Embed Size (px)
Citation preview

VOLUME 79, NUMBER 22 P H Y S I C A L R E V I E W L E T T E R S 1 DECEMBER 1997
fulethanhemald.
Mechanics of Microtubule-Based Membrane Extension
Deborah Kuchnir Fygenson,1,* John F. Marko,1,† and Albert Libchaber1,2
1Center for Studies in Physics and Biology, Rockefeller University, New York, New York 100212NEC Research, 4 Independence Way, Princeton, New Jersey 08540
(Received 30 September 1996)
We observe quasistatic deformation of lipid vesicles from within, due to the polymerization oconfined microtubules. A pair of long, narrow membrane sleeves appears, sheathing the microtubends as they grow. Spontaneous buckling reveals that the force generated can be greater2 pN. The evolution of shape and magnitude of force are consistent with a simple theory for tmembrane free energy. We consider a model of the force generating mechanism in which therfluctuations of the membrane are “rectified” by the binding of tubulin dimers to the microtubule en[S0031-9007(97)04450-5]
PACS numbers: 87.22.Bt, 47.20.Hw, 68.10.Cr, 82.65.Dp
].
r-ve
pe
in
r-gs.
lay
icle
Since it was discovered that the boundaries of and withliving cells are composed of lipid bilayer membrane[1] the material properties of such membranes and tmorphology of closed bilayers, or vesicles, have capturthe imagination of physicists. Early studies based on rblood cells [2], which have a two dimensional proteinetwork anchored to the membrane, have been superseby studies of cell-sized artificial vesicles of controlled lipidcomposition and no protein skeleton [3]. In such pursystems it has been possible to qualitatively reprodubiological phenomena such as budding and fission usidistinctly nonbiological changes in temperature, pH, anmembrane composition [4]. Technological advancesdirect manipulation of single vesicles using micropipette[5] and optical tweezers [6] has focused attention osurprising material and dynamic properties and away frothe complications of biological relevance. In this Lettewe turn again toward the world of proteins and cells anstudy vesicles deformed from within by the polymerizatioof one of the ubiquitous cytoskeletal fibers, microtubule
Microtubules by themselves have remarkable physicproperties [7,8]. These extraordinarily stiff, crystallinecylindrical aggregates of the protein tubulin are at thlimit of detectability with the light microscope and displayan intriguing nonequilibrium polymerization instabilitythat is essential to cellular locomotion and division [9Encapsulation of microtubules inside vesicles was firdemonstrated by Hotani and Miyamoto [10]. Similaexperiments have been done using actin filaments, anotimportant cytoskeletal fiber [11].
In this Letter, we observe and explain changes in veicle shape due to the growth of confined microtubuleWe place a lower bound on the mechanical force geneated and propose a model of the force generating meanism. The essential technique is an encapsulationpurified protein within vesicles of controlled composition, about 10mm in diameter. Our best results wereachieved with the freeze-thaw method [12] in which veicles (1–5 mgyml 60% DOPCy40% DOPS, Avanti) aremixed with 30mM tubulin in solution, rapidly frozen by
0031-9007y97y79(22)y4497(4)$10.00
insheedednded
ecengdinsnmr,dns.al,e
].strher
s-s.r-
ch-of
-
s-
immersion in liquid nitrogen, and thawed on ice [13We use temperature (4±C–37±C) to control microtubulenucleation and assembly [8] and video-enhanced diffeential interference contrast (DIC) microscopy to obsermicrotubules inside a vesicle and its subsequent shachanges [14].
The characteristic evolution of shapes is shownFig. 1. One or few microtubules initially deform a
FIG. 1. (a) A phospholipid vesicle deformed by 1 to3 microtubules (observed floating freely prior to vesicle defomation). The number at lower right is the length of the lonaxis in microns. This vesicle fortuitously stuck to the glasslide (spot below center), restricting its rotational diffusionThe final frame has a different vesicle since the other neverin the focal plane as af shape. (b) Numerical minimizationof the membrane free energy generates the observed vesshapes. See Table I.
© 1997 The American Physical Society 4497

VOLUME 79, NUMBER 22 P H Y S I C A L R E V I E W L E T T E R S 1 DECEMBER 1997
bye.
e
lyxr
-lyd-
o-ed
er
ed)
s
TABLE I. Observed vesicle shapes.Z is the length of thehorizontal axis (i.e., microtubule),a is the proportion of areastored in membrane undulations,r is the radius of the end capsandf ≠Fy≠Z is the force on the microtubule ends. Detailof the calculation are described in the caption of Fig. 3.
Z smmd a r smmd f (pN)
ellipsoid 5.00 0.16 1.05 0.22sphericylinder 5.99 0.11 0.92 0.25sphericylinder 7.00 0.048 0.84 0.27pointed prolate 7.99 0.022 0.49 0.72f 10.99 0.018 0.21 1.80f 14.96 0.015 0.13 2.94
near-spherical vesicle into an ellipsoid. Their rategrowth ,2 mmymin is slow compared to the slowest relaxation velocities of the vesicle,4 mmysec, so the de-formation is quasistatic and Brownian motion aligns thmicrotubules along a common axis [15]. In flaccid veicles, the ellipsoid gives way to a sphericylindrical shapAs visible thermal fluctuations diminish, the vesicle assumes a taut, pointed, prolate shape with the microtubualigned along the axis of rotational symmetry. Despiapparent end-on contact with the membrane, microtubgrowth does not slow. The vesicle develops regionsnegative curvature (“necks”) and collapses into a pairnarrow membrane tubes around the microtubule extreities. The resulting shape has a profile resembling tGreek letterf (Fig. 1). Thef shape accommodates awide range of microtubule lengths, from 2 to at lea10 times the initial vesicle diameter. As the arms etend, the central portion becomes increasingly sphericbut the overall morphology is preserved. Occasionally tcharacteristic microtubule dynamic instability [9] is observed, but surprisingly often, particularly at long lengthgrowth slows until the microtubules appear constantlength [16].
The membrane does not rupture. The microtubulesnot break. The vesicle regains its initial shape whethe microtubules disassemble completely. Variationsthe initial shape and size of the vesicle have sligeffects: Initially prolate vesicles bypass the ellipsoishape; initially taut vesicles bypass the sphericylindricshape; large vesicles (.20 mm diameter) are distorteddifferently because microtubules that span their diameare individually unstable to buckling [see Eq. (2)]; smavesicles (,0.5 mm diameter) are not distorted becausthe microtubule that spans the diameter depletestubulin supply so muchs.67%d that further growth andnucleation are suppressed.
Pulling on a flaccid vesicle with optical tweezers resulin a similar extension and relaxation of membrane sha(Fig. 2), indicating that long, narrow membrane extensiodo not require an attractive interaction between membraand microtubule [17]. We therefore look to understand tvesicle shapes from membrane elasticity alone.
The free energy of a vesicle has two components, odue to curvature and the other due to surface area exp
4498
,s
of-
es-e.-lesteuleofofm-he
stx-al,
he-s,in
doninhtdal
terllethe
tspensne
he
nean-
FIG. 2. (a) Closeup of a membrane extension generatedseveral growing microtubules encapsulated within a vesiclScale bar: 10mm. (b) A membrane extension drawn from afluctuating vesicle using optical tweezers (830 nm single-moddiode, 200 mW). Scale bar: 10mm. Arrowhead indicatesthe location of the tweezer. The vesicle is in an osmoticalmatched NaCl solution which creates a large refractive indemismatch with the interior for better contrast and easietweezing. The diameter of the extension is,1 mm.
sion [5,18]. At finite temperatures it is convenient to consider the contribution of large scale curvature separatefrom that of microscopic membrane undulations exciteby thermal energy. Equipartition yields an effective surface tension [18] which reflects the cost of increasing prjected area at the expense of flattening thermally excitundulations. A general expression for the free energy is
F 12 kc
Zs2Hd2d2r
1
(A0st0ygde2ga , a $ 0 ,A0st0yg 2 t0a 1
12 ksa2d, a # 0 ,
(1)
where H is the mean curvature,kc s,15kBT 6 3
10213 ergsd is the membrane bending modulus [5],A0
is the true membrane area [given by the (fixed) numbof lipid molecules at their preferred density], anda ;sA0 2 AdyA0 is a measure of the extent to which thetrue area is greater or less than the apparent (projectmembrane areaA. When A0 . A, a is the fraction ofmembrane area “stored” in undulations. WhenA0 ,
A, a is the relative area dilation. Equation (1) use
VOLUME 79, NUMBER 22 P H Y S I C A L R E V I E W L E T T E R S 1 DECEMBER 1997
ne
e
r
eles-e-
-h.
-
-
e
tn,
an empirical interpolation to preserve continuity of thfunction and its first derivative ata 0 [19]. Of theremaining parameters,g 8pkcykBT is a dimensionlessconstant, the prefactort0yg represents the membranetension ata 0 [18], and ks s,6 3 107kBTymm2 250 ergsycm2d is the membrane stretching modulus [5The initial a of a vesicle depends on the ratio of surfacto volume with which the vesicle formed.
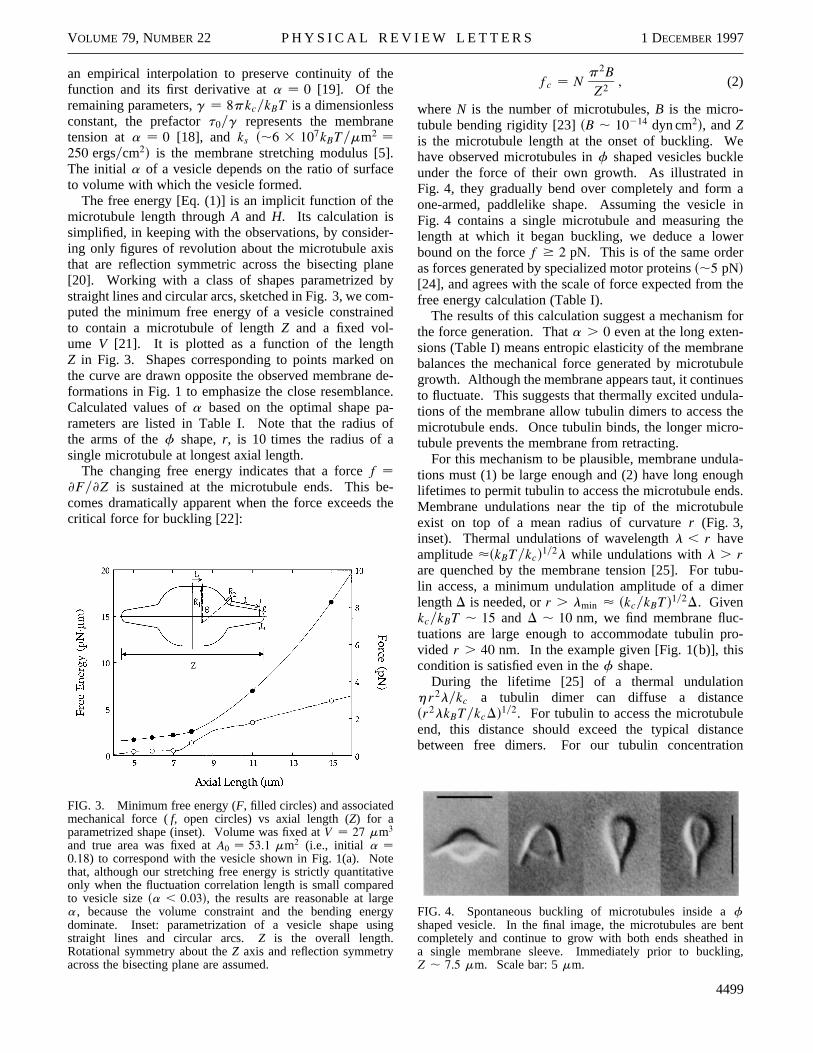
The free energy [Eq. (1)] is an implicit function of themicrotubule length throughA and H. Its calculation issimplified, in keeping with the observations, by consideing only figures of revolution about the microtubule axithat are reflection symmetric across the bisecting pla[20]. Working with a class of shapes parametrized bstraight lines and circular arcs, sketched in Fig. 3, we coputed the minimum free energy of a vesicle constrainto contain a microtubule of lengthZ and a fixed vol-ume V [21]. It is plotted as a function of the lengthZ in Fig. 3. Shapes corresponding to points markedthe curve are drawn opposite the observed membraneformations in Fig. 1 to emphasize the close resemblanCalculated values ofa based on the optimal shape parameters are listed in Table I. Note that the radiusthe arms of thef shape,r, is 10 times the radius of asingle microtubule at longest axial length.
The changing free energy indicates that a forcef ≠Fy≠Z is sustained at the microtubule ends. This bcomes dramatically apparent when the force exceedscritical force for buckling [22]:
FIG. 3. Minimum free energy (F, filled circles) and associatedmechanical force (f, open circles) vs axial length (Z) for aparametrized shape (inset). Volume was fixed atV 27 mm3
and true area was fixed atA0 53.1 mm2 (i.e., initial a 0.18) to correspond with the vesicle shown in Fig. 1(a). Notthat, although our stretching free energy is strictly quantitationly when the fluctuation correlation length is small compareto vesicle sizesa , 0.03d, the results are reasonable at larga, because the volume constraint and the bending enedominate. Inset: parametrization of a vesicle shape usstraight lines and circular arcs.Z is the overall length.Rotational symmetry about theZ axis and reflection symmetryacross the bisecting plane are assumed.
e
].e
r-sney
m-ed
onde-ce.-of
e-the
eved
ergying
fc Np2BZ2 , (2)
whereN is the number of microtubules,B is the micro-tubule bending rigidity [23]sB , 10214 dyn cm2d, andZis the microtubule length at the onset of buckling. Wehave observed microtubules inf shaped vesicles buckleunder the force of their own growth. As illustrated inFig. 4, they gradually bend over completely and form aone-armed, paddlelike shape. Assuming the vesicle iFig. 4 contains a single microtubule and measuring thlength at which it began buckling, we deduce a lowerbound on the forcef $ 2 pN. This is of the same orderas forces generated by specialized motor proteinss,5 pNd[24], and agrees with the scale of force expected from thfree energy calculation (Table I).
The results of this calculation suggest a mechanism fothe force generation. Thata . 0 even at the long exten-sions (Table I) means entropic elasticity of the membranbalances the mechanical force generated by microtubugrowth. Although the membrane appears taut, it continueto fluctuate. This suggests that thermally excited undulations of the membrane allow tubulin dimers to access thmicrotubule ends. Once tubulin binds, the longer microtubule prevents the membrane from retracting.
For this mechanism to be plausible, membrane undulations must (1) be large enough and (2) have long enouglifetimes to permit tubulin to access the microtubule endsMembrane undulations near the tip of the microtubuleexist on top of a mean radius of curvaturer (Fig. 3,inset). Thermal undulations of wavelengthl , r haveamplitudeøskBTykcd1y2l while undulations withl . rare quenched by the membrane tension [25]. For tubulin access, a minimum undulation amplitude of a dimerlengthD is needed, orr . lmin ø skcykBT d1y2D. GivenkcykBT , 15 and D , 10 nm, we find membrane fluc-tuations are large enough to accommodate tubulin provided r . 40 nm. In the example given [Fig. 1(b)], thiscondition is satisfied even in thef shape.
During the lifetime [25] of a thermal undulationhr2lykc a tubulin dimer can diffuse a distancesr2lkBTykcDd1y2. For tubulin to access the microtubuleend, this distance should exceed the typical distancbetween free dimers. For our tubulin concentration
FIG. 4. Spontaneous buckling of microtubules inside afshaped vesicle. In the final image, the microtubules are bencompletely and continue to grow with both ends sheathed ia single membrane sleeve. Immediately prior to bucklingZ , 7.5 mm. Scale bar: 5mm.
4499

VOLUME 79, NUMBER 22 P H Y S I C A L R E V I E W L E T T E R S 1 DECEMBER 1997
rte
n
of 30 mM, the constraint isr . 80 nm. Again, thiscondition is satisfied in our example [Fig. 1(b)].
We conclude that a force-generating mechaniswhereby thermal undulations of the membrane are retified by microtubule growth is likely to be responsiblefor force generation in this system. Specifically, wpropose that the binding energy of tubulin to the end othe microtubule drives the system out of equilibrium. Itrue, there should be a well-defined length, determinedthe initial ratio of surface area to volume of the vesicleat which microtubule growth will stop. Microtubules inf shaped vesicles that appear “stalled” indicate that supredictions can be tested experimentally [26].
The relevance of this work to processes in living cellswhere microtubule assembly rates and membrane elasticare quantitatively distinct from our model system, remainan open question. Observations of microtubule-basmembrane extensions in cells designed to generate excmicrotubules [27], and in cell extracts [28], as well aobservations of microtubules buckling in cells [29], invitespeculation that a similar mechanism may pertainin vivo.
We thank M. Betterton for early freeze-thaw andbuckling studies, M. Elbaum for the optical tweezesetup, and M. Goulian and H.-G. Döbereiner for helpfudiscussions. This work was supported in part by thNational Science Foundation through Grant No. Phy9408905, by the Mathers Foundation, and by the MeyFoundation (J. F. M.)
*Present address: University of Southern California, Department of Molecular Biology, Los Angeles, CA90087-1340.
†Present address: University of Illinois at ChicagoDepartment of Physics, 845 West Taylor Street, ChicagIL 60607-7059.
[1] A. D. Bangham and R. W. Horne, J. Mol. Biol.8, 660(1964).
[2] R. P. Rand, Biophys. J.4, 303 (1964); P. B. Canham,J. Theor. Biol.26, 61 (1970); E. Evans, R. Waugh, andL. Melnik, Biophys. J.16, 585 (1976).
[3] R. Lipowsky, Nature (London)349, 475 (1991).[4] K. Berndl et al., Europhys. Lett.13, 659 (1990); H.-G.
Döbereineret al., Biophys. J.65, 1396 (1993); E. Fargeand P. F. Devaux, Biophys. J.61, 347 (1992).
[5] E. Evans and W. Rawicz, Phys. Rev. Lett.64, 2094(1990); J. Song and R. E. Waugh, J. Biomech. Eng.112,235 (1990).
[6] R. Bar-Ziv, T. Frisch, and E. Moses, Phys. Rev. Lett75, 3481 (1995); G. Martinot-Lagardeet al., Ann. Phys.(France)20, 127 (1995).
[7] See, e.g., H. Flyvbjerg, T. Holy, and S. Leibler, PhysRev. Lett. 73, 2372 (1994); B. Houchmandzadeh andM. Vallade, Phys. Rev. E53, 6320 (1996).
[8] D. Kuchnir Fygenson, E. Braun, and A. LibchaberPhys. Rev. E50, 1579 (1994); D. Kuchnir Fygensonet al., Phys. Rev. E51, 5058 (1995); these references
4500
mc-
ef
fby,
ch
,itys
edesss
rle-
er
-
,o,
.
.
,
describe our experimental conditions, chosen to suppomarginal spontaneous nucleation of microtubules insidthe vesicles.
[9] T. Mitchison and M. Kirschner, Nature (London)312, 237(1984); M. Caplow, Curr. Opin. Cell Biol.4, 58 (1992).
[10] H. Hotani and H. Miyamoto, Adv. Biophys.26, 135(1990).
[11] J. D. Corteseet al., Proc. Natl. Acad. Sci. U.S.A.86, 5773(1989); H. Miyata and H. Hotani, Proc. Natl. Acad. Sci.U.S.A. 89, 11 547 (1992).
[12] R. R. C. New,Liposomes: a Practical Approach(Oxford,New York, 1990), pp. 58–60.
[13] Vesicles are swelled in buffer (100 mM Pipes, 2 mMEGTA, 2 mM MgSO4, 1 mM GTP, pH 6.9) at 37±C andchilled on ice before mixing with tubulin.
[14] Details of the apparatus are the same as in [8].[15] The relaxation velocity for membrane curvature on a
vesicle of radiusR in a medium of viscosityh is y kcyhR2. For our vesicles,kc , 10212 erg, h , 1022 P,and R , 5 mm. Transient shapes are observed whemicrotubule lengths change at a rate 10mmymin, ascommonly occurs during shortening.
[16] In contrast, microtubules in free solution rapidly shortenwhen growth is inhibited. R. A. Walker, N. K. Pryer, andE. D. Salmon, J. Cell Biol.114, 73 (1991).
[17] Extensions of much smaller diameters,50 nm “tethers”)result from pulling vesicles with fluid drag, beads, ormicropipettes. See, e.g., R. E. Waughet al., Biophys. J.61, 974 (1992).
[18] W. Helfrich, Z. Naturforsch.28C, 693 (1973); W. Helfrichand R. M. Servuss, Nuovo Cimento Soc. Ital. Fis.3D, 137(1984).
[19] t0 ksyg enforces continuity ofsd2Fyda2d at a 0,and gives good empirical agreement with data [5].
[20] S. Svetina and B. Zeks, Eur. Biophys. J.17, 101(1989).
[21] Solutes inside the vesicles.0.2 Md generate osmoticpressure which prevents volume change.1%.
[22] L. D. Landau and E. M. Lifshitz,Theory of Elasticity(Pergamon Press, Oxford, 1980), 3rd ed.
[23] M. Elbaum, D. Kuchnir Fygenson, and A. Libchaber,Phys. Rev. Lett.76, 4078 (1996), and references therein.
[24] J. T. Finer, R. M. Simmons, and J. A. Spudich, Nature(London) 368, 113 (1994); E. Meyhöfer and J. Howard,Proc. Natl. Acad. Sci. U.S.A.92, 574 (1995).
[25] S. T. Milner and S. A. Safran, Phys. Rev. A36, 4371(1987). This estimate does not account for local lipiddensity dynamics in each monolayer [U. Seifert andS. A. Langer, Europhys. Lett.23, 71 (1993)]. Onscales,100 nm, local density fluctuations retard bilayerrelaxation, reducing the lower boundr . 80 nm to r .
50 nm [26].[26] J. F. Marko and D. K. Fygenson (to be published).[27] K. Edson, B. Weisshaar, and A. Matus, Development117,
689 (1993); J. Knopset al., J. Cell Biol. 114, 725 (1991).[28] C. M. Waterman-Storeret al., J. Cell Biol. 130, 1161
(1995).[29] J. L. Travis and S. S. Bowser, Protoplasma154, 184
(1990); J. B. Tuckeret al., Cell Motil. Cytoskeleton25,49 (1993).




























