Embed Size (px)
Citation preview

ORIGINAL ARTICLE Andrology
Mast cell tryptase stimulatesproduction of decorin by humantesticular peritubular cells: possiblerole of decorin in male infertility byinterfering with growth factor signalingM. Adam1, J.U. Schwarzer2, F.M. Kohn3, L. Strauss4, M. Poutanen4,and A. Mayerhofer1,*1Anatomy and Cell Biology, Center for Integrated Protein Science Munich (CIPSM) Ludwig-Maximilians-University Munich, D-80802 Munich,Germany 2Praxis Urology-Andrology, D-85356 Freising, Germany 3Andrologicum, D-80331 Munich, Germany 4Department of Physiologyand Turku Center for Disease Modeling, Institute of Biomedicine, University of Turku, 20520 Turku, Finland
*Correspondence address. Fax: +49-89-397035; Email: [email protected]
Submitted on January 27, 2011; resubmitted on June 15, 2011; accepted on June 27, 2011
background: Myofibroblastic, peritubular cells in the walls of seminiferous tubules produce low levels of the extracellular matrix (ECM)protein decorin (DCN), which has the ability to interfere with growth factor (GF) signaling. In men with impaired spermatogenesis, fibroticremodeling of these walls and accumulation of tryptase-positive mast cells (MCs) occur.
methods: Human testicular biopsies with normal and focally impaired spermatogenesis (mixed atrophy) were subjected to immunohis-tochemistry and laser micro-dissection followed by RT–PCR. Primary human testicular peritubular cells (HTPCs), which originate fromnormal and fibrotically altered testes (HTPC-Fs), were studied by qRT–PCR, western blotting, enzyme-linked immunosorbent assaymeasurements and Ca2+ imaging. Phosphorylation and viability/proliferation assays were performed.
results: Immunohistochemistry revealed DCN deposits in the walls of tubules with impaired spermatogenesis. Mirroring the situation invivo, HTPC-Fs secreted more DCN than HTPCs (P , 0.05). In contrast to HTPCs, HTPC-Fs also responded to the main MC product, tryp-tase, and to a tryptase receptor (PAR-2) agonist by further increased production of DCN (P , 0.05). Several GF receptors (GFRs) areexpressed by HTPCs and HTPC-Fs. DCN acutely increased intracellular Ca2+-levels and phosphorylated epidermal GF (EGFR) withinminutes. Platelet-derived GF (PDGF) and EGF induced strong mitogenic responses in HTPC/-Fs, actions that were blocked by DCN,suggesting that DCN in the ECM interferes with GF/GFRs signaling of peritubular cells of the human testis.
conclusions: The data indicate that the increase in testicular DCN found in male infertility is a consequence of actions of MC-derivedtryptase. We propose that the increases in DCN may consequently imbalance the paracrine signaling pathways in human testis.
Key words: testis / decorin / paracrine / proliferation / fibrosis
IntroductionFlat, spindle-like peritubular cells and the extracellular matrix (ECM)build a wall, which separates the germinal epithelium of the seminifer-ous tubules from the interstitial compartment of the testis. The testi-cular peritubular cells (TPC) of this wall are myofibroblastic cells(Cigorraga et al., 1994), which form several layers in humans(HTPC) and are thought to be important for the transport of spermand testicular fluid (Rossi et al., 2002; Romano et al., 2005; Zhanget al., 2006; Welsh et al., 2009; Schell et al., 2010). In addition,
peritubular cells secrete a number of paracrine factors, participatingin the control of testicular functions (Schell et al., 2008, 2010; Spinnleret al., 2010).
Testicular peritubular cells are assumed to also secrete componentsof the ECM of the peritubular wall (Maekawa et al., 1996). Surprisingly,the full spectrum of these proteins and the exact composition of theECM of the peritubular wall in man have not been defined but type IVcollagen, heparin sulfate proteoglycan, fibronectin and laminin havebeen observed in ECM (Santoro et al., 2000; Dobashi et al., 2003).Of these, collagen type IV is a product of HTPC, and so is the
& The Author 2011. Published by Oxford University Press on behalf of the European Society of Human Reproduction and Embryology. All rights reserved. For Permissions, please email:[email protected]
Human Reproduction, Vol.26, No.10 pp. 2613–2625, 2011
Advanced Access publication on July 25, 2011 doi:10.1093/humrep/der245
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018

proteoglycan decorin (DCN; Ungefroren et al., 1995). Ungefrorenet al. (1995) located DCN gene expression to cells of the peritubularwall and interstitial fibroblastic cells of normal adult human testes.Interestingly, in other species, testicular DCN has not been examinedor is absent in developing mouse gonads (Miqueloto and Zorn, 2007).
DCN belongs to the family of secreted small leucine-rich proteogly-cans. Originally the name was given because it can ‘decorate’ collagenfibrils, thereby regulating collagen fibrillogenesis—a key mechanism ofECM assembly and homeostasis (Reed and Iozzo, 2002; Zhu et al.,2005; Schaefer and Iozzo, 2008).
In addition, two other actions of DCN have been recognized,linking DCN to fundamental biological functions, including cellmigration, adhesion and proliferation (Nili et al., 2003; Ferdouset al., 2010). First, DCN binds a variety of soluble and insolubleligands, thus regulating their activity and availability. This is welldocumented, for instance, for transforming growth factor-beta(TGF-b; Macri et al., 2007) and platelet-derived GF (PDGF; Niliet al., 2003). Second, DCN itself can act as a non-selectiveligand for several growth factor receptors (GFRs), including epider-mal GFR (EGFR; Schaefer and Iozzo, 2008), insulin-like GFR 1(IGF1R; Schaefer and Iozzo, 2008), vascular endothelial GFR(VEGFR; Iacob et al., 2008) and hepatocyte GFR (HGFR; Goldoniet al., 2009). It appears that DCN may cause initial activation ofthe respective signaling pathway, followed by its inactivation asshown, for example, for EGFR (Zhu et al., 2005).
Could DCN interfere with GF/GFR signaling also in the humantestis and in the peritubular wall? The answer to this question couldbe of potential interest for the understanding of testicular develop-ment (Mariani et al., 2002; Puglianiello et al., 2004; Ricci et al.,2004; Nurmio et al., 2007; Basciani et al., 2010), for the understandingof the adult human testis and for human male infertility (Nakazumiet al., 1996). In human male infertility, striking alterations of the cellularcomponents of the peritubular wall and of the ECM (Davidoff et al.,1990; Haider et al., 1999; Gulkesen et al., 2002) are evident;however, surprisingly little is known about this remodeling process.We recently reported hypertrophy and loss of contractility markersin peritubular cells of men with impaired spermatogenesis, indicatinga switch of the cellular phenotype of peritubular cells from contractilecells to synthesizing cell types (Schell et al., 2010). Whether this switchmay also affect the secretion of DCN by peritubular cells (Ungefrorenet al., 1995) is not known but in a number of other human fibroticremodeling processes (e.g. pulmonary and nephrogenic systemic fibro-sis; van Straaten et al., 1999; Fadic et al., 2006; Gambichler et al.,2009), increases in DCN production were reported.
What induces the fibrotic remodeling, called a hallmark of humanmale infertility (De Kretser and Baker, 1996), remains unknown butmast cells (MCs) and macrophages may be involved via their directand indirect products. This assumption is based on observations invivo and on results of cell culture experiments. It is established nowthat many more immune cells are present in the peritubular wall intestes with fibrotic changes (Meineke et al., 2000; Frungieri et al.,2002a,b; Spiess et al., 2007; Matzkin et al., 2010). The secreted pro-ducts of MCs and macrophages, namely tryptase and tumor necrosisfactor alpha (TNF-a), and also a prostaglandin metabolite, have beenshown to regulate the cellular phenotype and secretory behavior ofhuman fibroblasts and/or of HTPCs (Frungieri et al., 2002a; Albrechtet al., 2005; Schell et al., 2008).
The successful isolation of HTPCs, and recently also of the corre-sponding cells from testes of men with existing fibrotic changes(HTPC-Fs; Spinnler et al., 2010), has opened a door to the examin-ation of the role of this unexplored human testicular cell type. Wemade use of these unique models for human testicular cells in thepresent study. HTPCs and HTPC-Fs are comparable in mostaspects and both produce DCN and, as we found, both are targetsfor DCN, which acts as a ligand for GF receptors. Yet, HTPCs andHTPC-Fs differ with respect to the levels of DCN production andits regulation, and thus mirror the in vivo situation of the humantestis in health and disease.
Materials and Methods
Isolation of HTPC/HTPC-Fs, cell cultureand human testicular samplesIsolation of HTPC/-Fs was performed as described previously (Albrechtet al., 2006; Schell et al., 2008, 2010). HTPCs originate from patients dis-playing normal spermatogenesis, whereas HTPC-Fs were isolated frommen suffering from impaired spermatogenesis and testicular fibrosis. Allparticipants granted written informed consent. The local ethics committeeapproved the study.
All cells, passages 3–12, were maintained in Dulbecco’s modifiedEagle’s medium (DMEM) supplemented with 10% fetal calf serum (FCS;both from PAA GmbH, Colbe, Germany), unless indicated otherwise.After initial screening with several concentrations in order to determineoptimal response, treatments with 100 ng/ml human recombinant(rhSkin b) tryptase (Promega GmbH, Mannheim, Germany) and 10 mMprotease activated receptor (PAR-2) agonist peptide SLIGKV (NeoMPS,Strasbourg, France) were performed for 72 h, as detailed later. Solvent(H2O) for these substances was included as control. Cells were culturedin the presence of 10 mg/ml DCN (human recombinant, ImmundiagnostikAG, Bensheim, Germany or bovine articular cartilage, Sigma Aldrich, Dei-senhofen, Germany), 50 ng/ml EGF (human recombinant, New EnglandBiolabs, Frankfurt, Germany) or/and 5 ng/ml PDGF-BB (the BB homodi-meric form, human recombinant, Sigma) for 24 h, as described later.These concentrations were chosen based on the results of dose–responsetests (not shown). PDGF-BB was dissolved in 4 mM HCl containing 0.1%bovine serum albumin (BSA; stock; PAA GmbH) and was used at a 1:1000dilution (4 mM). Hence the stock solution without PDGF was added to themedia as a control.
For immunohistochemical studies, human testicular samples from menwith normal or pathologically altered spermatogenesis were used, asdescribed previously (Meineke et al., 2000; Frungieri et al., 2002a,b;Albrecht et al., 2006; Schell et al., 2010). We focused on samples ofmixed atrophy (MA) patients, which can show unaltered peritubularwalls as well as focally altered areas in the same sections.
Immunohistochemistry andimmunocytochemistryImmunohistochemistry and immunofluorescence methods were per-formed, as previously described (Albrecht et al., 2006; Schell et al.,2008, 2010). A monoclonal goat anti-human DCN antibody (1:100;R&D Systems, Wiesbaden-Nordenstadt, Germany) was used to staindeparaffinized 5-mm-thick sections. Cells from at least three differentpatients (HTPC-Fs and HTPCs) were used for each immunofluorescenceexperiment. Monoclonal mouse antibodies against smooth muscle actin(SMA; 1:2000; clone 1A4, Sigma), CD90 (1:100; clone AS02; DianovaGmbH, Hamburg; Germany) and collagen type I (1:200; clone I-8H5;
2614 Adam et al.
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018

Acris Antibodies GmbH; Hiddenhausen, Germany) were employed. Incu-bation with normal non-immune serum instead of the specific antibody, oromission of the primary antibody, served as controls.
Isolation of RNA, RT–PCR, semi-quantitativeRT–PCR and quantitative (q)RT–PCRHTPCs and HTPC-Fs from at least three different donors were grown tosub-confluence and incubated with/without tryptase (100 ng/ml) orSLIGKV (10 mM) in DMEM medium containing 10% FCS for 72 h. Sub-sequently cells were washed twice with DMEM without FCS and sus-pended in RLT-buffer (QIAGEN GmbH, Hilden, Germany) containing1% b-mercaptoethanol.
QIAGEN RNeasy minikit was used for RNA-isolation, followed byreverse transcription using random hexamer primers. Four hundred nano-grams of total RNA were used for further semi-quantitative RT–PCRexperiments (Frungieri et al., 2002a; Schell et al., 2010). For informationabout primers, annealing temperatures and product sizes, see Schellet al. (2010) and Table I. PCR products were visualized by ethidiumbromide staining in agarose gels. As a positive control, commercially avail-able human testes cDNA (Clontech, Palo Alto, CA, USA) was used. Iden-tities of all PCR products were verified by sequencing (Schell et al., 2010).
Synthesis of cDNAs for qRT–PCR was performed by using theDyNAmo two-step SYBR Green qRT–PCR kit (Finnzymes Oy, Espoo,Finland) and the Engine Opticon system (MJ Research, Inc., Waltham,MA, USA) for continuous fluorescent detection was used. Samples wereanalyzed in triplicate. The expression levels were determined in pro-portion to the ribosomal protein L19 (RPL19) housekeeping gene (Liet al., 2006; Strauss et al., 2009). Cells from at least three patients ineach group were examined.
Western blottingHTPC/-Fs grown on 60-mm dishes (Nunc GmbH & Co. KG, Wiesbaden,Germany) were incubated in the presence or absence of tryptase (100 ng/ml) or SLIGKV (10 mM) in DMEM medium containing 10% FCS for 72 h.We performed immunoblots, as described in previous studies (Frungieriet al., 2002a), using the same monoclonal DCN (1:1,000, R&D Systems)and SMA (1:500; Sigma) antibodies as for immunohistochemistry. Theuse of a beta-actin antibody (1:5000, Sigma) allowed corrections forsmall differences in loading. Western blot bands were detected with che-miluminescent reagents (SuperSignalw West Femto Maximum SensitivitySubstrate, Pierce, Thermo Scientific, Rockford, IL, USA) and analyzed den-sitometrically with ImageJ Software (National Institutes of Health,Bethesda, MD, USA; version 1.40g) and the results normalized to thoseobtained for b-actin. We repeated the experiments with cells from atleast three different patients of each group, HTPCs and HTPC-Fs, andalso used four freshly obtained biopsy samples.
Enzyme-linked immunosorbent assayDCN was detected in conditioned media by using a commercial immu-noassay (R&D Systems) according to the manufacturer’s protocol(100 ml of the conditioned media diluted 1:1000). The medium super-natants (total 2 ml per 60-mm dish) of the same tryptase/SLIGKV-treatedand untreated control cells that were used in the western blot exper-iments, were examined. The results were expressed as secreted DCNper mg total cellular protein.
Phosphorylation assays: screening usingproteome profiler and western blottingA total of 106 cells (HTPCs) were seeded on a 60-mm dish and treated for5 min with 10 mg/ml DCN, i.e. a concentration, which induced robust
increases in intracellular Ca2+. For controls, phosphate-buffered saline(PBS) was added to the culture medium. Cellular protein was extractedand used in an antibody-based array for human phospho-receptortyrosin kinases (RTKs), according to the manufacturer’s protocol (Pro-teome Profiler; R&D Systems; IGEPAL, Sigma). In brief, cell lysates wereprepared with a Nonidet P-40 lysis buffer, added to nitrocellulose mem-branes on which antibodies were spotted, and incubated overnight at48C. After washing steps, membranes were incubated with ananti-phospho-tyrosine-horse-radish peroxidase detection antibody for2 h at room temperature on a rocking platform shaker. The wash stepwas repeated. Chemiluminiscent reagent (Piercew ECL Western BlottingSubstrate, Thermo Scientific) was added and images were captured.Results were evaluated with ImageJ Software. The experiment wasrepeated twice.
The ability of DCN, EGF and PDGF-BB to phosphorylate EGFR andPDGFR-b was further studied by western blotting. HTPC/-Fs were incu-bated with 10 mg/ml DCN, 50 ng/ml EGF or 5 ng/ml PDGF-BB for10 min in DMEM without FCS. To exclude non-specific protein effects,cells from each patient were treated with 10 mg/ml BSA (pH ¼ 7.8;PAA GmbH). Other control cells were incubated in the presence ofPBS. Immunoblotting was performed using 0.5 mg/ml rabbitanti-phospho-PDGFR-b (1:100) or 0.4 mg/ml anti-phospho-EGFR sol-ution (1:250; antibodies were identical to the ones used in the ProteomeProfiler assay). For control purposes, monoclonal rabbit anti-human EGFRantibody (1:100; Thermo Fisher Scientific Inc., Fremont, CA, USA) andpolyclonal rabbit anti-human PDGFR antiserum (1:200; Santa Cruz Bio-technology Inc., Santa Cruz, CA, USA) were also used.
Measurements of intracellular Ca21
Intracellular Ca2+ levels were measured as described in previous studies(Fohr et al., 1993; Albrecht et al., 2006). In brief, cells were grown oncover slips for 24 h in DMEM supplemented with 10% FCS, loaded with5 mM fluo-4, acetoxy-methylester (fluo-4AM; Molecular Probes, Eugene,OR, USA) for 30 min at 378C and 5% CO2. After washing the cells withFCS-free DMEM, the cover slip was transferred into a recordingchamber mounted on a TCS SP2 confocal microscope (Leica Microsys-tems, Bensheim, Germany). Fluorescence was monitored at 500–540 nm (lex ¼ 488 nm) every 2 s and the intensity was quantified oversingle cells. To assess whether HTPC/-Fs may possess functionalPAR-2, changes of intracellular Ca2+ levels were recorded on-line whileapplying 100 ng/ml tryptase or 10 mM agonist peptide SLIGKV to cells(of at least three different patients for each group). In pilot experiments,10 mg/ml bovine articular cartilage DCN, as well as human recombinantDCN, were compared. No difference in the ability of the DCN prep-aration to mobilize intracellular Ca2+ was observed. Subsequently,unless indicated otherwise, bovine DCN was generally used but foreach experiment human recombinant DCN was tested as well tocontrol for any possible difference in actions. Experiments evaluating theactions of different concentrations of EGF (50 ng/ml, 250 ng/ml and1 mg/ml) and PDGF-BB (5 and 10 ng/ml) were also conducted. Inadditional experiments, cells (n ¼ 3 HTPCs and n ¼ 2 HTPC-Fs) werepretreated with 10 mg/ml DCN for 24 h before measuring changes inCa2+ levels with acute stimulation by EGF, PDGF or DCN. In addition,the response to histamine (100 mM) was tested (see Albrecht et al.,2006). Measurements of Ca2+ levels after adding 10 ml/ml BSA (pH ¼7.8) were performed to exclude non-specific protein effects.
Cell viability and proliferationViability of cells was assessed using the CellTiter-Glow Luminescent CellViability Assay (Promega GmbH, Mannheim, Germany). Cells wereseeded in quadruplicate in 24-well tissue culture plates and incubated
Decorin and human male infertility 2615
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
.............................................................................................................................................................................................
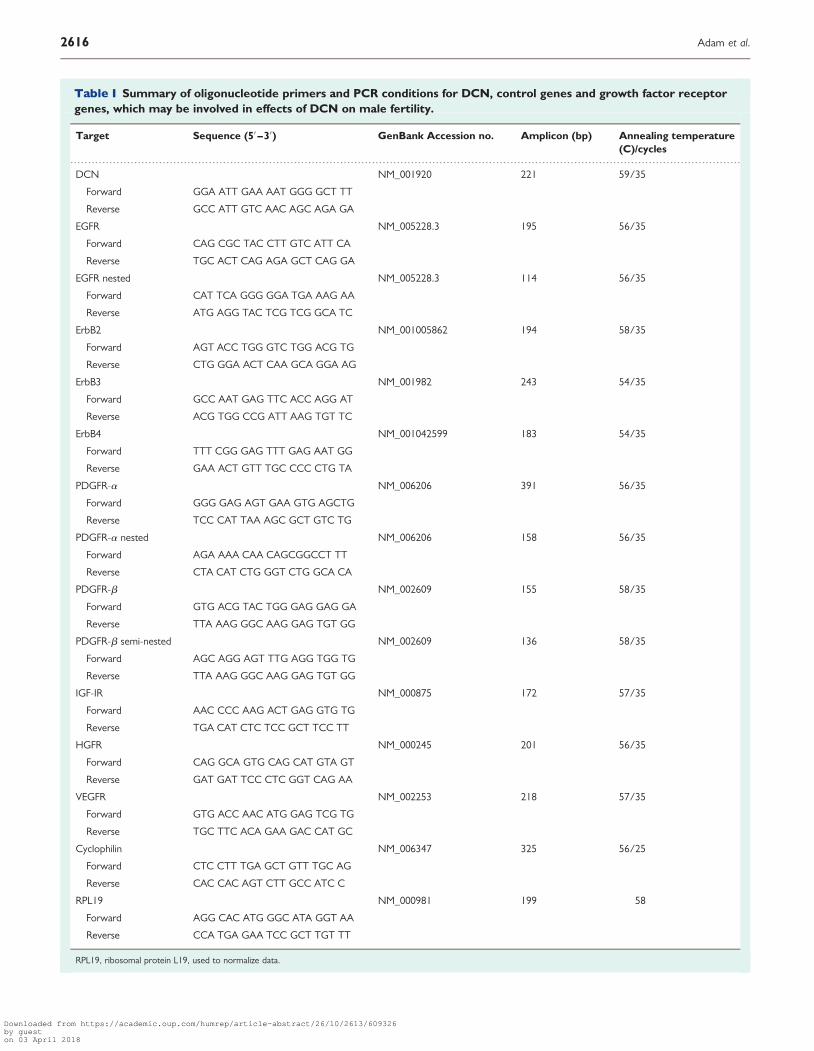
Table I Summary of oligonucleotide primers and PCR conditions for DCN, control genes and growth factor receptorgenes, which may be involved in effects of DCN on male fertility.
Target Sequence (5′ –3′) GenBank Accession no. Amplicon (bp) Annealing temperature(C)/cycles
DCN NM_001920 221 59/35
Forward GGA ATT GAA AAT GGG GCT TT
Reverse GCC ATT GTC AAC AGC AGA GA
EGFR NM_005228.3 195 56/35
Forward CAG CGC TAC CTT GTC ATT CA
Reverse TGC ACT CAG AGA GCT CAG GA
EGFR nested NM_005228.3 114 56/35
Forward CAT TCA GGG GGA TGA AAG AA
Reverse ATG AGG TAC TCG TCG GCA TC
ErbB2 NM_001005862 194 58/35
Forward AGT ACC TGG GTC TGG ACG TG
Reverse CTG GGA ACT CAA GCA GGA AG
ErbB3 NM_001982 243 54/35
Forward GCC AAT GAG TTC ACC AGG AT
Reverse ACG TGG CCG ATT AAG TGT TC
ErbB4 NM_001042599 183 54/35
Forward TTT CGG GAG TTT GAG AAT GG
Reverse GAA ACT GTT TGC CCC CTG TA
PDGFR-a NM_006206 391 56/35
Forward GGG GAG AGT GAA GTG AGCTG
Reverse TCC CAT TAA AGC GCT GTC TG
PDGFR-a nested NM_006206 158 56/35
Forward AGA AAA CAA CAGCGGCCT TT
Reverse CTA CAT CTG GGT CTG GCA CA
PDGFR-b NM_002609 155 58/35
Forward GTG ACG TAC TGG GAG GAG GA
Reverse TTA AAG GGC AAG GAG TGT GG
PDGFR-b semi-nested NM_002609 136 58/35
Forward AGC AGG AGT TTG AGG TGG TG
Reverse TTA AAG GGC AAG GAG TGT GG
IGF-IR NM_000875 172 57/35
Forward AAC CCC AAG ACT GAG GTG TG
Reverse TGA CAT CTC TCC GCT TCC TT
HGFR NM_000245 201 56/35
Forward CAG GCA GTG CAG CAT GTA GT
Reverse GAT GAT TCC CTC GGT CAG AA
VEGFR NM_002253 218 57/35
Forward GTG ACC AAC ATG GAG TCG TG
Reverse TGC TTC ACA GAA GAC CAT GC
Cyclophilin NM_006347 325 56/25
Forward CTC CTT TGA GCT GTT TGC AG
Reverse CAC CAC AGT CTT GCC ATC C
RPL19 NM_000981 199 58
Forward AGG CAC ATG GGC ATA GGT AA
Reverse CCA TGA GAA TCC GCT TGT TT
RPL19, ribosomal protein L19, used to normalize data.
2616 Adam et al.
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
for 24 h with/without DCN (10 mg/ml). The kit reagents were addeddirectly to the cells, and luminescence of the luciferase reaction wasmeasured in a FLUOstar Optima (BMG Labtech GmbH, Offenburg,Germany) reader. Cells from five men with normal testis and five patientswith fertility problems were used.
To evaluate mitotic figures of cultured HTPC/-Fs, DAPI(4′,6′-diamidino-2′-phenylindole; 1.5 mg/ml; Vectashield mountingmedium, Vector laboratories, Burlingame, CA, USA) was used, asdescribed (Kunz et al., 2006). For this study, HTPCs and HTPC-Fs from3 to 12 different men were used. Different time points (6, 18 and 24 h)were studied. Only at 24 h was a significant increase of proliferativeevents in treatment groups over untreated controls observed. At 20×magnification, all nuclei (.200/slide) were counted and the percentagesof metaphase, anaphase and telophase figures were determined.
Laser capture microdissectionSections (5 mm) of human-paraffin-embedded biopsies were mountedonto thin polyethylene naphthalene membrane, as previously described(Frungieri et al., 2002a, b). The sections were deparaffinized, lightlystained with hamalaune/eosin and subjected to laser capture microdissec-tion (LMD; P.A.L.M. GmbH Mikrolaser Technologie, Bernried, Germany).This allowed the isolation of peritubular walls from surrounding tissue. Thesamples were kept frozen (2808C) until RT–PCR was performed. RNAwas extracted using the RNeasy FFPE kit (QIAGEN) and subjected to RT–PCR using oligo-dT primers. In order to ensure that the sections containedsuitable RNA for this procedure, a whole deparaffinized consecutivesection was also scraped from the slide and processed as described. Forinformation on primers and PCR conditions (see Table I). Amplified pro-ducts were sequenced.
Data analysis and statisticsResults obtained were analyzed using PRISM 4.0 (GraphPad Software, Inc.,San Diego, CA, USA). t-Test or one-way analysis of variance (ANOVA)tests were performed, as indicated. Differences between the groupswere evaluated with the appropriate post hoc tests (e.g. Newman–Keuls Multiple Comparison) and all data are expressed as mean+ SEM.Statistical significance was set at P , 0.05.
Results
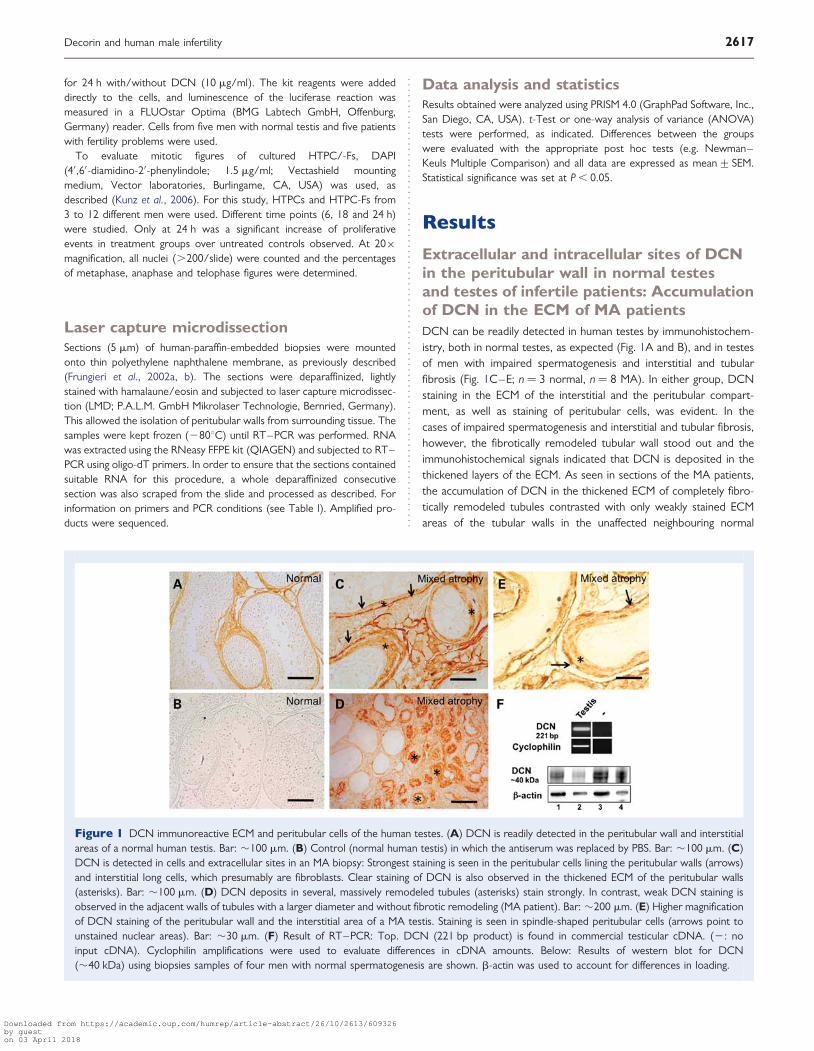
Extracellular and intracellular sites of DCNin the peritubular wall in normal testesand testes of infertile patients: Accumulationof DCN in the ECM of MA patientsDCN can be readily detected in human testes by immunohistochem-istry, both in normal testes, as expected (Fig. 1A and B), and in testesof men with impaired spermatogenesis and interstitial and tubularfibrosis (Fig. 1C–E; n ¼ 3 normal, n ¼ 8 MA). In either group, DCNstaining in the ECM of the interstitial and the peritubular compart-ment, as well as staining of peritubular cells, was evident. In thecases of impaired spermatogenesis and interstitial and tubular fibrosis,however, the fibrotically remodeled tubular wall stood out and theimmunohistochemical signals indicated that DCN is deposited in thethickened layers of the ECM. As seen in sections of the MA patients,the accumulation of DCN in the thickened ECM of completely fibro-tically remodeled tubules contrasted with only weakly stained ECMareas of the tubular walls in the unaffected neighbouring normal
Figure 1 DCN immunoreactive ECM and peritubular cells of the human testes. (A) DCN is readily detected in the peritubular wall and interstitialareas of a normal human testis. Bar: �100 mm. (B) Control (normal human testis) in which the antiserum was replaced by PBS. Bar: �100 mm. (C)DCN is detected in cells and extracellular sites in an MA biopsy: Strongest staining is seen in the peritubular cells lining the peritubular walls (arrows)and interstitial long cells, which presumably are fibroblasts. Clear staining of DCN is also observed in the thickened ECM of the peritubular walls(asterisks). Bar: �100 mm. (D) DCN deposits in several, massively remodeled tubules (asterisks) stain strongly. In contrast, weak DCN staining isobserved in the adjacent walls of tubules with a larger diameter and without fibrotic remodeling (MA patient). Bar: �200 mm. (E) Higher magnificationof DCN staining of the peritubular wall and the interstitial area of a MA testis. Staining is seen in spindle-shaped peritubular cells (arrows point tounstained nuclear areas). Bar: �30 mm. (F) Result of RT–PCR: Top. DCN (221 bp product) is found in commercial testicular cDNA. (2: noinput cDNA). Cyclophilin amplifications were used to evaluate differences in cDNA amounts. Below: Results of western blot for DCN(�40 kDa) using biopsies samples of four men with normal spermatogenesis are shown. b-actin was used to account for differences in loading.
Decorin and human male infertility 2617
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
tubules (Fig. 1D). As the cytoplasm of peritubular cells also stained(Fig. 1E), these cells are the likely to synthesize DCN. InterstitialECM deposits were also evident and the staining pattern obtainedsuggests that interstitial fibroblasts are the likely source of DCN(Fig. 1D). Controls with non-immune serum and omission ofprimary antibody were negative (Fig. 1B).
Expression and identity of DCN were confirmed by RT–PCR(Fig. 1F) and sequencing (data not shown) in normal human testesand by western blotting using four biopsy samples of men withnormal spermatogenesis (Fig. 1F).
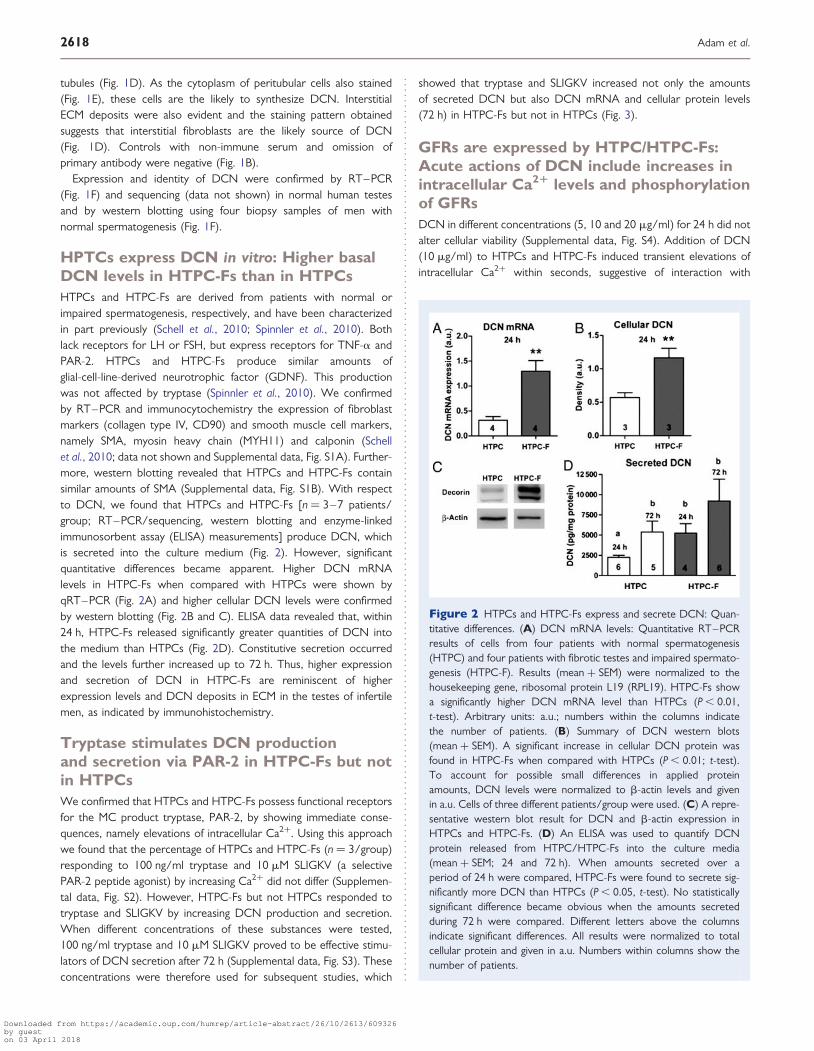
HPTCs express DCN in vitro: Higher basalDCN levels in HTPC-Fs than in HTPCsHTPCs and HTPC-Fs are derived from patients with normal orimpaired spermatogenesis, respectively, and have been characterizedin part previously (Schell et al., 2010; Spinnler et al., 2010). Bothlack receptors for LH or FSH, but express receptors for TNF-a andPAR-2. HTPCs and HTPC-Fs produce similar amounts ofglial-cell-line-derived neurotrophic factor (GDNF). This productionwas not affected by tryptase (Spinnler et al., 2010). We confirmedby RT–PCR and immunocytochemistry the expression of fibroblastmarkers (collagen type IV, CD90) and smooth muscle cell markers,namely SMA, myosin heavy chain (MYH11) and calponin (Schellet al., 2010; data not shown and Supplemental data, Fig. S1A). Further-more, western blotting revealed that HTPCs and HTPC-Fs containsimilar amounts of SMA (Supplemental data, Fig. S1B). With respectto DCN, we found that HTPCs and HTPC-Fs [n ¼ 3–7 patients/group; RT–PCR/sequencing, western blotting and enzyme-linkedimmunosorbent assay (ELISA) measurements] produce DCN, whichis secreted into the culture medium (Fig. 2). However, significantquantitative differences became apparent. Higher DCN mRNAlevels in HTPC-Fs when compared with HTPCs were shown byqRT–PCR (Fig. 2A) and higher cellular DCN levels were confirmedby western blotting (Fig. 2B and C). ELISA data revealed that, within24 h, HTPC-Fs released significantly greater quantities of DCN intothe medium than HTPCs (Fig. 2D). Constitutive secretion occurredand the levels further increased up to 72 h. Thus, higher expressionand secretion of DCN in HTPC-Fs are reminiscent of higherexpression levels and DCN deposits in ECM in the testes of infertilemen, as indicated by immunohistochemistry.
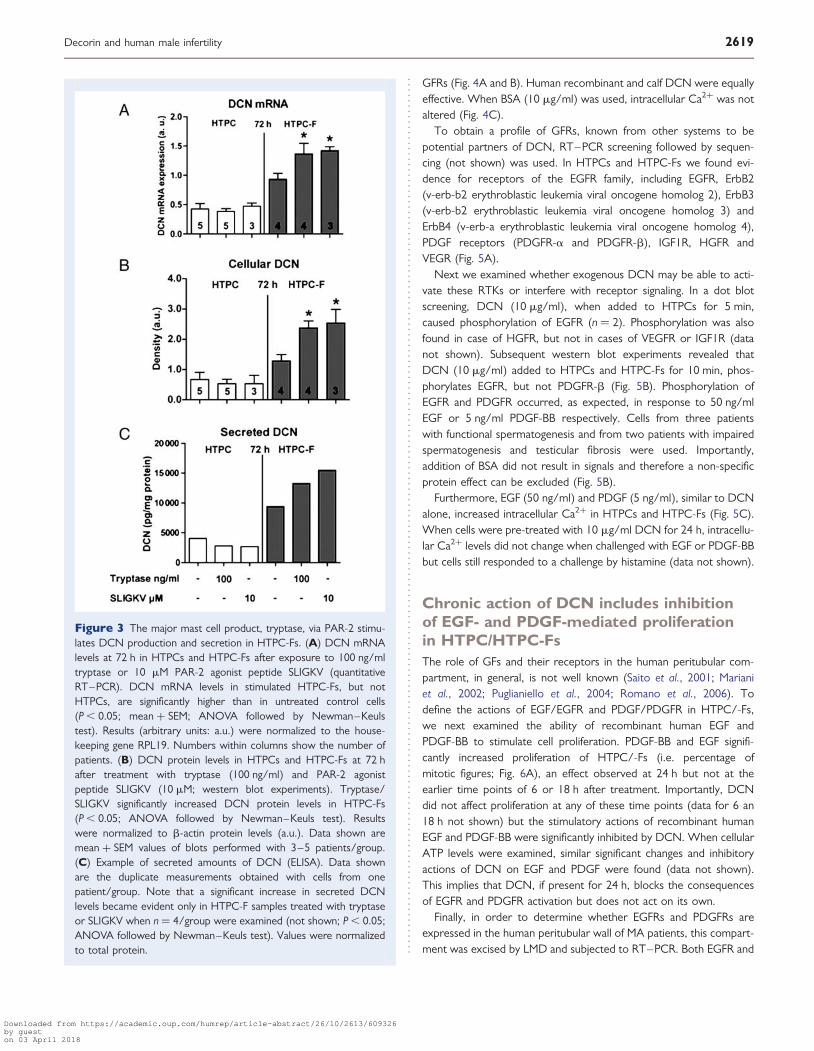
Tryptase stimulates DCN productionand secretion via PAR-2 in HTPC-Fs but notin HTPCsWe confirmed that HTPCs and HTPC-Fs possess functional receptorsfor the MC product tryptase, PAR-2, by showing immediate conse-quences, namely elevations of intracellular Ca2+. Using this approachwe found that the percentage of HTPCs and HTPC-Fs (n ¼ 3/group)responding to 100 ng/ml tryptase and 10 mM SLIGKV (a selectivePAR-2 peptide agonist) by increasing Ca2+ did not differ (Supplemen-tal data, Fig. S2). However, HTPC-Fs but not HTPCs responded totryptase and SLIGKV by increasing DCN production and secretion.When different concentrations of these substances were tested,100 ng/ml tryptase and 10 mM SLIGKV proved to be effective stimu-lators of DCN secretion after 72 h (Supplemental data, Fig. S3). Theseconcentrations were therefore used for subsequent studies, which
showed that tryptase and SLIGKV increased not only the amountsof secreted DCN but also DCN mRNA and cellular protein levels(72 h) in HTPC-Fs but not in HTPCs (Fig. 3).
GFRs are expressed by HTPC/HTPC-Fs:Acute actions of DCN include increases inintracellular Ca21 levels and phosphorylationof GFRsDCN in different concentrations (5, 10 and 20 mg/ml) for 24 h did notalter cellular viability (Supplemental data, Fig. S4). Addition of DCN(10 mg/ml) to HTPCs and HTPC-Fs induced transient elevations ofintracellular Ca2+ within seconds, suggestive of interaction with
Figure 2 HTPCs and HTPC-Fs express and secrete DCN: Quan-titative differences. (A) DCN mRNA levels: Quantitative RT–PCRresults of cells from four patients with normal spermatogenesis(HTPC) and four patients with fibrotic testes and impaired spermato-genesis (HTPC-F). Results (mean + SEM) were normalized to thehousekeeping gene, ribosomal protein L19 (RPL19). HTPC-Fs showa significantly higher DCN mRNA level than HTPCs (P , 0.01,t-test). Arbitrary units: a.u.; numbers within the columns indicatethe number of patients. (B) Summary of DCN western blots(mean + SEM). A significant increase in cellular DCN protein wasfound in HTPC-Fs when compared with HTPCs (P , 0.01; t-test).To account for possible small differences in applied proteinamounts, DCN levels were normalized to b-actin levels and givenin a.u. Cells of three different patients/group were used. (C) A repre-sentative western blot result for DCN and b-actin expression inHTPCs and HTPC-Fs. (D) An ELISA was used to quantify DCNprotein released from HTPC/HTPC-Fs into the culture media(mean + SEM; 24 and 72 h). When amounts secreted over aperiod of 24 h were compared, HTPC-Fs were found to secrete sig-nificantly more DCN than HTPCs (P , 0.05, t-test). No statisticallysignificant difference became obvious when the amounts secretedduring 72 h were compared. Different letters above the columnsindicate significant differences. All results were normalized to totalcellular protein and given in a.u. Numbers within columns show thenumber of patients.
2618 Adam et al.
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
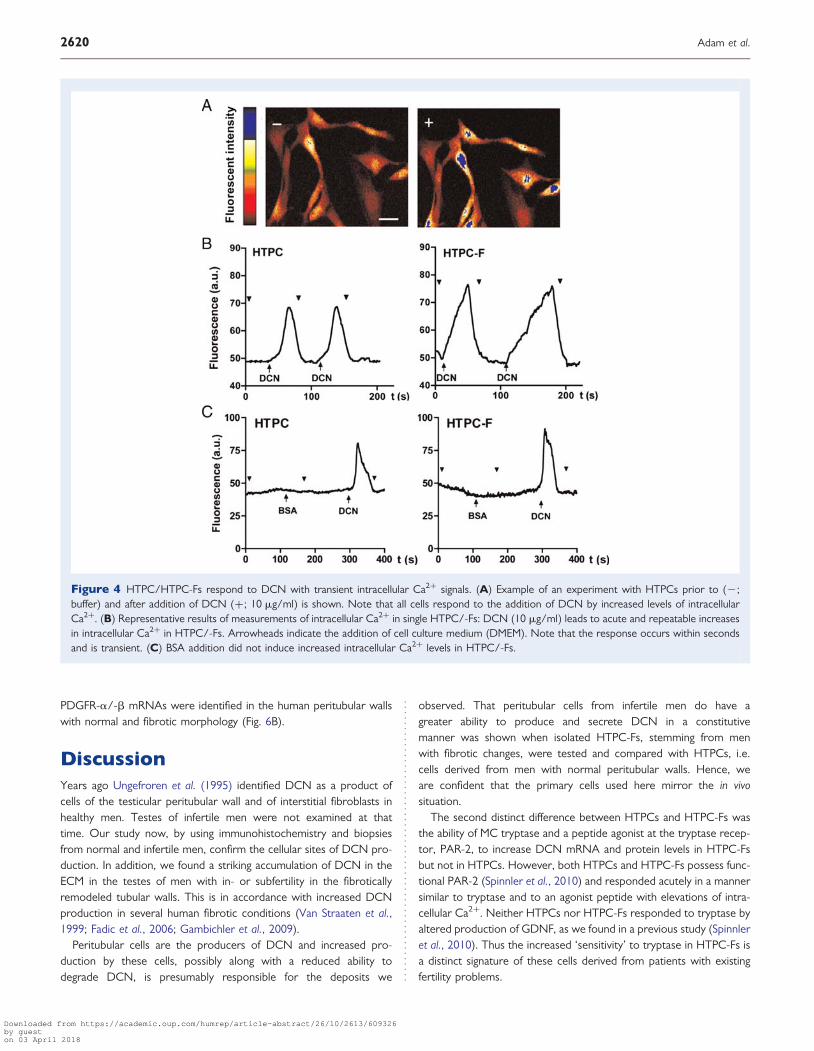
GFRs (Fig. 4A and B). Human recombinant and calf DCN were equallyeffective. When BSA (10 mg/ml) was used, intracellular Ca2+ was notaltered (Fig. 4C).
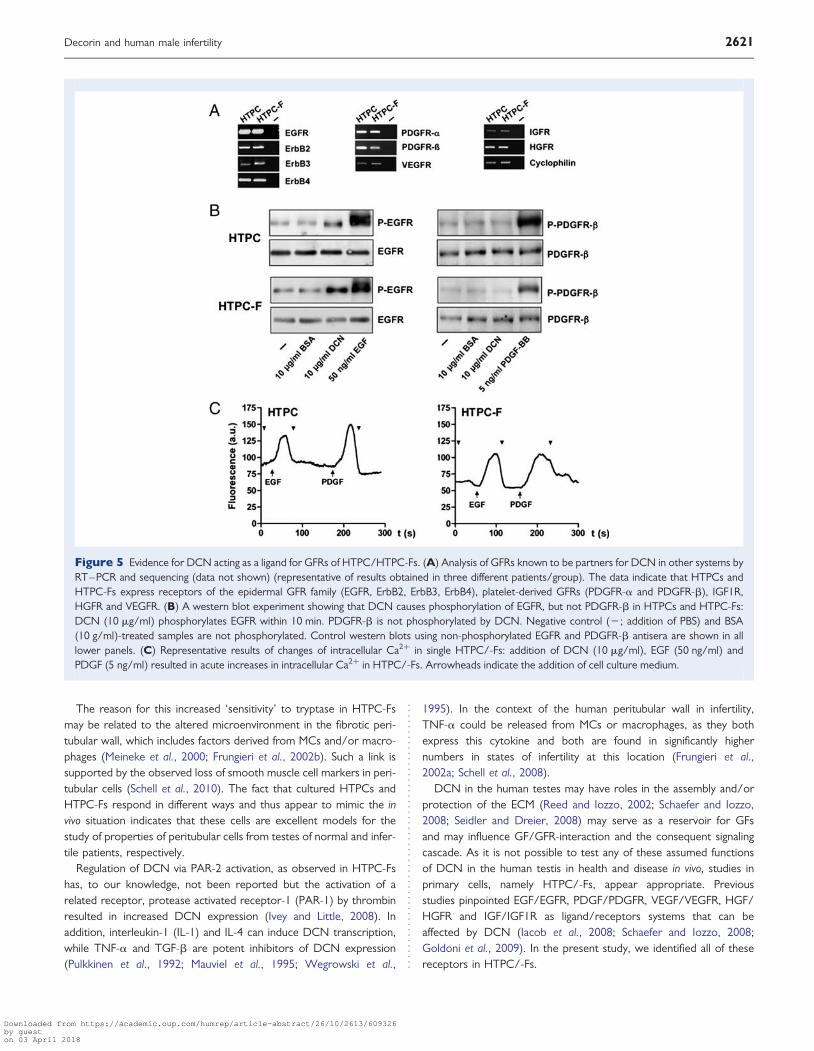
To obtain a profile of GFRs, known from other systems to bepotential partners of DCN, RT–PCR screening followed by sequen-cing (not shown) was used. In HTPCs and HTPC-Fs we found evi-dence for receptors of the EGFR family, including EGFR, ErbB2(v-erb-b2 erythroblastic leukemia viral oncogene homolog 2), ErbB3(v-erb-b2 erythroblastic leukemia viral oncogene homolog 3) andErbB4 (v-erb-a erythroblastic leukemia viral oncogene homolog 4),PDGF receptors (PDGFR-a and PDGFR-b), IGF1R, HGFR andVEGR (Fig. 5A).
Next we examined whether exogenous DCN may be able to acti-vate these RTKs or interfere with receptor signaling. In a dot blotscreening, DCN (10 mg/ml), when added to HTPCs for 5 min,caused phosphorylation of EGFR (n ¼ 2). Phosphorylation was alsofound in case of HGFR, but not in cases of VEGFR or IGF1R (datanot shown). Subsequent western blot experiments revealed thatDCN (10 mg/ml) added to HTPCs and HTPC-Fs for 10 min, phos-phorylates EGFR, but not PDGFR-b (Fig. 5B). Phosphorylation ofEGFR and PDGFR occurred, as expected, in response to 50 ng/mlEGF or 5 ng/ml PDGF-BB respectively. Cells from three patientswith functional spermatogenesis and from two patients with impairedspermatogenesis and testicular fibrosis were used. Importantly,addition of BSA did not result in signals and therefore a non-specificprotein effect can be excluded (Fig. 5B).
Furthermore, EGF (50 ng/ml) and PDGF (5 ng/ml), similar to DCNalone, increased intracellular Ca2+ in HTPCs and HTPC-Fs (Fig. 5C).When cells were pre-treated with 10 mg/ml DCN for 24 h, intracellu-lar Ca2+ levels did not change when challenged with EGF or PDGF-BBbut cells still responded to a challenge by histamine (data not shown).
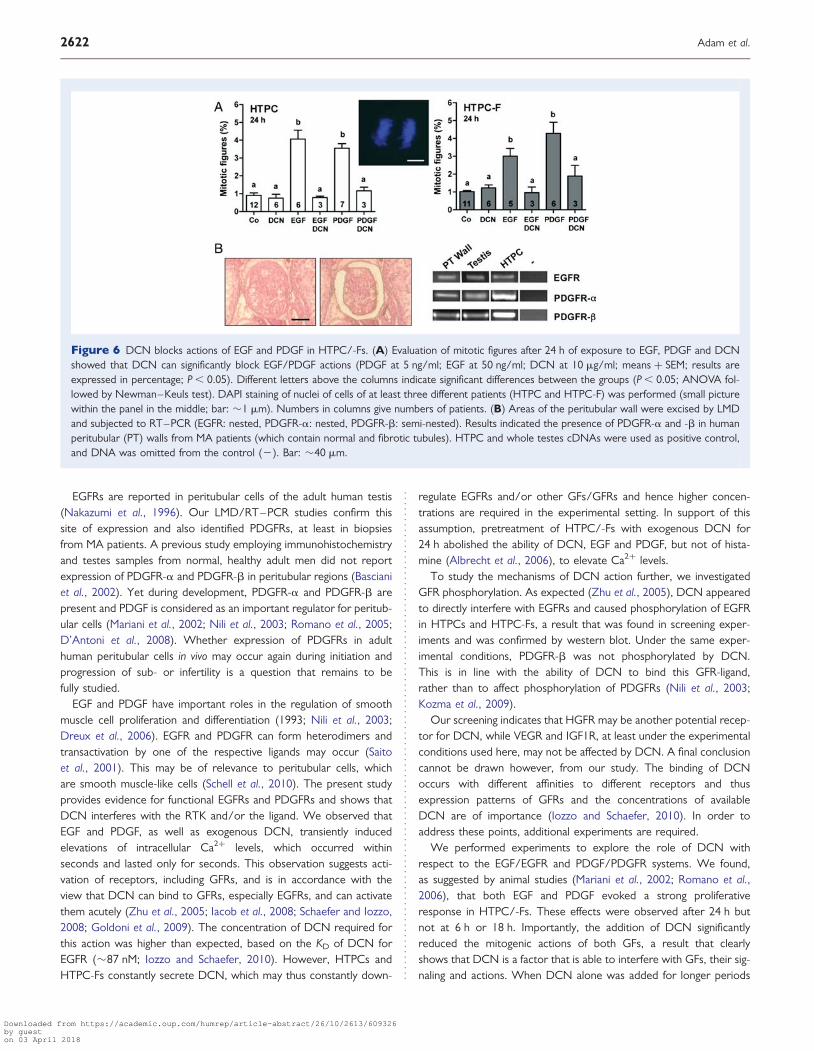
Chronic action of DCN includes inhibitionof EGF- and PDGF-mediated proliferationin HTPC/HTPC-FsThe role of GFs and their receptors in the human peritubular com-partment, in general, is not well known (Saito et al., 2001; Marianiet al., 2002; Puglianiello et al., 2004; Romano et al., 2006). Todefine the actions of EGF/EGFR and PDGF/PDGFR in HTPC/-Fs,we next examined the ability of recombinant human EGF andPDGF-BB to stimulate cell proliferation. PDGF-BB and EGF signifi-cantly increased proliferation of HTPC/-Fs (i.e. percentage ofmitotic figures; Fig. 6A), an effect observed at 24 h but not at theearlier time points of 6 or 18 h after treatment. Importantly, DCNdid not affect proliferation at any of these time points (data for 6 an18 h not shown) but the stimulatory actions of recombinant humanEGF and PDGF-BB were significantly inhibited by DCN. When cellularATP levels were examined, similar significant changes and inhibitoryactions of DCN on EGF and PDGF were found (data not shown).This implies that DCN, if present for 24 h, blocks the consequencesof EGFR and PDGFR activation but does not act on its own.
Finally, in order to determine whether EGFRs and PDGFRs areexpressed in the human peritubular wall of MA patients, this compart-ment was excised by LMD and subjected to RT–PCR. Both EGFR and
Figure 3 The major mast cell product, tryptase, via PAR-2 stimu-lates DCN production and secretion in HTPC-Fs. (A) DCN mRNAlevels at 72 h in HTPCs and HTPC-Fs after exposure to 100 ng/mltryptase or 10 mM PAR-2 agonist peptide SLIGKV (quantitativeRT–PCR). DCN mRNA levels in stimulated HTPC-Fs, but notHTPCs, are significantly higher than in untreated control cells(P , 0.05; mean + SEM; ANOVA followed by Newman–Keulstest). Results (arbitrary units: a.u.) were normalized to the house-keeping gene RPL19. Numbers within columns show the number ofpatients. (B) DCN protein levels in HTPCs and HTPC-Fs at 72 hafter treatment with tryptase (100 ng/ml) and PAR-2 agonistpeptide SLIGKV (10 mM; western blot experiments). Tryptase/SLIGKV significantly increased DCN protein levels in HTPC-Fs(P , 0.05; ANOVA followed by Newman–Keuls test). Resultswere normalized to b-actin protein levels (a.u.). Data shown aremean + SEM values of blots performed with 3–5 patients/group.(C) Example of secreted amounts of DCN (ELISA). Data shownare the duplicate measurements obtained with cells from onepatient/group. Note that a significant increase in secreted DCNlevels became evident only in HTPC-F samples treated with tryptaseor SLIGKV when n ¼ 4/group were examined (not shown; P , 0.05;ANOVA followed by Newman–Keuls test). Values were normalizedto total protein.
Decorin and human male infertility 2619
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
PDGFR-a/-b mRNAs were identified in the human peritubular wallswith normal and fibrotic morphology (Fig. 6B).
DiscussionYears ago Ungefroren et al. (1995) identified DCN as a product ofcells of the testicular peritubular wall and of interstitial fibroblasts inhealthy men. Testes of infertile men were not examined at thattime. Our study now, by using immunohistochemistry and biopsiesfrom normal and infertile men, confirm the cellular sites of DCN pro-duction. In addition, we found a striking accumulation of DCN in theECM in the testes of men with in- or subfertility in the fibroticallyremodeled tubular walls. This is in accordance with increased DCNproduction in several human fibrotic conditions (Van Straaten et al.,1999; Fadic et al., 2006; Gambichler et al., 2009).
Peritubular cells are the producers of DCN and increased pro-duction by these cells, possibly along with a reduced ability todegrade DCN, is presumably responsible for the deposits we
observed. That peritubular cells from infertile men do have agreater ability to produce and secrete DCN in a constitutivemanner was shown when isolated HTPC-Fs, stemming from menwith fibrotic changes, were tested and compared with HTPCs, i.e.cells derived from men with normal peritubular walls. Hence, weare confident that the primary cells used here mirror the in vivosituation.
The second distinct difference between HTPCs and HTPC-Fs wasthe ability of MC tryptase and a peptide agonist at the tryptase recep-tor, PAR-2, to increase DCN mRNA and protein levels in HTPC-Fsbut not in HTPCs. However, both HTPCs and HTPC-Fs possess func-tional PAR-2 (Spinnler et al., 2010) and responded acutely in a mannersimilar to tryptase and to an agonist peptide with elevations of intra-cellular Ca2+. Neither HTPCs nor HTPC-Fs responded to tryptase byaltered production of GDNF, as we found in a previous study (Spinnleret al., 2010). Thus the increased ‘sensitivity’ to tryptase in HTPC-Fs isa distinct signature of these cells derived from patients with existingfertility problems.
Figure 4 HTPC/HTPC-Fs respond to DCN with transient intracellular Ca2+ signals. (A) Example of an experiment with HTPCs prior to (2;buffer) and after addition of DCN (+; 10 mg/ml) is shown. Note that all cells respond to the addition of DCN by increased levels of intracellularCa2+. (B) Representative results of measurements of intracellular Ca2+ in single HTPC/-Fs: DCN (10 mg/ml) leads to acute and repeatable increasesin intracellular Ca2+ in HTPC/-Fs. Arrowheads indicate the addition of cell culture medium (DMEM). Note that the response occurs within secondsand is transient. (C) BSA addition did not induce increased intracellular Ca2+ levels in HTPC/-Fs.
2620 Adam et al.
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
The reason for this increased ‘sensitivity’ to tryptase in HTPC-Fsmay be related to the altered microenvironment in the fibrotic peri-tubular wall, which includes factors derived from MCs and/or macro-phages (Meineke et al., 2000; Frungieri et al., 2002b). Such a link issupported by the observed loss of smooth muscle cell markers in peri-tubular cells (Schell et al., 2010). The fact that cultured HTPCs andHTPC-Fs respond in different ways and thus appear to mimic the invivo situation indicates that these cells are excellent models for thestudy of properties of peritubular cells from testes of normal and infer-tile patients, respectively.
Regulation of DCN via PAR-2 activation, as observed in HTPC-Fshas, to our knowledge, not been reported but the activation of arelated receptor, protease activated receptor-1 (PAR-1) by thrombinresulted in increased DCN expression (Ivey and Little, 2008). Inaddition, interleukin-1 (IL-1) and IL-4 can induce DCN transcription,while TNF-a and TGF-b are potent inhibitors of DCN expression(Pulkkinen et al., 1992; Mauviel et al., 1995; Wegrowski et al.,
1995). In the context of the human peritubular wall in infertility,TNF-a could be released from MCs or macrophages, as they bothexpress this cytokine and both are found in significantly highernumbers in states of infertility at this location (Frungieri et al.,2002a; Schell et al., 2008).
DCN in the human testes may have roles in the assembly and/orprotection of the ECM (Reed and Iozzo, 2002; Schaefer and Iozzo,2008; Seidler and Dreier, 2008) may serve as a reservoir for GFsand may influence GF/GFR-interaction and the consequent signalingcascade. As it is not possible to test any of these assumed functionsof DCN in the human testis in health and disease in vivo, studies inprimary cells, namely HTPC/-Fs, appear appropriate. Previousstudies pinpointed EGF/EGFR, PDGF/PDGFR, VEGF/VEGFR, HGF/HGFR and IGF/IGF1R as ligand/receptors systems that can beaffected by DCN (Iacob et al., 2008; Schaefer and Iozzo, 2008;Goldoni et al., 2009). In the present study, we identified all of thesereceptors in HTPC/-Fs.
Figure 5 Evidence for DCN acting as a ligand for GFRs of HTPC/HTPC-Fs. (A) Analysis of GFRs known to be partners for DCN in other systems byRT–PCR and sequencing (data not shown) (representative of results obtained in three different patients/group). The data indicate that HTPCs andHTPC-Fs express receptors of the epidermal GFR family (EGFR, ErbB2, ErbB3, ErbB4), platelet-derived GFRs (PDGFR-a and PDGFR-b), IGF1R,HGFR and VEGFR. (B) A western blot experiment showing that DCN causes phosphorylation of EGFR, but not PDGFR-b in HTPCs and HTPC-Fs:DCN (10 mg/ml) phosphorylates EGFR within 10 min. PDGFR-b is not phosphorylated by DCN. Negative control (2; addition of PBS) and BSA(10 g/ml)-treated samples are not phosphorylated. Control western blots using non-phosphorylated EGFR and PDGFR-b antisera are shown in alllower panels. (C) Representative results of changes of intracellular Ca2+ in single HTPC/-Fs: addition of DCN (10 mg/ml), EGF (50 ng/ml) andPDGF (5 ng/ml) resulted in acute increases in intracellular Ca2+ in HTPC/-Fs. Arrowheads indicate the addition of cell culture medium.
Decorin and human male infertility 2621
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018
EGFRs are reported in peritubular cells of the adult human testis(Nakazumi et al., 1996). Our LMD/RT–PCR studies confirm thissite of expression and also identified PDGFRs, at least in biopsiesfrom MA patients. A previous study employing immunohistochemistryand testes samples from normal, healthy adult men did not reportexpression of PDGFR-a and PDGFR-b in peritubular regions (Bascianiet al., 2002). Yet during development, PDGFR-a and PDGFR-b arepresent and PDGF is considered as an important regulator for peritub-ular cells (Mariani et al., 2002; Nili et al., 2003; Romano et al., 2005;D’Antoni et al., 2008). Whether expression of PDGFRs in adulthuman peritubular cells in vivo may occur again during initiation andprogression of sub- or infertility is a question that remains to befully studied.
EGF and PDGF have important roles in the regulation of smoothmuscle cell proliferation and differentiation (1993; Nili et al., 2003;Dreux et al., 2006). EGFR and PDGFR can form heterodimers andtransactivation by one of the respective ligands may occur (Saitoet al., 2001). This may be of relevance to peritubular cells, whichare smooth muscle-like cells (Schell et al., 2010). The present studyprovides evidence for functional EGFRs and PDGFRs and shows thatDCN interferes with the RTK and/or the ligand. We observed thatEGF and PDGF, as well as exogenous DCN, transiently inducedelevations of intracellular Ca2+ levels, which occurred withinseconds and lasted only for seconds. This observation suggests acti-vation of receptors, including GFRs, and is in accordance with theview that DCN can bind to GFRs, especially EGFRs, and can activatethem acutely (Zhu et al., 2005; Iacob et al., 2008; Schaefer and Iozzo,2008; Goldoni et al., 2009). The concentration of DCN required forthis action was higher than expected, based on the KD of DCN forEGFR (�87 nM; Iozzo and Schaefer, 2010). However, HTPCs andHTPC-Fs constantly secrete DCN, which may thus constantly down-
regulate EGFRs and/or other GFs/GFRs and hence higher concen-trations are required in the experimental setting. In support of thisassumption, pretreatment of HTPC/-Fs with exogenous DCN for24 h abolished the ability of DCN, EGF and PDGF, but not of hista-mine (Albrecht et al., 2006), to elevate Ca2+ levels.
To study the mechanisms of DCN action further, we investigatedGFR phosphorylation. As expected (Zhu et al., 2005), DCN appearedto directly interfere with EGFRs and caused phosphorylation of EGFRin HTPCs and HTPC-Fs, a result that was found in screening exper-iments and was confirmed by western blot. Under the same exper-imental conditions, PDGFR-b was not phosphorylated by DCN.This is in line with the ability of DCN to bind this GFR-ligand,rather than to affect phosphorylation of PDGFRs (Nili et al., 2003;Kozma et al., 2009).
Our screening indicates that HGFR may be another potential recep-tor for DCN, while VEGR and IGF1R, at least under the experimentalconditions used here, may not be affected by DCN. A final conclusioncannot be drawn however, from our study. The binding of DCNoccurs with different affinities to different receptors and thusexpression patterns of GFRs and the concentrations of availableDCN are of importance (Iozzo and Schaefer, 2010). In order toaddress these points, additional experiments are required.
We performed experiments to explore the role of DCN withrespect to the EGF/EGFR and PDGF/PDGFR systems. We found,as suggested by animal studies (Mariani et al., 2002; Romano et al.,2006), that both EGF and PDGF evoked a strong proliferativeresponse in HTPC/-Fs. These effects were observed after 24 h butnot at 6 h or 18 h. Importantly, the addition of DCN significantlyreduced the mitogenic actions of both GFs, a result that clearlyshows that DCN is a factor that is able to interfere with GFs, their sig-naling and actions. When DCN alone was added for longer periods
Figure 6 DCN blocks actions of EGF and PDGF in HTPC/-Fs. (A) Evaluation of mitotic figures after 24 h of exposure to EGF, PDGF and DCNshowed that DCN can significantly block EGF/PDGF actions (PDGF at 5 ng/ml; EGF at 50 ng/ml; DCN at 10 mg/ml; means + SEM; results areexpressed in percentage; P , 0.05). Different letters above the columns indicate significant differences between the groups (P , 0.05; ANOVA fol-lowed by Newman–Keuls test). DAPI staining of nuclei of cells of at least three different patients (HTPC and HTPC-F) was performed (small picturewithin the panel in the middle; bar: �1 mm). Numbers in columns give numbers of patients. (B) Areas of the peritubular wall were excised by LMDand subjected to RT–PCR (EGFR: nested, PDGFR-a: nested, PDGFR-b: semi-nested). Results indicated the presence of PDGFR-a and -b in humanperitubular (PT) walls from MA patients (which contain normal and fibrotic tubules). HTPC and whole testes cDNAs were used as positive control,and DNA was omitted from the control (2). Bar: �40 mm.
2622 Adam et al.
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018

(up to 24 h) to cultured HTPC/-Fs, it did not alter cellular viability orcell proliferation, a result in line with studies in human airway smoothmuscle cells (D’Antoni et al., 2008). This lack of action of DCN after24 h also indicates that an initial activation, as shown by transientlyincreased Ca2+ levels or phosphorylation of the EGFR, is followedby a down-regulation of EGR signaling (Csordas et al., 2000; Zhuet al., 2005). In the case of PDGF, we suggest that DCN bound theGF and thus abolished its proliferative potential.
It is worth mentioning that the present investigation revealed DCNstaining also in the interstitial compartment of the human testis andthat several GFRs have been described in Leydig cells (Gnessi et al.,1995; Basciani et al., 2002). Thus, the ability of DCN to interferewith GF actions in the human testis may extend to Leydig as well asother cell types.
In summary, the present study provides insights into the roles ofDCN as a factor involved in testicular paracrine signaling. Results ofour study using cultured human testicular cells from normal and sub/-infertile men support such actions. Furthermore our results indicatethat in the testes of men with impaired spermatogenesis, the typicalfibrotic changes include increased amounts of the ECM proteinDCN, which we suggest consequently may lead to an imbalance in tes-ticular GF signaling via direct actions and/or by serving as a reservoirof GFs. Although we observed increased levels of DCN in HTPC-Fs,we could detect no difference between HTPCs and HTPC-Fs withregard to the Ca2+ increase, mitotic activity and phosphorylation ofEGFRs and PDGFRs. Despite this, we cannot rule out the possibilitythat increased levels of DCN upset the normal balance of testicularfunction, including the actions of GFs. In men with impaired sperma-togenesis, testicular MCs are increased in number and our studyshows that their major product, tryptase, can act as a regulator ofDCN production by peritubular cells. Hence, the results also indicatethat the functions of testicular MCs may be intrinsically linked to tes-ticular alterations in infertile men. In the search for novel therapeuticinterventions in male infertility MCs may be a key cell type to target(Meineke et al., 2000; Frungieri et al., 2002b; Schell et al., 2008, 2010).
Supplementary dataSupplementary data are available at http://humrep.oxfordjournals.org/.
Authors’ rolesThis work was done in partial fulfillment of the requirements of a Drrer nat. thesis of M.A. M.A. performed the majority of the work withperitubular cells and analyzed the results. J.U.S. and F.M.K. contributedessential biopsies and helped in the interpretation of the data andwriting of the paper. L.S. and M.P. provided access to qRT–PCR,helped analyzing qRT–PCR data and drafting of the manuscript.A.M. conceived of the study and designed the research. Togetherwith M.A. he wrote the manuscript. All authors contributed to thefinal version of the paper and approved it.
AcknowledgementsWe thank Astrid Tiefenbacher, Daniel Einwang, Sandra Raffael andSimone Spillner for excellent technical assistance, Karin Metzrath for
assistance with writing the manuscript and Dr Martin Albrecht andNimisha Desai for their input and discussions in the beginning of thisstudy.
FundingThis study was supported by grants from Deutsche Forschungsge-meinschaft (DFG) MA 1080/16–3, Deutscher Akademischer Aus-tauschdienst (DAAD) and the Academy of Finland.
ReferencesAlbrecht M, Frungieri MB, Kunz L, Ramsch R, Meineke V, Kohn FM,
Mayerhofer A. Divergent effects of the major mast cell productshistamine, tryptase and TNF-alpha on human fibroblast behaviour. CellMol Life Sci 2005;62:2867–2876.
Albrecht M, Ramsch R, Kohn FM, Schwarzer JU, Mayerhofer A. Isolationand cultivation of human testicular peritubular cells: a new model forthe investigation of fibrotic processes in the human testis and maleinfertility. J Clin Endocrinol Metab 2006;91:1956–1960.
Basciani S, Mariani S, Arizzi M, Ulisse S, Rucci N, Jannini EA, Della Rocca C,Manicone A, Carani C, Spera G et al. Expression of platelet-derivedgrowth factor-A (PDGF-A), PDGF-B, and PDGF receptor-alpha and-beta during human testicular development and disease. J ClinEndocrinol Metab 2002;87:2310–2319.
Basciani S, Mariani S, Spera G, Gnessi L. Role of platelet-derived growthfactors in the testis. Endocr Rev 2010;31:916–939.
Cigorraga SB, Chemes H, Pellizzari E. Steroidogenic and morphogeniccharacteristics of human peritubular cells in culture. Biol Reprod 1994;51:1193–1205.
Csordas G, Santra M, Reed CC, Eichstetter I, McQuillan DJ, Gross D,Nugent MA, Hajnoczky G, Iozzo RV. Sustained down-regulation ofthe epidermal growth factor receptor by decorin. A mechanism forcontrolling tumor growth in vivo. J Biol Chem 2000;275:32879–32887.
D’Antoni ML, Torregiani C, Ferraro P, Michoud MC, Mazer B, Martin JG,Ludwig MS. Effects of decorin and biglycan on human airway smoothmuscle cell proliferation and apoptosis. Am J Physiol Lung Cell MolPhysiol 2008;294:L764–L771.
Davidoff MS, Breucker H, Holstein AF, Seidl K. Cellular architecture of thelamina propria of human seminiferous tubules. Cell Tissue Res 1990;262:253–261.
De Kretser D, Baker H. Human infertility: the male factor. In: Adashi EY,Rock JA, Rosenwaks Z (eds) Reproductive endocrinology, surgery andtechnology. Philadelphia: Lippincott-Raven, 1996,2031–2062.
Dobashi M, Fujisawa M, Naito I, Yamazaki T, Okada H, Kamidono S.Distribution of type IV collagen subtypes in human testes and theirassociation with spermatogenesis. Fertil Steril 2003;80 Suppl 2:755–760.
Dreux AC, Lamb DJ, Modjtahedi H, Ferns GA. The epidermal growthfactor receptors and their family of ligands: their putative role inatherogenesis. Atherosclerosis 2006;186:38–53.
Fadic R, Mezzano V, Alvarez K, Cabrera D, Holmgren J, Brandan E.Increase in decorin and biglycan in Duchenne Muscular Dystrophy:role of fibroblasts as cell source of these proteoglycans in the disease.J Cell Mol Med 2006;10:758–769.
Ferdous Z, Peterson SB, Tseng H, Anderson DK, Iozzo RV,Grande-Allen KJ. A role for decorin in controlling proliferation,adhesion, and migration of murine embryonic fibroblasts. J BiomedMater Res A 2010;93:419–428.
Fohr KJ, Mayerhofer A, Sterzik K, Rudolf M, Rosenbusch B, Gratzl M.Concerted action of human chorionic gonadotropin andnorepinephrine on intracellular-free calcium in human granulosa-lutein
Decorin and human male infertility 2623
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018

cells: evidence for the presence of a functional alpha-adrenergicreceptor. J Clin Endocrinol Metab 1993;76:367–373.
Frungieri MB, Calandra RS, Lustig L, Meineke V, Kohn FM, Vogt HJ,Mayerhofer A. Number, distribution pattern, and identification ofmacrophages in the testes of infertile men. Fertil Steril 2002a;78:298–306.
Frungieri MB, Weidinger S, Meineke V, Kohn FM, Mayerhofer A.Proliferative action of mast-cell tryptase is mediated by PAR2,COX-2, prostaglandins, and PPAR. PNAS 2002b;99:15072–15077.
Gambichler T, Kreuter A, Skrygan M, Burkert B, Altmeyer P, Schieren G.Decorin is significantly overexpressed in nephrogenic systemic fibrosis.Am J Clin Pathol 2009;132:139–143.
Gnessi L, Emidi A, Jannini EA, Carosa E, Maroder M, Arizzi M, Ulisse S,Spera G. Testicular development involves the spatiotemporal controlof PDGFs and PDGF receptors gene expression and action. J Cell Biol1995;131:1105–1121.
Goldoni S, Humphries A, Nystrom A, Sattar S, Owens RT, McQuillan DJ,Ireton K, Iozzo RV. Decorin is a novel antagonistic ligand of the Metreceptor. J Cell Biol 2009;185:743–754.
Gulkesen KH, Erdogru T, Sargin CF, Karpuzoglu G. Expression ofextracellular matrix proteins and vimentin in testes of azoospermicman: an immunohistochemical and morphometric study. Asian J Androl2002;4:55–60.
Haider SG, Talati J, Servos G. Ultrastructure of peritubular tissue inassociation with tubular hyalinization in human testis. Tissue Cell 1999;31:90–98.
Iacob D, Cai J, Tsonis M, Babwah A, Chakraborty C, Bhattacharjee RN,Lala PK. Decorin-mediated inhibition of proliferation and migration ofthe human trophoblast via different tyrosine kinase receptors.Endocrinology 2008;149:6187–6197.
Iozzo RV, Schaefer L. Proteoglycans in health and disease: novel regulatorysignaling mechanisms evoked by the small leucine-rich proteoglycans.FEBS J 2010;277:3864–3875.
Ivey ME, Little PJ. Thrombin regulates vascular smooth muscle cellproteoglycan synthesis via PAR-1 and multiple downstream signallingpathways. Thromb Res 2008;123:288–297.
Kozma EM, Wisowski G, Olczyk K. Platelet derived growth factor BB is aligand for dermatan sulfate chain(s) of small matrix proteoglycans fromnormal and fibrosis affected fascia. Biochimie 2009;91:1394–1404.
Kunz L, Ramsch R, Krieger A, Young KA, Dissen GA, Stouffer RL,Ojeda SR, Mayerhofer A. Voltage-dependent K+ channel acts as sexsteroid sensor in endocrine cells of the human ovary. J Cell Physiol2006;206:167–174.
Li X, Strauss L, Kaatrasalo A, Mayerhofer A, Huhtaniemi I, Santti R,Makela S, Poutanen M. Transgenic mice expressing p450 aromataseas a model for male infertility associated with chronic inflammation inthe testis. Endocrinology 2006;147:1271–1277.
Macri L, Silverstein D, Clark RA. Growth factor binding to the pericellularmatrix and its importance in tissue engineering. Adv Drug Deliv Rev 2007;59:1366–1381.
Maekawa M, Kamimura K, Nagano T. Peritubular myoid cells in the testis:their structure and function. Arch Histol Cytol 1996;59:1–13.
Mariani S, Basciani S, Arizzi M, Spera G, Gnessi L. PDGF and the testis.Trends Endocrinol Metab 2002;13:11–17.
Matzkin ME, Mayerhofer A, Rossi SP, Gonzalez B, Gonzalez CR,Gonzalez-Calvar SI, Terradas C, Ponzio R, Puigdomenech E,Levalle O et al. Cyclooxygenase-2 in testes of infertile men: evidencefor the induction of prostaglandin synthesis by interleukin-1beta. FertilSteril 2010;94:1933–1936.
Mauviel A, Santra M, Chen YQ, Uitto J, Iozzo RV. Transcriptionalregulation of decorin gene expression. Induction by quiescence andrepression by tumor necrosis factor-alpha. J Biol Chem 1995;270:11692–11700.
Meineke V, Frungieri MB, Jessberger B, Vogt H, Mayerhofer A. Humantesticular mast cells contain tryptase: increased mast cell number andaltered distribution in the testes of infertile men. Fertil Steril 2000;74:239–244.
Miqueloto CA, Zorn TM. Characterization and distribution of hyaluronanand the proteoglycans decorin, biglycan and perlecan in the developingembryonic mouse gonad. J Anat 2007;211:16–25.
Nakazumi H, Sasano H, Maehara I, Orikasa S. Transforming growthfactor-alpha, epidermal growth factor, and epidermal growth factorreceptor in human testis obtained from biopsy and castration:immunohistochemical study. Tohoku J Exp Med 1996;178:381–388.
Nili N, Cheema AN, Giordano FJ, Barolet AW, Babaei S, Hickey R,Eskandarian MR, Smeets M, Butany J, Pasterkamp G et al. Decorininhibition of PDGF-stimulated vascular smooth muscle cell function:potential mechanism for inhibition of intimal hyperplasia after balloonangioplasty. Am J Pathol 2003;163:869–878.
Nurmio M, Toppari J, Zaman F, Andersson AM, Paranko J, Soder O,Jahnukainen K. Inhibition of tyrosine kinases PDGFR and C-Kit byimatinib mesylate interferes with postnatal testicular development inthe rat. Int J Androl 2007;30:366–376.
Puglianiello A, Campagnolo L, Farini D, Cipollone D, Russo MA,Siracusa G. Expression and role of PDGF-BB and PDGFR-beta duringtestis morphogenesis in the mouse embryo. J Cell Sci 2004;117:1151–1160.
Pulkkinen L, Alitalo T, Krusius T, Peltonen L. Expression of decorin inhuman tissue and cell lines and chromosomal assignment of the genelocus (DCN). Cytogenet Cell Genet 1992;60:107–111.
Reed CC, Iozzo RV. The role of decorin in collagen fibrillogenesis and skinhomeostasis. Glycoconj J 2002;19:249–255.
Ricci G, Catizone A, Galdieri M. Embryonic mouse testis development:role of platelet derived growth factor (PDGF-BB). J Cell Physiol 2004;200:458–467.
Romano F, Tripiciano A, Muciaccia B, De Cesaris P, Ziparo E, Palombi F,Filippini A. The contractile phenotype of peritubular smooth musclecells is locally controlled: possible implications in male fertility.Contraception 2005;72:294–297.
Romano F, Chiarenza C, Palombi F, Filippini A, Padula F, Ziparo E, DeCesaris P. Platelet-derived growth factor-BB-induced hypertrophy ofperitubular smooth muscle cells is mediated by activation of p38MAP-kinase and of Rho-kinase. J Cell Physiol 2006;207:123–131.
Rossi F, Ferraresi A, Romagni P, Silvestroni L, Santiemma V. Angiotensin IIstimulates contraction and growth of testicular peritubular myoid cells invitro. Endocrinology 2002;143:3096–3104.
Saito Y, Haendeler J, Hojo Y, Yamamoto K, Berk BC. Receptorheterodimerization: essential mechanism for platelet-derived growthfactor-induced epidermal growth factor receptor transactivation. MolCell Biol 2001;21:6387–6394.
Santoro G, Romeo C, Impellizzeri P, Gentile C, Anastasi G, Santoro A.Ultrastructural and immunohistochemical study of basal lamina of thetestis in adolescent varicocele. Fertil Steril 2000;73:699–705.
Schaefer L, Iozzo RV. Biological functions of the small leucine-richproteoglycans: from genetics to signal transduction. J Biol Chem 2008;283:21305–21309.
Schell C, Albrecht M, Mayer C, Schwarzer JU, Frungieri MB, Mayerhofer A.Exploring human testicular peritubular cells: identification of secretoryproducts and regulation by tumor necrosis factor-alpha. Endocrinology2008;149:1678–1686.
Schell C, Albrecht M, Spillner S, Mayer C, Kunz L, Kohn FM, Schwarzer U,Mayerhofer A. 15-deoxy-delta-Prostaglandin-J2 (15dPGJ2) induceshypertrophy and loss of contractility in human testicular peritubularcells: Implications for human male fertility. Endocrinology 2010;151:1257–1268.
2624 Adam et al.
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018

Seidler DG, Dreier R. Decorin and its galactosaminoglycan chain:extracellular regulator of cellular function? IUBMB Life 2008;60:729–733.
Spiess AN, Feig C, Schulze W, Chalmel F, Cappallo-Obermann H,Primig M, Kirchhoff C. Cross-platform gene expression signature ofhuman spermatogenic failure reveals inflammatory-like response. HumReprod 2007;22:2936–2946.
Spinnler K, Kohn FM, Schwarzer U, Mayerhofer A. Glial cell line-derivedneurotrophic factor is constitutively produced by human testicularperitubular cells and may contribute to the spermatogonial stem cellniche in man. Hum Reprod 2010;25:2181–2187.
Strauss L, Kallio J, Desai N, Pakarinen P, Miettinen T, Gylling H,Albrecht M, Makela S, Mayerhofer A, Poutanen M. Increasedexposure to estrogens disturbs maturation, steroidogenesis, andcholesterol homeostasis via estrogen receptor alpha in adult mouseLeydig cells. Endocrinology 2009;150:2865–2872.
Ungefroren H, Ergun S, Krull NB, Holstein AF. Expression of the smallproteoglycans biglycan and decorin in the adult human testis. BiolReprod 1995;52:1095–1105.
Van Straaten JF, Coers W, Noordhoek JA, Huitema S, Flipsen JT,Kauffman HF, Timens W, Postma DS. Proteoglycan changes in theextracellular matrix of lung tissue from patients with pulmonaryemphysema. Mod Pathol 1999;12:697–705.
Wegrowski Y, Paltot V, Gillery P, Kalis B, Randoux A, Maquart FX.Stimulation of sulphated glycosaminoglycan and decorin production inadult dermal fibroblasts by recombinant human interleukin-4. BiochemJ 1995;307:673–678.
Welsh M, Saunders PT, Atanassova N, Sharpe RM, Smith LB. Androgenaction via testicular peritubular myoid cells is essential for malefertility. FASEB J 2009;23:4218–4230.
Zhang C, Yeh S, Chen YT, Wu CC, Chuang KH, Lin HY, Wang RS,Chang YJ, Mendis-Handagama C, Hu L et al. Oligozoospermia withnormal fertility in male mice lacking the androgen receptor in testisperitubular myoid cells. Proc Natl Acad Sci USA 2006;103:17718–17723.
Zhu JX, Goldoni S, Bix G, Owens RT, McQuillan DJ, Reed CC, Iozzo RV.Decorin evokes protracted internalization and degradation of theepidermal growth factor receptor via caveolar endocytosis. J BiolChem 2005;280:32468–32479.
Decorin and human male infertility 2625
Downloaded from https://academic.oup.com/humrep/article-abstract/26/10/2613/609326by gueston 03 April 2018





























