Embed Size (px)
Citation preview

Full Communication
Matrix Metalloproteinase-1 Inhibitory Activity of Kaempferia pandurata Roxb.
Jae-Seok Shim,1 Eun-Jung Choi,2 Chan-Woo Lee,3 Han-Sung Kim,3 and Jae-Kwan Hwang1,2
Departments of 1Biomaterials Science and Engineering and 2Biotechnology, Yonsei University, Seoul;and 3R & D Center, Amore-Pacific Corporation, Kyounggi-do, Republic of Korea
ABSTRACT Matrix metalloproteinase (MMP)-1 is a superfamily of zinc-dependent endopeptidases that are capable of
degrading all components of the extracellular matrix. Kaempferia pandurata extract (0.01–0.5 mg=mL) significantly reduced
the expression of MMP-1 and induced the expression of type 1 procollagen at the protein and mRNA levels in a dose-
dependent manner. Ultraviolet (UV)-induced MMP-1 initiates cleavage of fibrillar collagen. Once cleaved by MMP-1,
collagen can be further degraded by elevated levels of MMP-3 and MMP-9. It was found that increased MMP-1 expression
due to UV irradiation was mediated by activation of mitogen-activated protein kinases such as extracellular-regulated kinase
(ERK), Jun N-terminal kinase ( JNK), and p38 kinase. Treatment of K. pandurata extract in the range of 0.01–0.5 mg=mL
inhibited the UV-induced phosphorylations of ERK, JNK, and p38, respectively. Moreover, inhibition of phosphorylated
ERK, JNK, and p38 by K. pandurata extract resulted in decreased c-Fos expression and c-Jun phosphorylation induced by UV
light. The results strongly suggest that K. pandurata is potentially useful for the prevention and treatment of skin aging.
KEY WORDS: � fibroblasts � Kaempferia pandurata � matrix metalloproteinase-1 � mitogen-activated protein kinase� type 1 procollagen � ultraviolet irradiation
INTRODUCTION
Ultraviolet (UV) irradiation is well known to in-duce photodamage and premature skin aging.1 UV ir-
radiation induces the synthesis of matrix metalloproteinases(MMPs) in fibroblasts; up-regulation of some MMPs is re-sponsible for the enhanced degradation of dermal collagenduring UV-induced skin aging.2
MMPs are a family of structurally related matrix-degrading enzymes that play important roles in variousdestructive processes, including inflammation, tumor inva-sion, and skin aging.3 Moreover, the expression of variousUV-induced MMPs in dermal fibroblasts leads to thebreakdown of collagen and other extracellular matrix pro-teins and is thus related to photoaging in human skin.4 Type 1collagen is the main structural component of the extracel-lular matrix, which is known to perform a pivotal function inthe maintenance of the structure of the skin dermis.5
Exposure of human skin in vivo to UV irradiation alsoactivates multiple cell surface cytokine and growth factorreceptors as well as mitogen-activated protein kinase(MAPK) signaling molecules such as extracellular signal-regulated kinase (ERK), Jun N-terminal kinase ( JNK), and
p38 kinase, which rapidly induces activator protein-1 (AP-1)activity.6 Transcription of several MMPs is regulated byAP-1. For AP-1 complex formation, Jun proteins formhomodimers or heterodimers with Fos proteins. Moreover,increased AP-1 activity is responsible for the degradation ofextracellular matrix proteins, such as collagen, by inducingMMPs.7
Kaempferia pandurata Roxb., a perennial herb of theZingiberaceae family, is mainly cultivated in tropical coun-tries, including Indonesia and Thailand. The fresh rhizomehas been used as a food material and also as a folk medicinefor the treatment of colic disorder, dry cough, rheumatism,and muscular pains and as an aphrodisiac.8 Several studieshave reported various biological activities of K. pandurata,including anti-inflammatory, antitumor, antidiarrhea, anti-dysentery, antiflatulence, and antiepidermophytid effects.9,10
However, its anti-aging effect has not yet been examined todate. In this study, we investigated the MMP-1 inhibition andsuppressive cellular mechanisms of K. pandurata extract inUV-irradiated human skin fibroblasts.
MATERIALS AND METHODS
Plant material
Dried rhizomes of K. pandurata were collected in Jakarta,Indonesia, and identified by Dr. Nam-In Baek, Departmentof Oriental Medicinal Materials and Processing, KyungheeUniversity, Yongin, Republic of Korea. A voucher specimen
Manuscript received 9 March 2008. Revision accepted 24 November 2008.
Address correspondence to: Prof. Jae-Kwan Hwang, Department of Biotechnology,Yonsei University, 134 Shinchon-dong, Seodaemun-gu, Seoul 120-749, Republic ofKorea, E-mail: [email protected]
JOURNAL OF MEDICINAL FOODJ Med Food 12 (3) 2009, 601–607# Mary Ann Liebert, Inc. and Korean Society of Food Science and NutritionDOI: 10.1089=jmf.2007.1041
601

is deposited in the Department of Biotechnology, YonseiUniversity, Seoul, Republic of Korea.
Preparation of plant extract
Dried rhizomes were ground and extracted with 5 vol-umes of 95% ethanol for 2 days at room temperature. Theextract was obtained by filtering and evaporating the sol-vent, which was dissolved in dimethyl sulfoxide at a con-centration of 10% to prepare the stock solution. For testing,the stock solution was diluted with cell culture medium.
Cell culture and cell viability
Human skin fibroblast cells (CCD-986sk) were purchasedfrom American Type Culture Collection (Manassas, VA).Cells were cultured in Dulbecco’s Modified Eagle’s Med-ium (Gibco, Grand Island, NY) supplemented with antibi-otics (100 U=mL penicillin A and 100 U=mL streptomycin)and 10% heat-inactivated fetal bovine serum (Gibco). Cellswere maintained at 378C in a humidified incubator con-taining 5% CO2. The tetrazolium dye colorimetric test [3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium (MTT)test] was used to determine the viability of fibroblast cells.11
The MTT assay is based on the ability of functional mito-chondria to catalyze the reduction of MTT bromide toinsoluble formazan, the concentration of which can bemeasured spectrophotometrically.
UV irradiation
The fibroblast cells were grown in 6-cm-diameter culturedishes (SPL, Seoul), and subsequently the medium was re-placed by 2 mL of phosphate-buffered saline. Then the cellswere exposed to UV (20 mJ=cm2) light. After irradiation,the cells were washed with phosphate-buffered saline andcultured for 48 hours in serum-free medium with or withoutsamples. The control cells were maintained under the sameconditions, but without UV irradiation.
Western blot analysis
Fibroblast-conditioned medium was collected, and pro-tein concentrations were then determined with protein assayreagents (kit based on the Bradford assay [Bio-Rad La-boratories Inc., Hercules, CA]). For the western blotting,equal amount of proteins were boiled for 3 minutes andchilled on ice, subjected to 10% sodium dodecyl sulfate-polyacrylamide gel electrophoresis, and electrophoreticallytransferred to a nitrocellulose membrane (Amersham Inter-national, Little Chalfont, UK). The membranes were satu-rated with 5% powdered skim milk in TBST (10 mM Tris[pH 7.5], 100 mM NaCl, and 0.1% Tween 20), and then blotswere incubated with 1:1,000-diluted primary antibody for2 hours. After washing with TBST, the membranes wereprobed with 1:2,000-diluted secondary antibody for 1 hourand washed with TBST three times. Blotted antibody wasvisualized using a chemiluminescence (ECL) detectionsystem (Amersham International). The densities of bands
were measured by RFLPscan version 2.1 software program(Scanalytics Inc., Fairfax, VA).
Reverse transcription (RT)-polymerase chainreaction (PCR)
Total RNA was isolated with Trizol reagent (Invitrogen,Madison, WI) from human skin fibroblast cells. The oligo-nucleotide primers of MMP-1, type 1 collagen, and glycer-aldehyde 3-phosphate dehydrogenase (GAPDH) target geneswere designed using a PCR primer selection program at thewebsite of the Virtual Genomic Center (Ithaca, NY) from theGenBank database. The reaction solution (final volume,25mL) contained 0.5mL of AMV reverse transcriptase (5units), 12.5mL of AccessQuick� RT-PCR System MasterMix (2�) (Promega, Madison), and each primer at 100 pM.The total RNA was reverse-transcribed and amplified ac-cording to the manufacturer’s instructions with a thermalcycler (PerkinElmer PCR Thermal Cycler, PerkinElmer,Wellesley, MA). PCR consisted of 25 amplification cycles(948C, 30 seconds; 508C, 1 minute; 728C, 1 minute) forMMP-1 and 28 amplification cycles (948C, 30 seconds; 608C,1 minute; 728C, 1 minute) for type 1 collagen using the oli-gonucleotide primer sets detailed in Table 1. In parallel, theGAPDH housekeeping gene was amplified in each RNAsample. Reaction products were electrophoresed in 1% aga-rose gels and visualized with ethidium bromide.
Detection of reactive oxygen species (ROS) production
Fibroblast cells were pretreated with the indicatedconcentrations of sample for 24 hours, washed withphosphate-buffered saline, and stained with 50mM 20,70-dichlorofluorescein diacetate (DCFH-DA; Sigma-AldrichCo., St. Louis, MO) for 30 minutes. The cells were then ir-radiated with UV light (20 mJ=cm2) and analyzed using aFACScan� flow cytometer (FACStar�, Becton-Dickinson,San Jose, CA).
Statistics
Each experiment was performed at least in triplicate. Alldata are given mean� standard deviation (SD) values. Thedata analysis was performed using the paired-samples t test.The differences between treated and control groups werealso analyzed by analysis of variance and Duncan’s test
Table 1. Characteristics of Specific Primers Used
for RT-PCR Analysis
Gene Primers (from 50 to 30)Size(bp)
MMP-1 Forward: AGCGTGTGACAGTAAGCTAA 409Reverse: GTTTTCCTCAGAAAGAGCAGCAT
Type 1 Forward: CTCGAGGTGGACACCACCCT 366collagen Reverse: CAGCTGGATGGCCACATCGG
GAPDH Forward: ATTGTTGCCATCAATGACCC 565Reverse: AGTAGAGGCAGGGATGAT
602 SHIM ET AL.
(SPSS version 12.0, SPSS, Chicago, IL). Values of P< .05were considered statistically significant.
RESULTS
Effect on UV-induced production of MMP-1
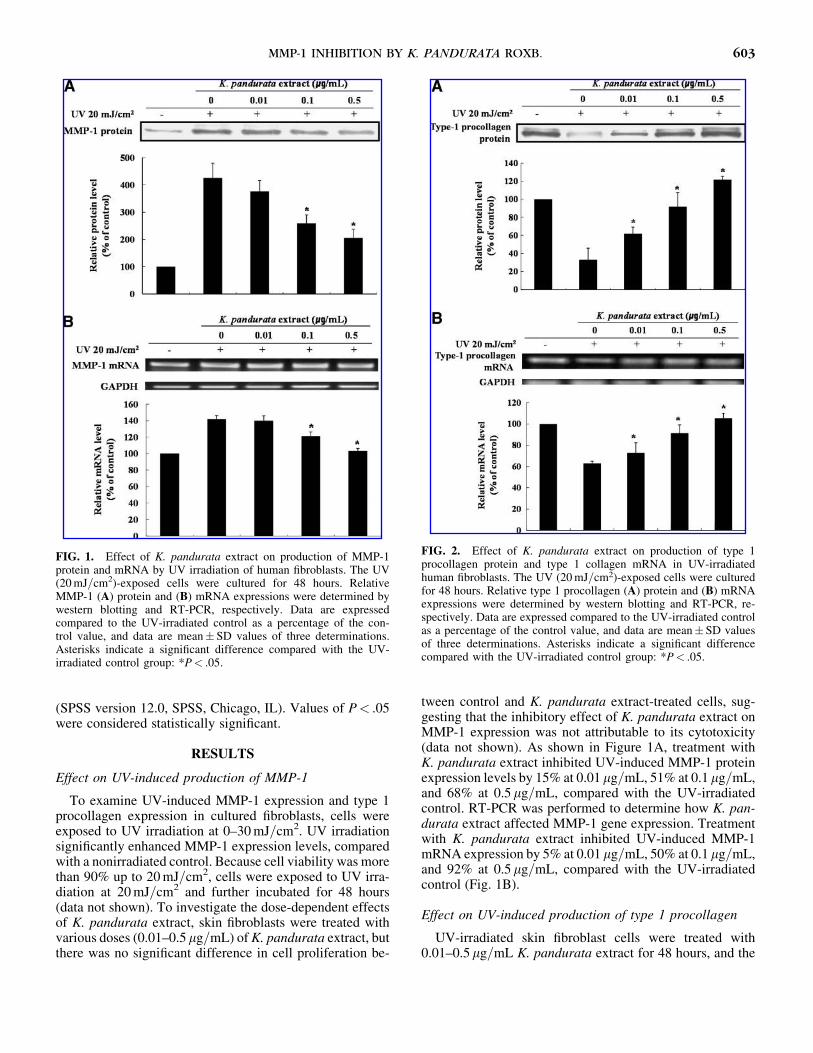
To examine UV-induced MMP-1 expression and type 1procollagen expression in cultured fibroblasts, cells wereexposed to UV irradiation at 0–30 mJ=cm2. UV irradiationsignificantly enhanced MMP-1 expression levels, comparedwith a nonirradiated control. Because cell viability was morethan 90% up to 20 mJ=cm2, cells were exposed to UV irra-diation at 20 mJ=cm2 and further incubated for 48 hours(data not shown). To investigate the dose-dependent effectsof K. pandurata extract, skin fibroblasts were treated withvarious doses (0.01–0.5mg=mL) of K. pandurata extract, butthere was no significant difference in cell proliferation be-
tween control and K. pandurata extract-treated cells, sug-gesting that the inhibitory effect of K. pandurata extract onMMP-1 expression was not attributable to its cytotoxicity(data not shown). As shown in Figure 1A, treatment withK. pandurata extract inhibited UV-induced MMP-1 proteinexpression levels by 15% at 0.01mg=mL, 51% at 0.1mg=mL,and 68% at 0.5mg=mL, compared with the UV-irradiatedcontrol. RT-PCR was performed to determine how K. pan-durata extract affected MMP-1 gene expression. Treatmentwith K. pandurata extract inhibited UV-induced MMP-1mRNA expression by 5% at 0.01mg=mL, 50% at 0.1mg=mL,and 92% at 0.5mg=mL, compared with the UV-irradiatedcontrol (Fig. 1B).
Effect on UV-induced production of type 1 procollagen
UV-irradiated skin fibroblast cells were treated with0.01–0.5mg=mL K. pandurata extract for 48 hours, and the
FIG. 1. Effect of K. pandurata extract on production of MMP-1protein and mRNA by UV irradiation of human fibroblasts. The UV(20 mJ=cm2)-exposed cells were cultured for 48 hours. RelativeMMP-1 (A) protein and (B) mRNA expressions were determined bywestern blotting and RT-PCR, respectively. Data are expressedcompared to the UV-irradiated control as a percentage of the con-trol value, and data are mean� SD values of three determinations.Asterisks indicate a significant difference compared with the UV-irradiated control group: *P< .05.
FIG. 2. Effect of K. pandurata extract on production of type 1procollagen protein and type 1 collagen mRNA in UV-irradiatedhuman fibroblasts. The UV (20 mJ=cm2)-exposed cells were culturedfor 48 hours. Relative type 1 procollagen (A) protein and (B) mRNAexpressions were determined by western blotting and RT-PCR, re-spectively. Data are expressed compared to the UV-irradiated controlas a percentage of the control value, and data are mean� SD valuesof three determinations. Asterisks indicate a significant differencecompared with the UV-irradiated control group: *P< .05.
MMP-1 INHIBITION BY K. PANDURATA ROXB. 603

type 1 procollagen protein expression levels were deter-mined in the culture medium by western blot analysis. Asshown in Figure 2A, treatment with K. pandurata extractincreased the type 1 procollagen protein expression levelsby 86% at 0.01 mg=mL, 176% at 0.1mg=mL, and 266% at0.5mg=mL, compared with the UV-irradiated control. Thelevel of type 1 collagen mRNA was analyzed by RT-PCR todetermine the regulatory activity mechanism of type 1 col-lagen production by K. pandurata extract in UV-irradiatedskin fibroblast cells. Treatment with K. pandurata extractincreased the type 1 collagen mRNA levels by 15% at0.01mg=mL, 45% at 0.1mg=mL, and 67% at 0.5 mg=mL,compared with the UV-irradiated control (Fig. 2B).
Effect on UV-induced production of intracellular ROS
To investigate whether K. pandurata extract inhibits UV-induced MMP-1 expression via an antioxidant effect, totalintracellular ROS levels were measured after K. pandurataextract treatment. As shown in Figure 3, UV irradiationmarkedly increased ROS generation compared with thecontrol; however, K. pandurata extract treatment inhibited
FIG. 3. Effect of K. pandurata extract on production of intracel-lular ROS by UV irradiation of human fibroblasts. Fibroblasts werepretreated with K. pandurata extract for 24 hours and then stainedwith 50mM DCFH-DA for 30 minutes. DCFH-DA-stained cells wereUV-irradiated (20 mJ=cm2), and 30 minutes after UV irradiation ROSlevels were analyzed using a FACScan flow cytometer. Data areexpressed compared to the UV-irradiated control as a percentage ofthe control value, and data are mean� SD values of three determi-nations. Asterisks indicate a significant difference compared with theUV-irradiated control group: *P< .05.
FIG. 4. Effect of K. pandurata extract on UV-induced phosphorylation of MAPKs. The fibroblasts were pretreated with indicated concen-trations of K. pandurata extract for 24 hours and UV-irradiated (20 mJ=cm2), and then the cells were further cultured for 1 hour. The phos-phorylation (p-) of MAPKs was detected by western blotting. Data are expressed compared to the UV-irradiated control as a percentage of thecontrol value, and data are mean� SD values of three determinations. Asterisks indicate a significant difference compared with the UV-irradiatedcontrol group: *P< .05.
604 SHIM ET AL.

UV-induced intracellular ROS levels compared with theUV-irradiated control at a concentration of 0.5mg=mL.
Effect on UV-induced phosphorylation of MAPKs
To investigate whether K. pandurata extract inhibits UV-induced MAPK (ERK, JNK, and p38) family activation,expression of phosphorylated forms of the MAPK familywere detected by western blotting. UV-irradiated fibroblastshave been shown to have elevated expressions of thephosphorylated MAPK family of enzymes. As shown inFigure 4, K. pandurata extract inhibited UV-inducedphosphorylation of ERK, JNK, and p38 kinase comparedwith the UV-irradiated control in a dose-dependent mannerwithout altering total ERK, JNK, and p38 kinase. The resultssuggested that K. pandurata extract may regulate the ex-pression of MMP-1 by UV irradiation via the ERK, JNK,and p38 kinase pathways.
Effect on UV-induced expression of c-Jun and c-Fos
We investigated the effects of K. pandurata extract onUV-induced c-Jun phosphorylation and c-Fos expression infibroblasts. Treatment with K. pandurata extract inhibitedUV-induced c-Jun phosphorylation by 15% at 0.01mg=mL,24% at 0.1mg=mL, and 37% at 0.5mg=mL, respectively,compared with the UV-irradiated control. Treatment with K.pandurata extract also inhibited c-Fos expression by 3% at0.01mg=mL, 15% at 0.1mg=mL, and 39% at 0.5mg=mL, re-spectively, compared with the UV-irradiated control (Fig. 5).
DISCUSSION
Up-regulation of MMP-1 is responsible for the enhanceddegradation of dermal collagen during chronological andUV-induced skin aging. With increasing age, collagensynthesis decreases, and MMP-1 levels increase in humanskin.12 Therefore, the regulation of MMP-1 activity mightbe a potential strategy for prevention and treatment ofUV-induced skin damage. In our study, it was shown thatK. pandurata extract at concentrations of 0.01–0.5mg=mL,which was not toxic to human skin fibroblast cells in vitro,significantly decreased MMP-1 expression and increasedtype 1 procollagen expression as shown by protein andmRNA levels in a dose-dependent manner (Figs. 1 and 2).
Traditional herbal medicines provide an interesting, lar-gely unexplored source for development of new drugs. Thepotential use of traditional herbal medicines for devel-opment of new skin care cosmetics has been emphasizedrecently.13 Natural products from medicinal plants are po-tential sources as MMP-1 inhibitors and collagen synthe-sizers. For example, green tea,14 pomegranate fractions,15
Camellia japonica oil,16 and rosemary,17 among others, werereported to regulate MMP-1 and type 1 collagen expressionlevels. However, these natural sources were usually exam-ined at high concentrations in the ranges of 10–1,000mg=mLand had little effect on MMP-1 and type 1 collagen expres-sion levels.
UV light is particularly associated with oxidative pro-cesses involved in photoaging. ROS generation is initiatedfollowing UV irradiation, which results in the up-regulation
FIG. 5. Effect of K. pandurata extract onUV-induced expression of c-Jun and c-Fos.The fibroblasts were pretreated with indicatedconcentrations of K. pandurata extract for24 hours and irradiated with UV (20 mJ=cm2),and then the cells were further cultured for2 hours. The phosphorylation (p-)=expressionof c-Jun and c-Fos was detected by westernblotting. Data are expressed compared to theUV-irradiated control as a percentage of thecontrol value, and data are mean� SD valuesof three determinations. Asterisks indicatea significant difference compared with theUV-irradiated control group: *P< .05.
MMP-1 INHIBITION BY K. PANDURATA ROXB. 605

of MMP-1 and the degradation of dermal collagen.18 ROSgeneration also plays a critical role in the MAPK-mediatedsignal transduction triggered by UV light. In this study, itwas found that K. pandurata extract only at a concentrationof 0.5mg=mL decreased intracellular ROS levels (Fig. 3).Based on these findings, the regulatory activity of K. pan-durata extract on MMP-1 and type 1 procollagen expressionlevels might be attributable in part to inhibition of ROSgeneration in UV-irradiated fibroblast cells. Therefore, theMMP-1 inhibitory effect shown by K. pandurata extract isnot mediated by its antioxidant effect and may be mediatedby the inhibition of the intracellular signal transduction re-sponse.
Signaling pathways involving MAPKs play importantroles in regulating gene expression, cell proliferation, cellmotility, and cell survival and death in eukaryotic cells.Three distinct groups of MAPKs have been identified inmammalian cells: JNKs, ERKs, and p38 kinase.19 A previ-ous study demonstrated that UV irradiation activates theMAPK cascade and that this activation is then succeeded byan increase in the expression of c-Jun and c-Fos, which formthe AP-1 complex.20 Based on these findings, K. pandurataextract inhibits UV-induced MMP-1 expression in fibro-blasts, and this inhibitory effect of K. pandurata extracton MMP-1 expression may be mediated by inhibition ofthe ERK-, JNK-, and p38 kinase-dependent pathways(Fig. 4).
The downstream effectors of the MAPKs include severaltranscription factors, such as c-Fos and c-Jun. The tran-scription of MMPs is regulated by AP-1, which is one ofseveral transcription factors activated by UV light.21 TheAP-1 complex is composed of Jun and Fos family members,and Jun proteins form either a homodimer or a heterodimerwith Fos protein to produce the AP-1 complex.22 In thisstudy, we found that K. pandurata extract treatment in-hibited UV-induced c-Jun phosphorylation and c-Fos ex-pression (Fig. 5).
Taken together with previous data, the present studydemonstrates that K. pandurata extract significantly inhibitsUV-induced MMP-1 expression by suppressing UV-inducedc-Jun phosphorylation and c-Fos expression, which may bemediated by inhibition of ERK, JNK, and p38 kinase path-ways in cultured fibroblasts. These results indicate thatK. pandurata extract can be employed as a potential candi-date for the prevention and treatment of skin aging. Isolationand structural elucidation of the active constituents fromK. pandurata extract will provide useful leads in the devel-opment of anti-aging agents.
ACKNOWLEDGMENTS
This work was supported partly by the Yonsei Biomole-cule Research Initiative of the Brain Korea 21 Project.
AUTHOR DISCLOSURE STATEMENT
No competing financial interests exist.
REFERENCES
1. Fisher GJ, Datta SC, Talwar HS, Wang ZQ, Varani J, Kang S,
Voorhees JJ: Molecular basis of sun-induced premature skin
aging and retinoid antagonism. Nature 1996;379:335–339.
2. Moon HI, Lee JK, Zee OP, Chung JH: The effect of flavonol
glycoside on the expressions of matrix metalloproteinase-1 in
ultraviolet-irradiated cultured human skin fibroblasts. J Ethno-
pharmacol 2005;101:176–179.
3. Vincenti MP, Brinckerhoff CE: Transcriptional regulation of
collagenase (MMP-1, MMP-13) genes in arthritis: integration of
complex signaling pathways for the recruitment of gene-specific
transcription factors. Arthritis Res 2002;4:157–164.
4. Brenneisen P, Wenk J, Klotz LO, Wlaschek M, Briviba K, Krieg
T, Seis H, Scharffetter-Kochanek K: Central role of ferrous=ferric
iron in the ultraviolet B irradiation-mediated signaling pathway
leading to increased interstitial collagenase (matrix-degrading
metalloprotease (MMP)-1) and stromelysin-1 (MMP-3) mRNA
levels in cultured human dermal fibroblasts. J Biol Chem 1998;
273:5279–5287.
5. Varani J, Warner RL, Gharaee-Kermani M, Phan SH, Kang S,
Chung JH, Wang ZQ, Datta SC, Fisher GJ, Voorhees JJ: Vitamin A
antagonizes decreased cell growth and elevated collagen-degrading
matrix metalloproteinases and stimulates collagen accumulation in
naturally aged human skin. J Invest Dermatol 2000;114:480–486.
6. Xu Y, Fisher GJ: Ultraviolet (UV) light irradiation induced signal
transduction in skin photoaging. J Dermatol Sci Suppl 2005;1:
S1–S8.
7. Huang C, Schmid PC, Ma WY, Schmid HH, Dong Z: Phos-
phatidylinositol-3 kinase is necessary for 12-O-tetra-decanoyl-
phorbol-13-acetate-induced cell transformation and activated
protein 1 activation. J Biol Chem 1997;272:4187–4194.
8. Tuchinda P, Reutrakul V, Claeson P, Pongprayoon U, Sematong
T, Santisuk T: Anti-inflammatory cyclohexenyl chalcone deriv-
atives in Boesenbergia pandurata. Phytochemistry 2002;59:169–
173.
9. Song MS, Shim JS, Gwon SH, Lee CW, Kim HS, Hwang JK:
Antibacterial activity of panduratin A and isopanduratin A iso-
lated from Kaempferia pandurata Roxb. against acne-causing
microorganisms. Food Sci Biotechnol 2008;17:1357–1360.
10. Shim JS, Han YS, Hwang JK: The effect of 4-hydroxypanduratin
A on the mitogen-activated protein kinase-dependent activation
of matrix metalloproteinase-1 expression in human skin fibro-
blasts. J Dermatol Sci 2009;53:129–134.
11. Mosmann T: Rapid colorimetric assay for cellular growth and
survival: application to proliferation and cytotoxicity assays.
J Immunol Methods 1983;65:55–63.
12. Soriani M, Rice-Evans C, Tyrrell RM: Modulation of the UVA
activation of heme oxygenase, collagenase, and cyclooxygenase
gene expression by epigallocatechin in human skin cells. FEBS
Lett 1998;439:253–257.
13. Kiken DA, Cohen DE: Contact dermatitis to botanical extracts.
Am J Contact Dermatol 2002;13:148–152.
14. Katiyar SK, Elmets CA: Green tea polyphenolic antioxidants and
skin photoprotection. Int J Oncol 2001;18:1307–1313.
15. Muhammad NA, Ephraim PL, Varani J: Pomegranate as a cos-
meceutical source: pomegranate fractions promote proliferation
and procollagen synthesis and inhibit matrix metalloproteinase-1
production in human skin cells. J Ethnopharmacol 2006;103:
311–318.
606 SHIM ET AL.

16. Jung ES, Lee JS, Baek JH, Jung KS, Lee JY, Huh SR, Kim SB,
Koh JS, Park DH: Effect of Camellia japonica oil on human type
1 procollagen production and skin barrier function. J Ethno-
pharmacol 2007;112:127–131.
17. Tokuda H, Ohigashi H, Koshimizu K, Ito Y: Inhibitory effects
of ursolic acid and oleanic acid on skin tumor promotion by 12-
O-tetradecanoylphorbol-13-acetate. Cancer Lett 1986;33:279–
285.
18. Stetler-Stevenson WG, Yu AE: Proteases in invasion: matrix
metalloproteinase. Semin Cancer Biol 2001;11:143–152.
19. Robinson MJ, Cobb MH: Mitogen-activated protein kinase
pathways. Curr Opin Cell Biol 1997;9:180–186.
20. Rittie L, Fisher GJ: UV-light-induced signal cascades and skin
aging. Ageing Res Rev 2002;1:705–720.
21. Hirano T, Higa S, Arimitsu J, Naka T, Okada A, Shima Y: Lu-
teolin, a flavonoid, inhibits AP-1 activation by basophils. Bio-
chem Biophys Res Commun 2006;340:1–7.
22. Waskiewicz AJ, Cooper JA: Mitogen and stress response path-
ways: MAP kinase cascades and phosphatase regulation in
mammals and yeast. Curr Opin Cell Biol 1995;7:798–805.
MMP-1 INHIBITION BY K. PANDURATA ROXB. 607

This article has been cited by:
1. Soo Hwa Kim, Hye-Sook Seo, Bo-Hyoung Jang, Yong-Cheol Shin, Seong-Gyu Ko. 2014. The effect of Rhus verniciflua Stokes(RVS) for anti-aging and whitening of skin. Oriental Pharmacy and Experimental Medicine . [CrossRef]
2. Azadeh Manayi, Soodabeh Saeidnia, Ahmad Reza Gohari, Mohammad Abdollahi. 2014. Methods for the discovery of new anti-aging products – targeted approaches. Expert Opinion on Drug Discovery 9:4, 383-405. [CrossRef]
3. Sungkyung Kim, Hyun-In Oh, Jae-Kwan Hwang. 2012. Oral administration of fingerroot (Boesenbergia pandurata) extractreduces ultraviolet b-induced skin aging in hairless mice. Food Science and Biotechnology 21:6, 1753-1760. [CrossRef]
4. Pulok K. Mukherjee, Niladri Maity, Neelesh K. Nema, Birendra K. Sarkar. 2011. Bioactive compounds from natural resourcesagainst skin aging. Phytomedicine . [CrossRef]
5. Junoh Kim, Chan-Woo Lee, Eun Kyung Kim, Su-Jin Lee, Nok-Hyun Park, Han-Sung Kim, Han-Kon Kim, KookheonChar, Young Pyo Jang, Jin-Woong Kim. 2011. Inhibition effect of Gynura procumbens extract on UV-B-induced matrix-metalloproteinase expression in human dermal fibroblasts. Journal of Ethnopharmacology 137:1, 427-433. [CrossRef]
6. Chen Zhang, Yong Li, Zhong-Ji Qian, Sang-Hoon Lee, Yong-Xin Li, Se-kwon Kim. 2011. Dieckol from Ecklonia cava RegulatesInvasion of Human Fibrosarcoma Cells and Modulates MMP-2 and MMP-9 Expression via NF-κB Pathway. Evidence-BasedComplementary and Alternative Medicine 2011, 1-8. [CrossRef]





























