2011)Depression in adults Alzheimer’s disease van de Ven et al. (2012)
Immune system Autoimmunity Chang (2012) and Hodkinson et al. (2009)Skin Myxedema Aamir et al. (2010) and Safer (2011)Cardiovascular system Heart rhythm problems Donangelo and Braunstein (2011) and Klein and Danzi (2007)Reproductive system Reduced fertility Krassas et al. (2010) and Unuane et al. (2011)
Impaired reproductive development (onset of puberty)
Mann and Plant (2010)
Ca-homeostasis, skeleton Osteoporosis; fracture risks Bassett et al. (2010), Greenspan and Greenspan (1999) and Waung et al. (2012)Brown adipose tissue Adaptation to cold stress; obesity Bianco (2011), Himms-Hagen (1989), Obregon (2008) and Silva (2011)
1322 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
defective TH function as well as data from various animal models corroborate evidence for the hypothesis that transient or persisten tTHD could alter maintenanc e of homeostasi s within the hypothal- amus–pituitary–thyroid-periphery (HPTP) axis and modulate the peripheral thyroid hormone depende nt functions. This might lead to temporary loss of homeostas is or even alter set points leading to long-term TH dysregul ation and physiologica l conseque nces, including thyroid pathology and altered metabolism and perinatal developmen t. Alterations of the TH system are associate d with sev- eral serious human diseases (Table 1).
TH are particularly important in perinatal development. They are involved in several critical processes for neurodevelop ment: neuronal proliferation, migration, synaptogenes is, synaptic plastic- ity and myelination processes (Horn and Heuer, 2010; Howdeshell ,2002). In humans, TH production starts at approximat ely 11 gesta- tional weeks and increases with the developmen t of the fetal HPTP axis (Howdeshel l, 2002 ). Trans-placenta l transfer of maternal TH to the fetus is critical for neurodevelopm ent (Bernal, 2007; Zoeller and Rovet, 2004 ), as impaired psychomotor developmen t, behav- ioural changes and effects on visuo-spa tial processing have been observed in children born to mothers with (subclinical) hypothy- roidism (de Cock et al., 2012; Haddow et al., 1999; Pop et al., 1999). Because alterations in TH balance can lead to altered devel- opment, even temporary THD during the perinatal period can have long-term conseque nces on human health (Zoeller and Rovet, 2004). But also later in life alterations in the TH system have been shown to play a role in several mental illnesses such as Alzheimer’s disease, bipolar disorder and major depressive disorder (Baueret al., 2008; Carta et al., 2002; Cooper-Kazaz and Lerer, 2008; de Jong et al., 2009; Hogervorst et al., 2008; Lovell et al., 2008; Tan and Vasan, 2009 ).
THDCs may interact with a number of molecular components of the HPTP axis and the functioning of the peripheral tissues: TH synthesis, TH storage and release by the thyroid gland, feedback mechanism s within the HPT, protein-bind ing and TH distribution ,cellular TH uptake, intracellul ar TH metabolism, catabolism of TH, classical ‘nuclear’ T3 receptor binding, as well as other target proteins (‘receptors’) in the cell membrane, mitochondria and other subcellular structures (Brix et al., 2011; Brucker-Davis, 1998; Capen, 1994; Cheng et al., 2010; DeVito et al., 1998; Köhrle,2008; Miller et al., 2009 ). THDCs may also react with more than one component of the TH system, possibly at different internal
concentr ations, which may give an overall in vivo effect at blood concentr ations that are lower than in vitro studies might suggest. This is expected to be the case with polychlor inated biphenyls (PCBs) and polybrom inated diphenyl ethers (PBDEs) in the blood of mothers and infants that have been associated with changes in thyroid hormone status and developmen tal endpoints and fertility (Chevrier et al., 2010; Harley et al., 2010; Koopman-Esse boom et al., 1994; Langer, 1998; Langer et al., 1998; Morse et al., 1993 ).
In addition to the direct impact of chemical s on the TH system, biologica l and environm ental stressors may also increase the sensitivit y of a number of physiologica l processes to THDCs. Iodine deficiency, which is commonly observed worldwide (Andersso n et al., 2012; de Benoist et al., 2008; Walker et al., 2007; Zimmermann and Andersson, 2012 ), is a well-known risk factor for increased sensitivity to adverse effects from thyroid-dis- rupting chemicals (Blount et al., 2006; Pearce and Braverman, 2009). Such effects are known as well from co-exposure to goitro- gens originating from the diet (e.g., (iso-)flavones like genistein in soy, goitrin contained in millet and cassava) in areas of iodine defi-ciency (Delange, 1994; Doerge and Sheehan, 2002; Köhrle, 2008 ).
1.2. Environm ental chemicals and THD
Various environmental contaminants have been shown to dis- rupt thyroid homeostasi s via several mechanism s (Brucker-Davi s, 1998; Capen, 1997; Cavalieri and Pitt-River s, 1981; Crofton, 2008; Hurley, 1998; Jugan et al., 2010; Köhrle, 2008; Surks and Sievert, 1995; Zoeller, 2007 ). Disruption of the TH system has been shown as one of the major toxic effects for chemicals ranging from halogena ted aromatic chemicals such as PCBs to inorganic anions such as perchlorate and nitrate (Brouwer et al., 1998; Brucker- Davis, 1998; Hallgren and Darnerud, 2002; Wolff, 1998 ), arsenic (Ciarrocca et al., 2012 ) or perfluorinated chemicals (Lopez-Espin-osa et al., 2012 ). This results in alteration of important biologica lprocesses under control of THs (Lopez-Juarez et al., 2012; Pascual and Aranda, 2012; Sirakov et al., 2012 ).
Historical ly, informat ion on the potency of THDCs has been de- rived from animal studies, mostly using rodents or amphibians (Biegel et al., 1995; Christenson et al., 1996; Hallgren and Darnerud, 2002; Davey et al., 2008; Grimaldi et al., 2012 ). How- ever, this information is limited to a small number of the 10,000+ chemicals that need assessme nts of potential risk (EPA,

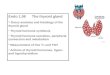
Fig. 1. Schematic representation of relevant aspects of thyroid hormone (TH) regulation and action (boxes 1-6), aspects of regulation of availability of TH disrupting compounds, and physiological consequences and related diseases of TH disruption. For further explanation see text.
AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1323
201 2; Jud so n et al ., 200 9; NRC , 19 84, 200 7; Wagne r, 20 00 ). Perf orm -in g in viv o exp eri me nts to as se ss poss ib le THD for such a la rge num- ber of che mi ca ls is unl ik ely to ha pp en in th e near futu re for pra cti ca l,as well as ethic al and financia l re aso ns. In add iti on , a sc ie ntific argu -me nt to repl ace anim al exp eri me nts is that api ca l endp oin ts pr ovid eli tt le in sig ht in unde rl yin g mec hanis ms (NIE HS, 20 07 ), ma y not bepr ed ic tiv e for the huma n si tua tio n or may not id entif y rel eva ntli fe -st ag e sp ecific effec ts, es pe cia ll y si nce un exp ec ted sp ecie s di ffer- ence s may exis t, an d curr ent ro de nt exp er im ents are no t opt im ize dfor the ide nti fica tio n of TH D. In addi tio n, the fact that THD C could re act wit h mo re tha n on e comp on ent of the TH sy st em, pos si bly atdi ff ere nt int ern al con centr at ion s, cou ld resu lt in conf usi ng non -mo noto nic in viv o do se –re sp onse cur ve s. A mor e me chani st ic all ydr iv en in vit ro app ro ac h can he lp to dis cri mi nate be tween chemi cals wit hou t in di cat io n for THD, wi th a str on g in di cat io n for THD and wit h wea k or not conclu si ve in di ca tio n for THD.
The relevance of such an approach is illustrated by the fact that the U.S. EPA is considering screening a universe of approximat ely 10,000 chemicals for endocrine disrupting properties as part of the endocrin e disruptor screening program. To facilitate rapid screening of endocrine-related endpoint s, further priority setting will be done using high throughput in vitro assays and in silico models (EPA, 2012 ). In Europe, different regulatio ns contain spe- cific provisions regarding endocrine disruptors: REACH (EU,2007), plant protectio n products (EU, 2009a ), cosmetics (EU,2009b) and biocides (EU, 2012 ). The OECD has developed several test methodol ogies and testing strategie s relating to EDCs but
not all of these methods are currently part of the standard informa- tion requiremen ts of the above mentioned regulations. Considering the large number of substances that might need to be prioritized for endocrine-related endpoint s, both in the US and in the EU, there is a need for an efficient, less-animal intensive screening method to identify potential THDCs.
Over the last two decades there has been a growing interest in defining the mechanism s of toxicity in in vitro systems (Eisenbrandet al., 2002 ). Furthermore, there has been an explosion of technol- ogy that has greatly increased not only the breadth of information that can be extracted from biological samples, but also the throughput by which samples can be analysed and interpreted. Thus, genomics, proteomics and metabolomics are becoming com- monplac e tools in biologica l research, as are genetically modifiedcells, transgenic animal models, model organisms such as zebra fish, and stem cell models. Many of these approaches have been adapted to the process of drug discovery (Houck and Kavlock, 2008), which has facilitated their adoption in toxicological studies including endocrine disruption, based on knowledge of the interac- tion of chemicals with biological pathways. While there has been much attention to develop pathway-based assays for estrogen and androgen function, approaches to the assessment of thyroid function have lagged behind because of the multiplicity of ways in which TH function can (and has) been altered by environm ental chemical s (OECD, 2006 ). This paper focuses on the impleme ntation of these approaches in testing strategies to the assessment of the effect of chemical s on the thyroid system.

Table 2This table describes potential targets for thyroid hormone disrupting chemicals plus available non-vertebrate test systems. The numbers of the different sub-sections of the table refer to the Boxes with relevant aspects of TH regulation and action as depicted in Fig. 1.
Target Assay Model References Comme nts
Box 1. Central regulation of the thyroid system via the hypothalamic–pituitary–thyroid axis TRH prod uction hypothalamus Genomic array indicating TRH
production Human glioblastoma–astrocytoma (U373MG) cells Garcia et al. (2000) Validation of assay and HTS application to be
develope dTRH receptor activation pituitary TRH receptor binding and calcium
measurem ent (functional assay)Stab le ValiScreen TRH (human) cell line www.perkinelmer.com/
GPCRcomplete (2012)96-well and 384-well plate format
TSH expression and secretion Mou se pituitary (TaT1) cells Zatelli et al. (2010) Validation of assay and HTS application to be develope d
TSH receptor activation thyroid gland
TSH-mediate d cAMP production cAMP measurement via CNG channe l-coupled cAMP assay
Human embryonic kidney (HEK293) cells stably express ing TSH receptor coupled to a cyclic nucleoti de gated ion channel as a biosensor
Titus et al. (2008) Publ ished HTS assay in 1536-well plate, requires aspecific cell line with expression of CNG channel. Assay kit available at Cisbio, www.htrf.com
TSH-mediate d cAMP production (direct cAMP measurement)
Chinese hamster ovary (CHO) cells stably express ing TSH receptor
Santini et al. (2003), Van Sande et al. (2003) and Zimmermann-Belsing et al. (2002)
Sensit ive and rapid assays using novel (colorimetric,luminometric , dye, etc.) readouts not requiring isotopes or immunoassays for cAMP detection
TSH-depe ndent cell proliferation Rat thyroid (FRTL5) cell with endogenous TSH receptor
Jomaa et al. (2013) Validation of assay is needed; non-TSH mediated cell proliferati on canno t be excluded
Box 2. Thyroid hormone production and biokinetics NIS-mediated iodide uptake Radioiod ide uptake assay Rat thyroid (FRTL5) cells Arturi et al. (2002) and Lecat-Guillet
et al. (2008)Assay is usable for MTS (96-well) and has been applied for many chemicals, disadvantage: radio iodide use
Iodide uptake assay (non-radioiso tope)
Rat thyroid (FRTL5) cells Waltz et al. (2010) and Renko and Köhrle (unpublished)
Assay is usable for MTS, not tested in HTS
Iodide uptake assay (use of halide- sensitiv e fluorescence of yellow fluorescent protein (YFP))
Rat thyroid (FRTL5) cells stably express ing YFP- H1 48Q/I152L
Di Bernardo et al. (2011) and Rhoden et al. (2007)
Not yet validated as HTS; changes in YFP fluorescencenot strictly specific for cellular iodine
Iodide retention and efflux via NIS Transf ected FTC-133 (human thyroid cancer cell line)
Schroder-van der Elst et al. (2003) FTC-1 33 transfected with NIS; not validated for HTS
TPO enzyme activity TPO-me diated oxidation of iodide or alternative substrates (e.g.,guaiacol)
Cell-fr ee enzymatic assay Schmutzler et al. (2007a) Oxidativ e activity of TPO is determined by spectr ophotometric detection systems. Iodide oxidation assays are likely to capture more inhibitors than guaiacol-based assays. Not yet validated as HTS
TPO-me diated iodination of tyrosine
Cell-fr ee enzymatic assay Freyberger and Ahr (2006) and Schmutzler et al. (2007a)
Tyrosin e iodination by TPO is determined spectr ophoto-metrically. Not yet validated as HTS
Thyroid hormone production Measur ement of TH with antibod ies production in follicle cells LC-MS/MS analysis of TH production
Zebrafish eleutheroembryos Antonica et al. (2012), Hornung et al. (2010), Kunisue et al. (2011) and Raldua et al. (2008)
This assay covers effects on all factors involved in TH synthesi s, e.g. NIS, Tg, TPO and DUOX. It requires intact thyroid follicles and still is ex vivo , not in vitro . The LC- MS/MS method determine s several TH metab olites simultaneous ly from a single sample of thyroid tissue or follicles in vitro , Not suitable for HTS, for Tier 2
DUOX -mediated H2O2
production Determination of H2O2 generation by DUOX/DUOXA
PCCl3 rat thyroid cells, CHO cells stably transfected with DUO X2/DUOXA2
Massart et al. (2011) H2O2 generation is determined spectrophotomet ric analysis of cell medium (conversion of homovanillic acid). Not yet validated as HTS
Box 3. Systemic distribution and transport of thy roid hormones T4 binding proteins TTR and TBG Competit ion with T4 for TTR and
TBG bin ding Cell-fr ee biosensor assay (Biacore) with T4 bound to chip
Marchesini et al. (2008) Per formance of Biacor e chips variable
Fluorescent microtiter method Purified human TTR and TBG Cao et al. (2011) Assay is disrupted by autofluorescent chemicals TTR or TBG Anilinosulfonic acid (ANSA) displacement assay
Purified human TTR and TBG Montano et al. (2012) Microtite r method; ANSA, potent ligand, substitute for T4, fluorescent when bound to TTR or TBG
TH membrane transport placenta and blood brain barrier
Transf er over cell monolayer in transwell system
Human placental choriocarcinoma (JEG) cells; human choriocarcinoma (JAR) cells; bovine brain cells
Kovo and Golan (2008), Patel et al. (2011a), Powell et al. (2000) and Vandenhaute et al. (2011)
This system is developed for transport of drugs; appli cability for THs and THDs to be determined
1324A
lberTinka J. Murk et al. /Toxicology in V
itro 27 (2013)1320–
1346

Box 4. Metabolism and excretion of thyroid hormones (De-)Glucuronidation Inhibition or upregulatio n of
cellular enzyme activity of: Tests are performed in intact (transfected) cells (over-)expressing the enzymes; Human hepatom a(HepaRG) cells
Martin et al. (2012), Tong et al. (2007),Wu et al. (2005)
This also includes effects on enzyme production, unless the cells overexpress the enzymes; in that case only inhibition of enzyme activity can be determined – Glucuronidases
– UDP-glucuronosyltrans ferases Inhibition of: Human recombinant enzymes; or enzymes
prepared from human and mouse livers, purifiedenzyme preparation
Hamers et al. (2006), Schuur et al. (1998a,b,c) and Wu et al. (2005)
Can be performed with recombinant enzymes or microsomal preparations , not yet validated as HTS
– Glucuronidases – UDP-glucuronosyltrans ferases
(De-)Sulfation Inhibition of enzymatic activities related to (de)sulfation:
Human recombinant enzyme Ekuase et al. (2011) Human cell lines with the relevant enzyme activities remain to be developed
– Sulfotransferases – Sulfatases
Deiodination Upregulation of Dio1/2/3 activities Human, rat, mouse cell lines Dentice and Salvatore (2011), Köhrle(2002, 2007, 2008) and Leonard and Rosenberg (1980)
Performed in cellular systems; sensitivity of non- radioactive alternativ es to be determined
Inhibition of Dio1/2/3 activities Human recombinant enzyme; human, rat, mouse cell lines; purified enzyme preparation
Dentice and Salvatore (2011), Hotz et al. (1996), Köhrle (2002, 2007, 2008), Leonard and Rosenb erg (1980),Renko et al. (2012) and Schmutzler et al. (2007b)
Can be performed in tissue homogenates or cellular systems; sensitivity of non-radioactive alternatives to be determined
TH deiodination profile (mainlyDio1)
Enzyme preparation; can be pooled human liver microsomes
Butt et al. (2011) Mass spectrometry- based method for measuring the activity of Dio, allows for analysis of TH profile
Box 5. (Sub-)Cellular distribution of thyroid hormones by membrane transporters and deiodinases TH membrane transporters TH transport assay via TH
membrane transporters Transfected COS, HEK293, MDCK, JEG, Hela, GH3 and other cells (over) express ing membrane transporters MCT8/10, LAT1/2, OATP1c1. MDRs also play a role in rat FRTL-5 thyroid cells and mouse NIH- 3T3 cells
Cavalieri et al. (1999), Kinne et al. (2009), Mitchell et al. (2005), van der Deure et al. (2010) and Westholm et al. (2009, 2010)
Stably transfected cells expressing individual TH transporters are available. Not yet validated as HTS. Transporters cell type specific. Not yet validated as HTS
Up- or down regulation of OATPs Rat hepatoma (H4) cells Chalmers et al. (1993) and Westholm et al. (2009)
Can only be tested in cells not over-expressing OATPs. Needs further development
TH membrane transport placenta and blood brain barrier
Transfer over cell mo nolayer in transwell system
Human placental choriocarcinoma (JEG) cells; human choriocarcinoma (JAR) cells ; bovine brain cells
Kovo and Golan (2008), Patel et al. (2011a), Powell et al. (2000) and Vandenhaute et al. (2011)
This system is developed for transport of drugs; applicability for THs and THDs to be determined
Peripheral deiodination Inhibition of peripheral Dio1/2/3 enzyme activity
Recombinant enzymes, tissue homogenates, subcellular fractions, cellular systems
Hotz et al. (1996), Köhrle (2002),Leonard and Rosenberg (1980), Renko et al. (2012), Schuur et al. (1998a,b,c)and Visser et al. (1983)
Radioligand, well-charac terized deiodinases are tissue-specific not yet fully in vitro (ex vivo )
Upregulation of peripheral Dio1/2/ 3 enzymes activity
In vivo (non-vertebrate) or in cellular system Köhrle (2002), Leonard and Rosenberg (1980), Renko et al. (2012) and Visser et al. (1983)
T3 is a strong inducer of hepatic Dio1 and also Dio3 activity in several tissues. T4 inactivates Dio2 by posttranslational mechanism, sensitivity of non- radioactive alternativ es to be determined
Box 6. Cellular responses to thyroid hormones Nuclear TR binding Molecular docking/ in silico
modelling of binding to nuclear TR In silico approach Schapira et al. (2003) >250,000 chemicals were screened, validation by
transfection and ligand bin ding TH–TR competitive binding assay (radioligand)
Whole cell uptake, nuclear extracts of TH responsive cells, overexpressed TRs
Kitamura et al. (2002, 2005a,b),Schapira et al. (2003) and You et al. (2006)
Disadvantages: Radioligand (125I) based, potential for high rate of false negatives; cannot distinguish between agonists and antagonists; folding of TR in vitro. Advantage s: HTS possible; solid -state binding assays available
Nuclear TR activation Stable luciferase reporte r gene assay for endogenous TR- activation and antagonism
Rat GH3 cell line based reporte r line (GH3.TRE-Luc); Lentivirus transduced Xenopus reporter line (XL58-TRE-LUC)
Freitas et al. (2011) and Sugiyama et al. (2005b)
Both TR a/b endogenously expressed. Published HTS (1536 well); not clear if line can be propagated as astable integrant, specific for amphibian cell conditions
Stable reporter gene assay for specific TR a or TR b receptor activation and antagonism
Rat PC12 cells express ing chicken TR a1 (PC-DR-LUC) Human (TRa, HeLa-Luc cells) NR1A1 and NR1A2 indigobioscie nces.com
Jugan et al. (2007, 2009) A variety of cell lines available, may need optimization/validation for HTS; with overexpressed TRs reporte r gene
Stable co-transfected gal4 TR expression vectors and UAS (upstream activating sequence)
Human embryonic kidney (HEK293T) cells stably transfected with gal4 TRb fusion, UAS b lactamase reporter (GeneBLAzer cell, Invitrogen, Inc.)
Huang et al. (2011) and Invitrog en (2012)
TR b-specific; Human based published HTS
(continued on next page )
AlberTinka J. M
urk et al. /Toxicology in Vitro 27 (2013)
1320–1346
1325

Table 2 (continued)
Target Assay Model References Comments
based reporters Transient transfection reporter gene assay to investigate nuclear TR transactivation and antagonism
Overexpressed TRs, Gal4-TR fusions or endog enous TRs in a variet y of cell types co-transfected with TRE or UAS based reporter genes
Hofmann et al. (2009), Ibhazehiebo et al. (2011), Kitamura et al. (2005a,b)and You et al. (2006)
Applicable to a variety of cell lines, several overexpressed TRs, reporter gene not fully wrapped in native chromatin which is important for part of the TR role in repression/activation
Stable reporter gene assay in yeast Full length TR or gal4 TR Iwasaki et al. (2002), Kitamura et al. (2005a), Kudo et al. (2006), Moriyama et al. (2002), Sugiyama et al. (2005a,b)
HTS possible; not mammalian; yeast cell wall permeability to chemicals limited, high incidence of false negatives
Liquid chemiluminescent DNA pull-down assay for association of TH receptor with TRE
Cell-free bead assay Ibhazehiebo et al. (2011) Novel endpoint; binding to TRE does not determine transactivation or trans-repression
Co-activator or co-repressor binding assay
PathHunter NHR co-activator assay based on b galactosidase complementation (DiscoveRx)
CHO-K1 cells stably transfected with TR a or TR band steroid receptor coactivator peptide (SRCP)
Patel et al. (2009) HTS possible, cell-based in vitro assay available at DiscoveRx
Human TR-alpha activation (yeasttwo-hybrid assay)
S. cervisiae (Y190) cells into which human TR a and coactivator TIF2 have been introduced
Arulmozhiraja et al. (2005) HTS possible; chemical accessibility across yeast cell wall a concern
LanthaScreen TR-FRET TRbeta coactivator assay (Invitrogen)
Human recombinant TR ligand-binding domain D’Souza et al. (2008) HTS possible, in vitro assay available at Invitrogen
CoA-BAP system; protein–proteininteractions detected via coactivator tagged alkaline phosphatase activity
Recombinant NR ligand-bi nding domain Kanayama et al. (2003) Completely in vitro assay, HTS possible; limited use for TRs to date (PPARs and other NRs)
Coactivator or corepressor peptide interaction assay based on fluorescence polarization
Human TR b LBD protein, SRC2 derived peptide; Human TR a1 LBD protein, SRC1 or NCoR derived peptides
Johnson et al. (2011) and Levy-Bimbot et al. (2012)
HTS adapted, >250,000 chemicals screened; cell free, robust assay; corepressor inter actions, or coactivato rinteractions acting via the amino terminal transactivation domain are not included ; HTS possible, includes corepressor interaction assay, isoform specificity not included
TR, coregulato r stabilization and autoregulation
Western blot, immunoprecipitation, quantitative PCR to investigate TR and coregulator modification,turnover, localization and autoregulation
e.g. Rat pituitary tumour (GH3) cells (or other cell types expressing endogenous or tagged receptors and coregulators)
Ball et al. (1997), Baumann et al. (2001), Hong et al. (2001) and Misiti et al. (1998)
Not clear how reflective of in vivo situation, possible role for arsenic in SMRT localization. Research gap
Mitochondrial TR- and non-TR mediated responses
Integrin binding CV-1 cells express alpahVbeta3 integrin but not nuclear TR
Bergh et al. (2005), Blanchet et al. (2012) and Freindorf et al. (2012)
Importance to overall TH physiology in progress, needs action; radioligand assay; Mitochondrial thyroid hormone effects mediated by the p43 TR a1 form
Cell type specific response s TH-dependent cell proliferation (T-screen)
Rat pituitary tumour cell line (GH3) cells, others Gutleb et al. (2005), Kitamura et al. (2005b), Medina and Santisteban (2000) and Schriks et al. (2006)
Can distinguish between agonist and antagonist; also non-TR-medi ated TH-related effects included; non- TH-related proliferation cannot be excluded (falsepositive)
1326A
lberTinka J. Murk et al. /Toxicology in V
itro 27 (2013)1320–
1346

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1327
1.3. Aim of the workshop
The current review is based on the outcome of a worksho p or- ganized within the framework of the ASAT initiative (AssuringSafety without Animal Testing) (Fentem et al., 2004 ). The general aim of the ASAT initiative is to develop ‘‘a radical new approach to assessing the risk posed by exposure to chemicals that would not involve testing of animals, taking advantage of the rapid ad- vances in science and technology.’’ Experts in thyroid endocrin ol- ogy, toxicologists with experience with endocrin e disruption and neurotoxicol ogy, computational experts, high-throughp ut screen- ing (HTS) and regulatory experts reviewed the state of science for (1) known mechanism s for THD plus examples of THDC; (2)in vitro THD tests currently available or under development related to these mechanis ms; and (3) in silico methods for estimating the blood levels of THDC.
Based on this scientific review, the panel recommends a battery of test methods to be able to classify chemicals as of less or high concern for further hazard and risk assessment for THD. In addi- tion, research gaps and needs are identified in order to optimize and validate the targeted THD in vitro test battery. This validation can lead to a mechanism -based strategy used for deciding on whether to opt out or to proceed with further THD testing.
To this aim, an inventory was taken of (1) the most relevant molecular targets and pathways that underlie thyroid functional disturbance s, which should be included in THD testing, (2) pres- ently available or preliminar y in vitro and in silico approach es that cover the most relevant mechanisms, and (3) areas in which assay developmen t is needed to fill data gaps in a comprehensive in vitro THD testing strategy.
2. Thyroid hormone system and endpoints for THD
Current knowledge indicates that the majority of THD effects are mediated via influences on the HPTP axis rather on direct inter- ference with nuclear receptor function in the target tissues. For aconceptual framework, the workshop expanded earlier systems biology models of thyroid disruption (Capen, 1997 ) using an ap- proach recently develope d (Keune et al., 2012; Ravnum et al., 2012; Smita et al., 2012; Zimmer et al., 2012 ) (Boxes 1–6; Fig. 1).THDC can interfere with the central regulatio n and feedback mech- anisms of the HPTP axis (Box 1); at the site of TH synthesis (Box 2, e.g., by inhibition of iodide uptake or iodination of thyroglobul in);with the TH distribution via the blood (e.g., by competition for high affinity binding of TH to transthyretin (TTR) and TH binding glob- ulin (TBG)) and across placenta and blood brain barrier (Box 3);with central metabolism and excretion of TH (Box 4; e.g., by hepa- tic deiodinases , and conjugating and deconjugating enzymes);with cellular uptake by selective TH transporters of the cell mem- brane, and intracellular activation or inactivati on of TH (Box 5);and finally some THDC interact with membrane, mitochond rial and nuclear TR receptors (Box 6).
An in vitro test battery to identify THDC should include the most relevant targets known for THD. For each component of the model in Fig. 1 the most relevant endpoints are briefly discussed. More detailed information can be found in the additional references gi- ven. For the selected endpoints, a panel of potential in vitro bioas-says was identified (Table 2) that should cover the biologically or toxicologica lly most relevant endpoints for THD. Table 2 includesexisting HTS bioassays (96- or even higher density well formats)as well as assays for which a higher throughput format still needs to be developed. While many more techniqu es and assays may provide valuable mechanis tic information on pathways and mech- anisms of TDHCs, only assays that can be performed in cell lines or with isolated proteins, and therefore amendable to medium to high throughput screening (MTS–HTS) are included.
2.1. Box 1. Central regulation
The tight regulation of circulating TH levels, a prerequisite for essential physiological control, is achieved by complex control mechanis ms along the hypothal amus–pituitary–thyroid (HPT)axis. Thyroid- stimulating hormone (TSH) is the primary physiolog- ical regulator of thyroid gland function and growth (Vassart and Dumont, 1992 ). The production and secretion of TSH by the thyro- trophs, a specialized pituitary cell type, is under a multifact orial control. A central regulator of thyrotroph activity is thyrotropi n- releasing hormone (TRH), synthesized by neurons residing in the paraventr icular nucleus of the hypothal amus (Nillni, 2010 ). TRH binding to its membrane receptor in thyrotrophs activates phos- pholipase C signalling via Gq11 leading to increased gene expres- sion of TSH a and b subunits. In addition to stimulating TSH production, TRH has also been shown to affect the glycosyla tion pattern of TSH thereby affecting its biologica l activity (Persani,1998). Importantly, as a target of several key pathways, these hyp- ophysiotr ophic TRH producing neurons integrate inputs from var- ious neural circuitrie s to provide exact set points for the thyroid axis depending on negative feedback by circulating TH, nutritional status, illness and even circadian rhythms (Costa-e-S ousa and Hollenber g, 2012 ).
The neuroendocrine control of thyroidal TH synthesis and secretion is very sensitive to negative feedback exerted by circulat- ing TH, both at the level of the pituitary and the hypothalam us (Fliers et al., 2006 ), and involves various molecular mechanisms such as specific expressions patterns of TR genes, TH transporter sand deiodinas es (Chiamolera and Wondisford, 2009; Fekete and Lechan, 2007; Fliers et al., 2006 ) that are discussed in the following boxes.
Thus, TSH and TRH receptor binding is central to HPT control. Assays which may be used to detect chemicals that interfere with signalling through these receptors are described below.
2.1.1. Endpoint: TRH receptor signalling
Overall, very little is known to date about the potential of envi- ronmental chemicals to specifically affect the neuroendoc rine con- trol mechanisms of thyroid gland function via alteration in TRH receptor signalling. Several drugs including somatostati n ana- logues and synthetic rexinoids (retinoid X receptor (RXR)-selectiveretinoids ) have been shown to suppress TSH levels and cause cen- tral hypothyroidism in humans as well as in experime ntal rodent models (Haugen, 2009; Sharma et al., 2006; Zatelli et al., 2010 ).The rexinoid LG 268, for example, specifically suppressed expres- sion of TSH b and DIO2 genes in thyrotrophs without affecting hypothal amic TRH mRNA expression (Sharma et al., 2006 ). Such findings are particularly interesting in the light of recent observa- tions that certain environmental chemical s (e.g., bispheno l A (BPA),2,4-dichlorop henol, p,p 0-dichloro diphenyltrichlor oethane, tributyl- tin chloride) can also act as disrupters of retinoid X receptor (RXR)function (Grun and Blumberg, 2006; Li et al., 2008 ). RXR typically from heterodimers with TR on TR responsive elements of TH regu- lated genes and thus RXR ligands modulate TR action on gene expression (Putcha et al., 2012 ).
The de ve lo pm ent of se ns iti ve tool s and as say s to char acte ri zeneur oend oc ri ne hy po thal am ic an d pi tu it ar y end oc ri ne ce ll re -sp on se s to pu tat iv e THDC s cl earl y re pr ese nt s a ma jo r re sea rc h need .
2.1.2. In vitro assays for TRH receptor signalling One cell line that holds promise for in vitro toxicological studies
is the pituitary thyrotroph TaT1 cell line. This cell line has been proven a valid and physiologica lly relevant tool to dissect various responses of thyrotrophs to TH as well as TRH (Chiamolera et al., 2012; Janssen et al., 2011 ).

1328 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
TRH receptor is a G protein-coupled receptor. In pituitary cells, TRH binds to its receptor and activates phospholipase C leading to an increase of inositol 1,4,5-trisphos phates (IP3) that mobilizes intracellular calcium (Gershengorn, 1993 ). There are at least two available assays for directly evaluating TRH receptor binding, bind- ing and activation of TRH, and intracellular calcium measurement. Both assays are commerc ially available (www.per kinelmer.com/ GPCRcompl ete, 2012 ).
2.1.3. Endpoint: TSH receptor signalling
At the thyrocyte level, stimulation of the TSH receptor (TSHR)by TSH activates several second messenger signalling cascades leading to increased iodide uptake, TH synthesis and secretion (Ro-ger et al., 2010; Vassart and Costagliola, 2011 ). Among the various TSHR-regula ted second messenger signalling cascades, the adenyl- ate cyclase-cAMP- protein kinase A pathway has been most inten- sively studied and the increase in cellular cAMP content is aclassical hallmark of TSHR stimulati on in thyrocyte s. A few studies are available that specifically analysed the interference of environ- mental chemicals with TSHR function (Picchietti et al., 2009; Rossi et al., 2007, 2009; Santini et al., 2003 ). Examples include the insec- ticide DDT and the PCB mixture Arochlor 1254 which were shown to inhibit both basal and TSH-stimulate d accumulation of cAMP in TSHR-transf ected cells (Rossi et al., 2007; Santini et al., 2003 ). Also several estrogen receptor agonists and antagoni sts such as diethyl- stilbestrol, quercetin, bisphenol A, and 17 b-estradiol have been shown to partially reduce TSH-induced cAMP accumulati on (Rossiet al., 2009 ).
2.1.4. In vitro assays for TSH receptor signalling In humans the TSHR activates both the cAMP and phospho li-
pase C-PIP2 cascades, which are both important for TH biosynthe- sis (Song et al., 2010 ). There are several automate d assays for TSHR binding by stimulati ng or blocking antibodies, there are small li- gands activating and blocking TSH binding, and there are highly selective monoclonal antibodie s to compete with TSH binding both blocking or stimulating the TSHR (Basaria and Cooper, 2005; Kreuchwig et al., 2011; Nunez Miguel et al., 2012; Titus et al., 2008). Measurem ents of cAMP accumulation in TSH-stimul ated thyroid cells (or TSHR-expres sing non-thyroid cells) can be used to evaluate chemicals for their potential to interfere with TSHR function (Rossi et al., 2007, 2009 ).
A quantitat ive HTS assay has been developed for the identifica-tion of TSHR agonists (Neumann et al., 2008, 2009; Titus et al., 2008). This assay utilizes a cyclic nucleotide gated ion channel (CNG)-coupled approach to measure changes in intracellular cAMP accumulation. In this cell system, TSHR stimulation by TSH leads to increased intracellular cAMP levels which activate a CNG cation channel leading to the influx of cations such as sodium and calcium and subsequently to a membrane depolarizati on. It is this mem- brane depolarization that is detected by means of a membran e po- tential dye in live cells. Changes in indirect measurements such as cAMP must be interpreted with caution. Xenobiotics may alter cAMP concentratio ns via a variety of mechanism , one of which may be TSH receptor activity. The same holds true for the recently developed TSH mediated cell proliferation assays in FRTL-5 rat thy- roid cells (Jomaa et al., 2013 ).
2.2. Box 2. TH synthesis and secretion by the thyroid gland
In response to TSH stimulation, the thyroid tissue produces and releases the T4 and to a lesser extent T3. Thyroid follicles represent the functional subunit of thyroid tissue. Each follicle is lined by a single epithelial cell layer and is filled with a thyroglob ulin- containing colloidal mass. A dense capillary network surrounds
individua l follicles providing intimate contact of thyroid follicular cells (thyrocytes) to the blood stream. Because TH are iodothyro- nine derivatives, uptake of iodide from the blood stream represents a critical step in their biosynthesis. Iodine is concentrated in thyro- cytes by a factor of 20–40 over blood plasma iodide concentratio ns (Dohan et al., 2003 ) by the sodium–iodide (Na+/I�) symporter (NIS)protein, a membran e glycoprotein located on the basolateral side of thyroid follicular cells (Dohan et al., 2003 ).
At the apical pole of thyrocyte s the ion channel pendrin exports iodide into the follicular colloid lumen (Taylor et al., 2002; van den Hove et al., 2006 ). Organification of iodide takes place at the apical membran e of thyrocytes (Regard and Mauchamp, 1973 ). Localized at the thyrocyte /colloid interface, the thyroid peroxidase (TPO) en- zyme oxidises iodide for organification of iodine, it catalyzes the iodination of tyrosyl residues in the thyroglobul in polypeptide chain to yield the TH precursor s monoiodoty rosine (MIT) and diio- dotyrosin e (DIT). TPO also catalyzes the subsequent coupling of MIT and DIT to produce T3 and T4 which remain covalently bound to the thyroglobul in matrix (Taurog, 1970 ). In addition to thyro- globulin and iodide, TPO requires H2O2 as a third factor to carry out the abovementioned reactions . H2O2 production is likely arate-limiti ng step during TH generation (Song et al., 2007 ). Two NAD PH -d epe nde nt oxi do re du ctas es , te rm ed dua l oxi das es (DUOX 1,DUO X2 ), ha ve bee n id en ti fied as im po rta nt co mp onen ts of th e H2O2-gene ra ti ng sy ste m in thy ro cyt es .
Iodinated thyroglobul in is stored extracellularly in the colloidal compartme nt. Upon demand, thyroglobul in is endocytosed from the colloid, enzymatical ly cleaved within the thyrocytes by cathep- sin enzymes and finally TH is deliberated and actively secreted into the blood stream by monocarboxyl ate transporter 8 (MCT8) and other TH transporters (Brix et al., 2011; Di Cosmo et al., 2010; Dunn and Dunn, 2001; Trajkovic-A rsic et al., 2010; Wirth et al., 2011). Several steps during TH synthesis are still not well under- stood such as the transport of the small charged iodine molecule which cannot easily diffuse through plasma membran es and the actual route of T4 and T3 release from the thyrocyte into the bloodstream .
The observation of hypothyroid ism and goitre in patients carry- ing mutations of genes coding for NIS, TPO, DUOX2, DEHAL and TSHR indicate that chemically- induced alterations in the activity of these molecules would also result in thyroid dysfunction. Fur- thermore , the thyroid gland is one of the most vascularized tissues of our body, which allows for significant exposure of thyrocytes to THDC in the blood (Gerard et al., 2009 ). The effect of THDCs on TH synthesis and secretion is well established with experimental data for inhibitors of NIS (perchlorate) and TPO (thiocarbamide drugs)function. Also the unique continuo us and life-long production of H2O2 and ROS (reactive oxygen species) for adequate TH biosyn- thesis (Poncin et al., 2008 ) renders the thyroid gland highly vulner- able for low molecula r weight agents, which may interfere with for instance H2O2-catalyzed oxidation and activation (Divi and Doerge, 1994; Jeong et al., 2005; Köhrle, 2008; Schmutzler et al., 2007a,b ).
A central problem of in vitro detection of chemicals altering TH biosynth esis is the fact that the most important processes occur extracellul arly in the colloidal compartme nt of thyroid follicles and therefore require that thyrocytes are organized in a follicular structure . Thus, typical monolayer cell culture techniques are rarely useful for examining TH synthesis. Recently, however, mouse embryonic stem-cell s were stimulated to different iate into thyroid follicular cells in vitro . These cells formed functiona l folli- cles, which accumulate iodide, produce thyroglobul in and even organify iodide on thyroglobulin, hence mimicking functional thy- roid tissue (Antonica et al., 2012 ). The application of this system in in vitro assays needs to be further evaluated, but clearly represents a breakthr ough in in vitro methods that may lead to improved analysis of THDCs in the near future. Thus far the alternativ es,

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1329
dispersed thyroid cell preparations from primary thyrocyte cul- tures as well immortalize d thyroid cell lines had lacked the ability of efficient iodide organification and TH synthesis . Although proto- cols exist to obtain follicle preparations from freshly dissected thy- roid tissue or to reconstitute thyroid follicles under in vitro conditions, these techniques are not readily applicable to a HTS ap- proach and have only recently been used to detect chemical inter- ference with TH biosynthesis (Hornung et al., 2010 ). Until an integrated in vitro testing model for HTS of chemical s interfering with TH synthesis, such as the functional follicles described above, is available, we recomme nd testing the two main aspects of TH biosynthesis , NIS and TPO activities, in specifically tailored in vitro assays that are currently available (Table 2, Box 2).
2.2.1. Endpoint: NIS-medi ated iodide uptake
Active uptake of iodide by thyroid follicular cell is indispen sable for TH synthesis . The pivotal role of NIS in mediating thyroidal io- dide uptake and thus facilitating TH synthesis is highlighted by several key findings. Competitive inhibition of NIS-mediated iodide uptake by specific anions, for example, blocks not only thyroidal iodide uptake but impairs TH synthesis (Alexander and Wolff, 1966; De Groef et al., 2006; Tonacchera et al., 2004; Wolff, 1998). In the human, mutations in the NIS protein are associated with congenital iodide transport defect, a condition characterized by low iodide uptake, hypothyroid ism and goitre (Bizhanova and Kopp, 2009; De La Vieja et al., 2000; Pohlenz and Refetoff, 1999 ).
2.2.2. In vitro assays for NIS-mediate d iodide uptake The classical assay to test the ability of a chemical to interfere
with NIS-medi ated iodide uptake is based on the measureme nt of radioiodine (125I�) uptake in NIS-expressi ng cells (Atterwilland Fowler, 1990; Schmutz ler et al., 2007b ). This type of assay is usually performed with a thyroid cell line (e.g., FRTL5 cells) but non-thyroid cell lines transfected with NIS have also been success- fully used (Lecat-Gu illet et al., 2007 ). Once a chemical shows inhib- itory effects on radioiodine uptake, further analyses are necessary to confirm a specific effect on NIS function and to identify false positives. Some chemicals might even increase iodide uptake be- cause of increased iodide retention resulting from inhibitory ef- fects on iodide efflux rates or interference with pendrin (Eliseiet al., 2006; Lecat-Guille t and Ambroise , 2009 ).
A fully automate d radioiodine uptake assay was develope d for rapid and quantitat ive screening of test chemicals in a 96-well for- mat using HEK293 cells transfected with human NIS (Lecat-Guille tet al., 2007, 2008; Lindenthal et al., 2009 ). This method has been used to screen a chemical library of 17,020 structures (Lecat-Guillet et al., 2008 ). A nonradioacti ve iodide uptake assay was re- cently develope d using FRLT5 cells (Waltz et al., 2010 ). The assay was robust and had a similar sensitivity to the classical radioiodine uptake assay using FRTL5 cells (Waltz et al., 2010 ). Recently also afluorescent assay for cellular iodide uptake was develope d on ayellow fluorescent protein variant, YFP-H148Q/I152L , which is sen- sitive to halide and several other voluminous anions and thus can be employed as a biosenso r to monitor the cellular uptake of nat- ural and EDC NIS substrates. FRTL-5 cells expressing high NIS levels as well as NIS transfected other cells can be used to study transport kinetic parameters such as maximal velocity of NIS-medi ated up- take as well as the rate constant for passive efflux (Cianchettaet al., 2010 ; Di Bernardo et al., 2011 ).
To date, several inhibitors of NIS-mediated iodide uptake have been identified, including inorganic monovalent anions such as perchlorate , thiocyanate , fluoroborates or nitrate (Dohan et al., 2003; Van Sande et al., 2003 ). Due to their similarity to iodide in charge and size, these voluminous anions (perchlorate, etc.) inter- fere with iodide uptake by competitive inhibition of iodide trans-
port by the NIS. Perchlorate itself is also known to be transported by NIS into thyroid cells and is concentrated in the thyroid gland (Clewell et al., 2004; Paroder-Belenits ky et al., 2011 ). This is likely true for other competitive inhibitors of NIS, as well. Recent work using an HTS NIS uptake inhibition assay revealed a novel class of NIS inhibitors, imidazothiazol es (Lecat-Guillet and Ambroise, 2009).
2.2.3. Endpoint: TPO inhibition
To date, inhibitors of TPO activity make a large group of THDC affecting the thyroidal TH synthesis. TPO-inhibiti ng chemical s are a structura lly diverse group of chemical s including thiourea deriv- atives (e.g., propylth iouracil (PTU)), flavonoids (e.g., genistein ),m-dihydroxybenzene derivatives (e.g., resorcinol), tetracycline drugs(e.g., minocycline) and other structurally unrelated chemical s (e.g.,amitrole, kojic acid). Potent TPO inhibitors such as the clinically used thyrostatic drugs PTU and MMI can cause an almost complete inhibition of TH synthesis in vivo reflecting the crucial role of TPO activity for normal TH synthesis to occur.
2.2.4. In vitro assays for TPO inhibition TPO-inhibiti ng chemical s could interfere with any of the dis-
tinct steps of the TPO-catalyz ed iodide organification process and a variety of enzyme assays have been historical ly used to evaluate effects on iodide oxidation, tyrosine iodination and iodotyros ine coupling . Classical assays to evaluate the effects of chemicals on io- dide oxidation use relatively simple spectrophot ometric detection systems to monitor the TPO-catalyz ed oxidation of iodide or alter- native substrates (e.g., guaiacol, azino-bis-(3-ethylbenzothia zo- line-6-su lfonic acid)) in the presence of H2O2. Iodination assays, in turn, measure the TPO-catalyzed iodination of tyrosine to yield MIT and DIT which are usually determined spectrophotom etrically or by chromatographi c techniqu es (Divi et al., 1997; Freyberger and Ahr, 2006 ). The least straightforwar d type of assays are those that aim to specifically assess chemical interfere nce with the coupling reaction (Divi et al., 1997 ). Notably, the existence of dif- ferent molecular mechanism s of TPO inhibition bears important implication s regarding the suitability of different assays to sensi- tively detect TPO inhibitors. For example, many TPO inhibitors like propylth iouracil (PTU), methimazole (MMI) and resorcinol deriva- tives can be readily detected in assays measuring the TPO- mediated oxidation of guaiacol. Still other potent TPO inhibitors such as ethylenethi ourea show no effects in the classical guaiacol oxidation assay but their inhibitory activity can be detected using iodide oxidation or tyrosine iodination assays as readout for TPO interfere nce (Freyberger and Ahr, 2006 ).
Common to all currently available TPO inhibition assays is that they are enzymatic assays performed in a cell-free environm ent. Although the enzymatic assays used to demonstrate TPO inhibition bear the potential for the developmen t of HTS protocols, no such efforts have yet been reported. A major difficulty with regard to TPO inhibition assays is the identification of a stable and reliable source of TPO for systematic screening approaches. Historically, most studies used partially purified TPO preparations from hog and bovine thyroids or from human goitre samples. However, re- cent studies started to validate the use of TPO preparations derived from thyroid cancer lines stably transfected with human TPO expression vectors for use in TPO inhibition assays, the results of which appear very promising (Schmutz ler et al., 2007b ). Currently, the most important endpoints of TH production can only be inves- tigated in separate assays. However , the number of assays for ini- tial screening for THDCs could be reduced with the developmen t of a TH producing in vitro follicular system. When an effect is ob- served in such an integrated system, underlying mechanisms could

1330 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
then be further investigated in a second tier of specific in vitro assays.
2.3. Box 3. Transport of thyroid hormones
After TPO catalyzed production, TH still remain integral parts of thyroglobul in deposited in the lumen of the thyroid follicle. Next, thyroglobul in molecules are internalized into the thyrocytes by pinocytosis and intracellular ly degraded by cathepsin s (Brixet al., 2001; Friedrichs et al., 2003 ). The TH are subsequent ly ex- ported via TH transporters (e.g., MCT8) across the basolateral thy- rocyte membrane into the circulation (Di Cosmo et al., 2010; Trajkovic-A rsic et al., 2010; Wirth et al., 2011 ). In the blood, most of the TH are bound to TH binding proteins, leaving approximat ely 0.03% of T4 and 0.3% T3 unbound, the ‘free’ hormone fraction (Yen,2001). In humans, the protein with highest affinity to TH is TBG, which carries the majority of the TH in the circulation. The role of TBG in TH transport is not only species-, but also age specific,since it is undetectabl e in adult, but present in serum of young, pregnant and obese rats (Savu et al., 1987, 1989 ). The second high-affinity TH binding protein is TTR (reviewed by Blake et al. (1978), Robbins (2000), Schreiber (2002) and Schussler (2000)).Albumin is the most abundant TH-binding protein, but the binding is unspecific and with low affinity (Schussler, 2000 ).
TTR is synthesized in liver, brain, pancreas, retina (Buxbaumand Reixach, 2009 ) and placenta (Mortime r et al., 2012 ), and is in- volved in the transport of T4 and retinol across the blood–brain barrier and to the fetus from the placenta. TTR may be an impor- tant target for THDCs, as many hydroxylated metaboli tes of persis- tent organic pollutants show high binding affinity for TTR that can be even higher than those of the endogenous ligand T4 (Ghoshet al., 2000; Hamers et al., 2006; Lans et al., 1993; Meerts et al., 2000a). Inhibitio n of T4-binding to TTR results in disruption of the TTR-retinol binding protein complex followed by reduced plas- ma levels of TH and reduced retinol levels.
2.3.1. Endpoint: Binding to transport proteins TTR and TBG
Given the relevance of TTR and TBG for transport of T4 in the plasma, and the relevance of TTR for T4-transport over the placen- tal and blood–brain barriers, competit ion of THDC for binding to these transport proteins should be included in a comprehensive THDC test battery. In vitro evaluation of TBG binding should also be considered when in vivo rodent studies are used, as TBG-bindin gis not assessed in rodent studies. Binding to albumin is unspecificand no competition for TH binding by xenobiotics has been re- ported. Apart from TH binding proteins TTR, TBG and albumin, TH transport over the placental and blood–brain barrier as well as across other membran es strongly depends on their expression levels of various TH transporters (MCT8, MCT10, OATP1c, LAT1, LAT2, etc. for transport and export of TH.
Several halogenated phenolic mono- and polycyclic chemicals such as hydroxylated PCB or PBDE metabolites (Brouwer et al., 1990; Gutleb et al., 2010; Hamers et al., 2006; Lans et al., 1993, 1994; Marchesini et al., 2008; Meerts et al., 2000a ) or natural and synthetic (iso-)flavonoids (Lueprasitsakul et al., 1990; Radovic et al., 2006 ) have been shown to bind to TTR with high affinityresulting in displacement of T4. In contrast to TTR, the interference of xenobiotics with binding of TH to TBG has not been studied as intensively (Cao et al., 2010; Marchesini et al., 2006, 2008; Meerts et al., 2000a ).
In addition to TTR, placental TH plasma membrane transporters play a role in the transplacen tal passage of TH from mother to fe- tus, including the TH transporter s MCT8, MCT10, L-type amino acid transporter (LAT) LAT1, LAT2, organic anion transporting peptide (OATP) OATP1A2 and OATP4A1 (Loubiere et al., 2010 ). In the brain
and in the blood brain barrier, two important transporter mole- cules are MCT8 and OATP1C1. MCT8 is also expressed in heart, kid- ney, liver, and skeletal muscle (Roberts et al., 2008 ).
The relevance of cell membrane transporters and assays for this endpoint are further discussed in Box 5 below on local cellular concentr ations.
2.3.2. In vitro assays for binding to transport proteins TTR and TBG Several TH binding assays have been developed and published,
most of them applying radioactive T4 displacemen t from TTR and to a lesser extent from TBG (Brouwer and van den Berg, 1986; Cheek et al., 1999; Hallgren and Darnerud, 2002; Hamers et al., 2006; Lans et al., 1994 ). An alternativ e method is the detection of the TH transport disruptors with a biosensor based on the sur- face plasmon resonance (SPR) technique (Marchesini et al., 2006 ).SPR biosenso rs such as Biacore measures binding as a change in the refractive index at the surface of the sensor caused by accumu- lation of mass within the surface layer which in this case is loaded with T4 to which TTR or TBG can bind (Marchesini et al., 2006, 2008). This method is fast and can be adapted to analyse the inter- action of THDC with multiple TH binding sites in parallel. Recently a 96-well non-radioactiv e fluorescence displacement assay has been developed based on competition of T4-like chemicals with the fluorescent probe ANSA 8-anilino-1- naphtalenesulfon ic acid ammoniu m salt) (Montano et al., 2012 ). An alternativ e new devel- opment makes use of immunoma gnetic microbeads followed by screening with flow cytometry and identification of THDCs with nano-liqu id chromatograp hy mass spectrometry (Aqai et al., 2012 ).
2.4. Box 4. Metabolism and excretion
The major pathways of TH metabolism resulting in activation, inactivati on or excretion from the body are (1) deiodination, (2)alanine side chain modification, (3) sulfation and (4) glucuronida -tion. Deiodination is the most important metabolic pathway in both quantitative and quantitative terms for normal physiological control of TH homeostasis. THDCs could interfere with both the activity of enzymes as well as the production of the enzymes by inhibiting or inducing their expression.
Inclusion of catabolic enzymes and cellular transporters , e.g., UGTs, SULTs, OATPs, MCT, and multi-drug efflux transporters, as molecula r targets for endocrine-disrup ting chemicals is clearly warranted due to the critical role these proteins play in regulating concentr ations of circulating thyroid hormones. Indeed, there are many published reports that correlate upregulation of these pro- teins with increased biliary eliminati on of glucuroni dated or sul- fated T4 or T3 following exposure to xenobiotics (see for example, Christenson et al., 1995, 1996 ), and/or to systemic decreases in thyroid hormones . Xenobiot ic nuclear receptors including AhR, CAR, PXR, and PPAR transcrip tionally regulate the expression of these proteins, and many endocrine-disrup ting xenobiotics are known to activate these receptors (Boas et al., 2006; Crofton, 2008; Kretschm er and Baldwin, 2005; Miller et al., 2009; Timsit and Negishi, 2007; Zhang et al., 2004 ). For example, activators of CAR or PXR are known to upregula te transcrip tion of a wide variety of Phase I–III enzymes, which increase hepatic catabolism and sometimes bioactivation of not only pharmaceutical s (Kretschm er and Baldwin, 2005; Sinz et al., 2006 ) and other chemicals, but also endogen ous thyroid hormones (Visser, 1996 ). Some of the relevant Phase II enzymes capable of thyroid hormone metabolism include UGT1A1, UGT1A6, and SULT2A1 (Kato et al., 2008 ). Phase III influxand efflux transporters including, but not limited to, MCT8, OAT- P1A4, and MRP2, may also be important for increased excretion of thyroid hormones and its conjugat es, thus enabling an overall increase in systemic clearance (Lecureux et al., 2009; Morimoto et al., 2008; van der Deure et al., 2010; Visser et al., 2010; Wong

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1331
et al., 2005 ). Therefore, any in vitro screening assay battery for thy- roid-disrupti ng chemicals should include biomarkers of nuclear receptor activation, because they represent upstream biological targets that indicate potential for modulation of the regulation of hepatic (and other tissues) proteins important for thyroid hormone homeostasi s (Sinz et al., 2006 ).
There are already several HTS assays for nuclear receptors (Huang et al., 2011; Kavlock et al., 2012; Raucy and Lasker, 2010; Schoonen et al., 2009 ) that are relevant for other toxic pathways as well. Therefore they will not be reviewed herein. Importantly, data from these assays must be included in any systems ap- proaches to determine the actions of chemical s on thyroid hor- mone pathways . Additional measurement of expression and/or activity UGT, SULT, or transporter s would serve as a downstream confirmation of one or more chemical -nuclear receptor interac- tions, and also an important marker of potential increases in thy- roid hormone turnover.
2.4.1. Endpoint: Deiodinat ion
In most species, T4 is the main TH secreted by the thyroid gland under normal conditions. The bioactive hormone T3 is largely pro- duced by enzymatic outer ring deiodination (ORD) of T4 in periph- eral tissues. Inner ring deiodination (IRD) of T4 produces the inactive metabolite rT 3. T3 is degraded by IRD to 3,3 0-T2, an inactive TH metabolite which is also generated by ORD of rT 3. Three iodo- thyronine deiodinases (DIO1, DIO2, DIO3) are involved in the deio- dination of TH. DIO1 is expressed predomin antly in the liver, the kidneys and the thyroid (Gereben et al., 2008 ). It catalyzes the ORD and/or IRD of a variety of iodothyronine derivatives, but it is most effective in the ORD of rT 3. Hepatic DIO1 is a major site for the production of plasma T3. The expression of DIO1 which codes for DIO1 is positively regulated by T3 at the transcriptio nal level. DIO2 is expressed primarily in brain, anterior pituitary, brown adi- pose tissue (BAT), thyroid and skeletal muscle (Gereben et al., 2008; Larsen, 2009 ). In brain, DIO2 is expressed predomin antly in astrocytes. DIO2 only catalyzes the ORD of iodothyronines, with a preference for T4 > rT 3. DIO2 plays an important role in the local production of T3 from T4 in brain, pituitary and BAT. DIO2 ex-pressed in the anterior pituitary and hypothalamus is important for the negative feed-back of T4 on TSH and TRH secretion. In gen- eral, DIO2 activity is negatively regulated by TH at the posttransla- tional level (Gereben et al., 2008 ). DIO3 is expresse d in different human tissues, i.e. brain, skin, liver, and intestine; DIO3 activities are much higher in fetal than in adult tissues (Gereben et al., 2008). DIO3 is also abundantly expressed in the placenta and the pregnant uterus. DIO3 has only IRD activity and catalyzes the inac- tivation of T3 and T4 and is thought to protect the developing fetal tissues against undue exposure to T3. Hence, fetal serum T3 levelsare low. DIO2 and DIO3 show complex spatio-temp oral expression patterns in fetal and neonatal brain, which are critical for optimal brain developmen t. In the brain, DIO3 expression is stimulated by T3, probably at the transcriptio nal level (Gereben et al., 2008 ).Thus, tissue specific expression of deiodinases and regulation of DIO expression through HPTP feedback mechanis ms are important regulators of TH activity in tissues.
2.4.2. Endpoint: Alanine side chain modification
Iodothyroni nes are also metaboli zed by decarboxylati on of the alanine side-chain to iodothyrona mines which could have acute and dramatic effects on heart rate, body temperat ure and physical activity (Scanlan, 2009 ). Presumably by further conversio n of iod- othyronami nes, the iodothyroac etic acid metabolites Tetrac (TA4)and Triac (TA3) are generated (Wood et al., 2009 ). TA 3 has signifi-cant TH activity (Freitas et al., 2011 ), and TA 4 is a strong inhibitor
of the non-genom ic effects of TH exerted at the cell membrane through binding to the avb3 integrin (Davis et al., 2011 ). There is, however, little or no information that environmental chemicals induce or inhibit alanine side chain modification of iodothyroni nes, therefore we do not suggest including this endpoint in an in vitro THD test battery.
2.4.3. Endpoint: Sulfation
Iodothyr onines are phenolic chemicals and thus readily metab- olized by conjugat ion of the phenolic hydroxyl group with sulfate or glucuroni c acid. Sulfation appears to be a primary step leading to the irreversibl e inactivation of TH (Kester and Visser, 2005; Wu et al., 2005 ). The IRD of T4S to rT 3S and of T3S to 3,3 0-T2S are orders of magnitude faster than the IRD of non-sulfa ted T4 andT3, whereas ORD of T4S to T3S is completely blocked. Normally, plasma levels of iodothyronine sulfates such as T4S and T3S are very low, as these conjugates are rapidly deiodinated by D1 by IRD. Plasma T4S and T3S are increased if D1 activity is inhibited during fetal development, non-thyroidal illness and fasting. Phenol sulfotran sferases (SULT) with significant activity towards iodothy- ronines include human SULT1A1, 1A2, 1A3, 1B1 and 1C2. They show substrate preference for 3,3 0-T2� T3, rT 3� T4. Iodothyr o- nines are also sulfated by human estrogen sulfotransferase (SULT1E1), in particular T4 (Kester et al., 1999 ). Drugs such as PTU can inhibit D1-activity, while in rodents it has been shown that phenoba rbital may accelerate TH metabolism by induction of sulfotransfe rases through activation of the constitutive andro- stane receptor. The importance of this mechanism in humans is still unknown (Qatanani et al., 2005 ).
2.4.4. Endpoint: Glucuronidation
In addition to sulfation, iodothyronines are metabolized by con- jugation of the phenolic hydroxyl group with glucuronic acid. Iod- othyroni ne glucuronides are rapidly excreted in the bile, constituti ng the first step in the enterohepatic cycle of TH (Visser,2008; Wu et al., 2005 ). In humans, about 20% of daily T4 productionappears in the faeces, probably through incomplete intestina lhydrolysi s and reabsorption of the biliary-excr eted conjugates. Glucuroni dation of T4 and T3 is catalyzed by different members of the UDP-glucuronyl transferase 1A family, i.e. UGT1A1, 1A3, 1A7-10 (Kato et al., 2008; Tong et al., 2007 ). Usually, this involves the glucuronida tion of the hydroxyl group, but human UGT1A3 also catalyzes the glucuroni dation of the carboxyl group of iodo- thyronin e derivatives.
In rodents, T4 metabolism is accelerated by different classes of chemical s, including barbiturates , fibrates, PCBs and PBDEs, which is associated with the induction of hepatic T4 UGT activities (Morseet al., 1996; Hallgren et al., 2001; Hood et al., 2003; Visser et al., 1993; Zhou et al., 2001 ). In rodent studies, induction of UGT activ- ity is a commonly proposed mechanis m of interference for a num- ber of environmental contaminan ts (Axelrad et al., 2005; Hurley, 1998). This may result in hypothyrox inemia or hypothyroidism if the thyroid gland is not able to compensate for the increased hor- mone loss. In humans, hypothyroxi nemia may occur via induction of T4 glucuroni dation by anti-epilept ics, but overt hypothyroidism is rare (Benedett i et al., 2005 ).
2.4.5. Inhibition of enzyme functioning
The deiodination of iodothyronines by the different deiodinases is inhibited by a variety of structurally related chemicals, including iodinated X-ray contrast agents, amiodarone and its metaboli tes, and OH-PBDEs (Butt et al., 2011; Köhrle, 2011 ). However, the iod- othyroni ne deiodinas es have markedly higher affinities for iodin-

1332 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
ated phenols than for brominated or chlorinat ed phenols, suggest- ing that environm ental levels of OH-PBDEs and OH-PCBs will affect TH deiodination less if at all (Butt et al., 2011 ) (T.J. Visser, unpub- lished work). The thyrostatic drug PTU potently inhibits DIO1 by reacting with a selenenyl iodide intermedi ate but it has no effect on DIO2 and DIO3 (Gereben et al., 2008 ). The sulfation of iodothy- ronines by different SULTs is inhibited by OH-PCBs , OH-PDBEs and other hydroxylate d metaboli tes of polyhalogenated aromatic hydrocarbo ns (OH-PHAHs) (Kester et al., 2002; Schuur et al., 1998b). This is especially the case for the estrogen sulfotransfe rase SULT1E1, which is potently inhibited by a variety of OH-PHAHs.
2.4.6. In vitro assays for induction and inhibition of enzymes for TH metabolism
Both enzyme induction and inhibition of enzyme activity can have important implication s for the availability of TH. The induc- tion of enzymes can only be tested in vitro in cell-based assays, the inhibition of enzyme activities in cell lysates containing these enzyme activities or isolated liver enzymes (Schuur et al., 1998a,b,c; Hamers et al., 2008 ). Since Phase II and III metabolism are partially controlled by specific nuclear receptors, assays for these receptors could be used as surrogates for induction of the en- zymes (Kavlock et al., 2012 ).
While there are many published methods that have been used to measure expression of UGT and SULT mRNA and activity (Biegelet al., 1995; Brucker-Davis, 1998; Hood et al., 2003; Hood and Klaassen, 2000a,b; Liu and Klaassen, 1996; Paul et al., 2012 ), there are only limited reports of the use of medium or HTS assays for measuring upregulation of these enzymes. Rotroff and colleagues (Rotroff et al., 2010 ), using quantitative nuclease protection assays (qNPA) for UGT1A1, SULT2A1 screened 309 chemicals as part of ToxCast Phase I (Sinz et al., 2006 ).
There is a clear need to improve and develop high-thro ughput assays for inhibition of Phase II enzymes that are involved in TH metabolism.
2.5. Box 5. Cellular concentrations and transporters
Cellular TH uptake is mediated by specific plasma membrane transporters which may differ between cells from different tissues such as liver, pituitary heart, placenta and blood brain barrier and showing preferential transport of T4 or T3 or their metabolites (Hennemann et al., 2001 ). Using transporter-speci fic ligands and inhibitors, it has been demonstrated that TH uptake by different cell types is mediated at least in part by amino acid transporter sshowing preference for large, neutral amino acids (L-type) or aro- matic amino acids (T-type). A number of transporters capable of transporting TH have been identified, including different members of the OATP family, 2 L-type amino acid transporter s (LAT1,2), and 2 members of the monocarboxy late transporter (MCT8,10) family (Butt et al., 2011; Friesema et al., 2005; Köhrle, 2011; Visser et al., 2008 ).
The human OATP family consists of 11 members, many of which have been shown to transport iodothyronine derivatives (Hagenbuch, 2007; Obaidat et al., 2012 ). OATPs are glycoprotei ns and the human OATP1 subfamily contains four members (OAT-P1A2, 1B1, 1B3, 1C1) that are capable of transporting a variety of anionic, neutral and even cationic ligands. OATP1A2, 1B1 and 1B3 show preference for sulfated over non-sulfated iodothyronines as ligands (van der Deure et al., 2009 ). OATP1A1 is expresse d in dif- ferent tissues but OATP1B1 and 1B3 are expressed only in liver. OATP1C1 shows a high preferenc e for T4 as the ligand and is almost exclusively expressed in brain capillaries and choroid plexus, sug- gesting that it is very important for T4 transport into the brain (Hagenbuch, 2007; Obaidat et al., 2012 ).
T4 and T3 are also transported by 2 members of the hetero- dimeric amino acid transporters: LAT1 and LAT2 (Taylor and Ritchie, 2007 ). Both LAT1 and LAT2 facilitate the bidirectional transport of a variety of aliphatic and aromatic amino acids as well as iodothyronines over the plasma membrane.
The MCT family contains 14 members. MCT8 (SLC16A2) and MCT10 (SLC16A10) have recently been identified as important TH transporters which can be affected by THDC (Braun et al., 2012; Friesema et al., 2006b, 2008; Halestrap, 2012; Kinne et al., 2010). So far, only iodothyronines have been identified as trans- ported substrates for MCT8 but MCT10 also transports aromatic amino acids (Trp, Tyr, Phe). They are non-glycosylated proteins showing high homology and wide but different tissue distribu- tions. In brain, MCT8 is importantly expressed in choroid plexus, capillaries and neurons in different brain areas. MCT8 is thought to be critical for T3 uptake in central neurons and, thus, for the ac- tion of TH during brain developmen t. Patients with mutations in MCT8 suffer severe psychomotor retardation in combination with highly elevated serum T3 levels (Friesema et al., 2006a ).
2.5.1. Endpoint: cellular TH uptake
An important endpoint to include in an HTS battery is inhibition of cellular uptake of iodothyronines, in particular of T4 and T3. TH uptake experime nts are usually done by incubation of radioiodine -labelled TH with cells transiently or stably transfected with atransporter such as MCT8, MCT10 or OATPC1. After incubation and washing, cells are lysed and counted for radioactivi ty (Friesema et al., 2006a, 2008 ), The assay may be adapted to measure the facilitation of intracellular TH metaboli sm in cells co-transf ected with transporter and one of the deiodinas es (Friesema et al., 2006a, 2008 ). However, to our knowledge HTS assays have not been develope d for the identification of chemical sthat inhibit cellular TH transport. The development of transporter assays is a recommended area for assay developmen t.
2.6. Box 6. Cellular responses
The best understood cellular actions of TH are carried out via receptors found constituti vely in the cell nucleus. Ligand-activ ated receptors directly control the transcrip tion of specific sets of target genes that carry out cell and developmen tal stage specific re- sponses. There are two well-conserved TH receptor gene loci in all vertebrates , designated TR a and TR b. Several TR isoforms gener- ated by alternative splicing and use of different initiation sites are known and expressed in a tissue specific manner (i.e. TR a: TR a1and the non-hormone binding TR a2; TR b: TR b1 and TR b2). These have been reported in various species and cell types with both overlappi ng and specific actions reported (reviewed in Flamantand Gauthier (2012)). The nuclear TRs control transcription of spe- cific genes directly through binding to thyroid hormone response elements (TREs). Often, the TRs are found in a complex with the related retinoid-X receptor; in the heterodimer complex with TRs the RXR is usually a ‘‘silent partner’’ but in some cell types and/ or TRE configurations it may be activated by various retinoids (Castillo et al., 2004 ). In the absence of ligand, co-repres sor com- plexes repress transcription and upon ligand binding, co-represso rcomplexes are replaced by co-activator complexes that initiate transcrip tion (Rosenfeld et al., 2006; Shi, 2009 ). Most of the studies of TR action have examined up-regulate d genes, since far less is understo od about direct ligand induced down-regulati on. How- ever, understand ing the mechanism of down-regul ation is impor- tant for understanding negative feedback on TRH and TSH expression, a common endpoint for TH disruption assays in intact animals.

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1333
Recently, TH have been shown to also directly act both at the cell membrane, in the cytoplasm (Cheng et al., 2010 ) and in mito- chondria as well (Chocron et al., 2012; Pessemesse et al., 2012 ),which is of interest given the important influence of TH on mito- chondrial function in warm-blooded vertebrates. Often, these effects are referred to as ‘‘nongenomi c’’ vs. the nuclear receptor- mediated ‘‘genomic’’ effects described above, although nongenom- ic actions may ultimately influence the transcriptio n of specificgenes, and interface with nuclear TR actions as well. The actions of TH at the cell membrane, in the cytoplasm or in mitochondria, may be mediated by a bona fide TR isoform acting outside the nu- cleus, an amino terminal truncated form of the TR a, or a distinct receptor for TH altogethe r. For example T4 (and to some extent T3) can stimulate angiogenesis in various models via a membran ereceptor (integrin avb3) mediated signalling pathway. T4 has been shown to bind directly to the avb3 integrin and activate down- stream signalling including induction of genes linked to blood ves- sel formation (Cheng et al., 2010 ). In addition, TR b1 has been reported to bind to PI3 kinase in the cytoplasm, inducing down- stream responses including gene activation linked to responses to hypoxia, such as HIF1 a (Moeller et al., 2005 ), although a classical ‘‘genomic’’ pathway to HIF1 a induction has also been proposed (Otto and Fandrey, 2008 ). Clarification of relative importance of these alternate modes of action in the homeostatic and develop- mental roles for TH will be important to determine whether these are relevant targets of concern for TH disrupting chemical s and as- say developmen t.
2.6.1. Endpoint: TR binding and transcriptiona l activity
While most TH disruption has been shown to occur at pre- receptor steps as discussed, the TRs are the point in the response pathway closest to the endpoints that result in altered cellular re- sponses: the proper transcriptio nal control of specific sets of genes. The relatively snug fit of T3 in the TR ligand binding domain only translates to those chemicals with a structure closely resembling that of the hormone itself and which specifically interact with the TR ligand binding domains with reasonable affinities (DeVitoet al., 1999; Zoeller, 2005 ). Examples are T3-like OH-PCBs and OH-PBDEs (Freitas et al., 2011 ). A functional consequence of ago- nist binding to the TR ligand binding domain is co-activators recruitment , and the few antagoni sts that have been described for TRs act at least in part via disruption of this interaction (Webbet al., 2002 ). In principle, assays that measure coactivator vs. core- pressor interactions may have more predictive value than simple ligand binding. Both cell free and cell based assays are amenable to HTS of TR-coregula tor interactio ns (Table 2). For the nuclear TRs, however, measurement of their transcriptional regulatory activity in their native environment at target genes, including in native chromati n context, should be most relevant for predicting THD acting via this pathway.
It should also be emphasized that multiple means for disrupting TR function are possible beyond direct binding to the receptors and altering coactivator/ corepressor interactio ns, given the complexi- ties of transcriptio nal control by these receptors. Interactions with RXRs may affect a subset of important TH target tissues or specific genes in specific cells. Further, the TRs themselv es and various coactivator and corepress or proteins are subject to post- translationa l modifications such as phosphoryla tion that may be a target for endocrin e disruption. TR and coactivator stability, nuclear localization , and interactions with other cofactors may be additional targets of THD chemical s. Arsenic trioxide exposure, for example, activates a MAP kinase cascade that leads to phos- phorylation of the corepressor SMRT and its dissociat ion from its nuclear receptor partners including the TRs, leading to de- repression of target genes (Hong et al., 2001 ).
Ultimatel y, changes in gene expression mediated by the nuclear TRs result in observable cellular phenotypes that are specific for agiven target cell type. For example, the rat pituitary tumour GH 3cell line proliferates in response to T3. T3 treated primary cultures of cerebella r Purkinje cells show increased dendritic branching (Heuer and Mason, 2003; Ibhazehiebo et al., 2011 ), cardiomyocytes self-organized into three dimensional heart muscle alter their con- tractility properties (Khait and Birla, 2008 ), and primary liver cells show altered expression of metabolic enzymes (Attia et al., 2010; Hwang et al., 2011 ) as observed in vivo . Although primary cell as- says may have more predictive value for effects in vivo than any of the other models discussed here, animals would still be used as source material and non-TR mediated mechanisms make the mode of action less certain. Miniaturizing the primary cell assays, or expansion of the primary cell population prior to assay, would greatly reduce the number of animals needed, and developmen tof sensitive and specific assays for sets of key, direct TR target genes in these cell lines may help with mode of action uncertainty. For Tier 1 HTS, however , this is not ideal with current technology.
Finally, as more is learned about ‘‘non-genomic ’’ TH action at the membrane, in the cytoplasm, or in mitochondr ia, in vitro assayswill need to be developed for these endpoints as well, but at the moment the relevance of non-genom ic pathways in THD remains unclear.
2.6.2. In vitro assays for detecting disruption of TR activity Ligand binding assays. Several ligand binding assays have been re- ported to detect chemicals binding to the TRs in vitro (Kitamuraet al., 2002, 2005a; You et al., 2006 ). Typically, these assays involve over-expr essed TR ligand binding domains or intact proteins in bacteria or are obtained from mammali an cell extracts, and detect the efficiency of displacemen t of radiolabel ed T3. The advantag es are speed and relative simplicit y, and the ability to demonst rate a direct interaction of a TH disrupting chemical and an important defined target. Further, since these are typically cell free assays, there are no confounding issues with cytotoxicity of the chemical in question. Beyond the less desirable use of radioiodine -labelled ligands, the primary disadvantag es are that binding does not pre- dict in vivo agonism or antagonism per se , and isolated receptors or receptor fragments are not in their native environment where proper folding or stability may be quite different. For example, re- cent studies with the glucocorti coid receptor have demonstrat ed an important allosteric effect of various response element se- quences on ligand binding that has not been widely appreciated for other receptors (Meijsing et al., 2009 ). Improveme nts in in silico methods to predict ligand binding of the TRs are an important complemen tary approach to the direct binding assays (Park et al., 2010).
Coregulat or interaction assays. Another related approach uses the recruitment of interaction domain peptides derived from nuclear receptor coactivators or corepressor s to the TR ligand binding do- main. These peptides are labelled and recruitment or displacemen tcan be followed by fluorescence polarization (Levy-Bimbot et al., 2012); furthermore , such assays have been adapted for HTS (Johnson et al., 2011 ).
Reporter gene assays. To be of highest concern, THDCs should alter TR transcriptio nal activity in a cellular context. To address this question transient and stable transfection assays are deployed to develop TRE driven reporter gene assays. Reporter gene assays for TR activity have taken advantage of cells with endogen ous receptor expression (Freitas et al., 2011; Sugiyama et al., 2005b ),co-transf ected specific TR expression vectors with the TRE based reporters (Jugan et al., 2007; Freitas et al., 2012b ), or co-transf ected gal4 TR expression vectors and UAS (upstream activating

1334 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
sequence) based reporters (Huang et al., 2011; Invitrogen, 2010; Tiefenbach et al., 2010 ) of which the response is relatively more specific for TRs so for example RXR activation would be less likely to contribute to a Gal4 based assay. T3 induces reporter gene activ- ity from a low baseline and the assay can be fairly sensitive and reproducibl e. Multiplexing responses with a reporter for non- receptor specific cytotoxicity can enhance the assay efficiency(commercial assays from Promega, and Indigo, no scientific publi- cations yet), especially when confirming antagonistic action. Tran- sient transfecti on allows flexible analysis of multiple reporter configurations, multiple receptor isoforms and in various cell lines and has been used to demonstrate the effect of chemical s like BPA, TBBPA, and various PCB and PBDE variants and their metabolites on TR transcrip tional activity (reviewed by Zoeller (2005)), and some other examples are listed here (Hofmann et al., 2009; Ibhazehiebo et al., 2011; You et al., 2006 ). A limitatio n of transient transfection assays for the use in HTS is that the transfection of DNA has to be performed again for each experiments which is time-consumi ng and results in relatively great variation s in re- sponses. Another disadvantag e is that the reporter gene in a tran- siently transfected cell is not fully wrapped in chromatin like endogenous genes are; this may be an important distinction with stably transfected cells given the TR’s role in modifying chromati nas part of its transcriptio nal repression/a ctivation properties. Therefore stably transfected reporter gene assays are much more suited for HTS applicati ons. Full-leng th endogen ous or co- transfected receptors have the advantag e of the full range of interactions with a native response element configuration and co-regulator y proteins, while the gal4 fusion system may be some- what more specific and show a wider dynamic range of regulation. Yeast based reporter assays have also been reported for full length TRs or to measure TR ligand binding domain-coacti vator interac- tions (Arulmozhiraja et al., 2005; Shiizaki et al., 2010 ), but there have been concerns raised about ligand availability in those sys- tems and the absence of a full complement of the TH signalling pathway and the typically minimal nature of the reporter gene reg- ulatory elements.
Induced cellular phenotypes. While further developmen t of primary cell based assays is desirable, to date few continuous cell-line- based assays for THD have been develope d. One is the so-called ‘‘T screen’’ that relies on a T3 induced proliferative response in rat somatotr ophic tumour cell line GH 3 (Ghisari and Bonefeld-Jor- gensen, 2009; Gutleb et al., 2005; Schriks et al., 2006 ). The assay has shown the ability to detect agonistic and antagoni stic chemi- cals presumably acting via a TR mediated mechanism (Table 2).
In terms of assay development and validation for TR binding and transcriptional activation endpoints, beyond using the native ligands T3 and T4, several synthetic thyromimetics have been de- scribed that are quite useful for this purpose. For example, the T3-analogue GC-1 is a TR b selective iodine free thyromimeti c with high affinity and works well across species (Chiellini et al., 1998; Furlow et al., 2004 ). Tetrac is another interesting chemical for con- sideration since it induced as a weak agonist the nuclear TR (Freitaset al., 2011 ), possibly after it is deiodinated to Triac in the cell, but Tetrac itself acts as antagonist at the avb3 integrin membrane receptor (Davis et al., 2009 ). Unfortunate ly, few antagonists have been developed for the TRs; nevertheless , NH-3 is an interesting chemical that induces release of corepressor but blocks coactivator recruitment to the TR ligand binding domain (Nguyen et al., 2002 ).However, in these in vitro assays (and also in vivo ) the endogenous level of T3 available for TR binding (i.e. hypo-, eu-, or hyperthyroid condition) will influence whether some of the T3 analogue s act as agonists or antagoni sts as illustrated by the case of NH 3 (Figueiraet al., 2011; Grover et al., 2007 ). The hydroxylated versions of PCBs and PBDEs are most active in TR binding and in cell based transac-
tivation assays (Freitas et al., 2011; Gauger et al., 2007; Li et al., 2010; You et al., 2006 ). A summary of suggested chemicals is found in Table 3.
3. Chemical bio-activa tion and availability in vitro
In addition to chemicals directly interfering with endpoints of the TH system, other potential THDCs such as PCBs and PBDEs have to undergo metabolic activation to become a THD hazard. This bio- activation into hydroxylated chemicals occurs in vivo by Phase Imetaboli c enzymes (Marsh et al., 2006; Morse and Brouwer, 1995). These OH-metabolite s can show remarkable structura l sim- ilarities with TH thereby causing direct effects on the TH homeo- stasis by mimicking T4 and to a lesser extent T3 (Freitas et al., 2011; Meerts et al., 2002 ). As was explained above, this can inter- fere with T4 and T3 transport, action, feedback mechanis ms and metaboli sm. In mammals including humans, OH-metabolite s, bound with high affinity to specific transport proteins like TTR and TGB (Gutleb et al., 2010; Lans et al., 1994; Marchesini et al., 2006; Meerts et al., 2000a ), can accumulate in blood plasma reach- ing levels similar or even much higher concentr ations than of the respective parent chemicals (Berg et al., 2010; Bergman et al., 1994; Gebbink et al., 2008a,b; Sjodin et al., 2000 ) Montano et al., submitte d for publication . In addition, the TH-like OH-meta bolites can be selectively transported over barriers such as the placenta and blood brain barrier (Meerts et al., 2002; Morse et al., 1993 ).
Given the high plasma levels and the toxicological relevance of themetabolite effects, it is very important to include quantification of thetoxic potencies of the OH-chemicals for toxicological hazard assess-ment. However, cell-based in vitro test systems almost all lack PhaseI metabolism. Therefore, this bio-activation step needs to be addedfor example by using in vitro biotransformation techniques withmicrosomal fractions, although biotransformation efficiencies, canbe low and matrix components co-extracted from the metabolizingsystem may interfere with the assay (Hamers et al., 2008; Meertset al., 2000a; Schriks et al., 2006). Recently a new method has beendeveloped to separate the OH-metabolites from the parent followedby a non-destructive clean-up to remove potentially interfering ma-trix chemicals such as microsomal fatty acids from the extract (Mont-ano et al., 2012). Alternatively, an in vitro system can be exposeddirectly to the metabolite of interest, if known and available.
Another important issue to take into account is the availabili ty of the chemicals in the test conditions. In cell-based assays with at least 5% serum lipophilic chemicals will remain available for the cell (Li et al., 2010; Weiss et al., 2009 ), although it also is argued that when they bind to serum proteins their bioavailabili ty for cells might decrease. In tests without serum or other carrier proteins or lipids relatively lipophilic chemical s may be lost to the plastic of the test plates. This can also be an issue when chemicals are added to aqueous test media before proteins such as enzymes or recep- tors are present. Depending on their chemical nature, chemical scould also be lost during exposure due to oxidation or light- induced degradation. These issues should get ample attention in the validation phase of every in vitro bioassay to prevent overesti- mation of the exposure concentr ation.
Finally it is important to characteri ze the phase 2 and cellular efflux mechanisms in cell based systems, as hydroxylate d chemi- cals may undergo rapid phase 2 metabolism and excretion from the cells resulting in an overestimation of the potency (Vandenb erg et al., 2010 ).
4. Suggested testing strategy
The goal of this review was to identify a mechanism -based battery of non-animal tests that can be applied for THD hazard

Table 3Examples of chemicals reported to alter critical elements of thyroid hormone regulation or action. This list is not exhaustive, nor all the results are critically checked in detail.
Thyroid regulation target Effect on thyroid regulation or action
Examples of active chemicals References
TRH receptor signalling Interfere with TSH production by thyrotrophic cells
Somatostatin analogues and synthetic rexinoids Haugen (2009), Sharma et al. (2006) and Zatelli et al. (2010)
TSH receptor signalling Decreased stimulation of thyrocytes
DDT, Aroclor 1254; plant extracts (e.g.3,4-dihydroxycinnamic acid derivatives)
Auf’mkolk et al. (1985) and Santini et al. (2003)
NIS-mediated iodide uptake
Decreased thyroidal synthesis of T3 and T4
Perchlorate, nitrate, thiocyanate, 4-methylbenzylidene-camphor, imidazothiazoles
Lecat-Guillet and Ambroise (2009), Schmutzler et al. (2007b), Van Sande et al. (2003) and Wolff (1998)
TPO inhibition Decreased thyroidal synthesis of T3 and T4
Methimazole, propylthiourea, amitrole, mancozeb, soy isoflavones, benzophenone 2, 1-methyl- 3-propyl-imidazole-2-thionem, kojic acid, 4-nonophenol
Biegel et al. (1995), Capen (1997), Doerge and Sheehan (2002), Higa et al. (2002), Jarry et al. (2004)and Schmutzler et al. (2007b)
Binding to serum transport proteins TTR and TBG
Decreased serum levels Hydroxy-PCBs and -PBDEs, EMD 49209; pentachlorophenol, HCB, phenols
Lans et al. (1993), Marchesini et al. (2008), Meerts et al. (2000b), Montano et al. (2012), Schröder-vander Elst et al. (1997) and van den Berg (1990)
Cellular TH transporters Altered cellular availability of TH for target cells, metabolism, action and elimination
Antidepressants, amino acids, aromatic dyes, flavonoids
Kinne et al. (2010) and Roth et al. (2010)
Inhibition of deiodination enzyme activity
Decreased peripheral synthesis of T3
FD&C Red Dye #3, PTU, PCBs, octyl- methoxycinnamate, 4-methylbenzylidene- camphor, iodinated X-ray contrast agents, halogenated aromatic chemicals
Capen (1998), Klammer et al. (2007), Köhrle et al. (1986), Morse et al. (1993), Renko et al. (2012) and Visser et al. (1979)
Induction of sulfation and glucuronidation enzyme activity
Increased biliary elimination of T3 and T4
Acetochlor, phenobarbital, 3-methylcolanthrene, PCBs, 1-methyl-3-propyl-imidazole-2-thione
Crofton (2008) and Vansell and Klaassen (2002)
Inhibition of sulfation and glucuronidation enzyme activity
Decreased biliary elimination of T3 and T4
Hydroxy PCBs and –PBDEs Hamers et al. (2008) and Schuur et al. (1999)
Hepatic Cellular Transporters
Altered biliary elimination of T3 and T4
TCPOBOP, pregnenolone-16- a-carbonitrile, TCDD, rifampicin, phenobarbital, oltipraz, di-n-butyl phthalate, n-butylbenzyl phthalate, dicofol
Guo et al. (2002), Jigorel et al. (2006), Petrick and Klaassen (2007), Shimada and Yamauchi (2004) and Staudinger et al. (2001)
TRbinding Altered activation of TH dependent gene transcription
T3-like hydroxy PCBs and -PBDEs, linuron, Tetrabromobisphenol A, arsenic
Davey et al. (2008), Freitas et al. (2011), Gauger et al. (2004), Hong et al. (2001), Kitamura et al. (2005b),Moriyama et al. (2002) and Schmutzler et al. (2007b)
AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1335
assessment. To minimize the risk of false negatives as well as of false positives, the in vitro assays should cover all relevant end- points of THD to distinguish between chemicals of less concern and chemicals that need further study. Prioritization of chemicals based on a well-chosen in vitro assay battery, possibly extended with in silico approaches, will be much more specific compared to in vivo tests, will improve the efficiency of testing methods by providing empirica l data on still untested chemicals and will in- crease the ability of risk managers to protect human health. Testing of chemical s for THD activity will likely need to be done in a tiered fashion, and in contrast to most proposed tiered approach es (Dixet al., 2007; Willett et al., 2011 ), we suggest that the first 2 tiers can be done fully in vitro. After every tier there is then an option not to proceed with further testing of a chemical.
4.1. Tier 1
A major goal of the pathway- based chemical hazard assessme nt is to be able to focus resource s, both laboratory and regulatory, on those chemical s of highest concern. Thus, the first function of acomprehens ive battery of in vitro tests is to prioritize the thou- sands of poorly characterized chemical s. Table 2 presents the range of endpoints covering the most important mechanism s of TH sys- tem regulation (Fig. 1) that currently are known to be sensitive for THDC and are now or in the near future, applicable to an in vitro HTS test battery. As current Tier 1 testing is aimed at pro- tection of humans from THDC, in vitro human cell-based assays are preferentiall y applied. Neverthel ess the high evolutionary preser- vation of THD targets clearly allows inclusion of other cell types, while on the other hand results obtained with human cells are usable for ecotoxicological purposes.
Importan tly, any in vitro approach should account for possible bio-activ ation of the test chemical to prevent false negatives, as several chemical s only become hormone disrupting after meta- bolic activation.
Once the test battery would be validated and accepted at OECD level, a Tier 1 in vitro battery screening outcome can be 1of 3 classifications. (1) No indication for THD effects even at high concentr ations. The chemical can then be classified as of less con- cern and does not need to be selected for further testing. (2)Strong indications for THD in the in vitro test battery. Further testing is needed at exposure concentrations that approach ex- pected in vivo concentrations (based on exposure assessment and toxicokineti c modellin g) unless hazard-based decisions are taken. (3) The responses in the Tier 1 in vitro test battery may not be conclusive or only weak and these chemicals should go to Tier 2 screening .
During optimizati on and in vivo validation of the in vitro testbattery, clearly defined triggers need to be developed, in order to be able to classify the chemicals as of less concern, high concern or inconclusive. While the classification ‘of less concern’ does not mean ‘‘no need for evaluation’’ category, the reality is that chemi- cals with low priority may never see testing because of limited resource s and test capacity. Therefore, it is important to minimize the number of false negatives, while not allowing the numbers of false positives to increase too much.
4.2. Tier 2
The purpose of this Tier 2 testing is to further identify and char- acterize the interactions of putative THDCs with the TH system. As an intermediate step between the in vitro Tier 1 assay battery and

Table 4In vitro bioassay battery recommended for Tier 1 screenin g of chemicals for THD. Summary of MTS and HTS assay usage and recomm endations for assay development by target. The last three assays are not specific for THD, but can indirectly induce a THD effect. Bioactivation of the test chemicals is needed to prevent false negative responses.
Target Relevance Status State of science References
TRH receptor signalling
Feedback control of TH synthesis
Technology available and research gap
1. Commercially available CHO-K1 cells with stable rhTRH1 (PerkinElmer). No literature reports on use
Boutin et al. (2012)
2. LTS assay using HEK-EM293 cells stably expressing mouse TRH-R1 or TRH-R2 that uses
TSH receptor signalling
Feedback control of TH synthesis
In use and research gap 1. HTS assay available using HEK293 cell line stably transfected with the TSHR coupled to acyclic nucleotide gated ion channel as abiosensor. Potential confounds by chemicals that interfere with cAMP pathways
Jomaa et al. (2013) and Titus et al. (2008)
2. MTS assay available for TSH mediated FRTL-5 cells rat thyroid cell proliferation
NIS-mediated iodide uptake
TH synthesis In use 1. HTS assays using HEK293 cells stably transfected with hNIS. Requires use of radiolabeled chemical
Cianchetta et al. (2010), Lecat-Guillet et al. (2007) and Waltz et al. (2010)
2. MTS assays available include FRTL-5 cells with yellow cerium indicator or with yellow fluorescent protein (YFP) variant YFP-H148Q/ I152L. Could be adapted for HTS
TPO inhibition TH synthesis Potential for adaptation for MTS or HTS, research gap: complex Fe-dependent mechanisms of action
1. LTS assays published that may be amendable to MTS or HTS. Both employ FTC-238 cells transfected with hrTPO
Schmutzler et al. (2007a,b),Song et al. (2011) and Verhaeghe et al. (2008)
2. HTS assay available that uses vanadium haloperoxidases. Will require comparison to vertebrate TPO
Binding to serum transport proteins TTR and TBG
TH availability in tissues, THDC transport, serum hormone levels
In use, needs optimization and validation
1. HTS assay available that uses Surface plasmon resonance based biosensor
Marchesini et al. (2008),Montano et al. (2012) and Yamauchi et al. (2003)
2. MTS available based on non-radioactive fluorescence displacement assay
Inhibition of deiodination enzyme activity
T3/T4 ratio Potential for adaptation for MTS or HTS and Research Gap
1. MTS assay available for D1 based on tissue or cell lysates, also applicable to intact cells
Kunisue et al. (2011), Piehl et al. (2008) and Renko et al. (2012)
Tissue hormone levels 2. Mass spectrometry of tissue thyroid hormone and metabolite profile
Inhibition of sulfation and glucuronidation enzyme activity
Hormone turnover Potential for adaptation for MTS or HTS
1. MTS assays currently being used. Very limited coverage of the biology and requires primary cells or enzyme preparations.
Hamers et al. (2008), Kavlock et al. (2012) and Paul et al. (2010)
2. Commercial assays from CellzDirect (UGT1A1,SULT2A1)
Tissue flux of THs via membrane transporters
Tissue hormone levels Research gap Low-throughput assays available that use radiolabeled transporter substrates
Freitas et al. (2011), Morimoto et al. (2008), van der Deure et al. (2010) and Visser et al. (2010)
TR binding and transcription
Receptor activation/ inhibition
In use 1. MTS assay for T3 induced proliferation of GH3 cells. Potential confounds by chemicals that interfere with non-T3 mediated proliferation pathways.
Freitas et al. (2011), Gutleb et al. (2005), Johnson et al. (2011), Rotroff et al. (2012)and Schriks et al. (2006)
2. HTS assays currently use a variety of assay technologies for a TR a and TR b as well as a TR b-SCR2 coactivator assay 3. HTS assay for TR activation (TR-GH3.Luc)
Activation of other nuclear receptors that heterodimerize with TRs
Influences TRE-mediated gene transcription
In use HTS assays currently use a variety of assay technologies for a number of nuclear receptors (e.g., CAR, PXR, PPAR) known to interfere with TR-activation
Huang et al. (2011), Knudsen et al. (2011) and Shah et al. (2011)
Upregulation of enzymes involved in TH conjugation or deiodination enzymes
Influences hormone turnover
In use HTS assays currently use a variety of assay technologies for a number of nuclear receptors (e.g., CAR, PXR, PPAR) known to be involved in the regulation of hepatic Phase 2 catabolic enzymes and Phase 3 cellular transporters
Huang et al. (2011), Knudsen et al. (2011) and Shah et al. (2011)
In vitro bioactivation of parent chemicals to test in the other in vitro bioassays
Bioactivation of THDC crucial in vivo but absent or not potent enough in in vitro assays
Potential for adaptation for HTS and research gap
In vitro metabolism followed by selective extraction of metabolites with limited co- extraction of disturbing matrix chemicals. This method needs further optimization and validation for HTS application.
Montano et al. (2012)
1336 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
in vivo bioassays, dedicated and modern tissue engineering tech- niques may be used to further select and refine the choice of chem- icals that need full Tier 3 testing in vivo . Tier 2 testing should allow
more focusing and targeting on more time-con suming but also more physiologica lly based assays e.g., with primary tissue culture, functiona l thyroid follicular cells different iated from mouse

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1337
embryonic stem-cell s (Antonica et al., 2012 ), embryonic zebrafisheleuthero-em bryos tissue to identify chemicals impairing thyroid gland morphogenes is (Raldua et al., 2008 ), or with fish- or amphib- ian embryos, targeting endpoints suggested by the results in Tier 1.
Once Tier 2 assays qualify the chemical as a potential THD haz- ard, evaluation could proceed to Tier 3 in vivo testing. However, be- fore this takes place, the toxicokinetics of the chemicals could be determined based on in vitro metabolism studies combined with in silico modellin g, as this has been shown to allow translation of in vitro results to in vivo effects (Punt et al., 2013 ). From the in vitro EC50 (which statistically is most robust) it is then possible to predict not only the in vivo EC50, but also the in vivo EC10 or BMD10 (benchmark dose at which a benchmark response equiva- lent to a 10% effect is induced) which is more relevant for the hu- man situation (Louisse et al., 2010 ).
Tier 3 in vivo experiments should preferably make use of assays that need to be performed anyhow for other ED endpoint s, and THD-relevan t endpoints indicated by the in vitro assays should be added to the standard in vivo tests.
With both Tier 1 and Tier 2 testing, the challenge is to have abattery of assays that comprehens ively evaluate the pathways re- lated to phenotype of interest together with a weight of evidence approach to ranking chemicals. The ToxCast program has devel- oped one such approach, termed the ‘‘ToxPi’’ that takes large amounts of information about chemical interactions with particu- lar biological pathways and displays them in a visual format for decision makers to use (Reif et al., 2010 ).
5. Advantages of in vitro testing
Although in vivo bioassays are generally perceived as being the most reliable tests for assessing the hazard of chemicals, including THDC, there also are several limitations for this approach. The functioning of the TH system depends on life-stage, species and health condition. Healthy animals may cope with certain exposures by compensati on via HPT feedback and effects may be unnoticed, while people with specific health conditions, such as hypo- or hyperthyr oidism or malnutrition, may be more vulnera- ble. Some animals may have targets for THD that are not relevant for humans and vice versa (e.g., Capen, 1997 ). For example TBG is an important human TH plasma transport protein while this is not the case for adult male rats. Life-stage specific differences may also result in under-estim ating hazards for humans. For example, most in vivo animal studies are conducted using adult euthyroid ani- mals, and the developing, or hypo- or hyperthyroid organisms may be more susceptible to THDC. A well-chosen in vitro test bat- tery should cover mechanism s and targets that are relevant for multiple life stages and conditions. This way a much broader range of chemical s can be assessed for potential perturbations of THD pathways of relevance for human biology in a shorter time, with virtually no animals and at lower costs. To be able to make the step from hazard to risk assessme nt based on in vitro test batteries more fundamenta l knowledge is needed to reveal the relationship between the extent and timing of exposure, with critical pathway perturbation s, physiolog ical effects, and disease outcomes. With insight in new mechanism s, in vitro test battery results may offer greater ability to predict which chemical s are more relevant for specifically sensitive groups or conditions. In vitro test results also offer more possibilities to predict THD effects for other species than humans, once more is known about possible species specificdifferences in cellular and molecula r targets, and physiologica l sys- tems. Comparing in vitro and in vivo responses will help to deter- mine specific mechanisms of THD chemicals and further develop better predictive research models for use in regulatory toxicolog y. To allow in vitro-in vivo comparis ons, it is very important to deter-
mine the blood plasma levels of the chemical s and/or its bioactive metaboli tes in the in vivo experiments .
6. Further outlook: using computationa l modelling to interpret in vitro assays for in vivo effects
Although it was not the ambition of this review to present an in vitro approach to be able to predict in vivo effects, this could at a later stage be further elaborated. Despite the many advantages of an in vitro testing battery, estimating in vivo toxicity from in vitro results can be a daunting task, as it is not possible to fully reproduce the in vivo situation in vitro . There are two major uncer- tainties in extrapolati on from in vitro to in vivo . One is the need to determine the ability of simple, in vitro cell culture or isolated pro- tein preparations to fully reflect the complex biology found in whole organisms. The second is the complexity in extrapolating in vitro concentr ations to in vivo pharmac okinetics (Blaauboe r, 2010). The absence of whole-body pharmacokine tic processes (absorption, distribution , metabolism and excretion) in the in vitro assay system is a challenge in implementi ng in vitro basedrisk assessments for chemicals. Integration of data on the toxic mode of action of a chemical with data on its pharmac okinetic behaviou r is essential for the interpretation of in vitro studies on the toxicity of a chemical in the whole animal. With physiologi- cally based biokinetic (PBBK) models, a better interpretation of in vivo relevance of in vitro results can be made by linking an in vitro concentratio n–response curve to an in vivo dose–response curve. These approaches are also known as IVIVE or reverse dosim- etry (Forsby and Blaauboe r, 2007; Judson et al., 2011; Louisse et al., 2010; Punt et al., 2013; Verwei et al., 2006; Wetmore et al., 2012 ).IVIVE is a process for estimating the environmental exposures to achemical that could produce tissue exposures at the site of toxicity in humans equivalent to those associate d with effects in an in vitro toxicity test (e.g., an EC 50, a Benchma rk concentratio n, etc.) (Louis-se et al., 2010 ). IVIVE can provide an estimate of the likelihood of harmful effects from expected environmental exposures to chemi- cals by integrati ng diverse information from targeted in vitro toxic-ity and kinetic assays using a computational modellin g approach .Pharmacoki netic modelling plays a pivotal role in this quantitative extrapolation, by incorporating the in vivo bioavailabili ty, distribu- tion, and clearance of the chemical into the process.
Computational systems biology pathway (CSBP) models are also a crucial component in the transition away from animal testing in toxicolog y (Krewski et al., 2009 ). CSBP models provide quantitative descriptions of molecular circuits responsible for the basal opera- tion of toxicity pathways (i.e., thyroid disruption) and characterize their alteration with chemical exposure (Zhang et al., 2010b ). Using quantitat ive descriptions of chemical interactio ns with specificreceptors or other molecular targets, these models describe the dose–response behaviou r observed in the various assays. These dose response models provide points of departure concentratio ns to serve as inputs for IVIV extrapolation and predictio n of human risk from environmental exposures . An especially attractive oppor- tunity exists in linking these cellular models across multiple cell types, representative of different tissues within the body (e.g., liver, hypothal amus, pituitary, gonads, etc.). CSBP models can also help explainin g non-monoto nic dose–response relationship s that are sometimes observed in in vivo experiments , as these might be the result of the combined result of multiple pathways with differ- ent dose–response relationships in different tissues and over time.
7. Recommendati ons for assay use and developmen t
The overall goal of this paper is to summarize the potential thy- roid targets of environmental chemical s and in vitro assays that

1338 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
cover the important mechanism s for TH disruption by environ- mental chemicals. From the above review it is clear that the state of the science for these assays ranges from those already being used in medium-through put screening (MTS) or HTS, to potential targets that lack cost efficient testing methods. To help focus research and developmen t efforts we provide a list of recommen- dations for assay use and developmen t (Table 4). These recommen- dations fall into three basic categories ; assays already in use for MTS and HTS that are assays not currently used but show promise for adaption to MTS or HTS, and endpoints for important mecha- nisms that either lack in vitro assays or available assays are not easily amenable to adaptation for MTS or HTS.
The first category contains assays that target known molecular events important for thyroid processes and are already employed in MTS or HTS of environmental chemicals. These include the Tox- Cast and Tox21 assays (Kavlock et al., 2012; Freitas et al., 2012a ).Some other assays are currently in use (Lecat-Guille t et al., 2008 )or could be used (Marchesini et al., 2008; Montano et al., 2012 ).Data from some of these assays has begun to be analysed (Johnsonet al., 2011; Rotroff et al., 2012 ) and will provide important infor- mation on assay performanc e and utility in screening environmen- tal chemicals for potential interactions with receptors and receptor signalling systems. Incorpora tion of existing HTS assays for NIS and TTR would expand coverage of screening systems to include important targets for THD chemicals. While there is little evidence that many xenobioti cs target TRH and TSH, there has also been very little testing. Therefore, existing HTS assays could be used to determine whether these are important targets for inclusion in HTS screening batteries.
The second group consists of assays that may be suitable for MTS or HTS. These assays require research to determine whether they could be adapted from current low-through put technology to allow for efficient testing of larger number of chemicals. For example, stable reporter cell lines with human recombinant TPO have been developed, but require adaptation testing. There are guidelines available that list requiremen ts for use in HTS screening programs (e.g., NCATS, 2012 ). Due to the large number and variety of chemicals known to inhibit TPO, a high priority should be given to developmen t of MTS or HTS TPO assays.
The third group includes endpoints for which no screening as- says currently exist that could be easily modified for MTS or HTS. These will require assay developmen t research to devise basic in vitro methods that could then be adapted to MTS or HTS. This in- cludes a number of the THDC targets reviewed above. For example, there are currently no MTS in vitro assays for cellular uptake and efflux of TH or TH metabolites . Current in vitro methods for detect- ing alteration s in TH secretion are very low throughput and involve the use of primary tissues from animals. The critical nature of deio- dinases in generating and degrading the active hormone T3, regu- lating intracellular TH homeostasis, and evidence that some chemicals inhibit activity, makes development of MTS and HTS as- says for interfere nce with normal deiodinase function a high priority.
In order to evaluate the predictabi lity and optimize the size of the in vitro test battery, we recommend that the proposed first tier assays be challenged with a well-chosen set of chemicals with known in vivo THD potential and preferably those with known mechanism of action. In Table 3 we present some examples of ac- tive chemicals that could be further investigated during the assay validation process.
8. Summarizing conclusi ons
This review presents a ‘best expert judgement’ for a set of in vitro testing assays that together will cover the most important
mechanis ms for TH disruption by chemicals. For these endpoints, currently available or desired in vitro assays are discussed that are or could be suitable for medium to high throughput testing. Bio-activ ation of the tested chemicals is a critical consideration in the in vitro testing, as several cases are known when testing of the parent chemical alone results in false negative responses. The applicati on of an in vitro test battery in a tiered testing approach for THDC is discussed as well as the outlook for combination with in silico alternativ es that eventually may further reduce the use of animals for testing chemical s for THD while enhancin g the knowl- edge of the mechanisms of action of the chemicals. Technical and in vivo validation of the in vitro test battery will be necessary to in- crease confidence for use of its results in regulatory decisions .
Conflict of interest statement
There are no conflicts of interest issues.
Disclaimer
The views expressed in this paper are those of the authors and do not necessarily reflect the views or policies of the U.S. Environ- mental Protection Agency. Mention of trade names or commercial products does not constitute endorsemen t or recommend ation for use.
Acknowled gements
This paper is based on the results of a workshop organized by the Institute for Risk Assessment Sciences, Utrecht University, for which financial support was kindly provided by ‘‘Proefdiervri j’’ (the Dutch Society for the Replacement of Animal Testing (dsRAT)www.pro efdiervrij.nl/engl ish ) and the ASAT Foundation (www.asat-initi ative.eu/ ). We want to thank Maurico Montaño for the nicer version he made of Fig. 1.
References
Aamir, I.S., Tauheed, S., Majid, F., Atif, A., 2010. Frequency of autoimmune thyroid disease in chronic urticaria. JCPSP – J. Coll. Phys. Surg. Pakistan 20, 158–161.
Alexander, W.D., Wolff, J., 1966. Thyroidal iodide transport. VIII. Relation between transport, goitrogenic and Antigoitrogenic properties of certain anions. Endocrinology 78, 581–590.
Andersson, M., Karumbunathan, V., Zimmermann, M.B., 2012. Global iodine status in 2011 and trends over the past decade. J. Nutr. 142, 744–750.
Antonica, F., Kasprzyk, D.F., Opitz, R., Iacovino, M., Liao, X.H., Dumitrescu, A.M., Refetoff, S., Peremans, K., Manto, M., Kyba, M., Costagliola, S., 2012. Generation of functional thyroid from embryonic stem cells. Nature.
Aqai, P., Fryganas, C., Mizuguchi, M., Haasnoot, W., Nielen, M.W., 2012. Triple bioaffinity mass spectrometry concept for thyroid transporter ligands. Anal. Chem. 84, 6488–6493.
Arturi, F., Presta, I., Scarpelli, D., Bidart, J.M., Schlumberger, M., Filetti, S., Russo, D., 2002. Stimulation of iodide uptake by human chorionic gonadotropin in FRTL-5 cells: effects on sodium/iodide symporter gene and protein expression. Eur. J. Endocrinol. 147, 655–661.
Arulmozhiraja, S., Shiraishi, F., Okumura, T., Iida, M., Takigami, H., Edmonds, J.S., Morita, M., 2005. Structural requirements for the interaction of 91 hydroxylated polychlorinated biphenyls with estrogen and thyroid hormone receptors. Toxicol. Sci. 84, 49–62.
Atterwill, C.K., Fowler, K.F., 1990. A comparison of cultured rat FRTL-5 and porcine thyroid cells for predicting the thyroid toxicity of xenobiotics. Toxicol. In Vitro 4, 369–374.
Attia, R.R., Connnaughton, S., Boone, L.R., Wang, F., Elam, M.B., Ness, G.C., Cook, G.A., Park, E.A., 2010. Regulation of pyruvate dehydrogenase kinase 4 (PDK4) by thyroid hormone: role of the peroxisome proliferator-activated receptor gamma coactivator (PGC-1 alpha). J. Biol. Chem. 285, 2375–2385.
Auf’mkolk, M., Amir, S.M., Kubota, K., Ingbar, S.H., 1985. The active principles of plant extracts with antithyrotropic activity: oxidation products of derivatives of 3,4-dihydroxycinnamic acid. Endocrinology 116, 1677–1686.
Axelrad, D.A., Baetcke, K., Dockins, C., Griffiths, C.W., Hill, R.N., Murphy, P.A., Owens, N., Simon, N.B., Teuschler, L.K., 2005. Risk assessment for benefits analysis: framework for analysis of a thyroid-disrupting chemical. J. Toxicol. Environ. Health A – Curr. Issues 68, 837–855.

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1339
Ball, S.G., Sokolov, J., Chin, W.W., 1997. 3,5-Diiodo- L-thyronine (T2) has selective thyromimetic effects in vivo and in vitro. J. Mol. Endocrinol. 19, 137–147.
Basaria, S., Cooper, D.S., 2005. Amiodarone and the thyroid. Am. J. Med. 118, 706–714.
Bassett, J.H., Boyde, A., Howell, P.G., Bassett, R.H., Galliford, T.M., Archanco, M., Evans, H., Lawson, M.A., Croucher, P., St Germain, D.L., Galton, V.A., Williams, G.R., 2010. Optimal bone strength and mineralization requires the type 2iodothyronine deiodinase in osteoblasts. Proc. Natl. Acad. Sci. USA 107, 7604–7609.
Bauer, M., Goetz, T., Glenn, T., Whybrow, P.C., 2008. The thyroid–brain interaction in thyroid disorders and mood disorders. J. Neuroendocrinol. 20, 1101–1114.
Baumann, C.T., Maruvada, P., Hager, G.L., Yen, P.M., 2001. Nuclear cytoplasmic shuttling by thyroid hormone receptors. Multiple protein interactions are required for nuclear retention. J. Biol. Chem. 276, 11237–11245.
Baxter, J.D., Webb, P., 2009. Thyroid hormone mimetics: potential applications in atherosclerosis, obesity and type 2 diabetes. Nat. Rev. Drug. Discov. 8, 308–320.
Benedetti, M.S., Whomsley, R., Baltes, E., Tonner, F., 2005. Alteration of thyroid hormone homeostasis by antiepileptic drugs in humans: involvement of glucuronosyltransferase induction. Eur. J. Clin. Pharmacol. 61, 863–872.
Berg, C., Gyllenhammar, I., Kvarnryd, M., 2009. Xenopus tropicalis as a test system for developmental and reproductive toxicity. J. Toxicol. Environ. Health A 72, 219–225.
Berg, V., Lyche, J.L., Gutleb, A.C., Lie, E., Skaare, J.U., Aleksandersen, M., Ropstad, E., 2010. Distribution of PCB 118 and PCB 153 and hydroxylated PCB metabolites (OH-CBs) in maternal, fetal and lamb tissues of sheep exposed during gestation and lactation. Chemosphere 80, 1144–1150.
Bergh, J.J., Lin, H.Y., Lansing, L., Mohamed, S.N., Davis, F.B., Mousa, S., Davis, P.J., 2005. Integrin alphaVbeta3 contains a cell surface receptor site for thyroid hormone that is linked to activation of mitogen-activated protein kinase and induction of angiogenesis. Endocrinology 146, 2864–2871.
Bergman, A., Klasson-Wehler, E., Kuroki, H., 1994. Selective retention of hydroxylated PCB metabolites in blood. Environ. Health Perspect. 102, 464–469.
Bernal, J., 2007. Thyroid hormone receptors in brain development and function. Nat. Clin. Pract. Endocrinol. Metab. 3, 249–259.
Bianco, A.C., 2011. Minireview: cracking the metabolic code for thyroid hormone signaling. Endocrinology 152, 3306–3311.
Biegel, L.B., Cook, J.C., O’Connor, J.C., Aschiero, M., Arduengo 3rd, A.J., Slone, T.W., 1995. Subchronic toxicity study in rats with 1-methyl-3-propylimidazole-2- thione (PTI): effects on the thyroid. Fundam. Appl. Toxicol. 27, 185–194.
Bizhanova, A., Kopp, P., 2009. Minireview: the sodium–iodide symporter NIS and pendrin in iodide homeostasis of the thyroid. Endocrinology 150, 1084–1090.
Blaauboer, B.J., 2010. Biokinetic modeling and in vitro-in vivo extrapolations. J. Toxicol. Environ. Health B Crit. Rev. 13, 242–252.
Blake, C.C., Geisow, M.J., Oatley, S.J., Rerat, B., Rerat, C., 1978. Structure of prealbumin: secondary, tertiary and quaternary interactions determined by Fourier refinement at 1.8 A. J. Mol. Biol. 121, 339–356.
Blanchet, E., Bertrand, C., Annicotte, J.S., Schlernitzauer, A., Pessemesse, L., Levin, J., Fouret, G., Feillet-Coudray, C., Bonafos, B., Fajas, L., Cabello, G., Wrutniak- Cabello, C., Casas, F., 2012. Mitochondrial T3 receptor p43 regulates insulin secretion and glucose homeostasis. FASEB J. 26, 40–50.
Blount, B.C., Pirkle, J.L., Osterloh, J.D., Valentin-Blasini, L., Caldwell, K.L., 2006. Urinary perchlorate and thyroid hormone levels in adolescent and adult men and women living in the United States. Environ. Health Perspect. 114, 1865–1871.
Boas, M., Feldt-Rasmussen, U., Skakkebaek, N.E., Main, K.M., 2006. Environmental chemicals and thyroid function. Eur. J. Endocrinol. 154, 599–611.
Boutin, A., Allen, M.D., Neumann, S., Gershengorn, M.C., 2012. Persistent signaling by thyrotropin-releasing hormone receptors correlates with G-protein and receptor levels. FASEB J. 26, 3473–3482.
Braun, D., Kim, T.D., le Coutre, P., Köhrle, J., Hershman, J.M., Schweizer, U., 2012. Tyrosine kinase inhibitors noncompetitively inhibit MCT8-mediated iodothyroninetransport. J. Clin. Endocrinol. Metab. 97, E100–E105.
Brent, G.A., 2012. Mechanisms of thyroid hormone action. J. Clin. Invest. 122, 3035–3043.
Brix, K., Linke, M., Tepel, C., Herzog, V., 2001. Cysteine proteinases mediate extracellular prohormone processing in the thyroid. Biol. Chem. 382, 717–725.
Brix, K., Führer, D., Biebermann, H., 2011. Molecules important for thyroid hormone synthesis and action – known facts and future perspectives. Thyroid Res. 4(Suppl. 1), S9.
Brouwer, A., van den Berg, K.J., 1986. Binding of a metabolite of 3,4,3 0 ,40-tetrachlorobiphenyl to transthyretin reduces serum vitamin A transport by inhibiting the formation of the protein complex carrying both retinol and thyroxin. Toxicol. Appl. Pharmacol. 85, 301–312.
Brouwer, A., Klasson-Wehler, E., Bokdam, M., Morse, D.C., Traag, W.A., 1990. Competitive inhibition of thyroxin binding to transthyretin by monohydroxy metabolites of 3,4,3 0 ,40-tetrachlorobiphenyl. Chemosphere 20, 1257.
Brouwer, A., Morse, D.C., Lans, M.C., Schuur, A.G., Murk, A.J., Klasson-Wehler, E., Bergman, A., Visser, T.J., 1998. Interactions of persistent environmental organohalogens with the thyroid hormone system: mechanisms and possible consequences for animal and human health. Toxicol. Ind. Health 14, 59–84.
Brucker-Davis, F., 1998. Effects of environmental synthetic chemicals on thyroid function. Thyroid 8, 827–856.
Butt, C.M., Wang, D., Stapleton, H.M., 2011. Halogenated phenolic contaminants inhibit the in vitro activity of the thyroid-regulating deiodinases in human liver. Toxicol. Sci. 124, 339–347.
Buxbaum, J.N., Reixach, N., 2009. Transthyretin: the servant of many masters. Cell. Mol. Life Sci. 66, 3095–3101.
Cao, J., Lin, Y., Guo, L.H., Zhang, A.Q., Wei, Y., Yang, Y., 2010. Structure-based investigation on the binding interaction of hydroxylated polybrominated diphenyl ethers with thyroxine transport proteins. Toxicology 277, 20–28.
Cao, J., Guo, L.H., Wan, B., Wei, Y., 2011. In vitro fluorescence displacement investigation of thyroxine transport disruption by bisphenol A. J. Environ. Sci. (China) 23, 315–321.
Capen, C.C., 1994. Mechanisms of chemical injury of thyroid gland. Prog. Clin. Biol. Res. 387, 173–191.
Capen, C.C., 1997. Mechanistic data and risk assessment of selected toxic end points of the thyroid gland. Toxicol. Pathol. 25, 39–48.
Capen, C.C., 1998. Correlation of mechanistic data and histopathology in the evaluation of selected toxic endpoints of the endocrine system. Toxicol. Lett. 102–103, 405–409.
Carta, M.G., Hardoy, M.C., Boi, M.F., Mariotti, S., Carpiniello, B., Usai, P., 2002. Association between panic disorder, major depressive disorder and celiac disease: a possible role of thyroid autoimmunity. J. Psychosom. Res. 53, 789–793.
Castillo, A.I., Sanchez-Martinez, R., Moreno, J.L., Martinez-Iglesias, O.A., Palacios, D., Aranda, A., 2004. A permissive retinoid X receptor/thyroid hormone receptor heterodimer allows stimulation of prolactin gene transcription by thyroid hormone and 9-cis-retinoic acid. Mol. Cell. Biol. 24, 502–513.
Cavalieri, R.R., Pitt-Rivers, R., 1981. The effects of drugs on the distribution and metabolism of thyroid hormones. Pharmacol. Rev. 33, 55–80.
Cavalieri, R.R., Simeoni, L.A., Park, S.W., Baxter, J.D., Scharschmidt, B.F., Ribeiro, R.C., Lomri, N., 1999. Thyroid hormone export in rat FRTL-5 thyroid cells and mouse NIH-3T3 cells is carrier-mediated, verapamil-sensitive, and stereospecific.Endocrinology 140, 4948–4954.
Chakrabarti, S., 2011. Thyroid functions and bipolar affective disorder. J. Thyroid Res. 2011, 306367.
Chalmers, D.K., Scholz, G.H., Topliss, D.J., Kolliniatis, E., Munro, S.L., Craik, D.J., Iskander, M.N., Stockigt, J.R., 1993. Thyroid hormone uptake by hepatocytes: structure–activity relationships of phenylanthranilic acids with inhibitory activity. J. Med. Chem. 36, 1272–1277.
Chang, C., 2012. Neonatal autoimmune diseases: a critical review. J. Autoimmun. 38, J223–J238.
Cheek, A.O., Kow, K., Chen, J., McLachlan, J.A., 1999. Potential mechanisms of thyroid disruption in humans: interaction of organochlorine compounds with thyroid receptor, transthyretin, and thyroid-binding globulin. Environ. Health Perspect. 107, 273–278.
Cheng, S.Y., Leonard, J.L., Davis, P.J., 2010. Molecular aspects of thyroid hormone actions. Endocr. Rev. 31, 139–170.
Chevrier, J., Harley, K.G., Bradman, A., Gharbi, M., Sjodin, A., Eskenazi, B., 2010. Polybrominated diphenyl ether (PBDE) flame retardants and thyroid hormone during pregnancy. Environ. Health Perspect. 118, 1444–1449.
Chiamolera, M.I., Wondisford, F.E., 2009. Minireview: thyrotropin-releasing hormone and the thyroid hormone feedback mechanism. Endocrinology 150, 1091–1096.
Chiamolera, M.I., Sidhaye, A.R., Matsumoto, S., He, Q., Hashimoto, K., Ortiga- Carvalho, T.M., Wondisford, F.E., 2012. Fundamentally distinct roles of thyroid hormone receptor isoforms in a thyrotroph cell line are due to differential DNA binding. Mol. Endocrinol. 26, 926–939.
Chiellini, G., Apriletti, J.W., Yoshihara, H.A., Baxter, J.D., Ribeiro, R.C., Scanlan, T.S., 1998. A high-affinity subtype-selective agonist ligand for the thyroid hormone receptor. Chem. Biol. 5, 299–306.
Chocron, E.S., Sayre, N.L., Holstein, D., Saelim, N., Ibdah, J.A., Dong, L.Q., Zhu, X., Cheng, S.Y., Lechleiter, J.D., 2012. The trifunctional protein mediates thyroid hormone receptor-dependent stimulation of mitochondria metabolism. Mol. Endocrinol. 26, 1117–1128.
Chr is tens on , W.R. , Beck er , B. D., Hoa ng , H. D., Wahle , B. S. , Moo re , K.D ., Dass , P. D., La ke ,S.G. , Stua rt , B. P. , Van Go ethe m, D.L ., Sang ha , G. K., et al. , 19 95. Ex tr athy ro id al ly media ted ch ang es in cir cul atin g th yr oi d hor mon e con cent ra ti ons in the mal e ra tfol lo win g adm in ist ra ti on of an exp eri ment al ox yac etamid e (FO E 5043 ). To xi co l.App l. Ph arm acol . 13 2, 253 –26 2.
Christenson, W.R., Becker, B.D., Wahle, B.S., Moore, K.D., Dass, P.D., Lake, S.G., Van Goethem, D.L., Stuart, B.P., Sangha, G.K., Thyssen, J.H., 1996. Evidence of chemical stimulation of hepatic metabolism by an experimental acetanilide (FOE 5043) indirectly mediating reductions in circulating thyroid hormone levels in the male rat. Fundam. Appl. Toxicol. 29, 251–259.
Cianchetta, S., di Bernardo, J., Romeo, G., Rhoden, K.J., 2010. Perchlorate transport and inhibition of the sodium iodide symporter measured with the yellow fluorescent protein variant YFP-H148Q/I152L. Toxicol. Appl. Pharmacol. 243, 372–380.
Ciarrocca, M., Tomei, F., Caciari, T., Cetica, C., Andrè, J.C., Fiaschetti, M., Schifano, M.P., Scala, B., Scimitto, L., Tomei, G., Sancini, A., 2012. Exposure to arsenic in urban and rural areas and effects on thyroid hormones. Inhal Toxicol. 24 (9),589–598.
Cioffi, F., Lanni, A., Goglia, F., 2010. Thyroid hormones, mitochondrial bioenergetics and lipid handling. Curr. Opin. Endocrinol. Diabetes Obes. 17, 402–407.
Clewell, R.A., Merrill, E.A., Narayanan, L., Gearhart, J.M., Robinson, P.J., 2004. Evidence for competitive inhibition of iodide uptake by perchlorate and translocation of perchlorate into the thyroid. Int. J. Toxicol. 23, 17–23.
Cooper-Kazaz, R., Lerer, B., 2008. Efficacy and safety of triiodothyronine supplementation in patients with major depressive disorder treated with

1340 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
specific serotonin reuptake inhibitors. Int. J. Neuropsychopharmacol. 11, 685–699.
Costa-e-Sousa, R.H., Hollenberg, A.N., 2012. Minireview: the neural regulation of the hypothalamic–pituitary–thyroid axis. Endocrinology.
Crofton, K.M., 2008. Thyroid disrupting chemicals: mechanisms and mixtures. Int. J. Androl. 31, 209–223.
Crofton, K.M., Zoeller, R.T., 2005. Mode of action: neurotoxicity induced by thyroid hormone disruption during development-hearing loss resulting from exposure to PHAHs. Crit. Rev. Toxicol. 35, 757–769.
Danzi, S., Klein, I., 2012. Thyroid hormone and the cardiovascular system. Med. Clin. North Am. 96, 257–268.
Davey, J.C., Nomikos, A.P., Wungjiranirun, M., Sherman, J.R., Ingram, L., Batki, C., Lariviere, J.P., Hamilton, J.W., 2008. Arsenic as an endocrine disruptor: arsenic disrupts retinoic acid receptor-and thyroid hormone receptor-mediated gene regulation and thyroid hormone-mediated amphibian tail metamorphosis. Environ. Health Perspect. 116, 165–172.
Davis, P.J., Davis, F.B., Lin, H.Y., Mousa, S.A., Zhou, M., Luidens, M.K., 2009. Translational implications of nongenomic actions of thyroid hormone initiated at its integrin receptor. Am. J. Physiol. Endocrinol. Metab. 297, E1238–E1246.
Davis, P.J., Davis, F.B., Mousa, S.A., Luidens, M.K., Lin, H.Y., 2011. Membrane receptor for thyroid hormone: physiologic and pharmacologic implications. Annu. Rev. Pharmacol. Toxicol. 51, 99–115.
de Benoist, B., McLean, E., Andersson, M., Rogers, L., 2008. Iodine deficiency in 2007: global progress since 2003. Food Nutr. Bull. 29, 195–202.
de Cock, M., Maas, Y.G., van de Bor, M., 2012. Does perinatal exposure to endocrine disruptors induce autism spectrum and attention deficit hyperactivity disorders? Review. Acta Paediatr. 101, 811–818.
De Groef, B., Decallonne, B.R., Van der Geyten, S., Darras, V.M., Bouillon, R., 2006. Perchlorate versus other environmental sodium/iodide symporter inhibitors: potential thyroid-related health effects. Eur. J. Endocrinol. 155, 17–25.
de Jong, F.J., Masaki, K., Chen, H., Remaley, A.T., Breteler, M.M., Petrovitch, H., White, L.R., Launer, L.J., 2009. Thyroid function, the risk of dementia and neuropathologic changes: the Honolulu-Asia aging study. Neurobiol. Aging 30, 600–606.
De La Vieja, A., Dohan, O., Levy, O., Carrasco, N., 2000. Molecular analysis of the sod iu m/i od id e sy mpor ter : impa ct on thy ro id and ex tr at hy ro id path op hys io log y. Ph ys io l. Re v. 80 , 1083–110 5.
Delange, F., 1994. The disorders induced by iodine deficiency. Thyroid 4, 107–128.Dentice, M., Salvatore, D., 2011. Deiodinases: the balance of thyroid hormone: local
impact of thyroid hormone inactivation. J. Endocrinol. 209, 273–282.DeVito, M.J., Ross, D.G., Dupuy Jr., A.E., Ferrario, J., McDaniel, D., Birnbaum, L.S.,
1998. Dose–response relationships for disposition and hepatic sequestration of polyhalogenated dibenzo-p-dioxins, dibenzofurans, and biphenyls following subchronic treatment in mice. Toxicol. Sci. 46, 223–234.
DeVito, M., Biegel, L., Brouwer, A., Brown, S., Brucker-Davis, F., Cheek, A.O., Christensen, R., Colborn, T., Cooke, P., Crissman, J., Crofton, K., Doerge, D., Gray, E., Hauser, P., Hurley, P., Kohn, M., Lazar, J., McMaster, S., McClain, M., McConnell, E., Meier, C., Miller, R., Tietge, J., Tyl, R., 1999. Screening methods for thyroid hormone disruptors. Environ. Health Perspect. 107, 407–415.
Di Bernardo, J., Iosco, C., Rhoden, K.J., 2011. Intracellular anion fluorescence assay for sodium/iodide symporter substrates. Anal. Biochem. 415, 32–38.
Di Cosmo, C., Liao, X.H., Dumitrescu, A.M., Philp, N.J., Weiss, R.E., Refetoff, S., 2010. Mice deficient in MCT8 reveal a mechanism regulating thyroid hormone secretion. J. Clin. Invest. 120, 3377–3388.
Divi, R.L., Doerge, D.R., 1994. Mechanism-based inactivation of lactoperoxidase and thyroid peroxidase by resorcinol derivatives. Biochemistry 33, 9668–9674.
Divi, R.L., Chang, H.C., Doerge, D.R., 1997. Anti-thyroid isoflavones from soybean: isolation, characterization, and mechanisms of action. Biochem. Pharmacol. 54, 1087–1096.
Dix, D.J., Houck, K.A., Martin, M.T., Richard, A.M., Setzer, R.W., Kavlock, R.J., 2007. The ToxCast program for prioritizing toxicity testing of environmental chemicals. Toxicol. Sci. 95, 5–12.
Doerge, D.R., Sheehan, D.M., 2002. Goitrogenic and estrogenic activity of soy isoflavones. Environ. Health Perspect. 110 (Suppl. 3), 349–353.
Dohan, O., De la Vieja, A., Paroder, V., Riedel, C., Artani, M., Reed, M., Ginter, C.S., Carrasco, N., 2003. The sodium/iodide symporter (NIS): characterization, regulation, and medical significance. Endocr. Rev. 24, 48–77.
Donangelo, I., Braunstein, G.D., 2011. Update on subclinical hyperthyroidism. Am. Fam. Phys. 83, 933–938.
D’Souza, P.C., Ally, N., Agarwal, A., Rani, P., Das, M., Sarma, D., 2008. Demonstration of LanthaScreen TR-FRET-based nuclear receptor coactivator recruitment assay using PHERAstar, a multi-detection HTS microplate reader. Indian J. Pharmacol. 40, 89–90.
Dunn, J.T., Dunn, A.D., 2001. Update on intrathyroidal iodine metabolism. Thyroid 11, 407–414.
Eisenbrand, G., Pool-Zobel, B., Baker, V., Balls, M., Blaauboer, B.J., Boobis, A., Carere, A., Kevekordes, S., Lhuguenot, J.C., Pieters, R., Kleiner, J., 2002. Methods of in vitro toxicology. Food Chem. Toxicol. 40, 193–236.
Ekuase, E.J., Liu, Y., Lehmler, H.J., Robertson, L.W., Duffel, M.W., 2011. Structure–activity relationships for hydroxylated polychlorinated biphenyls as inhibitors of the sulfation of dehydroepiandrosterone catalyzed by human hydroxysteroid sulfotransferase SULT2A1. Chem. Res. Toxicol. 24, 1720–1728.
Elisei, R., Vivaldi, A., Ciampi, R., Faviana, P., Basolo, F., Santini, F., Traino, C., Pacini, F., Pinchera, A., 2006. Treatment with drugs able to reduce iodine efflux
significantly increases the intracellular retention time in thyroid cancer cells stably transfected with sodium iodide symporter complementary deoxyribonucleic acid. J. Clin. Endocrinol. Metab. 91, 2389–2395.
EPA, U.S., 2012. Endocrine Disruptor Screening Program Comprehensive Management Plan, June 2012. <http://www.epa.gov/endo/pubs/EDSP-comprehensive-management-plan.pdf>.
EU, 2007. Regulation (EC) No. 1907/2006 of the European Parliament and of the Council on the Registration, Evaluation, Authorisation and Restriction of Chemicals (REAHC), corrigendum. Official Journal of the European Union L136/3.
EU, 2009a. Regulation (EC) No. 1107/2009 of the European Parliament and of the Council of 21 October 2009 concerning the placing of plant protection products on the market. Official Journal of the European Union L309/1.
EU, 2009b. Regulation (EC) No 1223/2009 of the European Parliament and of the Council of 30 November 2009 on cosmetic products. Official Journal of the European Union L675/59.
EU, 2012. Regulation (EU) No 528/2012 of the European Parliament and of the Council of 22 May 2012 concerning the making available on the market and use of biocidal products. Official Journal of the European Union L167/1.
Fekete, C., Lechan, R.M., 2007. Negative feedback regulation of hypophysiotropic thyrotropin-releasing hormone (TRH) synthesizing neurons: role of neuronal afferents and type 2 deiodinase. Front. Neuroendocrinol. 28, 97–114.
Fentem, J., Chamberlain, M., Sangster, B., 2004. The feasibility of replacing animal testing for assessing consumer safety: a suggested future direction. Altern. Lab. Anim. 32, 617–623.
Figueira, A.C., Saidemberg, D.M., Souza, P.C., Martinez, L., Scanlan, T.S., Baxter, J.D., Skaf, M.S., Palma, M.S., Webb, P., Polikarpov, I., 2011. Analysis of agonist and antagonist effects on thyroid hormone receptor conformation by hydrogen/ deuterium exchange. Mol. Endocrinol. 25, 15–31.
Flamant, F., Gauthier, K., 2012. Thyroid hormone receptors: the challenge of elucidating isotype-specific functions and cell-specific response. Biochim. Biophys. Acta.
Fliers, E., Unmehopa, U.A., Alkemade, A., 2006. Functional neuroanatomy of thyroid hormone feedback in the human hypothalamus and pituitary gland. Mol. Cell. Endocrinol. 251, 1–8.
Forrest, D., Swaroop, A., 2012. Minireview: the role of nuclear receptors in photoreceptor differentiation and disease. Mol. Endocrinol. 26, 905–915.
Forsby, A., Blaauboer, B., 2007. Integration of in vitro neurotoxicity data with biokinetic modelling for the estimation of in vivo neurotoxicity. Hum. Exp. Toxicol. 26, 333–338.
Freindorf, M., Furlani, T.R., Kong, J., Cody, V., Davis, F.B., Davis, P.J., 2012. Combined QM/MM study of thyroid and steroid hormone analogue interactions with alphavbeta3 integrin. J. Biomed. Biotechnol. 2012, 959057.
Freitas, J., Cano, P., Craig-Veit, C., Goodson, M.L., Furlow, J.D., Murk, A.J., 2011. Detection of thyroid hormone receptor disruptors by a novel stable in vitro reporter gene assay. Toxicol. In Vitro 25, 257–266.
Freyberger, A., Ahr, H.J., 2006. Studies on the goitrogenic mechanism of action of N,N,N0 , N0-tetramethylthiourea. Toxicology 217, 169–175.
Friedrichs, B., Tepel, C., Reinheckel, T., Deussing, J., von Figura, K., Herzog, V., Peters, C., Saftig, P., Brix, K., 2003. Thyroid functions of mouse cathepsins B, K, and L. J. Clin. Invest. 111, 1733–1745.
Friesema, E.C., Jansen, J., Milici, C., Visser, T.J., 2005. Thyroid hormone transporters. Vitam. Horm. 70, 137–167.
Friesema, E.C., Jansen, J., Heuer, H., Trajkovic, M., Bauer, K., Visser, T.J., 2006a. Mechanisms of disease: psychomotor retardation and high T3 levels caused by mutations in monocarboxylate transporter 8. Nat. Clin. Pract. Endocrinol. Metab. 2, 512–523.
Friesema, E.C., Kuiper, G.G., Jansen, J., Visser, T.J., Kester, M.H., 2006b. Thyroid hormone transport by the human monocarboxylate transporter 8 and its rate- limiting role in intracellular metabolism. Mol. Endocrinol. 20, 2761–2772.
Friesema, E.C., Jansen, J., Jachtenberg, J.W., Visser, W.E., Kester, M.H., Visser, T.J., 2008. Effective cellular uptake and efflux of thyroid hormone by human monocarboxylate transporter 10. Mol. Endocrinol. 22, 1357–1369.
Fr eit as, J., Mil le r, N. , Xia , M., Hua ng , R. , Hou ck, K., IMC M Rie tj ens, JD Fu rl ow , AJ Mur k.20 12a . Hig h-t hro ug hp ut id ent ifica tio n of thy ro id ho rm on e re cept or ago ni sts andan ta gon is ts . J. of Bi om ol ecul ar Scr ee nin g, su bmit te d for pu bli caton .
Freitas J., Hernandez-Moreno, D., Schophuizen, C., Spenkelink, A., van den Berg, J.H.J., Furlow, D ., Murk, A.J., 2012b. Human thyroid hormone receptor alpha reporter gene cell line for characterization of endocrine disrupting compounds. Toxicology in vitro, submitted for publication.
Furlow, J.D., Yang, H.Y., Hsu, M., Lim, W., Ermio, D.J., Chiellini, G., Scanlan, T.S., 2004. Induction of larval tissue resorption in Xenopus laevis tadpoles by the thyroid hormone receptor agonist GC-1. J. Biol. Chem. 279, 26555–26562.
Garcia, S.I., Porto, P.I., Martinez, V.N., Alvarez, A.L., Finkielman, S., Pirola, C.J., 2000. Expression of TRH and TRH-like peptides in a human glioblastoma–astrocytomacell line (U-373-MG). J. Endocrinol. 166, 697–703.
Gauger, K.J., Kato, Y., Haraguchi, K., Lehmler, H.J., Robertson, L.W., Bansal, R., Zoeller, R.T., 2004. Polychlorinated biphenyls (PCBs) exert thyroid hormone-like effects in the fetal rat brain but do not bind to thyroid hormone receptors. Environ. Health Perspect. 112, 516–523.
Gauger, K.J., Giera, S., Sharlin, D.S., Bansal, R., Iannacone, E., Zoeller, R.T., 2007. Polychlorinated biphenyls 105 and 118 form thyroid hormone receptor agonists after cytochrome P4501A1 activation in rat pituitary GH3 cells. Environ. Health Perspect. 115, 1623–1630.
Gebbink, W.A., Sonne, C., Dietz, R., Kirkegaard, M., Born, E.W., Muir, D.C.G., Letcher, R.J., 2008a. Target tissue selectivity and burdens of diverse classes of

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1341
brominated and chlorinated contaminants in polar bears (Ursus maritimus ) from East Greenland. Environ. Sci. Technol. 42, 752–759.
Gebbink, W.A., Sonne, C., Dietz, R., Kirkegaard, M., Riget, F.F., Born, E.W., Muir, D.C., Letcher, R.J., 2008b. Tissue-specific congener composition of organohalogen and metabolite contaminants in East Greenland polar bears (Ursus maritimus ).Environ. Pollut. 152, 621–629.
Gerard, A.C., Poncin, S., Audinot, J.N., Denef, J.F., Colin, I.M., 2009. Iodide deficiency-induced angiogenic stimulus in the thyroid occurs via HIF- and ROS-dependent VEGF-A secretion from thyrocytes. Am. J. Physiol. Endocrinol. Metab. 296, E1414–E1422.
Gereben, B., Zavacki, A.M., Ribich, S., Kim, B.W., Huang, S.A., Simonides, W.S., Zeold, A., Bianco, A.C., 2008. Cellular and molecular basis of deiodinase-regulated thyroid hormone signaling. Endocr. Rev. 29, 898–938.
Gershengorn, M.C., 1993. Thyrotropin-releasing hormone receptor: cloning and regulation of its expression. Recent Prog. Horm. Res. 48, 341–363.
Ghisari, M., Bonefeld-Jorgensen, E.C., 2009. Effects of plasticizers and their mixtures on estrogen receptor and thyroid hormone functions. Toxicol. Lett. 189, 67–77.
Ghosh, M., Meerts, I.A., Cook, A., Bergman, A., Brouwer, A., Johnson, L.N., 2000. Structure of human transthyretin complexed with bromophenols: a new mode of binding. Acta Crystallogr. D Biol. Crystallogr. 56, 1085–1095.
Greenspan, S.L., Greenspan, F.S., 1999. The effect of thyroid hormone on skeletal integrity. Ann. Intern. Med. 130, 750–758.
Grimaldi, A., Buisine, N., Miller, T., Shi, Y.B., Sachs, L.M., 2012. Mechanisms of thyroid hormone receptor action during development: lessons from amphibian studies. Biochim. Biophys. Acta.
Grover, G.J., Dunn, C., Nguyen, N.H., Boulet, J., Dong, G., Domogauer, J., Barbounis, P., Scanlan, T.S., 2007. Pharmacological profile of the thyroid hormone receptor antagonist NH3 in rats. J. Pharmacol. Exp. Ther. 322, 385–390.
Grun, F., Blumberg, B., 2006. Environmental obesogens: organotins and endocrine disruption via nuclear receptor signaling. Endocrinology 147, S50–S55.
Guo, G.L., Choudhuri, S., Klaassen, C.D., 2002. Induction profile of rat organic anion transporting polypeptide 2 (oatp2) by prototypical drug-metabolizing enzyme inducers that activate gene expression through ligand-activated transcription factor pathways. J. Pharmacol. Exp. Ther. 300, 206–212.
Gutleb, A.C., Meerts, I.A., Bergsma, J.H., Schriks, M., Murk, A.J., 2005. T-Screen as atool to identify thyroid hormone receptor active compounds. Environ. Toxicol. Pharmacol. 19, 231–238.
Gutleb, A.C., Cenijn, P., Velzen, M., Lie, E., Ropstad, E., Skaare, J.U., Malmberg, T., Bergman, A., Gabrielsen, G.W., Legler, J., 2010. In vitro assay shows that PCB metabolites completely saturate thyroid hormone transport capacity in blood of wild polar bears (Ursus maritimus ). Environ. Sci. Technol. 44, 3149–3154.
Haddow, J.E., Palomaki, G.E., Allan, W.C., Williams, J.R., Knight, G.J., Gagnon, J., O’Heir, C.E., Mitchell, M.L., Hermos, R.J., Waisbren, S.E., Faix, J.D., Klein, R.Z., 1999. Maternal thyroid deficiency during pregnancy and subsequent neuropsychological development of the child. N. Engl. J. Med. 341, 549–555.
Hagenbuch, B., 2007. Cellular entry of thyroid hormones by organic anion transporting polypeptides. Best Pract. Res. Clin. Endocrinol. Metab. 21, 209–221.
Halestrap, A.P., 2012. The monocarboxylate transporter family – structure and functional characterization. IUBMB Life 64, 1–9.
Hallgren, S., Darnerud, P.O., 2002. Polybrominated diphenyl ethers (PBDEs),polychlorinated biphenyls (PCBs) and chlorinated paraffins (CPs) in rats- testing interactions and mechanisms for thyroid hormone effects. Toxicology 177, 227–243.
Hallgren, S., Sinjari, T., Hakansson, H., Darnerud, P.O., 2001. Effects of polybrominated diphenyl ethers (PBDEs) and polychlorinated biphenyls (PCBs) on thyroid hormone and vitamin A levels in rats and mice. Arch. Toxicol. 75, 200–208.
Hamers, T., Kamstra, J.H., Sonneveld, E., Murk, A.J., Kester, M.H., Andersson, P.L., Legler, J., Brouwer, A., 2006. In vitro profiling of the endocrine-disrupting potency of brominated flame retardants. Toxicol. Sci. 92, 157–173.
Hamers, T., Kamstra, J.H., Sonneveld, E., Murk, A.J., Visser, T.J., Van Velzen, M.J., Brouwer, A., Bergman, A., 2008. Biotransformation of brominated flameretardants into potentially endocrine-disrupting metabolites, with special attention to 2,2 0 ,4,40-tetrabromodiphenyl ether (BDE-47). Mol. Nutr. Food Res. 52, 284–298.
Harley, K.G., Marks, A.R., Chevrier, J., Bradman, A., Sjodin, A., Eskenazi, B., 2010. PBDE concentrations in women’s serum and fecundability. Environ. Health Perspect. 118, 699–704.
Haugen, B.R., 2009. Drugs that suppress TSH or cause central hypothyroidism. Best Pract. Res. Clin. Endocrinol. Metab. 23, 793–800.
Hennemann, G., Docter, R., Friesema, E.C., de Jong, M., Krenning, E.P., Visser, T.J., 2001. Plasma membrane transport of thyroid hormones and its role in thyroid hormone metabolism and bioavailability. Endocr. Rev. 22, 451–476.
Heuer, H., Mason, C.A., 2003. Thyroid hormone induces cerebellar Purkinje cell dendritic development via the thyroid hormone receptor alpha1. J. Neurosci. 23, 10604–10612.
Higa, Y., Ohkubo, A., Kitajima, S., Moriyasu, M., Kariya, K., 2002. Effects of kojic acid on thyroidal functions in rats by single-dose administration and in cultured rat thyroid cells (FRTL-5 cells). J. Toxicol. Sci. 27, 423–431.
Himms-Hagen, J., 1989. Brown adipose tissue thermogenesis and obesity. Prog. Lipid Res. 28, 67–115.
Hodkinson, C.F., Simpson, E.E., Beattie, J.H., O’Connor, J.M., Campbell, D.J., Strain, J.J., Wallace, J.M., 2009. Preliminary evidence of immune function modulation by thyroid hormones in healthy men and women aged 55–70 years. J. Endocrinol. 202, 55–63.
Hofmann, P.J., Schomburg, L., Köhrle, J., 2009. Interference of endocrine disrupters with thyroid hormone receptor-dependent transactivation. Toxicol. Sci. 110, 125–137.
Hogervorst, E., Huppert, F., Matthews, F.E., Brayne, C., 2008. Thyroid function and cognitive decline in the MRC cognitive function and ageing study. Psychoneuroendocrinology 33, 1013–1022.
Hong, S.H., Yang, Z., Privalsky, M.L., 2001. Arsenic trioxide is a potent inhibitor of the interaction of SMRT corepressor with Its transcription factor partners, including the PML-retinoic acid receptor alpha oncoprotein found in human acute promyelocytic leukemia. Mol. Cell. Biol. 21, 7172–7182.
Hood, A., Klaassen, C.D., 2000a. Differential effects of microsomal enzyme inducers on in vitro thyroxine (T(4)) and triiodothyronine (T(3)) glucuronidation. Toxicol. Sci. 55, 78–84.
Hood, A., Klaassen, C.D., 2000b. Effects of microsomal enzyme inducers on outer- ring deiodinase activity toward thyroid hormones in various rat tissues. Toxicol. Appl. Pharmacol. 163, 240–248.
Hood, A., Allen, M.L., Liu, Y., Liu, J., Klaassen, C.D., 2003. Induction of T(4) UDP-GT activity, serum thyroid stimulating hormone, and thyroid follicular cell proliferation in mice treated with microsomal enzyme inducers. Toxicol. Appl. Pharmacol. 188, 6–13.
Horn, S., Heuer, H., 2010. Thyroid hormone action during brain development: more questions than answers. Mol. Cell. Endocrinol. 315, 19–26.
Hornung, M.W., Degitz, S.J., Korte, L.M., Olson, J.M., Kosian, P.A., Linnum, A.L., Tietge, J.E., 2010. Inhibition of thyroid hormone release from cultured amphibian thyroid glands by methimazole, 6-propylthiouracil, and perchlorate. Toxicol. Sci. 118, 42–51.
Hotz, C.S., Belonje, B., Fitzpatrick, D.W., L’Abbe, M.R., 1996. A method for the determination of type I iodothyronine deiodinase activity in liver and kidney using 125I-labelled reverse triiodothyronine as a substrate. Clin. Biochem. 29, 451–456.
Houck, K.A., Kavlock, R.J., 2008. Understanding mechanisms of toxicity: insights from drug discovery research. Toxicol. Appl. Pharmacol. 227, 163–178.
Howdeshell, K.L., 2002. A model of the development of the brain as a construct of the thyroid system. Environ. Health Perspect. 110 (Suppl. 3), 337–348.
Huang, R., Xia, M., Cho, M.H., Sakamuru, S., Shinn, P., Houck, K.A., Dix, D.J., Judson, R.S., Witt, K.L., Kavlock, R.J., Tice, R.R., Austin, C.P., 2011. Chemical genomics profiling of environmental chemical modulation of human nuclear receptors. Environ. Health Perspect. 119, 1142–1148.
Hurley, P.M., 1998. Mode of carcinogenic action of pesticides inducing thyroid follicular cell tumors in rodents. Environ. Health Perspect. 106, 437–445.
Hwang, J.Y., Huang, W., Arnold, L.A., Huang, R., Attia, R.R., Connelly, M., Wichterman, J., Zhu, F., Augustinaite, I., Austin, C.P., Inglese, J., Johnson, R.L., Guy, R.K., 2011. Methylsulfonylnitrobenzoates, a new class of irreversible inhibitors of the interaction of the thyroid hormone receptor and its obligate coactivators that functionally antagonizes thyroid hormone. J. Biol. Chem. 286, 11895–11908.
Ibha ze hie bo , K. , Iwasa ki , T. , Kim ur a-K ur od a, J., Miy aza ki, W., Shim ok awa , N., Ko ib uc hi ,N., 2011. Dis ru pt io n of thy roi d ho rm on e re cep tor -m edi ated tra ns cri pti on and thy ro id hor mon e-i ndu ced Pur ki nje ce ll dend ri te arb or iz ati on by po ly br omin ated dip he ny l ethe rs . En vir on . Healt h Pe rs pe ct. 119, 168–175.
Invitrogen, 2010. GeneBLAzer � TR Alpha HEK 293T DA and TR Alpha-UAS-bla HEK 293T Cell-based Assay. <http://tools.invitrogen.com/content/sfs/manuals/geneblazer_TRalpha_UASblaHEK293T_man. pdf >.
Invitrogen, 2012. GeneBLAzer� TR Alpha DA Assay Kit. <http://products.invitrogen.com/ivgn/product/K1385>.
Iwasaki, T., Miyazaki, W., Takeshita, A., Kuroda, Y., Koibuchi, N., 2002. Polychlorinated biphenyls suppress thyroid hormone-induced transactivation. Biochem. Biophys. Res. Commun. 299, 384–388.
Janssen, J.S., Sharma, V., Pugazhenthi, U., Sladek, C., Wood, W.M., Haugen, B.R., 2011. A rexinoid antagonist increases the hypothalamic–pituitary–thyroid set point in mice and thyrotrope cells. Mol. Cell. Endocrinol. 339, 1–6.
Jarry, H., Christoffel, J., Rimoldi, G., Koch, L., Wuttke, W., 2004. Multi-organic endocrine disrupting activity of the UV screen benzophenone 2 (BP2) in ovariectomized adult rats after 5 days treatment. Toxicology 205, 87–93.
Jeong, S.H., Kim, B.Y., Kang, H.G., Ku, H.O., Cho, J.H., 2005. Effects of butylated hydroxyanisole on the development and functions of reproductive system in rats. Toxicology 208, 49–62.
Jigorel, E., Le Vee, M., Boursier-Neyret, C., Parmentier, Y., Fardel, O., 2006. Differential regulation of sinusoidal and canalicular hepatic drug transporter expression by xenobiotics activating drug-sensing receptors in primary human hepatocytes. Drug Metab. Dispos. 34, 1756–1763.
Joh ns on , R. L. , Hw ang , J.Y ., Ar no ld , L. A. , Hu ang , R. , Wic ht erm an, J., Aug us tin ait e, I., Aus tin , C. P., Ingl ese , J., Guy , R. K., Hua ng , W., 201 1. A qua nt it ati ve hi gh- thr ou gh put scr ee n id ent ifies nov el in hi bit or s of th e in tera ct io n of thy roi d re cept or beta wit h apept id e of st ero id rec epto r coa ct iv ato r 2. J. Bio mol . Scre en. 16 , 618–627 .
Jomaa, B., Aarts, J.M.M.J.G., de Haan, L.H.J., Peijnenburg, A.A.C.M., Bovee, T.F.H., Murk, A.J., Rietjens, I.M.C.M., 2013. In vitro pituitary and thyroid cell proliferation assays and their relevance as alternatives to animal testing. ALTEX.
Judson, R., Richard, A., Dix, D.J., Houck, K., Martin, M., Kavlock, R., Dellarco, V., Henry, T., Holderman, T., Sayre, P., Tan, S., Carpenter, T., Smith, E., 2009. The toxicity data landscape for environmental chemicals. Environ. Health Perspect. 117, 685–695.
Judson, R.S., Houck, K.A., Kavlock, R.J., Knudsen, T.B., Martin, M.T., Mortensen, H.M., Reif, D.M., Rotroff, D.M., Shah, I., Richard, A.M., Dix, D.J., 2010. In vitro screening of environmental chemicals for targeted testing prioritization: the ToxCast project. Environ. Health Perspect. 118, 485–492.

1342 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
Judson, R.S., Kavlock, R.J., Setzer, R.W., Hubal, E.A., Martin, M.T., Knudsen, T.B., Houck, K.A., Thomas, R.S., Wetmore, B.A., Dix, D.J., 2011. Estimating toxicity- related biological pathway altering doses for high-throughput chemical risk assessment. Chem. Res. Toxicol. 24, 451–462.
Jugan, M.L., Levy-Bimbot, M., Pomerance, M., Tamisier-Karolak, S., Blondeau, J.P., Levi, Y., 2007. A new bioluminescent cellular assay to measure the transcriptional effects of chemicals that modulate the alpha-1 thyroid hormone receptor. Toxicol. In Vitro 21, 1197–1205.
Jugan, M.L., Oziol, L., Bimbot, M., Huteau, V., Tamisier-Karolak, S., Blondeau, J.P., Levi, Y., 2009. In vitro assessment of thyroid and estrogenic endocrine disruptors in wastewater treatment plants, rivers and drinking water supplies in the greater Paris area (France). Sci. Total Environ. 407, 3579–3587.
Jugan, M.L., Levi, Y., Blondeau, J.P., 2010. Endocrine disruptors and thyroid hormone physiology. Biochem. Pharmacol. 79, 939–947.
Kanayama, T., Mamiya, S., Nishihara, T., Nishikawa, J., 2003. Basis of a high- throughput method for nuclear receptor ligands. J. Biochem. 133, 791–797.
Kato, Y., Ikushiro, S., Emi, Y., Tamaki, S., Suzuki, H., Sakaki, T., Yamada, S., Degawa, M., 2008. He pa tic UDP- gl uc ur on os yl tra ns fera se s re sp on sib le for gl ucu ron id at io nof thy ro xi ne in hu mans. Dru g Metab. Dis po s. 36, 51–55.
Kavlock, R., Chandler, K., Houck, K., Hunter, S., Judson, R., Kleinstreuer, N., Knudsen, T., Martin, M., Padilla, S., Reif, D., Richard, A., Rotroff, D., Sipes, N., Dix, D., 2012. Update on EPA’s ToxCast program: providing high throughput decision support tools for chemical risk management. Chem. Res. Toxicol. 25, 1287–1302.
Kester, M.H.A., Visser, T.J., 2005. Sulfation of thyroid hormones. In: Pacifici, G.M., Coughtrie, M.W.H. (Eds.), Human Cytosolic Sulfotransferases. CRC Press, Boca Raton, pp. 121–134.
Kester, M.H., van Dijk, C.H., Tibboel, D., Hood, A.M., Rose, N.J., Meinl, W., Pabel, U., Glatt, H., Falany, C.N., Coughtrie, M.W., Visser, T.J., 1999. Sulfation of thyroid hormone by estrogen sulfotransferase. J. Clin. Endocrinol. Metab. 84, 2577–2580.
Kester, M.H., Kaptein, E., Van Dijk, C.H., Roest, T.J., Tibboel, D., Coughtrie, M.W., Visser, T.J., 2002. Characterization of iodothyronine sulfatase activities in human and rat liver and placenta. Endocrinology 143, 814–819.
Keune, H., Gutleb, A.C., Zimmer, K.E., Ravnum, S., Yang, A., Bartonova, A., Krayer von Krauss, M., Ropstad, E., Eriksen, G.S., Saunders, M., Magnanti, B., Forsberg, B., 2012. We’re only in it for the knowledge? A problem solving turn in environment and health expert elicitation. Environ. Health 11 (Suppl. 1), S3.
Khait, L., Birla, R.K., 2008. Effect of thyroid hormone on the contractility of self- organized heart muscle. In Vitro Cell. Dev. Biol. Anim. 44, 204–213.
Kim, W.G., Cheng, S.Y., 2012. Thyroid hormone receptors and cancer. Biochim. Biophys. Acta.
Kinne, A., Roth, S., Biebermann, H., Köhrle, J., Grüters, A., Schweizer, U., 2009. Surface translocation and tri-iodothyronine uptake of mutant MCT8 proteins are cell type-dependent. J. Mol. Endocrinol. 43, 263–271.
Kinne, A., Kleinau, G., Hoefig, C.S., Grüters, A., Köhrle, J., Krause, G., Schweizer, U., 2010. Essential molecular determinants for thyroid hormone transport and firststructural implications for monocarboxylate transporter 8. J. Biol. Chem. 285, 28054–28063.
Kitamura, S., Jinno, N., Ohta, S., Kuroki, H., Fujimoto, N., 2002. Thyroid hormonal activ it y of the flame re ta rd ant s tetra br om obi sph enol A and te tra chl or ob is ph enol A. Bio che m. Bi op hy s. Re s. Com mun . 293, 554–559 .
Kitamura, S., Kato, T., Iida, M., Jinno, N., Suzuki, T., Ohta, S., Fujimoto, N., Hanada, H., Kashiwagi, K., Kashiwagi, A., 2005a. Anti-thyroid hormonal activity of tetrabromobisphenol A, a flame retardant, and related compounds: affinity to the mammal ia n th yr oi d ho rm on e re ce pto r, and eff ect on ta dpo le metamor ph osi s. Lif e Sci . 76, 15 89–160 1.
Kitamura, S., Suzuki, T., Sanoh, S., Kohta, R., Jinno, N., Sugihara, K., Yoshihara, S., Fu ji mot o, N. , Wat ana be, H. , Ohta , S., 2005b. Co mpara ti ve stu dy of th e end ocr in e-dis ru pt in g act iv it y of bis ph eno l A and 19 re la ted com pou nd s. To xi col . Sci . 84, 249 –259.
Klammer, H., Schlecht, C., Wuttke, W., Schmutzler, C., Gotthardt, I., Köhrle, J., Jarry, H., 2007. Effects of a 5-day treatment with the UV-filter octyl- methoxycinnamate (OMC) on the function of the hypothalamo–pituitary–thyroid function in rats. Toxicology 238, 192–199.
Klein, I., Danzi, S., 2007. Thyroid disease and the heart. Circulation 116, 1725–1735.Knudsen, T.B., Houck, K.A., Sipes, N.S., Singh, A.V., Judson, R.S., Martin, M.T.,
Weissman, A., Kleinstreuer, N.C., Mortensen, H.M., Reif, D.M., Rabinowitz, J.R., Setzer, R.W., Richard, A.M., Dix, D.J., Kavlock, R.J., 2011. Activity profiles of 309 ToxCast chemicals evaluated across 292 biochemical targets. Toxicology 282, 1–15.
Köhrle, J., 2002. Iodothyronine deiodinases. Methods Enzymol. 347, 125–167.Köhrle, J., 2007. Thyroid hormone transporters in health and disease: advances in
thyroid hormone deiodination. Best Pract. Res. Clin. Endocrinol. Metab. 21, 173–191.
Köhrle, J., 2008. Environment and endocrinology: the case of thyroidology. Ann. Endocrinol. (Paris) 69, 116–122.
Köhrle, J., 2011. Environmental factors. In: Wass, J.A.H., Stewart, P.M., Amiel, A.S., Davies, M.C. (Eds.), Oxford Textbook of Endocrinology and Diabetes, second ed. Oxford University Press.
Köhrle, J., Auf’mkolk, M., Rokos, H., Hesch, R.D., Cody, V., 1986. Rat liver iodothyronine monodeiodinase. Evaluation of the iodothyronine ligand- binding site. J. Biol. Chem. 261, 11613–11622.
König, S., Moura Neto, V., 2002. Thyroid hormone actions on neural cells. Cell. Mol. Neurobiol. 22, 517–544.
Koopman-Esseboom, C., Morse, D.C., Weisglas-Kuperus, N., Lutkeschipholt, I.J., Van der Paauw, C.G., Tuinstra, L.G., Brouwer, A., Sauer, P.J., 1994. Effects of dioxins
and polychlorinated biphenyls on thyroid hormone status of pregnant women and their infants. Pediatr. Res. 36, 468–473.
Kovo, M., Golan, A., 2008. In vitro models using the human placenta to study fetal exposure to drugs. Clin. Med.: Reprod. Health 2, 15–24.
Krassas, G.E., Poppe, K., Glinoer, D., 2010. Thyroid function and human reproductive health. Endocr. Rev. 31, 702–755.
Kretschmer, X.C., Baldwin, W.S., 2005. CAR and PXR: xenosensors of endocrine disrupters? Chem. Biol. Interact. 155, 111–128.
Kreuchwig, A., Kleinau, G., Kreuchwig, F., Worth, C.L., Krause, G., 2011. Research resource: update and extension of a glycoprotein hormone receptors web application. Mol. Endocrinol. 25, 707–712.
Kre ws ki , D., And er se n, M.E ., Ma ntu s, E. , Zei se , L. , 2009. Tox ic it y test in g in the 21 st ce nt ur y: impl ic atio ns for hum an he alt h ri sk ass ess ment . Ris k An al. 29, 47 4–479.
Kudo, Y., Yamauchi, K., Fukazawa, H., Terao, Y., 2006. In vitro and in vivo analysis of the thyroid system-disrupting activities of brominated phenolic and phenol compounds in Xenopus laevis . Toxicol. Sci. 92, 87–95.
Kunisue, T., Fisher, J.W., Kannan, K., 2011. Determination of six thyroid hormones in the brain and thyroid gland using isotope-dilution liquid chromatography/ tandem mass spectrometry. Anal. Chem. 83, 417–424.
Langer, P., 1998. Minireview: polychlorinated biphenyls and the thyroid gland. Endocr. Regul. 32, 193–203.
Langer, P., Tajtakova, M., Fodor, G., Kocan, A., Bohov, P., Michalek, J., Kreze, A., 1998. Increased thyroid volume and prevalence of thyroid disorders in an area heavily polluted by polychlorinated biphenyls. Eur. J. Endocrinol. 139, 402–409.
Lans, M.C., Klasson-Wehler, E., Willemsen, M., Meussen, E., Safe, S., Brouwer, A., 1993. Structure-dependent, competitive interaction of hydroxy-polychlorobiphenyls,-dibenzo-p-dioxins and -dibenzofurans with human transthyretin. Chem. Biol.Interact. 88, 7–21.
Lans, M.C., Spiertz, C., Brouwer, A., Koeman, J.H., 1994. Different competition of thyroxine binding to transthyretin and thyroxine-binding globulin by hydroxy- PCBs, PCDDs and PCDFs. Eur. J. Pharmacol. 270, 129–136.
Larsen, P.R., 2009. Type 2 iodothyronine deiodinase in human skeletal muscle: new insights into its physiological role and regulation. J. Clin. Endocrinol. Metab. 94, 1893–1895.
Lecat-Guillet, N., Ambroise, Y., 2009. Synthesis and evaluation of imidazo[2,1- b]thiazoles as iodide efflux inhibitors in thyrocytes. ChemMedChem 4, 1819–1830.
Lecat-Guillet, N., Merer, G., Lopez, R., Pourcher, T., Rousseau, B., Ambroise, Y., 2007. A 96-well automated radioiodide uptake assay for sodium/iodide symporter inhibitors. Assay Drug Dev. Technol. 5, 535–540.
Lecat-Guillet, N., Merer, G., Lopez, R., Pourcher, T., Rousseau, B., Ambroise, Y., 2008. Small-molecule inhibitors of sodium iodide symporter function. ChemBioChem 9, 889–895.
Lecu re ux , L. , Diet er, M.Z ., Nels on , D.M., Wats on , L. , Won g, H. , Gemzi k, B. , Kla ass en, C.D ., Le hm an- McKe eman, L. D., 2009. He pa tob il ia ry dis pos it io n of thy ro id ho rm one in Mr p2-d eficie nt TR - ra ts: re du ce d bil ia ry exc re tio n of thy rox in e gl ucu ro ni de doe sno t pre ve nt xe no bio tic -i nd uc ed hyp oth yr oi dis m. To xi co l. Sci. 108, 482–491 .
Leonard, J.L., Rosenberg, I.N., 1980. Iodothyronine 50-deiodinase from rat kidney: substrate specificity and the 50-deiodination of reverse triiodothyronine. Endocrinology 107, 1376–1383.
Levy-Bimbot, M., Major, G., Courilleau, D., Blondeau, J.P., Levi, Y., 2012. Tetrabromobisphenol-A disrupts thyroid hormone receptor alpha function in vitro: use of fluorescence polarization to assay corepressor and coactivator peptide binding. Chemosphere 87, 782–788.
Li, J., Ma, M., Wang, Z., 2008. A two-hybrid yeast assay to quantify the effects of xenobiotics on retinoid X receptor-mediated gene expression. Toxicol. Lett. 176, 198–206.
Li, F., Xie, Q., Li, X., Li, N., Chi, P., Chen, J., Wang, Z., Hao, C., 2010. Hormone activity of hydroxylated polybrominated diphenyl ethers on human thyroid receptor-beta: in vitro and in silico investigations. Environ. Health Perspect. 118, 602–606.
Lindenthal, S., Lecat-Guillet, N., Ondo-Mendez, A., Ambroise, Y., Rousseau, B., Pourcher, T., 2009. Characterization of small-molecule inhibitors of the sodium iodide symporter. J. Endocrinol. 200, 357–365.
Liu, Y.Y., Brent, G.A., 2010. Thyroid hormone crosstalk with nuclear receptor signaling in metabolic regulation. Trends Endocrinol. Metab. 21, 166–173.
Liu, L., Klaassen, C.D., 1996. Regulation of hepatic sulfotransferases by steroidal chemicals in rats. Drug Metab. Dispos. 24, 854–858.
Lopez-Espinosa, M.J., Mondal, D., Armstrong, B., Bloom, M.S., Fletcher, T., 2012. Thyroid function and perfluoroalkyl acids in children living near a chemical plant. Environ. Health Perspect. 120, 1036–1041.
Lopez-Juarez, A., Remaud, S., Hassani, Z., Jolivet, P., Pierre Simons, J., Sontag, T., Yoshikawa, K., Price, J., Morvan-Dubois, G., Demeneix, B.A., 2012. Thyroid hormone signaling acts as a neurogenic switch by repressing Sox2 in the adult neural stem cell niche. Cell Stem Cell 10, 531–543.
Loubiere, L.S., Vasilopoulou, E., Bulmer, J.N., Taylor, P.M., Stieger, B., Verrey, F., McCabe, C.J., Franklyn, J.A., Kilby, M.D., Chan, S.Y., 2010. Expression of thyroid hormone transporters in the human placenta and changes associated with intrauterine growth restriction. Placenta 31, 295–304.
Louisse, J., de Jong, E., van de Sandt, J.J., Blaauboer, B.J., Woutersen, R.A., Piersma, A.H., Rietjens, I.M., Verwei, M., 2010. The use of in vitro toxicity data and physiologically based kinetic modeling to predict dose–response curves for in vivo developmental toxicity of glycol ethers in rat and man. Toxicol. Sci. 118, 470–484.
Lovell, M.A., Lynn, B.C., Xiong, S., Quinn, J.F., Kaye, J., Markesbery, W.R., 2008. An aberrant protein complex in CSF as a biomarker of Alzheimer disease. Neurology 70, 2212–2218.

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1343
Lueprasitsakul, W., Alex, S., Fang, S.L., Pino, S., Irmscher, K., Köhrle, J., Braverman, L.E., 1990. Flavonoid administration immediately displaces thyroxine (T4) from serum transthyretin, increases serum free T4, and decreases serum thyrotropin in the rat. Endocrinology 126, 2890–2895.
Luidens, M.K., Mousa, S.A., Davis, F.B., Lin, H.Y., Davis, P.J., 2010. Thyroid hormone and angiogenesis. Vascul. Pharmacol. 52, 142–145.
Mann, D.R., Plant, T.M., 2010. The role and potential sites of action of thyroid hormone in timing the onset of puberty in male primates. Brain Res. 1364, 175–185.
Montaño, M., Gutleb, A.C., Murk, A.J., Persistent toxic burdens of halogenated phenolic compounds in humans and wildlife, Environmental Science and Technology, submitted for publication.
Marchesini, G.R., Meulenberg, E., Haasnoot, W., Mizuguchi, M., Irth, H., 2006. Biosensor recognition of thyroid-disrupting chemicals using transport proteins. Anal. Chem. 78, 1107–1114.
Marchesini, G.R., Meimaridou, A., Haasnoot, W., Meulenberg, E., Albertus, F., Mizuguchi, M., Takeuchi, M., Irth, H., Murk, A.J., 2008. Biosensor discovery of thyroxine transport disrupting chemicals. Toxicol. Appl. Pharmacol. 232, 150–160.
Marsh, G., Athanasiadou, M., Athanassiadis, I., Sandholm, A., 2006. Identification of hydroxylated metabolites in 2,2 0 ,4,40-tetrabromodiphenyl ether exposed rats. Chemosphere 63, 690–697.
Martin, L.A., Wilson, D.T., Reuhl, K.R., Gallo, M.A., Klaassen, C.D., 2012. Polychlorinated biphenyl congeners that increase the glucuronidation and biliary excretion of thyroxine are distinct from the congeners that enhance the serum disappearance of thyroxine. Drug Metab. Dispos. 40, 588–595.
Massart, C., Hoste, C., Virion, A., Ruf, J., Dumont, J.E., Van Sande, J., 2011. Cell biology of H2O2 generation in the thyroid: investigation of the control of dual oxidases (DUOX) activity in intact ex vivo thyroid tissue and cell lines. Mol. Cell. Endocrinol. 343, 32–44.
Medina, D.L., Santisteban, P., 2000. Thyrotropin-dependent proliferation of in vitro rat thyroid cell systems. Eur. J. Endocrinol. 143, 161–178.
Meerts, I.A., van Zanden, J.J., Luijks, E.A., van Leeuwen-Bol, I., Marsh, G., Jakobsson, E., Bergman, A., Brouwer, A., 2000a. Potent competitive interactions of some brominated flame retardants and related compounds with human transthyretin in vitro. Toxicol. Sci. 56, 95–104.
Meerts, I.A.T.M., van Zanden, J.J., Luijks, E.A.C., van Leeuwen-Bol, I., Marsh, G., Jakobsson, E., Bergman, A., Brouwer, A., 2000b. Potent competitive interactions of some brominated flame retardants and related compounds with human transthyretin in vitro. Toxicol. Sci. 56, 95–104.
Meerts, I.A., Assink, Y., Cenijn, P.H., Van Den Berg, J.H., Weijers, B.M., Bergman, A., Koeman, J.H., Brouwer, A., 2002. Placental transfer of a hydroxylated polychlorinated biphenyl and effects on fetal and maternal thyroid hormone homeostasis in the rat. Toxicol. Sci. 68, 361–371.
Meijsing, S.H., Pufall, M.A., So, A.Y., Bates, D.L., Chen, L., Yamamoto, K.R., 2009. DNA binding site sequence directs glucocorticoid receptor structure and activity. Science 324, 407–410.
Miller, M.D., Crofton, K.M., Rice, D.C., Zoeller, R.T., 2009. Thyroid-disrupting chemicals: interpreting upstream biomarkers of adverse outcomes. Environ. Health Perspect. 117, 1033–1041.
Misiti, S., Schomburg, L., Yen, P.M., Chin, W.W., 1998. Expression and hormonal regulation of coactivator and corepressor genes. Endocrinology 139, 2493–2500.
Mitchell, A.M., Tom, M., Mortimer, R.H., 2005. Thyroid hormone export from cells: contribution of P-glycoprotein. J. Endocrinol. 185, 93–98.
Moeller, L.C., Dumitrescu, A.M., Refetoff, S., 2005. Cytosolic action of thyroid hormone leads to induction of hypoxia-inducible factor-1alpha and glycolytic genes. Mol. Endocrinol. 19, 2955–2963.
Montano, M., Cocco, E., Guignard, C., Marsh, G., Hoffmann, L., Bergman, A., Gutleb, A.C., Murk, A.J., 2012. New approaches assess the TTR binding capacity of bio- activated thyroid hormone disruptors. Toxicol. Sci. 130, 94–105.
Morimoto, E., Kanai, Y., Kim do, K., Chairoungdua, A., Choi, H.W., Wempe, M.F., Anzai, N., Endou, H., 2008. Establishment and characterization of mammalian cell lines stably expressing human L-type amino acid transporters. J. Pharmacol. Sci. 108, 505–516.
Moriyama, K., Tagami, T., Akamizu, T., Usui, T., Saijo, M., Kanamoto, N., Hataya, Y., Shimatsu, A., Kuzuya, H., Nakao, K., 2002. Thyroid hormone action is disrupted by bisphenol A as an antagonist. J. Clin. Endocrinol. Metab. 87, 5185–5190.
Morse, D.C., Brouwer, A., 1995. Fetal, neonatal, and long-term alterations in hepatic retinoid levels following maternal polychlorinated biphenyl exposure in rats. Toxicol. Appl. Pharmacol. 131, 175–182.
Morse, D.C., Groen, D., Veerman, M., van Amerongen, C.J., Koeter, H.B., Smits van Prooije, A.E., Visser, T.J., Koeman, J.H., Brouwer, A., 1993. Interference of polychlorinated biphenyls in hepatic and brain thyroid hormone metabolism in fetal and neonatal rats. Toxicol. Appl. Pharmacol. 122, 27–33.
Mor se , D.C ., Wess eli ng , W., Koe man, J.H ., Br ou wer, A., 199 6. Al te ra tio ns in ra t br ain thy roi d hor mon e st atus fol lo win g pr e and pos tna tal exp os ur e to po lyc hl or in ate dbip heny ls (Ar oc lo r 125 4). Tox ic ol . Ap pl .Ph arm acol . 13 6, 26 9–279.
Mortimer, R.H., Landers, K.A., Balakrishnan, B., Li, H., Mitchell, M.D., Patel, J., Richard, K., 2012. Secretion and transfer of the thyroid hormone binding protein transthyretin by human placenta. Placenta 33, 252–256.
NCATS, 2012. High-Throughput Screening Assay Guidance Criteria. Neumann, S., Kleinau, G., Costanzi, S., Moore, S., Jiang, J.K., Raaka, B.M., Thomas, C.J.,
Krause, G., Gershengorn, M.C., 2008. A low-molecular-weight antagonist for the human thyrotropin receptor with therapeutic potential for hyperthyroidism. Endocrinology 149, 5945–5950.
Neumann, S., Huang, W., Titus, S., Krause, G., Kleinau, G., Alberobello, A.T., Zheng, W., Southall, N.T., Inglese, J., Austin, C.P., Celi, F.S., Gavrilova, O., Thomas, C.J., Raaka, B.M., Gershengorn, M.C., 2009. Small-molecule agonists for the thyrotropin receptor stimulate thyroid function in human thyrocytes and mice. Proc. Natl. Acad. Sci. USA 106, 12471–12476.
Nguyen, N.H., Apriletti, J.W., Cunha Lima, S.T., Webb, P., Baxter, J.D., Scanlan, T.S., 2002. Rational design and synthesis of a novel thyroid hormone antagonist that blocks coactivator recruitment. J. Med. Chem. 45, 3310–3320.
NIEHS, 2007. Committee Proposes Paradigm Shift in Toxicity Testing, Environmental Factor. NIEHS. <http://www.niehs.nih.gov/news/newsletter/2007/september/toxicitytesting.cfm>.
Nillni, E.A., 2010. Regulation of the hypothalamic thyrotropin releasing hormone (TRH) neuron by neuronal and peripheral inputs. Front. Neuroendocrinol. 31, 134–156.
NRC, 1984. Toxicity Testing: Strategies to Determine Needs and Priorities. The National Academies Press, Washington, DC.
NRC, 2007. Toxicity Testing in the 21st Century: A Vision and a Strategy. National Academies Press, Washington, DC.
Nunez Miguel, R., Sanders, J., Sanders, P., Young, S., Clark, J., Kabelis, K., Wilmot, J., Evans, M., Roberts, E., Hu, X., Furmaniak, J., Rees Smith, B., 2012. Similarities and differences in interactions of thyroid stimulating and blocking autoantibodies with the TSH receptor. J. Mol. Endocrinol. 49, 137–151.
Obaidat, A., Roth, M., Hagenbuch, B., 2012. The expression and function of organic anion transporting polypeptides in normal tissues and in cancer. Annu. Rev. Pharmacol. Toxicol. 52, 135–151.
Obregon, M.J., 2008. Thyroid hormone and adipocyte differentiation. Thyroid 18, 185–195.
OEC D, 2006 . Detail ed Re vie w Pa pe r on Th yr oi d Hor mon e Dis ru pt io n Ass ays , pp. 1–43 4.Oetting, A., Yen, P.M., 2007. New insights into thyroid hormone action. Best Pract.
Res. Clin. Endocrinol. Metab. 21, 193–208.Otto, T., Fandrey, J., 2008. Thyroid hormone induces hypoxia-inducible factor 1alpha
gene expression through thyroid hormone receptor beta/retinoid x receptor alpha-dependent activation of hepatic leukemia factor. Endocrinology 149, 2241–2250.
Park, S.J., Kufareva, I., Abagyan, R., 2010. Improved docking, screening and selectivity prediction for small molecule nuclear receptor modulators using conformational ensembles. J. Comput. Aided Mol. Des. 24, 459–471.
Paroder-Belenitsky, M., Maestas, M.J., Dohan, O., Nicola, J.P., Reyna-Neyra, A., Follenzi, A., Dadachova, E., Eskandari, S., Amzel, L.M., Carrasco, N., 2011. Mechanism of anion selectivity and stoichiometry of the Na +/I� symporter(NIS). Proc. Natl. Acad. Sci. USA 108, 17933–17938.
Paschke, R., 2011a. Molecular pathogenesis of nodular goiter. Langenbecks Arch Surg 396, 1127–1136.
Paschke, R., 2011b. Nodulogenesis and goitrogenesis. Ann. Endocrinol. (Paris) 72, 117–119.
Pascual, A., Aranda, A., 2012. Thyroid hormone receptors, cell growth and differentiation. Biochim. Biophys. Acta.
Patel, A., Murray, J., McElwee-Whitmer, S., Bai, C., Kunapuli, P., Johnson, E.N., 2009. Acombination of ultrahigh throughput PathHunter and cytokine secretion assays to identify glucocorticoid receptor agonists. Anal. Biochem. 385, 286–292.
Patel, J., Landers, K., Li, H., Mortimer, R.H., Richard, K., 2011a. Delivery of maternal thyroid hormones to the fetus. Trends Endocrinol. Metab. 22, 164–170.
Patel, J., Landers, K., Li, H., Mortimer, R.H., Richard, K., 2011b. Thyroid hormones and fetal neurological development. J. Endocrinol. 209, 1–8.
Paul, K.B., Hedge, J.M., DeVito, M.J., Crofton, K.M., 2010. Short-term exposure to triclosan decreases thyroxine in vivo via upregulation of hepatic catabolism in Young Long-Evans rats. Toxicol. Sci. 113, 367–379.
Paul, K.B., Hedge, J.M., Bansal, R., Zoeller, R.T., Peter, R., DeVito, M.J., Crofton, K.M., 2012. Developmental triclosan exposure decreases maternal, fetal, and early neonatal thyroxine: a dynamic and kinetic evaluation of a putative mode-of- action. Toxicology 300, 31–45.
Pearce, E.N., Braverman, L.E., 2009. Environmental pollutants and the thyroid. Best Pract. Res. Clin. Endocrinol. Metab. 23, 801–813.
Persani, L., 1998. Hypothalamic thyrotropin-releasing hormone and thyrotropin biological activity. Thyroid 8, 941–946.
Pessemesse, L., Schlernitzauer, A., Sar, C., Levin, J., Grandemange, S., Seyer, P., Favier, F.B., Kaminski, S., Cabello, G., Wrutniak-Cabello, C., Casas, F., 2012. Depletion of the p43 mitochondrial T3 receptor in mice affects skeletal muscle development and activity. FASEB J. 26, 748–756.
Petrick, J.S., Klaassen, C.D., 2007. Importance of hepatic induction of constitutive androstane receptor and other transcription factors that regulate xenobiotic metabolism and transport. Drug Metab. Dispos. 35, 1806–1815.
Picchietti, S., Belardinelli, M., Taddei, A.R., Fausto, A.M., Pellegrino, M., Maggio, R., Rossi, M., Giorgi, F., 2009. Thyroid disruptor 1,1,1-trichloro-2,2-bis(p-chlorophenyl)ethane (DDT) prevents internalization of TSH receptor. Cell Tissue Res. 336, 31–40.
Piehl, S., Heberer, T., Balizs, G., Scanlan, T.S., Köhrle, J., 2008. Development of avalidated liquid chromatography/tandem mass spectrometry method for the distinction of thyronine and thyronamine constitutional isomers and for the identification of new deiodinase substrates. Rapid Commun. Mass Spectrom. 22, 3286–3296.
Piekielko-Witkowska, A., Nauman, A., 2011. Iodothyronine deiodinases and cancer. J. Endocrinol. Invest. 34, 716–728.
Pohlenz, J., Refetoff, S., 1999. Mutations in the sodium/iodide symporter (NIS) gene as a cause for iodide transport defects and congenital hypothyroidism. Biochimie 81, 469–476.

1344 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
Poncin, S., Gerard, A.C., Boucquey, M., Senou, M., Calderon, P.B., Knoops, B., Lengele, B., Many, M.C., Colin, I.M., 2008. Oxidative stress in the thyroid gland: from harmlessness to hazard depending on the iodine content. Endocrinology 149, 424–433.
Pop, V.J., Kuijpens, J.L., van Baar, A.L., Verkerk, G., van Son, M.M., de Vijlder, J.J., Vulsma, T., Wiersinga, W.M., Drexhage, H.A., Vader, H.L., 1999. Low maternal free thyroxine concentrations during early pregnancy are associated with impaired psychomotor development in infancy. Clin. Endocrinol. (Oxford) 50, 149–155.
Powell, K.A., Mitchell, A.M., Manley, S.W., Mortimer, R.H., 2000. Different transporters for tri-iodothyronine (T(3)) and thyroxine (T(4)) in the human choriocarcinoma cell line, JAR. J. Endocrinol. 167, 487–492.
Punt, A., Brand, W., Murk, A.J., van Wezel, A.P., Schriks, M., Heringa, M.B., 2013. Effect of combining in vitro estrogenicity data with kinetic characteristics of estrogenic compounds on the in vivo predictive value. Toxicol. In Vitro 27, 44–51.
Putcha, B.D., Wright, E., Brunzelle, J.S., Fernandez, E.J., 2012. Structural basis for negative cooperativity within agonist-bound TR:RXR heterodimers. Proc. Natl. Acad. Sci. USA 109, 6084–6087.
Qatanani, M., Zhang, J., Moore, D.D., 2005. Role of the constitutive androstane receptor in xenobiotic-induced thyroid hormone metabolism. Endocrinology 146, 995–1002.
Radovic, B., Mentrup, B., Köhrle, J., 2006. Genistein and other soya isoflavones are potent ligands for transthyretin in serum and cerebrospinal fluid. Br. J. Nutr. 95, 1171–1176.
Raldua, D., Andre, M., Babin, P.J., 2008. Clofibrate and gemfibrozil induce an embryonic malabsorption syndrome in zebrafish. Toxicol. Appl. Pharmacol. 228, 301–314.
Raucy, J.L., Lasker, J.M., 2010. Current in vitro high throughput screening approaches to assess nuclear receptor activation. Curr. Drug Metab. 11, 806–814.
Ravnum, S., Zimmer, K.E., Keune, H., Gutleb, A.C., Murk, A.J., Koppe, J.G., Magnanti, B., Lyche, J.L., Eriksen, G.S., Ropstad, E., Skaare, J.U., Kobernus, M., Yang, A., Bartonova, A., Krayer von Krauss, M., 2012. Policy relevant results from an expert elicitation on the human health risks of decabromodiphenyl ether (decaBDE) and hexabromocyclododecane (HBCD). Environ. Health 11 (Suppl. 1),S7.
Regard, E., Mauchamp, J., 1973. Development of peroxidase activity and organification of iodide in the thyroid gland of Xenopus laevis (Daudin) during metamorphosis. CR Acad. Sci. Hebd Seances Acad. Sci. D 276, 121–123.
Reif, D.M., Martin, M.T., Tan, S.W., Houck, K.A., Judson, R.S., Richard, A.M., Knudsen, T.B., Dix, D.J., Kavlock, R.J., 2010. Endocrine profiling and prioritization of environmental chemicals using ToxCast data. Environ. Health Perspect. 118, 1714–1720.
Reinehr, T., 2010. Obesity and thyroid function. Mol. Cell. Endocrinol. 316, 165–171.Renko, K., Hoefig, C.S., Hiller, F., Schomburg, L., Köhrle, J., 2012. Identification of
iopanoic acid as substrate of type 1 deiodinase by a novel nonradioactive iodide-release assay. Endocrinology 153, 2506–2513.
Rhoden, K.J., Cianchetta, S., Stivani, V., Portulano, C., Galietta, L.J., Romeo, G., 2007. Cell-based imaging of sodium iodide symporter activity with the yellow fluorescent protein variant YFP-H148Q/I152L. Am. J. Physiol. Cell Physiol. 292, C814–C823.
Ribeiro, M.O., 2008. Effects of thyroid hormone analogs on lipid metabolism and thermogenesis. Thyroid 18, 197–203.
Robbins, J., 2000. Thyroid hormone transport proteins and the physiology of hormone binding. In: Braverman, L.E., Utiger, R.D. (Eds.), Werner and Ingbar’s, The Thyroid, eighth ed. Lippincott Williams & Wilkins, Philadelphia, pp. 105–120.
Roberts, L.M., Woodford, K., Zhou, M., Black, D.S., Haggerty, J.E., Tate, E.H., Grindstaff, K.K., Mengesha, W., Raman, C., Zerangue, N., 2008. Expression of the thyroid hormone transporters monocarboxylate transporter-8 (SLC16A2) and organic ion transporter-14 (SLCO1C1) at the blood–brain barrier. Endocrinology 149, 6251–6261.
Roger, P.P., van Staveren, W.C., Coulonval, K., Dumont, J.E., Maenhaut, C., 2010. Signal transduction in the human thyrocyte and its perversion in thyroid tumors. Mol. Cell. Endocrinol. 321, 3–19.
Rosenfeld, M.G., Lunyak, V.V., Glass, C.K., 2006. Sensors and signals: a coactivator/ corepressor/epigenetic code for integrating signal-dependent programs of transcriptional response. Genes Dev. 20, 1405–1428.
Rossi, M., Dimida, A., Dell’anno, M.T., Trincavelli, M.L., Agretti, P., Giorgi, F., Corsini, G.U., Pinchera, A., Vitti, P., Tonacchera, M., Maggio, R., 2007. The thyroid disruptor 1,1,1-trichloro-2,2-bis(p-chlorophenyl)-ethane appears to be an uncompetitive inverse agonist for the thyrotropin receptor. J. Pharmacol. Exp. Ther. 320, 465–474.
Rossi, M., Dimida, A., Ferrarini, E., Silvano, E., De Marco, G., Agretti, P., Aloisi, G., Simoncini, T., Di Bari, L., Tonacchera, M., Giorgi, F., Maggio, R., 2009. Presence of a putative steroidal allosteric site on glycoprotein hormone receptors. Eur. J. Pharmacol. 623, 155–159.
Roth, S., Kinne, A., Schweizer, U., 2010. The tricyclic antidepressant desipramine inhibits T3 import into primary neurons. Neurosci. Lett. 478, 5–8.
Rotroff, D.M., Beam, A.L., Dix, D.J., Farmer, A., Freeman, K.M., Houck, K.A., Judson, R.S., LeCluyse, E.L., Martin, M.T., Reif, D.M., Ferguson, S.S., 2010. Xenobiotic- metabolizing enzyme and transporter gene expression in primary cultures of human hepatocytes modulated by ToxCast chemicals. J. Toxicol. Environ. Health B Crit. Rev. 13, 329–346.
Rotroff, D.M., Dix, D.J., Houck, K.A., Knudsen, T.B., Martin, M.T., McLaurin, K.W., Reif, D.M., Crofton, K.M., Singh, A.V., Xia, M., Huang, R., Judson, R.S., 2012. Using
in vitro high throughput screening assays to identify potential endocrine- disrupting chemicals. Environ. Health Perspect.
Safer, J.D., 2011. Thyroid hormone action on skin. Dermatoendocrinology 3, 211–215.
Santini, F., Vitti, P., Ceccarini, G., Mammoli, C., Rosellini, V., Pelosini, C., Marsili, A., Tonacchera, M., Agretti, P., Santoni, T., Chiovato, L., Pinchera, A., 2003. In vitro assay of thyroid disruptors affecting TSH-stimulated adenylate cyclase activity. J. Endocrinol. Invest. 26, 950–955.
Savu, L., Vranckx, R., Maya, M., Nunez, E.A., 1987. A thyroxine binding globulin (TBG)-like protein in the sera of developing and adult rats. Biochem. Biophys. Res. Commun. 148, 1165–1173.
Savu, L., Vranckx, R., Maya, M., Gripois, D., Blouquit, M.F., Nunez, E.A., 1989. Thyroxine-binding globulin and thyroxine-binding prealbumin in hypothyroid and hyperthyroid developing rats. Biochim. Biophys. Acta 992, 379–384.
Scanlan, T.S., 2009. Minireview: 3-Iodothyronamine (T1AM): a new player on the thyroid endocrine team? Endocrinology 150, 1108–1111.
Schapira, M., Raaka, B.M., Das, S., Fan, L., Totrov, M., Zhou, Z., Wilson, S.R., Abagyan, R., Samuels, H.H., 2003. Discovery of diverse thyroid hormone receptor antagonists by high-throughput docking. Proc. Natl. Acad. Sci. USA 100, 7354–7359.
Schmutzler, C., Bacinski, A., Gotthardt, I., Huhne, K., Ambrugger, P., Klammer, H., Schlecht, C., Hoang-Vu, C., Grüters, A., Wuttke, W., Jarry, H., Köhrle, J., 2007a. The ultraviolet filter benzophenone 2 interferes with the thyroid hormone axis in rats and is a potent in vitro inhibitor of human recombinant thyroid peroxidase. Endocrinology 148, 2835–2844.
Schmutzler, C., Gotthardt, I., Hofmann, P.J., Radovic, B., Kovacs, G., Stemmler, L., Nobis, I., Bacinski, A., Mentrup, B., Ambrugger, P., Grüters, A., Malendowicz, L.K., Christoffel, J., Jarry, H., Seidlova-Wuttke, D., Wuttke, W., Köhrle, J., 2007b. Endocrine disruptors and the thyroid gland – a combined in vitro and in vivo analysis of potential new biomarkers. Environ. Health Perspect. 115 (Suppl. 1),77–83.
Schoonen, W.G., Westerink, W.M., Horbach, G.J., 2009. High-throughput screening for analysis of in vitro toxicity. EXS 99, 401–452.
Schreiber, G., 2002. The evolutionary and integrative roles of transthyretin in thyroid hormone homeostasis. J. Endocrinol. 175, 61–73.
Schriks, M., Vrabie, C.M., Gutleb, A.C., Faassen, E.J., Rietjens, I.M., Murk, A.J., 2006. T- screen to quantify functional potentiating, antagonistic and thyroid hormone- like activities of poly halogenated aromatic hydrocarbons (PHAHs). Toxicol. In Vitro 20, 490–498.
Schröder-van der Elst, J.P., van der Heide, D., Rokos, H., Köhrle, J., Morreale de Escobar, G., 1997. Different tissue distribution, elimination, and kinetics of thyroxine and its conformational analog, the synthetic flavonoid EMD 49209 in the rat. Endocrinology 138, 79–84.
Schroder-van der Elst, J.P., Smit, J.W., Romijn, H.A., van der Heide, D., 2003. Dietary flavonoids and iodine metabolism. BioFactors 19, 171–176.
Schussler, G.C., 2000. The thyroxine-binding proteins. Thyroid 10, 141–149.Schuur, A.G., Brouwer, A., Bergman, A., Coughtrie, M.W., Visser, T.J., 1998a.
Inhibition of thyroid hormone sulfation by hydroxylated metabolites of polychlorinated biphenyls. Chem. Biol. Interact. 109, 293–297.
Schuur, A.G., Legger, F.F., van Meeteren, M.E., Moonen, M.J., van Leeuwen-Bol, I., Bergman, A., Visser, T.J., Brouwer, A., 1998b. In vitro inhibition of thyroid hormone sulfation by hydroxylated metabolites of halogenated aromatic hydrocarbons. Chem. Res. Toxicol. 11, 1075–1081.
Schuur, A.G., van Leeuwen-Bol, I., Jong, W.M., Bergman, A., Coughtrie, M.W., Brouwer, A., Visser, T.J., 1998c. In vitro inhibition of thyroid hormone sulfation by polychlorobiphenylols: isozyme specificity and inhibition kinetics. Toxicol. Sci. 45, 188–194.
Schuur, A.G., Bergman, A., Brouwer, A., Visser, T.J., 1999. Effects of pentachlorophenol and hydroxylated polychlorinated biphenyls on thyroid hormone conjugation in a rat and a human hepatoma cell line. Toxicol. In Vitro 13, 417–425.
Shah, I., Houck, K., Judson, R.S., Kavlock, R.J., Martin, M.T., Reif, D.M., Wambaugh, J., Dix, D.J., 2011. Using nuclear receptor activity to stratify hepatocarcinogens. PLoS ONE 6, e14584.
Sharlin, D.S., Visser, T.J., Forrest, D., 2011. Developmental and cell-specificexpression of thyroid hormone transporters in the mouse cochlea. Endocrinology 152, 5053–5064.
Sharma, V., Hays, W.R., Wood, W.M., Pugazhenthi, U., St Germain, D.L., Bianco, A.C., Krezel, W., Chambon, P., Haugen, B.R., 2006. Effects of rexinoids on thyrotrope function and the hypothalamic–pituitary–thyroid axis. Endocrinology 147, 1438–1451.
Shi, Y.B., 2009. Dual functions of thyroid hormone receptors in vertebrate development: the roles of histone-modifying cofactor complexes. Thyroid 19, 987–999.
Shiizaki, K., Asai, S., Ebata, S., Kawanishi, M., Yagi, T., 2010. Establishment of yeast reporter assay systems to detect ligands of thyroid hormone receptors alpha and beta. Toxicol. In Vitro 24, 638–644.
Shimada, N., Yamauchi, K., 2004. Characteristics of 3,5,3 0-triiodothyronine (T3)-uptake system of tadpole red blood cells: effect of endocrine-disrupting chemicals on cellular T3 response. J. Endocrinol. 183, 627–637.
Silva, J.E., 2011. Physiological importance and control of non-shivering facultative thermogenesis. Front. Biosci. (Schol. Ed.) 3, 352–371.
Sinz, M., Kim, S., Zhu, Z., Chen, T., Anthony, M., Dickinson, K., Rodrigues, A.D., 2006. Evaluation of 170 xenobiotics as transactivators of human pregnane X receptor (hPXR) and correlation to known CYP3A4 drug interactions. Curr. Drug Metab. 7, 375–388.

AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346 1345
Sirakov, M., Skah, S., Nadjar, J., Plateroti, M., 2012. Thyroid hormone’s action on progenitor/stem cell biology: new challenge for a classic hormone? Biochim. Biophys. Acta.
Sjodin, A., Hagmar, L., Klasson-Wehler, E., Bjork, J., Bergman, A., 2000. Influence of the consumption of fatty Baltic Sea fish on plasma levels of halogenated environmental contaminants in Latvian and Swedish men. Environ. Health Perspect. 108, 1035–1041.
Smita, S., Gupta, S.K., Bartonova, A., Dusinska, M., Gutleb, A.C., Rahman, Q., 2012. Nanoparticles in the environment: assessment using the causal diagram approach. Environ. Health 11 (Suppl. 1), S13.
Song, Y., Driessens, N., Costa, M., De Deken, X., Detours, V., Corvilain, B., Maenhaut, C., Miot, F., Van Sande, J., Many, M.C., Dumont, J.E., 2007. Roles of hydrogen peroxide in thyroid physiology and disease. J. Clin. Endocrinol. Metab. 92, 3764–3773.
Song, Y., Massart, C., Chico-Galdo, V., Jin, L., De Maertelaer, V., Decoster, C., Dumont, J.E., Van Sande, J., 2010. Species specific thyroid signal transduction: conserved physiology, divergent mechanisms. Mol. Cell. Endocrinol. 319, 56–62.
Song, M., Kim, Y.J., Song, M.K., Choi, H.S., Park, Y.K., Ryu, J.C., 2011. Identification of classifiers for increase or decrease of thyroid peroxidase activity in the FTC-238/ hTPO recombinant cell line. Environ. Sci. Technol. 45, 7906–7914.
Staudinger, J., Liu, Y., Madan, A., Habeebu, S., Klaassen, C.D., 2001. Coordinate regulation of xenobiotic and bile acid homeostasis by pregnane X receptor. Drug Metab. Dispos. 29, 1467–1472.
Sugiyama, S., Miyoshi, H., Yamauchi, K., 2005a. Characteristics of a thyroid hormone responsive reporter gene transduced into a Xenopus laevis cell line using lentivirus vector. Gen. Comp. Endocrinol. 144, 270–279.
Sugiyama, S., Shimada, N., Miyoshi, H., Yamauchi, K., 2005b. Detection of thyroid system-disrupting chemicals using in vitro and in vivo screening assays in Xenopus laevis . Toxicol. Sci. 88, 367–374.
Surks, M.I., Sievert, R., 1995. Drugs and thyroid function. N. Engl. J. Med. 333, 1688–1694.
Tan, Z.S., Vasan, R.S., 2009. Thyroid function and Alzheimer’s disease. J. Alzheimers Dis. 16, 503–507.
Taurog, A., 1970. Thyroid peroxidase-catalyzed iodination of thyroglobulin; inhibition by excess iodide. Arch. Biochem. Biophys. 139, 212–220.
Taylor, P.M., Ritchie, J.W., 2007. Tissue uptake of thyroid hormone by amino acid transporters. Best Pract. Res. Clin. Endocrinol. Metab. 21, 237–251.
Taylor, J.P., Metcalfe, R.A., Watson, P.F., Weetman, A.P., Trembath, R.C., 2002. Mutations of the PDS gene, encoding pendrin, are associated with protein mislocalization and loss of iodide efflux: implications for thyroid dysfunction in Pendred syndrome. J. Clin. Endocrinol. Metab. 87, 1778–1784.
Tiefenbach, J., Moll, P.R., Nelson, M.R., Hu, C., Baev, L., Kislinger, T., Krause, H.M., 2010. A live zebrafish-based screening system for human nuclear receptor ligand and cofactor discovery. PLoS ONE 5, e9797.
Timsit, Y.E., Negishi, M., 2007. CAR and PXR: the xenobiotic-sensing receptors. Steroids 72, 231–246.
Titus, S., Neumann, S., Zheng, W., Southall, N., Michael, S., Klumpp, C., Yasgar, A., Shinn, P., Thomas, C.J., Inglese, J., Gershengorn, M.C., Austin, C.P., 2008. Quantitative high-throughput screening using a live-cell cAMP assay identifies small-molecule agonists of the TSH receptor. J. Biomol. Screen. 13, 120–127.
Tonacchera, M., Pinchera, A., Dimida, A., Ferrarini, E., Agretti, P., Vitti, P., Santini, F., Crump, K., Gibbs, J., 2004. Relative potencies and additivity of perchlorate, thiocyanate, nitrate, and iodide on the inhibition of radioactive iodide uptake by the human sodium iodide symporter. Thyroid 14, 1012–1019.
Tong, Z., Li, H., Goljer, I., McConnell, O., Chandrasekaran, A., 2007. In vitro glucuronidation of thyroxine and triiodothyronine by liver microsomes and recombinant human UDP-glucuronosyltransferases. Drug Metab. Dispos. 35, 2203–2210.
Trajkovic-Arsic, M., Muller, J., Darras, V.M., Groba, C., Lee, S., Weih, D., Bauer, K., Visser, T.J., Heuer, H., 2010. Impact of monocarboxylate transporter-8 deficiencyon the hypothalamus–pituitary–thyroid axis in mice. Endocrinology 151, 5053–5062.
Unuane, D., Tournaye, H., Velkeniers, B., Poppe, K., 2011. Endocrine disorders &female infertility. Best Pract. Res. Clin. Endocrinol. Metab. 25, 861–873.
van de Ven, A.C., Muntjewerff, J.W., Netea-Maier, R.T., de Vegt, F., Ross, H.A., Sweep, F.C., Kiemeney, L.A., Vos, P.E., Buitelaar, J.K., Hermus, A.R., den Heijer, M., Janz in g, J.G ., 2012 . As so cia ti on be twe en thy roi d fun ctio n, thy ro id aut oi mmuni ty, and state and trait factors of depression. Acta Psychiatr. Scand. 126, 377–384.
van den Berg, K.J., 1990. Interaction of chlorinated phenols with thyroxine binding sites of human transthyretin, albumin and thyroid binding globulin. Chem. Biol. Interact. 76, 63–75.
van den Hove, M.F., Croizet-Berger, K., Jouret, F., Guggino, S.E., Guggino, W.B., Devuyst, O., Courtoy, P.J., 2006. The loss of the chloride channel, ClC-5, delays apical iodide efflux and induces a euthyroid goiter in the mouse thyroid gland. Endocrinology 147, 1287–1296.
van der Deure, W., Peeters, R., Visser, T., 2009. Molecular aspects of thyroid hormone transporters, including MCT8, MCT10 and OATPs, and the effects of genetic variation in these transporters. J. Mol. Endocrinol.
van der Deure, W.M., Peeters, R.P., Visser, T.J., 2010. Molecular aspects of thyroid hormone transporters, including MCT8, MCT10, and OATPs, and the effects of genetic variation in these transporters. J. Mol. Endocrinol. 44, 1–11.
Van Sande, J., Massart, C., Beauwens, R., Schoutens, A., Costagliola, S., Dumont, J.E., Wolff, J., 2003. Anion selectivity by the sodium iodide symporter. Endocrinology 144, 247–252.
Vandenberg, L.N., Chahoud, I., Padmanabhan, V., Paumgartten, F.J., Schoenfelder, G., 2010. Biomonitoring studies should be used by regulatory agencies to assess human exposure levels and safety of bisphenol A. Environ. Health Perspect. 118, 1051–1054.
Vandenhaute, E., Dehouck, L., Boucau, M.C., Sevin, E., Uzbekov, R., Tardivel, M., Gosselet, F., Fenart, L., Cecchelli, R., Dehouck, M.P., 2011. Modelling the neurovascular unit and the blood–brain barrier with the unique function of pericytes. Curr. Neurovasc. Res. 8, 258–269.
Vansell, N.R., Klaassen, C.D., 2002. Effect of microsomal enzyme inducers on the biliary excretion of triiodothyronine (T(3)) and its metabolites. Toxicol. Sci. 65, 184–191.
Vargas, F., Moreno, J.M., Rodriguez-Gomez, I., Wangensteen, R., Osuna, A., Alvarez- Guerra, M., Garcia-Estan, J., 2006. Vascular and renal function in experimental thyroid disorders. Eur. J. Endocrinol. 154, 197–212.
Vassart, G., Costagliola, S., 2011. G protein-coupled receptors: mutations and endocrine diseases. Nat. Rev. Endocrinol. 7, 362–372.
Vassart, G., Dumont, J.E., 1992. The thyrotropin receptor and the regulation of thyrocyte function and growth. Endocr. Rev. 13, 596–611.
Verhaeghe, E., Buisson, D., Zekri, E., Leblanc, C., Potin, P., Ambroise, Y., 2008. Acolorimetric assay for steady-state analyses of iodo- and bromoperoxidase activities. Anal. Biochem. 379, 60–65.
Verwei, M., van Burgsteden, J.A., Krul, C.A., van de Sandt, J.J., Freidig, A.P., 2006. Prediction of in vivo embryotoxic effect levels with a combination of in vitro studies and PBPK modelling. Toxicol. Lett. 165, 79–87.
Visser, T.J., 1996. Pathways of thyroid hormone metabolism. Acta Med. Austriaca 23, 10–16.
Visser, T.J., 2008. Hormone metabolism. In: DeGroot, L. (Ed.), Thyroid Disease Manager.
Visser, T.J., van Overmeeren, E., Fekkes, D., Docter, R., Hennemann, G., 1979. Inhibition of iodothyronine 50-deiodinase by thioureylenes; structure–activityrelationship. FEBS Lett. 103, 314–318.
Visser, T.J., Kaplan, M.M., Leonard, J.L., Larsen, P.R., 1983. Evidence for two pathways of iodothyronine 50-deiodination in rat pituitary that differ in kinetics, propylthiouracil sensitivity, and response to hypothyroidism. J. Clin. Invest. 71, 992–1002.
Visser, T.J., Kaptein, E., Gijzel, A.L., de Herder, W.W., Ebner, T., Burchell, B., 1993. Glucuronidation of thyroid hormone by human bilirubin and phenol UDP- glucuronyltransferase isoenzymes. FEBS Lett. 324, 358–360.
Visser, W.E., Friesema, E.C., Jansen, J., Visser, T.J., 2008. Thyroid hormone transport in and out of cells. Trends Endocrinol. Metab. 19, 50–56.
Visser, W.E., Wong, W.S., van Mullem, A.A., Friesema, E.C., Geyer, J., Visser, T.J., 2010. Study of the transport of thyroid hormone by transporters of the SLC10 family. Mol. Cell. Endocrinol. 315, 138–145.
Wagner, W.E., 2000. The Precautionary Principle and Chemical Regulation in the US. Hum. Ecol. Risk Assess.: Int. J. 6, 459–477.
Wagner, M.S., Wajner, S.M., Maia, A.L., 2008. The role of thyroid hormone in testicular development and function. J. Endocrinol. 199, 351–365.
Walker, S.P., Wachs, T.D., Gardner, J.M., Lozoff, B., Wasserman, G.A., Pollitt, E., Carter, J.A., 2007. Child development: risk factors for adverse outcomes in developing countries. Lancet 369, 145–157.
Waltz, F., Pillette, L., Ambroise, Y., 2010. A nonradioactive iodide uptake assay for sodium iodide symporter function. Anal. Biochem. 396, 91–95.
Warner, A., Mittag, J., 2012. Thyroid hormone and the central control of homeostasis. J. Mol. Endocrinol. 49, R29–R35.
Waung, J.A., Bassett, J.H., Williams, G.R., 2012. Thyroid hormone metabolism in skeletal development and adult bone maintenance. Trends Endocrinol. Metab. 23, 155–162.
Webb, P., Nguyen, N.H., Chiellini, G., Yoshihara, H.A., Cunha Lima, S.T., Apriletti, J.W.,Ribeiro, R.C., Marimuthu, A., West, B.L., Goede, P., Mellstrom, K., Nilsson, S., Kushner,P.J., Fletterick, R.J., Scanlan, T.S., Baxter, J.D., 2002. Design of thyroid hormonereceptor antagonists from first principles. J. Steroid Biochem. Mol. Biol. 83, 59–73.
Weiss, J.M., Andersson, P.L., Lamoree, M.H., Leonards, P.E., van Leeuwen, S.P., Hamers, T., 2009. Competitive binding of poly- and perfluorinated compounds to the thyroid hormone transport protein transthyretin. Toxicol. Sci. 109, 206–216.
Westholm, D.E., Stenehjem, D.D., Rumbley, J.N., Drewes, L.R., Anderson, G.W., 2009. Competitive inhibition of organic anion transporting polypeptide 1c1-mediated thyroxine transport by the fenamate class of nonsteroidal antiinflammatorydrugs. Endocrinology 150, 1025–1032.
Westholm, D.E., Marold, J.D., Viken, K.J., Duerst, A.H., Anderson, G.W., Rumbley, J.N., 2010. Evidence of evolutionary conservation of function between the thyroxine transporter Oatp1c1 and major facilitator superfamily members. Endocrinology 151, 5941–5951.
Wetmore, B.A., Wambaugh, J.F., Ferguson, S.S., Sochaski, M.A., Rotroff, D.M., Freeman, K., Clewell 3rd, H.J., Dix, D.J., Andersen, M.E., Houck, K.A., Allen, B., Judson, R.S., Singh, R., Kavlock, R.J., Richard, A.M., Thomas, R.S., 2012. Integration of dosimetry, exposure, and high-throughput screening data in chemical toxicity assessment. Toxicol. Sci. 125, 157–174.
Willett, C.E., Bishop, P.L., Sullivan, K.M., 2011. Application of an integrated testing strategy to the US EPA endocrine disruptor screening program. Toxicol. Sci. 123, 15–25.
Wirth, E.K., Sheu, S.Y., Chiu-Ugalde, J., Sapin, R., Klein, M.O., Mossbrugger, I., Quintanilla-Martinez, L., de Angelis, M.H., Krude, H., Riebel, T., Rothe, K., Köhrle,J., Schmid, K.W., Schweizer, U., Grüters, A., 2011. Monocarboxylate transporter 8deficiency: altered thyroid morphology and persistent high triiodothyronine/ thyroxine ratio after thyroidectomy. Eur. J. Endocrinol. 165, 555–561.

1346 AlberTinka J. Murk et al. / Toxicology in Vitro 27 (2013) 1320–1346
Wolff, J., 1998. Perchlorate and the thyroid gland. Pharmacol. Rev. 50, 89–105.Won g, H. , Le hm an-M cK eem an, L. D., Gr ubb , M.F ., Gr os sman, S.J ., Bh ask ara n, V.M ., Sol on ,
E. G. , She n, H.S ., Ge rs on , R. J., Ca r, B. D., Zh ao, B. , Gem zik , B. , 2005. Inc re ase dhe pato bi lia ry cle ar an ce of un con ju ga ted thy ro xin e de term in es DMP 904- ind uce dalt era tio ns in thy ro id ho rm on e ho meos tasi s in ra ts. To xic ol . Sci . 84, 23 2–242.
Woo d, W.J ., Ge ra ci, T. , Nil se n, A. , DeBa rb er , A. E. , Scanl an, T. S., 20 09. Iod ot hy ro na min esare ox id ativ ely de amin ated to io do thy ro ac etic aci ds in vi vo. Che mBi oC hem 10 ,361–365 .
Wu, S.Y., Green, W.L., Huang, W.S., Hays, M.T., Chopra, I.J., 2005. Alternate pathways of thyroid hormone metabolism. Thyroid 15, 943–958.
www.perkinelmer.com/GPCRcomplete, 2012. GPCRcomplete, <www.perkinelmer.com/GPCRcomplete>.
Yamauchi, K., Ishihara, A., Fukazawa, H., Terao, Y., 2003. Competitive interactions of chlorinated phenol compounds with 3,3 0 ,5-triiodothyronine binding to transthyretin: detection of possible thyroid-disrupting chemicals in environmental waste water. Toxicol. Appl. Pharmacol. 187, 110–117.
Yen, P.M., 2001. Physiological and molecular basis of thyroid hormone action. Physiol. Rev. 81, 1097–1142.
You, S.H., Gauger, K.J., Bansal, R., Zoeller, R.T., 2006. 4-Hydroxy-PCB106 acts as adirect thyroid hormone receptor agonist in rat GH3 cells. Mol. Cell. Endocrinol. 257–258, 26–34.
Zatelli, M.C., Gentilin, E., Daffara, F., Tagliati, F., Reimondo, G., Carandina, G., Ambrosio, M.R., Terzolo, M., Degli Uberti, E.C., 2010. Therapeutic concentrations of mitotane (o,p0-DDD) inhibit thyrotroph cell viability and TSH expression and secretion in a mouse cell line model. Endocrinology 151, 2453–2461.
Zhang, J., Huang, W., Qatanani, M., Evans, R.M., Moore, D.D., 2004. The constitutive androstane receptor and pregnane X receptor function coordinately to prevent bile acid-induced hepatotoxicity. J. Biol. Chem. 279, 49517–49522.
Zhang, L., Cooper-Kuhn, C.M., Nannmark, U., Blomgren, K., Kuhn, H.G., 2010a. Stimulatory effects of thyroid hormone on brain angiogenesis in vivo and in vitro. J. Cereb. Blood Flow Metab. 30, 323–335.
Zhang, Q., Bhattacharya, S., Andersen, M.E., Conolly, R.B., 2010b. Computational systems biology and dose–response modeling in relation to new directions in toxicity testing. J. Toxicol. Environ. Health B Crit. Rev. 13, 253–276.
Zhou, T., Ross, D.G., DeVito, M.J., Crofton, K.M., 2001. Effects of short-term in vivo exposure to polybrominated diphenyl ethers on thyroid hormones and hepatic enzyme activities in weanling rats. Toxicol. Sci. 61, 76–82.
Zimmer, K.E., Gutleb, A.C., Ravnum, S., Krayer von Krauss, M., Murk, A.J., Ropstad, E., Skaare, J.U., Eriksen, G.S., Lyche, J.L., Koppe, J.G., Magnanti, B.L., Yang, A., Bartonova, A., Keune, H., 2012. Policy relevant results from an expert elicitation on the health risks of phthalates. Environ. Health 11 (Suppl. 1), S6.
Zimmermann, M.B., Andersson, M., 2012. Update on iodine status worldwide. Curr. Opin. Endocrinol. Diabetes Obes. 19, 382–387.
Zimmermann-Belsing, T., Rasmussen, A.K., Feldt-Rasmussen, U., Bog-Hansen, T.C., 2002. The influence of alpha1-acid glycoprotein (orosomucoid) and its glycoforms on the function of human thyrocytes and CHO cells transfected with the human TSH receptor. Mol. Cell. Endocrinol. 188, 241–251.
Zoeller, R.T., 2005. Environmental chemicals as thyroid hormone analogues: new studies indicate that thyroid hormone receptors are targets of industrial chemicals? Mol. Cell. Endocrinol. 242, 10–15.
Zoeller, R.T., 2007. Environmental chemicals impacting the thyroid: targets and consequences. Thyroid 17, 811–817.
Zoeller, R.T., Rovet, J., 2004. Timing of thyroid hormone action in the developing brain: clinical observations and experimental findings. J. Neuroendocrinol. 16, 809–818.
![Atypical Thyroid Function Tests, Thyroid Hormone ... · Atypical Thyroid Function Tests, Thyroid Hormone Resistance [Atipik Tiroid Fonksiyon Testleri: Tiroid Hormon Direnci] Soner](https://img.pdfslide.us/doc/110x75/5c83755009d3f2be2a8b56f6/atypical-thyroid-function-tests-thyroid-hormone-atypical-thyroid-function.jpg)