Embed Size (px)
Citation preview

23
Physiological and molecular mecha-nisms of resistance to drought inVitis vinifera L: aspects of primaryand secondary metabolisms andadaptation of genotypes
Abstract. In this review, we address the mainaspects of grapevine (Vitis vinifera L.) resistance towater stress. We point to examine physiological andmolecular answers, involving both primary and se-condary metabolisms. The grapevine has been gene-rally classified as drought-avoiding or as “pessimistic”following the ecological classification of plants into“pessimists” and “optimists”, even if differentgrapevine cultivars show different behaviours ofdefence against drought. This ecological classificationis analogous to the physiological classification intoisohydric and anisohydric plants and fundamentallylinked to stomatal behaviour. Stomatal regulation ofgrapevine is under hormone (ABA) and hydraulic con-trol, the latter linked to embolism formation and reco-very in xylem tissues up-stream the stomata. Wereview ABA effects on stomata, and their interrelation-ship with hydraulics of the plant. To this aim, we showthat in grapevine, xylem embolism occurs and repairsduring diurnal cycles, extreme water potential causingembolism, whereas an almost full recovery of waterpotential is needed to promote repair mechanisms. Inthis phase, an ABA-induced transpiration control isproposed. Aquaporins play fundamental role on waterstress control, and both early or late contribution(especially in root) are discussed. When drought-avoidance mechanisms are limiting in further plantdehydration avoidance (earlier in anisohydric varie-ties, but in all grape cultivars during late and/or severedrought stages) metabolic mechanisms of dehydra-tion tolerance arise. In all tissues solutes accumulateto prevent water loss, by minimizing interferences withcellular function. Grapevines accumulate one or moretypes of compatible solutes (osmotic adjustment).Metabolite profiling reveals that there are higher con-centrations of glucose, malate, proline, phenolics andflavour precursors upon water stress. The metabolitedifferences are linked to differences in transcript
abundance of many genes involved in energy metab-olism, particularly photosynthesis and photorespira-tion. Droughted grapevines appear to have a higherdemand than non-stressed plants to detoxify free radi-cals (reactive oxygen species), and to cope with pho-toinhibition. Results of these defence strategies accu-mulate in berries, during the ripening period, and areat the base of grape quality. Berry metabolites accu-mulated in several cultivars, as a response of con-trolled levels of water stress applied to the grapevine,are shown. Berry (skin, pulp and seed) phenolic frac-tions and flavours are reviewed and linked to pertur-bations of their molecular pathways, when known.Physiological, molecular and metabolic peculiaritiesgenotype-specific are underlined. Future perspectivesof research on the reviewed topic are proposed, asfocused on the interrelationship between ABA meta-bolism and molecular aspects of water control (aqua-porins).
Key words: aquaporin, abscisic acid (ABA), pheno-lics, flavour, transcriptomics.
Introduzione
La vite è la pianta da frutto più diffusa al mondo,sia in termini di produzione che di valore economico(66.271.676 Mt prodotte su 7.501.872 ha nel 2007, daFAOSTAT, http://faostat.fao.org/). Si tratta di unacoltura per tradizione non irrigua, diffusasi su un’am-pia area, anche in presenza di ecosistemi aridi esemiaridi. Il rendimento produttivo e qualitativo dellavite, da cui dipende la sua importanza economica, èstrettamente legato all’adattabilità di questa colturaalla siccità. Per la vite lo stress idrico non va pertantoesclusivamente considerato in termini negativi, secorrettamente gestito. Il controllo del livello di stressè infatti alla base di pratiche agronomiche affermate –uso del portinnesto, inerbimenti e lavorazioni control-lati, tecniche irrigue di soccorso quali la RDI, regula-ted deficit irrigation (Keller, 2005), o la PRD, partialrootzone drying (Dry e Loveys, 1999) – per inibire lacrescita vegetativa della pianta a vantaggio della qua-
Review n. 9 – Italus Hortus 16 (1), 2009: 23-44
Meccanismi fisiologici e molecolari di resistenza a stress idrico in Vitisvinifera L: aspetti del metabolismo primario e secondario e adattamentidi genotipi diversiAlessandra Ferrandino, Irene Perrone, Sara Tramontini e Claudio Lovisolo*
Dipartimento Colture Arboree, Università di Torino, via Leonardo da Vinci 44, Grugliasco (TO)
Ricezione: 28 novembre 2008; Accettazione: 12 gennaio 2009

Ferrandino et al.
24
lità del frutto. Lo stress idrico moderato sembra inol-tre promuovere la colonizzazione radicale da parte difunghi micorrizici arbuscolari (Schreiner et al., 2007).
In queste pagine verranno approfonditi alcuniaspetti della resistenza di Vitis vinifera a condizioni dicarenza idrica, con particolare attenzione alle risposteecofisiologiche e molecolari legate al metabolismoprimario e secondario, soprattutto a livello di bacca.
Nell’ultima parte saranno presi in esame gli aspettilegati alle peculiarità varietali, a tutti i livelli d’indagi-ne considerati e fino ai più recenti risultati pubblicati.
Meccanismi di elusione o tolleranza dello stressidrico
Come schematizzato in figura 1, il deficit idricoproduce svariate reazioni, a seconda che agisca nelbreve o nel lungo periodo. Nel primo caso, può favori-re la chiusura degli stomi, idropassiva ed idroattiva(quest’ultima controllata dall’acido abscissico), limi-tante la fotosintesi, l’abscissione fogliare e la rotturadella colonna d’acqua sotto tensione nello xilema(cavitazione). Nel secondo caso, può frenare lo svilup-po dell’area fogliare complessiva, agendo su numero edimensioni delle foglie, stimolare l’estensione dell’ap-parato radicale, provocare la sintesi di una spessa cuti-cola sulla superficie fogliare (Kramer e Boyer, 1995).
La vite bilancia lo stress idrico, per lo più prodottoda condizioni di siccità, eccessiva salinità e gelo, conun meccanismo di resistenza composto dielusione/tolleranza allo stress (da una definizione diLevitt, 1972). Con l’elusione la pianta tenta di mante-nere un alto potenziale idrico anche quando è sottopo-sta a condizioni di aridità. Quando questa condizionenon può più essere mantenuta e i tessuti soffrono lostress idrico, la pianta reagisce o mantenendo alto ilcontenuto idrico dei tessuti o tollerandone l’abbassa-mento. L’elusione agisce nel breve periodo sulla chiu-sura della rima stomatica e nel lungo su altri meccani-smi capaci di influenzare più o meno direttamente lagestione della riserva idrica a disposizione della pian-ta. Nel lungo periodo, perciò, si possono osservarecrescita degli elementi sotterranei (Soar e Loveys,2007) e abscissione volontaria delle foglie (con incre-mento del rapporto radici/fusto), modifiche dell’orien-tamento fogliare (Palliotti et al., 2008), variazioni deipigmenti fotosintetici, di dimensione e densità deglistomi (Gomez-Del-Campo et al., 2003) e degli ele-menti xilematici (Lovisolo e Schubert, 1998), incre-mento della capacità di immagazzinamento dell’acquanei tessuti (Patakas e Noitsakis, 1999), riduzione disuperficie e spessore fogliare (diretta conseguenzadelle limitazioni a divisione ed espansione cellulare),
inspessimento della cuticola, aumento della pubescen-za, variazioni nell’elasticità della parete cellulare(Patakas e Noitsakis, 1997), variazioni della composi-zione lipidica della foglia (Toumi et al., 2008) evariazioni nella permeabilità idrica della superficiefogliare (Keller, 2005). In tal modo e per condizionidi siccità temporanee o leggere, l’elusione può da solagarantire le performances della pianta.
Quando poi lo stress si acutizza, la regolazionedella fotosintesi è determinata da meccanismi nonstomatici: principalmente dalla riduzione dell’attivitàfotochimica, dalla depressione dell’attività degli enzi-mi fotosintetici, dall’abbassamento della conduttanzadel mesofillo e del tasso di trasporto elettronico sullamembrana del tilacoide e dalla ridotta capacità dirigenerazione della RuBisCO (Medrano et al., 2003).Oltre una certa soglia di stress, però, la pianta devecontare sulla propria tolleranza alla disidratazione.
I diversi eventi molecolari e le corrispondentirisposte metaboliche della pianta allo stress non pos-sono essere classificati come solo di tolleranza o solodi elusione, poiché agiscono in un continuum su varifronti in contemporanea, senza una progressionelineare su durata o intensità dello stress e regolati daun network di risposte complesse (Verslues et al.,2006); tuttavia verranno qui di seguito descritti inparagrafi distinti per favorirne la comprensione.
Meccanismi di elusione: la regolazione stomaticaLa prima reazione della vite allo stress idrico si
verifica a livello degli stomi, la cui apertura e chiusu-ra sono regolate attraverso meccanismi di tipo idrauli-co, ormonale e chimico. Allo scopo, entrano in giocovarie componenti: l’ABA (acido abscissico) inviatodalle radici con il flusso xilematico o redistribuito alivello fogliare, la domanda evaporativa, l’efficienzatraspirativa, la formazione di embolismi, la rapidità dicrescita fogliare, le componenti esogene origine deldeficit, nonché una certa influenza dovuta alla varia-bilità genetica.
Proprio in funzione del comportamento di control-lo stomatico, le specie vegetali possono essere classi-ficate come isoidriche o anisoidriche (Tardieu eSimonneau, 1998), riprendendo da un punto di vistafisiologico i concetti ecologici di piante “pessimiste”ed “ottimiste”. In condizioni limitanti, le prime tendo-no a modificare crescita e fisiologia per salvaguardareil più a lungo possibile il proprio stato idrico e le tem-poranee risorse, mentre le seconde a subire gli stresssenza evidenti reazioni di controllo del contenutod’acqua nei tessuti. Il meccanismo isoidrico, tipicodelle piante ecologicamente “pessimiste”, agisce sottostretto controllo stomatico.

Meccanismi di resistenza a stress idrico della vite
25
La vite è stata genericamente classificata comespecie isoidrica, capace di elevata regolazione stoma-tica sotto il controllo di segnale ABA, tuttavia esisteuna diffusa variabilità varietale che determina scosta-menti anche considerevoli da questo tipo di comporta-mento.
Il delicato meccanismo di regolazione stomaticache nella vite si attiva in risposta a condizioni dimoderate carenze idriche del suolo fa della conduttan-za stomatica stessa un indicatore dello stress idricopiù preciso e sensibile di altri parametri di misuradelle relazioni idriche (Cifre et al., 2005), primo fratutti il potenziale idrico.
Fig. 1 - Meccanismi di elusione o tolleranza dello stress idrico: il deficit idrico produce svariate reazioni, a seconda che agiscanel breve o nel lungo periodo.
Fig. 1 - Avoidance and tolerance of water stress in the grapevine: different responses are related to short or long term strategies.

Ferrandino et al.
26
La regolazione stomatica influenza direttamentel’assimilazione della CO2 fotosintetica. È probabileche, in condizioni di moderato stress idrico, la foto-sintesi nella vite venga depressa quasi esclusivamentea livello degli stomi, a conferma dell’incremento diefficienza intrinseca nell’uso dell’acqua, cosa peraltro presumibilmente comune a varie specie vegetali.In condizioni più limitanti, però, l’inibizione dellafotosintesi è provocata da fattori non direttamentelegati all’attività delle cellule di guardia (Flexas et al.,2004). Sebbene, infatti, l’effetto stomatico giochi unruolo determinante nella risposta della vite allo stressidrico, è stato dimostrato come effetti permanenti nonstomatici divengano più importanti mano a mano chele condizioni ambientali si fanno più limitanti(Escalona et al., 1999).
L’acido abscissico. Gli stomi si chiudono per indu-zione ormonale originata dalle radici le quali, in penu-ria d’acqua, inviano messaggeri biochimici, in parti-colare l’ABA, verso gli organi fogliari attraverso ilflusso di linfa grezza del legno. L’ABA, in sinergiacon la concentrazione cellulare di ioni Ca2+, ha uneffetto stimolante sulle strutture radicali, mentre suquelle stomatiche favorisce la chiusura delle cellule diguardia (Quintero et al., 1999), e non produce nessuneffetto su sviluppo e funzionalità idraulica dei tessutixilematici (Lovisolo et al., 2002).
In presenza di stress idrico, l’ABA è ridistribuitonella foglia a seguito dell’alcalinizzazione del succoxilematico. In condizioni naturali, infatti, il succoxilematico è debolmente acido, cosa che favoriscel’assorbimento della forma indissociata di ABA(ABAH) da parte delle cellule del mesofillo. Al verifi-carsi dello stress, il succo xilematico diventa debol-mente alcalino, favorendo la dissociazionedell’ABAH in ABA-, che non permea la membranacellulare. Di conseguenza le cellule del mesofilloassorbono meno ABA, rendendone disponibile unaquantità maggiore che raggiunge le cellule di guardia,dotate di canali ionici specifici. Gli effetti del segnaledi ABA sono avvertiti dagli stomi della vite non soloin stress idrico (vedi bibliografia in Lovisolo et al.,2002), ma anche nelle prime fasi della reidratazione,quando i flussi xilematici dilavano l’elevato ABAresiduo radicale e lo trasportano verso le rime stoma-tiche (Lovisolo et al., 2008a).
L’importanza della relazione tra ABA, conduttan-za stomatica e stato idrico della pianta (potenzialeidrico) si può evincere da un modello di simulazione(Tardieu, 2003) col quale è stato confrontato il com-portamento di quattro genotipi virtuali che presentinodifferenti abilità nella sintesi di ABA in condizioni di
stress idrico. Nel virtuale wild type il potenziale idricofogliare non varia nella prima settimana di carenzad’acqua per poi decrescere progressivamente fino a -1,5 MPa nelle successive due settimane, secondoun trend piuttosto comune in differenti esperienzereali su vite. Al contrario, il potenziale quasi nonvaria in un genotipo virtuale capace di un tasso di sin-tesi ABA doppio, mentre crolla in cinque giorni o indue giorni a -2,0 MPa in due genotipi virtuali la cuisintesi di ABA è rispettivamente la metà o un quintodel controllo wild type.
Formazione e recupero degli embolismi. L’ABAsintetizzato in condizioni di stress idrico, e più ingenerale i segnali di tipo chimico, controllano peròsolo parzialmente la risposta stomatica nella vite:sembrano essere i segnali di tipo idraulico i primi adessere avvertiti (Rodrigues et al., 2008).
La struttura dello xilema garantisce una semplicevia di transito a bassa resistenza attraverso cui l’acquapuò muoversi liberamente in risposta a gradienti dipressione o tensione, richiedendo una forza motrice1010 volte inferiore rispetto a quella necessaria a spo-stare l’acqua attraverso la cellula. Struttura e dimen-sioni delle diverse subunità xilematiche, poi, sonodeterminanti nell’efficienza della conduttività(Lovisolo e Schubert, 1998). L’acqua scorre all’inter-no dello xilema in minima parte sospinta dalla pres-sione radicale positiva (quasi solo al germogliamento)ed in massima parte attirata verso l’apice dalla fortepressione idrostatica negativa (tensione) che la traspi-razione produce. Quest’elevata tensione, che tende-rebbe a far collassare gli elementi di trasporto, vienesopportata grazie alla lignificazione dei tessuti vasali,ma con un’importante conseguenza sul liquido: favo-risce la separazione dei gas che vi sono disciolti. Ècosì che si formano le bolle di gas all’interno dei tes-suti xilematici per il fenomeno detto di cavitazione.Secondo l’ipotesi più accreditata (Zimmermann,1983), i gas che penetrano nei condotti xilematiciattraverso i micropori, una volta all’interno, produr-rebbero la perdita di coesione fra le molecole d’ac-qua. Questo meccanismo è frequente e minimizzabiledalla pianta: infatti i piccoli pori che connettono levarie unità costituenti dello xilema impediscono allabolla di espandersi all’infinito, grazie alla tensionesuperficiale che l’acqua adiacente oppone a questolivello.
Se la vite non è in grado di evitare l’embolizzazio-ne, ha i mezzi per limitare il fenomeno: l’embolizza-zione, infatti, causa in primo luogo una riduzionedella conduttività idraulica, che a sua volta, limitato ilflusso idrico, provoca la chiusura degli stomi

Meccanismi di resistenza a stress idrico della vite
27
(Schultz, 2003). Ne risultano un abbassamento del-l’efficienza traspirativa, quindi della tensione disuzione dell’acqua e, in ultima analisi, delle condizio-ni favorevoli al (ri-)verificarsi dell’evento.
Esistono poi vie alternative, fornite dai capillarivicini e comunicanti, che permettono al flusso idricodi proseguire aggirando l’ostacolo, e meccanismi direcupero dell’embolismo. Inoltre, quando durante leore pomeridiane e la notte la traspirazione si riduce,anche la tensione dello xilema scende e i gas sonoriassorbiti, almeno parzialmente, nel liquido circolan-te. La dinamica di recupero degli embolismi nella vitesegue un modello proposto di recente (Lovisolo et al.,2008a) secondo tre punti principali: i) il recuperoavviene in primo luogo a livello di fusto e poi in radi-ce e piccioli fogliari, ma solo a seguito di risalita delpotenziale idrico; ii) il recupero avviene anche duran-te le ore del giorno, mentre la vite traspira; ma iii) latraspirazione nelle fasi di recupero è ridotta da unsegnale di ABA, residuale dello stato di stress.
Ruolo delle acquaporine. L’acqua si muove all’in-terno delle strutture vegetali attraverso tre diverse vie:apoplastica, simplastica e transmembrana. La via apo-plastica individua il passaggio del liquido attraverso ilcontinuum di pareti cellulari e spazi aeriferi intercellu-lari che incontra nei tessuti della pianta, senza pene-trare la membrana cellulare. Questo tipo di trasporto èguidato da forze fisiche e regolato principalmente dadifferenze di potenziale tra suolo, pianta e atmosfera(Lovisolo et al., 2002). La via transmembrana, concui l’acqua attraversando il citoplasma percorre alme-no due volte, in entrata ed in uscita la membrana e lavia simplastica, creata dai plasmodesmi quale passag-gio preferenziale fra due cellule contigue, sono invecedifficilmente distinguibili sperimentalmente e comu-nemente riunite sotto la più ampia definizione di viacellulare (Steudle, 2000).
La conduttanza della via cellulare è fortementeinfluenzata dalla presenza e attività delle acquaporine(Vandeleur et al., 2008). Un aspetto interessante delleacquaporine vegetali consiste nella loro abbondanza ecomplessità: basti pensare che solo in Vitis sp. nesono state identificate 28, attraverso analisi del geno-ma sequenziato (Fouquet et al., 2008). Come giàaccennato, è intuitivo arrivare a comprendere l’impor-tanza fisiologica che le acquaporine sono in grado dirivestire per le piante, in tutti i processi fondamentaliin cui entra in gioco il trasporto idrico, dall’assorbi-mento radicale, alla crescita cellulare (Maurel et al.,2008), alla traspirazione e al controllo dell’embolizza-zione (Kaldenhoff et al., 2008). Il fatto che questeproteine facilitino il flusso di molecole d’acqua attra-
verso la membrana, non è, di per sé, garanzia di unmiglior funzionamento del sistema: piante di tabaccoin cui la sintesi di una specifica acquaporina, precisa-mente la PIP1, era stata geneticamente incrementata,hanno mostrato sì un aumento di efficienza fotosinte-tica e traspirativa, ma anche di velocità di appassi-mento (Aharon et al., 2003).
Questi risultati possono essere interpretati comeutili delucidazioni sulle reciproche proporzioni entrocui l’acqua utilizza nella pianta il cammino endo- edesocellulare. Se quest’ultimo, infatti, è strettamentegovernato da forze fisiche, controllato dai salti dienergia potenziale esistenti fra suolo, pianta e atmo-sfera e stimolato dalla traspirazione, il primo dipendeprevalentemente da numero e funzionalità delleacquaporine. Una pianta con “troppi” canali per l’ac-qua, come osservabile in individui mutanti, potrebbetraspirare di più, producendo un flusso idrico di mag-gior portata; in tal caso, all’alta traspirazione corri-sponderanno un’elevata fotosintesi e un aspetto vigo-roso. Un sistema così esigente non è però sempre ingrado di far fronte a condizione di stress idrico e/osalino.
Il mezzo con cui questi canali facilitano il flussodell’acqua è la riduzione dell’energia di attivazioneper il trasporto. Le molecole d’acqua che utilizzanoquesto cammino preferenziale, infatti, possono supe-rare l’alta barriera energetica prodotta dall’acquaincanalata all’interno della fase idrofobica del doppiostrato fosfolipidico attraverso un flusso passivo. Se lafunzionalità delle acquaporine viene meno, l’energiadi attivazione cresce immediatamente, secondo uneffetto che può essere provocato artificialmente ricor-rendo all’uso di reagenti mercuriali. Il mercurio inte-ragisce con i legami solfidrilici presenti nella strutturaproteica e dovuti ai residui di cisteina, posti in specifi-ci punti della molecola. Il blocco così ottenuto èreversibile per somministrazione di composti riducen-ti, come il ß-mercaptoetanolo. L’utilizzo di questesostanze presenta però dei limiti: prima di tutto, lasensibilità agli agenti mercuriali non è una caratteri-stica comune a tutte le acquaporine, mentre lo è adaltri trasportatori di membrana; poi, il mercurio pro-duce una serie di effetti negativi, come la depolarizza-zione delle membrane e l’inibizione del metabolismoin generale, non sempre riconoscibili e quantificabili(Tyerman et al., 2002). Ciononostante, continua adessere un utile supporto in indagini sperimentali sutessuti e organismo.
In vite, l’uso del mercurio ha dimostrato che ilrecupero degli embolismi necessita di apporto meta-bolico e non si verifica se l’attività cellulare, e quelladelle acquaporine, viene inibita (Lovisolo e Schubert,

Ferrandino et al.
28
2006). Inoltre, a livello radicale, ha permesso di sug-gerire che i portinnesti ibridi di Vitis rupestris conferi-scono resistenza a stress idrico maggiore che gli ibrididi Vitis riparia, grazie ad un miglior controllo del-l’embolizzazione (Lovisolo et al., 2008b).
Il ruolo delle acquaporine è nella vite funzione deltipo di tessuto, dello stadio di sviluppo e delle condi-zioni ambientali (Galmés et al., 2007): in effetti, lapermeabilità all’acqua, può essere anche una caratteri-stica che la pianta rifugge, come avviene in caso dicondizioni idriche limitanti. Per far fronte alle esigen-ze ambientali, l’attività dei canali per l’acqua è rego-lata in fase di trascrizione genica in funzione di mes-saggi di tipo ormonale (Kaldenhoff et al., 2008), e alivello post-traduzionale attraverso reazioni di fosfori-lazione e defosforilazione (Maurel et al., 2008).L’espressione di alcuni geni è stimolata dalla siccità edagli stress salini (Yamada et al., 1997) mentre, peraltri, non subisce alcuna influenza di tipo ambientale.In condizioni avverse, la capacità delle cellule radicalidi interrompere chimicamente questo flusso trans-membrana preferenziale potrebbe essere un mezzo perlimitare le perdite d’acqua, quando il potenziale idricodel suolo è inferiore a quello delle radici (Martre etal., 2001). Considerando che nelle radici la condutti-vità idraulica del trasporto attraverso le strutture cellu-lari (perciò tramite le vie transmembrana e simplasti-ca) è circa tre volte quella della via apoplastica(Siefritz et al., 2002), l’importanza del suddetto mec-canismo risulta evidente.
Non si può infine escludere che le acquaporineseguano speciali modelli d’espressione spaziali e tem-porali la cui attivazione, indotta da stress abiotici, siverifica solo in specifici tessuti e fasi di sviluppo(Aharon et al., 2003).
Le acquaporine della vite vengono differentementeespresse e/o attivate in condizioni di stress idrico,come discusso più avanti.
Meccanismi di tolleranza dello stress idricoLo stress idrico limita la fotosintesi non soltanto
attraverso la chiusura degli stomi, ma anche causandoalterazioni metaboliche dell’apparato fotosintetizzan-te, tanto più evidenti quanto più lo stato di stress siprotrae nel tempo (Escalona et al., 1999; Chaves,2002). Nella vite la regolazione non stomatica dellafotosintesi in condizioni di severo stress idrico è evi-denziata da parecchi indizi. All’aumentare dellacarenza idrica, il corrispondente calo nei parametrifotosintetici (assimilazione netta di CO2, assimilazio-ne in condizioni ottimali di luce e di CO2, tasso di tra-sporto elettronico, efficienza di carbossilazione) èinfatti molto inferiore alla diminuzione di conduttanza
stomatica (Cifre et al., 2005). Inoltre, in condizionicolturali tipiche, lo stress idrico si associa spesso adalte temperature ed irraggiamento, condizioni cheesasperano fotorespirazione e fotoinibizione (Gamone Pearcy, 1990 a, b).
La prima, insieme a retroregolazione del fotosiste-ma II e assimilazione fotosintetica di CO2, contrastal’eccessiva fotoriduzione di O2, e perciò il danno ossi-dativo a membrane cellulari e apparato fotosintetico,causato dalle specie attive dell’ossigeno prodottedurante il ciclo perossidatico di Mehler-Asada, di ces-sione di elettroni all’ossigeno (Osmond et al., 1997;Asada, 1999). Efficienza ed efficacia del meccanismofotorespirativo e perciò resistenza della pianta allecondizioni di stress, sembrano essere in V. viniferastrettamente dipendenti dal tipo di cultivar in esame(Guan et al., 2004).
La seconda invece è conseguenza del danneggia-mento dell’apparato enzimatico fotosintetico prodottoda un eccesso di fotoni e porta a una riduzione del-l’efficienza fotochimica e perciò della capacità foto-sintetica della pianta (Long et al., 1994). Per altro,data la particolare efficienza in V. vinifera di fotosin-tesi e fotorespirazione nel dissipare l’energia in ecces-so (Medrano et al., 2002) e trattandosi di un meccani-smo largamente studiato in condizioni controllate,perciò di ancor dubbia significatività per le piante incampo (Bertamini e Nedunchezhian, 2002), una stret-ta causalità e/o sinergia tra danni da stress idrico efotoinibizione non è certa.
A livello cellulare si innescano meccanismi didifesa per evitare la disidratazione cellulare, o per tol-lerarla. I primi sono alla base della regolazione osmo-tica, ovvero dell’accumulo di soluti compatibili, chenon interferiscono col metabolismo cellulare, ma con-tribuiscono a mantenere un buon equilibrio osmoticoin condizioni di stress (Bertamini et al., 2006). Nellavite, oltre gli zuccheri, è la prolina il più studiato ditali soluti (Keller, 2005, Wada et al., 2008). I mecca-nismi di tolleranza dello stato di disidratazione ruota-no invece intorno alla biosintesi di soluti e/o proteinedi protezione (ad es. deidrine, LEA Late-Embryogenesis-Abundant proteins) o di difesa (odetossificazione) dalle specie reattive dell’ossigeno.Tali meccanismi vengono esaminati in vite discuten-done le vie biosintetiche nel paragrafo successivo,mentre qui di seguito riportiamo gli effetti metabolicidi tali difese nell’acino d’uva.
Effetti dello stress idrico sui principali metabolitisecondari della bacca. I primi lavori riguardanti glieffetti del regime idrico sulla qualità dell’uva risalgo-no agli anni ottanta del secolo scorso. All’epoca l’in-

Meccanismi di resistenza a stress idrico della vite
29
teresse era soprattutto focalizzato sullo studio deiparametri base della qualità dell’uva (contenuto zuc-cherino, acidità, pH) poi, via via, anche grazie all’affi-narsi ed alla diffusione di tecniche analitiche più sofi-sticate ed efficaci, si sono approfonditi gli effetti dellostato idrico della pianta sui metaboliti secondari dellabacca, polifenoli ed aromi in particolare.
Visto che numerosi metaboliti secondari come icomposti di natura fenolica, i carotenoidi, i terpeni, lemetossipirazine e altri ancora sono di fatto molecoledi protezione di funzioni metaboliche vitali della cel-lula, la cui concentrazione può variare a seguito distress biotici e/o abiotici, lo studio delle loro variazio-ni diventa fondamentale per la valutazione di ciò cheè definita come ‘qualità’ dell’uva, che ha necessaria-mente importanti ricadute applicative a livello di tec-nologia della vinificazione e di ‘qualità’ del vino.
Polifenoli. Alla fine degli anni ‘80, Matthews eAnderson (1988) in uno studio su Cabernet franc inCalifornia misero in evidenza gli effetti del deficitidrico sulla qualità dell’uva, distinguendo gli effettiimputabili all’applicazione di stress idrico in epocaanticipata (prima dell’invaiatura) rispetto a quellidovuti all’applicazione dello stesso posticipatamente(dopo l’invaiatura). La concentrazione dei polifenolidelle bucce incrementava se la pianta era sottoposta acondizioni di stress idrico, indipendentemente dall’e-poca in cui esso veniva applicato; ciò veniva messo inrelazione con la diminuzione del volume delle baccheche si registrava in condizione di stress idrico e, quin-di, ad un effetto concentrazione legato al decrementodella fase solvente nell’acino. Gli stessi autori peròrilevavano che, anche quando si esprimevano i polife-noli come quantità per superficie di buccia (e quindi siescludeva l’effetto concentrazione dovuto alla ridu-zione del volume dell’acino), si registrava comunqueun incremento del quantitativo di polifenoli. Il deficitidrico influenzava anche il contenuto di antociani delmosto e del vino e la formazione del colore risultavapiù rapida durante le prime due settimane dopo l’in-vaiatura, soprattutto qualora le condizioni di stressidrico fossero applicate in epoca anticipata.
Gli effetti dello stress idrico sull’accumulo deimetaboliti secondari della bacca, polifenoli totali e/oantociani, in particolare, furono descritti anche inmolti altri lavori su vitigni quali Cabernet Sauvignon(Bravdo et al., 1985; Dry et al., 2001; Kennedy et al.,2002), Cabernet Franc (Matthews et al., 1990),Moscato di Alessandria (Pedreira dos Santos et al.,2007), Sangiovese (Poni et al., 2007).
Ampia discussione venne intrapresa a propositodel fatto che il deficit idrico potesse alterare diretta-
mente le numerose vie biosintetiche che portanoall’accumulo dei diversi metaboliti secondari o se illoro accumulo fosse prevalentemente una conseguen-za della riduzione della dimensione delle bacche.
Maggiori elementi di chiarezza a proposito dell’in-fluenza dello stress idrico sull’accumulo di alcuneclassi di metaboliti secondari, sulla dimensione degliacini e sulla loro interazione, vennero apportati daRoby e collaboratori (2004). In quell’esperienza, supiante di Cabernet Sauvignon innestate su 110 Richtere sottoposte a condizioni di eccesso di acqua (64l/pianta/settimana), a condizioni di normalità(32l/pianta/settimana) ed a condizioni di stress (som-ministrazione di 32/l/pianta/settimana non prima diaver raggiunto -1,5 MPa di potenziale idrico fogliare amezzogiorno), si rilevò la frequenza delle diverseclassi di dimensioni degli acini e si effettuarono deter-minazioni del contenuto in tannini dei semi ed in tan-nini e antociani delle bucce per ciascuna classe didimensione. Il contenuto in tannini dei semi, cheaumentava con l’aumento delle dimensioni dellabacca, del numero di semi e della massa totale di semiper bacca, risultava inferiore nelle bacche provenientida piante mantenute in condizioni di stress idrico,rispetto alle altre due tesi; questo aspetto venne spie-gato ammettendo l’accelerazione del tasso di degrada-zione dei tannini durante la maturazione. A conclusio-ni simili erano in precedenza pervenuti Ojeda e colla-boratori (2002) nel caso di uve provenienti da piantedi Syrah innestate su Fercal. Anche il contenuto ditannini e di antociani della buccia aumentava con l’au-mentare delle dimensioni della bacca, ma la concen-trazione di antociani diminuiva con l’aumentare delledimensioni del frutto. Analizzando il contenuto di talicomposti in bucce di bacche aventi la stessa dimensio-ne si verificò che sia le loro quantità sia le loro con-centrazioni erano maggiori nelle bacche provenientida piante sottoposte a stress idrico rispetto ai controllio alle piante irrigate con il volume di acqua superiore.
Roby e collaboratori (2004) spiegarono l’incre-mento della concentrazione di antociani e proantocia-nidine della buccia ammettendo che lo stress idricopotesse influire sulla crescita delle cellule della bucciamaggiormente di quanto non accadesse a carico dellecellule della polpa, il che portava all’incremento deiquantitativi dei soluti localizzati nella buccia, oltreche ipotizzare un possibile effetto diretto sulla biosin-tesi dei flavonoidi.
Recenti ricerche su Cabernet Sauvignon innestatosu 140R hanno dimostrato che l’espressione di alcunienzimi legati alla via biosintetica dei fenilpropanoidi,risultava incrementata in condizioni di stress idricosia se esso veniva applicato in epoca anticipata (dal-

Ferrandino et al.
30
l’allegagione alla fine dell’invaiatura, 77 giorni dopola fioritura), sia se esso veniva applicato più tardi dal-l’invaiatura (77 giorni dopo la fioritura) fino alla rac-colta (Castellarin et al., 2007a). Anche il profilo degliantociani risultava influenzato da condizioni di stressidrico in quanto veniva favorito l’accumulo delleantocianine tri-sostituite (delfinina, petunina, malvi-na) rispetto alle di-sostituite (cianina e peonina). Lamaggior concentrazione di antociani per acino erastrettamente legata all’incremento di espressione deglienzimi della biosintesi dei flavonoidi che si traduce inun reale incremento del loro accumulo e non ad uneffetto concentrazione legato alla riduzione del pesodella bacca. Lo stress idrico anticipato era in gradonon solo di incrementare il quantitativo di antocianialla raccolta, ma anche di anticipare l’accumulo delcolore nelle bacche.
I meccanismi che sono stati ipotizzati per giustifi-care l’incremento e l’anticipo dell’accumulo di anto-ciani sono complessi: si è ipotizzato che uno stressidrico prima dell’invaiatura, abbassando il turgore cel-lulare, possa incrementare l’afflusso di zuccheri eABA floematici, aumentando il delta di pressione trasorgenti e sink necessario per attivare i meccanismi dicaricamento e scaricamento del floema. La maggiorconcentrazione di zuccheri a sua volta attiverebbe l’e-spressione dei trasportatori degli zuccheri e i genilegati al metabolismo dei flavonoidi (Castellarin etal., 2007a).
Gli stessi autori su Merlot innestato su SO4 avevanorilevato che, in condizioni di stress idrico, molti genilegati alla biosintesi dei flavonoidi erano sovra-regolati,il che portava ad un significativo incremento del conte-nuto in antociani e ad un effetto sul loro profilo, nonsolo aumentando l’incidenza delle antocianine tri-rispetto alle di-sostituite, come già rilevato dagli stessiautori su Cabernet Sauvignon, ma anche favorendol’accumulo delle forme maggiormente metilate (malvi-na e peonina) rispetto a quelle con un più basso o nullolivello di metilazione (Castellarin et al., 2007b).
L’effetto del deficit idrico specificatamente riferitoa flavonoidi non antociani è stato meno indagato. Unlieve incremento dell’espressione della leucoantocia-nidina-riduttasi (enzima chiave che apre la via dellabiosintesi delle catechine) è indotto da condizioni dideficit idrico anticipato: a questo incremento non èperò corrisposto un significativo aumento della con-centrazione in proantocianidine (Castellarin et al.,2007a) anche se, a livello di tendenza, alcuni gruppidi lavoro su Syrah (Ojeda et al., 2002) hanno rilevatoche sia il deficit idrico anticipato che quello tardivodeterminano un incremento del contenuto in proanto-cianidine rispetto ai controlli. Ancora minori sono
risultati gli effetti dello stress idrico sulla flavonol-sintasi (FLS1) e quindi sull’accumulo dei flavonoli(Castellarin et al., 2007a).
Sono poco delineate, al momento, le conoscenzerelative all’effetto dello stress idrico sull’accumulodegli stilbeni, resveratrolo in particolare, il cui conte-nuto non è risultato influenzato dallo stress idrico inpiante di Barbera innestate su 3309 C (Vezzulli et al.,2007).
Composti aromatici. Attualmente i lavori relativiall’effetto dello stress idrico sulla quantità di compo-sti aromatici e sul loro profilo non sono ancora moltonumerosi. Qualche conoscenza in più si ha a proposi-to dell’effetto dello stress idrico sui carotenoidi per iquali la relativamente recente identificazione e carat-terizzazione di una specifica carotenoide diossigenasidell’uva, la cui attività è stata correlata in vivo conl’accumulo di 3-idrossi-ß-ionone, isoprenoide C-13,dal significativo impatto aromatico, ha definitivamen-te chiarito, anche in Vitis vinifera, l’origine dei C-13norisoprenoidi da precursori carotenoidi (Mathieu etal., 2005).
La mancanza di irrigazione su piante di Touriganacional innestate su 196-17 causava un incrementosignificativo del contenuto in carotenoidi rispetto altrattamento irriguo, in condizioni di suolo con scarsacapacità di trattenuta idrica, mentre nel suolo concapacità di trattenuta superiore le differenze tra condi-zioni irrigue e condizioni non irrigue risultavano sfu-mate, pur essendo ancora evidenti (Oliveira et al.,2003). L’influenza dello stato idrico è stata valutatasu piante di Cabernet Sauvignon sottoposte a partialroot drying (PRD) e su piante testimoni normalmenteirrigate su entrambi i lati del filare: l’influenza dellariduzione dell’apporto idrico incomincia a verificarsia partire da circa 10 e circa 30 giorni dall’invaiaturaper la luteina e per il ß-carotene, rispettivamente.Anche la concentrazione dei C-13 norisoprenoidi, inparticolare quella di ß-damascenone, ß-ionone e1,1,6-trimetil-1,2-diidronaftalene (TDN), era signifi-cativamente maggiore nelle piante sottoposte a condi-zioni di PRD; ciò si verificava anche su base di conte-nuto per acino e quindi parrebbe un effetto che, simil-mente a quanto già chiarito per i polifenoli, non è soloil risultato di un alterato rapporto buccia/polpa ma,presumibilmente, è legato ad un incremento dell’atti-vità enzimatica (Bindon et al., 2007). Una limitatadisponibilità idrica incrementava il contenuto degliaromi glico-coniugati nei vini prodotti a partire dauve della cv Agiorgitiko innestata su 41B, in partico-lare nelle condizioni di minor disponibilità idrica sideterminarono le maggiori concentrazioni di terpenil-

Meccanismi di resistenza a stress idrico della vite
31
glicosilati (Koundouras et al., 2006). Un comporta-mento simile è stato osservato su un altro vitigno, ilMoscato di Alessandria su 1103 Paulsen (Pedreira dosSantos et al., 2007), caso in cui, in condizioni dimoderata disponibilità idrica a seguito dell’applica-zione di condizioni di PRD e di non-irrigazione, sirilevò un incremento della concentrazione del glico-sil-glucosio (GG) che, nelle uve a bacca bianca, èessenzialmente ascrivibile alla presenza di metabolitisecondari glicosilati aromatici (Williams et al., 1995).L’effetto della ridotta disponibilità idrica è stato ancheosservato a carico di alcuni aromi tiolici, derivatidall’S-cisteina, in Sauvignon blanc: Peyrot desGachons e collaboratori (2005) hanno dimostrato checondizioni di ridotta disponibilità idrica accompagna-te da una non limitata dotazione azotata favoriscono ilmassimo accumulo del 4MMP (4-mercapto-4-metil-pentano-2-one), del 4MMPOH (4-mercapto-4-metil-pentan-2-olo) e del 3MH (3-mercaptoesan-1-olo).
È stato riportato che lo stress idrico ha favorito siaincrementi sia decrementi dell’accumulo dell’mRNA.Alcuni enzimi coinvolti nella sintesi dell’isopentenil-pirofosfato attraverso la via metabolica non mevalo-nato-dipendente, hanno mostrato una significativariduzione in condizioni di stress idrico; tra questi la 1-deossi-D-xylulosio 5-fosfato sintasi e la (E)-ßocimenesintasi che catalizzano la sintesi di olefina monoterpe-nica, un composto con importanti implicazioni dalpunto di vista del profumo e la zeaxantina epossidasiche porta alla formazione di xantofille. La β-caroteneidrossilasi, responsabile dell’idrossilazione del carote-ne e che è necessaria per la sintesi delle xantofille,risultava significativamente indotta nella polpa aseguito di stress idrico. Anche 5 metil-transferasi del-l’acido caffeico, coinvolte, almeno alcune, nella for-mazione di sostanze aromatiche mostravano incre-mento di mRNA nella buccia e nella polpa di bacchein stress idrico (Grimplet et al., 2007).
In tabella 1 (a: polifenoli; b: composti aromatici)sono elencati in forma riassuntiva i risultati ottenutiper i metaboliti descritti.
Risposte molecolari allo stress idrico
In questi ultimi anni l’uso della genomica funzio-nale ha permesso di comprendere le risposte moleco-lari agli stress in maniera più completa rispetto allealtre tecniche molecolari tradizionali, osservando lacontemporanea variazione di espressione, e l’eventua-le correlazione, di migliaia di trascritti o centinaia diproteine, e non solo di pochi geni d’interesse. Oggisono ormai disponibili chip che contengono l’intero
genoma di una pianta; in un solo esperimento si pos-sono quindi ricavare migliaia di informazioni.
Esiste una quantità di studi molecolari di genomicafunzionale condotti su piante sottoposte a condizionidi deficit idrico, molti dei quali hanno sfruttato la tec-nologia microarray come strumento per l’analisi tra-scrittomica, quali quelli di Seki e collaboratori in ara-bidopsis (2002) o di Rabbani e collaboratori in riso(2003). Nel 2007 Cramer et al. si sono serviti di que-sta potente tecnologia informativa per indagare sullerisposte molecolari precoci e tardive della vite (cvCabernet Sauvignon) a stress di tipo idrico e salino.L’elegante disegno sperimentale utilizzato ha permes-so di separare le risposte dovute agli effetti dei duetipi di stress. Durante l’esperimento, le piante sottopo-ste a stress idrico non sono state irrigate, mentre quel-le sottoposte a stress salino sono state irrigate con unasoluzione di sali tale da portarle ad avere valori dipotenziale idrico del fusto simili alle piante trattatecon deficit di acqua. Al sedicesimo giorno di tratta-mento, tutte le piante stressate mostravano un valorebasso di potenziale idrico del fusto rispetto al control-lo normalmente irrigato, ma solo le piante in stressidrico apparivano avvizzite. Il lavoro molecolare e dicaratterizzazione dei metaboliti eseguito in paralleloha permesso di dare delle risposte a questo diversocomportamento e permette di sottolineare quanto siaimportante una corretta integrazione di studi di tipofisiologico, molecolare e metabolico per avere unavisione globale del processo studiato. Entrambi i tipidi stress impongono cambiamenti osmotici nella pian-ta, ma quello salino, oltre ad un abbassamento delpotenziale idrico, provoca un accumulo di ioni nellazona radicale. La compartimentalizzazione di questiioni nel vacuolo delle cellule appare essere un vantag-gio per le piante, che dispongono in questo modo diosmoliti “a basso costo”, ossia che non devono sinte-tizzare, per bilanciare gli equilibri osmotici. Al con-trario le viti in stress idrico analizzate possiedono unamaggior concentrazione di prolina, glucosio e acidomalico che devono sintetizzare con dispendio di ener-gia (Raven, 1985). È interessante notare come l’acidomalico, secondo più abbondante acido organico invite, mostra un notevole incremento in risposta allostress idrico rispetto a quello salino (rispettivamente473 e 182 % rispetto al controllo).
A livello del profilo trascrittomico, questo studiosembra dimostrare come in vite la risposta allo stressidrico sia più immediata rispetto a quella allo stresssalino, anche se per rilevare cambiamenti nell’espres-sione genica piuttosto importanti bisogna attenderealmeno dieci giorni in entrambi i casi. Nei primi diecigiorni di stress, i meccanismi di elusione, di controllo
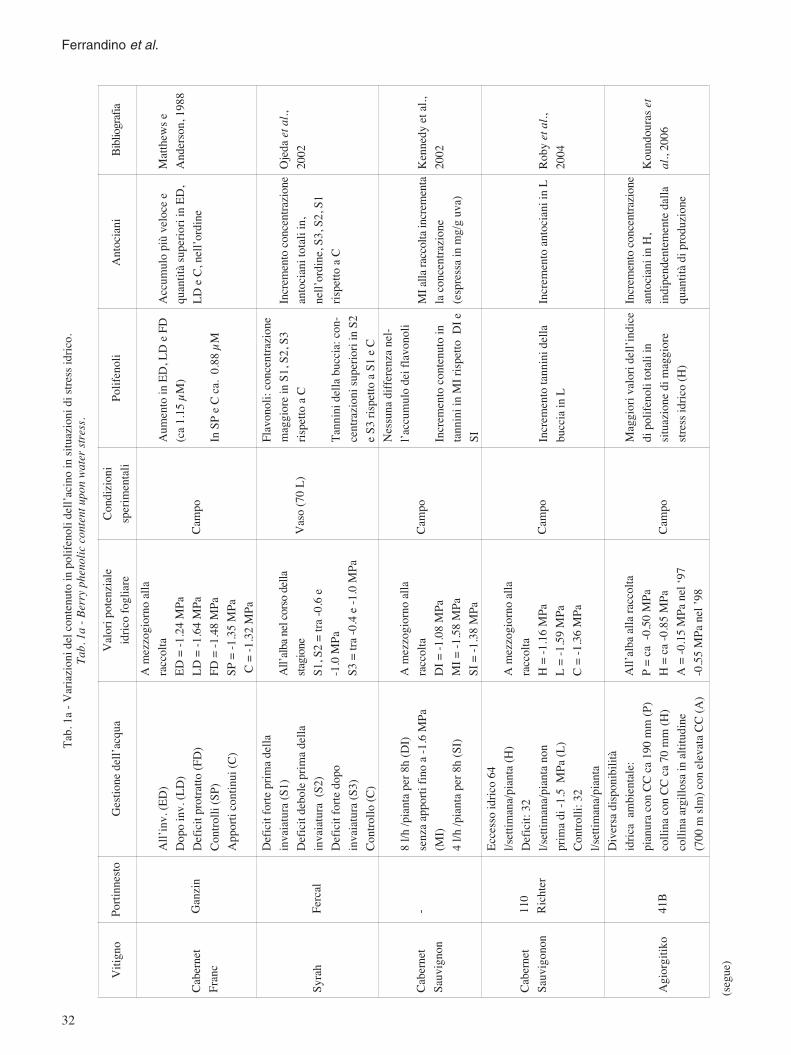
Ferrandino et al.
32
Vitig
no
Cabe
rnet
Fran
c
Syrah
Cabe
rnet
Sauv
ignon
Cabe
rnet
Sauv
igono
n
Agior
gitiko
Ganz
in
Ferca
l
- 110
Rich
ter
41B
All’i
nv.(E
D)Do
poinv
.(LD)
Defic
itprot
ratto
(FD)
Contr
olli(
SP)
Appo
rtico
ntinu
i(C)
Defic
itfort
eprim
adell
ainv
aiatur
a(S1
)De
ficitd
ebole
prima
della
invaia
tura
(S2)
Defic
itfort
edop
oinv
aiatur
a(S3
)Co
ntroll
o(C)
8l/h
/pian
tape
r8h(
DI)
senza
appo
rtifin
oa-1.
6MPa
(MI)
4l/h
/pian
tape
r8h(
SI)
Ecce
ssoidr
ico64
l/sett
iman
a/pian
ta(H
)De
ficit:
32l/s
ettim
ana/p
ianta
non
prima
di-1.
5M
Pa(L
)Co
ntroll
i:32
l/sett
iman
a/pian
taDi
versa
dispo
nibilit
àidr
icaam
bienta
le:pia
nura
conC
Cca
190m
m(P
)co
llinac
onCC
ca70
mm(H
)co
llinaa
rgillo
sain
altitu
dine
(700m
slm)c
onele
vata
CC(A
)
Porti
nnest
oGe
stion
edell
’acqu
a
Ame
zzog
iorno
alla
racco
ltaED
=-1.2
4MPa
LD=-
1.64M
PaFD
=-1.4
8MPa
SP=-
1.35M
PaC
=-1.3
2MPa
All’a
lbane
lcors
odell
asta
gione
S1,S
2=tra
-0.6e
-1.0M
PaS3
=tra
-0.4e
-1.0M
Pa
Ame
zzog
iorno
alla
racco
ltaDI
=-1.0
8MPa
MI=
-1.58
MPa
SI=-
1.38M
Pa
Ame
zzog
iorno
alla
racco
ltaH
=-1.1
6MPa
L=-
1.59M
PaC
=-1.3
6MPa
All’a
lbaall
arac
colta
P=ca
-0.50
MPa
H=c
a-0.8
5MPa
A=-
0.15M
Pane
l‘97
-0.55
MPa
nel’
98
Accu
mulo
piùve
locee
quan
titàsu
perio
riin
ED,
LDeC
,nell
’ordin
e
Increm
ento
conc
entra
zione
antoc
ianito
taliin
,ne
ll’ord
ine,S
3,S2
,S1
rispe
ttoaC
MIa
llarac
colta
increm
enta
laco
ncen
trazio
ne(es
pressa
inmg
/guv
a)
Increm
ento
antoc
ianiin
L
Increm
ento
conc
entra
zione
antoc
ianiin
H,ind
ipend
entem
ente
dalla
quan
titàdi
produ
zione
Matt
hews
eAn
derso
n,19
88
Ojed
aeta
l.,20
02
Kenn
edye
tal.,
2002
Roby
etal.
,20
04
Koun
doura
set
al .,2
006
Aume
ntoin
ED,L
DeF
D(ca
1.15µ
M)
InSP
eCca
.0.88
µM
Flavo
noli:
conc
entra
zione
magg
iorei
nS1,
S2,S
3ris
petto
aC
Tann
inide
llabu
ccia:
con-
centr
azion
isup
eriori
inS2
eS3r
ispett
oaS1
eCNe
ssuna
differ
enza
nel-
l’acc
umulo
deif
lavon
oli
Increm
ento
conte
nuto
intan
niniin
MIr
ispett
oDI
eSI Inc
remen
totan
ninid
ella
bucc
iain
L
Mag
giori
valor
idell
’indic
edi
polif
enoli
totali
insit
uazio
nedi
magg
iore
stress
idrico
(H)
Camp
o
Vaso
(70L)
Camp
o
Camp
o
Camp
o
Valor
ipote
nzial
eidr
icofog
liare
Cond
izion
isp
erime
ntali
Polif
enoli
Antoc
iani
Bibli
ograf
ia
Tab.
1a-V
ariaz
ionid
elco
ntenu
toin
polif
enoli
dell’
acino
insit
uazio
nidi
stress
idrico
.Ta
b.1a
-Ber
ryph
enoli
ccon
tentu
ponw
aters
tress.
(segu
e)

Meccanismi di resistenza a stress idrico della vite
33
Vitig
no
Cabe
rnet
Sauv
ignon
Merl
ot
Mos
cato
diAl
essan
dria
Sang
iovese
(clon
e12T
)
140R
SO4
1103
P
SO4
Defic
itdaa
llega
gione
ainv
aiatur
a(ED
)De
ficitd
allaf
inede
ll’inv
aiatur
a(L
D)Co
ntroll
o(C)
2.5L/
m/hp
erma
ntene
reil
poten
ziale
tra-0.
2e-0.
6MPa
neic
ontro
lli
Irriga
zioni
diso
ccors
oper
evita
repe
rdita
dipro
duzio
nene
llepia
nteW
S
Noni
rriga
to(N
I)50
%ev
apotr
aspira
zione
reale
(ETc
),(DI
)50
%ET
c,su
latia
lterni
(PRD
)10
0%ET
c(FI
)ca
5L/va
so/gi
ornop
errip
ortare
allac
apac
itàdi
camp
o(W
W)
ca5L
/vaso
/giorn
osolo
in1d
ei2v
asialt
ernati
vame
nte(P
RD)
Porti
nnest
oGe
stion
edell
’acqu
a
Ame
zzog
iorno
alla
racco
ltaED
=fino
a-1.4
MP
ante
invaia
turap
oisim
ileaC
;LD
=sim
ileaC
ante
invaia
turap
oifin
oa-1.
4MPa
;C
=tra
-0.9e
-1.2M
PaPo
tenzia
lede
ltralc
ioPia
ntein
stress
(WS)
=fino
a-1.4
MPa
,nel
2004
;=f
inoa-
1.20M
Pane
l20
05ma
pert
empi
piùpro
lunga
tiinp
re-inv
aiatur
aco
ntroll
o(CT
)=tra
-0.2e
-0.6
MPa
inen
tramb
igli
anni
All’a
lbain
pre-ra
ccolt
aNI
=-0.6
4MPa
DI=-
0.45M
PaPR
D=-
0.40M
PaFI
=-0.2
2MPa
All’a
lbaall
arac
colta
WW
=-0.3
0MPa
PRD
=-0.5
5MPa
Increm
ento
conc
entra
zione
econ
tenuto
antoc
ianiin
defic
it
increm
ento
della
perce
ntuale
dian
tocian
inetri
-idros
silate
inED
Increm
ento
antoc
iani
totali
inW
S+37
%ne
l20
04,+
57%
nel2
005
Increm
ento
%de
llean
tocian
inetri
-sosti
tuite
inW
SInc
remen
tode
llive
llodi
mutila
zione
delle
antoc
ianine
inW
S
- Increm
ento
antoc
ianito
tali
(suba
sepe
sofre
sco)
Caste
llarin
etal.
,20
07a
Caste
llarin
etal.
,20
07b.
Pedre
irado
sSa
ntos e
tal.,
2007
Poni
etal.
,200
7.
Effet
tolim
itato
sull’
accu
mulo
delle
proan
tocian
idine
effett
onon
signif
icativ
osu
ll’ac
cumu
lode
iflav
onoli
- Aume
ntoin
NIeP
RD,
noni
mputa
bile,
almen
oin
PRD,
allar
iduzio
nede
llaqu
antità
dipro
duzio
ne
-
Camp
o
Camp
o
Camp
o
Vaso
(40L)
con
appa
rator
adica
lediv
isoin
due
conte
nitori
Valor
ipote
nzial
eidr
icofog
liare
Cond
izion
isp
erime
ntali
Polif
enoli
Antoc
iani
Bibli
ograf
ia

Ferrandino et al.
34
Vitig
no
Touri
gaNa
ciona
l
Sauv
ignon
Blan
c
Agior
gitiko
Cabe
rnet
Sauv
ignon
Mos
cato
diAl
essan
dria
196-1
7
Dive
rsipo
rtinn
esti
asec
onda
delv
igneto
41B
- 1103
P
Dive
rsadis
ponib
ilitài
drica
ambie
ntale:
suolo
A=c
apac
itàvo
lumetr
icaalt
a(irr
igato
=AI;
noni
rriga
to=
ANI)
suolo
B=c
apac
itàvo
lumetr
icaba
ssa(ir
rigato
=BI;
noni
rriga
to=B
NI)
Dive
rsadis
ponib
ilitài
drica
ambie
ntale:
capa
citàv
olume
trica
di67
mmpe
r1.0
mdi
profon
dità(
SG)
capa
citàv
olume
trica
di39
mmpe
r0.8
mdi
profon
dità(
GS)
capa
citàv
olume
trica
di93
mmpe
r1.4
mdi
profon
dità(
LSB)
Nessu
nalim
itazio
neidr
ica(L
HB)
Dive
rsadis
ponib
ilitài
drica
ambie
ntale:
pianu
raco
ncap
acità
volum
etrica
di19
0mm
(P)
collin
acon
capa
citàv
olume
trica
di70
mm(H
)co
llinaa
rgillo
sain
altitu
dine
(700m
slm)c
onele
vata
capa
cità
volum
etrica
(A)
1.19M
L/ha
nel2
001-0
2e1.3
7M
L/ha
nel2
002-0
3(C)
0.84M
L/ha
nel2
001-0
2e0.8
5M
L/ha
nel2
002-0
3(PR
D)
Noni
rriga
to(N
I)50
%ev
apotr
aspira
zione
reale
(ETc
),(DI
)50
%ET
c,su
latia
lterni
(PRD
)10
0%ET
c(FI
)
Porti
nnest
oGe
stion
edell
’acqu
a
- All’a
lba:
piante
noni
nstre
ssin
LSB
eLHB
;ap
artire
dalg
iorno
giulia
no23
0:-0.
5MPa
inGS
,-1.0
MPa
inSG
All’a
lbaall
arac
colta
P=ca
-0.50
MPa
H=c
a-0.8
5MPa
A=-
0.15M
Pane
l‘97
-0.55
MPa
nel’
98
Varie
misu
redu
rante
icicl
idi
irriga
zione
ampia
mente
discu
ssean
chei
nfun
zione
della
cond
uttan
zasto
matic
a
All’a
lbain
pre-ra
ccolt
aNI
=-0.6
4MPa
DI=-
0.45M
PaPR
D=-
0.40M
PaFI
=-0.2
2MPa
- - Conc
entra
zioni
magg
iori
dico
mpost
iC6(
es.eta
nolo,
eseno
lo),d
ifen
olivo
latili,
diter
peni
nelle
cond
izion
idi
magg
iores
tress
idrico
(H);d
iC13
-noris
opren
oidi
inun
anno
sudu
e
Aume
ntain
modo
netto
ilco
ntenu
tope
rbac
cadi
alcun
iC13
-noris
opren
oidi
( β-da
masce
none
,β-io
none
eTDN
)inc
ondiz
ionid
iPR
DIl
conte
nuto
digli
cosil
-glu
cosio
perb
acca
aume
ntain
nelle
cond
izion
idi
magg
iorstr
ess(N
IePR
D)
Olive
iraet
al.,
2003
Peyro
tdes
Gach
onet
al.,
2005
Koun
doura
s et
al .,2
006
Bind
onet
al.,
2007
Pedre
irado
sSa
ntos e
tal.,
2007
Ilco
ntenu
toin
carot
enoid
iinc
remen
tane
lleco
ndizi
oni
distr
essidr
ico,ta
ntopiù
quan
tomi
nore
èlac
apac
itàvo
lumetr
icade
lsuo
lo(in
Bpiù
chei
nA)
Iprec
ursori
aroma
tici
coniu
gatid
ell’S
-ciste
inaau
menta
noin
cond
izion
idi
mode
ratos
tress
idrico
Stress
idrici
severi
sembra
nolim
itare
illoro
accu
mulo
- Ilco
ntenu
toin
carot
enoid
iau
menta
inPR
Dris
petto
aC,
piùla
lutein
adel
β-ca
roten
e
-
Camp
o
Camp
o
Camp
o
Camp
o
Camp
o
Valor
ipote
nzial
eidr
icofog
liare
Cond
izion
isp
erime
ntali
Prec
ursori
d’arom
aAr
omi(
liberi
oinf
orma
glico
-coniu
gata)
Bibli
ograf
ia
Tab.
1b-V
ariaz
ionid
elco
ntenu
toin
aromi
eloro
precu
rsori
nell’
acino
insit
uazio
nidi
stress
idrico
.Ta
b.1b
-Ber
ryfla
vour
sand
flavo
urpr
ecur
sors
asaff
ected
bywa
terstr
ess.

Meccanismi di resistenza a stress idrico della vite
35
idraulico e stomatico hanno probabilmente ritardato lamaggior parte delle risposte metaboliche della pianta.Infatti i trascritti che hanno mostrato una variazione diespressione rispetto al controllo sono aumentati conl’intensificarsi degli stress e molti di tali trascrittivariano in entrambi i tipi di stress.
Questa ricerca ha rivelato inoltre come l’accresci-mento vegetativo della vite sembri più sensibile allostress idrico che ad un equivalente livello di stresssalino. Questo rafforza la tesi che l’assorbimento delsale possa contribuire al riequilibrio osmotico, facili-tando il richiamo di acqua e la crescita nei giovanigermogli. È interessante notare come, nel caso distress prolungati nel tempo, la vite mostri invece mag-gior adattabilità allo stress idrico e sia da sempre con-siderata una specie sensibile alla salinità (Hawker eWalker, 1978; Shani et al., 1993). Oggi possiamofinalmente comprendere come questa intolleranza siaprobabilmente dovuta all’incapacità della pianta ditener testa all’alta concentrazione interna di ioni e allaconseguente tossicità nel lungo periodo (Munns,2002).
Se nel lavoro appena descritto Cramer e collabora-tori indagano sulle risposte allo stress idrico a livellodella pianta intera, un altro studio, eseguito sempre suCabernet Sauvignon, con un approccio di tipo trascrit-tomico, servendosi della tecnologia microarray, spostal’attenzione direttamente sull’acino (Grimplet et al.,2007). Tale ricerca, condotta su diversi tessuti dell’a-cino e su acini raccolti da piante normalmente irrigateo poste in condizioni di stress idrico, mette in lucecome questo tipo di stress abbia profondi effetti sulprofilo di espressione di geni soprattutto a livello dipolpa e buccia. In questi tessuti il deficit idrico provo-ca l’aumento del livello di trascritti di diverse catego-rie funzionali, tra cui quelle associate alla biosintesidei fenilpropanoidi, della lignina, delle proantociani-dine così come dei metaboliti responsabili dell’aroma.Inoltre, come già discusso precedentemente, piante divite (cv Cabernet Sauvignon e cv Merlot) cresciute incondizioni di stress idrico mostrano un incremento diespressione dei geni coinvolti nelle vie di biosintesidei flavonoidi nell’acino (CHS, F3H, DFR, LDOX,F3’5’H, UFGT, GST, OMT) (Castellarin et al.,2007a, 2007b), con le importanti ricadute sulla qualitàdell’uva di cui si è detto.
Altrettanto interessanti sono i risultati di una ricer-ca che ha sfruttato un altro strumento della genomicafunzionale, l’analisi proteomica, già utilizzata perindividuare le variazioni enzimatiche nell’acinodurante la maturazione (Giribaldi et al., 2007). InChardonnay e Cabernet Sauvignon sono state indaga-te le differenze proteomiche di risposta allo stress
idrico e salino (Vincent et al., 2007). Le pratiche dimiglioramento genetico cercano da sempre di sfrutta-re variazioni genetiche tra cultivar per ottenere unavarietà d’élite più performante. Nel caso della vitesono invece proprio le differenze tra le diverse culti-var a guidare le preferenze del consumatore per unparticolare vino o una particolare uva da tavola; talidifferenze vanno quindi mantenute. Lo studio effet-tuato dimostra come, a livello fisiologico,Chardonnay sembra essere più tollerante di CabernetSauvignon sia allo stress idrico che a quello salino.Fra le proteine maggiormente attivate dagli stress inquest’ultima cultivar si ritrovano proteine coinvoltenel macchinario di proteolisi delle cellule, risultatoche sembra indicare una maggiore sofferenza fisiolo-gica agli stress. Parallelamente, in Chardonnay risul-tano attivati enzimi, quali xiloglucano-endotransglico-silasi, che modificano la parete nella fase di crescitacellulare (Wu et al., 2005). La sovra-espressione diquesti enzimi potrebbe essere correlata al migliorcomportamento della cv Chardonnay per quel cheriguarda l’accrescimento del fusto sotto stress.Risultato particolarmente interessante, l’analisi dellavarianza indica che è la differenza varietale la princi-pale componente che influisce sulla variazione dell’e-spressione proteica (40% degli spot mostra variazionesignificativa in risposta alla cv contro il 15% chevaria in risposta agli stress).
Nei paragrafi successivi riepiloghiamo, divise invarie categorie funzionali, quelle che sono le principa-li risposte molecolari della vite allo stress idrico cheemergono dagli studi presenti in letteratura.
Effetti sul metabolismo dei carboidratiL’aumentata produzione di soluti cellulari, princi-
palmente osmoliti protettori come zuccheri, ammine ezuccheri alcolici, è una delle prime risposte di difesaalla disidratazione e mira a mantenere inalterato ilpotenziale idrico cellulare e a limitare il più possibilela perdita di acqua (Bray, 1997). È risaputo inoltreche l’espressione di enzimi coinvolti nella degrada-zione di carboidrati complessi a zuccheri sempliciaumenta in risposta a stress idrico, salino e freddo(Taji et al., 2002). In giovani apici vegetativi diCabernet Sauvignon, quali quelli usati nel lavoro diCramer e collaboratori già citato, l’aumento dei tra-scritti coinvolti nella glicolisi e nel metabolismo dicomposti osmoprotettori è una delle prime risposteallo stress idrico. Diversamente, in foglie mature diviti (cv Savatiano) sottoposte a stress idrico, si èosservato come il riequilibrio osmotico è largamenteassociato all’aumento del contenuto di ioni inorganicie non di carboidrati solubili o di aminoacidi (Patakas

Ferrandino et al.
36
et al., 2002). Questa diversa risposta sembra essereassociata all’età dell’organo vegetativo preso in consi-derazione. Patakas (2000) ha infatti dimostrato come,in normali condizioni, i carboidrati solubili rappresen-tino circa il 70% dei soluti osmoticamente attivi ingiovani foglie di vite cv Savatiano. Nelle foglie matu-re, durante lo stress idrico, la concentrazione di talicarboidrati è ridotta forse perchè trasportati nei tessutisink più giovani (Cramer et al., 2007); per questomotivo le foglie più mature si servirebbero degli ioniinorganici per gestire il riequilibrio osmotico.
A livello dell’acino di Cabernet Sauvignon si èriscontrata una sottoregolazione dell’espressione ditrasportatori di zucchero seme- e polpa-specifici e diuna invertasi vacuolare polpa-specifica. Questopotrebbe suggerire un’ipotesi alternativa al carica-mento di esosi nell’acino durante lo stress idrico,ossia che l’aumento dei gradi Brix possa essere dovu-to ad un rimpicciolimento dell’acino (Grimplet et al.,2007).
Effetti sul trasporto di soluti e ioniI trasportatori, presenti soprattutto sul plasmalem-
ma e sul tonoplasto, hanno lo specifico compito diripristinare la pressione osmotica in condizioni distress imposte dall’ambiente esterno (Seki et al.,2002). In Cabernet Sauvignon lo stress idrico, comequello salino, provoca un incremento dei trascritti checodificano per una varietà di trasportatori di ioni, ami-noacidi, nucleotidi e peptidi (Cramer et al., 2007). Siritrova anche una pronunciata e precoce espressionedi un trasportatore di lipidi. Trasportatori di questotipo pare abbiano un ruolo nella protezione dellemembrane cellulari dal danno osmotico e sembra pos-sano mediare eventuali cambiamenti nella composi-zione dei fosfolipidi (Holmberg e Bülow, 1998). Unproblema che le piante in stress idrico si trovano adaffrontare è la riduzione dell’attività fotosintetica chepuò portare a problemi di fotoinibizione in condizionidi luce normali. Per tali motivi, l’organismo tenta direagire attivando la fotorespirazione e provvedendo asmaltire diversamente gli elettroni in eccesso; si spie-ga così l’incremento nei trascritti relativi ai geni coin-volti nella catena di trasporto di elettroni del fotosiste-ma I e II (Cramer et al., 2007).
In merito alla problematica dello stress idrico, leacquaporine, proteine canale finemente regolate chefacilitano il trasporto dell’acqua, meritano particolareattenzione. Alcune acquaporine sono espresse costitu-tivamente mentre l’espressione di altre è regolata dadifferenti stimoli quali lo stadio di sviluppo, la luce,gli ormoni o da avverse condizioni ambientali qualideficit idrico, stress salino, basse temperature (Vera-
Estrella et al., 2004). Studi di espressione genica invarie specie vegetali mostrano una risposta variabiledelle diverse isoforme di acquaporina allo stress idri-co, con sotto- o sovra-regolazione dei geni (Yamadaet al., 1997; Alexandersson et al., 2005; Secchi et al.,2007). Probabilmente per mantenere un corretto statoidrico in condizioni di stress abiotico, sono necessarisia l’incremento del trasporto dell’acqua in certi tes-suti che la riduzione in altri (Jang et al., 2004). I risul-tati delle analisi trascrittomiche sopra descritte metto-no in evidenza come diverse acquaporine risultinosotto-regolate sia in apice vegetativo che nella polpa enella buccia di acini di Cabernet Sauvignon in stressidrico. Un altro lavoro prende invece in considerazio-ne uno studio di espressione di sette geni di acquapo-rine in foglie e radici dell’ibrido Richter-110 (Vitisberlandieri x Vitis rupestris), portinnesto che mostraparticolare vigore e resistenza allo stress idrico(Galmés et al., 2007). In foglia tutti i geni consideratimostrano una significativa sottoregolazione in condi-zioni di stress moderato; al contrario nelle piante for-temente stressate l’espressione risulta uguale o piùalta rispetto al controllo, a seconda del particolaregene. Nelle radici non si osserva un trend generalenella variazione dei trascritti: PIP1;2, PIP2;1 e TIP2risultano sovra-espressi in condizioni di stress mode-rato, ma in una settimana la loro espressione ritorna ailivelli del controllo. Solo l’espressione di una PIP2;2risulta prolungata nel tempo.
Una recente ricerca di Vandeleur et al. (2008)mette in relazione la diversa espressione di dueacquaporine (VvPIP2;2 e VvPIP1;1) fortementeespresse in radice con la diversa strategia di difesa didue varietà di vite, Grenache e Chardonnay, nei con-fronti dello stress idrico. In generale le acquaporineradicali influiscono sulla via transcellulare e simpla-stica del flusso radiale dell’acqua in radice e, con laloro espressione regolata da vari fattori, condizionanola conduttanza idraulica di quest’organo. I risultati diquesto studio mostrano come VvPIP2;2 sia caratteriz-zata da un’espressione costitutiva in radice, inentrambe le cultivar ed anche in stress idrico, mentreil livello del trascritto VvPIP1;1 aumenta durante lostress idrico in Chardonnay ma non in Grenache.Questo si traduce in un contenimento della riduzionedi conduttanza idraulica radicale in Chardonnay incondizioni di stress idrico, a causa dell’aumento delcontributo transcellulare al trasporto radiale d’acquain radice. Questo indicherebbe perché tale cultivar siameno vulnerabile all’embolizzazione di Grenache(Alsina et al., 2007), riuscendo a mantenere differen-ze di potenziale più piccole fra suolo e xilema. Alcontrario, la strategia adottata dalla cv Grenache sem-

Meccanismi di resistenza a stress idrico della vite
37
bra essere opposta; la mancata variazione nei livellidei trascritti delle due più importanti acquaporineradicali si traduce in una conduttanza idraulica(soprattutto nella sua componente intracellulare) piùbassa in condizioni di deficit idrico e quindi in unamaggiore tolleranza alle condizioni di stress. Questastrategia è possibile solo grazie allo stretto controllodella regolazione stomatica che questa varietà possie-de, riuscendo così a prevenire potenziali xilematicieccessivamente negativi (Schultz, 2003; Soar et al.,2006). Grenache sembra quindi avere un approccioallo stress idrico simile a quello delle piante del deser-to, dove le acquaporine della radice vengono sotto-regolate per prevenire la perdita di acqua nel suolo instress idrico (North et al., 2004).
Effetti sulla risposta agli stimoli ormonaliL’acido abscissico (ABA) ricopre il ruolo di
importante molecola segnale nei meccanismi di rispo-sta a svariate condizioni di stress, come quelle provo-cate da deficit idrico (Wilkinson e Davies, 2002). Inapici vegetativi di piante di Cabernet Sauvignon sotto-poste a stress idrico, un enzima chiave regolatoredella biosintesi dell’ acido abscissico (ABA), quale il9-cis-epossicarotenoide diossigenasi (NCED), aumen-ta molto il suo livello di espressione rispetto al con-trollo, così come risulta incrementata l’espressione di25 diversi trascritti correlati a quest’ormone (Crameret al., 2007). Sappiamo che in foglie e radici l’ABAricopre il ruolo di importante molecola segnale neimeccanismi di risposta a svariate condizioni di stress,come quelle provocate da deficit idrico (Stoll et al.,2000; Wilkinson e Davies, 2002). Tale ruolo potrebbeessere invece trascurabile nell’acino. In Merlot si èvisto infatti come, a livello di trascritto, l’espressionedi quattro geni correlati al metabolismo dell’ABA(NCED1, NCED2, rd22 e ACPK1) non risultino parti-colarmente indotti nell’esocarpo di acini in stress idri-co (Castellarin et al., 2007b) mentre in acini diCabernet Sauvignon risulta incrementata l’espressionedi un gene relativo all’enzima NCED ma sotto-regola-ti geni rd22 che rispondono all’ABA e alla deidrata-zione (Grimplet et al., 2007).
È ben noto invece come l’ABA rappresenti unimportante segnale dell’inizio della maturazione degliacini, poiché la vite mostra un aumento della concen-trazione di quest’ormone nelle bucce delle baccheappena prima dell’invaiatura (Coombe e Hale, 1973;Davies et al., 1997). Evidenze sull’importanzadell’ABA in questo processo sono state ottenute stu-diando gli effetti di applicazioni esogene di ormonesugli acini. Questi trattamenti favoriscono l’accumulodegli antociani e l’espressione dei geni che presiedono
alla loro via biosintetica (Ban et al., 2003; Jeong etal., 2004). Come già discusso nei paragrafi preceden-ti, lo stress idrico porta ad un incremento di espressio-ne dei geni coinvolti nelle vie di biosintesi di antocia-ni e flavonoidi ma, alla luce di quanto detto e di quan-to discusso soprattutto da Castellarin e collaboratori,non sembra esserci un rapporto causale tra stress idri-co a livello di pianta intera, livelli di ABA nell’eso-carpo dell’acino e regolazione trascrizionale degliantociani. Lo stress idrico potrebbe servirsi di vie disegnalazione alternative a quella dell’ABA o di modi-ficazioni post-trascrizionali o di attività delle proteineper indurre l’aumento della biosintesi di antociani(Castellarin et al., 2007b).
L’etilene sembra giocare un ruolo critico durantelo stress idrico, anche sullo sviluppo e la maturazionedell’acino. Enzimi quali S-adenosil-L-metionina sin-tetasi 1 (SAM) o 1-aminociclopropano-1-carbossilato(ACC) ossidasi vengono sovra-espressi sia nell’acinoche nel tessuto vegetativo in seguito a questo tipo distress (Cramer et al., 2007; Grimplet et al., 2007).
Effetti sul metabolismo e sulle proprietà chimico-fisi-che della parete cellulare
Anche piccoli cambiamenti nello stato idrico dellapianta possono avere effetti rapidi e significativi sullacrescita (Passioura e Munns, 2000). L’inibizione del-l’espansione cellulare può risultare innanzitutto da unridotto turgore cellulare. Infatti la crescita per disten-sione dipende innanzitutto da fattori chimico-fisici,ossia dall’equilibrio tra la pressione di turgore e laforza meccanica della parete che si oppone a cambia-menti di forma. Si è prima discusso come il rallenta-mento della crescita sia meno evidente in viti sottopo-ste a stress salino che riescono a gestire meglio lapressione di turgore, avendo a disposizione osmoliti“a basso costo”. Per sostenere la crescita è però neces-sario un processo metabolicamente attivo che control-li l’estensibilità della parete cellulare. Tale processorichiede un’acidificazione di parete dovuta allaaumentata attività delle pompe protoniche ATPasichelocalizzate sulla membrana plasmatica, mediata dal-l’auxina. Il basso valore di pH rilassa i legami chetengono insieme i componenti di parete e nello stessotempo attiva gli enzimi che portano alla sintesi dinuovo materiale. In piante in stress idrico, la riduzio-ne dell’allungamento fogliare è correlata all’aumentodi pH del succo xilematico, aumento che causa ancheun incremento della concentrazione di ABA xilemati-co che agisce sulla conduttanza stomatica (Hartung etal., 1988; Bacon et al., 1998).
Inoltre gli studi microarray effettuati su tessutivegetativi di Cabernet Sauvignon in stress idrico

Ferrandino et al.
38
mostrano una riduzione dei trascritti relativi a geniche codificano per enzimi di parete (poligalatturonasi,pectinmetilesterasi, espansine, xiloglucano-endotran-sglicosilasi (XET)) (Cramer et al., 2007). Una rispo-sta simile si ritrova negli acini sottoposti a stress idri-co, in cui si verifica anche una diminuzione dei livellidi trascritti relativi a proteine indotte o che rispondo-no all’auxina; questo porterebbe a ipotizzare unabbassamento della concentrazione di auxina e unconseguente minor contributo all’acidificazione diparete (Grimplet et al., 2007).
Tutto questo si traduce in un rallentamento o unarresto della crescita in risposta allo stress idrico,anche se sono presenti differenze fra cultivar. La cre-scita in condizioni di stress sembra essere sostenutameglio da Chardonnay rispetto a Cabernet Sauvignon;questa differenza potrebbe essere dovuta all’elevataquantità di xiloglucano-endotransglicosilasi (XET)osservata in Chardonnay rispetto all’altra varietà(Vincent et al., 2007).
Effetti sul metabolismo energeticoCome sopra accennato, lo stress idrico induce un
calo dell’attività fotosintetica, in quanto, come attesta-to da diversi studi (Chaves, 2002), la chiusura deglistomi stimolata dai livelli endogeni di ABA, impedi-sce il normale assorbimento di CO2, limitando il fun-zionamento dei fotosistemi I e II. La RuBisCO attiva-si è in grado di modulare la fotosintesi agendo sullaRuBisCO; in condizioni normali solo l’85% del pooldi RuBisCO è attivato da quest’enzima in tabacco(Mott e Woodrow, 2000). Durante lo stress idrico invite si osserva un rapido aumento del livello di tra-scritto di questo enzima chiave che porta ad unmiglioramento dell’efficienza fotosintetica, incremen-tando la quantità di RuBisCO attivata.
Effetti sulla regolazione dei fattori di trascrizioneLa risposta agli stress osmotici è regolata da un
complesso network di fattori trascrizionali (Shinozakie Yamaguchi-Shinozaki, 2000). Il profilo di espres-sione di circa 200 fattori di trascrizione, presenti sulchip utilizzato per l’analisi trascrittomica in apicivegetativi di Cabernet Sauvignon, è significativamen-te influenzato da entrambi i tipi di stress abiotici con-siderati, soprattutto ad un livello elevato di stress. Trai geni indotti sono stati individuati fattori di trascrizio-ne appartenenti alla famiglia NAC. I membri di questanumerosa famiglia mostrano svariate funzioni cellula-ri (Olsen et al., 2005) e sono implicati nell’intricatoprocesso di trasduzione del segnale diretto dall’ABA.Il ruolo dei NAC sembra essere quello di aumentare lasensibilità della pianta alle concentrazioni endogene
di ABA, determinando di conseguenza una miglioretolleranza al danno osmotico (Fujita et al., 2004).Altre due importanti famiglie risultano attivate dallostress idrico: quella dei fattori trascrizionaliCBF/DREB, che giocano un ruolo importante anchenell’attivazione della risposta al freddo (Xiao et al.,2008) e quella denominata WRKY. L’espressione deigeni appartenenti a quest’ultima famiglia è localizzatanella buccia e nella polpa dell’acino e sembra esserecoinvolta nella risposta a stress abiotici e di difesa aipatogeni. Nell’acino infatti, in seguito a stress, siosserva l’aumento di trascritti corrispondenti a chitina-si, taumatine, β-1,3-glucanasi. L’aumento di tali pro-teine correlate alla patogenesi (PR) potrebbe esserespiegato se si pensa che le vie di risposta a stress bioti-ci e abiotici sono in parte sovrapponibili. Un’ipotesiche si può formulare, ma che necessita di evidenzesperimentali, è che lo stress idrico potrebbe aiutare amigliorare la resistenza a patogeni nell’acino(Grimplet et al., 2007). Sempre in quest’organo, infi-ne, questo studio ha messo in evidenza come i fattoridi trascrizione possano avere un’espressione tessuto-trattamento-specifica: un gene MYB che mostra unprofilo di espressione buccia-specifico in condizioni dibuona disponibilità di acqua viene invece espressospecificamente nella polpa in condizioni di stress idri-co.
DeidrineAnche le deidrine, proteine che si accumulano nei
tessuti vegetativi soggetti a vari tipi di stress abioticoe nei semi durante l’ultimo stadio di embriogenesi,vengono sovra-espresse durante il deficit idrico invite. Recentemente due geni molto simili tra loro checodificano per deidrine, DHN1a e DHN1b, sono statiisolati in Vitis vinifera e in Vitis riparia, specie que-st’ultima molto tollerante alle basse temperature(Xiao e Nassuth, 2006). DHN1b risulta essere espres-so solo in Vitis vinifera e quindi non sembra contri-buire alla maggiore tolleranza al freddo. Questo genepossiede 18pb in meno rispetto al DHN1a e forse unnon corretto ripiegamento della proteina, dovuto agliaminoacidi mancanti, impedisce l’attività della deidri-na o l’attivazione a basse temperature.
In tabella 2 sono elencate in forma riassuntiva lerisposte molecolari descritte.
Adattamenti di genotipi diversi allo stress idrico
Come descritto a riguardo dei differenti meccani-smi discussi, un interessante adattamento della vite,caratterizzata da un’incredibile diversità varietale,riguarda la strategia di risposta al deficit idrico, varia-

Meccanismi di resistenza a stress idrico della vite
39
Gene o composto di biosintesi
prolina, glucosio, malato
RuBisCO attivasi, citocromi, xantofille
chitinasi, taumatine, beta-1,3-glucanasi
fenilalanina ammonioliasi(PAL), stilbene sintasi, calcone sintasi, cinnamoilCoA-riduttasi
invertasi vacuolare
CHS, F3H, DFR, LDOX,F3’5’H, UFGT, GST, OMT
enzimi che modificano laparete cellulare (es. XET -xiloglucano-endotransglicosilasi )proteasomi, ubiquitine, chaperones molecolari
ioni inorganici
acquaporine
9-cis-epossicarotenoidediossigenasi (NCED)S-adenosil-L-metionina sintetasi 1 (SAM)1-aminociclopropano-1-carbossilato (ACC) ossidasitrasportatori di zuccheri,ioni, aminoacidi,nucleotidi,lipidi
NAC, CBF/DREB, WRKY,MYB
deidrine
osmolitigeni coinvolti nella regolazione fotosintetica e nella prevenzione del dannofoto-ossidativoproteine correlate allapatogenesi (PR)biosintesi dei fenilpropanoidi, dellalignina, delle proantocianidine e deimetaboliti responsabilidegli aromiscissione di zucchericomplessienzimi coinvolti nellabiosintesi di antocianine e flavonoidi
coinvolgimento nella crescita cellulare
coinvolgimento nel catabolismo delleproteineosmoliti
proteine trasportatrici diacqua
enzima coinvolto nellabiosintesi dell’ABA
enzimi coinvolti nellabiosintesi dell’etilene
famiglie di fattori trascrizionali coinvoltenelle vie di risposta astress abiotici e patogeniproteine accumulate incondizioni di stress;funzione non benconosciuta
Vitis vinifera L
Richter-110(Vitis berlandieri xVitis rupestris)
Vitis vinifera L
V. vinifera L,Vitis riparia
Cabernet Sauvignon
Cabernet Sauvignon
Cabernet SauvignonMerlot
ChardonnayCabernet Sauvignon
Cabernet Sauvignon
SavatianoCabernet Sauvignon
GrenacheChardonnay
Cabernet Sauvignon
Chardonnay
Cramer et al., 2007
Grimplet et al., 2007
Castellarin et al., 2007aCastellarin et al., 2007b
Vincent et al., 2007Grimplet et al., 2007Cramer et al., 2007
Patakas et al., 2002Grimplet et al., 2007Cramer et al., 2007Vandeleur et al., 2008
Galmés et al., 2007
Grimplet et al., 2007Cramer et al., 2007Castellarin et al., 2007b
Xiao et al., 2006a
Funzione Specie Cultivar Bibliografia
Tab. 2 - Risposte molecolari allo stress idrico.Tab. 2 - Molecular responses to water stress condition.

Ferrandino et al.
40
bile anche a livello intraspecifico (Vitis vinifera L) enon unicamente interspecifico (Schultz, 2003).
In effetti, non sorprende che una pianta conosciutae coltivata sin dall’antichità come la vite, con un area-le di distribuzione vastissimo così come il numero dicultivar accertate (10-20.000), possa essere fenotipi-camente molto eterogenea, ma è certamente peculiareil fatto che questa variabilità porti a tali differenzefisiologiche. Genericamente, Vitis vinifera L. è classi-ficata come ecologicamente “pessimista”, per viadella sua tendenza a ritardare le condizioni di stressrallentando traspirazione, crescita e metabolismi, sinoalla scomparsa del fenomeno limitante; in realtà non èsempre così. Cultivar originarie di zone diverse, carat-terizzate da condizioni climatiche distinte, mostranocomportamenti tanto peculiari da poter essere classifi-cate come anisoidriche e isoidriche.
Proprio a questa ampia varietà comportamentalefanno riferimento i risultati di Schultz (2003) relativialla diversità intraspecifica di Vitis vinifera, che indi-vidua in Syrah e Grenache due rappresentanti di com-portamenti fisiologici nettamente distinti, all’internodella medesima specie: anisoidrico e quasi isoidrico.Tali indicazioni sono emerse in seguito ad analisidegli scambi gassosi e del potenziale idrico in condi-zioni di siccità. Così che, se Grenache, originaria delbacino del Mediterraneo, tende a mantenere un poten-ziale fogliare pressoché costante durante il decorsodella giornata e delle stagioni (comportamento isoidri-co, o ecologicamente “pessimista”), Syrah, nativadella Valle del Reno, regola il potenziale idrico pro-porzionandolo, inversamente, al livello evaporativodettato dall’ambiente e dalle stagioni (comportamentoanisoidrico, o ecologicamente “ottimista”). Questadifferenza comportamentale per i medesimi genotipiviene descritta e confermata anche a livello dei tessutifogliari in una più recente pubblicazione di Soar et al.(2006), o descritta per altri vitigni, ad esempioSangiovese, isoidrico a bassi livelli di stress, ma ani-soidrico quando lo stress diventa più severo (Poni etal., 2007) e Chardonnay, anisoidrico (Vandeleur etal., 2008).
Le acquaporine, per attività, composizione edespressione, sembrano dare un importante contributoalle diverse risposte che le radici di Vitis vinifera pos-sono dare allo stress idrico. Cultivar “ottimiste”(Chardonnay) reagiscono con un blando abbassamen-to della conduttanza idraulica radicale, mantenendobasso il gradiente di potenziale idrico tra lo xilema e ilsuolo; ciò potrebbe spiegare perché molte cultivar ani-soidriche presentano anche una minor propensioneagli embolismi. D’altro canto, cultivar “pessimiste”riducono l’espressione delle acquaporine in stress
idrico, e proprio per prevenire conseguenze di poten-ziali idrici eccessivamente negativi a livello xilemati-co, necessitano di un maggior controllo stomatico(Vandeleur et al., 2008) e di fenomeni efficienti delrecupero dell’embolizzazione (Lovisolo et al.,2008a).
La discriminazione isotopica del carbonio come stru-mento di screening dell’adattamento varietale allostress idrico
Il rapporto tra gli isotopi 13C e 12C nei tessuti vege-tali è minore di quello atmosferico, ad indicare che lapianta discrimina a sfavore del 13C durante l’assimila-zione fotosintetica. La diffusione della 13CO2 attraver-so la rima stomatica è infatti inferiore a quella della12CO2 di un fattore 4,4‰. Inoltre, anche la ribulosiobisfosfato carbossilasi (RuBisCO) discrimina a favoredella 12CO2 rispetto alla 13CO2 (secondo un fattore di~27‰). In entrambi i casi, il processo discrimina l’i-sotopo pesante (Farquhar et al., 1989), quantitativa-mente meno presente nella miscela aria.
Quando negli spazi intercellulari del mesofillofogliare la concentrazione di CO2 diminuisce - e que-sto accade sia quando è maggiore l’assimilazione (A),che sottrae CO2 dall’apoplasto, sia quando è minore laconduttanza stomatica (gs), via d’ingresso della CO2 -anche il tasso di discriminazione verso l’isotopopesante diminuisce. Tale tasso viene comunementeespresso come ‰13C, che esprime il rapporto 13C/12Cin relazione ad un rapporto isotopico standard(PeeDee Belemnite) (Craig, 1957).
L’efficienza di uso dell’acqua (WUE, Water UseEfficiency) può essere espressa come rapporto tra A egs o tra A e la traspirazione (E), che è funzione dellags e del deficit di pressione di vapore nell’aria. Sullabase di quanto detto, la WUE è maggiore se diminui-sce la gs (cioè in presenza di chiusura stomatica) e/ose aumenta l’assimilazione (Pou et al., 2008).Chiusura stomatica ed assimilazione carbonica dimi-nuiscono la concentrazione di CO2 intercellulare e diconseguenza la discriminazione verso la 13CO2. Sia laWUE che la regolazione stomatica sono quindi corre-late al ‰13C. Per questo motivo il valore del ‰13Caccumulato nei tessuti fogliari è indice dal regime sto-matico (e fotosintetico) medio presente durante la vitadella foglia.
Durante la maturazione dell’acino, il saccarosio èfloematicamente traslocato dalle foglie ai frutti e rapi-damente convertito in glucosio e fruttosio (Davies eRobinson, 1996). Per questo motivo il rapporto traidue isotopi di carbonio negli zuccheri dell’acinomaturo dipende dalla discriminazione isotopica avve-nuta durante la fotosintesi nella foglia nel periodo

Meccanismi di resistenza a stress idrico della vite
41
della maturazione, come dimostrato da Di Marco etal. (1977). Partendo da questo presupposto,Gaudillère et al. (2002) hanno stabilito in 31 differentigenotipi una relazione tra il ‰13C nell’acino alla rac-colta e lo stress idrico avvertito dalla pianta nell’estatedel 1999 in un vigneto sperimentale a Ferrade, nelBordolese. Ordinando i 31 genotipi a ‰13C decre-scente, gli Autori hanno proposto un metodo permisurare la capacità di controllo stomatico dei diffe-renti genotipi. Nel loro ranking, Riesling e Grenachesuperano il Syrah, dimostrando una maggior efficien-za delle prime nel controllare stomaticamente lo stressidrico, come discusso dagli autori, o una miglioreWUE, forse causata da una migliore assimilazione. Incondizioni di stress idrico, si può quindi supporre cheil δ ‰13C nella foglia o nell’acino sia indice del com-portamento iso o anisoidrico dei differenti genotipi.
Sviluppi di ricerca futuri
Funzione delle acquaporine e trasduzione delsegnale di acido abscissico (ABA) sono interconnessedurante le situazioni di stress idrico (Tyerman et al.,2002; Zhu et al., 2005). Tale regolazione appare stra-tegica a livello di idraulica della radice (Vandeleur etal., 2008), di controllo dell’embolizzazione e delrecupero degli embolismi (Lovisolo et al., 2008a), diregolazione dei flussi verso l’acino durante la matura-zione (Foquet et al., 2008; Castellarin et al., 2007a), epiù in generale dei meccanismi iso ed anisoidrici dirisposta a stress idrico della vite. Oltre alla variabiltàintraspecifica di Vitis vinifera L, le relazioni idrichedel portinnesto (per lo più ibridi di Vitis spp.) condi-zionano biosintesi e flussi da radice a foglia di ABA eruolo delle acquaporine nella radice. In campo sidispone di innumerevoli combinazioni cultivar/portin-nesto, le quali all’atto pratico possono presentarerisposte assai peculiari a situazioni di stress idrico.Tali risposte potranno essere studiate in quest’otticafisiologica, per poter prevedere e quindi esasperare ocontenere le risposte della pianta, puntando ad otti-mizzare i fenomeni di tolleranza della pianta. Questonell’evidente prospettiva di migliorare gli aspetti delmetabolismo della pianta, e soprattutto dell’acino, intali situazioni.
Ringraziamenti
Gli autori ringraziano Andrea Schubert, nel labora-torio del quale hanno maturato l’esperienza nei diversiaspetti della fisiologia delle relazioni idriche dellavite.
Riassunto
In questa review, affrontiamo i principali aspettidella resistenza della vite (Vitis vinifera L) a stress idri-co, soffermandoci sulle risposte ecofisiologiche, fisio-logiche e molecolari legate al metabolismo primario esecondario, soprattutto a livello di acino. Esaminiamo imeccanismi di elusione dello stress, relativi al control-lo stomatico, al ruolo di segnale dell’acido abscissico,a quello del segnale idraulico (embolismi) e delleacquaporine. Descriviamo i diversi meccanismi di tol-leranza, sia dal punto di vista dell’induzione e regola-zione delle vie biosintetiche, sia dell’accumulo deimetaboliti derivati. In tutti gli aspetti esaminati, sottoli-neiamo le peculiarità varietali conosciute.
Parole chiave: acquaporine, acido abscissico (ABA),polifenoli, aromi, analisi trascrittomica.
BibliografiaAHARON R., SHAHAK Y., WININGER S., BENDOV R., KAPULNIK Y.,
GALILI G., 2003. Overexpression of a plasma membraneaquaporin in transgenic tobacco improves plant vigor underfavorable growth conditions but not under drought or saltstress. The Plant Cell 15: 439-447.
ALEXANDERSSON E., FRAYSSE L., SJOVALL-LARSEN S.,GUSTAVSSON S., FELLERT M., KARLSSON M., JOHANSON U.,KJELLBOM P., 2005. Whole gene family expression anddrought stress regulation of aquaporins. Plant MolecularBiology 59: 469-484.
ALSINA M.M., DE HERRALDE F., ARANDA X., SAVÈ R., BIEL C.,2007. Water relations and vulnerability to embolism are notrelated: experiments with eight grapevine cultivars. Vitis 46(1): 1-6.
ASADA K., 1999. The water-water cycle in chloroplasts: scaven-ging of active oxygens and dissipation of excess photons.Annual Review of Plant Physiology and Plant MolecularBiology 50: 601-639.
BACON M.A., WILKINSON S., DAVIES W.J., 1998. pH-regulatedleaf cell expansion in droughted plants is abscisic acid depen-dent. Plant Physiology 118: 1507-1515.
BAN T., ISHIMARU M., KOBAYASHI S., GOTO-YAMAMOTO S.N.,HORIUCHI S., 2003. Abscisic acid and 2,4-dichlorophenoxy-acetic acid affect the expression of anthocyanin biosyntheticpathway genes in ‘Kyoho’ grape berries. Journal ofHorticoltural Science and Biotechnology 78: 586-89.
BERTAMINI M., NEDUNCHEZHIAN N., 2002. Photoinhibition of pho-tosynthesis in Vitis berlandieri and Vitis rupestris leavesunder field conditions. Photosynthetica 40(4): 597-603.
BERTAMINI M., ZULINI L., MUTHUCHELIAN K., NEDUNCHEZHIAN N.,2006. Effect of water deficit on photosynthetic and other phy-siological responses in grapevine (Vitis vinifera L. cv.Riesling) plants. Photosynthetica 44(1): 151-154.
BINDON K.A., DRY P.R., LOVEYS B.R., 2007. Influence of plantwater status on the production of C13-norisoprenoid precur-sors in Vitis vinifera L. cv Cabernet Sauvignon grape berries.Journal of Agricultural and Food Chemistry 55: 4493-4500.
BRAVDO B., HEPNER Y., LOINGER S., COHEN S., TABACMAN H.,1985. Effect of irrigation and crop level on growth and winequality of Cabernet Sauvignon. American Journal of Enologyand Viticulture 36:132-139.

Ferrandino et al.
42
BRAY E.A., 1997. Plant responses to water deficit. Trends in PlantScience 2: 48-54.
CASTELLARIN S.D., MATTHEWS M.A., DI GASPERO G., GAMBETTAG.A., 2007a. Water deficits accelerate ripening and inducechanges in gene expression regulating flavonoid biosynthesisin grape berries. Planta 227:101-112.
CASTELLARIN S.D., PFEIFFER A., SIVILOTTI P., DEGAN M.,PETERLUNGER E., DI GASPERO G., 2007b. Transcriptional re-gulation of anthocyanin biosynthesis in ripening fruits ofgrapevine under seasonal water deficit. Plant, Cell andEnvironment 30 (11): 1381-1399.
CHAVES M., 2002. Water stress in the regulation of photosynthesisin the field. Annals of Botany 89: 907-916.
CIFRE J., BOTA J., ESCALONA J.M., MEDRANO H., FLEXAS J., 2005.Physiological tools for irrigation scheduling in grapevine(Vitis vinifera L). An open gate to improve water-use efficien-cy? Agriculture, Ecosystems and Environment 106: 159-170.
COOMBE B.G., HALE C.R., 1973. The hormone content of ripeninggrape berries and the effects of growth substance treatments.Plant Physiology 51: 629-634.
CRAIG H., 1957. Isotopic standards for carbon and oxygen andcorrelation factors for mass-spectrometric analysis of carbondioxide. Geochimical Acta 12:133-149.
CRAMER G.R., ERGUL A., GRIMPLET J. TILLET R.L., TATTERSALA.R., BOHLMAN M.C., VINCENT D., SONDEREGGER J., EVANSJ., OSBORNE C., QUILLICI D., SCHLAUCH K., SCHOOLEY D.A.,CUSHMAN J.C., 2007. Water and salinity stress in grapevines:early and late changes in transcript and metabolite profiles.Functional Integrative Genomics 7: 111-134.
DAVIES C., BOSS P.K., ROBINSON S.P., 1997. Treatment of grapeberries, a nonclimateric fruit with a synthetic auxin, retardsripening and alters the expression of developmentally regula-ted genes. Plant Physiology 115: 1155-61.
DAVIES C., ROBINSON S.P., 1996. Sugar accumulation in grapeberries. Cloning of two putative vacuolar invertase cDNAsand their expression in grapevine tissues. Plant Physiology111(1): 275–283.
DI MARCO G., GREGO S., TRICOLI D., TURI B., 1997. Carbon iso-tope ratios (13C/12C) in fractions of field-grown grape.Physiologia Plantarum 41: 139-141.
DRY P.R., LOVEYS B.R., MCCARTHY M.G., STOLL M., 2001.Strategic irrigation management in Australian vineyards.Journal international des Sciences de la Vigne et du Vin35(3): 29-139.
DRY P.R., LOVEYS B.R., 1999. Grapevine shoot growth and sto-matal conductance are reduced when part of the root systemis dried. Vitis 38: 151-156.
ESCALONA J.M., FLEXAS J., MEDRANO H., 1999. Stomatal andnon-stomatal limitations of photosynthesis under water stressin field-grown grapevines. Australian Journal of PlantPhysiology 26: 421-433.
FARQUHAR G.D., EHLERINGER J.R., HUBICK K.T, 1989. Carbonisotope discrimination and photosynthesis. Annual Review ofPlant Physiology and Plant Molecular Biology 40: 503-537.
FLEXAS J., BOTA J., LORETO F., CORNIC G., SHARKEY T.D., 2004.Diffusive and metabolic limitations to photosynthesis underdrought and salinity in C3 plants. Plant Biology 6: 1-11.
FOUQUET R., LEON C., OLLAT N., BARRIEU F., 2008. Identificationof grapevine aquaporins and expression analysis in develo-ping berries. Plant Cell Reports 27(9): 1541-1550.
FUJITA M., FUJITA Y., MARUYAMA K., SEKI M., HIRATSU K.,OHME-TAKAGI M., TRAN L.S., YAMAGUCHI-SHINOZAKI K.,SHINOZAKI K., 2004. A dehydration-induced NAC protein,RD26, is involved in a novel ABA-dependent stress-signallingpathway. Plant Journal 39: 863-876.
GALMÉS J., POU A., ALSINA M.M., TOMÀS M., MEDRANO H.,FLEXAS J., 2007. Aquaporin expression in response to diffe-rent water stress intensities and recovery in Richter-110 (Vitis
sp.): relationship with ecophysiological status. Planta 226:671-681.
GAMON J.A., PEARCY R.W., 1990a. Photoinhibition in Vitis cali-fornica: interactive effect of sunlight, temperature and waterstatus. Plant Cell and Environment 13: 267-275.
GAMON J.A., PEARCY R.W., 1990b. Photoinhibition in Vitis cali-fornica. The role of temperature during high-light treatment.Plant Physiology 92: 487-494.
GAUDILLÈRE J.P., VAN LEEUWEN C., OLLAT N., 2002. Carbon iso-tope composition of sugars in grapevine, an integrated indi-cator of vineyard water status. Journal of ExperimentalBotany 53(369): 757-763.
GIRIBALDI M., PERUGINI I., SAUVAGE S-X., SCHUBERT A., 2007.Analysis of protein changes during grape berry ripening by 2-DE and MALDI-TOF. Proteomics 7 (17): 3154–3170.
GOMEZ-DEL-CAMPO M., RUIZ C., BAEZA P., LISSARRAGUE J.R.,2003. Drought adaptation strategies of four grapevine culti-vars (Vitis vinifera L.): Modification of the properties of theleaf area. Journal international des Sciences de la Vigne et duVin 37 (3): 131-143.
GRIMPLET J., DELUC L.G., TILLETT R.L., WHEATLEY M.D.,SCHLAUCH K.A., CRAMER G.R., CUSKMAN J.C., 2007. Tissue-specific mRNA expression profiling in grape berry tissues.BMC Genomics 8 (187).
GUAN X.Q., ZHAO S.J., LI D.Q., SHU H.R., 2004. Photoprotectivefunction of photorespiration in several grapevine cultivarsunder drought stress. Photosynthetica 42(1): 31-36.
HARTUNG W., RADIN J.W., HENDRIX D.L., 1988. Abscisic acidmovement into the apoplastic solution of water-stressed cot-ton leaves: role of apoplastic pH. Plant Physiology 86: 908-913.
HAWKER J.S., WALKER R.R., 1978. The effect of sodium chlorideon the growth and fruiting of Cabernet Sauvignon vines.American Journal of Enology and Viticulture 29:172-176.
HOLMBERG N., BULOW L., 1998. Improving stress tolerance inplants by gene transfer. Trends in Plant Science 3: 61-66.
JANG J.Y., KIM D.G., KIM Y.O., KIM J.S., KANG H., 2004. Anexpression analysis of a gene family encoding plasma mem-brane aquaporins in response to abiotic stresses inArabidopsis thaliana. Plant Molecular Biology 54: 713-725.
JEONG S.T., GOTO-YAMAMOTO N., KOBAYASHI S., ESAKA M.,2004. Effects of plant hormones and shading on the accumu-lation of anthocyanins and the expression of anthocyaninbiosynthetic genes in grape berry skins. Plant Science 167:247-52.
KALDENHOFF R., RIBAS CARBO M., FLEXAS SANS J., LOVISOLO C.,HECKWOLF M., UEHLEIN N., 2008. Aquaporins and PlantWater Balance. Plant, Cell and Environment 31 (5): 658-666.
KELLER M., 2005. Deficit Irrigation and Vine Mineral Nutrition.American Journal of Enology and Viticulture 56 (3): 267-283.
KENNEDY J.A., MATTHEWS M.A., WATERHOUSE A.L., 2002. Effectof maturity and vine water status on grape skins and wineflavonoids. American Journal of Enology and Viticulture 53:268-274.
KOUNDOURAS S., MARINOS V., GKOULIOTI A., KOTSERIDIS Y., VANLEEUWEN C., 2006. Influence of vineyard location and vinewater status on fruit maturation of nonirrigated cvAgiorgitiko (Vitis vinifera L.) effects on wine phenolic andaroma components. Journal of Agricultural and FoodChemistry 54: 5077-5086.
KRAMER P.J., BOYER J.S., 1995. Water relations of plants andsoils. Academic Press (San Diego).
LEVITT J., 1972. Responses of plants to environmental stresses.Academic Press (New York).
LONG S.P., HUMPHRIES S., FALKOWSKI P.G., 1994:Photoinhibition of photosynthesis in nature. Annual Reviewof Plant Physiology and Plant Molecular Biology 45: 633-662.

Meccanismi di resistenza a stress idrico della vite
43
LOVISOLO C., HARTUNG W., SCHUBERT A., 2002. Whole-planthydraulic conductance and root-to-shoot flow of abscisic acidare independently affected by water stress in grapevines.Functional Plant Biology 29: 1349-1356.
LOVISOLO C., SCHUBERT A., 1998. Effects of water stress on vesselsize and xylem hydraulic conductivity in Vitis vinifera L.Journal of Experimental Botany 49 (321): 693-700.
LOVISOLO C., SCHUBERT A., 2006. Mercury hinders recovery ofshoot hydraulic conductivity during grapevine rehydration:evidence from a whole-plant approach. New Phytologist 172(3): 469-478.
LOVISOLO C., PERRONE I., HARTUNG W., SCHUBERT A., 2008a. Anabscisic acid-related reduced transpiration promotes gradualembolism repair when grapevines are rehydrated afterdrought. New Phytologist 180 (3): 642-651.
LOVISOLO C., TRAMONTINI S., FLEXAS J., SCHUBERT A., 2008b.Mercurial inhibition of root hydraulic conductance in Vitisspp. rootstocks under water stress. Environmental andExperimental Botany 63: 178-182.
MARTRE P., NORTH G.B., NOBEL P.S., 2001. Hydraulic conduc-tance and mercury-sensitive water transport of roots ofOpuntia acanthocarpa in relation to soil drying and rewet-ting. Plant Physiology 126: 352-362.
MATHIEU S., TERRIE N., PROCUREUR J., BIGEY F., GUNATA Z., 2005. Acarotenoid cleavage dioxygenase from Vitis vinifera L. func-tional characterization and expression during grape berrydevelopment in relation to C13-norisoprenoid accumulation.Journal of Experimental Botany 56: 2721-2731.
MATTHEWS M.A., ANDERSON M.M., 1988. Fruit ripening in Vitisvinifera L.: responses to seasonal water deficits. AmericanJournal of Enology and Viticulture 39 (4): 313-320.
MATTHEWS M.A., ISHII R., ANDERSON M.M., O’MAHONY M.,1990. Dependence of wine sensory attributes on vine waterstatus. Journal of the Science of Food and Agriculture, 51:321-335.
MAUREL C., VERDOUCQ L., LUU D-T, SANTONI V., 2008. Plantaquaporins: membrane channels with multiple integratedfunctions. Annual Review of Plant Biology 59: 595-624.
MEDRANO H., BOTA J., ABADIA A., SAMPOL B., ESCALONA J.M.,FLEXAS J., 2002. Effects of drought on light-energy dissipationmechanisms in high-light-acclimated, field-grown grapevines.Functional Plant Biology 29(10): 1197-1207.
MEDRANO H., ESCALONA J.M., CIFRE J., BOTA J., FLEXAS J., 2003.A ten-year study on the physiology of two Spanish grapevinecultivars under field conditions: effects of water availabilityfrom leaf photosynthesis to grape yield and quality.Functional Plant Biology 30: 607-619.
MOTT K.A., WOODROW I.E., 2000. Modelling the role of Rubiscoactivase in limiting non-steady-state photosynthesis. Journalof Experimental Botany 51: 399-406.
MUNNS R., 2002. Comparative physiology of salt and waterstress. Plant Cell and Environment 25: 239-250.
NORTH G.B., MARTRE P., NOBEL P.S., 2004. Aquaporins accountfor variations in hydraulic conductance for metabolicallyactive root regions of Agave desertii in wet dry and rewettedsoil. Plant Cell and Environment 27: 219-228.
OJEDA H., ANDARY C., KRAEVA E., CARBONNEAU A., DELOIRE A.,2002. Influence of pre- and postveraison water deficit on syn-thesis and concentration of skin phenolic compounds duringberry growth of Vitis vinifera cv Shiraz. American Journal ofEnology and Viticulture 53(4): 261-267.
OLIVEIRA C., SILVA FERREIRA A.C., MENDES PINTO M., HOGG T.,ALVES F., GUEDES DE PINHO P., 2003. Carotenoid compoundsin grapes and their relationship to plant water status. Journalof Agriculture and Food Chemistry 51:5967-5971.
OLSEN A.N., ERNST H.A., LEGGIO L.L., SKRIVER K., 2005. NACtranscription factors: structurally distinct, functionallydiverse. Trends in Plant Science 10: 79-87.
OSMOND B., BADGER M., MAXWELL K., BJORKMAN O., LEEGOODR., 1997. Too many photons: photorespiration, photoinhibitionand photooxidation. Trends in Plant Science 2(4): 119-121.
PALLIOTTI A., SILVESTRONI O., PETOUMENOU D., VIGNAROLI S.,BERRIOS J.G., 2008. Evaluation of low-energy demand adapti-ve mechanisms in Sangiovese grapevine during drought.Journal international des Sciences de la Vigne et du Vin 42(1): 41-47.
PASSIOURA J.B., MUNNS R., 2000. Rapid environmental changesthat affect leaf water status induce transient surges or pausesin leaf expansion rate. Australian Journal of Plant Physiology27: 941-948.
PATAKAS A., 2000. Changes in the solutes contributing to osmoticpotential during leaf ontogeny in grapevine leaves. AmericanJournal of Enology and Viticulture 51: 223-226.
PATAKAS A., NIKOLAU N., ZIOZIOU E., RADOGLOU K., NOITSAKISB., 2002. The role of organic solute and ion accumulation inosmotic adjustment in drought-stressed grapevines. PlantScience 163: 361-367.
PATAKAS A., NOITSAKIS B., 1997. Cell wall elasticity as a mecha-nism to maintain favorable water relations during leafontogeny in grapevines. American Journal of Enology andViticulture 48(3): 352-356.
PATAKAS A., NOITSAKIS B., 1999. Osmotic adjustment and parti-tioning of turgor responses to drought in grapevines leaves.American Journal of Enology and Viticulture 50 (1): 76-80.
PEDREIRA DOS SANTOS T., LOPES C.M., RODRIGUES M.L., DESOUZA C.R., RICARDO-DA-SILVA J.M., MAROCO J.P., PEREIRAJ.S., CHAVES M.M., 2007. Effect of deficit irrigation strategieson cluster microclimate for improving composition ofMoscatel field-grown grapevines. Scientia Horticulturae 112:321-330.
PEYROT DES GACHONS C., VAN LEEUWEN C., TOMINAGA T, SOYERJ.P., GAUDILLÈRE J.P., DUBOURDIEU D., 2005. Influence ofwater and nitrogen deficit on fruit ripening and aroma poten-tial of Vitis vinifera L cv Sauvignon blanc in filed conditions.Journal of the Science of Food and Agriculture 85: 73-85.
PONI S., BERNIZZONI F., CIVARDI S., 2007. Response of‘Sangiovese’ grapevines to partial root-zone drying: gas-exchange, growth and grape composition. ScientiaHorticulturae 114: 96-103.
POU A., FLEXAS J., ALSINA M.M., BOTA J., CARAMBULA C., DEHERRALDE F., GALMÉS J., LOVISOLO C., JIMÉNEZ M., RIBAS-CARBO M., RUSJAN D., SECCHI F., TOMAS M., ZSÓFI Z.,MEDRANO H., 2008. Adjustments of water-use efficiency bystomatal regulation during drought and recovery in thedrought-adapted Vitis hybrid Richter-110 (V. berlandieri xV.rupestris). Physiologia plantarum 134 (2): 313 – 323.
QUINTERO J.M., FOURNIER J.M., BENLLOCH M., 1999. Water trans-port in sunflower root systems: effects of ABA, Ca2+ statusand HgCl2. Journal of Experimental Botany 50: 1607-1612.
RABBANI M.A., MARUYAMA K., HIRISHI A., KHAN M.A., KATSURAK., ITO Y., YOSHIWARA K., SEKI M., SHINOZAKI K.,YAMAGUCHI-SHINOZAKI K., 2003. Monitoring the expressionprofiles of rice genes under cold, drought and high-salinitystresses and abscisic acid application using cDNA microarrayand RNA gel-blot analyses. Plant Physiology 133: 1755-1767.
RAVEN J.A., 1985. Regulation of pH and generation of osmolarityin vascular plants: a cost-benefit analysis in relation to effi-ciency of use of energy, nitrogen and water. New Phytologist101: 25-77.
ROBY G., HABERSTON J.F., ADAMS D.A., MATTHEWS M.A., 2004.Berry size and vine water deficits as factors in winegrapecomposition: anthocyanins and tannins. Australian Journal ofGrape and Wine Research 10: 100-107.
RODRIGUES M.L., SANTOS T.P., RODRIGUES A.P., DE SOUZA C.R.,LOPES C.M., MAROCO J.P., PEREIRA J.S., CHAVES M.M., 2008.Hydraulic and chemical signalling in the regulation of sto-

Ferrandino et al.
44
matal conductance and plant water use in field grapevinesgrowing under deficit irrigation. Functional Plant Biology 35:565-579.
SCHREINER R.P., TARARA J.M., SMITHYMAN R.P., 2007. Deficitirrigation promotes arbuscular colonization of fine roots bymycorrhizal fungi in grapevines (Vitis vinifera L.) in an aridclimate. Mycorrhiza 17: 551-562.
SCHULTZ H.R., 2003. Differences in hydraulic architectureaccount for near isohydric and anisohydric behaviours of twofield-grown Vitis vinifera L. cultivars during drought. PlantCell and Environment 26: 1393-1405.
SECCHI F., LOVISOLO C., UEHLEIN N., KALDENHOFF R., SCHUBERTA., 2007. Isolation and functional characterization of threeaquaporins from olive (Olea europaea L.). Planta 225: 381-392.
SEKI M., NARUSAKA M., ISHIDA J., NANJO T., FUJITA M., OONO Y.,KAMIYA A., NAKAJIMA M., ENJU A., SAKURAI T., SATOU M.,AKIYAMA K., TAJI T., YAMAGUCHI-SHINOZAKI K., CARNINCI P.,KAWAI J., HAYASHIZAKI Y., SHINOZAKI K., 2002. Monitoringthe expression profiles of 7000 Arabidopsis genes underdrought, cold and high-salinity stresses using a full-lengthcDNA microarray. Plant Cell 31: 279-292.
SHANI U., WAISEL Y., ESHEL A., XUE S., ZIV G., 1993. Responsesto salinity of grapevine plants with split root systems. NewPhytologist 124: 695-701.
SHINOZAKI K., YAMAGUCHI-SHINOZAKI K., 2000. Molecularresponses to dehydration and low temperature: differencesand cross-talk between two stress signalling pathways.Current Opinion in Plant Biology 3: 217-223.
SIEFRITZ F., TYREE M.T., LOVISOLO C., SCHUBERT A.,KALDENHOFF R., 2002. PIP1 plasma membrane aquaporins intobacco: from cellular effects to function in plants. The PlantCell 14: 869-876.
SOAR C.J., LOVEYS, B.R., 2007. The effect of changing patterns insoil-moisture availability on grapevine root distribution, andviticultural implications for converting full-cover irrigationinto a point-source irrigation system. Australian Journal ofGrape and Wine Research 13 (1): 2-13.
SOAR C.J., SPEIRS J., MAFFEI S.M., PENROSE A.B., MCCARTHYM.G., LOVEYS B.R., 2006. Grape vine varieties Shiraz andGrenache differ in their stomatal response to VPD: apparentlinks with ABA physiology and gene expression in leaf tissue.Australian Journal of Grape and Wine Research (12):2-12.
STEUDLE E., 2000. Water uptake by plant roots: an integration ofviews. Plant and Soil 226: 45-56.
STOLL M., LOVEYS B., DRY P., 2000. Ormonal changes inducedby partial rootzone drying of irrigated grapevine. Journal ofExperimental Botany 51: 1627-1634.
TAJI T., OHSUMI C., IUCHI S., SEKI M., KASUGA M., KOBAYASHIM., YAMAGUCHI-SHINOZAKI K., SHINOZAKI K., 2002.Important roles of drought- and cold-inducible genes forgalactinol synthase in stress tolerance in Arabidopsisthaliana. The Plant Journal 29: 417-426.
TARDIEU F., 2003. Virtual plants: modelling as a tool for thegenomics of tolerance to water deficit. Trends in PlantScience 8 (1): 9-14.
TARDIEU F., SIMONNEAU T., 1998. Variability among species ofstomatal control under fluctuating soil water status and eva-porative demand: modelling isohydric and anisohydric beha-viours. Journal of Experimental Botany 49: 419-432.
TOUMI I., GARGOURI M., NOUAIRI I., MOSCHOU P.N., SALEM-FNAYOU A., MLIKI A., ZARROUK M., GHORBEL A., 2008.
Water stress induced changes in the leaf lipid composition offour grapevine genotypes with different drought tolerance.Biologia Plantarum 52 (1): 161-164.
TYERMAN S.D., NIEMIETZ C.M., BRAMLEY H., 2002. Plant aqua-porins: multifunctional water and solute channels withexpanding roles. Plant, Cell and Environment 25: 173-194.
VANDELEUR R.K., MAYO G., SHELDEN M.C., GILLIHAM M.,KAISER B.N., TYERMAN S.D., 2008. The role of PIP aquapor-ins in water transport through roots: diurnal and droughtstress responses reveal different strategies between isohydricand anisohydric cultivars of grapevine. Plant PhysiologyPreview November 5, 2008; 10.1104/pp.108.128645.
VERA-ESTRELLA R., BARKLA B.J., BOHNERT H.J., PANTOIA O.,2004. Novel regulation of aquaporins during osmotic stress.Plant Physiology 135: 2318-2329.
VERSLUES P.E., AGARWAL M., KATIYAR-AGARWAL S., ZHU J. ANDZHU J.K., 2006. Methods and concepts in quantifying resi-stance to drought, salt and freezing, abiotic stresses thataffect plant water status. The Plant Journal 45: 523-539.
VEZZULLI S., CIVARDI S., FERRARI F., BAVARESCO L., 2007.Methyl jasmonate treatment as a trigger of resveratrol syn-thesis in cultivated grapevine. American Journal of Enologyand Viticulture 58(4): 530-533.
VINCENT D., ERGUL A., BOHLMAN M.C., TATTERSALE E.A.R.,TILLETT R.L., WHEATLEY M.D., WOOLSEY R., QUILICI D.R.,JOETS J., SCHLAUCH K., SCHOLLEY D.A., CUSHMAN J.C.,CRAMER G.R., 2007. Proteomic analysis reveals differencesbetween Vitis vinifera L. cv Chardonnay and cv CabernetSauvignon and their responses to water deficit and salinity.Journal of Experimental Botany 58: 1873-1892.
WADA H., SHACKEL K.A, MATTHEWS M.A., 2008. Fruit ripeningin Vitis vinifera: apoplastic solute accumulation accounts forpre-veraison turgor loss in berries. Planta 227: 1351-1361.
WILKINSON S., DAVIES W.J., 2002. ABA-based chemical sig-nalling: the co-ordination of responses to stress in plants.Plant Cell and Environment 25: 195-210.
WILLIAMS P.J., CYNCAR W., FRANCIS I.L., GRAY J.D., ILAND P.G.,COOMBE B.G., 1995. Quantification of glycosides in grapes,juices and wines through a determination of glycosyl-glucose(G-G). Journal of Agriculture and Food Chemistry 43: 121-128.
WU Y., JEONG B.R., FRY S.C., BOYER J.S., 2005. Change in XETactivities, cell wall extensibility and hypocotyls elongation ofsoybean seedlings at low water potential. Planta 220: 593-601.
XIAO H., NASSUTH A., 2006. Stress- and development-inducedexpression of spliced and unspliced transcripts from twohighly similar dehydrin 1 genes in V. riparia and V. vinifera.Plant Cell Reports 25: 968-977.
XIAO H., TATTERSALE E.A.R., SIDDIQUA M.K., CRAMER G.R.,NASSUTH A., 2008. CBF4 is a unique member of the CBFtranscription factor family of Vitis vinifera and Vitis riparia.Plant Cell and Environment 31: 1-10.
YAMADA S., KOMORI T., MYERS P.N., KUWATA S., KUBO T.,IMASEKI H., 1997. Expression of plasma membrane waterchannel genes under water stress in Nicotiana excelsior.Plant and Cell Physiology 38:1226-1231.
ZHU C., SCHRAUT D., HARTUNG W., SCHÄFFNER A.R., 2005.Differential responses of maize MIP genes to salt stress andABA. Journal of Experimental Botany 56 (421): 2971-2981.
ZIMMERMANN M.H., 1983. Xylem structure and the ascent of sap.Springer Verlag (Berlin).























![Appalti Pubblici Pre-Commerciali - [MIDES] Forges...Sfide sociali S3 -Smart Puglia 2020 • Città e territori sostenibili (risorse idriche, controllo e gestione del territorio, air](https://img.pdfslide.us/doc/110x75/5faa8b071a302a14b140819e/appalti-pubblici-pre-commerciali-mides-forges-sfide-sociali-s3-smart-puglia.jpg)





